 Domingo C. Salazar-García
Domingo C. Salazar-García Robert C. Power3
Robert C. Power3 Joan Daura
Joan Daura- 1Departament de Prehistòria, Arqueologia i Història Antiga, Universitat de València, València, Spain
- 2Department of Geological Sciences, University of Cape Town, Cape Town, South Africa
- 3School of Archaeology, University College Dublin, Dublin, Ireland
- 4Grup de Recerca del Quaternari (GRQ)-SERP, Departament d'Història i Arqueologia, Universitat de Barcelona, Barcelona, Spain
The emergence of Neolithic societies was transformative, impacting many aspects of life, particularly diet. The process of Neolithization in Iberia is increasingly understood as the arrival of new people from the Central Mediterranean, who dispersed along the Iberian coasts introducing cereal production, herding, and Cardial pottery and associated material culture. Although research has clarified aspects of the cultigen-dominated economy of these new people, questions remain due to the limitations of conventional archaeobotanical and archaeozoological methods that tend to produce indirect evidence. The extent to which these early farmers adopted Mesolithic staples, which are often difficult to detect with other methods, remains unclear. Furthermore, questions surround the nature of methods of food preparation Cardial Neolithic people used when incorporating grains into their diet. In this study, we examined direct evidence of the diet from the Iberian Cardial Neolithic site of Cova Bonica (Vallirana, Baix Llobregat, Catalonia) using CN stable isotopes on bone and plant microremains trapped in dental calculus from six human individuals and associated fauna. Isotopes show a diet based on terrestrial C3 resources, with no isotopic evidence of aquatic or C4 resource consumption. Plant microremains (starches and phytoliths) provide evidence of cereal use, as well as of other plant foods. However, perhaps due to Bonica’s early farmers’ choice of grain variety, their grain processing methods, or due to specific dental calculus formation factors, the grain assemblages are rather limited and provide scarce information on food preparation.
1 Introduction
1.1 Combining stable isotope and plant microremain analysis for dietary studies
The study of stable isotopes on both bone collagen and plant microremains in dental calculus is useful for recovering information on past diets and has been used individually as a direct means to trace critical aspects of prehistoric populations such as dietary patterns. While carbon and nitrogen stable isotope analysis is a well-established and frequently used method to obtain information on average protein consumption (Lee-Thorp, 2008), the study of plant microremains on dental calculus to recover information on plant food consumption is still not as frequently used as the latter (Power et al., 2015); the combination of both proxies is even scarcer. However, by combining these two techniques, there is a potential for achieving a wider picture of dietary subsistence patterns during prehistoric times (Goude et al., 2019; Goude et al., 2020; Salazar-García et al., 2021).
Isotopic diet reconstruction is based on the principle that as animal and human tissue grows, it records the isotopic composition of the diet through predictable isotope fractionation (Schoeller, 1999). For this analysis, bone collagen is usually the preferred substrate because it is both the only considerable nitrogen source from skeletal remains (Salazar-García et al., 2014a) and has accepted quality indicators to assess its isotopic integrity (De Niro, 1985; Van Klinken, 1999). However, it is necessary to consider when interpreting results that stable isotope ratios from bone collagen reflect only the main dietary protein sources consumed several years prior to death (Hedges et al., 2007; Katzenberg, 2012) rather than that of a diet as a whole, especially for nitrogen (Ambrose and Norr, 1993) since carbon may be derived from other dietary macronutrients like sugars and fats (Howland et al., 2003; Jim et al., 2004).
On the other hand, dental calculus is an oral plaque mineralized by salivary calcium. During the accumulation process, dietary and environmental information from plant and animal foods, including starch grains, phytoliths, lipids, proteins, and DNA, can become trapped and embedded. Dietary remains retrieved from dental calculus are a useful additional source of information on human diet and behavior. Amongst dietary reconstruction methods, it is a helpful complement to other methods because it offers direct information on food consumed and substances that enter the mouth (Armitage 1975; Henry and Piperno 2008; Power et al., 2014; Warinner et al., 2014; Leonard et al., 2015; Power et al., 2015). In this sense, while isotope analysis gives a more quantitative approach on the origin of consumed protein, plant microremains on dental calculus give more qualitative data on specific foods consumed. Likewise, while stable isotope analysis gives long-term averaged dietary information on protein consumption (Lee-Thorp, 2008), plant microremains can give information on other types of consumed non-protein food resources, even if less frequently eaten (Leonard et al., 2015; Power et al., 2015). Therefore, combining these two techniques provides complementary information on past diets and should be implemented in dietary studies when possible (Salazar-García et al., 2021).
The application of these analytical techniques in Mediterranean Iberia is, however, unequal between them. While studies of stable isotopes have significantly increased during the past decade in prehistoric periods (Salazar-García et al., 2018) creating an important corpus of prehistoric dietary data from the Mesolithic onward in the region and its hinterland, plant microremain studies on dental calculus are almost non-existent. Isotope studies from the Mesolithic (García-Guixé et al., 2006; Fernández-López de Pablo et al., 2013; Salazar-García et al., 2014b), Neolithic–Chalcolithic (Fontanals-Coll et al., 2015, Fontanals-Coll et al., 2017; García-Borja et al., 2013; Gibaja et al., 2016; McClure et al., 2011; Salazar-García, 2009, Salazar-García, 2011, Salazar-García, 2014; Salazar-García et al., 2013, Salazar-García et al., 2016, Salazar-García et al., 2017; Villalba-Mouco et al., 2018a; Villalba-Mouco et al., 2018b; Villalba-Mouco et al., 2019), Bronze Age (McClure et al., 2011; García-Borja et al., 2013; Salazar-García et al., 2017), and Iron Age (Salazar-García et al., 2010) periods have been performed in Mediterranean Iberia. On the other hand, plant microremain studies on dental calculus have only been performed so far on one Paleolithic site (Salazar-García et al., 2013) and one Chalcolithic site (Power et al., 2014) in prehistoric Mediterranean Iberia.
1.2 The site studied: Cova Bonica
Cova Bonica (41°22′10.29″N, 1°53′38.64″E) is located in the vicinity of Vallirana, near Barcelona (northeast of the Iberian Peninsula). The site was discovered and first excavated in 1936; however, the archaeological artifacts were not published until the 70s (Baldellou, 1974). Current archaeological excavations are conducted by Grup de Recerca del Quaternari (Universitat de Barcelona) and produced genetic (Olalde et al., 2015) and archaeological data (Oms et al., 2017; Daura et al., 2019). Amongst the archaeological data published by Daura et al., 2019, macrobotanical evidence of diet from flotation consists only of the remains of Arbutus unedo charcoal, which produces edible fruits, and some taxa charcoal with aromatic and medicinal properties such as Laurus nobilis or Rosmarinus officinalis.
The cave consists of a principal chamber (SP) and two lateral chambers, one next to the entrance (SL1) and another in the inner area (SL2). The cave and sediments were partially destroyed by a sparry calcite mine, but intact Cardial infillings have been preserved in two separated layers: IV2 preserved in SP-Grao 1 and IV in SP (Figure 1).

FIGURE 1. Cova Bonica. (A) Location of Cova Bonica in the NE of the Iberian Peninsula. (B) Current view of the cave entrance and main areas. (C) Neolithic area excavated. (D) Plan of the site indicating the excavated area. (E) Stratigraphy of the site’s profiles (cave -a to d-, terrace -e-, and SL2 -f-).
The stratigraphic succession of Cova Bonica is complex and can be grouped into five episodes (1–5). Episodes 5 and 4 represent endokarstic deposits and a ceiling collapse with a restricted entrance and scarce archaeological materials. Episode 3 corresponds to the Cardial Neolithic horizon and is formed by layers IV and IV2. Layer IV is located in SL1, laying above the stalagmitic flowstone of layer V and contains a thermoaltered sediment labeled as EC1 and 14 post-holes, one of them yielding a radiocarbon age of 6,340 ± 34 BP. Layer IV contains scarce archaeological artifacts and human remains probably displaced from layer IV2; a sheep bone shows a14C age of 6,158 ± 32 BP, and one human individual shows an age of 6,395 ± 39 BP. Layer IV2 lies in a depression in the cave’s bedrock and between large collapsed speleothems and contains a minimum number of individuals (MNI) of 7 (ca. 315 bones): 6 individuals defined osteologically, and a seventh one defined by aDNA. The archaeological remains also include personal ornaments, lithics, bone tools, Cardial Neolithic pottery, faunal remains, and charcoal. Radiocarbon dating of the human remains recovered from layers IV and IV2 indicates a homogeneous assemblage and a short or synchronic time span for the deposition of the human corpses around ca. 6400 BP (5,470–5,320 ka cal. BP). Episode 2, recorded in several areas of the cave, represents archaeological occupation spanning from the late Neolithic to the Bronze Age.
2 Materials and methods
For this study, we analyzed for stable isotopes 12 faunal and 6 human remains and assessed 5 samples of human dental calculus, one sediment and two associated blank controls. Human and faunal remains analyzed in the present work were recovered during the 2008–2017 field seasons conducted by Grup de Recerca del Quaternari (University of Barcelona). Materials were previously identified and studied in the Guixera laboratory (Castelldefels City Council) (Figures 2, 3). Human remains were scattered and often it was not possible to associate elements (e.g., long bones, ribs, teeth) as pertaining to the same individual with certainty.
