 Raúl O. Gómez
Raúl O. Gómez Celeste M. Pérez-Ben
Celeste M. Pérez-Ben- 1CONICET-Departamento de Ciencias Geológicas, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
- 2Departamento de Biodiversidad y Biología Experimental, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina
Within the already peculiar Bauplan of anurans, pipid frogs have evolved an array of bizarre features that are commonly linked to their highly aquatic lifestyle. Among the latter, there are several distinctive sacro-caudo-pelvic features shared by extant pipids, which have been regarded as evolutionary novelties taking part of a specialized fore-aft-sliding ilio-sacral joint. Pipids and their kin (pipimorphs) have a rich fossil record documenting 130 million years of uninterrupted evolution in aquatic environments, which, along with our current understanding of their phylogeny and recently available musculoskeletal data, allows us to inquire on the patterns and processes that have led to their distinctive sacro-caudo-pelvic system with a deep-time perspective. Here, we take a phylomorphospace approach based on discrete character matrices and a scaffold tree derived from recent studies, making comparisons of morphospace occupation between pipids and other anurans, and morphospace occupation, disparity, levels of homoplasy, and shared evolving characters between different groups and/or over time across pipimorphs. In doing so, we focus on trends of morphological diversification and discuss the potential role that ecological and developmental constraints might have had in driving the evolution of the sacro-caudo-pelvic complex of pipid frogs. Our main findings reveal a pattern of continuous and parallel innovation early in the history of pipids, followed by arrested evolution of novel morphologies toward the Recent. The latter, in turn, is mirrored by rampant homoplasy in the ilio-sacral sliding joint among extant pipid frogs. This study highlights the importance of fossils in revealing macroevolutionary patterns that will be otherwise veiled based on neontological data alone.
Introduction
Anurans (frogs and toads) have a distinctive Bauplan (Handrigan and Wassersug, 2007) that makes them “the most easily diagnosed of all vertebrate groups” (Griffiths, 1963, p. 241). Within this body plan, the sacro-caudo-pelvic complex assembles many of the prominent musculo-skeletal fingerprints of adult frogs that are unique among vertebrates, including a single caudal element (urostyle) lying between very long ilia that posteriorly contact one another and with a compound ischiopubis, whereas anteriorly, they articulate with the ventral face of sacral diapophyses, which, in turn, result from the evolutionary integration of sacral ribs and vertebra into a sacrum (Jenkins and Shubin, 1998; Reilly and Jorgensen, 2011; Lires et al., 2016). This suite of features constitutes a well-integrated functional complex (Emerson, 1982) that has been largely conserved over the history of anurans (Jenkins and Shubin, 1998; Lires et al., 2016), although it has diversified into a relatively limited range of morphs in relation to locomotion and microhabitat use (Emerson, 1979; Reilly and Jorgensen, 2011). Perhaps one of the most extreme morphologies within this already peculiar Bauplan is displayed by the early diverging but derived frogs of the family Pipidae, bearing an array of bizarre features amongst anurans that has usually been linked to their highly aquatic lifestyle (Cannatella and Trueb, 1988; Trueb, 1996; Báez et al., 2012; Cannatella, 2015).
Extant pipid species, arranged in the three distinct subclades Pipa, Xenopodinae (Xe, Silurana and Xenopus), and Hymenochirini (Hymenochirus and Pseudhymenochirus), inhabit a variety of freshwater environments at both sides of the Atlantic Ocean (Bewick et al., 2012; Gómez, 2016). Pipa occurs in tropical South America and Panama, whereas xenopodines and hymenochirines are found in sub-saharan Africa (Frost, 2018). They share several characteristic sacro-caudo-pelvic features (Figure 1), including an urostyle solidly fused to a sacrum having broadly expanded, flat, bony-sided diapophyses, a ‘U’-shaped pelvic girdle resulting from a broad interiliac symphysis, with ilia often bearing well-developed crests, all of which have been regarded as novelties of extant pipids that together readily distinguish them from other anurans (Cannatella and Trueb, 1988; Trueb, 1996; Báez et al., 2008; Reilly and Jorgensen, 2011). However, some of these features entail more variation than it is commonly acknowledged (Přikryl et al., 2009; Báez et al., 2012; Gómez, 2016). In turn, this configuration of the sacro-caudo-pelvic complex takes part of a specialized system with a Type I ilio-sacral joint allowing extensive fore-aft-sliding movements of the pelvic girdle relative to the sacrourostyle (Emerson, 1979, 1982; Figure 1A), which has variously been regarded as functionally important in swimming locomotion, burrowing, prey capture, ventilation, hydrostatics, avoiding potential threats, or emetic behavior (Willem, 1938; Palmer, 1960; Whiting, 1961; Videler and Jorna, 1985; Naitoh et al., 1996; Van Dijk, 2002; Robovska-Havelková et al., 2014).
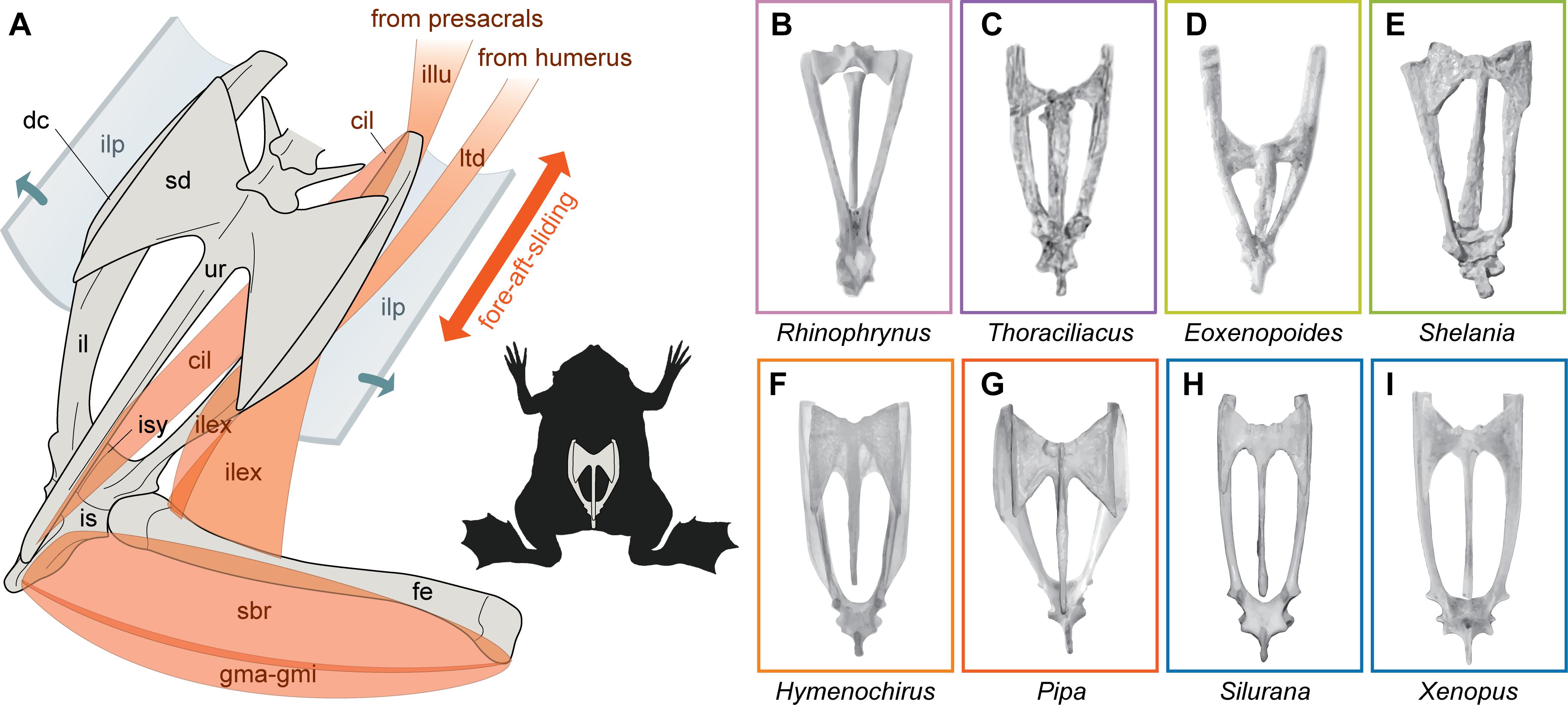
Figure 1. Sacro-caudo-pelvic complex of pipid frogs and their kin. Diagram of the fore-aft-sliding system in Pipa pipa (A), showing major distinct features of the sacrourostyle and pelvic girdle of pipids along with some relevant pelvic and thigh muscles (red-shaded) that attach to them, as well as the main direction of movement at the ilio-sacral joint (double arrow); the ilio-ilial ligamentous plate is shown dissected and unfolded (short arrows). Sacro-caudo-pelvic complex of extant and extinct xenoanurans (B-I): Rhinophrynus dorsalis CFA-An-275 (B), Thoraciliacus rostriceps† HUJZ F93 (C), Eoxenopoides reuningi† SAM K-4956 (D), Shelania pascuali† CPBA-V 12219 (E), Hymenochirus boettgeri MACN-He 42621 (F), Pipa pipa MACN-He 42612 (G), Silurana tropicalis MACN-He 42633 (H), Xenopus laevis AMNH 177087 (I); not to the same scale. †, extinct; cil, coccygeoiliacus; dc, dorsal crest; fe, femur; gma-gmi, gracilis major-gracilis minor; il, ilium; ilex, iliacus externus; illu, iliolumbaris; ilp, ilio-ilial ligamentous plate; is, ischium; isy, interiliac symphysis; ltd, latissimus dorsi; sbr, semimembranosus; sd, sacral diapophysis; ur, urostyle. Institutional abbreviations: AMNH, American Museum of Natural History, New York, NY, United States; CFA, Colección Anfibios, Fundación Félix de Azara-Universidad Maimónides, Buenos Aires, Argentina; CPBA-V, Palaeontología de Vertebrados, Departamento de Ciencias Geológicas, Universidad de Buenos Aires, Buenos Aires, Argentina; MACN-He, Herpetología, Museo Argentino de Ciencias Naturales ‘Bernardino Rivadavia,’ Buenos Aires, Argentina; SAM, Iziko South African Museum, Cape Town, South Africa.
Pipid frogs and their kin have a long history since they diverged from their sister group Rhinophrynus in the Jurassic at least 150 Ma ago (Roelants et al., 2007, 2011; Bewick et al., 2012; Cannatella, 2015; Gómez, 2016), the last 130 million years of which is documented by a rich fossil record scattered across South America and Africa, but also Europe and the Arabian Plate, documenting a continuous evolution in aquatic environments and a wider geographic distribution in the past (Báez, 2013; Cannatella, 2015; Gómez, 2016). As privileged bearers of time, but also a source of unique morphologies that might not be represented in present-day biodiversity, fossils may play an important role in systematics and other historical endeavors (e.g., Donoghue et al., 1989; Smith and Turner, 2005; Pyron, 2015). Taking benefit from these qualities, the fossil record of pipids and allies has been studied with reasonable success along with neontological, morphological and/or molecular, data to enable disentangling and dating of the phylogenetic and biogeographic history of pipids (Báez et al., 2012; Cannatella, 2015; Evans et al., 2015; Gómez, 2016). Moreover, the integration of fossils and neontological data in an explicit phylogenetic framework can also shed light on the origin and early evolution of morphological innovations or even entire Baupläne (Slater and Harmon, 2013; Brusatte et al., 2014; Lires et al., 2016), though the unique anatomy of pipids has not yet been explored with these aims.
The rich fossil record of pipids and their kin, along with our current understanding of their phylogeny and recently available data on morphological variation in the musculoskeletal system of extant and extinct forms, allow us to inquire on the patterns and processes that have led to their specialized sacro-caudo-pelvic system with a deep-time perspective. Here, we take a phylomorphospace approach (Sidlauskas, 2008) based on a discrete character matrix and a scaffold tree derived from recent studies, making comparisons of morphospace occupation between pipid frogs and other anurans, and morphospace occupation, disparity, levels of homoplasy and shared evolving characters between different groups and/or over time across pipimorphs. In doing so, we focus on trends of morphological diversification and discuss the potential role that ecological and developmental constraints might have had in driving the evolution of the sacro-caudo-pelvic complex of pipid frogs.
