 Jennifer L. Baltzer1*†
Jennifer L. Baltzer1*† Samuel Haché2†
Samuel Haché2† James Hodson3†
James Hodson3† M. Razu Ahmed4
M. Razu Ahmed4 Suzanne Carrière5,6
Suzanne Carrière5,6 Matthew Coyle7Eliot J. B. McIntire8,9Ashley McLaren7
Matthew Coyle7Eliot J. B. McIntire8,9Ashley McLaren7 Eamon Riordan-Short2Merritt R. Turetsky10
Eamon Riordan-Short2Merritt R. Turetsky10 Jurjen van der Sluijs4Joanna Wilson3
Jurjen van der Sluijs4Joanna Wilson3 Bradley K. Woodworth3
Bradley K. Woodworth3- 1Biology Department, Wilfrid Laurier University, Waterloo, ON, Canada
- 2Canadian Wildlife Service, Environment and Climate Change Canada, Yellowknife, NT, Canada
- 3Environment and Climate Change, Government of the Northwest Territories, Yellowknife, NT, Canada
- 4NWT Centre for Geomatics, Government of the Northwest Territories, Yellowknife, NT, Canada
- 5Arctic Institute of North America, University of Calgary, Calgary, AB, Canada
- 6Wek’èezhìı Renewable Resources Board, Yellowknife, NT, Canada
- 7Environment and Climate Change, Government of the Northwest Territories, Fort Smith, NT, Canada
- 8Pacific Forestry Centre, Canadian Forest Service, Natural Resources Canada, Victoria, BC, Canada
- 9Faculty of Forestry, Forest Resources Management, The University of British Columbia, Vancouver, BC, Canada
- 10Ecology and Evolutionary Biology Department, Renewable and Sustainable Energy Institute, University of Colorado Boulder, Boulder, CO, United States
The summer of 2023 was the first time the planet exceeded 1.5 °C above pre-industrial average temperatures and was recognized as the hottest year on record globally. In Canada, this translated to temperatures that were 2.2 °C above average, resulting in a record shattering fire season. Burning conditions were exceptional in many ways including large total burned area and individual fire size, vast swaths of short-interval reburns and severe combustion, fast-moving wildfires, and substantial burning outside of the typical fire season, reflecting climate warming-induced changes in fire regime. While we know how some components of biodiversity in the boreal biome have responded to historic burning, the 2023 fire season highlights that we are rapidly moving into a novel set of burning conditions and that we require better knowledge of how these conditions will alter landcover and the associated wildlife communities. Here, we synthesize our understanding of how changing fire regimes will affect (short- and long-term effects) boreal landscapes and the implications of these changes for terrestrial wildlife using the extreme burning conditions in subarctic forests of the Northwest Territories, Canada in 2023 as a case study. Our goals were to a) evaluate the impacts of the changing fire regime on habitat composition and/or structure in the short- and longer-term; and b) assess the potential responses of terrestrial vertebrates to these changes based on our understanding of their reliance on key aspects of habitat composition and/or structure. We describe impacts of changes in individual aspects of the fire regime on wildlife taxa but, overall, at the landscape level, changes in fire regime are altering the composition and structure of boreal forests, which will drive decreases in taxa requiring mature conifer forest and post-fire structural complexity while favoring early seral species or those that prefer broadleaf forests or open habitats. This review offers us a range of possibilities about the future landcover and northern wildlife communities under changing wildfires, however uncertainties about feedbacks and future conditions of this vast, diverse, and remote landscape present the greatest challenges for forecasting and other supports for adaptation planning.
Introduction
The boreal biome comprises ∼30% of the global forest area and is one of the most intact forest ecosystems, occupying high latitude, low productivity regions of Canada, Russia and Alaska (Brandt et al., 2013). This vast area contains 30%–40% of terrestrial carbon stocks and more freshwater than any other biome (reviewed in Gauthier et al., 2015). Human population densities are low meaning that ecological processes and natural disturbances operate across broad spatial scales, and the biodiversity of the biome is largely intact, supporting northern livelihoods and a wide range of ecosystem services (reviewed in Turetsky et al., 2017). Large wildfires are the primary natural disturbance in this system and have historically burned every approximately 50–500 years, depending on the region (Coops et al., 2018; de Groot et al., 2013; Johnstone et al., 2010a). Wildfires create heterogeneity at both the landscape level and within individual fire perimeters (Bergeron et al., 2004; Coogan et al., 2021; Johnson et al., 1998; Weir et al., 2000), which promotes biodiversity (Steel et al., 2024; Hekkala et al., 2023). This heterogeneity is owing to variation in fire behaviour and a wide range of factors that impact successional trajectories and maintain a mosaic of successional stages and forest composition (Perera et al., 2009; Perera and Buse, 2014; Day et al., 2020). Wildfires are also well known to influence important ecosystem functions such as biogeochemical cycling (Eckdahl et al., 2023; Eckdahl et al., 2024; Granath et al., 2021; Pellegrini et al., 2018; Walker et al., 2019; Wei et al., 2018), water quality and quantity (Carvalho-Santos et al., 2019; Granath et al., 2021; Robinne et al., 2020; Santos et al., 2015; Smith et al., 2011), and land surface - atmosphere energy exchanges (French et al., 2016; Zhao et al., 2024) among others.
High latitudes are warming approximately four times faster than the global mean rate (Chylek et al., 2022; Rantanen et al., 2022). Trends in precipitation are much less clear with studies reporting small increases which are often non-significant trends (Zhang et al., 2000; Schetselaar et al., 2023). In addition, small streams in the subarctic may be shifting towards a winter-flow dominated hydrological regime where the majority of streamflow occurs in winter rather than after spring snowmelt, reducing surface moisture during the growing season (Spence et al., 2025). Taken together for the growing season, the increase in evapotranspiration is greater than the increase in precipitation meaning that many high latitude regions are experiencing increasing moisture deficits (Mirabel et al., 2023; Wang et al., 2014; Zhang et al., 2019). This has important consequences for fire regimes (i.e., the characteristics of the fires that have occurred over a long period of time, including mean frequency, size, intensity, severity, ignition source, seasonality, type [ground, surface, or crown], and the variation around these means (Whelan, 1995). Warmer, drier conditions increase landscape flammability (Jain et al., 2024; Whitman et al., 2024) and the frequency of extreme fire weather (Wang et al., 2015), increasing fire activity and promoting more extreme burning conditions. Effective fire suppression and removal of Indigenous burning practices have simultaneously acted to lengthen fire return intervals and promote fuel build-up (e.g., Chavardès et al., 2018; Christianson et al., 2022; Parisien et al., 2023a). Coupled with greater amounts and flammability of fuels is an increase in lightning activity owing to increasing convective periods with climate change, meaning greater ignition potential (Veraverbeke et al., 2017).
Due to these changes, we have seen an increase in the average annual area burned in boreal forests over the last several decades (Coops et al., 2018; Soja et al., 2007; Walker et al., 2020). Notably, extreme wildfire years where area burned far exceeds the long-term average are becoming increasingly common and there has been clear attribution to climate change in regions where it has been investigated (Beverly and Schroeder, 2024; Boulanger et al., 2024; Parisien et al., 2023b). The hot, droughty conditions that support large fire years also facilitate other attributes of a more extreme fire regime such as wind events that further increase flammability and carry fire (Tavakol et al., 2020), longer fire seasons including overwinter smouldering (Hanes et al., 2019; Jain et al., 2024; Scholten et al., 2021), burning of parts of the landscape including young forests and peatlands/wetlands that would have historically served as fuel breaks or fire refugia (Kuntzemann et al., 2023; Whitman et al., 2024), and more severe combustion of aboveground biomass and the soil organic layer that serves as a critical carbon sink in the boreal biome and affects post-fire establishment conditions (Walker et al., 2020; 2019; Baltzer et al., 2021). Climate projections for the coming decades suggest that this increasing trend of fire conducive weather will continue (Wang et al., 2020; Wotton et al., 2017). Such changes in fire activity coupled with ongoing warming are expected to alter the successional trajectory of forests with implications for biodiversity and ecosystem function.
The summer of 2023 was the first time that global average summer temperatures exceeded 1.5 °C above pre-industrial average temperatures for 1850–1900. The United Nations Convention on Climate Change’s Paris Agreement stipulates that limiting warming to 1.5 °C should be our collective goal as we aim to reduce emissions and slow global warming; a goal that is now considered unachievable without greatly increased ambition and much more rapid implementation (Ou et al., 2021). Global mean summer temperatures of 1.5 °C above pre-industrial levels translated to much greater increases at high latitudes; for example, Canada was, on average, 2.2 °C above pre-industrial levels. During this exceptionally hot and dry summer, out-of-control wildfires burned across Canada leading to a burned area of more than 15 M ha, seven times the 1986–2022 average and more than double the previous record (Jain et al., 2024); this area was comparable to the record breaking 2021 wildfires in Siberia where more than 17 M ha of land burned (Kharuk et al., 2022). Most burning occurred in boreal forests and a range of fire behaviours and outcomes were observed that are novel disturbance conditions relative to historic burning (Jain et al., 2024; Turner and Seidl, 2023). Half of all individual fires were >100 ha and there were >800 “large fires” (>200 ha; Jain et al., 2024). The average number of large fires is 320 (1986–2022) and before 2023 only 1 year on record had >600 large fires (1989; Jain et al., 2024). The fire season started early in the spring and ended late in the fall (Jain et al., 2024) with hundreds of fires continuing to smoulder through winter 2024. Fire spread rate was unprecedented in many areas. For example, the largest fire complex in Canada in 2023 was in the subarctic boreal forests of Northwest Territories (NWT) (Figure 1), which saw a maximum daily growth rate of nearly 100,000 ha (Jain et al., 2024), this is extraordinary when compared to the Canadian average value of 571 ha/day (2001–2021; Barber et al., 2024). The extreme fire weather conditions (Fire Weather Index exceeding the 95th percentile for 1990–2021 at a location; Jain et al., 2024) in many parts of Canada resulted in the reburning of young forests (<20 years since last fire), wetlands, and peatlands that would historically serve as natural fuel breaks (Whitman et al., 2024). Extreme fires have been increasing disproportionately in boreal North America as evidenced by the 2023 fire season, and this is expected to continue with ongoing human-caused climate warming (Cunningham et al., 2024). The 2023 fire season was extraordinary in its early start and late finish. Fires were still burning very actively into the fall with almost 440,000 ha burning across Canada in a single day in September and active fires and fire management well into October (Jain et al., 2024). Effectively, the fire season lasted up to 7 months in some places and in western Canada the fire season practically never ended with more than 150 overwintering or ‘zombie’ fires smouldering through the winter months in British Columbia, Alberta, and NWT, leading to an early and intense start to the 2024 fire season (Baltzer et al., 2025).
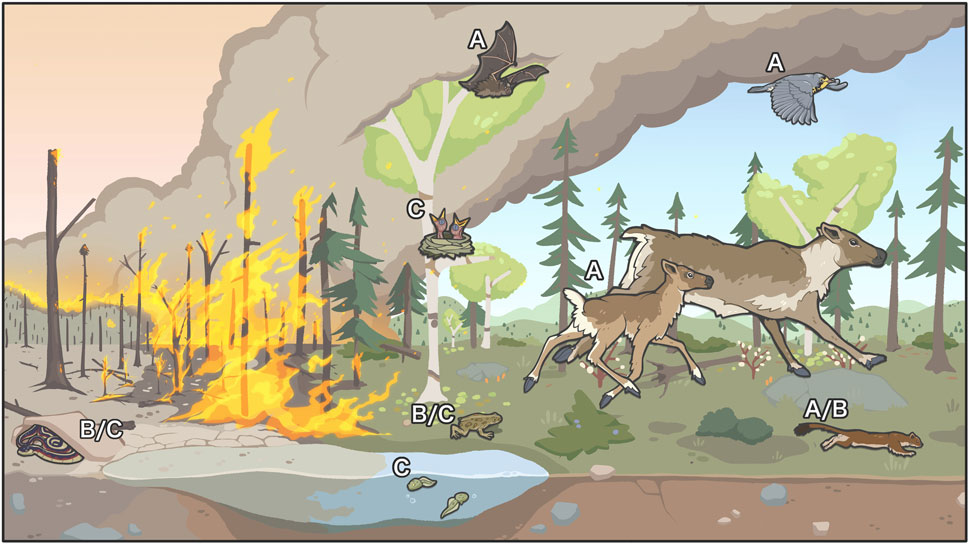
Figure 1. Illustration of examples of the acute effects of wildfire on vertebrate wildlife species including (A) forcing mobile species to flee: little brown myotis (Myotis lucifugus), boreal caribou (Rangifer tarandus caribou), Canada warbler (Cardellina canadensis), short-tailed weasel (Mustela erminea); (B) forcing burrowing/denning animals to seek shelter belowground to try to avoid mortality: Red-sided garter snake (Thamnophis sirtalis parietalis), short-tailed weasel, western toad (Anaxyrus boreas); (C) mortality of less mobile species: Red-sided garter snake, western toad, other aquatic species in land-locked water bodies, slow-moving species, nestling birds, etc. The species that avoid direct mortality are expected to experience physiological stress because of smoke inhalation, physical exertion, etc.
There is mounting evidence that current fire regimes are becoming misaligned with the plant communities inhabiting boreal regions with wide-ranging implications (Baltzer et al., 2021; Johnstone et al., 2016). After wildfire, various biological and structural legacies remain that shape the response of the system to current and future disturbances. These can include seeds and other reproductive structures, woody biomass, and residual organic soils. Many of the changes in fire regime described above have been shown to remove or alter these legacies, with implications for post-fire forest successional trajectory (reviewed in Johnstone et al., 2016). Examples include changes in: forest composition (including recruitment failure) (Baltzer et al., 2021; Burrell et al., 2022; Hayes and Buma, 2021; Johnstone et al., 2016; Whitman E. et al., 2019); mode of ground vegetation recovery with implications for plant community composition (Day et al., 2020; Hollingsworth et al., 2013); microbial community structure and function which affect biogeochemical processes with direct impacts on plant community composition and function (Day et al., 2019; Whitman E.et al., 2019; Eckdahl et al., 2023; Eckdahl et al., 2024); boreal carbon sequestration potential including large carbon emissions and losses of legacy carbon (Veraverbeke et al., 2021; Walker et al., 2019; 2018); and rates of permafrost thaw, which can further amplify vegetation-changes and modify other ecosystem functions (Bill et al., 2023; Gibson et al., 2018; Holloway et al., 2020). Across jurisdictions, there are concerns about the biodiversity and self-sustainability of wildlife populations, including species at risk, given wildfire-induced changes in wildlife habitat. More specifically, for Indigenous communities in northern boreal regions, such wildfire-induced changes affect the diversity, abundance, and access to “country” or subsistence food resources (e.g., wildlife) by altering habitat availability, removing traditional trails, and making land access more dangerous or less predictable (Spring et al., 2019). Understanding and forecasting these changes have been identified as priorities for many northern communities and as such forms the focus of this review.
In this review, we synthesize knowledge of how novel fire regimes are changing North American boreal forests and highlight known and anticipated effects of these changes on landcover (i.e., wildlife habitat) and the terrestrial wildlife that rely on these lands using examples of species that occur in NWT. More specifically, we consider how changes to four components of the fire regime (fire size and area burned; burn severity; short-interval reburning; and longer fire seasons) may impact vegetation structure and composition over the short-term (<10 years post-fire) and longer-term (30–40 years post-fire) and how these changes may differentially affect different groups of wildlife taxa. We use the 2023 burning conditions in NWT as a case study because it was the subarctic boreal region in Canada that was most severely impacted. Furthermore, the 2023 fires in NWT came on the heels of the second largest recorded fire year that occurred in the same area only 9 years earlier, in 2014 [2.85 M ha burned; Walker et al., 2018], leading to effects of compounded extreme fire years. This is not intended to be an exhaustive review of all wildlife taxa but to provide a survey of our understanding of responses across a range of trophic levels and functional groups of the ∼300 terrestrial vertebrate taxa in boreal NWT (Working Group on General Status of NWT Species, 2021). Few studies have tackled the impacts of individual components of fire regime on wildlife taxa (e.g., reviewed in Forney and Peacock, 2024; Volkmann and Hodges, 2024) so our goals were to a) evaluate the impacts of these changing components of the fire regime on habitat composition and/or structure in the short- and longer-term; and b) assess the potential responses of terrestrial vertebrates to these changes based on our understanding of their reliance on key aspects of habitat composition and/or structure. We focus predominantly on species of special conservation concern (species at risk, migratory birds, culturally important species) owing to greater availability of information. We did not focus on predators owing to disproportionately fewer studies evaluating predator responses to fire and because the response of predators to fire tends to be an indirect response to prey availability rather than the direct effects of changes in forest composition and structure that we are focused on (reviewed in Forney and Peacock, 2024; Volkmann and Hodges, 2024). We also identify key knowledge gaps to help guide wildlife–wildfire research in NWT and elsewhere in northern Canada. We end with ideas about wildlife–wildfire research and monitoring priorities and how we can best use this knowledge to support adaptation planning in jurisdictions and communities in the boreal forest. Canada’s 2023 wildfire season gave us a glimpse of the future in a warming, fierier boreal biome. It is imperative for adaptation planning that we work to understand and forecast these impacts.
Acute responses to wildfire
The immediate impacts of fire on wildlife populations includes direct mortality, which can be particularly important for small mammals, amphibians, juvenile birds, and reptiles that have limited capacity to flee approaching fire (Figure 1), however there are many unknowns. We will not deal comprehensively with acute fire impacts as these constitute a variety of physiological and behavioural responses beyond the scope of this review (reviewed in Sanderfoot et al., 2022). However, below we will touch briefly on some taxa that face heightened risk of direct mortality from fire in NWT.
Small mammals (mice, voles, shrews, squirrels, and hares) found in NWT forests have co-evolved with wildfire and some species of small mammals (e.g., deer mice [Peromyscus maniculatus], voles, and red squirrels [Tamiasciurus hudsonicus]) are likely able to survive the immediate impacts of wildfires by using burrows, logs, or crevices under rocks to shelter from passing fires (Banks et al., 2011; Hale et al., 2022; Shaw et al., 2021; Sutherland and Dickman, 1999) or survive within unburned residuals within the fire perimeter (Bond, 2015). Other small mammal species that nest in tree cavities (e.g., northern flying squirrel [Glaucomys sabrinus]) and birds nesting on a wide range of substrates may be more susceptible to mortality and loss of young during fires as these aboveground structures are readily consumed. Even though snowshoe hare (Lepus americanus) leverets are precocious at birth, they may be particularly susceptible to mortality during a fire event, as they are left unattended in aboveground, shallow, fur-lined bowls and may not be mobile enough to escape a fire (Aldous, 1937; O’Donoghue and Bergman; Rongstad and Tester, 1971). Although it has been suggested that bats could suffer some direct mortality during fires, little is known about the magnitude of this mortality (Loeb and Blakey, 2021). Typically, bats should be capable of leaving their roosts and flying away from an approaching fire although non-volant bat pups may be more susceptible (Dickinson et al., 2010). Similar to small mammals, snakes can take refuge in wetland habitats or belowground (in crevices, hollows, or rodent burrows) which can allow them to survive a fire but does not always provide complete protection (Cross, 2015; Russell et al., 1999; Smith et al., 2001); indeed, high levels of fire-induced mortality have been documented in snake populations (e.g., Erwin and Stasiak, 1979; Lyet et al., 2009). Fires can directly kill many amphibians while they are in terrestrial habitats (e.g., Bailey et al., 2025) and even while they are in burrows or other refuges, especially if the fire is intense (Beranek et al., 2023; Mahony et al., 2022). Combustion of downed woody debris and fires that burn deeper into the soil organic layer and tree root system will likely result in higher immediate mortality of ground-dwelling small mammal, amphibian, and snake species that have lower mobility and typically use protective features (e.g., burrows, hollows in logs) to survive fires in situ (Jager et al., 2021). In short, the ability and instinct to flee is a major factor determining whether wildfire will lead to mortality events in a population (reviewed comprehensively in Sanderfoot et al., 2022).
Area burned and fire size
A key component of any disturbance is its extent. In the case of fire, this includes total area burned in a region as well as the size distribution of individual fires. Both total area burned and individual fire sizes were unprecedented in 2023 (Jain et al., 2024). In NWT alone, more than 4 M ha of forested land burned, almost a third of total burned area in Canada in 2023 (Jain et al., 2024). NWT was also home to the largest fire complex in Canada since the 1950s, exceeding 1 M ha (Jain et al., 2024; Figure 2). This was the largest fire season by far for both Canada and NWT since records began in 1965.