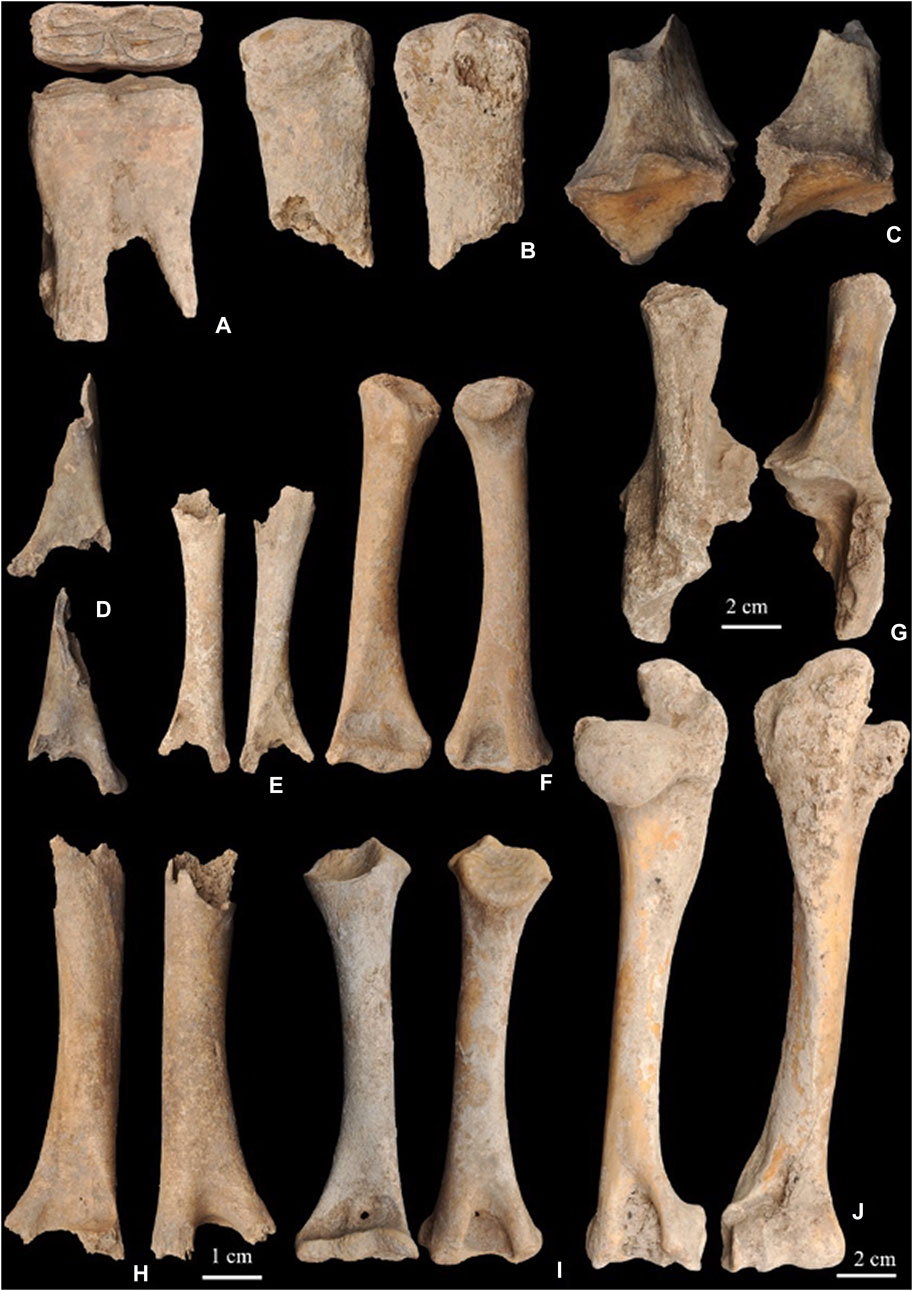
FIGURE 2. Faunal remains from layer IV2. (A) (Site # 3475) equid lower deciduous tooth (dp3). (B) (Site # 2969) brown bear metatarsal. (C) (Site # 3166) bovid calcaneum. (D) (Site # 3148) ovicaprid humerus. (E) (Site # 3147) ovicaprid humerus. (F) (Site # 2760) ovicaprid humerus. (G) (Site # 2444) bovid calcaneum. (H) (Site # 3167) ovicaprid humerus. (I) (Site # 2872) ovicaprid humerus. (J) (Site # 3279) ovicaprid humerus.
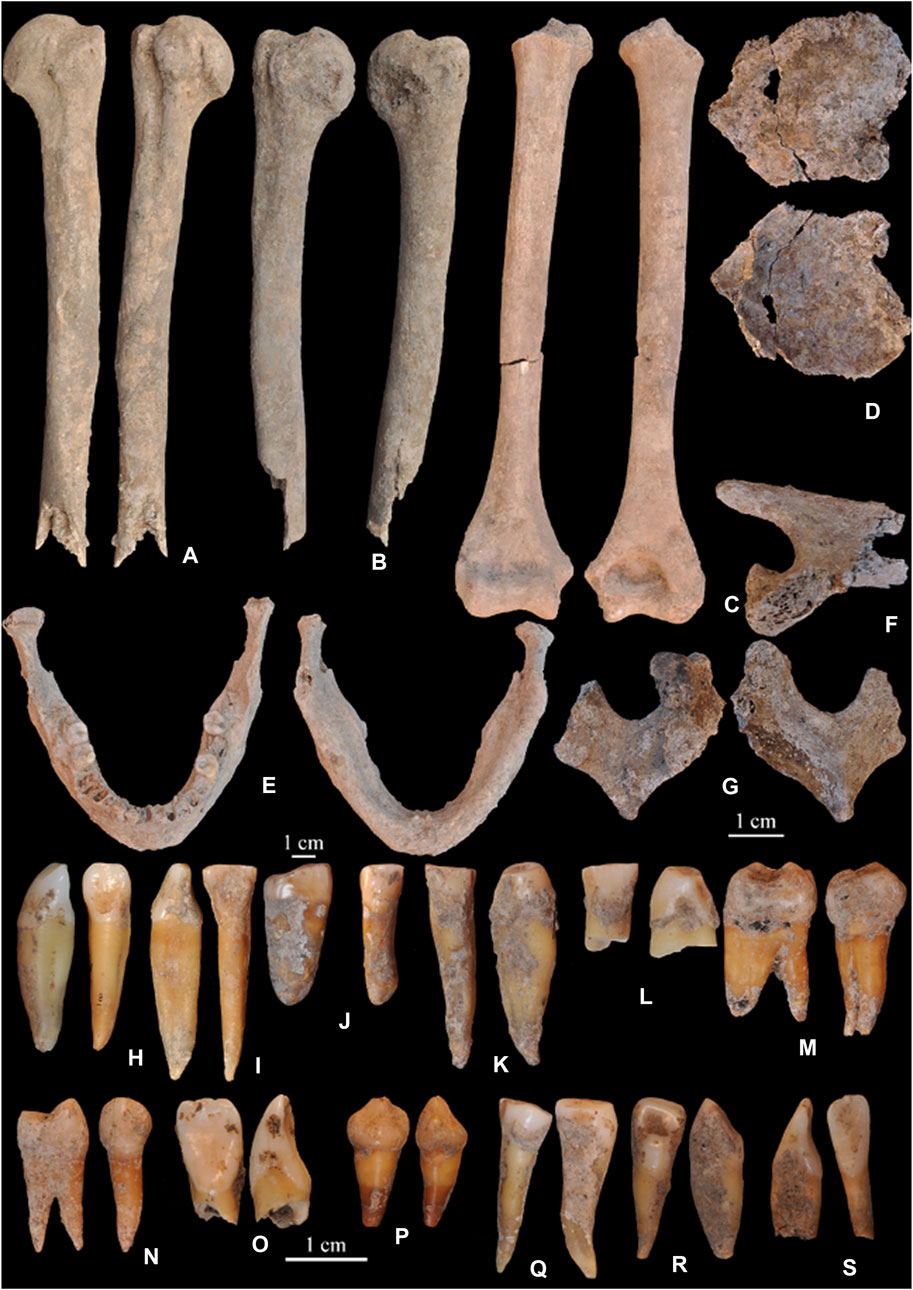
FIGURE 3. Human remains from layer IV2. (A) (Site # 3029) humerus. (B) (Site # 2996) humerus. (C) (Site # 3121) humerus. (D) (Site # 3066) frontal bone. (E) (Site # 3075) mandible. (F) (Site # 2517) scapula. (G) (Site # 2777) vertebra. (H) (Site # 2303) lower canine. (I) (Site # 2793) lower incisor. (J) (Site # 2595) upper premolar. (K) (Site # 3260) canine. (L) (Site # 3155) canine. (M) (Site # 2618) upper molar. (N) (Site # 2583) upper premolar. (O) (Site # 3334) upper incisor. (P) (Site # 2406) upper canine. (Q) (Site # 3439) premolar. (R) (Site # 3343) incisor. (S) (Site # 3158) upper incisor.
2.1 CN stable isotope analysis
Bone samples from 6 human individuals and 12 faunal specimens from five species were sampled from Cova Bonica’s Early Neolithic layer (IV2) (Table 1) to obtain information on their diet by carrying out C and N stable isotope ratio analysis. The consumption of C3 and C4 terrestrial resources is distinguishable by the δ13C stable isotope ratio (Van der Merwe and Vogel, 1978). Isotopic signals also help define the input in the diet of terrestrial and marine foods (Chisholm et al., 1982), although if freshwater or estuarine fish are involved, the interpretation of δ13C values becomes more complex (Salazar-García et al., 2014b). The δ15N stable isotope ratio increases by 3–5 ‰ up the food chain with each trophic level and is usually used to indicate the position of an organism in the food chain (Minagawa and Wada, 1984). Even if this quantification is less straightforward than previously thought (Hedges and Reynard, 2007), based on the exact values of the nitrogen ratio, it is potentially possible to differentiate between individuals that consumed more animal resources from those who consumed very little animal proteins (Fahy et al., 2013). Furthermore, the fact that aquatic food chains tend to contain more trophic levels than terrestrial ones, and therefore, show an increase in δ15N, helps to discriminate between the consumption of marine or C4 terrestrial foods when samples are 13C enriched (Schoeninger and DeNiro, 1984).