Materials and Methods
Anatomical Terminology and Systematic Nomenclature
Musculo-skeletal terminology of the sacro-caudo-pelvic complex (Figure 1A) follows those of Přikryl et al. (2009); Gómez and Turazzini (2016), and Gómez (2016), and references cited therein. Taxonomy of extant species, genera, and families follows Frost (2018), with the single exception of the genus Silurana, which is considered distinct from Xenopus. This distinction is based on different lines of evidence, including the recognition of diagnostic morphological features (Cannatella and Trueb, 1988; Evans et al., 2015; Gómez, 2016) as well as their deep genetic divergence and concomitant inferred deep divergence time (Bewick et al., 2012; Evans et al., 2015). Systematic nomenclature regarding high-rank taxa follows that used by Gómez (2016) and authors cited therein. In this regard, Pipidae was kept for the crown group defined by extant pipids, whereas for the respective total group we used the name Pipimorpha (sensu Ford and Cannatella, 1993). Similarly, we used Pipinae, Hymenochirini, Pipa, and Xe as crown-based names for the corresponding pipid subclades, whereas Pipinomorpha (Pm) and Xenopodinomorpha (Xm) are used for the stem-based groups that represent the main split of crown-group pipids (Supplementary Figure S1). Extinct taxa along the stem of Pipidae are hereby informally referred to as ‘basal pipimorphs (Bp).’
Taxon Sampling
Our sampling of Pipidae included all the extant and extinct species considered by Gómez (2016), plus several additional extant pipids, totalizing 25 extant species of the 41 currently recognized (eight of which have been erected during the last decade; Evans et al., 2015; Frost, 2018) plus 11 fossil forms. Extant taxa include all species of Pipa, almost all hymenochirines, and half of the species of Xenopodineae, which belong to different lineages all across this pipid subclade, so there is a good representation of extant pipid diversity. We also included all the extinct pipimorphs lying outside crown group Pipidae, as well as Rhinophrynus and its extinct allies, considered in recent phylogenetic studies of the group (e.g., Báez et al., 2012; Cannatella, 2015; Gómez, 2016). In addition, to better contextualize the unique morphology of pipids, we expanded the matrix by adding several non-pipoid anuran taxa including Ascaphus, Leiopelma, costatans (Discoglossus, Latonia, Alytes, Bombina, Barbourula), anomocoelans (Pelodytes, schaphiopodids, pelobatids, megophryids), and main neobatrachian clades (Heleophrynus, calyptocephalellid and nobleobatrachian hyloids, and microhylid, afrobatrachian, and natatanuran ranoids) (Supplementary Material).
Character Sampling
The morphology of the sacro-caudo-pelvic complex was represented by the discrete characters concerning the sacrum, urostyle, and pelvic girdle used by Gómez (2016) in his phylogenetic analyses of pipoid frogs, with the addition of three characters of the urostyle that were recently considered in the phylogenetic analysis of Báez and Gómez (2018). Also, several character states were added to incorporate the morphology exhibited by anuran taxa that have not been included in the analysis of Gómez (2016). The complete data matrix consists of 34 characters (8 of the sacrum; 5 of the urostyle; 21 of the pelvic girdle) scored for 49 and 95 terminal taxa, in its non-expanded and expanded versions, respectively (for character list and data matrix see Supplementary Material). It is noteworthy that most of these characters, although with some modifications, have also been considered by Báez and Gómez (2018) in their phylogenetic analysis, focused on neobatrachians. We are confident that this data matrix is also reliable for our present aims, since it is inclusive regarding characters of the sacro-caudo-pelvic complex, disrespecting whether they are invariable across pipoids or their variation is restricted to autapomorphies (see Lloyd, 2016).
Scaffold Time-Tree
We constructed a scaffold time-tree (Supplementary Figure S1) mainly derived from the parsimony results of Gómez (2016), in combination with hypotheses derived from different studies concerning the internal relationships of extant Hymenochirus, Pipa, and Xenopus (Cannatella and Trueb, 1988; Pyron, 2014; Evans et al., 2015). It is noteworthy that the general phylogenetic arrangement of extant lineages, as well as the position of different extinct taxa is relatively well supported, based on morphology alone, molecular data, or a combination of both (Báez et al., 2012; Bewick et al., 2012; Cannatella, 2015; Evans et al., 2015; Gómez, 2016). The ages of fossils were updated based on available data of the respective fossil-bearing stratigraphic units and under the current chronostratigraphic framework (Cohen et al., 2013, updated). Numerical ages of fossil terminals used for the time-tree were derived from the latter as the minimum (i.e., youngest) possible age of the oldest stratigraphic unit from which a particular extinct taxon is known. Node ages were derived from previous molecular-based estimates whenever possible (Bewick et al., 2012; Cannatella, 2015; Evans et al., 2015) or from the ages of fossils following the approach of Marjanović and Laurin (2014), assigning a minimum of 3 Myrs for internodes. After calibration, we pruned the extinct species that were scored for only 50% or less of the characters, namely Gracilibatrachus avallei, Pachycentrata taqueti; Vulcanobatrachus mandelai, Llankibatrachus truebae, Shelania laurenti, and Oumtkoutia anae. Because performed quantitative analyses require a fully dichotomous topology, we carried out all the analyses on the three possible solutions (referred below as trees 1, 2, and 3) of the single polytomy on the tree, including the extinct Kuruleufenia xenopoides and ‘Xenopus’ romeri together with crown group Xe. The lengths of the new branches that resulted from solving the polytomy were set to the minimum branch length found in the rest of the tree. To perform some of the analyses explained below, we pruned from each tree the fossils and the outgroup taxa, alternatively. Because the single polytomy includes two extinct species, pruning extinct taxa led to a completely resolved tree.
Ancestral State Reconstruction and Distance Matrices
Based on the non-expanded character matrix described above (Supplementary Material), we reconstructed the ancestral states of the characters for every node using Maximum-Likelihood (ML) methods implemented in the function AncStateEstMatrix of the R package Claddis (Lloyd, 2016). We also calculated the reconstructions based on the character matrices from which fossil or outgroup taxa, alternatively, were excluded. For the character matrix with all taxa and that without outgroups, the reconstruction was made three times, one for each of the three fully dichotomous trees. Multistate characters were treated as unordered because the data matrix is mainly derived from that of Gómez (2016) and since in that phylogenetic analysis multistate characters were treated as unordered, we opt to treat the characters as they were originally conceived for consistency. However, we reconstructed the ancestral states with the following multistate characters considered as ordered: 1, 3, 4, 15, 16, 19, 23, 28, and 29 for comparative purposes. From the three topologies, only tree 1 was considered for the reconstruction with ordered characters. We added the ancestors with their reconstructed states to the respective matrices of terminals and, from these combined matrices, we calculated distance matrices using two metrics: generalized Euclidean distance (GED) and maximum observable rescaled distance (MORD; Lloyd, 2016). Both distance metrics are designed to cope with missing data, which is expected when considering extinct taxa, as in this study. GED is similar to the raw Euclidean distance, but it uses the calculable distances to estimate those that are missing. In contrast, when MORD is applied, calculable distances are rescaled by dividing them by the maximum realizable distance based on the characters observed (Lloyd, 2016). It should be noted, however, that the total amount of missing data in the non-expanded data matrix (with a higher proportion of extinct taxa) is relatively low (15.8%). The distance matrices were calculated by the function MorphDistMatrixFast of Claddis (Lloyd, 2016), with an arcsine square root transformation when MORD was used as the distance metric, as suggested by Lloyd (2016). In addition, distance matrices using the same metrics were calculated for the expanded matrix, excluding fossils. All these analyses were carried out in R 3.3.0 (R Core Team, 2016).
Morphospace Occupation
We applied principal coordinates (PCO) to a subset of the distance matrices produced to ordinate the data in morphospaces where we compared the occupation of different sets of groups. To perform the PCOs, we used the cmdscale function of R, adding a correction to avoid the negative eigenvalues that could result when dealing with distance matrices based on unordered multivariate characters and/or missing data (Hopkins, 2016; Guillerme and Cooper, 2018), as the ones used here. In the morphospaces derived from distance matrices calculated from the expanded matrix, we tested whether extant pipids occupy a distinctive domain amongst anurans. In the phylomorphospaces derived from the non-expanded matrix, only including extant pipids and selected outgroup taxa, we compared the occupation of the main pipid subclades Xe and Pipinae Whereas, in those also including fossils, we made pairwise comparisons of the occupation between: outgroup taxa, extant pipids, extinct crown-group pipids, and Bp along the stem of Pipidae; each group encompassed terminal taxa and their respective nodes. We assessed the significance of differences in morphospace occupation using PERMANOVA, as implemented in the adonis function the R package vegan (Oksanen et al., 2016), applying the Holm-Bonferroni correction to the p-values for comparison. All axes of the PCOs were considered in the PERMANOVA. To account for the differences in sample sizes, we bootstrapped the samples 1000 times, rarefying them to the smallest sample size among the groups being compared. The PERMANOVA was carried out in each iteration, resulting in 1000 p-values. This bootstrapping was not performed for the comparison of the two living clades of pipids because the sample sizes were similar. The assumption of homogeneous multivariate dispersion among groups was tested by pairwise comparisons of groups using the functions betadisper and permutest of the same package. All pairwise comparisons of multivariate dispersion resulted as non-significant, except between extant pipids and the expanded sample of outgroup taxa. However, the heterogeneity in dispersions of these two groups was not problematic for the analysis with the rarefied samples because PERMANOVA is largely unaffected by heterogeneity for balanced designs (Anderson and Walsh, 2013). All these analyses were carried out in R 3.3.0 (R Core Team, 2016).
To visualize the general patterns of sacro-caudo-pelvic morphospace occupation, we performed non-metric Multidimensional Scaling analyses (NMDSNMDS). We chose this multivariate method for visualization because the axes of every PCO performed individually explained an extremely small amount of the total variance (Supplementary Figures S2–S5, S20–S23; plots of first PCOs in Supplementary Figures S6–S8, S24–S26) and, thus, the visualization of patterns would have required the inspection of a high number of dimensions. In contrast to PCO, NMDS is a method that seeks to optimize the fit between the data and a preselected number of axes, allowing visualization of multivariate data in few bivariate scatter plots. Morphospaces were visualized through NMDS from the distance matrices calculated from: (a) the expanded character matrix; (b) the non-expanded character matrix containing the total taxa and ancestral reconstructions; and (c) and the non-expanded character matrix excluding fossil species, and ancestral reconstructions. We acknowledge that reducing a large number of dimensions to only a few is a crude approximation of the original morphospace, thus we reduced the data to three dimensions, which was a trade-off between a low number of axes to visualize and low stress values (Supplementary Tables S1, S5). For each NMDS, we made bivariate scatter plots for every pair of axes and, for (b) and (c), we superimposed the respective tree topologies, generating phylomorphospaces to better understand the relationships among taxa. These plots were used for visualization only and all calculations were based on the PCOs. All these analyses were carried out in R 3.3.0 (R Core Team, 2016).
Disparity and Diversity
We applied PCO to the distance matrices built from the non-expanded character matrix without outgroups to generate multidimensional morphospaces within which measures of disparity could be calculated. We quantified disparity from the scores of tips and nodes on the totality of PCOs axes using the dispRity R package (Guillerme, 2018). Two disparity metrics were calculated: sum of variances (SOV) and sum of ranges (SOR). Disparity was calculated including terminal taxa and reconstructed ancestors and compared between groups and time bins. Groups included: all extant pipids, further subdivided into Xe and Pipinae, and fossils. Four-time bins were defined following chronological divisions: Early Cretaceous, Late Cretaceous, Paleogene, and Neogene–Quaternary. All these groupings have relatively low levels of missing data, ranging from 1% for Pipinae up to 15% for fossils, and from 5% for the Neogene–Quaternary up to 20% for the Late Cretaceous. We assessed the significance of differences in disparity between groups (i.e., groups of taxa and time bins) by the overlap or non-overlap of 95% bootstrap confidence intervals. These intervals were calculated by bootstrapping each group 1000 times from the PCO matrices and calculating the disparity metrics in each iteration. To account for the differences in sample size, the samples were rarefied to the smallest sample size among the groups being compared. All these analyses were carried out in R 3.3.0 (R Core Team, 2016).
Taxonomic diversity over time was estimated using the same time bins as for disparity and calculated, taking into account terminals and reconstructed ancestors, as well as ghost lineages implied by our preferred topology, following previous approximations (e.g., Marjanović and Laurin, 2008; Halliday and Goswami, 2016).
Character Evolution and Homoplasy
Patterns of repeated evolution of the same character state (i.e., homoplasy) in the stem-based clades of the main crown pipid dichotomy, Xm and Pm, lineages of crown group pipids, Xenopodinae (Xe), Pipa (Pi), and Hymenochirini (Hy; including extinct Singidella), and the paraphyletic array of Bp (Bp) lying outside crown group Pipidae were examined following an approach akin to that of Roelants et al. (2011). Computation of the homoplasy index (HI), complementary of the consistency index (Kluge and Farris, 1969), was done on the ML character optimization. Also, state-specific HIs were computed manually for each lineage. In all cases computation of indices was done excluding terminal branches to avoid autapomorphies. We conducted all pairwise comparisons between groups and the level of significance of the shared homoplasy level was estimated as in Roelants et al. (2011), by calculating the probability P(h) of observed shared homoplastic states: (h) if all changes had equal probability as well as the probability, P(e) of observed shared evolving characters, (e) if all characters are randomly drawn from the same set of 34 characters.