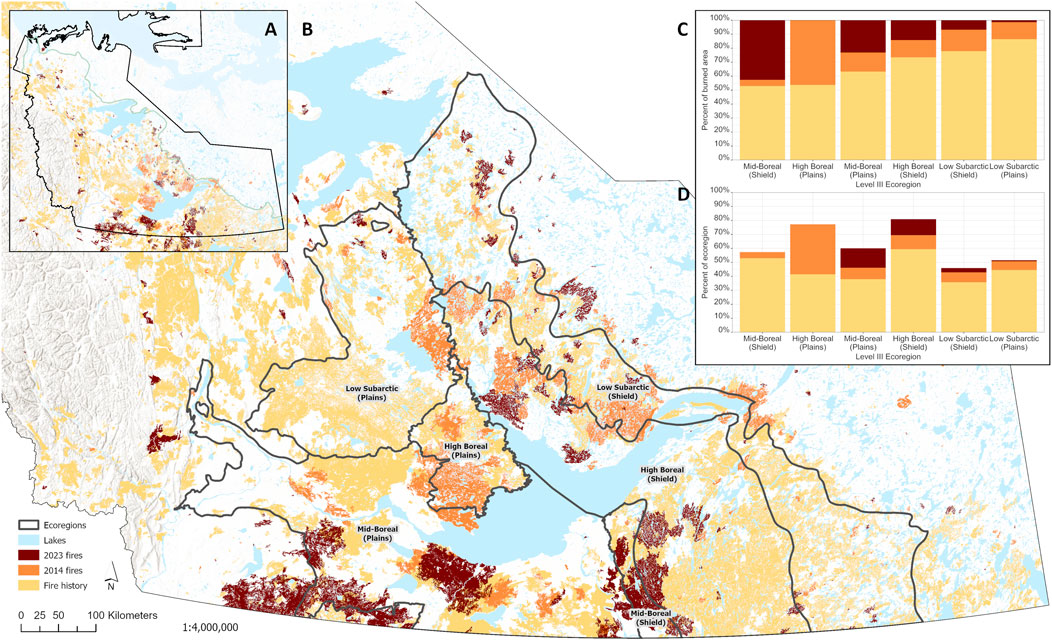
Figure 2. Extent of two consecutive mega-fire years (2014, 2023) in the Great Slave Lake region of Northwest Territories (NWT). Fires on a NWT-wide scale (A) and within the southern NWT where Level III Ecoregions are also demarcated (B). Ecoregion delineations are from Ecosystem Classification Group (2007 [rev. 2009]; 2008; 2010; 2012). Combined, these fire seasons burned approximately 7 M ha in the same region. The percent contribution of 2014 and 2023 wildfires to total burned area (C) and total area of Level III Ecoregions (D) is provided.
Large boreal wildfires in and of themselves are not necessarily problematic for wildlife. Large fire complexes burn over the course of many days to weeks and across large areas. Consequently, burning conditions vary owing to temporal changes in fire weather and spatial changes in fuel conditions. This means that the fire intensity and spread rate can vary substantially as the fire grows, leading to tremendous heterogeneity in residual vegetation and post-fire vegetation structure within large fire complexes (reviewed in Burton et al., 2008). These variable conditions are thought to emulate pre-suppression fire regimes to which the landscape may be better adapted (reviewed in Stephens et al., 2014). Variation in burn severity within large fire complexes may increase wildlife diversity because greater heterogeneity in the post-fire composition and structure provides a more diverse set of habitat conditions (reviewed in Burton et al., 2008; Jones and Tingley, 2022). Furthermore, large fires tend to have more complex edge structure which has been shown to be important for wildlife (reviewed in Burton et al., 2008). However, much of this will depend on the size, severity, and frequency of burning in these complexes. For example, large, high severity burns (see definition in burn severity section) will promote species that recover rapidly post-fire and can in fact reduce landscape level heterogeneity with associated negative consequences for the diversity of wildlife habitat (reviewed in Steel et al., 2024).
Short-term responses to larger fires
For species that are reliant on mature forest conditions, larger fires have negative short-term effects through reduction in available habitat. In NWT, the two largest fire seasons on record (2014 and 2023; 2.9 M and 4 M ha, respectively; Walker et al., 2018; Jain et al., 2024) impacted the same general region around Great Slave Lake in southern NWT resulting in vast tracts of disturbed forest accumulating across a 9-year period (Figure 2). As the climate warms and dries, we expect such repeated instances of extreme fire years like we have seen in NWT, but the extent of the impact on mature forest specialists is not yet fully known owing to the relative novelty of these large events. For example, boreal caribou (Rangifer tarandus caribou) are particularly reliant on mature forests for access to arboreal and terrestrial lichens which are their predominant winter food (Schaefer and Pruitt, 1991; Thompson et al., 2015). Reindeer lichens (Cladonia subgenus Cladina) are readily consumed by fire and, due to their extremely slow growth rates (∼4.9 mm/yr; McMullin and Rapai, 2020), can take several decades to recover to pre-fire biomass found in mature forest stands (McMullin et al., 2011; Silva et al., 2019; Greuel et al., 2021). We expect that increasing occurrence of both larger individual fires and higher total area burned will lead to winter habitat loss (Palm et al., 2022) and could lead to boreal caribou congregating at higher densities in small patches of remaining mature forest during winter. While boreal caribou avoid burns most strongly during winter (Konkolics et al., 2021), they may actually select for recent burns during calving (Silva et al., 2020) or summer (DeMars et al. unpublished data) likely due to the greater availability of protein-rich forage provided by early-successional vegetation that helps female caribou meet the nutritional demands of lactation (Denryter et al., 2017). When boreal caribou do use areas within burns, in all seasons they select areas closer to burn perimeters (Palm et al., 2022). Thus, a very large individual fire event may lead to greater habitat loss for caribou than several smaller fires amounting to an equivalent area due to edge effects (i.e., their avoidance of areas deeper within burns).
Similarly, there are many mature forest specialists in the landbird community that are vulnerable to widespread burning (e.g., Canada Warbler [Cardellina canadensis], Brown Creeper [Certhia americana], and Evening Grosbeak [Coccothraustes vespertinus]; Gillihan and Byers, 2020; Poulin et al., 2020; Reitsma et al., 2020; see also text in short-interval reburn section below). Alternatively, landbird species associated with early seral stages (e.g., Alder Flycatcher [Empidonax alnorum] and Dark-eyed Junco [Junco hyemalis]) or habitat generalists (e.g., American Robin [Turdus migratorius]) would thrive or not show a numerical response to larger fire events (Schieck and Song, 2006). Recently, Lane-Shaw et al. (unpublished data) used predictions from species distribution models for 144 landbird species generated from the Boreal Avian Modelling Center (Stralberg, et al., 2025) to quantify potential changes in abundance of these species breeding in mature forests of NWT following the 2023 fire season. Preliminary results suggest that the most negatively impacted species include the mature forest obligates Rusty Blackbird (Euphagus Carolinus), Ruby-crowned Kinglet (Corthylio Calendula), Golden-crowned Kinglet (Regulus Satrapa), and Bay-Breasted Warbler (Setophaga Castanea). For these species, an estimated ∼20% of their populations breeding in mature forest experienced lost or altered breeding habitat in NWT. In total, ∼59,000 breeding males had habitat in mature forests (>80 years) that burned in 2023 across NWT.
Fire affects foraging opportunities for bats. Recently burned habitat often experiences a “pulse” of higher insect productivity (Lacki et al., 2009; Malison and Baxter, 2010). Fire also changes forest structure by reducing understory vegetation, making flying and foraging easier for bat species that are less tolerant of clutter (Blakey et al., 2019); this can increase activity of bat species such as the little brown myotis (Myotis lucifugus) in burned landscapes (Buchalski et al., 2013; Blakey et al., 2019; Low et al., 2024). In contrast, northern myotis (Myotis septentrionalis) is more dependent on forest cover (Henderson et al., 2008) and better adapted to foraging in densely vegetated environments (Norberg and Rayner, 1987; Ratcliffe and Dawson, 2003). As such, northern myotis is expected to be more be negatively impacted by larger burned areas compared to the little brown myotis (Blakey et al., 2019; Snider et al., 2013; Bosso et al., 2018; Kotliar et al., 2002; Azeria et al., 2011; Doty et al., 2023).
Despite the potential for widespread mortality of small, ground dwelling mammals during a large fire event, their high reproductive capacity and ability for some individuals to survive the fire in situ, means that they can recolonize burned areas relatively quickly (Hale et al., 2022). In boreal North America, deer mice tend to be the most abundant species immediately post-fire, perhaps owing to their omnivorous diet, followed by red-backed voles (Clethrionomys gapperi) which may repopulate burns within the first 3 years (Olson et al., 2003; Simon et al., 1998 - reviewed in Fisher and Wilkinson, 2005; Zwolak and Foresman, 2007). Meadow voles (Microtus pennsylvanicus) and meadow jumping mice (Zapus hudsonicus) may also be more abundant than red-backed voles in the short-term due to their association with grassy cover that may recover quickly post-fire (Fisher and Wilkinson, 2005). Marten (Martes spp.) and weasels (Mustela spp.), which primarily prey on small mammals, may benefit from edge habitat created by fires that locally increase small mammal populations; however, large homogenous fires may decrease edge habitat and habitat suitability for marten as they avoid areas of low canopy closure and may be reluctant to forage far from the forest edge (Volkman and Hodges, 2024). Red squirrels, conifer seed specialists that are more common in mature forests, may similarly forage within the periphery of recent burns (Fisher, 1999), but are unlikely to recolonize the center of large burns until conifer forests recover.
Reptiles such as red-sided garter snakes (Thamnophis sirtalis parietalis) may experience indirect impacts of large boreal wildfires linked to post-fire changes in predator-prey dynamics and habitat suitability (Rochester et al., 2010). As species that rely on habitat structure to avoid predation, they may experience higher predation rates and lower survival in large, recent burns (Doherty et al., 2022; Howey et al., 2016; Russell et al., 1999; Webb and Shine, 2008; Wilgers and Horne, 2007). Reduced cover (i.e., removal of shrubs, woody debris and the litter layer, and opening of the forest canopy) can make it easier for avian predators to detect them (Doherty et al., 2022; Wilgers and Horne, 2007). Reptiles can benefit from the opening of forest canopies or dense shrub thickets that increase surface temperatures (Bury, 2004; Howey et al., 2016; Russell et al., 1999); however, widespread removal of vegetation, leaf litter and woody material can reduce the thermal heterogeneity that they require (Howey et al., 2016; Smith et al., 2001).
There are many impacts of fire on amphibian species, but those most impacted by large fires will likely be those reliant on cool, moist microhabitats and that use ground cover consumed by fire (e.g., leaf litter) (Rochester et al., 2010; Russell et al., 1999), such as the wood frog (Rana sylvatica) (Bailey et al., 2025; Constible et al., 2001). Reduced habitat structure, lower soil moisture and more variable soil temperatures in their terrestrial habitats present amphibians with increased risk of overheating, desiccation, and predation, reduced availability of habitat refugia and invertebrate prey, and barriers to movement (Browne et al., 2009; Bury, 2004; Cline and Hunter Jr, 2016; Hossack et al., 2013; Rochester et al., 2010; Russell et al., 1999; Schurbon and Fauth, 2003). Amphibians are also affected by changes to their aquatic habitats post-fire and watersheds that are completely combusted in large burns will see the biggest changes. Reductions in dissolved oxygen seem consistently negative for boreal amphibians whereas other chemical changes have been shown to negatively affect wood frog and boreal chorus frog (Pseudacris maculata) abundance but are better tolerated by western toad (Anaxyrus boreas; Jager et al., 2021; Browne et al., 2009). Increased solar radiation and contaminants (e.g., wildfire ash) in their aquatic habitats have negative effects on amphibians (Bancroft et al., 2008; Gomez Isaza et al., 2022; McDonald et al., 2018; Muñoz et al., 2019; Xu et al., 2024). Loss of vegetation surrounding wetlands can reduce protection from temperature extremes and predators, with negative effects on amphibian survival (Beranek et al., 2022) and reproductive success (Muñoz et al., 2019). Wetland habitats also experience changes in productivity, water temperature, and hydroperiods that affect amphibians (dos Anjos et al., 2021; Gomez Isaza et al., 2022; Schurbon and Fauth, 2003). However, certain amphibians are more resilient to large fire extents and might even show positive population responses (reviewed by dos Anjos et al., 2021). For example, western toads exhibit rapid tadpole development and are habitat generalists able to reproduce in almost any water body, which facilitates colonization of burned habitat (dos Anjos et al., 2021) but does not necessarily result in longer-term increases (Hossack et al., 2013).
Long-term responses to larger fires
Very large fires can alter post-fire forest composition through their effects on dispersal distances required for establishment. Depending on the pre-fire forest composition and combustion severity of canopy seed sources, soil seedbanks, and below/near-ground budding structures, on-site reproductive sources may be insufficient when distance to an unburned edge is large. This can lead to lagged recovery, post-fire forest compositional changes (e.g., conifer to deciduous), poor recruitment leading to forest structural changes (Girard et al., 2008), or recruitment failure (Johnstone et al., 2010b). For example, white spruce (Picea glauca) is a non-serotinous conifer; white spruce stands within a large burn complex may fail to self-replace owing to long distances to the nearest seed source coupled with relatively low dispersal distances of white spruce seeds (Dobbs, 1976). In contrast, trembling aspen (Populus tremuloides) seeds can travel as much as 10 km from the nearest seed source (Turner et al., 2003) and aspen produces seed prolifically in the summer following a large wildfire year (J. Baltzer, personal observation; Landhäusser et al., 2019). As such, large burn complexes, especially in conifer dominated areas, may be particularly susceptible to forest compositional change (Johnstone et al., 2010b). Although a few large fire complexes may be beneficial in supporting landscape heterogeneity, where many large fire complexes occur in proximity as has occurred in southern NWT, this change can create challenges. Changes in overstory composition alter the ground vegetation community composition (e.g., Day et al., 2020; Greuel et al., 2021) with implications for wildlife forage availability (Jorgensen et al., 2023) and habitat use (Palm et al., 2022) (Figure 3).
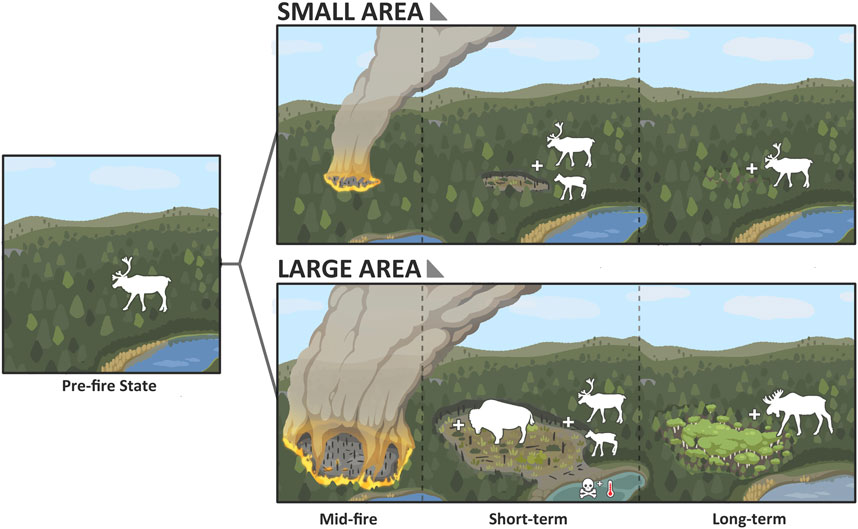
Figure 3. Illustration of an example of the effects of burn area on forest succession and some boreal wildlife. Note, plus signs denote habitat selection by the selected large mammal species, not a population response. Smaller burn areas in coniferous forest are more likely to be revegetated by the cones of surrounding trees, thus returning to a structure and composition similar to pre-fire conditions and in a shorter time frame. Larger burns are likely to experience compositional change, favouring long-distance dispersers, such as graminoids and broadleaf deciduous trees (e.g., Populus tremuloides). Early successional vegetation post-fire may provide boreal caribou (Rangifer tarandus caribou) with improved nutrition during calving. Conifer forest conversion to grass/shrubland or deciduous forest, will favour species including bison (Bison bison athabascae) and moose (Alces alces), whereas mature conifer forest specialists like boreal caribou will avoid these areas. Larger burn areas may also have profound impacts on adjacent aquatic environments, altering the aquatic species community. For example, large fires may alter water chemistry and thermal conditions of adjacent water bodies more profoundly.
For some wildlife species, changes in forest composition from conifer to deciduous dominance will be beneficial. For example, moose (Alces alces) occur throughout boreal North America and consume a variety of forage species including coniferous and deciduous trees and shrubs, graminoids, forbs, and aquatic plants (Joyal and Scherrer, 1978; Jung et al., 2018; Timmermann and McNicol, 1988). However, moose generally prefer deciduous trees and shrubs, notably willow (Salix spp.), birch (Betula spp.), and poplar (especially trembling aspen) which are commonly a dominant component of moose diet throughout the year (Risenhoover, 1989; Shively et al., 2019). For other wildlife species, such changes will be detrimental. For example, landbird species associated with mature conifer forests, such as Olive-sided Flycatcher (Contopus cooperi; Altman and Sallabanks, 2020), Canada Jay (Perisoreus canadensis; Strickland and Ouellet, 2020), Boreal Chickadee (Poecile hudsonicus; Ficken et al., 2020) and others are expected to decline in abundance as forests transition from conifer to deciduous. Conversion to deciduous forests would also likely be detrimental to boreal caribou as they avoid most age classes of deciduous stands relative to other landcover types from late-fall to late-winter (DeMars et al. unpublished data). However, species associated with mature deciduous or mixedwood forests such as the Warbling Vireo (Vireo Gilvus), Magnolia Warbler (Setophaga Magnolia), and Ovenbird (Seiurus Aurocapilla) will likely thrive for some time (see Short-interval reburning) because of such changes in forest composition (Scheick and Song 2006). Knaggs et al. (unpublished data) showed that, in NWT, early seral stage or habitat generalist species tend to be at a higher probability of occurrence between 3 and 10 years post-fire (e.g., Chipping Sparrow, Spizella passerina, White-throated sparrow, Zonotrichia albicollis, American Robin) and 11–30 years (e.g., Alder Flycatcher, Lincoln Sparrow, Melospiza lincolnii, White-crowned Sparrow, Zonotrichia leucophrys), while other species associated more with mature forests started peaking 11–30 (e.g., Hermit Thrush, Catharus guttatus), 31–50 (e.g., Swainson’s Thrush, Catharus ustulatus), or >51 years post-fire (e.g., Yellow-rumped Warbler, Setophaga coronata). Other bird species like the Ruffed Grouse (Bonasa umbellus) and Willow Ptarmigan (Lagopus lagopus) might be more abundant in recent burns due to increase in forage availability (Cringan, 1958; Rusch et al., 2020; Snow, 1996; Weeden, 1963), while Spruce Grouse (Canachites canadensis) would be more abundant in mature conifer forests (Cringan, 1958; Ellison, 1975).
While short-term post-fire changes in small mammals communities are fairly well documented (reviewed in Fisher and Wilkinson, 2005), the longer-term implications of larger fires are less clear. The combined effect of fire size and homogeneity of the burn with respect to fire severity is likely a more important driver of longer-term recovery of small mammal populations than fire size alone. Generally speaking, the abundance of small mammals increases with time after fire (Griffiths and Brook, 2014), and the relative abundance of different small mammal species within the community shifts with forest succession (Fisher and Wilkinson, 2005). For example, red-backed voles begin to replace deer mice as the most common species in the small mammal community once shrubby and herbaceous vegetation replaces grasses (Fisher and Wilkinson, 2005). American marten (Martes americana) used 21-year-old burns in NWT that had abundant standing snags and deadfall (Latour et al., 1994), suggesting they will make use of recent burns provided there is sufficient residual structure to provide subnivean access to prey and/or shelter. Populations of red squirrels and northern flying squirrels will take longer to recover within large burns as both are reliant on habitat features common to mature conifer forests such an abundant supply of conifer seeds (red squirrels), large live and dead trees used for dens, and as fungal and lichen food sources (flying squirrels) (Fisher and Wilkinson, 2005).
The extent of burns can affect patterns of amphibian occupancy in the longer term, through increased distance to source populations (Bailey et al., 2025; Hossack et al., 2013) and reduced habitat connectivity that impedes movement (Beranek et al., 2022; Cline and Hunter Jr, 2016). These effects would be influenced by heterogeneity and the existence of habitat refugia within large burn perimeters (Mahony et al., 2022; Schurbon and Fauth, 2003). For wood frogs, post-fire breeding persistence in wetlands can be high if unburned habitat is available nearby, but probability of persistence decreases when a greater proportion of the surrounding terrestrial habitat is burned, and it could be several years or decades before wood frogs return to breed in these areas (Bailey et al., 2025). A similar relationship was found for Columbia spotted frogs (Rana luteiventris) with effects on occupancy 7–21 years after wildfire (Hossack et al., 2013). Large, intense wildfires can also negatively affect persistence of amphibian species and communities across a broad geographic area and periods of time, and that even species with large populations and/or ranges can be affected (Beranek et al., 2023).
For snakes, habitat refugia (i.e., unburned areas) are important for persistence after fire and re-establishment in the longer term (Robinson et al., 2013; Beaupre and Douglas, 2012). Larger fires could reduce the number and proximity of habitat refugia on the landscape in boreal systems (McKenzie et al., 2004; Mackey et al., 2021), potentially limiting the ability of red-sided garter snakes to recolonize after fire (Santos et al., 2022), especially if large fires simultaneously affect many individuals and/or den sites, a situation that was observed in 2023 (J. Wilson, pers. obs.). Nevertheless, snake populations can persist in situ and be resilient to fire (Halstead et al., 2019; Maloney, 2024; Russell et al., 1999; Smith et al., 2001), even large fires (Santos et al., 2021).
In summary, responses to large burn areas or fire sizes vary considerably across taxa. Species reliant on large tracts of mature forest or nearby forest edges will fare poorly in the short- and longer-term as area burned increases and access to these habitats decreases. Many taxa are ambivalent to burned condition per se but have forest structural and compositional requirements for protective or thermal cover or other habitat attributes that will determine their response to the fire. Taxa preferring more open habitats or early post-fire vegetation types will fare well in the face of large burns. Longer-term land cover changes associated with large burns (i.e., conifer to broadleaf deciduous) will favour some species and disadvantage others, effectively altering the structure of terrestrial wildlife communities.