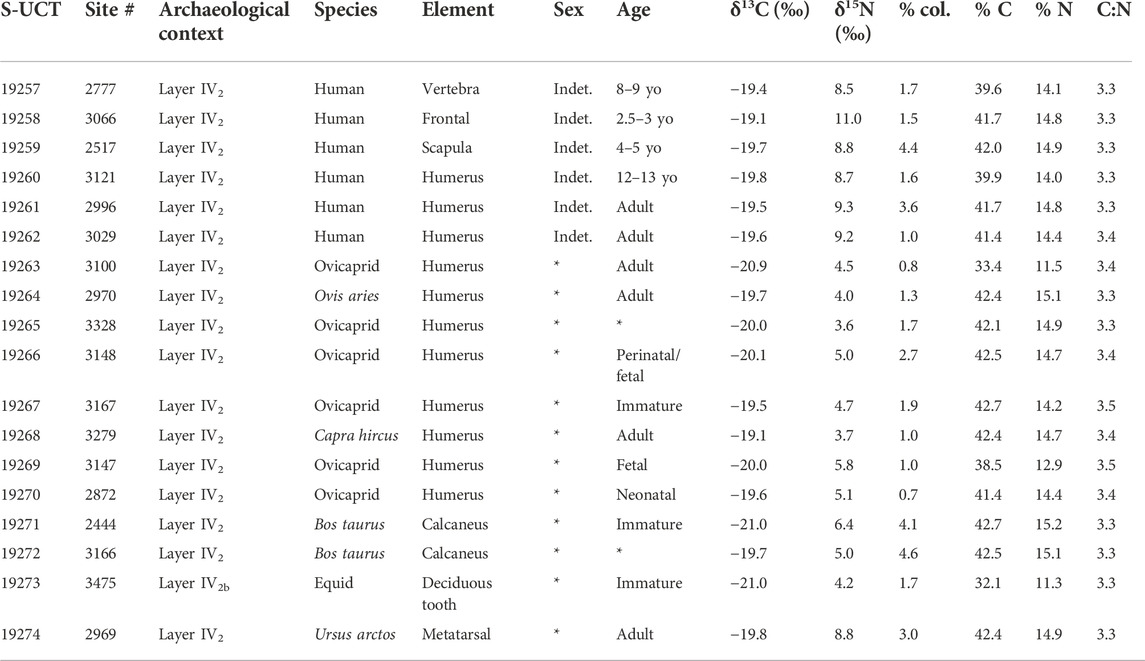
TABLE 1. Cova Bonica δ13C and δ15N values from fauna and humans, human sex and age, chronology, collagen control indicators (yield, % C, % N, and C:N), S-UCT number, sampled bone, and archaeological context.
Prior to analysis, visible contaminants were removed with aluminum oxide powder abrasion. Methods outlined by Richards and Hedges (1999) were followed to extract collagen for the C and N isotope ratio analysis at the Stable Isotope Facilities of the University of Cape Town (Cape Town, South Africa). Whole bone fragments weighing ca. 300 mg obtained from each of the specimens were demineralized in a 0.5 M HCl solution at 5°C. They were then rinsed three times with deionized water until the pH became neutral and gelatinized over 48 h at 70°C before being filtered and ultra-filtered using 9-ml EZEE© filters to remove small particles (<8 μm) and >30 kDa Amicon© ultrafilters, respectively. Finally, the purified solutions were frozen and lyophilized before being weighed into tin capsules and loaded into mass spectrometers.
The resultant collagen product was combusted to N2 and CO2. The carbon and nitrogen isotope ratios in collagen were measured in duplicate (reproducibility better than 0.1 ‰) using a Finnigan Delta plus XP continuous-flow isotope ratio mass spectrometer (Thermo Fisher Scientific, United States) after being combusted in an elemental analyzer Flash EA 1112 interfaced with it (Thermo Fisher Scientific, United States), at the Isotope Facilities of the University of Cape Town (South Africa). Stable carbon isotope ratios were expressed relative to the VPDB scale (Vienna PeeDee Belemnite) and stable nitrogen isotope ratios were measured relative to the AIR scale (atmospheric N2), using the delta notation (δ) in parts per thousand (‰). Repeated analysis of internal and international standards (methionine, Merk Gelatin, Valine, and seal bone) determined an analytical error <0.1 ‰ (1σ) for δ13C and δ15N.
2.2 Dental calculus microremains
For samples prepared for this study, teeth were evaluated and sampled for dental calculus with a scalar in the field (Table 2). Samples were subsampled in the Department of Human Evolution at the Max Planck Institute for Evolutionary Anthropology (Leipzig, Germany). When possible, we sampled chunks of supragingival dental calculus by prying them off, rather than scraping off a powder but a significant number of samples were of powder. The samples were then processed in a Heraeus safety-controlled airflow cabinet at the labs of the Department of Primatology in the Max Planck Institute for Evolutionary Anthropology. We used EDTA to decalcify the sample (Power et al., 2015; Tromp et al., 2017).
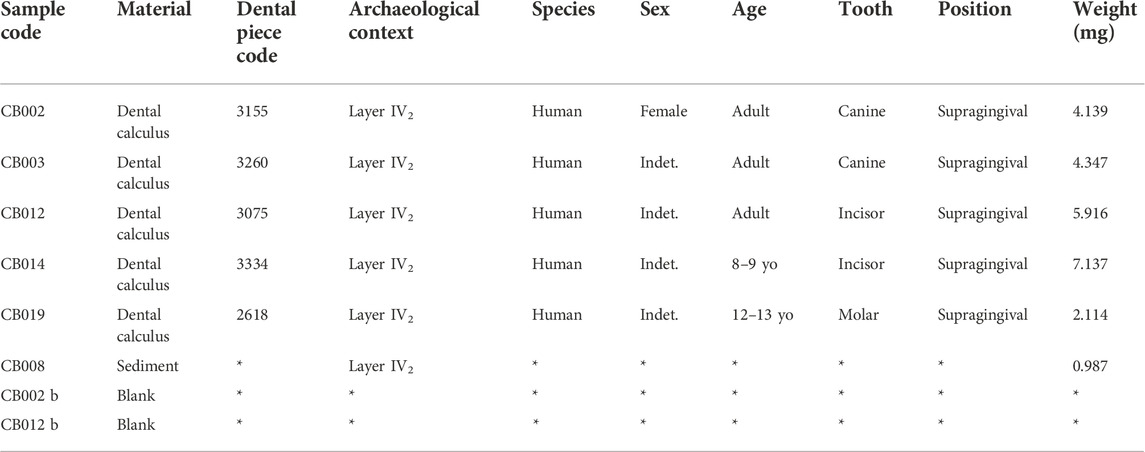
TABLE 2. Cova Bonica dental calculus samples taken for microremain microscopic analysis.
After the decalcification, the sample was rinsed twice with distilled water after centrifugation at 2,000×g for 10 min (Roth Mini-Centrifuge). Then, after the first decanting, the tubes were refilled with a 25% glycerin solution. Then we mounted 20 μL of the solution on a slide with an 18 × 18 mm or a 24 × 24 mm coverslip. During the preparation of samples, we prepared slides using an identical mounting procedure using 25% glycerin solution but without any dental calculus, to serve as a control blank to expose environmental contamination. We counted starches, phytoliths, and all other microremains irrespective of the type at 200–400x magnification using an absolute counting method with a light microscope under brightfield and cross-polarized light on an Axio Scope A1, Zeiss microscope with AxioVs40 V 4.8.2.0 software. Microremains were classified according to standard conventions (ICPN 2.0, Neumann et al., 2019).
3 Results
3.1 CN stable isotope analysis
Samples from six humans and 12 faunal specimens from five different species (Ovis aries, Capra hircus— ovicaprid when not assigned to species taxon—, Bos taurus, equid, and Ursus arctos) were taken for stable isotope analysis. All samples yielded sufficient collagen in the >30 kDa fraction for the δ13C and δ15N analysis in duplicate. All of them met published collagen quality controls: appropriate CN elemental percentages together with C:N ratios between 2.9 and 3.6 (De Niro, 1985; Ambrose, 1990; Van Klinken, 1999). All isotope ratio results from Cova Bonica are given in Table 1 and shown in Figure 4.
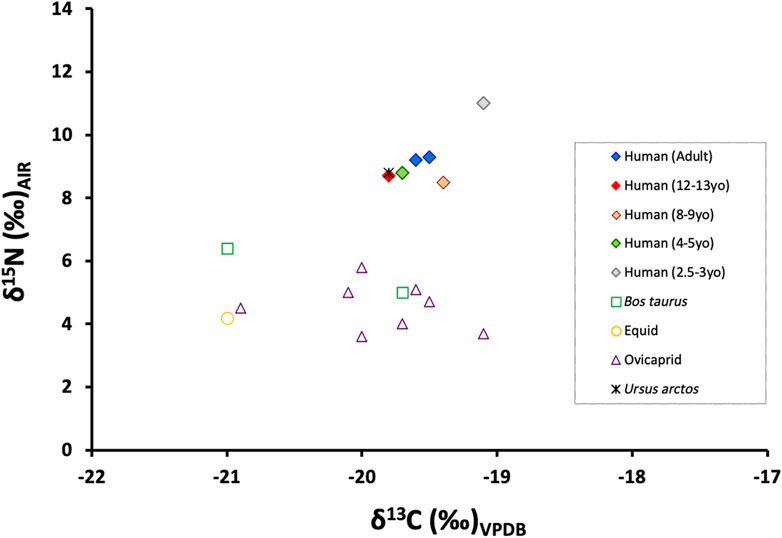
FIGURE 4. Plot of human and animal bone collagen δ13C and δ15N values from Cova Bonica.
Analyzing the carbon values of the 11 terrestrial herbivores, it can be seen that their δ13C mean value is -20.1 ± 0.6 (1σ) ‰ and its minimum and maximum values are -21.0 ‰ and -19.1 ‰, respectively. This herbivore value is compatible with a typical C3 terrestrial ecosystem. Analyzing the nitrogen values, the herbivore mean δ15N value is 4.7 ± 0.9 (1σ) ‰ and has minimum and maximum values of 3.6 and 6.4 ‰, respectively, establishing the trophic baseline for human subsistence. The only available omnivore carbon value is -19.8 ‰, which is also consistent with the general herbivore values and a terrestrial C3 food ecosystem. The δ15N value of the omnivore specimen analyzed is 8.8 ‰, which is 4.1 ‰ higher than the herbivore mean value and situates the omnivore (Ursus arctos) almost one complete step higher than the herbivores in the food chain. Unfortunately, no aquatic resources are available for this site, and thus, the marine and freshwater-specific “baseline” is lacking for Cova Bonica.