Results
Morphospace Occupation
Morphospaces constructed with different distance metrics (GED, MORD) and based on the three possible resolutions of the polytomy in the scaffold tree (i.e., nine resulting morphospaces) show overall similar results, and thus, only those based on GED and tree 1 are further commented on. All results are available in the Supplementary Material.
Qualitative inspection of the 3D NMDS plot for extant anurans shows that pipids occupy a distinct domain of the morphospace (Figure 2 and Supplementary Figures S9, S27), which is significantly separate from that of all other clades (Table 1), even considering other extant taxa distantly related to pipids that are also mostly aquatic or semi-aquatic or share the same overall type of sacroilial joint. This was anticipated since most of the discrete character matrix, upon which the morphospace is constructed, has originally been designed for resolving internal pipoid relationships (Gómez, 2016). Nevertheless, it should be noted that Rhinophrynus is in the antipode of pipids in this morphospace, in sharp contrast with their sister-group relationship. Moreover, a few characters and several extra character states were added to the original data matrix to cope with the variation present in the anuran taxa added herein, many of which do not cluster in the morphospace according to their phylogenetic affinities. Some anuran clades other than pipids occupy more or less restricted areas of the morphospace, including Ascaphus, Leiopelma, and anomocoelans among ‘archaeobatrachians’ and microhylids, natatanuran ranoids, and some hyloid clades among neobatrachians (Figure 2), whereas some pairs of closely related taxa (e.g., bombinatorids Bombina and Barbourula; calyptocephallelids Calyptocephalella and Telmatobufo) occupy relatively distant regions. In any case, most non-pipid anurans are restricted to a relatively distinct domain of the morphospace. Aside from pipids and Rhinophrynus, the most remarkable departure from their close relatives is perhaps that of the aquatic frog Barbourula, which most closely approximates the pipid condition among extant anurans. This occupation pattern is suggestive of a general historical constraint with a superimposed ecological signal and highlights the uniqueness of the pipid sacro-caudo-pelvic complex amongst anurans.
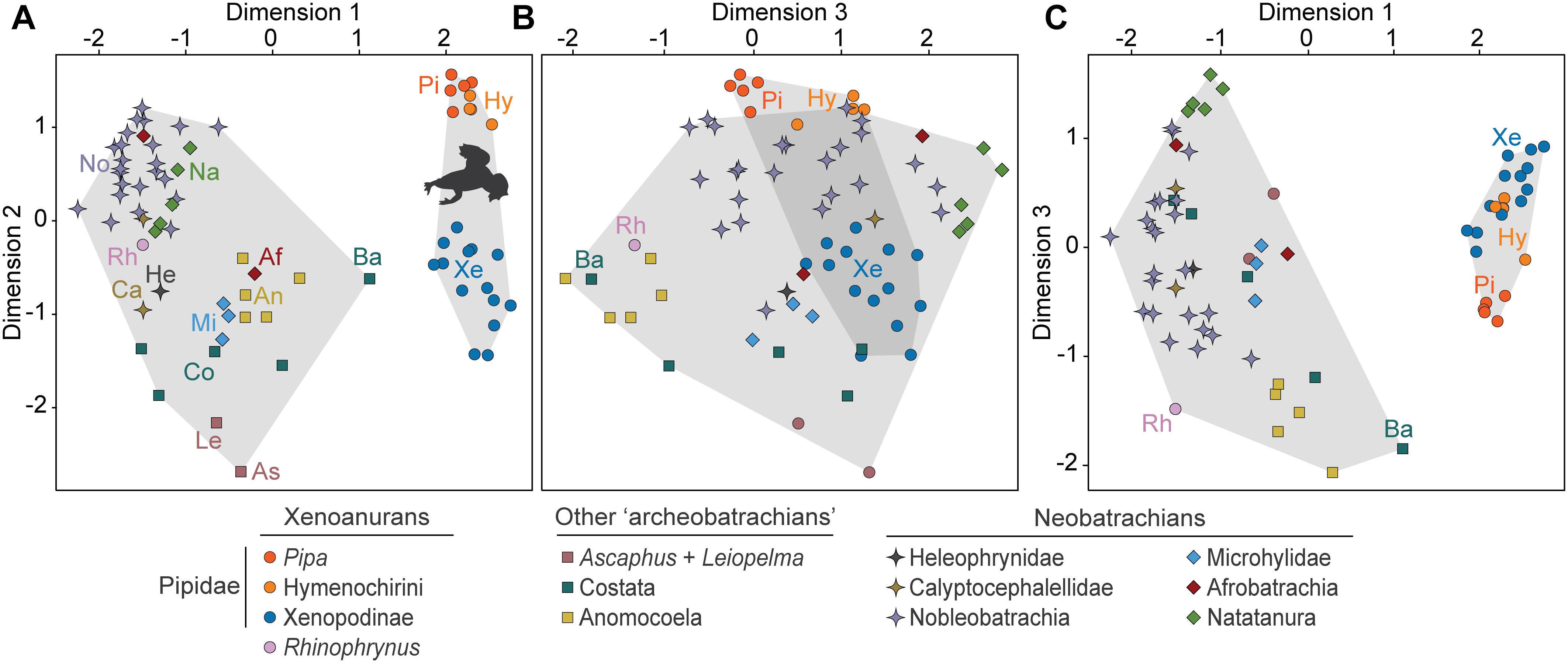
Figure 2. 3D Morphospace of the sacro-caudo-pelvic complex of extant pipids relative to other extant anurans (A-C) constructed by NMDS (Stress: 0.102) of the GED matrix. Gray-shaded polygons depict the groups for which morphospace occupation is compared. Af, Afrobatrachia; An, Anomocoela; As, Ascaphus; Ba, Barbourula; Ca, Calyptocephalellidae; Co, Costata; He, Heleophrynidae; Hy, Hymenochirini; Le, Leiopelma; Mi, Microhylidae; Na, Natatanura; No, Nobleobatrachia; Pi, Pipa; Rh, Rhynophrynus; Xe, Xenopodinae.
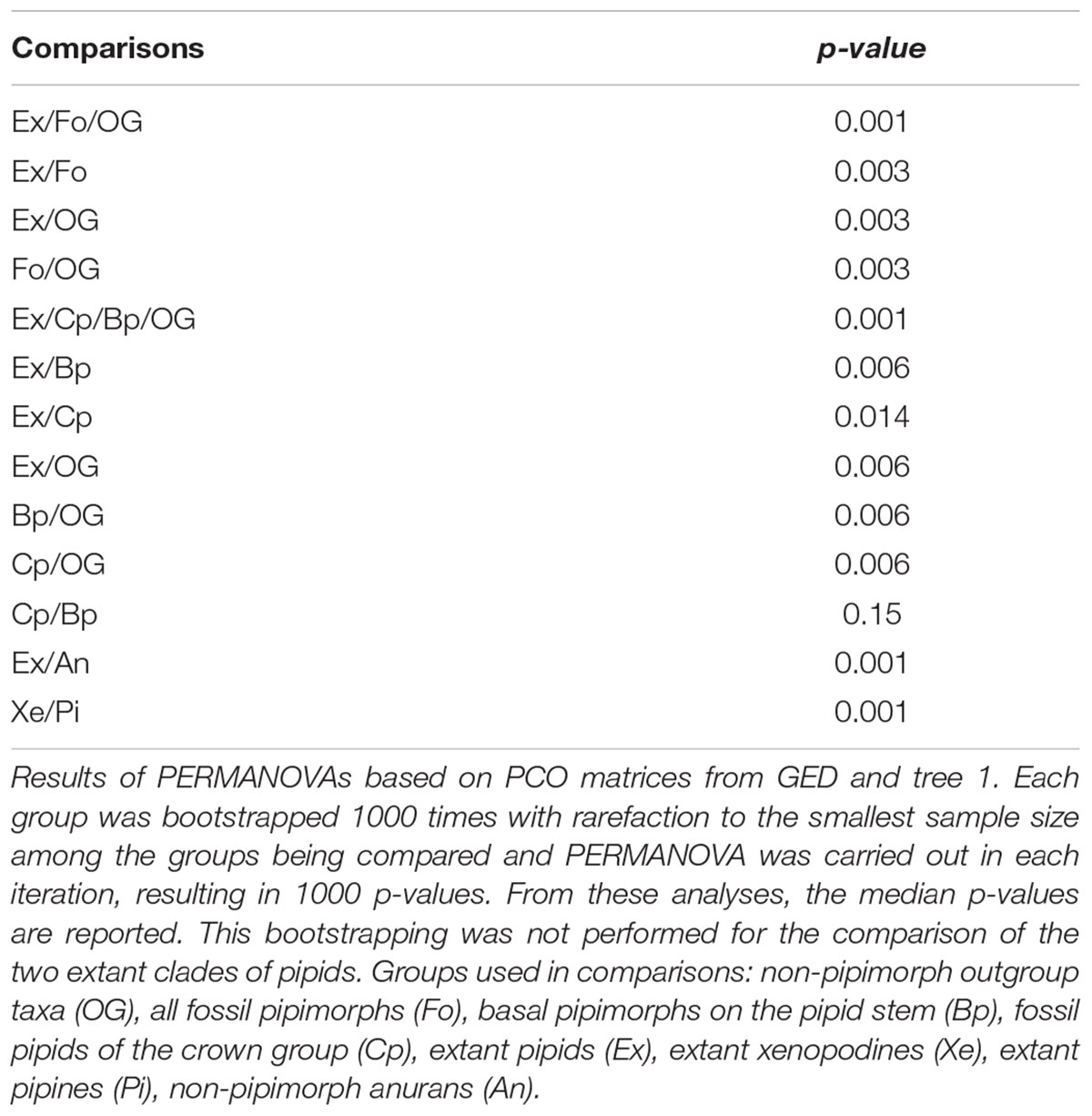
Table 1. Morphospace occupation comparisons between different anuran groups.
Visualization of the NMDS phylomorphospace constructed upon a subset of extant taxa including extant pipids, their sister group Rhinophrynus, a few basal anurans (non-pipanuran taxa), plus the ML reconstructed ancestors shows that different anuran lineages have explored the sacro-caudo-pelvic morphospace in opposite directions from the ancestral condition, with pipids set apart from the remaining extant taxa (Figures 3A–C and Supplementary Figures S10, S28). In this regard, the longest branch in this phylomorphospace is that leading to Pipidae, with their reconstructed common ancestor, as most internal nodes, falling within the morphospace envelope circumscribed by pipid terminal taxa. Additionally, the main divergence within Pipidae implied by the scaffold tree, namely that between Pipinae (Hymenochirini + Pipa) and Xe (Silurana + Xenopus), is also boldly reflected in morphospace occupation, in which each subclade occupies a significantly distinct region within the pipid domain (Table 1 and Supplementary Tables S2, S6). Unexpectedly, the reconstructed common ancestor of pipids appears noticeably nearer to Pipinae than to Xe, which mirrors the actual distances in the distance matrices and contrasts with the general perception of xenopodines as more representative of the ancestral pipid morphology than pipines (e.g., Cannatella and Trueb, 1988; Báez et al., 2012).