Burn severity
Burn severity characterizes the magnitude of the combustion of both aboveground (vegetation) and belowground (soil) organic matter (Keeley, 2009). Although annual burned area is increasing across the boreal biome with climate warming (Coops et al., 2018; Soja et al., 2007; Walker et al., 2020), the same trend in burn severity, as measured by dNBR (difference Normalized Burn Ratio; a remotely sensed measure of combustion severity comparing two time points) is not apparent (Guindon et al., 2021). This is consistent for NWT (Figure 4; Supplementary Table S2). Despite 2023 having the largest total area burned and largest total number of fires in recorded history (Figure 5), there is no evidence of elevated dNBR in 2023 compared to previous years and no relationship between annual area burned or number of fires and burn severity as measured by dNBR (Figure 4). As such, we do not expect a proportional increase in the effects of severe fire during large fire years. However, the total area experiencing high burn severity will be greater simply owing to the larger total burned area. Almost 50% of the total burned area experienced severe combustion in 2023 in NWT (Figure 4) meaning that there is nearly 2 M ha of severely burned land that will experience various short-term and longer-term changes. Post-fire NBR (Normalized Burn Ratio; a remotely sensed measure of combustion severity) and dNBR are closely related to canopy combustion severity and can reflect belowground combustion as well although this relationship is less clear (Soverel et al., 2010; Whitman E. et al., 2019). While dNBR demonstrated some potential as predictor for burn depth in a study conducted in the larch forests of Northeast Siberia (Delcourt et al., 2021), Fraser et al. (2017) reported stronger predictions for their char fraction (designed to derive the extent of burning of the surface organic layer) using post-fire NBR. The latter study also indicated how drone-based imagery may assist in clearly defining at fine spatial scales the extent of mineral soil where the entire surface organic layer had been consumed, potentially offering a bridging solution to overcome challenges with predicting the extent of exposed mineral soil directly through satellite imagery (Hudak et al., 2007). Taken together, our understanding of the relationship between satellite-derived metrics such as dNBR and belowground combustion in the boreal forests is still somewhat limited. Additionally, we lack clarity on whether and how dNBR applies to short interval reburns, which limits the inference of these findings and our ability to understand temporal trends in belowground combustion severity.
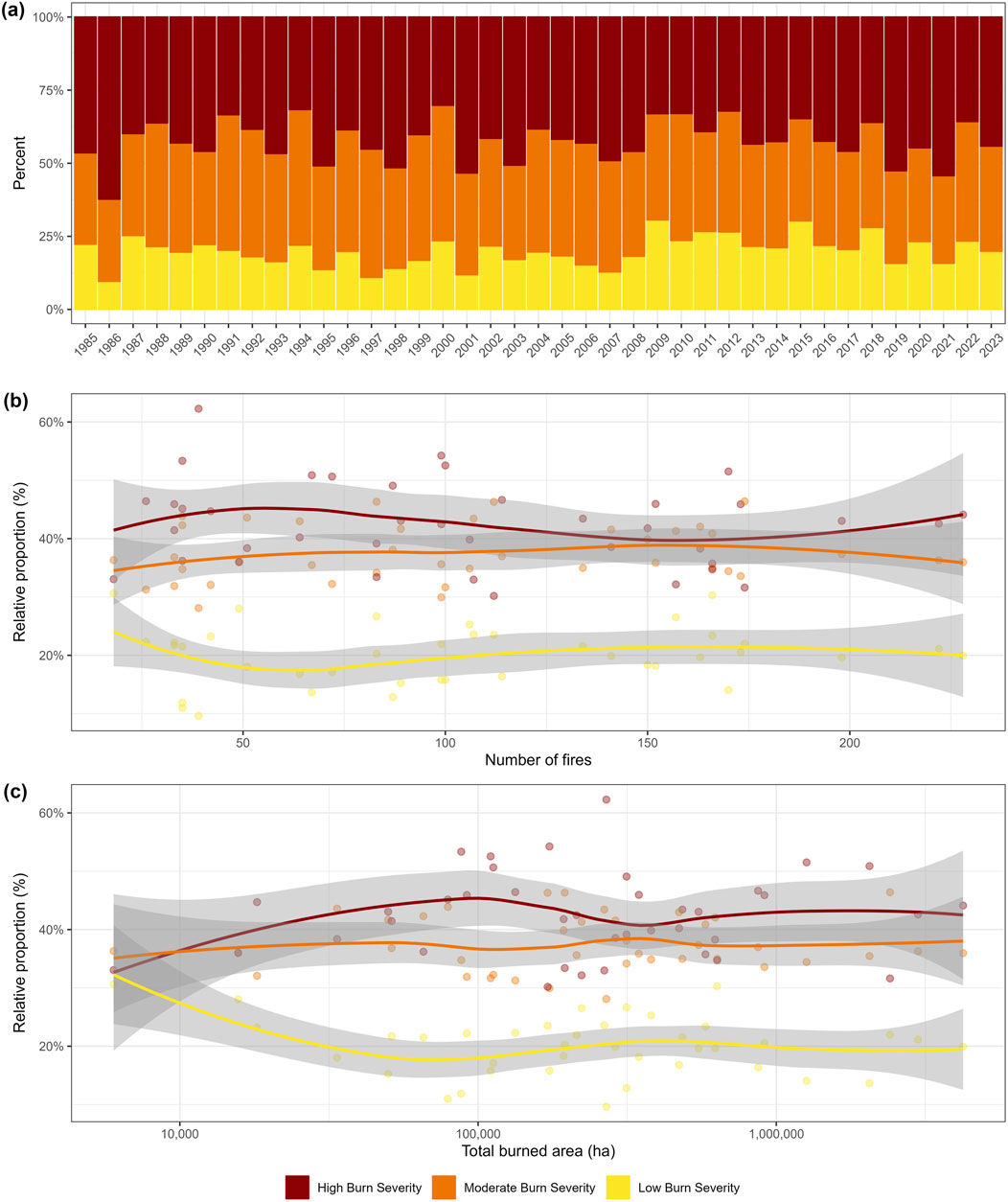
Figure 4. Summary of burn severity information for Northwest Territories including the percent of LandSat pixels experiencing high, medium and low severity burning (derived from dNBR thresholds; Hall et al., 2008) through the satellite record (a), as a function of total number of fires (b) and as a function of total burned area (c). There are no apparent trends in any of these measures and the statistics are presented in Supplementary Table S2. Methods for determination of dNBR and further details of this categorization are provided in Supplementary Table S1.
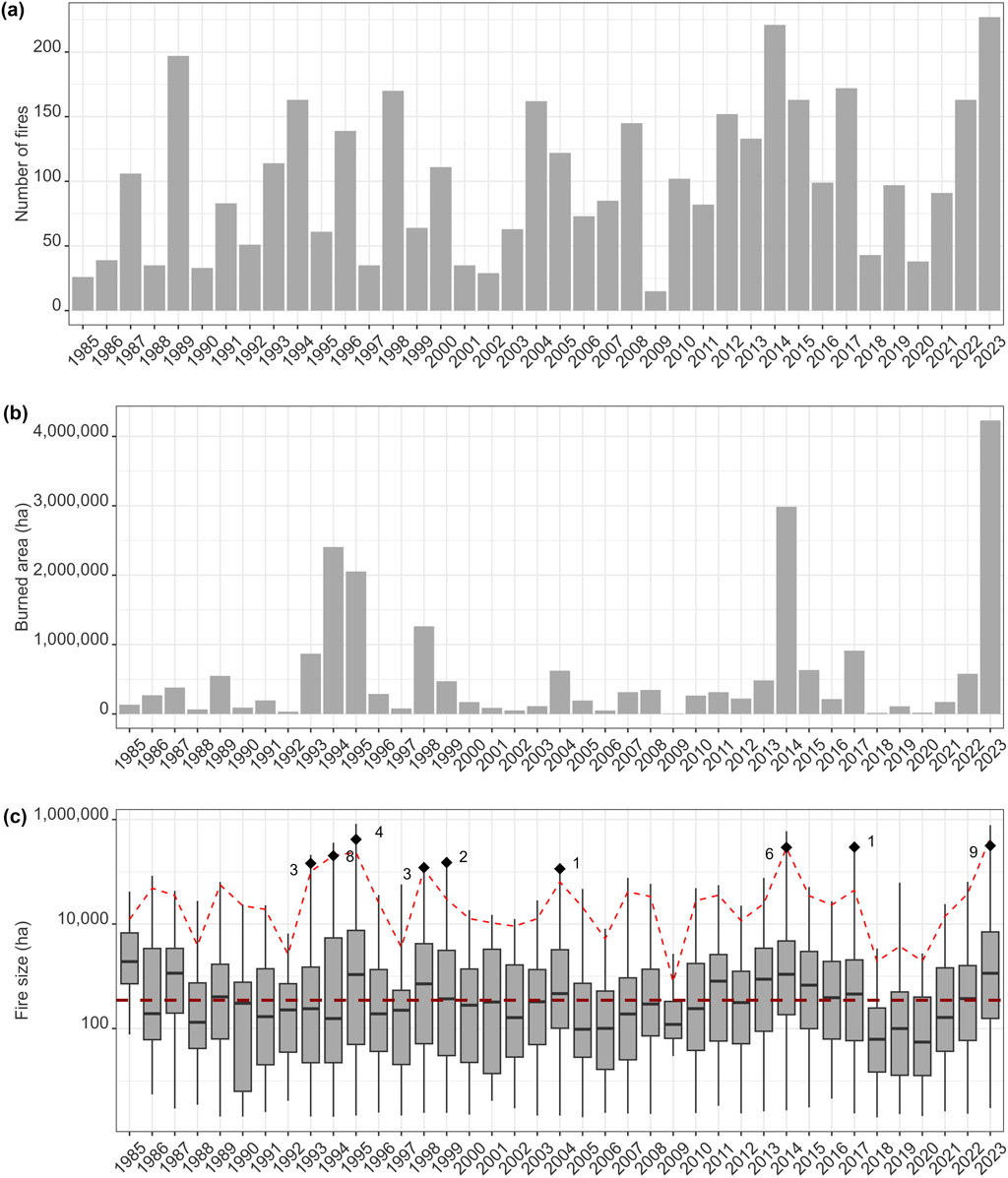
Figure 5. A comparison of the total number of fires (a), total burned area (b), and fire size (note the log scale) (c) of fire seasons from 1985 onward for the entire Northwest Territories. Note that 2023 had the largest number of fires, largest burned area, and is amongst the top fire years for largest mean fire size. In (c) the long-term median (1985–2022; 348 ha) is identified by the horizontal dark red line, whereas the mean area of the five largest fires for each year is indicated by the bright red line. Inset (c) also identifies the mean area (shown with solid diamond) and number (adjacent to solid diamond) of exceptionally large fire events, defined by exceeding the long-term mean +3 S.D. The statistics comparing 2023 to the long-term mean (1985–2022) are provided in Supplementary Table S2.
Short-term effects of more severe burning
More complete belowground combustion (i.e., severe burning of the soil organic layer and tree roots) can result in marked structural changes to the forest with implications for wildlife habitat conditions (Figure 6). Specifically, more severe belowground combustion should accelerate post-fire tree fall because the combustion of tree roots and the organic soil supporting them compromises the structural integrity of the trees. Dead fallen vs dead standing trees alter post-fire regeneration conditions (e.g., via shading), fuel structure, decomposition, and wildlife use (Palm et al., 2022; Parker et al., 1984; Parro et al., 2015). For example, increased deadfall in burns, either immediately post-fire or as standing dead trees that fall down over time, is thought to impede movements of boreal caribou (caribou avoid burns from ∼11–30 years post-fire; J. Hodson personal communication) and may be one of the mechanisms contributing to their avoidance of burns (Schaefer and Pruitt, 1991; Palm et al., 2022). More severe belowground combustion at the time of fire will accelerate tree fall and may therefore limit access to or movement through burns for a longer period of time and limit access to the protein-rich summer forage that caribou access in new burns. Combustion of roots and soil in 2023 was noticeably greater than in the next largest fire year in NWT (2014) and immediate post-fire tree fall was widespread, which was much less common in the 2014 fire season (J. Baltzer, personal observation).
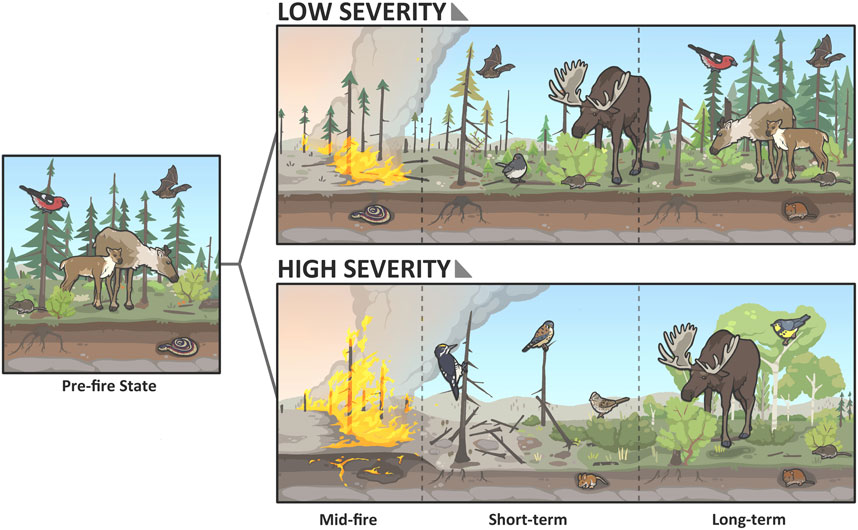
Figure 6. Illustration of the effects of high and low burn severity on forest structure and associated boreal species. High severity burns may burn deeper into the ground and completely remove the organic soil layer and more (if not all) of the surface litter, causing direct mortality in burrowing species. High severity burns experience slower forest recovery, and the removal of coniferous seed banks also encourages the replacement of coniferous forest by faster-growing deciduous trees, which will support a different community of boreal species. Low severity; left to Right: Red-sided garter snake (Thamnophis sirtalis parietalis), little brown myotis (Myotis lucifugus), dark-eyed junco (Junco hyemalis), masked shrew (Sorex cinereus), moose (Alces alces), white-winged crossbill (Loxia leucoptera), boreal caribou (Rangifer tarandus caribou) red-backed vole (Clethrionomys gapperi). High severity; left to right: Black-backed woodpecker (Picoides arcticus), American kestrel (Falco sparverius), deer mouse (Peromyscus maniculatus), Lincoln’s sparrow (Melospiza lincolnii), moose, Canada warbler (Cardellina canadensis), red-backed vole (Clethrionomys gapperi), masked shrew.
Severe burning directly impacts wildlife. For example, Knaggs et al. (2020) tested for the effects of fire severity (the first 2 years post-fire) on a community of 42 landbird species nesting in the large 2014 Birch Lake burn complex and surrounding unburned areas in southern NWT. Functional diversity, based on life history traits (foraging and migration strategy and foraging, breeding, and nesting substrate), decreased with increasing fire severity. The same pattern emerged with species richness, but only in peatlands. Unburned peatlands had higher species richness than unburned uplands, but species richness became more similar across habitat types as burn severity increased (Knaggs et al., 2020). Density or occupancy models were generated for 20 species and 86% of them showed significant burn severity effects (positive or negative), sometimes depending on prior vegetation (i.e., uplands vs peatlands). For example, the Olive-sided Flycatcher showed higher densities in recent burns irrespective of fire severity, while the Common Yellowthroat (Geothlypis trichas) showed the opposite response. Other species, such as the Black-backed Woodpecker (Picoides arcticus) and Three-toed Woodpecker (P. dorsalis), benefit from the emergence of nesting substrate and food resources because of large numbers of standing dead trees the first few years post-fire (Knaggs et al., 2020). Clearly there are differences in species responses, and we have insufficient knowledge of the responses of most wildlife to severe burning. Moose may respond negatively in the short-term to burn severity. Post-fire forage availability and burn severity were not strongly related across different soil moisture classes in a <5-year-old fire in north-central British Columbia, and moose used areas of low and medium burn severity more than areas of severe burning (McNay et al., 2021).
There are few published studies evaluating effects of fire severity on boreal small mammals, but studies from temperate forests found that, in the short-term, generalist species like deer mice were more abundant in areas of high burn severity (Culhane et al., 2022; Fontaine and Kennedy, 2012), whereas species dependent on the litter layer such as shrews were most negatively affected (Culhane et al., 2022). Fires that severely burn vegetation cover and seed or berry crops (in tree crowns, seed banks, and live shrubs) had negative impacts on foraging behaviour of specialist small mammals (e.g., voles [Microtus sp.] and Albert’s squirrel [Sciurus aberti]) but less so for generalist species (e.g., mice [Peromyscus sp.]) in a study in the southern United States (Morandini et al., 2023). In some cases, small mammal abundance does not differ across fire severity categories, but diversity and hence community structure are reduced after fires (Olson et al., 2003), especially after the most severe fires (Culhane et al., 2022). Terrestrial predators of small mammals like weasels select severe burns, which likely reflects greater availability of their preferred prey (deer mouse and meadow vole) in these areas (Volkman and Hodges, 2024). In contrast, Pacific marten (Martes caurina) favor lower severity burns which may reflect their need for areas with more abundant snags and downed woody debris for foraging and higher abundance of primary prey species like red-backed vole and red squirrel (Volkman and Hodges, 2024).
Because high severity burning generally results in greater loss of above- and below-ground biomass (Certini et al., 2021), severely burned areas are likely less suitable for amphibians and snakes owing to reduced cover and drier soil with more variable temperatures (Jager et al., 2021; Constible et al., 2001; Smith et al., 2001). Although lack of cover would typically increase predation risk to snakes, amphibians, and small mammals, it is possible that the risk from avian predators could be lower in severe burns with few or no remaining standing trees (i.e., perches for raptors such as American kestrel [Falco sparverius]; Widén, 1994) and substantial downed wood that can serve as protective cover. Changes to aquatic habitat for amphibians (e.g., oxygen levels; water temperature; concentrations of ash, sediment and nutrients; loss of vegetation around wetlands) are also exacerbated with severe fire (Jager et al., 2021; Gomez Isaza et al., 2022; Santos et al., 2022; Beranek et al., 2023).
Although fire can help to create roosting habitat for boreal bats by damaging or killing large trees (e.g., Johnson et al., 2009), these trees must remain standing after fire. Accelerated post-fire tree fall due to severe burning, combined with less residual vegetation (i.e., fewer green islands) in severe burns, would likely result in fewer roosts available on the landscape. Little brown myotis may be less impacted than northern myotis if they have access to alternate roosts (e.g., buildings). While reduction in vegetative clutter in forests can improve foraging opportunities for some bats (e.g., little brown myotis, big brown bat [Eptesicus fuscus]; Blakey et al., 2019; Low et al., 2024), severe burning can result in poor foraging habitat that is too open and well-lit for bats to fly (Jung, 2020). The short-term pulses in insect productivity that typically occurs after fire (Lacki et al., 2009; Malison and Baxter, 2010) are less likely to occur, or at least at a lower magnitude, in high severity burns with extensive combustion of above- and below-ground biomass; in fact, insect abundance may actually be lower post-fire (Dole et al., 2023). Few studies on the response of bats to fire have been conducted in the boreal forest where the availability of roost trees on the landscape is limited and bats’ foraging opportunities are already constrained by high latitude. However, a large, severe wildfire in Yukon created large patches of homogeneously burned areas that had negative effects on little brown myotis (Jung, 2020). Most importantly, the fire resulted in the nearly complete loss of suitable roost trees: although dead trees (snags) remained in burned uplands, none of them were sufficiently large (≥30 cm diameter at breast height) to meet the roosting requirements of this species (Crampton and Barclay, 1998; Jung et al., 2004). Sites where at least some large standing trees remain are likely very important for the persistence of bats post-fire (Jung, 2020).
Long-term effects of more severe burning
More severe burning increases the likelihood of post-fire forest compositional change (typically conifer to deciduous) and even complete recruitment failure (i.e., forest to non-forest; Johnstone et al., 2016; Whitman E. et al., 2019; Baltzer et al., 2021). Severe canopy combustion can lead to the death or combustion of seeds in aerial cone banks, reducing rates of establishment post-fire (Reid et al., 2023; Splawinski et al., 2019). Likewise, more complete combustion of the soil organic layer alters seedbed conditions by exposing underlying mineral soil, favoring faster growing, broadleaf deciduous taxa over the historically dominant spruce species (Johnstone et al., 2016; Whitman E. et al., 2019; Baltzer et al., 2021). Indeed, modelled projections of these processes demonstrate strong likelihood of landscape level forest compositional changes at high latitudes from conifer to broadleaf deciduous in response to climate warming and increasing fire (Mekonnen et al., 2019; Micheletti et al., 2021). Forest compositional changes or loss attributable to severe combustion can be further compounded by fire in permafrost environments such as those of NWT where fire accelerates thaw. Specifically, burning, but particularly severe burning where there is more complete combustion of the soil organic layer that protects permafrost (i.e., ground perennially at or below 0 °C) drives rapid permafrost thaw (Talucci et al., 2024; Holloway et al., 2020). This can lead to a range of outcomes, but in thaw sensitive landscapes (e.g., permafrost peatlands), both forest compositional changes and net forest loss are possible in response to thaw (e.g., Lara et al., 2016; Baltzer et al., 2014). As described above, a transition from conifer to deciduous may favor moose and lead to habitat losses for mature conifer forest specialists like boreal caribou (Stewart et al., 2023) and squirrels. Boreal bats and other tree-roosting species are associated with large and/or dead trees, therefore recruitment failure because of severe burning that turns forest into non-forest (Johnstone et al., 2016; Whitman E.et al., 2019; Baltzer et al., 2021) would reduce the amount of available habitat. Similar responses are anticipated for the woodpecker community and all other species associated with mature forests. Deep burning also modifies the post-fire soil microbial community with implications for biogeochemical cycling and plant community composition and dynamics, adding complexity to projections of post-fire successional dynamics (Day et al., 2019; Whitman T. et al., 2019; Eckdahl et al., 2023; Eckdahl et al., 2024).