All humans from the Early Neolithic period (n=6) have δ13C and δ15N mean values of −19.5 ± 0.2 (1σ) ‰ (min: −19.8 ‰ and max: −19.1 ‰) and 9.3 ± 0.9 (1σ) ‰ (min: 8.5 ‰ and max: 11.0 ‰), respectively. These δ13C values portray a diet based on terrestrial C3 resources, while the δ15N values place humans at almost one full trophic level over herbivores (4.6 ‰ higher) and only slightly higher than the one omnivore analyzed (0.5 ‰ higher). There is no isotopic evidence of marine protein consumption.
3.2 Microremain analysis
Microremains were limited in the control samples, and their assemblages were entirely composed of cellulose-type fibers. Plant remains were present in all the archaeological dental calculus samples (Table 2). They occur in significant numbers in most samples. Of the microremains, charcoal was the dominant group but starches and phytoliths were also common. Fibers, pollen, fungal spores, calcium oxalates, and a variety of other unknown particles were also present. Their abundance varied considerably between samples (Figure 5).
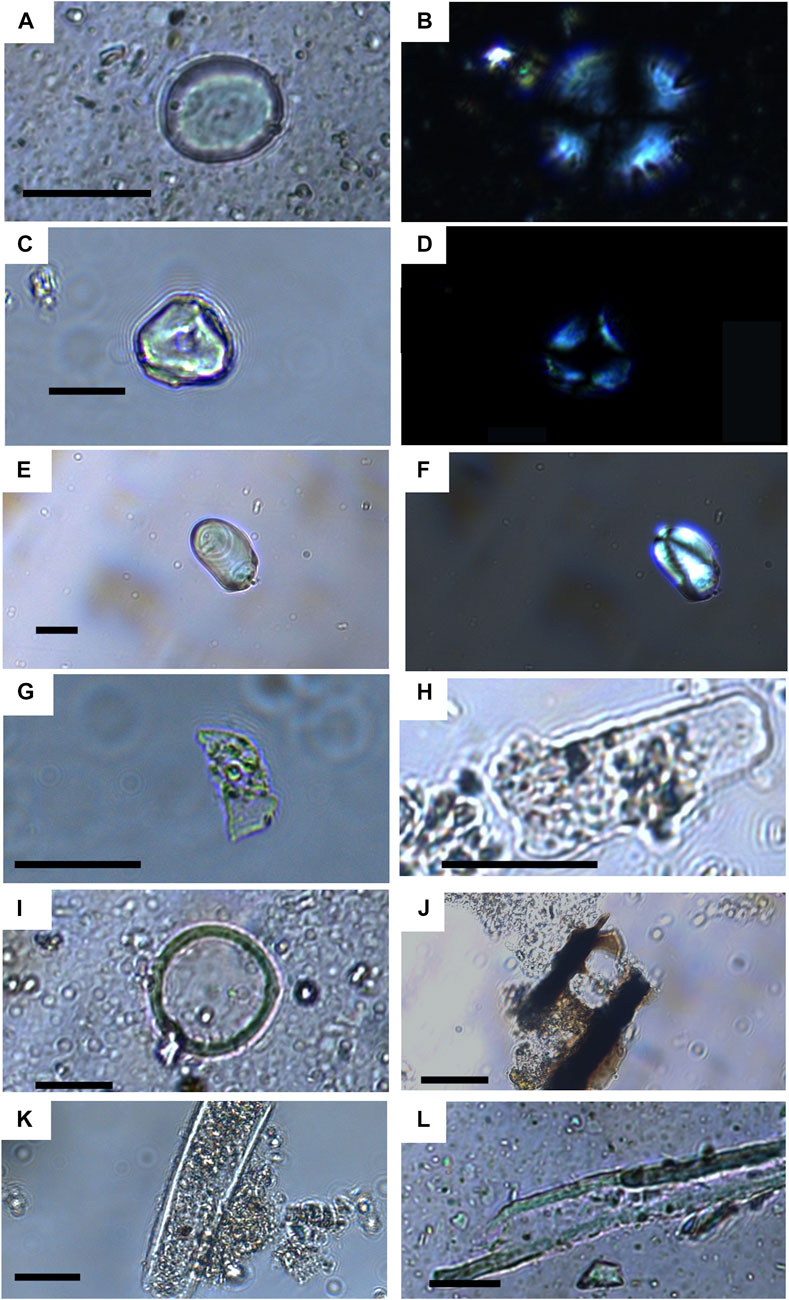
FIGURE 5. Micrographs of a selection of microremains. All scale bars represent 20 microns, (A) brightfield image of a lenticular starch in CB003, (B) cross-polarized image of the same starch, (C) brightfield image of faceted starch in CB003, (D) cross-polarized image of the same starch, (E) brightfield image of an eccentric starch in CB008, (F) cross-polarized image of the same starch, (G) trapezoidal (grass short-cell) phytolith in CB012, (H) elongate psilate thin phytolith, (I) indeterminate pollen from CB003, (J) charcoal particle in CB019, (K) broken probable non-human mammalian hair in CB003, and (L) plant fiber in CM003.
Phytoliths are well represented in the dental calculus samples, forming the most diverse category in the assemblage by occurring in five of the samples, totaling 23 phytoliths. Phytoliths from monocotyledon plants (wild grasses, domestic cereals, sedges, etc.) were the largest group, totaling 15 phytoliths (Table 3). Of these, Poaceae was dominant, comprising six phytoliths, present in five samples. A single Poaceae type was found in sediment (CB008). It cannot be determined if the assemblage is predominantly derived from the leaves/stems of wild or domestic. Grass short-cells (rondels and trapezoids) show grasses from the pooid subtribe which are consistent with, but not exclusive of, barleys and wheat (n =4). Spheroid ornate/echinate is present in two individuals. Eudicots are represented in only two samples and represent leafy matter.
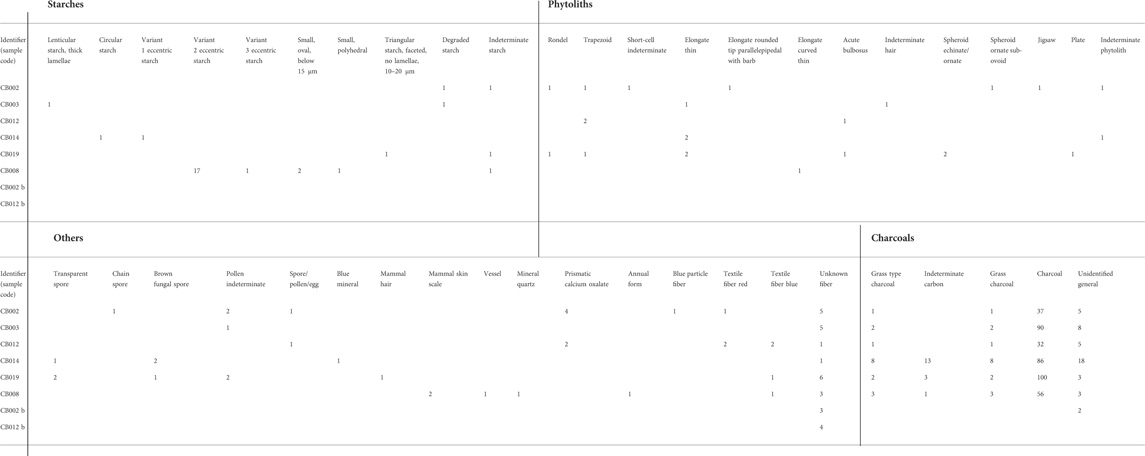
TABLE 3. Chart of total microremains. B samples represent controls. Eccentric starch variant 1: a slightly eccentric starch of variable sizes. Eccentric starch variant 2: a highly eccentric starch of variable sizes. Eccentric starch variant 3: an eccentric faceted starch over 20 microns in diameter.
Starches are widespread in the assemblages, being present in four dental calculus samples totaling six grains. They occur in small numbers (5 or fewer starch grains). In contrast, in sediment sample CB008, they occur in a large concentration. These in CB008 are largely comprised of a highly eccentric simple starch, sub-spherical sub-ovoid types, typically with a birefringence, representing a different origin than the dental calculus starches. Lenticular starch indicating Triticeae that could derive from cultivated wheat or barley was only found in CB003. Polyhedral-type starches also occurred in CB008, and a related form is found in CB019.
Pieces of combustion products are present in all five dental calculus samples, totaling 389. They include long thin pieces of charcoal, indicative of charred grasses, rather than wood or other materials (Crawford and Belcher, 2014). They also include unidentified combustion products and the largest category, nonspecific charcoal, which is probably mostly wood charcoal in origin.
A total of five grains of pollen were found in four samples. Grains of pollen could not be identified. Calcium oxalates (6) were found in CB002 and CB012. Fungal spores totaling 8 of an unknown family were found in three individuals. A non-silica plant vessel was found in CB008, but is non-diagnostic. A mixture of cellulose-type fibers is present in the samples, totaling 32, and 4 in the sediment samples. These included colored fibers dyed red and blue (Table 3). A probable mammalian hair, identified due to its birefringence, “surface scales,” and animal cell structure, was found in CB018. The medullar index indicates it is not human (Kshirsagar et al., 2009).
4 Discussion
Stable isotope and plant microremain studies are commonly used for past human dietary reconstructions. Until now, only few isotopic data of material directly dated to the earlier stages of the Neolithic are available in Eastern Iberia, unlike what is observed in Western Iberia (Guiry et al., 2016). No information on the diet inferred from plant microremains from dental calculus is available from the Early Neolithic. However, the Early Neolithic is a key period for understanding the process of Neolithization in Iberia, including dietary patterns and ecology. Mediterranean Iberia is also characterized by the low presence of human remains from this period compared with other regions, which makes the data from this study on the site of Cova Bonica even more valuable.
Previous paleogenetic data obtained from Cova Bonica (Olalde et al., 2015) support the evidence that new migrant populations introduced farming into western and central Europe. The presence of Cardial pottery, as documented in Cova Bonica, shows that the northeastern Iberian Neolithic is linked to the Italian Peninsula. It is in these regions that we see the earliest Cardial pottery and ovicaprid husbandry (Tresset and Vigne 2007; Vigne 2007) together with wheat and barley (Zapata et al., 2004; Antolín et al., 2015). Accordingly, the archaeological record (stratigraphy, radiocarbon dating, and artifacts) along the Mediterranean shoreline suggests a maritime colonization process (Zilhão 2001) that bypassed geographic obstacles using enclaves to rapidly settle lands optimal for farming (Bernabéu et al., 2015) (Figure 6A). Further insights into dietary patterns from these earlier stages of the Neolithic can help clarify the subsistence strategies of these first farming societies in the region.