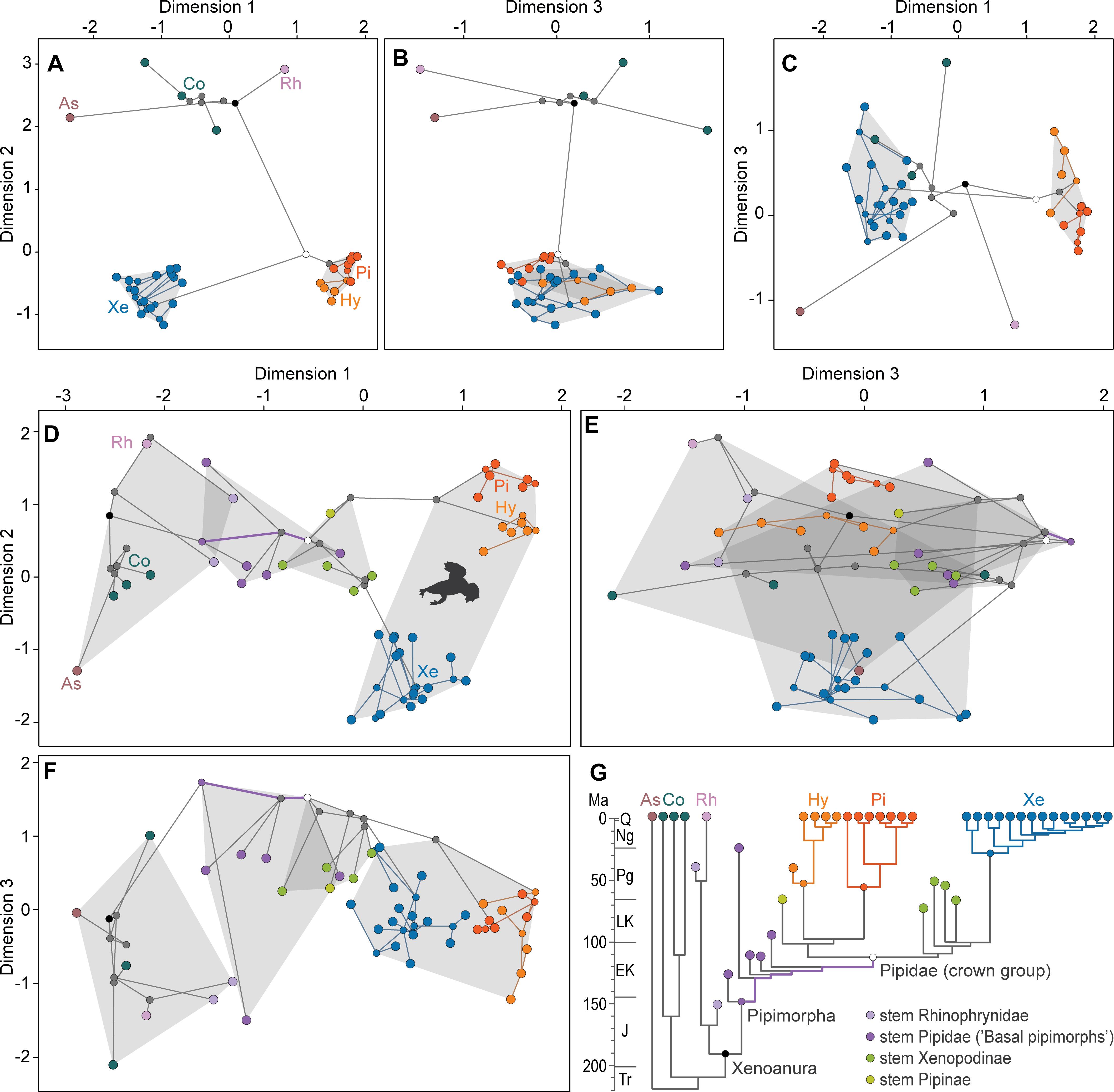
Figure 3. 3D phylomorphospaces of the sacro-caudo-pelvic complex of pipids constructed by NMDS of GED matrices. 3D NMDS plot (Stress: 0.050) of extant pipids and outgroup taxa (A-C) and 3D NMDS plot (Stress: 0.088) of pipimorphs, including fossils, and outgroup taxa (D–F). Gray-shaded polygons depict the groups, encompassing both terminal taxa and the respective nodes, for which morphospace occupation is compared. Key to colors of terminal taxa, branches, and nodes are in the schematic scaffold time-tree (G). EK, Early Cretaceous; J, Jurassic; LK, Late Cretaceous; Ng, Neogene; Pg, Paleogene; Q, Quaternary; Tr, Triassic. Other abbreviations as in Figure 2.
This broad morphological gap between extant pipids and their sister group Rhinophrynus and other extant anurans is partially bridged when fossils are considered, but extant pipids still occupy a distinct domain (Figures 3D–F and Supplementary Figures S11, S29). It is noteworthy that when fossils are considered the domain occupied by crown-group, pipids considerably expands relative to that defined by extant taxa alone (Figures 3A–C and Supplementary Figures S10, S28), with their last common ancestor (node Pipidae; Figure 3G) now occupying a completely different region of the morphospace, separate from the domain of extant pipids (Figures 3D–F and Supplementary Figures S11, S29). Four broad non-monophyletic groupings, namely: (1) outgroup taxa including Rhinophrynus and allies, (2) Bps lying on the stem of Pipidae, (3) fossil crown-group pipids that are part of the stems of Pipinae and Xe, and (4) the crown groups Pipinae and Xe together, occupy more or less distinct domains of this phylomorphospace, although those occupied by fossils on the stem and crown of Pipidae do not differ statistically (Table 1, Supplementary Tables S2, S6, and Supplementary Figures S12, S13, S30, S31). Remarkably, these four groupings are arranged in a roughly linear sequence along the first axis of the NMDS, which is better revealed by the evolutionary path depicted by the deepest reconstructed ancestors (Figures 3D–F and Supplementary Figures S11, S29). This pattern of continuous innovation in deep time toward a distinct domain of the morphospace, after the divergence of pipimorphs from their sister lineage is striking and matches the respective minimal spanning tree across these same nodes. This long-term evolutionary trend over most of the history of pipimorphs somewhat contrasts with the pattern depicted by the different crown subclades of pipids, namely Xe, Hymenochirini, and Pipa, showing that subsequent morphological change over the last 30 Ma occurs within more limited ranges of the morphospace, which is also depicted by the increasing clumpiness in each subclade (Figures 3D–F and Supplementary Figures S11, S29).
Phylogenetic Disparity and Diversity
The non-monophyletic grouping of extant pipids shows a higher disparity than the fossil pipids when SOV is used as disparity metric, regardless the distance metric (GED or MORD) used, the tree topology considered in the analyses, or whether the calculations are carried out with rarefied and unrarefied samples. When disparity is measured as SOR, the same results are recovered when the samples are unrarefied. In contrast, with rarefied samples, extant pipids present a higher disparity than the fossil group, except for the tree 3 when MORD is used as the distance metric (Table 2, Supplementary Tables S3, S7, Figures 4A,B, and Supplementary Figures S14–S16, S32). The differences between the results obtained from SOR with rarefied and unrarefied samples are expected, given that range-based metrics of disparity are sensitive to sample size biases (e.g., Foote, 1992; Ciampaglio et al., 2001; Butler et al., 2012). The opposite results obtained between SOV and SOR with rarefied samples are most likely due to differences in the morphospace occupation pattern of extant and fossil groups, since both SOR and SOV raises with increasing bifurcation of lineages. However, with changes in shape, SOR decreases and SOV increases (Ciampaglio et al., 2001). This is also reflected in the progressively increasing, yet non-significant, rarefied SOR and SOV disparity values over the Cenozoic (see below), which might be due to increasing bifurcation between the main pipid clades represented today.
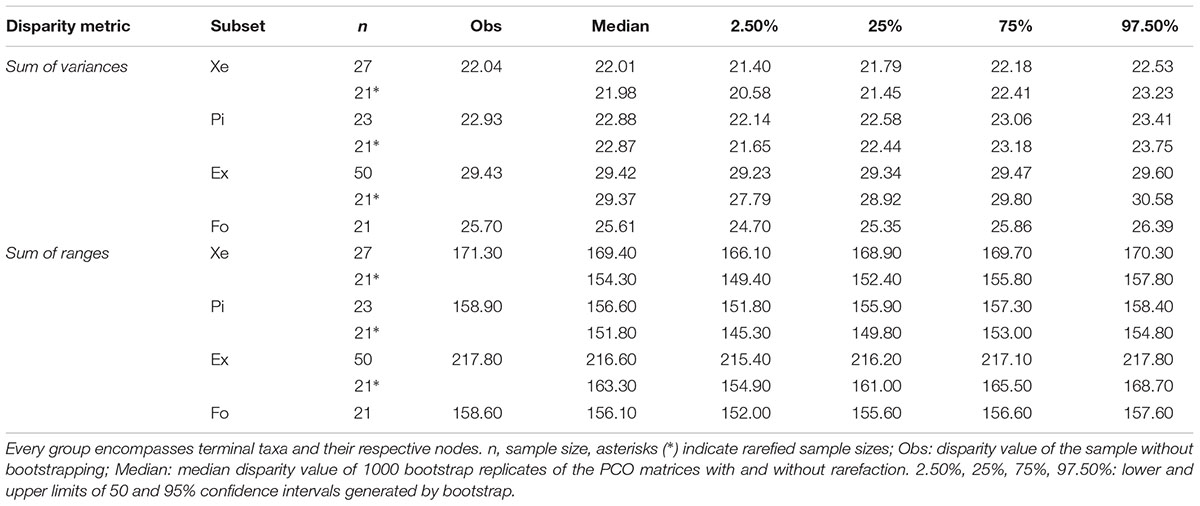
Table 2. Disparity of the sacro-caudo-pelvic complex of extant Pipidae (Ex), its subclades Xenopodinae (Xe) and Pipinae (Pi), and fossil pipimorphs (Fo) based on the tree 1 and GED matrix.
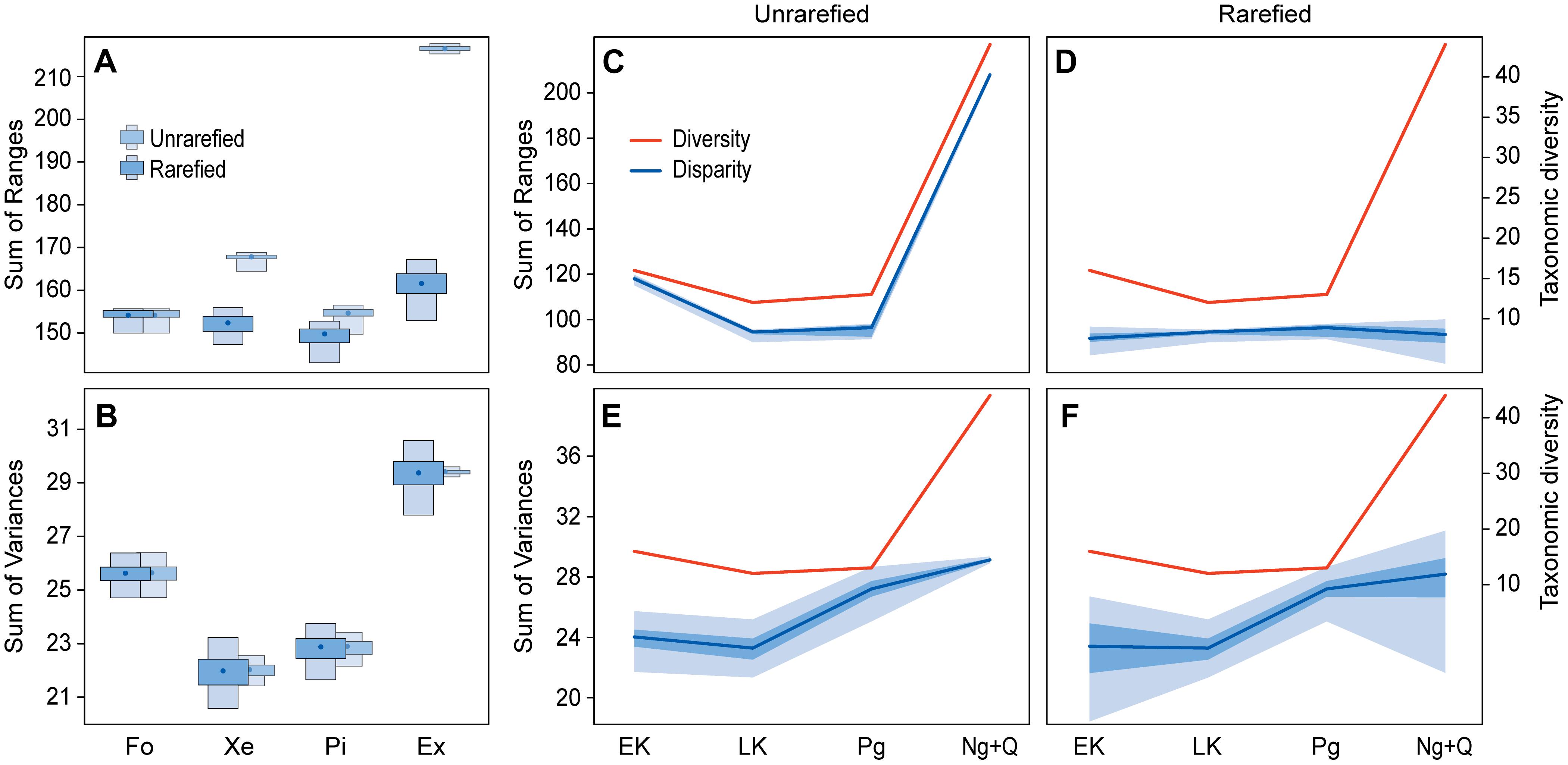
Figure 4. Patterns of disparity in the sacro-caudo-pelvic complex and taxonomic diversity of pipimorphs. Disparity across pipimorphs (A,B) as estimated by sum of ranges (A) and sum of variances (B); black dots indicate medians, boxes show 50 and 90% confidence intervals, and unrarefied values are depicted lighter at the back. Taxonomic diversity (red) and disparity (blue) over time (C–F) as estimated by unrarefied and rarefied sum of ranges (C,D) and sum of variances (E,F); blue lines indicate the central tendency of disparity following the medians of each time bin and bands show 50 and 90% confidence intervals. The disparity of every group and time bin encompasses terminal taxa and nodes and their calculations were based on tree 1 and GED matrices. Ex, extant pipids; Fo, fossil pipimorphs; Ng+Q, Neogene–Quaternary; Pi, Pipinae; Xe, Xenopodinae. Other abbreviations as in Figure 3.