Boreal caribou is a species at risk for which we have a better understanding of long-term impacts of severe burning. During winter, boreal caribou avoid areas of higher burn severity for up to 30 years, whereas in summer they exhibit weak avoidance of high and low severity burn areas (Palm et al., 2022), or even similar selection to unburned forest (A. Kelly, personal communication). Areas selected within burns corresponded to areas with higher percent cover of lichens, and remaining lichen cover within burns was negatively related to burn severity (Palm et al., 2022; Pinno and Errington, 2016), highlighting the importance of variable burn severity and associated post-fire residual vegetation. Negative relationships between winter movement speeds of barren-ground caribou and burn severity have also been found which may indicate greater foraging activity within areas of lower burn severity. However, movement speeds in all burns were still higher than in unburned forests suggesting recent burns were primarily used for movement rather than foraging (Rickbeil et al., 2018). On the other hand, moose may benefit from high severity fires, though this appears to depend on time after fire and time of year. Moose forage biomass was up to three times greater in high severity burns than low severity burns in a 14-year-old fire in Alaska, and moose consumed more forage biomass and a higher proportion of available forage in these high severity burns (Lord and Kielland, 2015). However, areas of lower burn severity may provide a combination of both forage and cover that favors use by moose. For example, in a 20-year old fire in Alaska, moose selected areas of low burn severity in winter, and high burn severity in summer (Brown et al., 2018).
Lower severity burning leads to greater likelihood of self-replacement in conifer stands (Baltzer et al., 2021). Notably, spruce establishes well on thicker residual soil organic layers, which is not the case for faster growing broadleaf deciduous trees or pine (Johnstone et al., 2010a; Baltzer et al., 2021). Lower severity fire also corresponds with a greater frequency of residual vegetation (i.e., “green islands”; Perera et al., 2009), which may enhance landscape connectivity of burned areas, helping to accelerate regeneration, maintain structural diversity, and promote use by species known to avoid wildfires (reviewed in Sommers and Flannigan, 2022). For example, boreal caribou have been shown to use residual patches within burns for calving demonstrating their importance in reducing impacts of fire on this species (Skatter et al., 2017).
Similarly to the short-term effects, many of the longer-term effects of fire on ground-dwelling, less mobile species described earlier (see “Area burned and fire size”) will be more pronounced with higher severity burning. Widespread combustion of features that these species use as refugia, like logs, hollow trees or burrows, could slow or prevent the recovery of local populations (Robinson et al., 2013) and potentially reduce their resilience to future fires. Species dependent on cool, moist microhabitats, like wood frogs, may show reduced persistence and time-lagged declines in occupancy, especially where a large proportion of their terrestrial habitat is severely burned (Bailey et al., 2025; Hossack et al., 2013; but see Hunter, 2022). Changes to amphibian breeding wetlands (e.g., productivity, hydroperiod, contaminants) are more dramatic when burning is severe (dos Anjos et al., 2021; Gomez Isaza et al., 2022).
In summary, critical impacts of severe burning in the short term relate to post-fire forest structure that affects landscape permeability, availability of large snags for roosting and foraging, and protective cover from predators and thermal variation. Longer term land cover changes associated with severe burning (i.e., conifer to broadleaf deciduous) will favor some species and disadvantage others, altering the structure of terrestrial wildlife communities in similar ways as large fires but likely more consistently reinforcing forest compositional change than is the case for large fires.
Short-interval reburning
Young forest stands have historically served as natural fire breaks owing to fuel limitations (Thompson et al., 2017) and less flammable fuel types (Hanes et al., 2020), which has resulted in strong fire self-regulation on the landscape (Heon et al., 2014; Prichard et al., 2017). Climate drivers are leading to extreme fire weather that reduces this fire self-regulation mechanism (Whitman et al., 2024). This was evident in NWT in 2023 with more than 400,000 ha of forested land experiencing short-interval reburning (i.e., reburning after only 20 years; Figure 7). There is evidence that short-interval reburns (forests younger than 20 years) have been increasing slightly through time from about 2% to 5% of annual burned area in Alaska since the early 1990s (Buma et al., 2022). In contrast, approximately 10% of forest that burned in 2023 in NWT was <20 years old (400K ha short-interval reburn in 4 M ha of total burned area), more than double the reported maximum proportional burned area as was found in Buma et al. (2022). This is in keeping with evidence that short-interval reburning can become much more common during hot, dry fire seasons (Heon et al., 2014; Whitman et al., 2024). In 2023, short-interval reburning occurred across a diversity of landscape positions from treed wetlands to dry upland forests (Figure 8) with implications for forest composition, structure, and function and the wildlife using these lands.
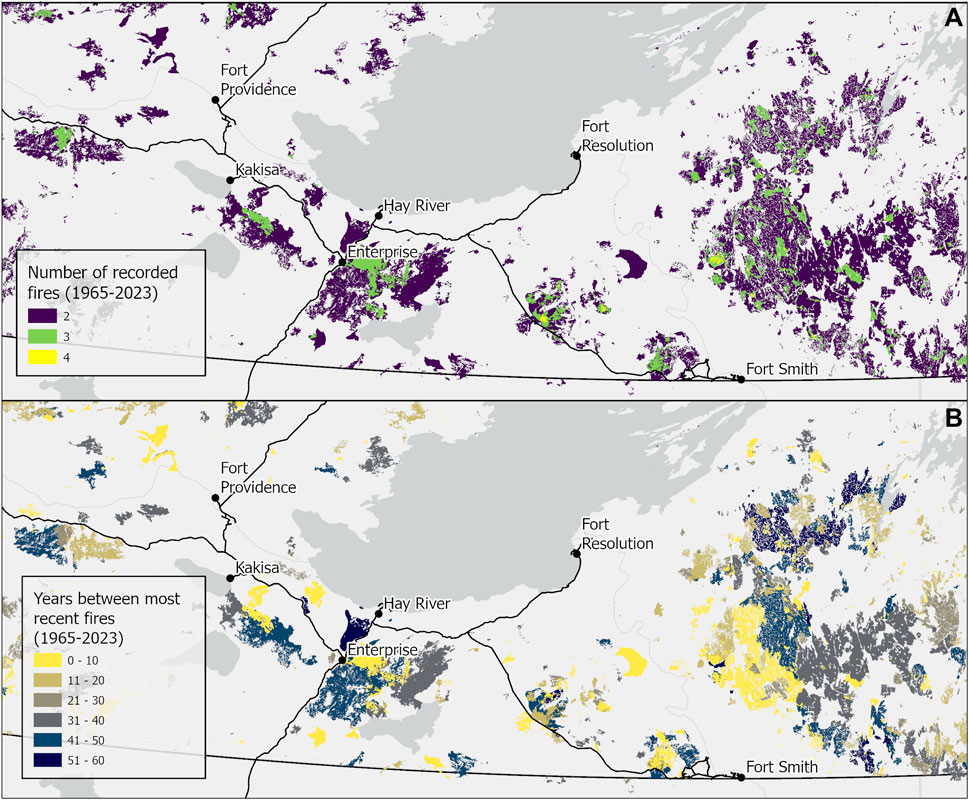
Figure 7. Short-interval reburning in the South Slave and Dehcho regions of the Northwest Territories where this was most common in 2023. (A) shows the fires in each area since 1965 when the recorded fire history for this region begins. (B) shows the fire return interval in the same region where fire return interval is less than 60 years.

Figure 8. Pictures of short-interval reburns from the 2023 wildfires in Northwest Territories taken in summer 2024 demonstrating loss of material legacies. All locations burned in 2014 and reburned in 2023. Examples include (A) a treed fen near Kakisa, NWT; (B) a dry, upland forest near Fort Smith, NWT; and (C) a mesic forest near Hay River, NWT. In (C) note the “shadows” of downed stems from the 2014 fires that were combusted during the 2023 fires. Photo credits: J. Baltzer.
Short-term effects of short-interval reburning
Short-term consequences of short-interval reburning may mirror those of high severity burning for several taxa. These include more complete combustion of material legacies including the soil organic layer (Hoy et al., 2016; Walker et al., 2018; 2019) and standing and downed woody debris (Turner et al., 2019), as well as delayed regrowth of vegetation (Day et al., 2020; Hollingsworth et al., 2013). These changes will create challenging conditions for wildlife species requiring soil moisture, standing trees, vegetation structure, and cover (e.g., amphibians, red-sided garter snake, bats, woodpeckers; Figure 9), with potential impacts on foraging, survival, reproductive success and dispersal (Bailey et al., 2025; Beranek et al., 2023; Bury, 2004; Doherty et al., 2022; Hossack et al., 2013; Howey et al., 2016; Mahony et al., 2022; Muñoz et al., 2019; Rochester et al., 2010; Russell et al., 1999; Schurbon and Fauth, 2003; Webb and Shine, 2008; Wilgers and Horne, 2007; Thomas and Jung, 2019; Jung, 2020; Grindal and Brigham, 1999; Patriquin and Barclay 2003; Henderson and Broders, 2008). Where short-interval reburning is low severity, we expect more woody biomass to remain on site, including live trees in many cases and the soil organic layer should be more intact. As such, the short-term consequences of short interval fire will depend strongly on the severity of that fire.
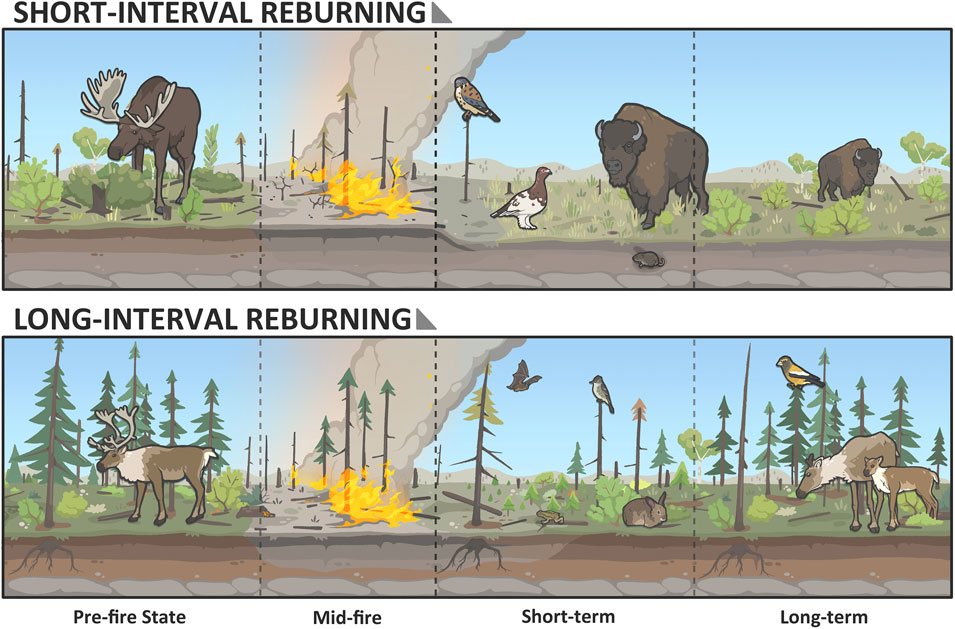
Figure 9. Illustration of an example of the effects of the interval between burns on forest structure and associated boreal species. Repeated burns combust trees’ reproductive structures (cones, rhizomes) and the soil organic layer and shift the plant community towards grass/shrublands and animal species that occupy them, such as: American kestrel (Falco sparverius), meadow vole (Microtus pennsylvanicus), and as shrubby cover regenerates, willow ptarmigan (Lagopus lagopus) and wood bison (Bison bison athabascae). A longer interval between burns increases the likelihood and speed of recovery to the original plant community and leaves more ground cover/litter and residual standing trees. These trees are vital for perching and foraging for olive-sided flycatcher (Contopus cooperi) and roosting for little brown myotis (Myotis lucifugus). Species such as boreal chorus frog (Pseudacris maculata) and snowshoe hare (Lepus americanus) are dependent on the remaining/regenerating cover. Return to coniferous-dominant forest will favour species such as evening grosbeak (Coccothraustes vespertinus) and boreal caribou (Rangifer tarandus caribou).
Long-term effects of short-interval reburning
In the longer term, short-interval reburning has a variety of consequences for regeneration. When fires are too close together in time, reproductive structures (cones, rhizomes) are damaged or destroyed causing forest compositional change, commonly conifer to broadleaf deciduous forest, or with multiple short-interval fires, failed regeneration leading to shrub thickets or grassland (Hayes and Buma, 2021; Whitman E. et al., 2019). Short-interval reburning results in more complete combustion of the soil organic layer (Hoy et al., 2016; Walker et al., 2018; 2019), which negatively impacts rhizomatous species and can lead to a shift toward reproduction from off-site seed sources (Day et al., 2020; Hollingsworth et al., 2013), leading to longer vegetation recovery times. These changes may not be unprecedented in the history of this area. Indeed, there is evidence from paleoecological studies that more widespread burning has reduced forest cover dramatically in the past (e.g., Girardin et al., 2024). Whether the type of short interval burning we are seeing today will move the system back toward this state is uncertain but initial reconnaissance in the southern NWT suggests that transitions from forest to prairie or aspen parkland ecosystems may be common in response to short interval reburning in southern NWT (Figures 10a,b). Where short-interval reburning is low severity, there may be many live trees remaining (e.g., Figure 10c) and recovery of ground vegetation may be much more rapid owing to the maintenance of on-site seed sources and intact belowground structures. However, very few data are available for either short or long-term impacts of short interval burning, the variability in its outcomes, or the vegetation conditions that are most likely to support this fire behaviour. This is a major gap in our understanding of changes in fire behaviour.

Figure 10. Short-interval reburns that occurred in the 2023 fire season near Fort Smith, Northwest Territories. Panels (A) and (B) depict stands that were forested pre-fire but show little evidence of tree regeneration and appear to be transitioning to grasslands dominated in this case by wild rye (Elymus innovatus) though other graminoid taxa can also be common. (C) shows a jack pine tree (Pinus banksiana) with multiple fire scars indicating repeated low severity (i.e., not stand-replacing) burning. The forest is open with groundcover dominated by caribou lichen (Cladonia spp.) and lingonberry (Vaccinium vitis-idaea). Photo credits: J. Baltzer.
Forest compositional change, longer recovery time, or failed regeneration due to short-interval reburning would have direct consequences for the wildlife taxa that rely on mature conifer forests. Reduced conifer forest recovery on the landscape would decrease habitat availability for species reliant on that forest type for all or part of their resource requirements. For example, as lichen biomass in NWT forests may take more than 70 years to recover to half that found in late-successional forests (Greuel et al., 2021), decreasing fire return intervals will for the most part lead to a loss of winter habitat for boreal caribou. In contrast, short-interval reburns leading to conversion from conifer to broadleaf deciduous stands may benefit species such as moose that predominantly forage on deciduous browse during winter (Fisher and Wilkinson, 2005; Maier et al., 2005); however, burns in peatlands may not lead to an increase in food availability for moose (DeMars et al., 2019). Similarly, conversion to grasslands associated with recruitment failure may benefit wood bison (Bison bison athabascae) that prefer to forage on grasses and sedges (Larter and Gates, 1991; Redburn et al., 2008) and meadow voles whose abundance and survival is positively correlated with grass cover (Adler and Wilson, 1989). Black bears (Ursus americanus), which are a generalist omnivore species, may also benefit from conversion to grasslands or deciduous vegetation as they tend to consume grasses, forbs and deciduous plants during spring and summer (Mosnier et al., 2008; Romain et al., 2013; Lesmerises et al., 2015; Jorgensen, 2021). Black bears tend to shift towards using younger conifer-dominated stands later in the summer and fall (Tomchuk, 2019), likely reflecting greater availability of berries in these habitats (Nelson et al., 1983; Tomchuk, 2019), and bears may thus benefit from shorter-interval reburns if they increase the prevalence of these stands on the landscape. When short-interval reburning is not stand replacing, fire will remove ground fuels and can leave an open canopy of mature trees, a fire behaviour that commonly occurs in mature jack pine stands (Figure 10c; Stocks, 1989). Anecdotally, many of these stands support high biomass of caribou lichen (J. Baltzer pers. obs.; M. Parisien pers. comm.), which would be beneficial for caribou winter habitat.
Repeat burning can also reduce the quality of habitat conditions in fresh burns by consuming material legacies from the previous fire. Notably, short-interval reburning can lead to the complete combustion of standing and downed woody debris, which reduces protective cover and habitat for a variety of wildlife and modifies the regeneration microenvironment dramatically for plants (Figure 8). For example, although snowshoe hare favour deciduous browse during winter, their need for protective cover (usually provided by conifer forest) might preclude them from benefiting from widespread conversion to deciduous-dominated stands or shrublands (Hutchen and Hodges, 2019). Similarly, lynx (Lynx canadensis) avoid travelling through open areas in recent burns and rely on the presence of preferred prey (snowshoe hare) meaning likely avoidance of short-interval reburns (Vanbianchi et al., 2017). Predators that rely on downed woody debris for subnivean access to prey and shelter, such as American marten (Viau et al., 2024), would also likely be negatively impacted.
Landbirds associated with mature forests such as Canada Warbler (Cardellina canadensis) (Reitsma et al., 2020) and Evening Grosbeak (Coccothraustes vespertinus) (Gillihan and Byers, 2020) are expected to experience important nesting habitat loss in response to shorter fire return intervals. The Olive-sided Flycatcher (Contopus cooperi) is an aerial insectivore often considered a “fire specialist”, but it also thrives at mature coniferous/recent burn ecotones and open mature conifer stands (Altman and Sallabanks, 2020). Thus, the anticipated loss of these edges and adjacent mature stands owing to very large fires and short fire return intervals will likely result in important breeding habitat loss for this species and other species associated with mature forests. For example, the entire woodpecker community will likely also experience important declines owing to loss of standing snags critical for feeding and nesting. Similar responses are anticipated for other species (e.g., bats and northern flying squirrels) using large, mature and dead trees (Senior et al., 2021; Loeb and Blakey, 2021). Similarly, woody debris used by amphibians and reptiles for habitat and refugia (Robinson et al., 2013) may not be available if wildfire occurs too soon after the last fire (Hossack and Pilliod, 2011), and deep accumulations of leaf litter used by amphibians can take years to recover from fire (Schurbon and Fauth, 2003). If more frequent large fires lead to a shift in the age-class distribution of the landscape towards higher prevalence of younger stands, we could see a shift in the boreal small mammal community towards dominance of early-successional or non-forest species like deer mice and meadow voles and lower populations of red squirrels.
In summary, most of the documented impacts of short-interval burning are those of loss of aboveground biomass and conversion of forests from conifer to broadleaf deciduous dominated or to a non-forested state. These changes will drive marked shifts in the wildlife community occupying areas of short-interval fire. However, the impacts, both short and longer term, are mediated by the severity of the fire (see Figure 11) such that low severity, short interval fire may have relatively modest or even beneficial effects in some cases.
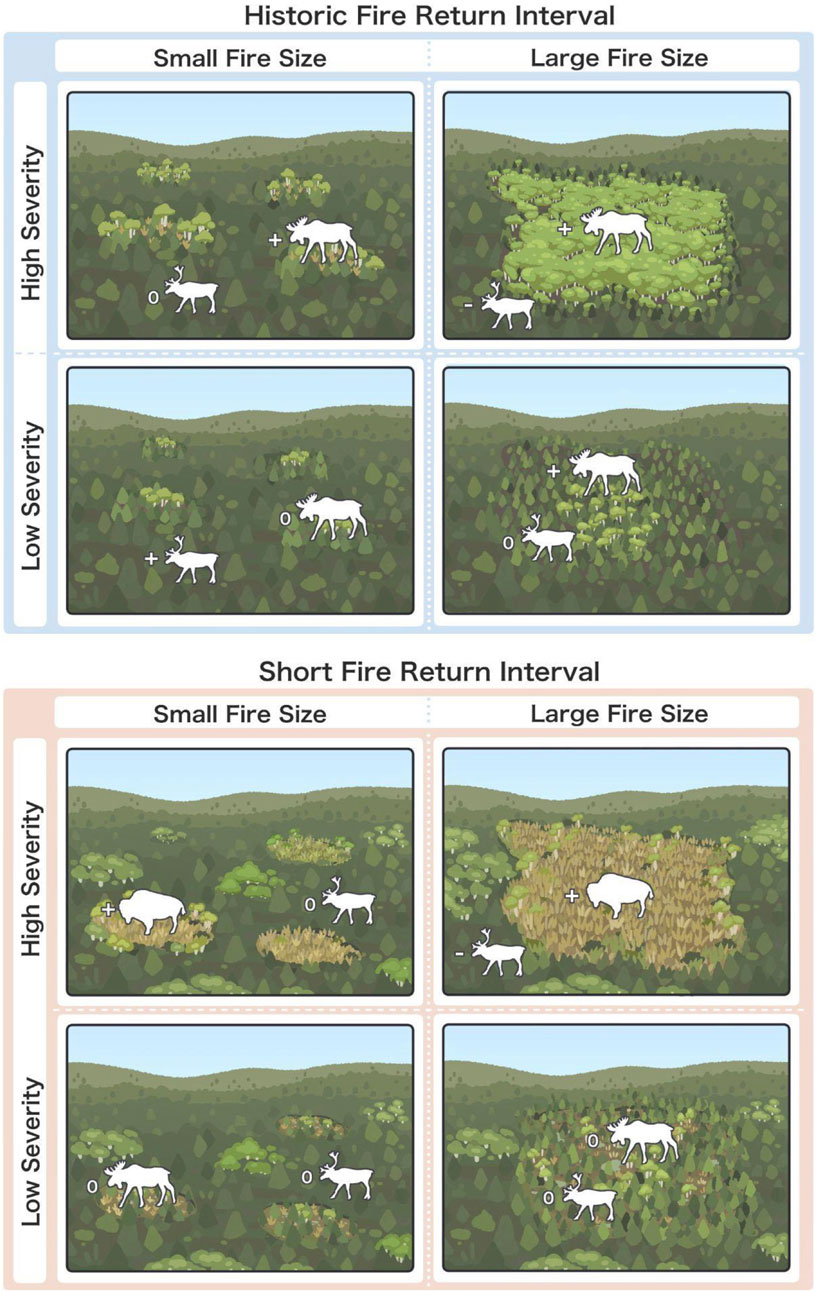
Figure 11. Illustration of expected long-term changes in forest landscape composition and patch structure in the NWT under different combinations of burn severity (High vs Low), fire size (Large vs Small), and fire return interval (Historic vs Short). Examples of corresponding changes in abundance (increase (+), decrease (−) or remain unchanged (‘o’)) of large terrestrial mammals - boreal caribou (Rangifer tarandus caribou), moose (Alces alces) and wood bison (Bison bison athabascae). For example, under a fire regime consisting primarily of large, high severity fires that reburn within short intervals (<20 years), we expect to see an increase in the prevalence of grassland and open shrubland habitat, with corresponding increases in wood bison abundance, and decreases in boreal caribou abundance. Under a fire regime consisting primarily of large, high severity fires that reburn at historic intervals (70–100 years), we would expect to see a higher prevalence of deciduous stands, with a corresponding increase in moose abundance and decrease in boreal caribou abundance.