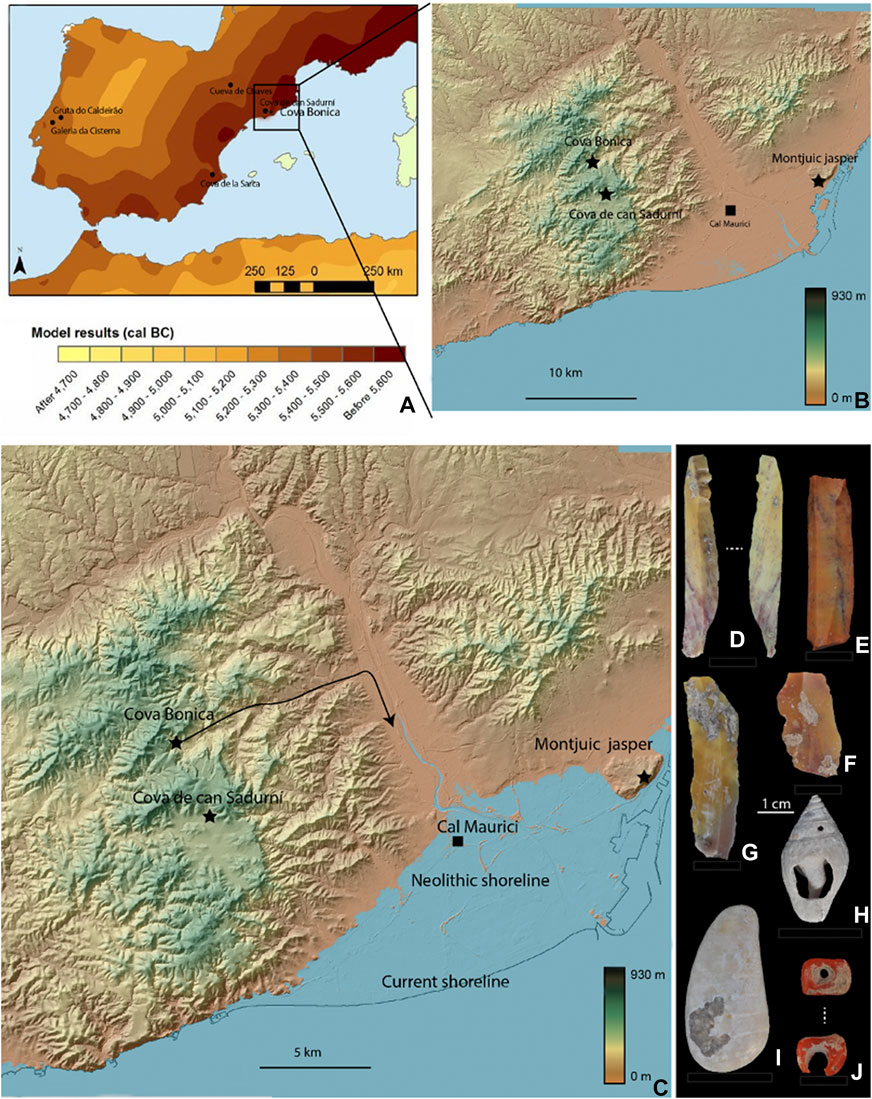
FIGURE 6. Cova Bonica in the context of regional settings. (A): Model predictions for the Neolithic expansion: leapfrog voyaging with jumps of about 350 km (Isern et al., 2017). (B–C): Digital elevation model (DEM) constructed using GIS version 3.24.2 and signaling current coastline (B) and neolithic shoreline (C). Neolithic shoreline has been established using data provided from the Cal Maurici site (Daura et al., 2016). (D–G): Jasper blades and flakes recovered at layer IV2 coming from Montjuic (Barcelona). (H–J): Marine ornaments recovered at layer IV2 elaborated from Columbella rustica (H), polished shell (I), and red coral (J).
The Early Neolithic individuals from Cova Bonica show a tight range of carbon isotope ratios, portraying a homogeneous protein diet based on C3 terrestrial resources. While this is not unexpected for a Neolithic population, what is surprising is that there is no isotopic evidence whatsoever of marine protein consumption by any of the individuals. Taking into consideration that Cova Bonica is only 12 km away from the coastline (Figures 6.B,C), there is evidence at the same layers of the site of ornaments crafted from marine resources (Figures 6.H–J), and that there is jasper from the coast as well on the site (Figures 6.D–G), this population was aware of the marine environment. Of course, that there is no isotopic evidence of marine consumption does not mean necessarily that they did not eat any sea resources but that, if they ate them, they did so in such low amounts as to not be indicated by the collagen signal of their bones.
Having discarded any marine signal from the δ13C values, the differences observed in the δ15N values (range 8.5–11.0 ‰) must be considered under another light. This difference cannot be, at this point, attributed to sex, as all human individuals are of indeterminate sex. However, the age of the different individuals analyzed should be considered, especially since four of the six total human individuals are not adults. As can be seen in Figure 3, the youngest individual analyzed (2.5–3 yo) is the one with the highest δ15N value (ca. 2 ‰ higher than the adults) as well as a slightly higher δ13C value. All other sub-adult individuals (ages ranging from 4–13 yo) have isotopic values similar to those of the adults analyzed. This pattern could suggest that at the age of 2.5–3 yo, the individuals were still breastfed and in the process of being weaned (Fogel et al., 1989; Fuller et al., 2006; Herring et al., 1998). Although the sample is small, there is scarce information about these social practices from Early Neolithic people which highlights the relevance of this finding.
For all individuals from 4 years old onward, nitrogen isotope ratios show that humans possess a higher trophic level than herbivores, but have similar δ15N values to the omnivore (brown bear). The archaeological record of Cova Bonica identified butchery activities mainly in adult ovicaprines, but also in other taxa, such as large bovids (Daura et al., 2019), demonstrating meat processing of domesticates. Moreover, the cave was used as a sheepfold indicating the relevance of husbandry practices for these individuals. All of this suggests that they consumed herbivore animal protein (possibly mainly ovicaprids), but also that they consumed plant foods (enough as to have similar values to the omnivore analyzed). While isotope analysis cannot shed further light on the type of plant resources consumed, the study of plant microremains trapped in dental calculus can.
The dental calculus assemblages show a distinct pattern that contrasts with the sparse remains in the control samples. This indicates the substantial integrity of the finds and that the microremains are archaeological with few phytoliths in the sediment sample. Poaceae is the dominant group of the assemblage, but counts are low relative to studies of many comparable grain-based economies (Scott et al., 2021). The only other Late prehistoric site from Mediterranean Iberia from which microremains from dental calculus have been studied is Camino del Molino (Murcia, Spain), although the site is from the Chalcolithic period (Power et al., 2014). The example of Camino del Molino does show a parallel with a relatively low-density assemblage missing expected morphotypes such as dendritics. At Camino del Molino, we do find some individuals with relatively higher numbers, as well as rare non-phytolith microremains such as sponge spicules and others. Given the purported grain-based Neolithic economy, the microremain counts in the Cova Bonica dental calculus are less than expected. They also lack unique husk types. The low Poaceae microremains count can be considered cryptic, but it could relate to limited cereal cultivation, free threshing versus non-free threshing varieties, or growing conditions that did not encourage phytolith formation in plant cells (Jenkins et al., 2011). Alternatively, it may be a product of specific grain processing methods that minimized phytolith intake.
There is evidence of eudicots, but they are rare and it is unclear if they had dietary significance. The occasional calcium oxalates we found may have also been derived from these eudicots, but they also could have originated from angiosperms. The starch assemblage is comparatively more numerous than the phytolith assemblage, but the non-dietary starches in CB008 drive this. Unidentified eccentric starches are predominant across the assemblage, representing plant storage organs, presumably from the C3 pathway. Triticeae starch occurs, reinforcing the data from phytoliths of cereal use. The rich presence of charcoal indicates close contact with fires (Hardy et al., 2012; Buckley et al., 2021). It is unclear how close this contact was and whether these particles represent airborne charcoal inhaled, water-carried charcoal that was drunk, or the consumption of charcoal-covered or other sources.
The Cova Bonica isotope values complement the picture that the isotope analysis has already portrayed for Mediterranean Iberia from the Mesolithic up to the Chalcolithic (Table 4). Especially useful for this are the Early Neolithic individuals, as only very few have been published before for this chronology in the region. As we can see in the table, the values from all chronological periods (Mesolithic, Early Neolithic, Middle Neolithic, Late Neolithic, and Chalcolithic) are partially overlapping. Regarding marine protein, Mesolithic hunter-gatherers show intermixed isotopic values whether from coastal or inland sites, which is expected from a population with high mobility through different environments. Examples of this are the inland sites of Santa Maira and Cingle del Mas Nou and the coastal site of El Collado, as previous studies from all three reported individuals with both a full terrestrial isotopic signature and others with enough marine protein consumption to have it recorded in the collagen isotopic signal. However, during the Early Neolithic, not even individuals from coastal sites have a clear marine isotopic signal. All Early Neolithic individuals from the region studied, whether from the coastal (Cova Bonica, Costamar, and Cueva de Nerja) or inland sites (Feixa del Moro), show a terrestrial isotopic signature. It is from the Middle Neolithic onward, and especially during the last stages of the Neolithic and the Chalcolithic periods, when people start to live in more stable settlements and we could consider that the amount of marine resource consumption is many times linked to the proximity to the sea rather than to chronology or archaeological culture. Suggestive of this pattern is that most individuals reported with marine protein input in the diet are from sites directly located on the coastline (e.g., Tossal de les Basses). This same diachronic pattern is observed even in individuals from different time periods of a single same coastal site, Costamar, where its Early Neolithic individuals have a full terrestrial isotopic signal while its Late Neolithic individuals show isotopic evidence of marine protein consumption.