Xenopodines and pipines do not show differences in disparity when samples are rarefied, but their disparities do differ in some cases when samples are unrarified (Table 2, Supplementary Tables S3, S7, Figures 4A,B, and Supplementary Figures S14–S16, S32). Both extant clades consistently show a significantly lower SOV than the fossil grouping, regardless the distance metric (GED or MORD) used, the tree topology considered in the analyses, or whether the calculations are carried out with rarefied and unrarefied samples. When SOR is used to measure disparity and samples are rarefied, there are not significant differences between the two extant clades and fossils. In contrast, most of the SOR analyses with unrarefied samples return higher disparity values for the living clades than for fossils (Table 2, Supplementary Tables S3, S7, Figures 4A,B, and Supplementary Figures S14–S16, S32), probably reflecting, as before, the sensitivity to sample size biases of range-based metrics of disparity.
Significant differences between consecutive time bins are recovered for: (1) unrarefied SOR between Paleogene and Late Cenozoic, which is expected because of the sample size bias; and (2) between Late Cretaceous and Paleogene for all analyses based on the topology 3 and those using SOR of MORD values based on topologies 1 and 2 (Table 3, Supplementary Tables S4, S8, Figures 4C–F, and Supplementary Figures S17–S19, S33). Properly scaled, general trends depicted by rarefied and unrarefied SOV mirror qualitatively the phylogenetic diversity (i.e., including ghost lineages) througout the Mesozoic–Early Cenozoic, with a partial decoupling in the Late Cenozoic when diversity distinctly rises but rarefied disparity values remain fairly constant (Figures 4E,F).
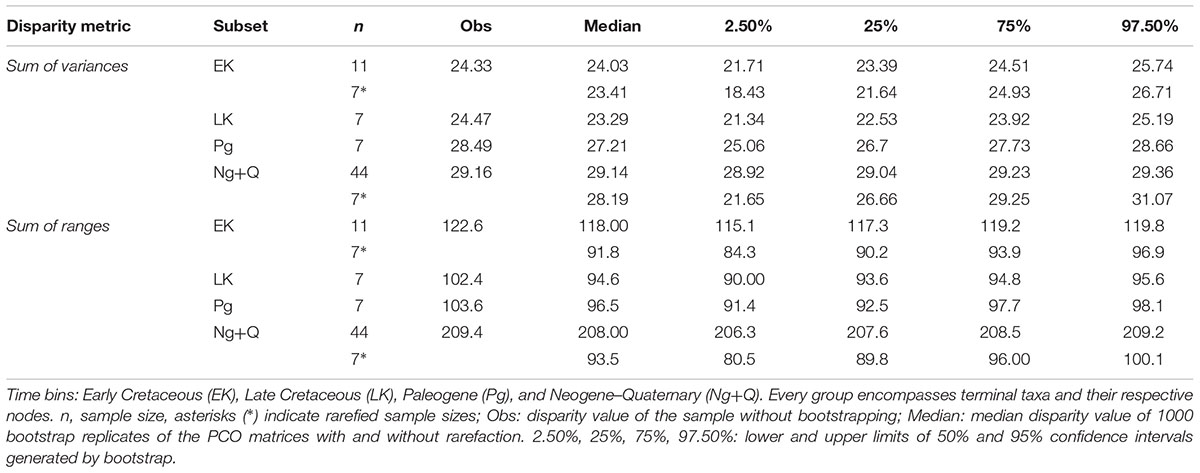
Table 3. Disparity of the sacro-caudo-pelvic complex of pipimorphs over time based on the tree 1 and GED matrix.
Character Evolution and Homoplasy
Levels of homoplasy across the pipimorph tree are relatively low for most characters as indicated by median values of zero for the HI (Figure 5A), although HI mean values appear to correlate with group size as has previously been noticed for the complementary Consistency Index (e.g., Archie, 1989). Nevertheless, state-specific HIs values are also relatively low in all groups with medians of two or less (Figure 5B). Bp show a lower HIs mean value than pipid lineages, whereas, those of the three crown-based pipid subclades (Xe, Pi, Hy) are higher than those of the more inclusive stem-based clades (Xm, Pm). This pattern may not be biased due to underrepresentation of autapomorphies in the original data matrix, since calculations do not take terminal branches into account (Roelants et al., 2011), and indicates an increase in homoplasy toward most recent branches of the pipimorph tree and over geological time. In partial correspondence with the HIs pattern, the number of homoplastic states shared (h) by all pairwise comparisons between lineages of crown-group pipids (Xe, Pi, Hy, Xm, Pm) considerably surpasses numbers expected by chance with high significance (P(h) < 0.01), whereas those shared by Bp and the latter are relatively low (Figure 5C). These values also highlight a raise in homoplasy after diversification of crown group pipids, but in addition denote that there is comparatively more homoplasy between than within the different crown group lineages, taking into account the relatively low HI and His values for all lineages and most characters.
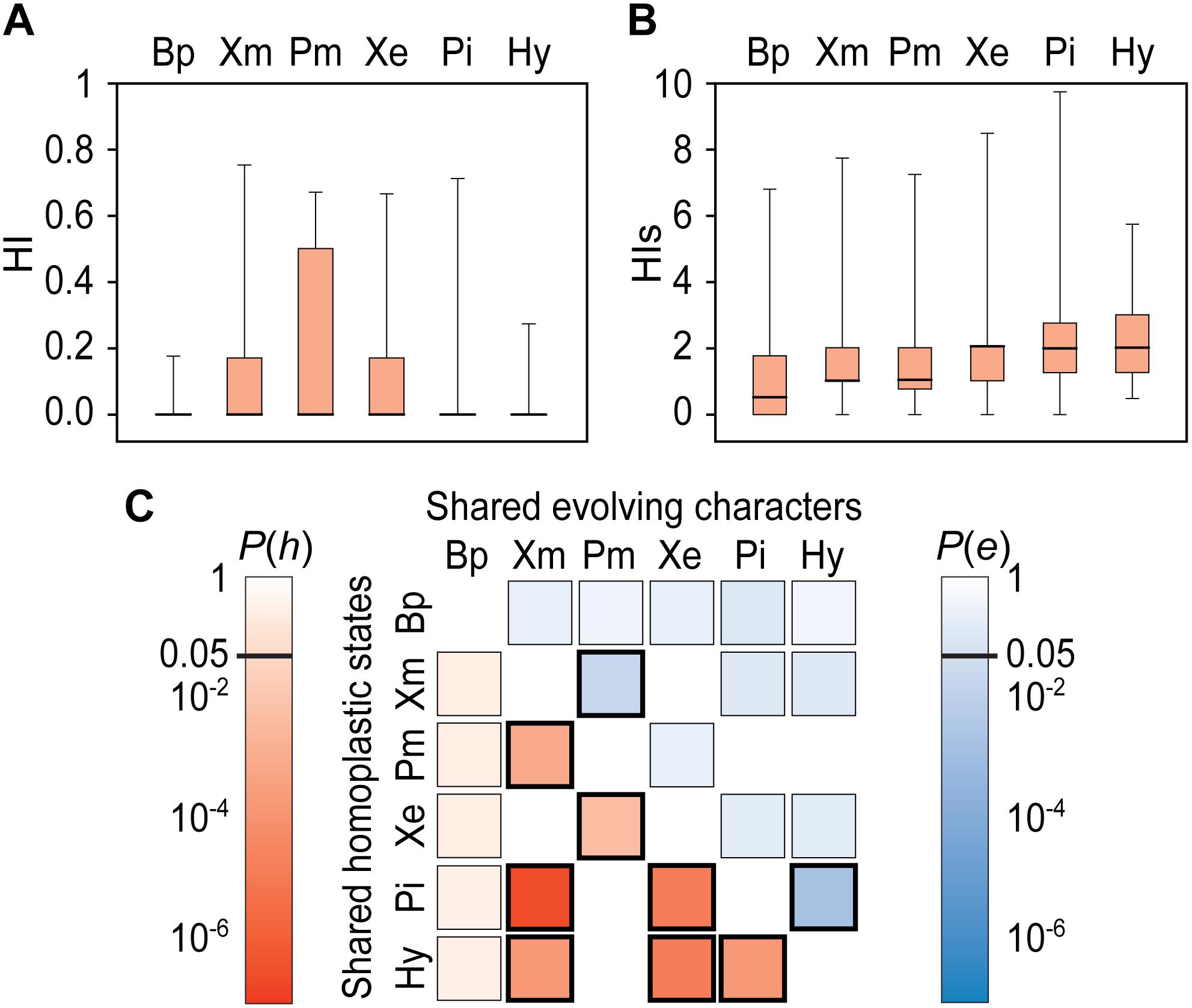
Figure 5. Homoplasy and evolving characters in the sacro-caudo-pelvic complex across the pipimorph clade. Box plots for HI (A) and HIs (B) values, where thick, black lines depict medians, boxes are percentiles 25–75, and whiskers are percentiles 5–95. Pairwise comparisons between groups (C) depict levels of shared homoplastic states in shades of red and shared evolving characters in shades of blue, according to their probability P(h) and P(e), respectively (see text for explanation); probabilities < 0.05 are denoted by thick frames. Bp, basal pipimorphs; Hy, Hymenochirini; Pi, Pipa; Pm, Pipinomorpha; Xe, Xenopodinae; Xm, Xenopodinomorpha.
If the same set of characters of the sacro-caudo-pelvic complex has been evolving across the pipimorph phylogeny, the low shared homoplasy between the Bp array and crown pipid subclades may either be due to state reversals and/or acquisition of new states of these same characters during diversification of the latter. However, results indicate that in all the involved pairwise comparisons between Bp and crown-group pipids (Xe, Pi, Hy, Xm, Pm), the number of shared evolving characters (e) does not exceed values expected when all characters are randomly drawn from the same set of 34 characters (Figure 5C), indicating a shift in the set of evolving characters before and after early cladogenesis of the pipid crown group. Interestingly, amongst all pairs of crown group subclades, significant high values of shared evolving characters are only shown by the main crown pipid split Xenopodinomorpha–Pipinomorpha and the pipine sister taxa Pipa-Hymenochirini [P(e) < 0.05; Figure 5C] and, hence, this suggests that only sister subclades also shared the same set of evolving characters. Moreover, this pattern may also suggest a temporal shift in the set of evolving characters between stem and crown-groups in the xenopodinomorph and pipinomorph lineages. On the other hand, the low number of shared evolving characters between the pairs of more distant crown pipid lineages, coupled with their high number of shared homoplastic states, points to rampant levels of homoplasy in the relatively few characters involved.
Discussion
Unraveling major patterns of morphological diversification and the origin of evolutionary novelties, as well as the underpinning processes, are an important part of ongoing research on paleontology and evolutionary biology, with several studies conducted in diverse groups of organisms and mainly focused on ecomorphology, adaptive radiations, and mass extinctions (e.g., Foote, 1997; Lupia, 1999; Ruta et al., 2006, 2013; Brusatte et al., 2008, 2014; Bapst et al., 2012; Lloyd et al., 2012; Hughes et al., 2013; Colombo et al., 2015; Halliday and Goswami, 2016). Most of these studies recovered a prevailing ‘disparity-first’ pattern of increased innovation and maximum disparity occurring early in the evolution of major clades or in the aftermaths of mass extinctions, followed by a subsequent abrupt rise in taxonomic diversity and deceleration of morphological diversification (e.g., Minter et al., 2017). These ‘early burst’ patterns of diversity have recurrently been explained in the context of an ‘empty ecospace’ hypothesis, whereby an ecological release, either produced by extinction events, the acquisition of morphological novelties (Dumont et al., 2012), or the assembly of a novel body plan (Brusatte et al., 2014), would have allowed the exploitation of a till, then inaccessible niche or resource (Hughes et al., 2013). Alternatively, slowdowns in morphological innovation have been variously interpreted as increasing stabilizing selection, guild structure, saturation of the available ecospace, or genetic/developmental constraints (Foote, 1997; Ciampaglio, 2002; Arthur, 2004; Wagner et al., 2006; Erwin, 2007; Gerber, 2013; Hughes et al., 2013; Minter et al., 2017). Most groups for which disparity is not highest early in their history and is stable or increases more gradually over time have been shown to be subclades of major clades that do exhibit an ‘early burst’ pattern (Foote, 1993, 1997; Wagner, 1997; Erwin, 2007) or to be truncated by the present time or mass extinction events (Hughes et al., 2013). However, it has been emphasized that the recognition of such patterns might ultimately depend upon the scale at which organisms are studied (Slater, 2015; Hopkins, 2016).