Fire season length and timing
Fire season length and timing are changing in multiple ways across Canada and the broader boreal biome. Spring wildfires are increasing in many parts of Canada and are most common when snowmelt is early and/or under fire prone weather conditions (Albert-Green et al., 2013; Hanes et al., 2019; Parisien et al., 2023a). The frequency of overwintering fires is related to annual area burned with very large fire years corresponding with years having greater frequency of overwinter smouldering (Scholten et al., 2021). The 2023 wildfire season was no exception with western North America experiencing unprecedented numbers of overwintering fires (Baltzer et al., 2025). Late season burning is also increasing across boreal North America, a pattern that is particularly pronounced in the west (Kasischke and Turetsky, 2006).
Although there is limited information on how longer-, earlier-, and later-burning fire seasons will impact the boreal biome, there has been some investigation of early and late season fires in other ecosystems which may provide guidance for relevant research directions in boreal forests as fire season length and timing changes. For example, early season fires in grasslands can favor plant species that grow and reproduce later in the season, which could lead to plant community compositional shifts (reviewed in Knapp et al., 2009) with implications for wildlife forage. Potential short-term impacts of early season fires also include overlap in the timing of fire with peak reproductive times in some taxa. For example, wood bison begin calving in April (Jung et al., 2018; Species at Risk Committee, 2016) and most boreal caribou in NWT have their calves during May (Species at Risk Committee, 2022). While caribou calves quickly become mobile after birth, they may be particularly susceptible to mortality from early season fires as they tend to remain at the calving site for about 1 week postpartum (Gustine et al., 2006). Negative impacts can also be expected for resident birds (early breeders) and add physiological stress to arriving migratory birds. However, investigations of fire timing on mobile organisms in other systems suggest that these animals may simply avoid the disturbance (e.g., Murphy et al., 2009). For less mobile or hibernating taxa, early and late season and overwintering fires may have more disruptive effects. Snakes and amphibians may experience higher mortality rates if they encounter wildfire during a season when they are more exposed and vulnerable (Beaupre and Douglas, 2012; Cross, 2015), for example, red-sided garter snakes when migrating between their underground hibernation sites and wetland foraging areas in the spring and fall (Gregory and Stewart, 1975; Larsen, 1987). Wood frogs and boreal chorus frogs overwinter in a dormant state, in shallow micro-sites such as leaf litter, vegetation, or under cover objects such as logs and rocks (MacArthur and Dandy, 1982; Regosin et al., 2003) and would probably be killed if exposed to any out of season fire that overlaps with their dormant period. Certain species’ overwintering strategies may offer some protection from the direct impacts of early and late season fires in the boreal forest. Animals overwintering underground (e.g., toads in burrows; red-sided garter snakes in dens; bats in caves or crevices) are likely buffered from the effects of fire at the surface. However, during intense fires, amphibians in underground burrows can still be harmed or killed (Beranek et al., 2023; Mahony et al., 2022), as can snakes (Cross, 2015; González-Fernández et al., 2024; Jordaan et al., 2020; Smith et al., 2001). Hibernating bats respond to fire-related cues such as smoke (Dickinson et al., 2010; Doty et al., 2018) and could experience energetically costly arousals from torpor. Where many individuals overwinter communally together in a single location (e.g., toads, red-sided garter snakes, bats), any harmful effects of fire that do occur have the potential to impact a large part of the population simultaneously (e.g., Shine et al., 2001).
As outlined above, many of the anticipated impacts of out of season fires are acute leading to mortality. The short-term impacts of out-of-season fires will almost certainly relate to their timing and severity. There is the potential for late season fires to burn more deeply owing to complete seasonal thaw and low late season soil moisture (Kasischke and Turetsky, 2006). Impacts of this would presumably be similar to severe burning described above. Spring wildfires occur when the forest, particularly deciduous taxa, have not had a chance to fully rehydrate, providing a ready source of fuel. As such they can be very fast moving and may result in large burn areas (reviewed in Parisien et al., 2023b). The longer-term impacts of changes in fire season length and timing and increased frequency of overwintering fires include changes in forest composition and may favour regeneration of deciduous trees (Baltzer et al., 2025); responses of wildlife taxa to these changes have been elaborated on above. That said, we have little information on the impact of out-of-season fires and effort should be made to understand responses of land cover and associated wildlife to this increasingly important fire behaviour.
Synthesizing impacts and help for planning
Novel wildfire conditions are altering boreal landscapes and ecosystems, creating challenges for northern communities and lifestyles, particularly through impacts on the safety and security of country or subsistence foods and threatening biodiversity and species at risk, with implications for ecosystem functions. Northern communities and governments require information to support adaptation planning in response to these changes. The preceding sections offer us a range of possibilities about the future of wildlife community structure under changing wildfires in northern boreal forests. We have synthesized what we see as likely outcomes to landscape scale changes resulting from changing wildfire based on published studies and experience of the authors in these systems and with the taxa that were considered (see Figure 11 for important large ungulates and Table 1 for a complete list of assessed taxa). However, these possible outcomes are difficult to use directly in planning owing to uncertainties arising from potential feedbacks between fire and landcover conditions as well as insufficient information on the responses of many northern taxa to wildfire. For example, will the increase in high severity burns and decrease in reburn interval cause widespread declines in bats, birds, snakes, small mammals, and boreal caribou because the boreal ecosystem will have widespread conversion to grasslands due to tree regeneration failures? Or will this ecosystem conversion indeed occur, only in a small fraction of the area, resulting in quantitatively modest changes to the wildlife? If these changes are widespread, when would these declines become more important, what might be the population impacts and the implications on the conservation status of different northern wildlife taxa? In the example of short-interval reburn, does this fire behaviour create a positive feedback with areas reburning ever more quickly, or does this lead to an eventual negative feedback with fewer fires by the end of the century owing to fuel limitation (e.g., Marchal et al., 2020) thereby stabilizing observed changes? Given the many outstanding questions reviewed above there is much work to do and we suggest three paths forward for wildlife-wildfire research in northern boreal regions that would help to fill key knowledge gaps and provide information in support of adaptation planning.
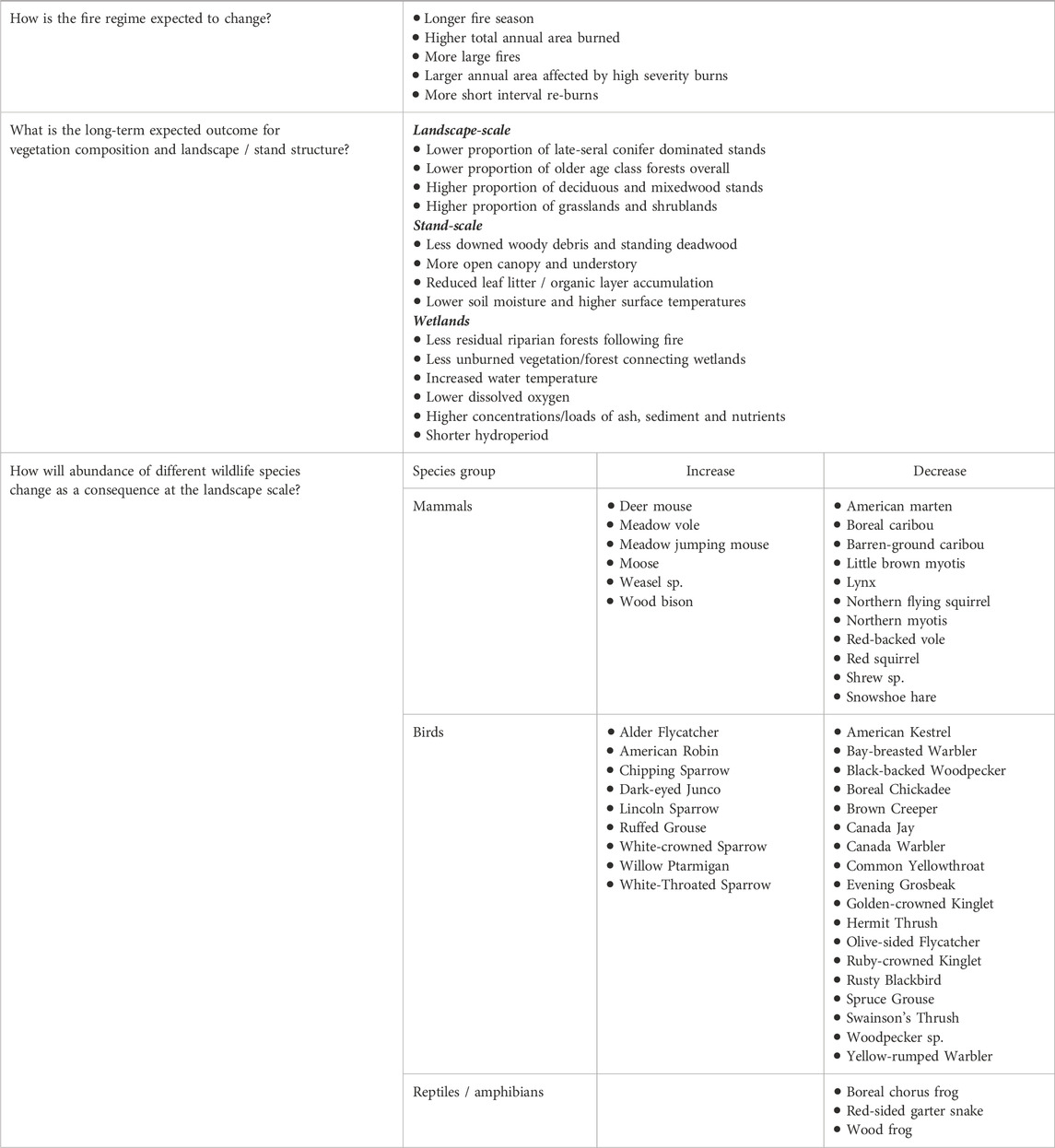
Table 1. Summary of expected changes in components of the NWT fire regime under climate change, resulting long-term (e.g., next 100 years) likely changes in composition and structure of forests at landscape and stand scales, including implications for wetlands, and examples of expected changes in abundance of terrestrial wildlife species based on our review of literature. Wildlife species for which it was uncertain whether the direction of change would be predominantly negative or positive are not included.
The first path is to make better use of existing and ongoing monitoring programs (collar data, cameras, audio recording units (ARUs), small mammal monitoring, studies of land cover change, etc.) to understand responses of northern wildlife to fire age, severity, and frequency, and where needed expand monitoring programs. Outside of boreal caribou and, to a lesser extent barren-ground caribou, there is limited information on population responses to wildfire and post-fire habitat selection for other large herbivores, including moose, muskox, and bison, all three of which are important to northern ecosystems and communities. Responses of furbearers, including semi-aquatic furbearers like beaver and muskrat, bears, and other predators were other major gaps. While existing individual-based collaring programs for bison and moose could be leveraged to fill knowledge gaps for these species, filling knowledge gaps for other less well-studied northern boreal species will require expanding research and monitoring initiatives. While there is comparatively better understanding of boreal caribou and barren-ground caribou responses, more work could be done for these species to compare habitat selection/use pre- and post-fire, responses to burn severity and short interval reburns, and on-the-ground work looking at caribou forage resources as a function of fire age and burn severity, tied to collar location data so we are actually sampling places that we know caribou are either selecting or avoiding.
A second path is to take advantage of remote-sensing and other emerging technologies to model and map fire refugia and track broad-scale and, where possible, fine-scale changes in wildlife habitat in near real-time. Northern regions, including the NWT, present a unique opportunity for wildlife-wildfire research given the relatively low human footprint and more limited fire suppression activities, potentially allowing for wildlife responses to wildfire to be more easily parsed out from other anthropogenic impacts. At the same time, the extensive and remote landscapes make in situ research and monitoring costly and resource limitations will always result in some species being under-monitored and researched and necessitates the coupling of in situ habitat change studies with remotely sensed data products. Given these challenges, ongoing development, ground truthing, and application of remote monitoring tools for measuring habitat and landscape changes in a timely manner will be important. This includes mapping and modeling areas on the landscape that are more resilient to climate change and climate change impacts, including fire refugia.
A third path is to use what is learned and developed in paths one and two to forecast species responses to future wildfire and landscape change. Indeed, making projections i.e., predictions of the future followed by testing, is a valuable means of converting our inferential knowledge to reliable actionable science (Houlahan et al., 2017). One promising pathway for planners and decision makers when facing unprecedented and changing wildfires is offered by the Ecological Forecasting community (Dietze, 2017; Dietze et al., 2018; McIntire et al., 2022). Evidence is mounting that forecasting surprises us by showing that simpler models (e.g., “the null” or “less mechanism”) are better at forecasting than more sophisticated models with more parameters (Barros et al., 2023; Wheeler et al., 2024). Each of the inferences outlined in this review can become part of quantitative forecasting efforts where we can evaluate multiple drivers and direct and indirect effects. For example, there have been initial attempts to build forecasts for these ecosystems with some of the processes and species outlined here (e.g., landbirds (Raymundo et al., 2024), and boreal caribou (Stewart et al., 2023)). Further investigations demonstrated that a forecasted increase in area burned caused only a modest forecasted change in landbirds, and that this effect was much smaller than direct climate effects (Micheletti et al., 2023). With this new knowledge, planners can see trade-offs more clearly and prioritize actions now to deal with the changing fire regimes in the context of other environmental changes.
In conclusion, there is an urgent need to improve our understanding of the impacts of the novel burning conditions experienced in 2023 on northern habitats and the associated wildlife communities. Some of the largest unknowns relate to the direct effects of the changing fire regime on wildlife taxa necessitating additional baseline data collection to better understand these impacts. Indeed, aside from some species at risk, long-term monitoring of northern wildlife populations is highly limited, negatively impacting our ability to accurately anticipate the impacts of increasing wildfire or other changes. Implications of climate change and wildfire-mediated boreal vegetation transitions for species range shifts and invasions is another major unknown. The development of ecological forecasting tools to anticipate both landcover changes and associated impacts on wildlife will be essential for informing conservation and management actions. Given the important knowledge gaps and risks associated with all this uncertainty, there is a great need to set wildlife research and monitoring priorities based on contributions of all co-management partners across the NWT and beyond, and to better coordinate resource allocations for more effective co-production of knowledge to inform wildlife conservation in a changing world.
Author contributions
JB: Conceptualization, Writing – original draft, Writing – review and editing. SH: Conceptualization, Writing – original draft, Writing – review and editing. JH: Conceptualization, Writing – original draft, Writing – review and editing. MA: Formal Analysis, Visualization, Writing – review and editing. SC: Visualization, Writing – review and editing. MC: Formal Analysis, Writing – review and editing. EM: Writing – original draft, Writing – review and editing. AM: Conceptualization, Writing – review and editing. ER: Visualization, Writing – review and editing. MT: Conceptualization, Writing – review and editing. JvdS: Formal Analysis, Visualization, Writing – review and editing. JW: Visualization, Writing – review and editing. BW: Conceptualization, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. JB was supported by the Canada Research Chairs program and a NSERC Discovery Grant.
Acknowledgements
The authors acknowledge valuable conversations with various colleagues that led to the development of ideas in this manuscript. We are grateful for field support from H. Cott in acquiring photos in Figure 8. E. Tattersall, I. Lane-Shaw, and M. Knaggs all provided input on responses of bird functional groups to wildfire.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2025.1504568/full#supplementary-material
References
Adler, G. H., and Wilson, M. L. (1989). Demography of the meadow vole along a simple habitat gradient. Can. J. Zool. 67, 772–774. doi:10.1139/z89-110
Albert-Green, A., Dean, C. B., Martell, D. L., and Woolford, D. G. (2013). A methodology for investigating trends in changes in the timing of the fire season with applications to lightning-caused forest fires in Alberta and Ontario, Canada. Can. J. For. Res. 43, 39–45. doi:10.1139/cjfr-2011-0432
Aldous, C. M. (1937). Notes on the life history of the snowshoe hare. J. Mammal. 18, 46–57. doi:10.2307/1374307
Altman, B., and Sallabanks, R. (2020). “Olive-sided flycatcher (Contopus cooperi),” in Birds of the world. Editor A. F. Poole (Ithaca, NY, USA: Cornell Lab of Ornithology).
Azeria, E. T., Ibarzabal, J., Hébert, C., Boucher, J., Imbeau, L., and Savard, J. P. L. (2011). Differential response of bird functional traits to post-fire salvage logging in a boreal forest ecosystem. Acta Oecol. 37, 220–229. doi:10.1016/j.actao.2011.02.005
Bailey, L. L., Henderson, R., Estes-Zumpf, W. A., Rhoades, C. C., Miller, E., Lujan, D., et al. (2025). Unburned habitat essential for amphibian breeding persistence following wildfire. Glob. Ecol. Conservation 57, e03389. doi:10.1016/j.gecco.2024.e03389
Baltzer, J. L., Veness, T., Sniderhan, A. E., Chasmer, L. E., and Quinton, W. L. (2014). Forests on thawing permafrost: fragmentation, edge effects, and net forest loss. Glob. Change Biol. 20, 824–834. doi:10.1111/gcb.12349
Baltzer, J. L., Day, N. J., Walker, X. J., Greene, D., Mack, M. C., Alexander, H. D., et al. (2021). Increasing fire and the decline of fire adapted black spruce in the boreal forest. Proc. Natl. Acad. Sci. U. S. A. 118, e2024872118. doi:10.1073/pnas.2024872118
Baltzer, J. L., Walker, X. J., Veraverbeke, S., Hessilt, T. D., Alfaro-Sanchez, R., van Gerrevink, M. J., et al. (2025). Overwintering fires can occur in both peatlands and upland forests with varying ecological impacts. Nat. Ecol. Evol. 9, 559–564. doi:10.1038/s41559-024-02630-2
Bancroft, B. A., Baker, N. J., and Blaustein, A. R. (2008). A meta-analysis of the effects of ultraviolet B radiation and its synergistic interactions with pH, contaminants, and disease on amphibian survival. Conserv. Biol. 22, 987–996. doi:10.1111/j.1523-1739.2008.00966.x
Banks, S. C., Dujardin, M., McBurney, L., Blair, D., Barker, M., and Lindenmayer, D. B. (2011). Starting points for small mammal population recovery after wildfire: recolonisation or residual populations? Oikos 120, 26–37. doi:10.1111/j.1600-0706.2010.18765.x
Barber, Q. E., Jain, P., Whitman, E., Thompson, D. K., Guindon, L., Parks, S. A., et al. (2024). The Canadian fire spread dataset. Sci. Data 11, 764. doi:10.1038/s41597-024-03436-4
Barros, C., Luo, Y., Chubaty, A. M., Eddy, I. M. S., Micheletti, T., Boisvenue, C., et al. (2023). Empowering ecological modellers with a PERFICT workflow: seamlessly linking data, parameterisation, prediction, validation and visualisation. Methods Ecol. Evol. 14, 173–188. doi:10.1111/2041-210X.14034
Beaupre, S. J., and Douglas, L. E. (2012). “Responses of timber rattlesnakes to fire: lessons from two prescribed burns,” in Proceedings of the 4th fire in eastern Oak forests conference. Editors D. C. Dey, M. C. Stambaugh, S. L. Clark, and C. J. Schweitzer (Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station), 2011 May 17–19; Springfield, MO. Gen. Tech. Rep. NRS-P-102. 192–204.
Beranek, C. T., Sanders, S., Clulow, J., and Mahony, M. (2022). Factors influencing persistence of a threatened amphibian in restored wetlands despite severe population decline during climate change driven weather extremes. Biodivers. Conservation 31, 1267–1287. doi:10.1007/s10531-022-02387-9
Beranek, C. T., Hamer, A. J., Mahony, S. V., Stauber, A., Ryan, S. A., Gould, J., et al. (2023). Severe wildfires promoted by climate change negatively impact forest amphibian metacommunities. Divers. Distributions 29 (6), 785–800. doi:10.1111/ddi.13700
Bergeron, Y., Gauthier, S., Flannigan, M., and Kafka, V. (2004). Fire regimes at the transition between mixedwood and coniferous boreal forest in Northwestern Quebec. Ecology 85, 1916–1932. doi:10.1890/02-0716
Beverly, J., and Schroeder, D. (2024). Alberta’s 2023 wildfires: context, factors and futures. Can. J. For. Res. 55, 1–19. doi:10.1139/cjfr-2024-0099
Bill, K. E., Dieleman, C. M., Baltzer, J. L., Degré-Timmons, G. É., Mack, M. C., Day, N. J., et al. (2023). Post-fire recovery of soil organic layer carbon in Canadian boreal forests. Ecosystems 26, 1623–1639. doi:10.1007/s10021-023-00854-0
Blakey, R. V., Webb, E. B., Kesler, D. C., Siegel, R. B., Corcoran, D., and Johnson, M. (2019). Bats in a changing landscape: linking occupancy and traits of a diverse montane bat community to fire regime. Ecol. Evol. 9, 5324–5337. doi:10.1002/ece3.5121
Bond, M. L. (2015). “Chapter 4. Mammals and mixed- and high-severity fire,” in The ecological importance of mixed-severity fires: nature's phoenix. Editors D. A. DellaSala, and C. Hanson (Boston, United States: Elsevier).