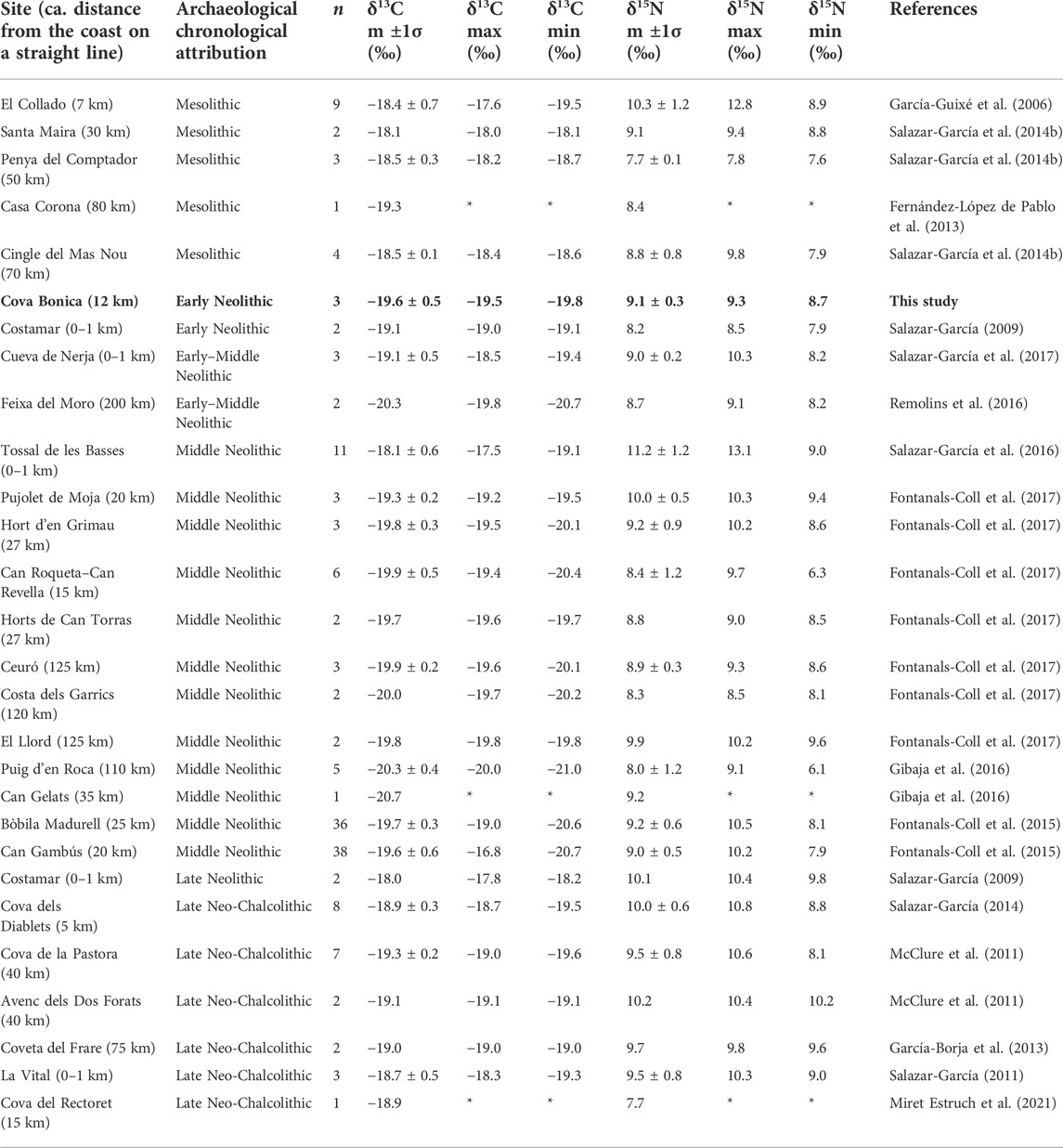
TABLE 4. List of all Mediterranean Iberia sites with published carbon and nitrogen isotopic ratio analysis on human bone collagen from the Mesolithic up to the Late Neolithic–Chalcolithic period. Only samples from human adults for which both C and N results are available which meet collagen quality controls have been considered here.
We must keep in mind that some of the differences observed between chronological periods and even sites of the same period could be due to non-dietary influences that modify the isotopic composition of resources consumed regardless of the resource type (i.e., factors that influence the food chain values themselves). For example, different environmental conditions between contemporary sites (e.g., coastal vs. inland; mountain vs. plain), or even between sites of a single geographic area but of different chronology, could have different isotopic food chain baselines (Goude and Fontugne, 2016). The use of complex husbandry practices and associated manuring in the region from the Late Neolithic onward could also influence crops, as well as domestic animals that feed on them, and, in turn, humans who consume the domestic crops or animals (Bogaard et al., 2007). Physiological traits of both the animals consumed (e.g. infantile vs adult specimens) and humans (age, nutritional status, and others), as well as the types of bones sampled (e.g., high vs. low collagen turnover skeletal elements), also influence the isotopic values obtained from human bone collagen. Some of these “non-dietary” influencing factors are difficult to assess, as the archaeological assemblages provide limited information. These issues caution us away from rigid quantified interpretations when interpreting isotopic studies applied to archaeological material.
Conclusion
The combination of stable isotopes and plant microremains trapped in dental calculus in Cova Bonica brings, for the first time, the combined direct evidence of the diet of these Early Neolithic farmers in the NE Mediterranean area. The human stable isotope analysis and plant microremains from the Cardial Neolithic of Cova Bonica document a diet based on terrestrial C3 resources. Although Cova Bonica is a mere 12 km from the current coastline, there is no isotopic evidence of marine protein consumption. The use of cereal and other plant foods is supported by plant microremains trapped in dental calculus.
The absence of marine and freshwater foods in the diet of Cova Bonica humans suggests that newcomers, at least initially, did not adapt to the fresh resources in the new land. In addition, the absence of a marine diet in people who moved through sea transport is, according to the coastal spread of the first farmers’ model, remarkable. Archaeological data from the early Neolithic suggest that farming people expand hand-in-hand with this “package:” pottery, domesticated plants, livestock, polished stones, and other archaeological materials. Thus, the Cova Bonica results support the notion that newcomers expanded bearing their own subsistence strategy, focused on terrestrial resources, such as husbandry of ovicaprines and domesticated plants, instead of exploring the alternatives that became available in new territories. This lifestyle is implemented in the regions that they arrived at with suitable lands (i.e., deltaic, estuarine, and lagoonal environments) for agriculture and animal husbandry.
Data availability statement
The original contributions presented in the study are included in the article; further inquiries can be directed to the corresponding author.
Author contributions
DS initiated and coordinated the study. JD and MS conducted the archaeological/anthropological work and the curation of the archeological material. DS, RP, JD, and MS participated in the sample management and gathering of contextual information. DS and RP organized the sample collection for analyses. DS and RP performed the laboratory work. All authors contributed to the interpretation of the data. DS wrote the first version of the manuscript, which was edited by RP, JD, and MS.
Funding
DS acknowledges funding for this analytical study from the Generalitat Valenciana (CIDEGENT/2019/061) and the Spanish government (PID 2019-111207GA-I00 and EUR2020-112213). This study is also part of the output of research project CLT/2022/ARQ001SOLC/128—“Les societats caçadores recol·lectores del Paleolític mitjà i superior: Estratègies de subsistència” i canvis climàtics (Generalitat de Cataluña), supported by projects 2017SGR-00011 (Generalitat de Catalunya) and PID2020-113960GB-100 (Ministerio de Ciencia e Innovación). JD and MS hold a Ramon y Cajal contract (RYC-2015-17667 and RYC-2021-032099-1).
Acknowledgments
The authors thank Ian Newton for technical assistance at UCT. They thank Roisin Murtagh and Tobias Deschner (MPI-EVA) for laboratory access. They also thank Ingelise Stuijts for comments on the findings. They acknowledge the Castelldefels City Council (La Guixera Laboratory) where the archaeological analyses were carried out. Fieldwork was also sponsored by the Ajuntament de Vallirana. They thank the three reviewers for the comments and feedback.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors, and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ambrose, S. H. (1990). Preparation and characterization of bone and tooth collagen for isotopic analysis. J. Archaeol. Sci. 17 (4), 431–451. doi:10.1016/0305-4403(90)90007-r
Ambrose, S. H., and Norr, L. (1993). “Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate,” in Prehistoric human bone (Berlin, Heidelberg: Springer), 1–37.
Antolín, F., Jacoment, S., and Buxó, R. (2015). The hard knock life: Archeobotanical data on farming practices during the Neolithic (5400–300 CAL B.C.) in the NE of Iberian Peninsula. J. Archaeol. Sci. 61, 90–104. doi:10.1016/j.jas.2015.05.007
Armitage, P. L. (1975). The extraction and identification of opal phytoliths from the teeth of ungulates. J. Archaeol. Sci. 2 (3), 187–197. doi:10.1016/0305-4403(75)90056-4
Baldellou, V. (1974). Los materiales arqueológicos de la «Cova Bonica» de Vallirana (Barcelona). Empúries Rev. món clàssic Antig. tardana 36, 1–19.
Bernabéu, J., Barton, M., Pardo, S., and Bergin, S. M. (2015). Modeling initial neolithic dispersal. The first agricultural groups in west mediterranean. Ecol. Model. 307, 22–31. doi:10.1016/j.ecolmodel.2015.03.015
Bogaard, A., Heaton, T. H. E., Poulton, P., and Merbach, I. (2007). The impact of manuring on nitrogen isotope ratios in cereals: archaeological implications for reconstruction of diet and crop management practices. J. Archaeol. Sci. 34, 335–343.