Previous work on anurans, which has relied almost exclusively on larval or adult morphology of living taxa, also identified an ‘early burst’ in morphospace occupation and widespread homoplasy over large spatial and temporal scales, suggesting a mix of ecological innovation and constraints along anuran evolution (Roelants et al., 2011; Jorgensen and Reilly, 2013; Moen et al., 2015). However, quite different patterns have been obtained based on the study of larvae or adults. Patterns based on larval characters have shown an early divergence of major lineages (including xenoanurans) toward a small number of distinct morphs prior to the Cretaceous with subsequent arrested morphological diversification around each theme, with the exception of microhylids that represent a new morph originated in the Late Cretaceous (Roelants et al., 2011). In contrast, based on sets of continuous characters of adults, a limited number of highly homoplastic ecomorphs might have evolved repeatedly within each major clade along the anuran history in relation to lineage shifts in locomotor mode (highly aquatic species not included; Jorgensen and Reilly, 2013) and/or microhabitat use (a single pipid species included; Moen et al., 2015). In the latter case, varying degrees of proximity of lineages to particular ‘adaptive optima’ have been related to historical time in the same environment and evolutionary rate of morphological change (Moen et al., 2015).
Pipid frogs, with their peculiar and presumably well-adapted skeletal morphology and good fossil record, documenting a conserved highly aquatic lifestyle for over 130 Ma, provide an interesting model to explore some of these scenarios. Our results depict pipids occupying a fundamentally distinct domain of the sacro-caudo-pelvic morphospace among extant anurans, broadly separated from other aquatic species or with the same overall type of ilio-sacral joint (Emerson, 1979; Reilly and Jorgensen, 2011), the water-dwelling bombinatorid Barbourula being the closest to pipids in morphospace. This agrees with the upheld view of regarding pipids as having a unique sacro-caudo-pelvic morphology easily distinguishable from that of other frogs (Cannatella and Trueb, 1988; Trueb, 1996; Gardner et al., 2010; Reilly and Jorgensen, 2011; Báez et al., 2012), and to some extent with the distinctive xenoanuran larvae (Orton, 1957; Roelants et al., 2011). In this way, pipids appears not to fit the generality of anurans of revisiting over and over the same adult morphologies explored by other lineages (Jorgensen and Reilly, 2013; Moen et al., 2015). It should be noted, however, that this pattern might support the hypothesis of Moen et al. (2015), explaining the differential proximity to ‘adaptive optima,’ since the pipimorph clade is likely to be the one that has continuously evolved in aquatic environments for the most time, followed by the semiaquatic/aquatic bombinatorids, which diversified around 50 Ma (Blackburn et al., 2010). Some ecomorphological signal in our results is also expected, since sacrum, urostyle, and pelvis constitute an integrated morphofunctional complex in anurans, whose major variation within the group has been linked to different locomotor modes (Emerson, 1982; Jenkins and Shubin, 1998; Reilly and Jorgensen, 2011; Jorgensen and Reilly, 2013).
It is remarkable that when fossils are taken into account there is no longer this ample gap between pipids and other anurans, showing a pattern of continuous innovation toward current pipid morphology that challenges the apparent ‘long-fuse’ pattern that would be inferred based on neontological data alone (Roelants et al., 2011; this paper). Apparently contradicting results based alternatively on paleontological and neontological evidence have previously been noticed for other groups (e.g., Harmon et al., 2010; Brusatte et al., 2014; Slater, 2015), stressing the need for proper integration of data from fossils and extant forms in a phylogenetic framework (Slater and Harmon, 2013; Lires et al., 2016). The evolutionary pathway recovered here depicts that the combination of traits that currently characterize the sacro-caudo-pelvic complex of adult pipids have evolved piecemeal up the pipimorph tree, which is also reflected in a temporal shift in the set of evolving characters. Relatively early in their history, Bp attained a compound sacrourostyle, which might account for the absence of coccygeosacralis muscles both in Xenopus and Pipa (Přikryl et al., 2009), and enlarged ischia, where in extant forms originate muscles that retract the femur and flex the knee joint during the swimming kick (Přikryl et al., 2009; Figure 1A). Remarkably, most blueprint features of the sacro-caudo-pelvic complex of pipids that set them apart from most other frogs (e.g., Trueb, 1996; Van Dijk, 2002; Báez et al., 2012), including the broad, flat, bony-sided diapophyses, parallel ilial shafts at their distal portion as a consequence of the morphology at the interiliac symphysis, and well-developed ilial crests where originate large thigh protractors (Přikryl et al., 2009; Figure 1A), evolved in parallel after the main split of crown-group pipids. Noteworthy, the latter features have often been related with the highly aquatic habits of pipids and an extreme fore-aft-sliding ilio-sacral joint (Emerson, 1979; Van Dijk, 2002; Reilly and Jorgensen, 2011). In all pipids, the latter also includes an extensive ilio-ilial ligamentous plate deriving from the dorsal fascia of other anurans and the same sets of muscles may be involved in the sliding motion (Emerson, 1979; Přikryl et al., 2009; Figure 1A). Therefore, it appears that an ilio-sacral joint allowing a fore-aft-sliding movement might have evolved early among pipimorphs, as also suggested by the extreme longitudinal displacement of the pelvis relative to the sacrum in several fossils (Van Dijk, 2002; Figures 1C,D). However, subsequent morphological novelties related to this mechanism might have evolved in parallel within crown group pipids, which is further supported by previous observations on the variation among pipids of these and other musculo-skeletal traits, including the ilial dorsal crest (Báez et al., 2012), the structure and sites of origin of the iliacus externus muscle (protractor of the femur; Figure 1A) on this crest and the ilial shaft (Přikryl et al., 2009).
The pattern of sustained innovation observed in the phylomorphospace (Figures 3D–F and Supplementary Figures S11, S29) and the PERMANOVA results (Table 1 and Supplementary Tables S2, S4) indicate what has been called a driven trend (McShea, 1994) or, more generally, an active trend (Wagner, 1996), showing a long-term displacement in the morphospace away from the ancestral morphologies, which are no longer represented over time (Hopkins, 2016). Even more striking is that fossils revealed that roughly parallel active trends also characterize the main lineages of crown-group pipids since their divergence in the Early Cretaceous more than 110 Ma ago, as depicted by: (1) their pathways in the morphospace; (2) the distinct domain occupied by extant pipids relative to extinct species and ancestors; (3) equal or lower disparity of main extant pipid subclades (i.e., Xe and Pipinae) relative to the paraphyletic array of fossils; (4) the high number of shared evolving characters between main sister subclades. It has been noted that when disparity remains constant or even decreases in particular subclades showing active trends, each might contribute to a global increase in morphological diversity at a higher level of their phylogeny (Hopkins, 2016), as is the case here in which the relatively low disparity of each pipid subclade contributes to a significantly higher disparity for extant pipids as a whole, relative to that of fossils (Figures 4A,B). Noteworthy, under most estimations of disparity (Ciampaglio et al., 2001), differences are expected to be even more overestimated if subclades show opposite, instead of parallel, active trends. This way, our data is in line with the view that recognition of different disparity patterns over the history of clades might be scale-dependent (Slater, 2015) and heavily influenced by the presence of active evolutionary trends (Hopkins, 2016).
In contrast to the continuous morphological innovation that the sacro-caudo-pelvic complex of pipimorphs as a whole experienced in the Cretaceous, an arrested morphological diversification of Xe, Hymenochirini, and Pipa is apparent during the Cenozoic, also reflected in their stagnated disparity and decoupled, growing phylogenetic diversity toward the Recent, and in the increased levels of homoplasy within and between the different crown group pipid subclades. This is intriguing, since the aftermath of the Cretaceous–Paleogene (K–Pg) mass extinction is recognized as a time of biotic turnover and ecological release and, as a result, it might be expected to be a rapid, relatively unrestricted diversification of lineages (Simpson, 1944; Foote, 1997; Erwin, 2007; Ruta et al., 2013). The slight trend, yet non-significant in most cases, of increased disparity across the K–Pg depicted by SOV measures that is only later followed by a rise in taxonomic diversity (Figures 4D,E) may suggest, though, that pipids conform to a ‘disparity-first’ pattern observed in many other groups after a mass extinction event (Benton, 2015; Minter et al., 2017). It has been proposed that this pattern might be the rule only in cases in which an ‘empty ecospace’ favors specialists over generalists (Grossnickle and Newham, 2016), which may be the case for pipids since they were likely among the most specialized frogs at that time. It is noteworthy, however, that early diversification of crown group Pipidae was well underway by the end-Cretaceous, which might have been triggered by the complete opening of the South Atlantic Ocean around 100 Ma (Bewick et al., 2012; Pyron, 2014; Gómez, 2016), but it was not until the Eocene onward that crown xenopodines, hymenochirines, and Pipa began to diversify (Pyron, 2014; Cannatella, 2015; Evans et al., 2015; this paper). This might reflect other potential causes for the observed disparity pattern than an ecological release in the aftermath of the K–Pg mass extinction event. In this regard, the oldest post K–Pg record of pipids is represented by ‘Xenopus’ romeri from Itaboraí (Estes, 1975). The age of the Itaboraian South American Land Mammal Age (SALMA) has long been debated (e.g., Marshall et al., 1997; Oliveira and Goin, 2011) and recent redimetric dates indicate that the Itaboraian SALMA in Patagonia might be restricted to the early Eocene (Krause et al., 2017). This way, pipids are still unknown from the early Paleocene, or even the entire Paleocene, and the bulk of disparity in the Paleogene resulted from fossils and reconstructed ancestors of late Paleocene–Eocene age. A candidate cause for such patterns might be the Paleocene–Eocene Thermal Maximum, a relatively brief global-warming event occurring around 55.8 Ma, which had lead to increased diversification in most marine and terrestrial ecosystems (Wilf et al., 2003; Keller et al., 2018). Increased sampling of fossil pipids along with a better temporal discrimination of this record will be necessary to further explore this hypothesis.
Hence, t appears that the patterns recovered here for pipids and their kin cannot be simply explained in the context of an ‘empty ecospace’ model. Stabilizing selection has previously been advanced for explaining stasis on anuran larval morphology after an initial adaptive radiation (Roelants et al., 2011), which might hold true for the respective crowns of Xe, Hymenochirini, and Pipa, although the active trends and disparity/diversity patterns exhibited by pipimorphs and early pipids neither reflect stasis nor the signature of an adaptive radiation. However, as aforementioned, the pattern of continuous innovation that proceed in parallel in South America and Africa might indeed indicate evolution in analogous Simpsonian landscapes (Simpson, 1944) with ‘adaptive optima’ that remains stable over deep time (Mahler et al., 2013; Moen et al., 2015). The continuous evolutionary history of pipimorphs in similar freshwater environments supports this scenario. On the other hand, the existence of long-term active trends and constant disparity over time, as well as decoupled taxonomic diversity and an arrested evolution of novel morphologies toward the Recent, strongly suggest an increasingly constrained morphological diversification in pipimorphs that might be better explained in the context of a ‘genetic/developmental’ hypothesis (Ciampaglio, 2002; Erwin, 2007; Hughes et al., 2013; Minter et al., 2017). Shift in evolving characters and patterns of homoplasy point in the same direction as they indicate a phylogenetic signal in evolvability, which becomes progressively more restricted and produces increasing clumpiness in morphospace occupation (Roelants et al., 2011), a pattern which in turn might indicate highly conserved structuring of developmental features (Erwin, 2007).