Bosso, L., Ancillotto, L., Smeraldo, S., D’Arco, S., Migliozzi, A., Conti, P., et al. (2018). Loss of potential bat habitat following a severe wildfire: a model-based rapid assessment. Int. J. Wildland Fire 27, 756–769. doi:10.1071/wf18072
Boulanger, D. Y., Arseneault, M. D., Bélisle, A. C., Bergeron, D. Y., Boucher, D. J., Boucher, D. Y., et al. (2024). The 2023 wildfire season in Québec: an overview of extreme conditions, impacts, lessons learned and considerations for the future. Can. J. For. Res. 55, 1–21. doi:10.1139/cjfr-2023-0298
Brandt, J. P., Flannigan, M. D., Maynard, D. G., Thompson, I. D., and Volney, W. J. A. (2013). An introduction to Canada’s boreal zone: ecosystem processes, health, sustainability, and environmental issues. Environ. Rev. 21, 207–226. doi:10.1139/er-2013-0040
Brown, C. L., Kielland, K., Euskirchen, E. S., Brinkman, T. J., Ruess, R. W., and Kellie, K. A. (2018). Fire-mediated patterns of habitat use by male moose (Alces alces) in Alaska. Can. J. Zool. 96, 183–192. doi:10.1139/cjz-2017-0069
Browne, C. L., Paszkowski, C. A., Foote, A. L., Moenting, A., and Boss, S. M. (2009). The relationship of amphibian abundance to habitat features across spatial scales in the Boreal Plains. Ecoscience 16 (2), 209–223. doi:10.2980/16-2-3220
Buchalski, M. R., Fontaine, J. B., Heady III, P. A., Hayes, J. P., and Frick, W. F. (2013). Bat response to differing fire severity in mixed-conifer forest California, USA. PLoS One 8 (3), e57884. doi:10.1371/journal.pone.0057884
Buma, B., Hayes, K., Weiss, S., and Lucash, M. (2022). Short-interval fires increasing in the Alaskan boreal forest as fire self-regulation decays across forest types. Sci. Rep. 12, 4901. doi:10.1038/s41598-022-08912-8
Burrell, A. L., Sun, Q., Baxter, R., Kukavskaya, E. A., Zhila, S., Shestakova, T., et al. (2022). Climate change, fire return intervals and the growing risk of permanent forest loss in boreal Eurasia. Sci. Total Environ. 831, 154885. doi:10.1016/j.scitotenv.2022.154885
Burton, P. J., Parisien, M.-A., Hicke, J. A., Hall, R. J., and Freeburn, J. T. (2008). Large fires as agents of ecological diversity in the North American boreal forest. Int. J. Wildland Fire 17, 754. doi:10.1071/WF07149
Bury, R. B. (2004). Wildfire, fuel reduction, and herpetofaunas across diverse landscape mosaics in northwestern forests. Conserv. Biol. 18, 968–975. doi:10.1111/j.1523-1739.2004.00522.x
Carvalho-Santos, C., Marcos, B., Nunes, J., Regos, A., Palazzi, E., Terzago, S., et al. (2019). Hydrological impacts of large fires and future climate: modeling approach supported by satellite data. Remote Sens. 11, 2832. doi:10.3390/rs11232832
Certini, G., Moya, D., Lucas-Borja, M. E., and Mastrolonardo, G. (2021). The impact of fire on soil-dwelling biota: a review. For. Ecol. Manag. 488, 118989. doi:10.1016/j.foreco.2021.118989
Chavardès, R. D., Daniels, L. D., Gedalof, Z., and Andison, D. W. (2018). Human influences superseded climate to disrupt the 20th century fire regime in Jasper National Park, Canada. Dendrochronologia 48, 10–19. doi:10.1016/j.dendro.2018.01.002
Christianson, A. C., Sutherland, C. R., Moola, F., Gonzalez Bautista, N., Young, D., and MacDonald, H. (2022). Centering Indigenous voices: the role of fire in the boreal forest of North America. Curr. For. Rep. 8, 257–276. doi:10.1007/s40725-022-00168-9
Chylek, P., Folland, C., Klett, J. D., Wang, M., Hengartner, N., Lesins, G., et al. (2022). Annual mean arctic amplification 1970–2020: observed and simulated by CMIP6 climate models. Geophys. Res. Lett. 49, e2022GL099371. doi:10.1029/2022GL099371
Cline, B. B., and Hunter Jr, M. L. (2016). Movement in the matrix: substrates and distance-to-forest edge affect postmetamorphic movements of a forest amphibian. Ecosphere 7 (2), e01202. doi:10.1002/ecs2.1202
Constible, J. M., Gregory, P. T., and Anholt, B. R. (2001). Patterns of distribution, relative abundance, and microhabitat use of anurans in a boreal landscape influenced by fire and timber harvest. Ecoscience 8 (4), 462–470. doi:10.1080/11956860.2001.11682676
Coogan, S. C. P., Daniels, L. D., Boychuk, D., Burton, P. J., Flannigan, M. D., Gauthier, S., et al. (2021). Fifty years of wildland fire science in Canada. Can. J. For. Res. 51, 283–302. doi:10.1139/cjfr-2020-0314
Coops, N. C., Hermosilla, T., Wulder, M. A., White, J. C., and Bolton, D. K. (2018). A thirty year, fine-scale, characterization of area burned in Canadian forests shows evidence of regionally increasing trends in the last decade. PLoS ONE 13, e0197218. doi:10.1371/journal.pone.0197218
Crampton, L. H., and Barclay, R. M. (1998). Selection of roosting and foraging habitat by bats in different-aged aspen mixedwood stands. Conserv. Biol. 12 (6), 1347–1358. doi:10.1046/j.1523-1739.1998.97209.x
Cringan, A. T. (1958). Influence of forest fires and fire protection on wildlife. For. Chron. 34 (1), 25–30. doi:10.5558/tfc34025-1
Cross, M. D., Root, K. V., Mehne, C. J., McGowan-Stinski, J., Pearsall, D., and Gillingham, J. C. (2015). Multi-scale responses of eastern massasauga rattlesnakes (Sistrurus catenatus) to prescribed fire. Am. Midl. Nat. 173 (2), 346–362. doi:10.1674/amid-173-02-346-362.1
Culhane, K., Sollmann, R., White, A. M., Tarbill, G. L., Cooper, S. D., and Young, H. S. (2022). Small mammal responses to fire severity mediated by vegetation characteristics and species traits. Ecol. Evol. 12, e8918. doi:10.1002/ece3.8918
Cunningham, C. X., Williamson, G. J., and Bowman, D. M. J. S. (2024). Increasing frequency and intensity of the most extreme wildfires on Earth. Nat. Ecol. Evol. 8, 1420–1425. doi:10.1038/s41559-024-02452-2
Day, N. J., Dunfield, K. E., Johnstone, J. F., Mack, M. C., Turetsky, M. R., Walker, X. J., et al. (2019). Wildfire severity reduces richness and alters composition of soil fungal communities in boreal forests of western Canada. Glob. Change Biol. 25, 2310–2324. doi:10.1111/gcb.14641
Day, N. J., White, A. L., Johnstone, J. F., Degré-Timmons, G. E., Cumming, S. G., Mack, M. C., et al. (2020). Fire characteristics and environmental conditions shape plant communities via regeneration strategy. Ecography 43, 1464–1474. doi:10.1111/ecog.05211
de Groot, W. J., Cantin, A. S., Flannigan, M. D., Soja, A. J., Gowman, L. M., and Newbery, A. (2013). A comparison of Canadian and Russian boreal forest fire regimes. For. Ecol. Manag. 294, 23–34. doi:10.1016/j.foreco.2012.07.033
Delcourt, C. J. F., Combee, A., Izbicki, B., Mack, M. C., Maximov, T., Petrov, R., et al. (2021). Evaluating the differenced normalized burn ratio for assessing fire severity using Sentinel-2 imagery in Northeast Siberian larch forests. Remote Sens. 13, 2311. doi:10.3390/rs13122311
DeMars, C. A., Serrouya, R., Mumma, M. A., Gillingham, M. P., McNay, R. S., and Boutin, S. (2019). Moose, caribou, and fire: have we got it right yet? Can. J. Zoology 97, 866–879. doi:10.1139/cjz-2018-0319
Denryter, K. A., Cook, R. C., Cook, J. G., and Parker, K. L. (2017). Straight from the caribou’s (Rangifer tarandus) mouth: detailed observations of tame caribou reveal new insights into summer–autumn diets. Can. J. Zool. 95, 81–94. doi:10.1139/cjz-2016-0114
Dickinson, M. B., Norris, J. C., Bova, A. S., Kremens, R. L., Young, V., and Lacki, M. J. (2010). Effects of wildland fire smoke on a tree-roosting bat: integrating a plume model, field measurements, and mammalian dose–response relationships. Can. J. For. Res. 40 (11), 2187–2203. doi:10.1139/x10-148
Dietze, M. C., Fox, A., Beck-Johnson, L. M., Betancourt, J. L., Hooten, M. B., Jarnevich, C. S., et al. (2018). Iterative near-term ecological forecasting: needs, opportunities, and challenges. Proc. Natl. Acad. Sci. U.S.A. 115, 1424–1432. doi:10.1073/pnas.1710231115
Dobbs, R. C. (1976). White spruce seed dispersal in Central British Columbia. For. Chron. 52, 225–228. doi:10.5558/tfc52225-5
Doherty, T. S., Geary, W. L., Jolly, C. J., Macdonald, K. J., Miritis, V., Watchorn, D. J., et al. (2022). Fire as a driver and mediator of predator–prey interactions. Biol. Rev. 97 (4), 1539–1558. doi:10.1111/brv.12853
Dole, H. E., Villamarin-Cortez, S., and Richards, L. A. (2023). Facing the flames: insect responses to megafires and changing fire regimes. Curr. Opin. Insect Sci. 60, 101129. doi:10.1016/j.cois.2023.101129
dos Anjos, A. G., Solé, M., and Benchimol, M. (2021). Fire effects on anurans: what we know so far? For. Ecol. Manag. 495, 119338. doi:10.1016/j.foreco.2021.119338
Doty, A. C., Currie, S. E., Stawski, C., and Geiser, F. (2018). Can bats sense smoke during deep torpor? Physiology Behav. 185, 31–38. doi:10.1016/j.physbeh.2017.12.019
Doty, A. C., Crawford, R. D., and Blakey, R. V. (2023). Bat roosting strategies and torpor expression in a wildfire-affected landscape during summer. Fire Ecol. 19 (1), 39. doi:10.1186/s42408-023-00199-y
Eckdahl, J. A., Kristensen, J. A., and Metcalfe, D. B. (2023). Climate and forest properties explain wildfire impact on microbial community and nutrient mobilization in boreal soil. Front. For. Glob. Change 6, 1136354. doi:10.3389/ffgc.2023.1136354
Eckdahl, J. A., Kristensen, J. A., and Metcalfe, D. B. (2024). Restricted plant diversity limits carbon recapture after wildfire in warming boreal forests. Commun. Earth Environ. 5, 186. doi:10.1038/s43247-024-01333-7
Ecosystem Classification Group (2007). Ecological regions of the Northwest Territories – Taiga plains. Yellowknife, NT, Canada: Environment and Natural Resources, Government of the Northwest Territories, viii + 173 pp. + folded insert map.
Ecosystem Classification Group. (2008). Ecological regions of the Northwest Territories – taiga shield. Yellowknife, NT, Canada: Environment and Natural Resources, Government of the Northwest Territories, viii + 146 pp. + insert map.
Ecosystem Classification Group (2010). Ecological regions of the Northwest Territories – cordillera. Yellowknife, NT, Canada: Environment and Natural Resources, Government of the Northwest Territories, X + 170 p. + insert map.
Ecosystem Classification Group (2012). Ecological regions of the Northwest Territories – southern Arctic. Yellowknife, NT, Canada: Environment and Natural Resources, Government of the Northwest Territories, x + 170 pp. + insert map.
Ellison, L. N. (1975). Density of Alaskan Spruce grouse before and after fire. The J. Wildlife Manage. 39 (3), 468–471. doi:10.2307/3800385
Erwin, W. J., and Stasiak, R. H. (1979). Vertebrate mortality during the burning of a reestablished prairie in Nebraska. Am. Midl. Nat. 101, 247–249. doi:10.2307/2424922
Ficken, M., McLaren, A., and Hailman, J., (2020). “Boreal chickadee (Poecile hudsonicus),” in Birds of the world, Editors A. F. Poole, and F. B. Gill Ithaca, NY: Cornell Lab of Ornithology.
Fisher, J. T. (1999). The influence of landscape structure on the distribution of the North American red squirrel (Tamiasciurus hudsonicus) in a heterogeneous boreal mosaic. MSc Thesis. Edmonton, AB: Department of Biological Sciences, University of Alberta.
Fisher, J. T., and Wilkinson, L. (2005). The response of mammals to forest fire and timber harvest in the North American boreal forest. Mammal Review 35, 51–81. doi:10.1111/j.1365-2907.2005.00053.x
Fontaine, J. B., and Kennedy, P. L. (2012). Meta-analysis of avian and small-mammal response to fire severity and fire surrogate treatments in US fire-prone forests. Ecol. Applicat. 22, 1547–1561. doi:10.1890/12-0009.1
Forney, R. R., and Peacock, M. M. (2024). The effects of fire on large-and medium-sized mammal communities: what do we know? A review. Mammal Rev. 54, 357–372. doi:10.1111/mam.12350
Fraser, R. H., Van der Sluijs, J., and Hall, R. J. (2017). Calibrating satellite-based indices of burn severity from UAV-derived metrics of a burned boreal forest in NWT, Canada. Remote Sensing 9, 279. doi:10.3390/rs9030279
French, N. H. F., Whitley, M. A., and Jenkins, L. K. (2016). Fire disturbance effects on land surface Abedo in Alaskan tundra. JGR Biogeosciences 121, 841–854. doi:10.1002/2015JG003177
Gauthier, S., Bernier, P., Kuuluvainen, T., Shvidenko, A. Z., and Schpaschenko, D. G. (2015). Boreal forest health and global change. Science 349, 819–822. doi:10.1126/science.aaa9092
Gibson, C. M., Chasmer, L. E., Thompson, D. K., Quinton, W. L., Flannigan, M. D., and Olefeldt, D. (2018). Wildfire as a major driver of recent permafrost thaw in boreal peatlands. Nat Commun 9, 3041. doi:10.1038/s41467-018-05457-1
Gillihan, S., and Byers, B. (2020). “Evening grosbeak (Coccothraustes vespertinus),” in Birds of the world. Editors A. F. Poole, and F. B. Gill (Ithaca, NY: Cornell Lab of Ornithology).
Girard, F., Payette, S., and Gagnon, R. (2008). Rapid expansion of lichen woodlands within the closed-crown boreal forest zone over the last 50 years caused by stand disturbances in eastern Canada. J Biogeography 35, 529–537. doi:10.1111/j.1365-2699.2007.01816.x
Girardin, M. P., Gaboriau, D. M., Ali, A. A., Gajewski, K., Briere, M. D., Bergeron, Y., et al. (2024). Boreal forest cover was reduced in the mid-Holocene with warming and recurring wildfires. Commun Earth Environ 5, 176. doi:10.1038/s43247-024-01340-8
Gomez Isaza, D. F., Cramp, R. L., and Franklin, C. E. (2022). Fire and rain: a systematic review of the impacts of wildfire and associated runoff on aquatic fauna. Global Change Biol. 28 (8), 2578–2595. doi:10.1111/gcb.16088
González-Fernández, A., Couturier, S., Dotor-Diego, R., Martínez-Díaz-González, R., and Sunny, A. (2024). Direct fire-induced reptile mortality in the Sierra Morelos natural protected area (Mexico). Herpetozoa 37, 213–226. doi:10.3897/herpetozoa.37.e116376
Granath, G., Evans, C. D., Strengbom, J., Fölster, J., Grelle, A., Strömqvist, J., et al. (2021). The impact of wildfire on biogeochemical fluxes and water quality in boreal catchments. Biogeosciences 18, 3243–3261. doi:10.5194/bg-18-3243-2021
Gregory, P. T., and Stewart, K. W. (1975). Long-distance dispersal and feeding strategy of the red-sided garter snake (Thamnophis sirtalis parietalis) in the Interlake of Manitoba. Can. J. of Zool. 53 (3), 238–245. doi:10.1139/z75-030
Greuel, R. J., Degré-Timmons, G. É., Baltzer, J. L., Johnstone, J. F., McIntire, E. J. B., Day, N. J., et al. (2021). Predicting patterns of terrestrial lichen biomass recovery following boreal wildfires. Ecosphere 12, e03481. doi:10.1002/ecs2.3481
Griffiths, A. D., and Brook, B. W. (2014). Effect of fire on small mammals: a systematic review. Inter. J. Wildland Fire 23, 1034–1043. doi:10.1071/wf14026
Grindal, S. D., and Brigham, R. M. (1999). Impacts of forest harvesting on habitat use by foraging insectivorous bats at different spatial scales. Ecoscience 6 (1), 25–34. doi:10.1080/11956860.1999.11952206
Guindon, L., Gauthier, S., Manka, F., Parisien, M.-A., Whitman, E., Bernier, P., et al. (2021). Trends in wildfire burn severity across Canada, 1985 to 2015. Can. J. For. Res. 51, 1230–1244. doi:10.1139/cjfr-2020-0353
Gustine, D. D., Parker, K. L., Lay, R. J., Gillingham, M. P., and Heard, D. C. (2006). Calf survival of woodland caribou in a multi-predator ecosystem. Wildlife Monogr. 165, 1–32. doi:10.2193/0084-0173(2006)165[1:csowci]2.0.co;2
Hale, S., Mendoza, L., Yeatman, T., Cooke, R., Doherty, T., Nimmo, D., et al. (2022). Evidence that post-fire recovery of small mammals occurs primarily via in situ survival. Div. Distribut. 28, 404–416. doi:10.1111/ddi.13283
Hall, R. J., Freeburn, J. T., De Groot, W. J., Pritchard, J. M., Lynham, T. J., and Landry, R. (2008). Remote sensing of burn severity: experience from western Canada boreal fires. Int. J. Wildland Fire 17, 476. doi:10.1071/WF08013
Halstead, B. J., Thompson, M. E., Amarello, M., Smith, J. J., Wylie, G. D., Routman, E. J., et al. (2019). Effects of prescribed fire on San Francisco gartersnake survival and movement. J. Wildlife Manage. 83 (1), 231–240. doi:10.1002/jwmg.21585
Hanes, C. C., Wang, X., Jain, P., Parisien, M.-A., Little, J. M., and Flannigan, M. D. (2019). Fire-regime changes in Canada over the last half century. Can. J. For. Res. 49, 256–269. doi:10.1139/cjfr-2018-0293
Hanes, C., Wotton, M., Woolford, D. G., Martell, D. L., and Flannigan, M. (2020). Preceding fall drought conditions and overwinter precipitation effects on spring wildland fire activity in Canada. Fire 3 (24), 24. doi:10.3390/fire3020024
Hayes, K., and Buma, B. (2021). Effects of short-interval disturbances continue to accumulate, overwhelming variability in local resilience. Ecosphere 12, e03379. doi:10.1002/ecs2.3379
Hekkala, A.-M., Jönsson, M., Kärvemo, S., Strengbom, J., and Sjögren, J. (2023). Habitat heterogeneity is a good predictor of boreal forest biodiversity. Ecol. Indicators 148, 110069. doi:10.1016/j.ecolind.2023.110069
Henderson, L. E., and Broders, H. G. (2008). Movements and resource selection of the northern long-eared myotis (Myotis septentrionalis) in a forest—agriculture landscape. J. Mammal. 89 (4), 952–963. doi:10.1644/07-mamm-a-214.1
Henderson, L. E., Farrow, L. J., and Broders, H. G. (2008). Intra-specific effects of forest loss on the distribution of the forest-dependent northern long-eared bat (Myotis septentrionalis). Biol. Conservat. 141 (7), 1819–1828. doi:10.1016/j.biocon.2008.04.028
Heon, J., Arseneault, D., and Parisien, M.-A. (2014). Resistance of the boreal forest to high burn rates. Proc. Natl. Acad. Sci. U. S. A. 111, 13888–13893. doi:10.1073/pnas.1409316111
Hollingsworth, T. N., Johnstone, J. F., Bernhardt, E. L., and Chapin, F. S. (2013). Fire severity filters regeneration traits to shape community assembly in Alaska’s boreal Forest. PLoS One 8, e56033. doi:10.1371/journal.pone.0056033
Holloway, J. E., Lewkowicz, A. G., Douglas, T. A., Li, X., Turetsky, M. R., Baltzer, J. L., et al. (2020). Impact of wildfire on permafrost landscapes: a review of recent advances and future prospects. Permafrost Periglacial 31, 371–382. doi:10.1002/ppp.2048
Hossack, B. R., and Pilliod, D. S. (2011). Amphibian responses to wildfire in the western United States: emerging patterns from short-term studies. Fire Ecol.y 7 (2), 129–144. doi:10.4996/fireecology.0702129
Hossack, B. R., Lowe, W. H., and Corn, P. S. (2013). Rapid increases and time-lagged declines in amphibian occupancy after wildfire. Conserv. Biol. 27 (1), 219–228. doi:10.1111/j.1523-1739.2012.01921.x
Houlahan, J. E., McKinney, S. T., Anderson, T. M., and McGill, B. J. (2017). The priority of prediction in ecological understanding. Oikos 126, 1–7. doi:10.1111/oik.03726
Howey, C. A., Dickinson, M. B., and Roosenburg, W. M. (2016). Effects of a landscape disturbance on the habitat use and behavior of the black racer. Copeia 104 (4), 853–863. doi:10.1643/ce-15-373
Hoy, E. E., Turetsky, M. R., and Kasischke, E. S. (2016). More frequent burning increases vulnerability of Alaskan boreal black spruce forests. Environ. Res. Lett. 11, 095001. doi:10.1088/1748-9326/11/9/095001
Hudak, A. T., Morgan, P., Bobbitt, M. J., Smith, A. M. S., Lewis, S. A., Lentile, L. B., et al. (2007). The relationship of multispectral satellite imagery to immediate fire effects. Fire Ecol. 3, 64–90. doi:10.4996/fireecology.0301064
Hunter, D. (2022). Effects of the Kenow wildfire on amphibian populations in Waterton Lakes National Park. Doctoral dissertation. Lethbridge, Canada. University of Lethbridge, Department of Biological Sciences, 68.