Buckley, S., Power, R. C., Andreadaki-Vlazaki, M., Akar, M., Becher, J., Belser, M., et al. (2021). Archaeometric evidence for the earliest exploitation of lignite from the bronze age Eastern Mediterranean. Sci. Rep. 11 (1), 24185. doi:10.1038/s41598-021-03544-w
Chisholm, B. S., Erle Nelson, D., and Schwarcz, H. P. (1982). Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science 216 (4550), 1131–1132. doi:10.1126/science.216.4550.1131
Crawford, A. J., and Belcher, C. M. (2014). Charcoal morphometry for paleoecological analysis: The effects of fuel type and transportation on morphological parameters. Appl. Plant Sci. 2 (8), 1400004. doi:10.3732/apps.1400004
Daura, J., Sanz, M., Ramos, J., Riera, S., Miras, Y., Allué, E., et al. (2016). Palaeoenvironmental record of the Cal Maurici wetland sediment archive in Barcelona (NE Iberian Peninsula) between c. 6000 and 4000 cal. yr BP. Holocene 26, 1020–1039. doi:10.1177/0959683616632891
Daura, J., Sanz, M., Oms, F. X., Pedro, M., Martínez, P., Mendiela, S., et al. (2019). Deciphering neolithic activities from a cardial burial site (cova Bonica) on the Western mediterranean coast. J. Archaeol. Sci. Rep. 23, 324–347. doi:10.1016/j.jasrep.2018.10.036
De Niro, M. J. (1985). Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 3317, 806–809. doi:10.1038/317806a0
Fahy, G. E., Richards, M., Riedel, J., Hublin, J. J., and Boesch, C. (2013). Stable isotope evidence of meat eating and hunting specialization in adult male chimpanzees. Proc. Natl. Acad. Sci. U. S. A. 110 (15), 5829–5833. doi:10.1073/pnas.1221991110
Fernández López de Pablo, J., Salazar-García, D. C., Subirà, M. E., Roca-Togores, C., Gómez Puche, M., Richards, M. P., et al. (2013). Late mesolithic burials at casa corona (villena, Spain): Direct radiocarbon and palaeodietary evidence of the last forager populations in eastern Iberia. J. Archaeol. Sci. 40, 671–680. doi:10.1016/j.jas.2012.09.005
Fogel, M., Tuross, N., and Owsley, D. W. (1989). Nitrogen isotope tracers of human lactation in modern and archaeological populations. Washington, DC: Geophysical Laboratory-Carnegie Institution.
Fontanals−Coll, M., Subirà, M. E., Díaz-Zorita Bonilla, M., Duboscq, S., and Gibaja, J. F. (2015). Investigating palaeodietary and social differences between two differentiated sectors of a Neolithic community, La Bòbila Madurell-Can Gambús (north−east Iberian Peninsula). J. Archaeol. Sci. Rep. 3, 160–170. doi:10.1016/j.jasrep.2015.06.013
Fontanals-Coll, M., Subirà, M. E., Díaz-Zorita, M., and Gibaja, J. F. (2017). First insight into the neolithic subsistence economy in the north-east iberian Peninsula: Paleodietary reconstruction through stable isotopes. Am. J. Phys. Anthropol. 162 (1), 36–50. doi:10.1002/ajpa.23083
Fuller, B. T., Fuller, J. L., Harris, D. A., and Hedges, R. E. M. (2006). Detection of breastfeeding and weaning in modern human infants with carbon and nitrogen stable isotope ratios. Am. J. Phys. Anthropol. 129, 279–293.
García Borja, P., Pérez Fernández, A., Biosca Cirujeda, V., Ribera i Gomes, A., and Salazar-García, D. C. (2013). “Los restos humanos de la Coveta del Frare (La Font de la Figuera, València),” in El Naiximent d'un Poble. Història i Arqueologia de la Font de la Figuera, Ajuntament de la Font de la Figuera. Editors P. García Borja, E. Revert Francés, A. Y. Ribera i Gomes, and V. Biosca Cirujeda, 47–60.
Garcia-Guixé, E., Richards, M. P., and Subirà, M. E. (2006). Palaeodiets of humans and fauna at the Spanish mesolithic site of El Collado. Curr. Anthropol. 47, 549–557. doi:10.1086/504170
Gibaja, J. F., Fontanals-Coll, M. F., Subosqu, S., Oms, F. X., Augés, A., Javier Santos, F., et al. (2016). Human diet and the chronology of neolithic societies in the north-east of the iberian Peninsula: The necropolises of puig d'en roca and can gelats (girona, Spain). Archaeol. Anthropol. Sci. 9, 903–913. doi:10.1007/s12520-015-0311-y
Goude, G., and Fontugne, M. (2016). Carbon and nitrogen isotopic variability in bone collagen during the Neolithic period: Influence of environmental factors and diet. J. Archaeol. Sci. 70, 117–131.
Goude, G., Salazar-García, D. C., Power, R. C., Terrom, J., Rivollat, M., Deguilloux, M. F., et al. (2019). A multidisciplinary approach to Neolithic life reconstruction. J. Archaeol. Method Theory 26 (2), 537–560. doi:10.1007/s10816-018-9379-x
Goude, G., Salazar-García, D. C., Power, R. C., Rivollat, M., Gourichon, L., Deguilloux, M. F., et al. (2020). New insights on Neolithic food and mobility patterns in Mediterranean coastal populations. Am. J. Phys. Anthropol. 173, 218–235. doi:10.1002/ajpa.24089
Guiry, E. J., Hillier, M., Boaventura, R., Silva, A. M., Oosterbeek, L., Tiago, T., et al. (2016). The transition to agriculture in south-western Europe: new isotopic insights from Portugal's Atlantic coast. Antiquity 90, 604–616.
Hardy, K., Buckley, S., Collins, M. J., Estalrrich, A., Brothwell, D., Copeland, L., et al. (2012). Neanderthal medics? Evidence for food, cooking, and medicinal plants entrapped in dental calculus. Naturwissenschaften 99 (8), 617–626. doi:10.1007/s00114-012-0942-0
Hedges, R. E. M., and Reynard., L. M. (2007). Nitrogen isotopes and the trophic level of humans in archaeology. J. Archaeol. Sci. 34 (8), 1240–1251. doi:10.1016/j.jas.2006.10.015
Hedges, R. E. M., Clement, J. G., Thomas, C. D. L., and O'Connell, T. C. (2007). Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. Am. J. Phys. Anthropol. 133, 808–816. doi:10.1002/ajpa.20598
Herring, D. A., Saunders, S. R., and Katzenberg, M. A. (1998). Investigating the weaning process in past populations. Am. J. Phys. Anthropol. 105, 425–439.
Henry, Amanda G., and Piperno, Dolores R. (2008). Using plant microfossils from dental calculus to recover human diet: A case study from tell al-raqā'i, Syria. J. Archaeol. Sci. 35, 1943–1950. doi:10.1016/j.jas.2007.12.005
Howland, M. R., Corr, L. T., Young, S. M. M., Jones, V., Jim, S., Van Der Merwe, N. J., et al. (2003). Expression of the dietary isotope signal in the compound-specific δ13C values of pig bone lipids and amino acids. Int. J. Osteoarchaeol. 13 (1–2), 54–65. doi:10.1002/oa.658
Isern, N., Zilhão, J., Fort, J., and Ammerman, A. J. (2017). Modeling the role of voyaging in the coastal spread of the early neolithic in the west mediterranean. Proc. Natl. Acad. Sci. U. S. A. 114, 897–902. doi:10.1073/pnas.1613413114
Jenkins, E., Jamjoum, K., and Nuimat, S. Al. (2011). “Irrigation and phytolith formation: An experimental study,” in Water, life and civilisation: Climate, environment and society in the Jordan valley. Editors S. Mithen, and E. Black (Cambridge University Press), 347–372.
Jim, S., Ambrose, S. H., and Evershed, R. P. (2004). Stable carbon isotopic evidence for differences in the dietary origin of bone cholesterol, collagen and apatite: Implications for their use in dietary reconstruction. Geochim. Cosmochim. Acta. 68, 61–72.
Katzenberg, M. A. (2012). “The ecological approach: Understanding past diet and the relationship between diet and disease,” in A companion to paleopathology. Editor A. L. Grauer (New Jersey: John Wiley & Sons), 97–113.
Kshirsagar, S. V., Singh, B., and Fulari, S. P. (2009). Comparative study of human and animal hair in relation with diameter and medullary index. Indian J. Forensic Med. Pathology 2 (3), 105–108.
Lee-Thorp, J. A. (2008). On isotopes and old bones. Archaeometry 50, 925–950. doi:10.1111/j.1475-4754.2008.00441.x
Leonard, C. A., Vashro, L., O'Connell, J. F., and Henry, A. G. (2015). Plant microremains in dental calculus as a record of plant consumption: A test with twe forager-horticulturalists. J. Archaeol. Sci. Rep. 2, 449–457. doi:10.1016/j.jasrep.2015.03.009
McClure, S., García-Puchol, O., Roca, C., Culleton, B., and Kennett, D. (2011). Osteological and paleodietary investigation of burials from Cova de la Pastora, Alicante, Spain. J. Archaeol. Sci. 38, 420–428. doi:10.1016/j.jas.2010.09.023
Minagawa, Masao, and Wada, Eitaro (1984). Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochimica Cosmochimica Acta 48 (5), 1135–1140. doi:10.1016/0016-7037(84)90204-7
Miret Estruch, C., Cardona Escrivà, J., Salazar-García, D. C., García Borja, P., Pérez Fernández, A., and Guerrero Blázquez, M. (2021). La collecció arqueológica preshitòrica de la cova del Rectoret (Gandia, la Safor, País Valencià). Recer. del Mus. d’Alcoi 30, 43–62.