Overall, the evolution of the entire pipimorph clade might be better explained by two concerted evolutionary processes: Continuous and parallel adaptive evolution toward stable ‘optima,’ channeled by increasing genetic/developmental constraints in deep time. This meets the expectation that genetic/developmental constraints channel phenotypic evolution along ‘genetic lines of least resistance’ (Schluter, 1996) and is further supported by recent studies in the Caribbean Anolis lizards, demonstrating that adaptive peaks and genetic constraints may be highly conserved over a macroevolutionary scale (Mahler et al., 2013; McGlothlin et al., 2018). In addition, the herein recovered pattern of pipids and their sister group, the fossorial Rhinophrynus, at the antipodes of anuran morphospace, contrasts with the unique and relatively conserved morphology of the xenoanuran larvae (Orton, 1957; Cannatella, 1999; Roelants et al., 2011). Given that pipids and Rhinophrynus share the same type of larva, but as adults differ sharply in habits, it is likely that different ecological constraints have subsequently shaped their adult morphology. Most notably, aside from Rhinophrynus and microhylids, our overall pattern of morphospace occupation strikingly echoes that of larval morphology (Roelants et al., 2011), suggesting that long-term developmental constraints might act upon adult morphology of a well-integrated sacro-caudo-pelvic complex, not only in pipimorphs, but in most anuran clades recalling the ‘default-frog’ hypothesis of Altig (2006). Further integration of neontological and paleontological data, including that from ontogeny of extant and fossil taxa (e.g., Roček and Van Dijk, 2006), as well as estimations of evolutionary rates, are warranted in order to explore this issue.
Data Availability
The datasets analyzed for this study can be found in the Supplementary Material.
Author Contributions
RG conceived the study and compiled the data. CP-B performed the statistical analyses. RG and CP-B wrote and approved the submitted version of the manuscript.
Funding
Research was funded by grants from Agencia Nacional de Promoción Científica y Tecnológica (PICT2017-1665) to RG and from Consejo Nacional de Investigaciones Cientificas y Técnicas (PIP-11220150100423CO ) to Ignacio Soto and RG.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank A. M. Báez (University of Buenos Aires/Museo Argentino de Ciencias Naturales), D. Cabaza (Museo Municipal de Lamarque), M. Fabrezi (Museo de Ciencias Naturales, Universidad Nacional de Salta), J. Williams (Museo de La Plata, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata), J. Faivovich (Museo Argentino de Ciencias Naturales), G. Schneider (Museum of Zoology, University of Michigan), J. Losos and J. Martinez (Museum of Comparative Zoology, Harvard University), R. Brown and R. Glor (Natural History Museum, University of Kansas), and S. Bogan (Fundación de Historia Natural ‘Félix de Azara’, Universidad Maimónides) for providing access to and/or photographs of specimens under their care. Thanks are extended to David Blackburn (University of Florida) and his team, whose CT scanning project MorphoSource provides access to invaluable skeletal data. We would also like to thank A. M. Báez, A. I. Lires, and G. F. Turazzini (University of Buenos Aires) for fruitful discussions as well as the valuable comments and suggestions of the Editor ML (Muséum National d’Histoire Naturelle, Paris) and the reviewers who certainly improved the manuscript. They also thank Facultad de Ciencias Exactas y Naturales-University of Buenos Aires (FCEN-UBA) and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) of Argentina for continuous support. RG is teaching assistant and researcher at FCEN-UBA and member of Carrera de Investigador Científico (CONICET), CP-B is postdoctoral fellow of CONICET.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/feart.2019.00056/full#supplementary-material
References
Altig, R. (2006). Tadpoles evolved and frogs are the default. Herpetologica 62, 1–10. doi: 10.1655/05-23.1
Anderson, M. J., and Walsh, D. C. (2013). PERMANOVA, ANOSIM, and the Mantel test in the face of heterogeneous dispersions: what null hypothesis are you testing? Ecol. Monogr. 83, 557–574. doi: 10.1890/12-2010.1
Archie, J. W. (1989). A randomization test for phylogenetic information in systematic data. Syst. Zool. 38, 239–252. doi: 10.2307/2992285
Arthur, W. (2004). The effect of development on the direction of evolution: toward a twenty-first century consensus. Evol. Dev. 6, 282–288. doi: 10.1111/j.1525-142X.2004.04033.x
Báez, A. M. (2013). Anurans from the Early Cretaceous Lagerstätte of Las Hoyas, Spain: new evidence on the Mesozoic diversification of crown-clade Anura. Cretac. Res. 41, 90–106. doi: 10.1016/j.cretres.2012.11.002
Báez, A. M., and Gómez, R. O. (2018). Dealing with homoplasy: osteology and phylogenetic relationships of the bizarre neobatrachian frog Baurubatrachus pricei from the Upper Cretaceous of Brazil. J. Syst. Palaeontol. 16, 279–308. doi: 10.1080/14772019.2017.1287130
Báez, A. M., Gómez, R. O., and Taglioretti, M. L. (2012). The archaic ilial morphology of an enigmatic pipid frog from the upper Pleistocene of the South American pampas. J. Vertebr. Paleontol. 32, 304–314. doi: 10.1080/02724634.2012.637591
Báez, A. M., Scanferla, C. A., Agnolin, F. L., Cenizo, M., and De Los Reyes, M. (2008). Pipid frog from the Pleistocene of the Pampas of southern South America. J. Vertebr. Paleontol. 28, 1195–1198. doi: 10.1671/0272-4634-28.4.1195
Bapst, D. W., Bullock, P. C., Melchin, M. J., Sheets, H. D., and Mitchell, C. E. (2012). Graptoloid diversity and disparity became decoupled during the Ordovician mass extinction. Proc. Natl. Acad. Sci. U.S.A. 109, 3428–3433. doi: 10.1073/pnas.1113870109
Benton, M. J. (2015). Exploring macroevolution using modern and fossil data. Proc. R. Soc. B Biol. Sci. 282:20150569. doi: 10.1098/rspb.2015.0569
Bewick, A. J., Chain, F. J., Heled, J., and Evans, B. J. (2012). The pipid root. Syst. Biol. 61, 913–926. doi: 10.1093/sysbio/sys039
Blackburn, D. C., Bickford, D. P., Diesmos, A. C., Iskandar, D. T., and Brown, R. M. (2010). An ancient origin for the enigmatic flat-headed frogs (Bombinatoridae: Barbourula) from the islands of Southeast Asia. PLoS One 5:e12090. doi: 10.1371/journal.pone.0012090
Brusatte, S. L., Benton, M. J., Ruta, M., and Lloyd, G. T. (2008). Superiority, competition, and opportunism in the evolutionary radiation of dinosaurs. Science 321, 1485–1488. doi: 10.1126/science.1161833
Brusatte, S. L., Lloyd, G. T., Wang, S. C., and Norell, M. A. (2014). Gradual assembly of avian body plan culminated in rapid rates of evolution across dinosaur-bird transition. Curr. Biol. 24, 2386–2392. doi: 10.1016/j.cub.2014.08.034
Butler, R. J., Brusatte, S. L., Andres, B., and Benson, R. B. J. (2012). How do geological sampling biases affect studies of morphological evolution in deep time? A case study of Pterosaur (Reptilia: Archosauria) disparity. Evolution 66, 147–162. doi: 10.1111/j.1558-5646.2011.01415.x
Cannatella, D. C. (1999). “Architecture: cranial and axial musculoskeleton,” in Tadpoles, the Biology of Anuran Larvae, eds R. W. McDiarmid and R. Altig (Chicago, IL: University of Chicago Press), 52–91.
Cannatella, D. C. (2015). Xenopus in space and time: fossils, node calibrations, tipedating, and paleobiogeography. Cytogenet. Genome Res. 145, 283–301. doi: 10.1159/000438910
Cannatella, D. C., and Trueb, L. (1988). Evolution of pipoid frogs: intergeneric relationships of the aquatic frog family Pipidae (Anura). Zool. J. Linn. Soc. Lond. 94, 1–38. doi: 10.1111/j.1096-3642.1988.tb00880.x
Ciampaglio, C. N. (2002). Determining the role that ecological and developmental constraints play in controlling disparity: examples from the crinoid and blastozoan fossil record. Evol. Dev. 4, 170–188. doi: 10.1046/j.1525-142X.2002.02001.x
Ciampaglio, C. N., Kemp, M., and McShea, D. W. (2001). Detecting changes in morphospace occupation patterns in the fossil record: characterization and analysis of measures of disparity. Paleobiology 27, 695–715. doi: 10.1666/0094-8373 (2001)027<0695:DCIMOP>2.0.CO;2
Cohen, K. M., Finney, S. C., Gibbard, P. L., and Fan, J.-X. (2013). The ICS international chronostratigraphic chart. Episodes 36, 199–204.
Colombo, M., Damerau, M., Hanel, R., Salzburger, W., and Matschiner, M. (2015). Diversity and disparity through time in the adaptive radiation of Antarctic notothenioid fishes. J. Evol. Biol. 28, 376–394. doi: 10.1111/jeb.12570
Donoghue, M. J., Doyle, J. J., Gauthier, J., Kluge, A. G., and Rowe, T. (1989). The importance of fossils in phylogeny reconstruction. Annu. Rev. Ecol. Syst. 20, 431–460. doi: 10.1146/annurev.es.20.110189.002243
Dumont, E. R., Dávalos, L. M., Goldberg, A., Santana, S. E., Rex, K., and Voigt, C. C. (2012). Morphological innovation, diversification and invasion of a new adaptive zone. Proc. R. Soc. B 279, 1797–1805. doi: 10.1098/rspb.2011.2005
Emerson, S. B. (1979). The ilio-sacral articulation in frogs: form and function. Biol. J. Linn. Soc. Lond. 11, 153–168. doi: 10.1111/j.1095-8312.1979.tb00032.x
Emerson, S. B. (1982). Frog postcranial morphology: identification of a functional complex. Copeia 1982, 603–613. doi: 10.2307/1444660
Erwin, D. H. (2007). Disparity: morphological pattern and developmental context. Palaeontology 50, 57–73. doi: 10.1111/j.1475-4983.2006.00614.x
Estes, R. (1975). Fossil Xenopus from the Paleocene of South America and the zoogeography of pipid frogs. Herpetologica 31, 263–278.
Evans, B. J., Carter, T. F., Greenbaum, E., Gvozdík, V., Kelley, D. B., McLaughlin, P. J., et al. (2015). Genetics, morphology, advertisement calls, and historical records distinguish six new polyploid species of African clawed frog (Xenopus, Pipidae) from West and Central Africa. PLoS One 10:e0142823. doi: 10.1371/journal.pone.0142823
Foote, M. (1992). Rarefaction analysis of morphological and taxonomic diversity. Paleobiology 18, 1–16. doi: 10.1017/S0094837300012185
Foote, M. (1993). Discordance and concordance between morphological and taxonomic diversity. Paleobiology 19, 185–204. doi: 10.1017/S0094837300015864
Foote, M. (1997). The evolution of morphological diversity. Annu. Rev. Ecol. Syst. 28, 129–152. doi: 10.1146/annurev.ecolsys.28.1.129
Ford, L., and Cannatella, D. C. (1993). The major clades of frogs. Herpetol. Monogr. 7, 94–117. doi: 10.2307/1466954
Frost, D. R. (2018). Amphibian Species of the World: an Online Reference. Version 6.0 Electronic Database. New York, NY: American Museum of Natural History.