Hutchen, J., and Hodges, K. E. (2019). Foraging behaviour of snowshoe hares (Lepus americanus) in conifer forests regenerating after fire. Can. J. Zool. 97, 446–452. doi:10.1139/cjz-2018-0222
Jager, H. I., Long, J. W., Malison, R. L., Murphy, B. P., Rust, A., Silva, L. G., et al. (2021). Resilience of terrestrial and aquatic fauna to historical and future wildfire regimes in western North America. Ecol. Evolut. 11, 12259–12284. doi:10.1002/ece3.8026
Jain, P., Barber, Q. E., Taylor, S. W., Whitman, E., Castellanos Acuna, D., Boulanger, Y., et al. (2024). Drivers and impacts of the record-breaking 2023 wildfire season in Canada. Nat. Commun. 15, 6764. doi:10.1038/s41467-024-51154-7
Johnson, E. A., Miyanishi, K., and Weir, J. M. H. (1998). Wildfires in the western Canadian boreal forest: landscape patterns and ecosystem management. J Vegetat. Sci. 9, 603–610. doi:10.2307/3237276
Johnson, J. B., Edwards, J. W., Ford, W. M., and Gates, J. E. (2009). Roost tree selection by northern myotis (Myotis septentrionalis) maternity colonies following prescribed fire in a Central Appalachian Mountains hardwood forest. For. Ecol. Manage. 258 (3), 233–242. doi:10.1016/j.foreco.2009.04.008
Johnstone, J. F., Chapin, F. S., Hollingsworth, T. N., Mack, M. C., Romanovsky, V., and Turetsky, M. (2010a). Fire, climate change, and forest resilience in interior Alaska. Can. J. For. Res. 40, 1302–1312. doi:10.1139/X10-061
Johnstone, J. F., McIntire, E. J. B., Pedersen, E. J., King, G., and Pisaric, M. J. F. (2010b). A sensitive slope: estimating landscape patterns of forest resilience in a changing climate. Ecosphere 1, 1–21. doi:10.1890/ES10-00102.1
Johnstone, J. F., Allen, C. D., Franklin, J. F., Frelich, L. E., Harvey, B. J., Higuera, P. E., et al. (2016). Changing disturbance regimes, ecological memory, and forest resilience. Front. Ecol. Environ. 14, 369–378. doi:10.1002/fee.1311
Jones, G. M., and Tingley, M. W. (2022). Pyrodiversity and biodiversity: a history, synthesis, and outlook. Div. Distribut. 28, 386–403. doi:10.1111/ddi.13280
Jordaan, P. R., Steyl, J. C., Hanekom, C. C., and Combrink, X. (2020). Fire-associated reptile mortality in Tembe Elephant Park, South Africa. Fire Ecol. 16, 3–6. doi:10.1186/s42408-019-0066-4
Jorgensen, A. (2021). Wildlife forage recovery following boreal wildfire. Theses and dissertations (comprehensive), 2411. Available online at: https://scholars.wlu.ca/etd/2411.
Jorgensen, A. G., Alfaro-Sánchez, R., Cumming, S. G., White, A. L., Degré-Timmons, G. É., Day, N., et al. (2023). The influence of postfire recovery and environmental conditions on boreal vegetation. Ecosphere 14, e4605. doi:10.1002/ecs2.4605
Joyal, R., and Scherrer, B. (1978). Summer movements and feeding by moose in western Quebec. Can. Field Nat. 92, 252–258. doi:10.5962/p.346688
Jung, T. S. (2020). Bats in the changing boreal forest: response to a megafire by endangered little brown bats (Myotis lucifugus). Ecoscience 27 (1), 59–70. doi:10.1080/11956860.2019.1687084
Jung, T. S., Thompson, I. D., and Titman, R. D. (2004). Roost site selection by forest-dwelling male Myotis in central Ontario, Canada. For. Ecol. Manage. 202, 325–335. doi:10.1016/j.foreco.2004.07.043
Jung, T. S., Larter, N. C., and Powell, T. (2018). Early and late births in high-latitude populations of free-ranging Bison (Bison bison). .Can. Field Nat. 132, 219–222. doi:10.22621/cfn.v132i3.1983
Kasischke, E. S., and Turetsky, M. R. (2006). Recent changes in the fire regime across the North American boreal region – spatial and temporal patterns of burning across Canada and Alaska. Geophys. Res. Lett. 33, L09703. doi:10.1029/2006gl025677
Keeley, J. E. (2009). Fire intensity, fire severity and burn severity: a brief review and suggested usage. Int. J. Wildland Fire 18, 116. doi:10.1071/WF07049
Kharuk, V. I., Dvinskaya, M. L., Im, S. T., Golyukov, A. S., and Smith, K. T. (2022). Wildfires in the Siberian arctic. Fire 5, 106. doi:10.3390/fire5040106
Knaggs, M., Haché, S., Nielsen, S. E., Pankratz, R. F., and Bayne, E. (2020). Avian response to wildfire severity in a northern boreal region. Forests 11, 1330. doi:10.3390/f11121330
Knapp, E. E., Estes, B. L., and Skinner, C. N. (2009). Ecological effects of prescribed fire season: a literature review and synthesis for managers (No. PSW-GTR-224). Albany, CA: U.S. Department of Agriculture, Forest Service, Pacific Southwest Research Station. doi:10.2737/PSW-GTR-224
Konkolics, S., Dickie, M., Serrouya, R., Hervieux, D., and Boutin, S. (2021). A burning question: what are the implications of forest fires for woodland caribou? J. Wildl. Manag. 85, 1685–1698. doi:10.1002/jwmg.22111
Kotliar, N. B., Hejl, S. J., Hutto, R. L., Saab, V. A., Melcher, C. P., and McFadzen, M. E. (2002). “Effects of fire and post-fire salvage logging on avian communities in conifer-dominated forests of the western United States”, in effects of habitat fragmentation on birds in western landscapes: contrasts with paradigms from the eastern United States (Studies in Avian Biology No. 25). Editors T. L. George and D. S. Dobkin (Camarillo, CA: Cooper Ornithological Society). 49–64.
Kuntzemann, C. E., Whitman, E., Stralberg, D., Parisien, M., Thompson, D. K., and Nielsen, S. E. (2023). Peatlands promote fire refugia in boreal forests of northern Alberta, Canada. Ecosphere 14, e4510. doi:10.1002/ecs2.4510
Lacki, M. J., Cox, D. R., Dodd, L. E., and Dickinson, M. B. (2009). Response of northern bats (Myotis septentrionalis) to prescribed fires in eastern Kentucky forests. J. Mammal. 90 (5), 1165–1175. doi:10.1644/08-mamm-a-349.1
Landhäusser, S. M., Pinno, B. D., and Mock, K. E. (2019). Tamm Review: Seedling-based ecology, management, and restoration in aspen (Populus tremuloides). For. Ecol. Manage. 432, 231–245. doi:10.1016/j.foreco.2018.09.024
Lara, M. J., Genet, H., McGuire, A. D., Euskirchen, E. S., Ahang, Y., Brown, D. R. N., et al. (2016). Thermokarst rates intensify due to climate change and forest fragmentation in an Alaskan boreal forest lowland. Global Change Biol. 22, 816–829. doi:10.1111/gcb.13124
Larsen, K. W. (1987). Movements and behavior of migratory garter snakes, Thamnophis sirtalis. Can. J. Zool. 65 (9), 2241–2247. doi:10.1139/z87-339
Larter, N. C., and Gates, C. C. (1991). Diet and habitat selection of wood bison in relation to seasonal changes in forage quantity and quality. Can. J. Zool. 69, 2677–2685. doi:10.1139/z91-376
Latour, P. B., Maclean, N., and Poole, K. G. (1994). Movements of Martens, Martes americana, in burned and unburned taiga in the Mackenzie Valley Northwest Territories. Can. Field-Nat. 108, 351–354. doi:10.5962/p.356802
Lesmerises, R., Rebouillat, L., Dussault, C., and St-Laurent, M.-H. (2015). Linking GPS telemetry surveys and scat analyses helps explain variability in black bear foraging strategies. PLoS One 10, e0129857. doi:10.1371/journal.pone.0129857
Loeb, S. C., and Blakey, R. V. (2021). Bats and fire: a global review. Fire Ecol. 17, 29–18. doi:10.1186/s42408-021-00109-0
Lord, R., and Kielland, K. (2015). Effects of variable fire severity on forage production and foraging behavior of moose in winter. Alces. 51, 23–54.
Low, E. B., Florko, K. R. N., Mahoney, H. K., and Barclay, R. M. R. (2024). Effects of a severe wildfire on a bat community in the Canadian Rocky Mountains. For. Ecol. Manage. 563, 121983. doi:10.1016/j.foreco.2024.121983
Lyet, A., Cheylan, M., Prodon, R., and Besnard, A. (2009). Prescribed fire and conservation of a threatened mountain grassland specialist: a capture–recapture study on the Orsini’s viper in the French alps. Ani. Conservat. 12 (3), 238–248. doi:10.1111/j.1469-1795.2009.00245.x
MacArthur, D. L., and Dandy, J. W. T. (1982). Physiological aspects of overwintering in the boreal chorus frog (Pseudacris triseriata maculata). Comp. Biochem. Physiol. Part A Physiol. 72 (1), 137–141. doi:10.1016/0300-9629(82)90022-6
Mackey, B., Lindenmayer, D., Norman, P., Taylor, C., and Gould, S. (2021). Are fire refugia less predictable due to climate change? Environ. Res. Lett. 16, 114028. doi:10.1088/1748-9326/ac2e88
Mahony, M., Gould, J., Beranek, C. T., Callen, A., Clulow, J., Clulow, S., et al. (2022). A trait-based analysis for predicting impact of wildfires on frogs. Austr. Zool. 42 (2), 326–351. doi:10.7882/az.2022.021
Maier, J. A., Ver Hoef, J. M., McGuire, A. D., Bowyer, R. T., Saperstein, L., and Maier, H. A. (2005). Distribution and density of moose in relation to landscape characteristics: effects of scale. Can. J. For. Res. 35, 2233–2243. doi:10.1139/x05-123
Malison, R. L., and Baxter, C. V. (2010). The fire pulse: wildfire stimulates flux of aquatic prey to terrestrial habitats driving increases in riparian consumers. Can. J. Fisher. Aqu. Sci. 67 (3), 570–579. doi:10.1139/f10-006
Maloney, A. J. (2024). Impacts of wildfire and windfarm construction and operation on herpetofauna community ecology. Doctoral dissertation. Sudbury, Ontario: Laurentian University, 97.
Marchal, J., Cumming, S. G., and McIntire, E. J. B. (2020). Turning down the heat: vegetation feedbacks limit fire regime responses to global warming. Ecosystems 23, 204–216. doi:10.1007/s10021-019-00398-2
McDonald, L. A., Grayson, K. L., Lin, H. A., and Vonesh, J. R. (2018). Stage-specific effects of fire: effects of prescribed burning on adult abundance, oviposition habitat selection, and larval performance of Cope’s Gray Treefrog (Hyla chrysoscelis). For. Ecol. Manage. 430, 394–402. doi:10.1016/j.foreco.2018.08.008
McIntire, E. J. B., Chubaty, A. M., Cumming, S. G., Andison, D., Barros, C., Boisvenue, C., et al. (2022). PERFICT: a re-imagined foundation for predictive ecology. Ecol. Lett. 25, 1345–1351. doi:10.1111/ele.13994
McKenzie, D., Gedalof, Z. E., Peterson, D. L., and Mote, P. (2004). Climatic change, wildfire, and conservation. Conservat. Biol. 18, 890–902. doi:10.1111/j.1523-1739.2004.00492.x
McMullin, R. T., and Rapai, S. (2020). A review of reindeer lichen (Cladonia subgenus Cladina) linear growth rates. Rangifer 40, 15–26. doi:10.7557/2.40.1.4636
McMullin, R. T., Thompson, I. D., Lacey, B. W., and Newmaster, S. G. (2011). Estimating the biomass of woodland caribou forage lichens. Can. J. For. Res. 41, 1961–1969. doi:10.1139/x11-108
McNay, K. G., McNay, R. S., Sittler, K., and Rea, R. V. (2021). Moose use of the Mount McAllister burn in North-Central Central British Columbia: influence of burn severity and soil moisture. Alces. 57, 1–22. Available online at: https://alcesjournal.org/index.php/alces/article/view/273
Mekonnen, Z. A., Riley, W. J., Randerson, J. T., Grant, R. F., and Rogers, B. M. (2019). Expansion of high-latitude deciduous forests driven by interactions between climate warming and fire. Nat. Plants 5, 952–958. doi:10.1038/s41477-019-0495-8
Micheletti, T., Stewart, F. E. C., Cumming, S. G., Haché, S., Stralberg, D., Tremblay, J. A., et al. (2021). Assessing pathways of climate change effects in SpaDES: an application to boreal landbirds of Northwest Territories Canada. Front. Ecol. Evol. 9, 679673. doi:10.3389/fevo.2021.679673
Micheletti, T., Haché, S., Stralberg, D., Stewart, F. E. C., Chubaty, A. M., Barros, C., et al. (2023). Will this umbrella leak? A caribou umbrella index for boreal landbird conservation. Conservat. Sci. Prac. 5, e12908. doi:10.1111/csp2.12908
Mirabel, A., Girardin, M. P., Metsaranta, J., Way, D., and Reich, P. B. (2023). Increasing atmospheric dryness reduces boreal forest tree growth. Nat. Commun. 14, 6901. doi:10.1038/s41467-023-42466-1
Morandini, M., Mazzamuto, M. V., and Koprowski, J. L. (2023). Foraging behavior response of small mammals to different burn severities. Fire 6, 367. doi:10.3390/fire6090367
Mosnier, A., Ouellet, J.-P., and Courtois, R. (2008). Black bear adaptation to low productivity in the boreal forest. Écoscience 15, 485–497. doi:10.2980/15-4-3100
Muñoz, A., Felicísimo, Á. M., and Santos, X. (2019). Assessing the resistance of a breeding amphibian community to a large wildfire. Acta Oecol. 99, 103439. doi:10.1016/j.actao.2019.06.002
Murphy, S., Legge, S., Heathcote, J., and Mulder, E. (2009). The effects of early and late-season fires on mortality, dispersal, physiology and breeding of red-backed fairy-wrens (Malurus melanocephalus). Wild. Res. 37, 145–155. doi:10.1071/wr09007
Nelson, R. A., Folk, G. E., Pfeiffer, E. W., Craighead, J. J., Jonkel, C. J., and Steiger, D. L. (1983). Behavior, biochemistry, and hibernation in black, grizzly, and polar bears. Bears Their Biol. Manage. 5, 284–290. doi:10.2307/3872551
Norberg, U. M., and Rayner, J. M. (1987). Ecological morphology and flight in bats (Mammalia; Chiroptera): wing adaptations, flight performance, foraging strategy and echolocation. Philosoph. Transact. B 316, 335–427. doi:10.1098/rstb.1987.0030
O'Donoghue, M., and Bergman, C. M. (1992). Early movements and dispersal of juvenile snowshoe hares. Can. J. Zool. 70, 1787–1791. doi:10.1139/z92-246
Olson, R. A., Perryman, B. L., Petersburg, S., and Naumann, T. (2003). Fire effects on small mammal communities in Dinosaur National Monument. Western North Am. Nat. 63, 50–55. doi:10.1071/WF14026_AC
Ou, Y., Iyer, G., Clarke, L., Edmonds, J., Fawcett, A. A., Hultman, N., et al. (2021). Can updated climate pledges limit warming well below 2 °C? Science 374, 693–695. doi:10.1126/science.abl8976
Palm, E. C., Suitor, M. J., Joly, K., Herriges, J. D., Kelly, A. P., Hervieux, D., et al. (2022). Increasing fire frequency and severity will increase habitat loss for a boreal forest indicator species. Ecol. Appl. 32, e2549. doi:10.1002/eap.2549
Parisien, M.-A., Barber, Q. E., Bourbonnais, M. L., Daniels, L. D., Flannigan, M. D., Gray, R. W., et al. (2023a). Abrupt, climate-induced increase in wildfires in British Columbia since the mid-2000s. Commun. Earth Environ. 4, 309. doi:10.1038/s43247-023-00977-1
Parisien, M.-A., Barber, Q. E., Flannigan, M. D., and Jain, P. (2023b). Broadleaf tree phenology and springtime wildfire occurrence in boreal Canada. Global Change Biol. 29, 6106–6119. doi:10.1111/gcb.16820
Parker, K. L., Robbins, C. T., and Hanley, T. A. (1984). Energy expenditures for locomotion by mule deer and elk. J Wild. Manage. 48, 474. doi:10.2307/3801180
Parro, K., Metslaid, M., Renel, G., Sims, A., Stanturf, J. A., Jõgiste, K., et al. (2015). Impact of postfire management on forest regeneration in a managed hemiboreal forest, Estonia. Estonia. Can. J. For. Res. 45, 1192–1197. doi:10.1139/cjfr-2014-0514
Patriquin, K. J., and Barclay, R. M. R. (2003). Foraging by bats in cleared, thinned and unharvested boreal forest. J. Appl. Ecol. 40 , 601–770. doi:10.1046/j.1365-2664.2003.00831.x
Pellegrini, A. F. A., Ahlström, A., Hobbie, S. E., Reich, P. B., Nieradzik, L. P., Staver, A. C., et al. (2018). Fire frequency drives decadal changes in soil carbon and nitrogen and ecosystem productivity. Nature 553, 194–198. doi:10.1038/nature24668
Perera, A. H., and Buse, L. J. (2014). Ecology of wildfire residuals in boreal forests. Hoboken, NJ: John Wiley and Sons, Ltd.
Perera, A. H., Dalziel, B. D., Buse, L. J., and Routledge, R. G. (2009). Spatial variability of stand-scale residuals in Ontario’s boreal forest fires. Can. J. For. Res. 39, 945–961. doi:10.1139/X09-024
Pinno, B., and Errington, R. (2016). Burn severity dominates understory plant community response to fire in xeric jack pine forests. Forests 7, 83. doi:10.3390/f7040083
Poulin, J., D'Astous, É., Villard, M., Hejl, S. J., Newlon, K. R., McFadzen, M. E., et al. (2020). “Brown Creeper (Certhia americana), version 1.0,” in Birds of the world. Editor A. F. Poole (Ithaca, NY: Cornell Lab of Ornithology). doi:10.2173/bow.brncre.01
Prichard, S. J., Stevens-Rumann, C. S., and Hessburg, P. F. (2017). Tamm review: shifting global fire regimes: lessons from reburns and research needs. For. Ecol. Manage. 396, 217–233. doi:10.1016/j.foreco.2017.03.035
Rantanen, M., Karpechko, A.Yu., Lipponen, A., Nordling, K., Hyvärinen, O., Ruosteenoja, K., et al. (2022). The Arctic has warmed nearly four times faster than the globe since 1979. Commun. Earth Environ. 3, 168. doi:10.1038/s43247-022-00498-3
Ratcliffe, J. M., and Dawson, J. W. (2003). Behavioural flexibility: the little brown bat, Myotis lucifugus, and the northern long-eared bat, M. septentrionalis, both glean and hawk prey. Ani. Behav. 66, 847–856. doi:10.1006/anbe.2003.2297
Raymundo, A., Micheletti, T., Haché, S., Stralberg, D., Stewart, F. E. C., Tremblay, J. A., et al. (2024). Climate-sensitive forecasts of marked short-term and long-term changes in the distributions or abundances of Northwestern boreal landbirds. Clim. Change Ecol. 7, 100079. doi:10.1016/j.ecochg.2023.100079
Redburn, M. J., Strong, W. L., and Gates, C. C. (2008). Suitability of boreal mixedwood clearcuts as wood bison (Bison bison athabascae) foraging habitat in north-central Alberta, Canada. For. Ecol. Manage. 255, 2225–2235. doi:10.1016/j.foreco.2007.12.033
Regosin, J. V., Windmiller, B. S., and Reed, J. M. (2003). Terrestrial habitat use and winter densities of the wood frog (Rana sylvatica). J. Herpetol. 37 (2), 390–394. doi:10.1670/0022-1511(2003)037[0390:thuawd]2.0.co;2
Reid, K. A., Day, N. J., Alfaro-Sánchez, R., Johnstone, J. F., Cumming, S. G., Mack, M. C., et al. (2023). Black spruce (Picea mariana) seed availability and viability in boreal forests after large wildfires. Ann. For. Sci. 80, 4. doi:10.1186/s13595-022-01166-4
Reitsma, L., Hallworth, M., McMahon, M., and Conway, C. (2020). “Canada warbler (Cardellina canadensis),” in Birds of the world. Editors P. G. Rodewald, and B. K. Keeney (Ithaca, NY: Cornell Lab of Ornithology).