Neumann, K., Stromberg, C. A. E., Ball, T., Albert, R. M., Vrydaghs, L., and Cummings, L. S. (2019). International code for phytolith nomenclature (ICPN) 2.0. Ann. Bot. 124 (2), 189–199. doi:10.1093/aob/mcz064
Olalde, I., Schroeder, H., Sandoval-Velasco, M., Vinner, L., Lobón, I., Ramirez, O., et al. (2015). A common genetic origin for early farmers from mediterranean cardial and central European LBK cultures. Mol. Biol. Evol. 32, 3132–3142. doi:10.1093/molbev/msv181
Oms, F. X., Daura, J., Sanz, M., Mendiela, S., Pedro, M., and Martínez, P. (2017). First evidence of collective human inhumation from the cardial neolithic (cova Bonica, Barcelona, NE iberian Peninsula). J. Field Archaeol. 42, 43–53. doi:10.1080/00934690.2016.1260407
Power, R. C., Salazar-García, D. C., Wittig, R. M., and Henry, A. G. (2014). Assessing use and suitability of scanning electron microscopy in the analysis of micro remains in dental calculus. J. Archaeol. Sci. 49, 160–169. doi:10.1016/j.jas.2014.04.016
Power, R. C., Salazar-García, D. C., Wittig, R. M., Freiberg, M., and Henry, A. G. (2015). Dental calculus evidence of Taï Forest Chimpanzee plant consumption and life history transitions. Sci. Rep. 5, 15161. doi:10.1038/srep15161
Remolins, G., Gibaja, J. F., Allièse, F., Fontanals-Coll, M., Martin, P., Masclans, A., et al. (2016). The neolithic necropolis of La feixa del Moro (juberri, Andorra): New data on the first farming communities in the pyrenees. Comptes Rendus Palevol 15 (5), 537–554. doi:10.1016/j.crpv.2015.11.005
Richards, M. P., and Hedges, R. E. M. (1999). Stable isotope evidence for similarities in the types of marine foods used by Late Mesolithic humans at sites along the Atlantic coast of Europe. J. Archaeol. Sci. 26 (6), 717–722. doi:10.1006/jasc.1998.0387
Salazar-García, D. C. (2009). “Estudio de la dieta en la población neolítica de Costamar. Resultados preliminares de análisis de isótopos estables de carbono y nitrógeno,” in Flors Ureña E. (Coord.) Torre la Sal (Ribera de Cabanes, Castellón). Evolución del paisaje antrópico desde la prehistoria hasta el medioevo. M.P.A.C. 8 (Castellón: Servicio de Investigaciones Arqueológicas y Prehistóricas-Diputación de Castellón), 411–418.
Salazar-García, D. C., Vives-Ferrándiz, J., Fuller, B., and Richards, M. P. (2010). Alimentación estimada de la población del Castellet de Bernabé (ss. V-III a.C.) mediante el uso de ratios de isótopos estables de C y N. Saguntum Extra 9, 313–322.
Salazar-García, D. C. (2011). “Aproximación a la dieta de la población calcolítica de La Vital a través del análisis de isótopos estables del carbono y del nitrógeno sobre restos óseos,” in La Vital (Gandia, Valencia). Vida y muerte en la desembocadura del Serpis durante el III y el I milenio a.C. T.V. 113. Editors G. Perez Jordá, J. Bernabeu Aubán, Y. Carrión Marco, O. García-Puchol, L. L. Molina Balaguer, and M. Gómez Puche (València: Museu de Prehistòria de València-Diputación de Valencia), 139–143.
Salazar-García, D. C. (2014). “Estudi de la dieta en la població de Cova dels Diablets mitjançant anàlisi d’isòtops estables del carboni i del nitrogen en collàgen ossi. Resultats preliminars,” in La Cova dels Diablets (Alcalà de Xivert, Castelló). Prehistòria a la Serra d’Irta. Castellón, Diputació de Castelló. Editors G. Aguilella Arzo, D. Roman i Monroig, and P. García Borja, 67–78.
Salazar-García, D. C., Power, R. C., Sanchis-Serra, A., Villaverde, V., Walker, M. J., and Henry, A. G. (2013). Neanderthal diets in central and southeastern Mediterranean Iberia. Quat. Int. 318, 3–18. doi:10.1016/j.quaint.2013.06.007
Salazar-García, D. C., Richards, M. P., Nehlich, O., and Henry, A. G. (2014a). Dental calculus is not equivalent to bone collagen for isotope analysis: A comparison between carbon and nitrogen stable isotope analysis of bulk dental calculus, bone and dentine collagen from same individuals from the medieval site of El raval (alicante, Spain). J. Archaeol. Sci. 47, 70–77. doi:10.1016/j.jas.2014.03.026
Salazar-García, D. C., Aura, E., Olària, C., Talamo, S., Morales, J. V., and Richards, M. P. (2014b). Isotope evidence for the use of marine resources in the Eastern Iberian Mesolithic. J. Archaeol. Sci. 42, 231–240. doi:10.1016/j.jas.2013.11.006
Salazar-García, D. C., Romero, A., García-Borja, P., Subirà, E., and Richards, M. P. (2016). A combined dietary approach using isotope and dental buccal-microwear analysis of human remains from the Neolithic, Roman and Medieval periods from the archaeological site of Tossal de les Basses (Alicante, Spain). J. Archaeol. Sci. Rep. 6, 610–619. doi:10.1016/j.jasrep.2016.03.002
Salazar-García, D. C., Pérez-Ripoll, M., García-Borja, P., Jordá Pardo, J. F., and Aura Tortosa, J. E. (2017). “A terrestrial diet close to the coast: A case study from the neolithic levels of Nerja cave (málaga, Spain),” in Times of neolithic transition along the western mediterranean. Editors D. C. Salazar-García, and O. García-Puchol (Springer), 281–307.
Salazar-García, D. C., Fontanals-Coll, M., Goude, G., and Subirà, E. (2018). To 'seafood' or not to 'seafood'?" an isotopic perspective on dietary preferences at the Mesolithic-Neolithic transition in the Western Mediterranean. Quat. Int. 470, 497–510. doi:10.1016/j.quaint.2017.12.039
Salazar-García, D. C., Power, R. C., Rudaya, N., Kolobova, K., Markin, S., Krivoshapkin, A., et al. (2021). Dietary evidence from central asian neanderthals: A combined isotope and plant microremains approach at chagyrskaya cave (altai, Russia). J. Hum. Evol. 156, 102985. doi:10.1016/j.jhevol.2021.102985
Schoeller, D. A. (1999). Isotope fractionation: Why aren’t we what we eat? J. Archaeol. Sci. 26 (6), 667–673. doi:10.1006/jasc.1998.0391
Schoeninger, M. J., and DeNiro, M. J. (1984). Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochimica Cosmochimica Acta 48 (4), 625–639. doi:10.1016/0016-7037(84)90091-7
Scott, A., Power, R. C., Altmann-Wendling, V., Artzy, M., Martin, M. A. S., Eisenmann, S., et al. (2021). Exotic foods reveal contact between South Asia and the Near East during the second millennium BCE. Proc. Natl. Acad. Sci. U. S. A. 118 (2), e2014956117. doi:10.1073/pnas.2014956117
Tresset, A., and Vigne, J. D. (2007). “Substitution of species, techniques and symbols at the mesolithic-neolithic transition in western europe,” in Going over: The mesolithic-neolithic Transition in north-west europe, proceedings of the British academy 144. Editors Wittle, and V. Cummings (Oxford: Oxford University Press), 189–201.
Tromp, M., Buckley, H., Geber, J., and Matisoo-Smith, E. (2017). EDTA decalcification of dental calculus as an alternate means of microparticle extraction from archaeological samples. J. Archaeol. Sci. Rep. 14, 461–466. doi:10.1016/j.jasrep.2017.06.035
Van der Merwe, N. J., and Vogel, J. C. (1978). 13C Content of human collagen as a measure of prehistoric diet in Woodland North America. Nature 276, 815–816. doi:10.1038/276815a0
Van Klinken, G. J. (1999). Bone collagen quality indicators for palaeodietary and radiocarbon measurements. J. Archaeol. Sci. 26 (6), 687–695. doi:10.1006/jasc.1998.0385
Vigne, J-D. (2007). “Exploitation des animaux et néolithisation en Méditerranée nord-occidentale,” in Pont de Roque-Haute. Nouveaux Regards sur la Néolithisation de la France Méditerranéenne. Editors J. Guilaine, C. Manen, and J. D. Vigne (Toulouse: Archives d’Écologie Préhistorique), 221–301.
Villalba-Mouco, V., Utrilla, P., Laborda, R., Lorenzo, J. I., Martínez-Labarga, C., and Salazar-García, D. C. (2018a). Reconstruction of human subsistence and husbandry strategies from the Iberian Early Neolithic using carbon and nitrogen stable isotope ratio analysis. Am. J. Phys. Anthropol. 167, 257–271.
Villalba-Mouco, V., Sauqué, V., Sarasketa-Gartzia, I., Pastor, M. V., le Roux, P. J., Vicente, D., et al. (2018b). Territorial mobility and subsistence strategies during the Ebro Basin Late Neolithic-Chalcolithic: A multi-isotope approach from San Juan cave (Loarre, Spain). Quat. Int. 481, 28–41.
Villalba-Mouco, V., Sarasketa-Gartzia, I., Utrilla, P., Oms, F. X., Mendiela, S., Lorenzo, J. I., et al. (2019). Stable isotope ratio analysis of bone collagen as indicator of different environments conditions and dietary habits in Northeastern Iberia during the 4th and 3rd millenium calBC. Archaeol. Anthropol. Sci. 11, 3931–3947.
Warinner, C., Rodrigues, J. F. M., Vyas, R., Trachsel, C., Shved, N., Grossmann, J., et al. (2014). Pathogens and host immunity in the ancient human oral cavity. Nat. Genet. 46 (4), 336–344. doi:10.1038/ng.2906
Zapata, L., Peña-Chocarro, L., Pérez-Jordà, G., and Stika, H. (2004). Early neolithic agriculture in the iberian Peninsula. J. World Prehist. 18 (4), 283–325. doi:10.1007/s10963-004-5621-4
Keywords: Neolithic, diet, stable isotopes, plant microremains, Western Mediterranean
Citation: Salazar-García DC, Power RC, Daura J and Sanz M (2022) Diet at the onset of the Neolithic in northeastern Iberia: An isotope–plant microremain combined study from Cova Bonica (Vallirana, Catalonia). Front. Earth Sci. 10:957344. doi: 10.3389/feart.2022.957344
Received: 31 May 2022; Accepted: 17 October 2022;
Published: 28 November 2022.
Edited by:
Li Liu, Stanford University, United StatesReviewed by:
Subir Bera, University of Calcutta, IndiaJingbo Li, Stanford University, United States
Huan Liu, South China Normal University, China
Copyright © 2022 Salazar-García, Power, Daura and Sanz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Domingo C. Salazar-García, ZG9taW5nb2Nhcmxvcy5zYWxhemFyQHV2LmVz