Gardner, J. D., Roček, Z., Přikryl, T., Eaton, J. G., Blob, R. W., and Sankey, J. T. (2010). Comparative morphology of the ilium of anurans and urodeles (Lissamphibia) and a re-assessment of the anuran affinities of Nezpercius dodsoni Blob et al., 2001. J. Vertebr. Paleontol. 30, 1684–1696. doi: 10.1080/02724634.2010.521605
Gerber, S. (2013). On the relationship between the macroevolutionary trajectories of morphological integration and morphological disparity. PLoS One 8:e63913. doi: 10.1371/journal.pone.0063913
Gómez, R. O. (2016). A new pipid frog from the Upper Cretaceous of Patagonia and early evolution of crown–group Pipidae. Cretac. Res. 62, 52–64. doi: 10.1016/j.cretres.2016.02.006
Gómez, R. O., and Turazzini, G. F. (2016). An overview of the ilium of anurans (Lissamphibia, Salientia), with a critical appraisal of the terminology and primary homology of main ilial features. J. Vertebr. Paleontol. 36:e1030023. doi: 10.1080/02724634.2015.1030023
Griffiths, I. (1963). The phylogeny of the Salientia. Biol. Rev. 38, 241–292. doi: 10.1111/j.1469-185X.1963.tb00784.x
Grossnickle, D. M., and Newham, E. (2016). Therian mammals experience an ecomorphological radiation during the Late Cretaceous and selective extinction at the K–Pg boundary. Proc. R. Soc. B Biol. Sci. 283:20160256. doi: 10.1098/rspb.2016.0256
Guillerme, T. (2018). dispaRity: a modular R package for measuring disparity. Methods Ecol. Evol. 9, 1755–1763. doi: 10.1111/2041-210X.13022
Guillerme, T., and Cooper, N. (2018). Time for a rethink: time sub-sampling methods in disparity-through-time analyses. Palaeontology 61, 481–493. doi: 10.1111/pala.12364
Halliday, T. J. D., and Goswami, A. (2016). Eutherian morphological disparity across the end-Cretaceous mass extinction. Biol. J. Linn. Soc. Lond. 118, 152–168. doi: 10.1111/bij.12731
Handrigan, G. R., and Wassersug, R. J. (2007). The anuran Bauplan: a review of the adaptive, developmental, and genetic underpinnings of frog and tadpole morphology. Biol. Rev. 82, 1–25. doi: 10.1111/j.1469-185X.2006.00001.x
Harmon, L. J., Losos, J. B., Jonathan Davies, T., Gillespie, R. G., Gittleman, J. L., Bryan Jennings, W., et al. (2010). Early bursts of body size and shape evolution are rare in comparative data. Evolution 64, 2385–2396. doi: 10.1111/j.1558-5646.2010.01025.x
Hopkins, M. J. (2016). Magnitude versus direction of change and the contribution of macroevolutionary trends to morphological disparity. Biol. J. Linn. Soc. Lond. 118, 116–130. doi: 10.1111/bij.12759
Hughes, M., Gerber, S., and Wills, M. A. (2013). Clades reach highest morphological disparity early in their evolution. Proc. Natl. Acad. Sci. U.S.A. 110, 13875–13879. doi: 10.1073/pnas.1302642110
Jenkins, F., and Shubin, N. (1998). Prosalirus bitis and the anuran caudopelvic mechanism. J. Vertebr. Paleontol. 18, 495–510. doi: 10.1080/02724634.1998.10011077
Jorgensen, M. E., and Reilly, S. M. (2013). Phylogenetic patterns of skeletal morphometrics and pelvic traits in relation to locomotor mode in frogs. J. Evol. Biol. 26, 929–943. doi: 10.1111/jeb.12128
Keller, G., Mateo, P., Punekar, J., Khozyem, H., Gertsch, B., Spangenberg, J., et al. (2018). Environmental changes during the Cretaceous-Paleogene mass extinction and Paleocene-Eocene Thermal Maximum: implications for the Anthropocene. Gondwana Res. 56, 69–89. doi: 10.1016/j.gr.2017.12.002
Kluge, A. G., and Farris, J. S. (1969). Quantitative phyletics and the evolution of anurans. Syst. Zool. 18, 1–32. doi: 10.2307/2412407
Krause, J. M., Clyde, W. C., Ibañez-Mejía, M., Schmitz, M. D., Barnum, T., Bellosi, E. S., et al. (2017). New age constraints for early Paleogene strata of central Patagonia, Argentina: implications for the timing of South American Land Mammal Ages. Geol. Soc. Am. Bull. 129, 886–903. doi: 10.1130/B31561.1
Lires, A. I., Soto, I. M., and Gómez, R. O. (2016). Walk before you jump: new insights on frog locomotion from the oldest known salientian. Palaeobiology 42, 612–623. doi: 10.1017/pab.2016.11
Lloyd, G. T. (2016). Estimating morphological diversity and tempo with discrete character-taxon matrices: implementation, challenges, progress, and future directions. Biol. J. Linn. Soc. Lond. 118, 131–151. doi: 10.1111/bij.12746
Lloyd, G. T., Wang, S. C., and Brusatte, S. L. (2012). Identifying heterogeneity in rates of morphological evolution: discrete character change in the evolution of lungfish (Sarcopterygii; Dipnoi). Evolution 66, 330–348. doi: 10.1111/j.1558-5646.2011.01460.x
Lupia, R. (1999). Discordant morphological disparity and taxonomic diversity during the Cretaceous angiosperm radiation: North American pollen record. Paleobiology 25, 1–28.
Mahler, D. L., Ingram, T., Revell, L. J., and Losos, J. B. (2013). Exceptional convergence on the macroevolutionary landscape in island lizard radiations. Science 341, 292–295. doi: 10.1126/science.1232392
Marjanović, D., and Laurin, M. (2008). Assessing confidence intervals for stratigraphic ranges of higher taxa: the case of Lissamphibia. Acta Palaeontol. Pol. 53, 413–432. doi: 10.4202/app.2008.0305
Marjanović, D., and Laurin, M. (2014). An updated paleontological timetree of lissamphibians, with comments on the anatomy of Jurassic crown-group salamanders (Urodela). Hist. Biol. 26, 535–550. doi: 10.1080/08912963.2013.797972
Marshall, L. G., Sempere, T., and Butler, R. F. (1997). Chronostratigraphy of the mammal-bearing Paleocene of South America. J. S. Am. Earth Sci. 10, 49–70. doi: 10.1016/S0895-9811(97)00005-9
McGlothlin, J. W., Kobiela, M. E., Wright, H. V., Mahler, D. L., Kolbe, J. J., Losos, J. B., et al. (2018). Adaptive radiation along a deeply conserved genetic line of least resistance in Anolis lizards. Evol. Lett. 2, 310–322. doi: 10.1002/evl3.72
McShea, D. W. (1994). Mechanisms of large-scale evolutionary trends. Evolution 48, 1747–1763. doi: 10.1111/j.1558-5646.1994.tb02211.x
Minter, N. J., Buatois, L. A., Mángano, M. G., Davies, N. S., Gibling, M. R., MacNaughton, R. B., et al. (2017). Early bursts of diversification defined the faunal colonization of land. Nat. Ecol. Evol. 1:0175. doi: 10.1038/s41559-017-0175
Moen, D. S., Morlon, H., and Wiens, J. J. (2015). Testing convergence versus history: convergence dominates phenotypic evolution for over 150 million years in frogs. Syst. Biol. 65, 146–160. doi: 10.1093/sysbio/syv073
Naitoh, T., Wassersug, R. J., and Leslie, R. A. (1996). The physiology, morphology, and ontogeny of emetic behavior in anuran amphibians. Physiol. Zool. 62, 819–843. doi: 10.1086/physzool.62.3.30157929
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’hara, R. B., et al. (2016). vegan: Community Ecology Package. R package version 2.0–7. 2013. Available at: http://CRANR-project.org/package=vegan
Oliveira, E. V., and Goin, F. J. (2011). A reassessment of bunodont metatherians from the Paleogene of Itaboraí (Brazil): systematics and age of the Itaboraian SALMA. Rev. Bras. Paleontol. 14, 105–136. doi: 10.4072/rbp.2011.2.01
Orton, G. I. (1957). The bearing of larval evolution on some problems in frog classification. Syst. Zool. 6, 79–86. doi: 10.2307/2411805
Palmer, M. (1960). Expanded ilio-sacral joint in the toad Xenopus laevis. Nature 187, 797–798. doi: 10.1038/187797a0
Přikryl, T., Aerts, P., Havelková, P., Herrel, A., and Roček, Z. (2009). Pelvic and thigh musculature in frogs (Anura) and origin of anuran jumping locomotion. J. Anat. 214, 100–139. doi: 10.1111/j.1469-7580.2008.01006.x
Pyron, R. A. (2014). Biogeographic analysis reveals ancient continental vicariance and recent oceanic dispersal in amphibians. Syst. Biol. 63, 779–797. doi: 10.1093/sysbio/syu042
Pyron, R. A. (2015). Post-molecular systematics and the future of phylogenetics. Trends Ecol. Evol. 30, 384–389. doi: 10.1016/j.tree.2015.04.016
R Core Team (2016). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Reilly, S. M., and Jorgensen, M. E. (2011). The evolution of jumping in frogs: morphological evidence for the basal anuran locomotor condition and the radiation of locomotor systems in crown group anurans. J. Morphol. 272, 149–168. doi: 10.1002/jmor.10902
Robovska-Havelková, P., Aerts, P., Roček, Z., Přikryl, T., Fabre, A.-C., and Herrel, A. (2014). Do all frogs swim alike? The effect of ecological specialization on swimming kinematics in frogs. J. Exp. Biol. 217, 3637–3644. doi: 10.1242/jeb.109991
Roček, Z., and Van Dijk, E. (2006). Patterns of larval development in Cretaceous pipid frogs. Acta Palaeontol. Pol. 51, 111–126.
Roelants, K., Gower, D. J., Wilkinson, M., Loader, S. P., Biju, S. D., Guillaume, K., et al. (2007). Global patterns of diversification in the history of modern amphibians. Proc. Natl. Acad. Sci. U.S.A. 104, 887–892. doi: 10.1073/pnas.0608378104
Roelants, K., Haas, A., and Bossuyt, F. (2011). Anuran radiations and the evolution of tadpole morphospace. Proc. Natl. Acad. Sci. U.S.A. 108, 8731–8736. doi: 10.1073/pnas.1100633108
Ruta, M., Botha-Brink, J., Mitchell, S. A., and Benton, M. J. (2013). The radiation of cynodonts and the ground plan of mammalian morphological diversity. Proc. R. Soc. B Biol. Sci. 280:20131865. doi: 10.1098/rspb.2013.1865
Ruta, M., Wagner, P. J., and Coates, M. I. (2006). Evolutionary patterns in early tetrapods. I. Rapid initial diversification followed by decrease in rates of character change. Proc. R. Soc. B Biol. Sci. 273, 2107–2111. doi: 10.1098/rspb.2006.3577
Schluter, D. (1996). Adaptive radiation along genetic lines of least resistance. Evolution 50, 1766–1774. doi: 10.1111/j.1558-5646.1996.tb03563.x
Sidlauskas, B. (2008). Continuous and arrested morphological diversification in sister clades of characiform fishes: a phylomorphospace approach. Evolution 62, 3135–3156. doi: 10.1111/j.1558-5646.2008.00519.x
Slater, G. J. (2015). Not-so-early bursts and the dynamic nature of morphological diversification. Proc. Natl. Acad. Sci. U.S.A. 112, 3595–3596. doi: 10.1073/pnas.1502204112
Slater, G. J., and Harmon, L. J. (2013). Unifying fossils and phylogenies for comparative analyses of diversification and trait evolution. Methods Ecol. Evol. 4, 699–702. doi: 10.1111/2041-210X.12091
Smith, N. D., and Turner, A. H. (2005). Morphology’s role in phylogeny reconstruction: perspectives from paleontology. Syst. Biol. 54, 166–173. doi: 10.1080/10635150590906000
Trueb, L. (1996). “Historical constraints and morphological novelties in the evolution of the skeletal system of pipid frogs (Anura: Pipidae),” in The Biology of Xenopus. Symposia of the Zoological Society of London 68, eds R. C. Tinsley and H. R. Kobel (Oxford: Clarendon Press), 350–377.
Van Dijk, D. E. (2002). Longitudinal sliding articulations in pipid frogs: research in action. S. Afr. J. Sci. 98, 555–556.
Videler, J. J., and Jorna, J. T. (1985). Functions of the sliding pelvis in Xenopus laevis. Copeia 1985, 251–254. doi: 10.2307/1444825
Wagner, P. J. (1996). Contrasting the underlying patterns of active trends in morphologic evolution. Evolution 50, 990–1007. doi: 10.1111/j.1558-5646.1996.tb02341.x
Wagner, P. J. (1997). Patterns of morphologic diversification among the Rostroconchia. Paleobiology 23, 115–150. doi: 10.1017/S0094837300016675
Wagner, P. J., Ruta, M., and Coates, M. I. (2006). Evolutionary patterns in early tetrapods. II. Differing constraints on available character space among clades. Proc. R. Soc. B Biol. Sci. 273, 2113–2118. doi: 10.1098/rspb.2006.3561
Wilf, P., Cúneo, N. R., Johnson, K. J., Hicks, J. F., Wing, S. L., and Obradovich, J. D. (2003). High plant diversity in Eocene South America: evidence from Patagonia. Science 5616, 122–125. doi: 10.1126/science.1080475
Keywords: anura, Pipidae, sacrourostyle, ilium, phylomorphospace, disparity, homoplasy, constraints
Citation: Gómez RO and Pérez-Ben CM (2019) Fossils Reveal Long-Term Continuous and Parallel Innovation in the Sacro-Caudo-Pelvic Complex of the Highly Aquatic Pipid Frogs. Front. Earth Sci. 7:56. doi: 10.3389/feart.2019.00056
Received: 07 November 2018; Accepted: 07 March 2019;
Published: 22 March 2019.
Edited by:
Michel Laurin, UMR7207 Centre de Recherche sur la Paléobiodiversité et les Paléoenvironnements (CR2P), FranceReviewed by:
Marcello Ruta, University of Lincoln, United KingdomSylvain Gerber, Muséum National d’Histoire Naturelle (France), France
Copyright © 2019 Gómez and Pérez-Ben. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Raúl O. Gómez, cmF1bG9yZW5jaW9nb21lekBnbWFpbC5jb20=