Rickbeil, G. J. M., Hermosilla, T., Coops, N. C., White, J. C., Wulder, M. A., and Lantz, T. C. (2018). Changing northern vegetation conditions are influencing barren ground caribou (Rangifer tarandus groenlandicus) post-calving movement rates. J. Biogeogr. 45, 702–712. doi:10.1111/jbi.13161
Risenhoover, K. L. (1989). Composition and quality of moose winter diets in Interior Alaska. J. Wild. Manage. 53, 568. doi:10.2307/3809178
Robinne, F.-N., Hallema, D. W., Bladon, K. D., and Buttle, J. M. (2020). Wildfire impacts on hydrologic ecosystem services in North American high-latitude forests: a scoping review. J. Hydrol. 581, 124360. doi:10.1016/j.jhydrol.2019.124360
Robinson, N. M., Leonard, S. W., Ritchie, E. G., Bassett, M., Chia, E. K., Buckingham, S., et al. (2013). REVIEW: refuges for fauna in fire-prone landscapes: their ecological function and importance. J. Appl. Ecol. 50 (6), 1321–1329. doi:10.1111/1365-2664.12153
Rochester, C. J., Brehme, C. S., Clark, D. R., Stokes, D. C., Hathaway, S. A., and Fisher, R. N. (2010). Reptile and amphibian responses to large-scale wildfires in southern California. J. Herpetol. 44 (3), 333–351. doi:10.1670/08-143.1
Romain, D. A., Obbard, M. E., and Atkinson, J. L. (2013). Temporal variation in food habits of the American black bear (Ursus americanus) in the boreal forest of northern Ontario. Can. Field-Nat. 127, 118–130. doi:10.22621/cfn.v127i2.1442
Rongstad, O. J., and Tester, J. R. (1971). Behavior and maternal relations of young snowshoe hares. J. Wild. Manage. 35, 338–346. doi:10.2307/3799610
Rusch, D. H., Destefano, S., Reynolds, M. C., and Lauten, D. (2020). Ruffed Grouse (Bonasa umbellus), version 1.0. birds of the world. Ithaca, NY: Cornell Lab of Ornithology. Available online at: https://birdsoftheworld-org.eu1.proxy.openathens.net/bow/species/rufgro/cur/introduction.
Russell, K. R., Van Lear, D. H., and Guynn, J. , D. C. (1999). Prescribed fire effects on herpetofauna: review and management implications. Wild. Soc. Bull. 27, 374–384. Available online at: http://www.jstor.org/stable/3783904
Sanderfoot, O. V., Bassing, S. B., Brusa, J. L., Emmet, R. L., Gillman, S. L., Swift, K., et al. (2022). A review of the effects of wildfire smoke on the health and behavior of wildlife. Environ. Res. Lett. 16, 123003. doi:10.1088/1748-9326/ac30f6
Santos, R. M. B., Sanches Fernandes, L. F., Pereira, M. G., Cortes, R. M. V., and Pacheco, F. A. L. (2015). Water resources planning for a river basin with recurrent wildfires. Sci. Total Environ. 526, 1–13. doi:10.1016/j.scitotenv.2015.04.058
Santos, X., Belliure, J., Gonçalves, J. F., and Pausas, J. G. (2021). Resilience of reptiles to megafires. Ecol. Appl. 32 (2), e2518. doi:10.1002/eap.2518
Santos, J. L., Sitters, H., Keith, D. A., Geary, W. L., Tingley, R., and Kelly, L. T. (2022). A demographic framework for understanding fire-driven reptile declines in the ‘land of the lizards. Global Ecol. Biogeogr. 31 (10), 2105–2119. doi:10.1111/geb.13520
Schaefer, J. A., and Pruitt, W. (1991). Fire and Woodland Caribou in Southeastern Manitoba. Wild. Monogr. 116, 3–39. Available online at: https://www.jstor.org/stable/3830581
Schetselaar, A., Andersen, T., and Burn, C. R. (2023). Performance of climate projections for Yukon and adjacent Northwest Territories, 1991–2020. Arctic 76, 244–264. doi:10.14430/arctic77263
Schieck, J., and Song, S. J. (2006). Changes in bird communities throughout succession following fire and harvest in boreal forests of western North America: literature review and meta-analyses. Can. J. For. Res. 36, 1299–1318. doi:10.1139/x06-017
Scholten, R. C., Jandt, R., Miller, E. A., Rogers, B. M., and Veraverbeke, S. (2021). Overwintering fires in boreal forests. Nature 593, 399–404. doi:10.1038/s41586-021-03437-y
Schurbon, J. M., and Fauth, J. E. (2003). Effects of prescribed burning on amphibian diversity in a southeastern US national forest. Conser. Biol. 17 (5), 1338–1349. doi:10.1046/j.1523-1739.2003.01514.x
Senior, K. L., Giljohann, K. M., McCarthy, M. A., Rainsford, F. W., and Kelly, L. T. (2021). Predicting mammal responses to pyrodiversity: from microbats to macropods. Biol. Conser. 256, 109031. doi:10.1016/j.biocon.2021.109031
Shaw, R. E., James, A. I., Tuft, K., Legge, S., Cary, G. J., Peakall, R., et al. (2021). Unburnt habitat patches are critical for survival and in situ population recovery in a small mammal after fire. J. Appl. Ecol. 58, 1325–1335. doi:10.1111/1365-2664.13846
Shine, R., LeMaster, M. P., Moore, I. T., Olsson, M. M., and Mason, R. T. (2001). Bumpus in the snake den: effects of sex, size, and body condition on mortality of red-sided garter snakes. Evolution 55, 598–604. doi:10.1111/j.0014-3820.2001.tb00792.x
Shively, R. D., Crouse, J. A., Thompson, D. P., and Barboza, P. S. (2019). Is summer food intake a limiting factor for boreal browsers? Diet, temperature, and reproduction as drivers of consumption in female moose. PLoS One 14, e0223617. doi:10.1371/journal.pone.0223617
Silva, J. A., Nielsen, S. E., Lamb, C. T., Hague, C., and Boutin, S. (2019). Modelling lichen abundance for woodland caribou in a fire-driven boreal landscape. Forests 10, 962. doi:10.3390/f10110962
Silva, J. A., Nielsen, S. E., McLoughlin, P. D., Rodgers, A. R., Hague, C., and Boutin, S. (2020). Comparison of pre-fire and post-fire space use reveals varied responses by woodland caribou (Rangifer tarandus caribou) in the Boreal Shield. Can. J. Zool. 98, 751–760. doi:10.1139/cjz-2020-0139
Simon, N. P., Schwab, F. E., Baggs, E. M., and McTaggart-Cowan, G. I. (1998). Distribution of small mammals among successional and mature forest types in western Labrador. Can. Field Nat. 112, 441–445. doi:10.5962/p.358446
Skatter, H. G., Charlebois, M. L., Eftestøl, S., Tsegaye, D., Colman, J. E., Kansas, J. L., et al. (2017). Living in a burned landscape: Woodland caribou (Rangifer tarandus caribou) use of postfire residual patches for calving in a high fire – low anthropogenic Boreal Shield ecozone. Can. J. Zool. 95, 975–984. doi:10.1139/cjz-2016-0307
Smith, L. J., Holycross, A. T., Painter, C. W., and Douglas, M. E. (2001). Montane rattlesnakes and prescribed fire. Southwestern Naturalist 46, 54–61. doi:10.2307/3672373
Smith, H. G., Sheridan, G. J., Lane, P. N. J., Nyman, P., and Haydon, S. (2011). Wildfire effects on water quality in forest catchments: a review with implications for water supply. J. Hydrol. 396, 170–192. doi:10.1016/j.jhydrol.2010.10.043
Snider, E. A., Cryan, P. M., and Wilson, K. R. (2013). Roost selection by western long-eared myotis (Myotis evotis) in burned and unburned piñon–juniper woodlands of southwestern Colorado. J. Mammal. 94 (3), 640–649. doi:10.1644/11-mamm-a-153.1
Snow, D. P. (1996). Effects of prescribed burning on willow ptarmigan in Newfoundland. M. Sc. dissertation. Stevens Point, WI: University of Wisconsin.
Soja, A. J., Tchebakova, N. M., French, N. H. F., Flannigan, M. D., Shugart, H. H., Stocks, B. J., et al. (2007). Climate-induced boreal forest change: predictions versus current observations. Global and Planet. Change 56, 274–296. doi:10.1016/j.gloplacha.2006.07.028
Sommers, M., and Flannigan, M. D. (2022). Green islands in a sea of fire: the role of fire refugia in the forests of Alberta. Environ. Rev. 30, 402–417. doi:10.1139/er-2021-0115
Soverel, N. O., Perrakis, D. D. B., and Coops, N. C. (2010). Estimating burn severity from Landsat dNBR and RdNBR indices across western Canada. Rem. Sens. Environ. 114, 1896–1909. doi:10.1016/j.rse.2010.03.013
Species at Risk Committee (2016). Species status report for wood Bison (Bison bison athabascae) in the Northwest Territories. Yellowknife, NT: Species at Risk Committee.
Species at Risk Committee (2022). Species status report for Boreal Caribou (Rangifer tarandus caribou) in the Northwest Territories. Yellowknife, NT: Species at Risk Committee.
Spence, C., Galloway, J. M., Hedstrom, N., Kokelj, S. V., Kokelj, S. A., Muise, P., et al. (2025). Process synchrony a key control of resilience in a subarctic freshwater system. Earth’s Future 13, e2024EF005518. doi:10.1029/2024ef005518
Splawinski, T. B., Greene, D. F., Michaletz, S. T., Gauthier, S., Houle, D., and Bergeron, Y. (2019). Position of cones within cone clusters determines seed survival in black spruce during wildfire. J. For. Res. 49, 121–127. doi:10.1139/cjfr-2018-0209
Spring, A., Skinner, K., Simba, M., Nelson, E., Baltzer, J., Swanson, H., et al. (2019). “Taking care of the land: an interdisciplinary approach to community-based food systems assessment in Kakisa, Northwest Territories, Canada,” in Sustainable food System assessment. Editor A. Blay- Palmer 1st edition (London, England: Taylor and Francis Group).
Steel, Z. L., Miller, J. E. D., Ponisio, L. C., Tingley, M. W., Wilkin, K., Blakey, R., et al. (2024). A roadmap for pyrodiversity science. J. Biogeogr. 51, 280–293. doi:10.1111/jbi.14745
Stephens, S. L., Burrows, N., Buyantuyev, A., Gray, R. W., Keane, R. E., Kubian, R., et al. (2014). Temperate and boreal forest mega-fires: characteristics and challenges. Front. Ecol. Environ. 12, 115–122. doi:10.1890/120332
Stewart, F. E. C., Micheletti, T., Cumming, S. G., Barros, C., Chubaty, A. M., Dookie, A. L., et al. (2023). Climate-informed forecasts reveal dramatic local habitat shifts and population uncertainty for northern boreal caribou. Ecol. Appl. 33, e2816. doi:10.1002/eap.2816
Stocks, B. J. (1989). Fire behavior in mature jack pine. Can. J. For. Res. 19, 783–790. doi:10.1139/x89-119
Stralberg, D., Solymos, P., Docherty, T. D. S., Crosby, A. D., Wilgenburg, S. L. W., Knight, E. C., et al. (2025). A generalized modeling framework for spatially extensive species abundance prediction and population estimation. Ecosphere 16, e70405. doi:10.1002/ecs2.70405
Strickland, D., and Ouellet, H. (2020). “Canada Jay (Perisoreus canadensis),” in Birds of the world. Editor A. F. Poole (Ithaca, NY: Cornell Lab of Ornithology).
Sutherland, E. F., and Dickman, C. R. (1999). Mechanisms of recovery after fire by rodents in the Australian environment: a review. Wild. Res. 26, 405–419. doi:10.1071/wr97045
Talucci, A., Loranty, M. M., Holloway, J. E., Rogers, B. M., Alexander, H. D., Baillargeon, N., et al. (2024). Permafrost-wildfire interactions: active layer thickness estimates for paired burned and unburned sites in northern high-latitudes. Earth Syst. Sci. Data 17, 2887–2909. doi:10.5194/essd-17-2887-2025
Tavakol, A., Rahmani, V., and Harrington, J. (2020). Temporal and spatial variations in the frequency of compound hot, dry, and windy events in the central United States. Sci. Rep. 10, 15691. doi:10.1038/s41598-020-72624-0
Thomas, J. P., and Jung, T. S. (2019). Life in a northern town: rural villages in the boreal forest are islands of habitat for an endangered bat. Ecosphere 10, e02563. doi:10.1002/ecs2.2563
Thompson, I. D., Wiebe, P. A., Mallon, E., Rodgers, A. R., Fryxell, J. M., Baker, J. A., et al. (2015). Factors influencing the seasonal diet selection by woodland caribou (Rangifer tarandus tarandus) in boreal forests in Ontario. Can. J. Zool. 93, 87–98. doi:10.1139/cjz-2014-0140
Thompson, D. K., Parisien, M. -A., Morin, J., Millard, K., Larsen, C. P. S., et al. (2017). Fuel accumulation in a high-frequency boreal wildfire regime: from wetland to upland. Can. J. For. Res. 47, 957–964. doi:10.1139/cjfr-2016-0475
Timmermann, H. R., and McNicol, J. G. (1988). Moose habitat needs. Forestry Chronicle 64, 238–245. doi:10.5558/tfc64238-3
Tomchuk, P. (2019). Differential habitat selection of black bears, gray wolves, and boreal caribou in the Boreal Shield of Saskatchewan. Doctoral dissertation. Saskatoon, SK: University of Saskatchewan.
Turetsky, M. R., Baltzer, J. L., Johnstone, J. F., Mack, M. C., McCann, K., and Schuur, E. A. G. (2017). Losing legacies, ecological release, and transient responses: key challenges for the future of northern ecosystem science. Ecosystems 20, 23–30. doi:10.1007/s10021-016-0055-2
Turner, M. G., and Seidl, R. (2023). Novel disturbance regimes and ecological responses. Annu. Rev. Ecol. Evol. Syst. 54, 63–83. doi:10.1146/annurev-ecolsys-110421-101120
Turner, M. G., Romme, W. H., Reed, R. A., and Tuskan, G. A. (2003). Post-fire aspen seedling recruitment across the Yellowstone (USA) Landscape. Lands. Ecol. 18, 127–140. doi:10.1023/a:1024462501689
Turner, M. G., Braziunas, K. H., Hansen, W. D., and Harvey, B. J. (2019). Short-interval severe fire erodes the resilience of subalpine lodgepole pine forests. Proc. Natl. Acad. Sci. U. S. A. 116, 11319–11328. doi:10.1073/pnas.1902841116
Vanbianchi, C. M., Murphy, M. A., and Hodges, K. E. (2017). Canada lynx use of burned areas: conservation implications of changing fire regimes. Ecol. Evolution. 7, 2382–2394. doi:10.1002/ece3.2824
Veraverbeke, S., Rogers, B. M., Goulden, M. L., Jandt, R. R., Miller, C. E., Wiggins, E. B., et al. (2017). Lightning as a major driver of recent large fire years in North American boreal forests. Nat. Clim. Change 7, 529–534. doi:10.1038/nclimate3329
Veraverbeke, S., Delcourt, C. J. F., Jujavskaya, E., Mack, M., Walker, X., Hessilt, T., et al. (2021). Direct and longer term carbon emissions from arctic-boreal fires: a short review of recent advances. Curr. Opin. Environ. Sci. Health 23, 100277. doi:10.1016/j.coesh.2021.100277
Viau, J. P., Sigouin, D., and St-Laurent, M. H. (2024). Seasonal shifts in the habitat selection patterns of male American Marten (Martes americana) at a fine spatial scale. J. Mammal. 105, 740–751. doi:10.1093/jmammal/gyae048
Volkmann, L. A., and Hodges, K. E. (2024). Effects of burn severity and postfire salvage logging on carnivore communities in montane forests. J. Mammal. 105, 390–403. doi:10.1093/jmammal/gyad135
Walker, X. J., Rogers, B. M., Baltzer, J. L., Cumming, S. G., Day, N. J., Goetz, S. J., et al. (2018). Cross-scale controls on carbon emissions from boreal forest megafires. Glob. Change Biol. 24, 4251–4265. doi:10.1111/gcb.14287
Walker, X. J., Baltzer, J. L., Cumming, S. G., Day, N. J., Ebert, C., Goetz, S., et al. (2019). Increasing wildfires threaten historic carbon sink of boreal forest soils. Nature 572, 520–523. doi:10.1038/s41586-019-1474-y
Walker, X. J., Rogers, B. M., Veraverbeke, S., Johnstone, J. F., Baltzer, J. L., Barrett, K., et al. (2020). Fuel availability not fire weather controls boreal wildfire severity and carbon emissions. Nat. Clim. Chang. 10, 1130–1136. doi:10.1038/s41558-020-00920-8
Wang, Y., Hogg, E. H., Price, D. T., Edwards, J., and Williamson, T. (2014). Past and projected future changes in moisture conditions in the Canadian boreal forest. For. Chronicle 90, 678–691. doi:10.5558/tfc2014-134
Wang, X., Thompson, D. K., Marshall, G. A., Tymstra, C., Carr, R., and Flannigan, M. D. (2015). Increasing frequency of extreme fire weather in Canada with climate change. Clim. Change 130, 573–586. doi:10.1007/s10584-015-1375-5
Wang, X., Studens, K., Parisien, M.-A., Taylor, S. W., Candau, J.-N., Boulanger, Y., et al. (2020). Projected changes in fire size from daily spread potential in Canada over the 21st century. Environ. Res. Lett. 15, 104048. doi:10.1088/1748-9326/aba101
Webb, J. K., and Shine, R. (2008). Differential effects of an intense wildfire on survival of sympatric snakes. J. Wild. Manage. 72 (6), 1394–1398. doi:10.2193/2007-515
Weeden, R. B. (1963). Management of ptarmigan in North America. J. Wild. Manage. 27 (4), 672–683. doi:10.2307/3798484
Wei, X., Hayes, D. J., Fraver, S., and Chen, G. (2018). Global pyrogenic carbon production during recent decades has created the potential for a large, long-term sink of atmospheric CO2. JGR Biogeosciences 123, 3682–3696. doi:10.1029/2018JG004490
Weir, J. M. H., Johnson, E. A., and Miyanishi, K. (2000). Fire frequency and the spatial age mosaic of the mixedwood boreal forest in western Canada. Ecol. Appl. 10, 1162–1177. doi:10.1890/1051-0761(2000)010[1162:ffatsa]2.0.co;2
Wheeler, K. I., Dietze, M. C., LeBauer, D., Peters, J. A., Richardson, A. D., Ross, A. A., et al. (2024). Predicting spring phenology in deciduous broadleaf forests: NEON phenology forecasting community challenge. Agricult. For. Meteorol. 345, 109810. doi:10.1016/j.agrformet.2023.109810
Whitman, E., Parisien, M.-A., Thompson, D. K., and Flannigan, M. D. (2019). Short-interval wildfire and drought overwhelm boreal forest resilience. Sci. Rep. 9, 18796. doi:10.1038/s41598-019-55036-7
Whitman, E., Barber, Q. E., Jain, P., Parks, S. A., Guindon, L., Thompson, D. K., et al. (2024). A modest increase in fire weather overcomes resistance to fire spread in recently burned boreal forests. Glob. Change Biol. 30, e17363. doi:10.1111/gcb.17363
Whitman, T., Whitman, E., Woolet, J., Flannigan, M. D., Thompson, D. K., and Parisien, M.-A. (2019). Soil bacterial and fungal response to wildfires in the Canadian boreal forest across a burn severity gradient. Soil Biol. Biochem. 138, 107571. doi:10.1016/j.soilbio.2019.107571
Widén, P. (1994). Habitat quality for raptors: a field experiment. J. Avian Biol. 25, 219–223. doi:10.2307/3677078
Wilgers, D. J., and Horne, E. A. (2007). Spatial variation in predation attempts on artificial snakes in a fire-disturbed tallgrass prairie. Southwestern Nat. 52 (2), 263–270. doi:10.1894/0038-4909(2007)52[263:svipao]2.0.co;2
Working Group on General Status of NWT Species (2021). NWT species 2021–2025 – general status ranks of wild species in the Northwest Territories. Yellowknife, NT: Department of Environment and Natural Resources, Government of the Northwest Territories, 389.
Wotton, B. M., Flannigan, M. D., and Marshall, G. A. (2017). Potential climate change impacts on fire intensity and key wildfire suppression thresholds in Canada. Environ. Res. Lett. 12, 095003. doi:10.1088/1748-9326/aa7e6e
Xu, M. D., Dong, W. J., Long, X. Z., Yang, X. W., Han, X. Y., Cui, L. Y., et al. (2024). Impact of wildfire ash on skin and gut microbiomes and survival of Rana dybowskii. J. Hazar. Mater. 474, 134729. doi:10.1016/j.jhazmat.2024.134729
Zhang, X., Vincent, L. A., Hogg, W. D., and Niitsoo, A. (2000). Temperature and precipitation trends in Canada during the 20th century. Atmosphere-Ocean 38, 395–429. doi:10.1080/07055900.2000.9649654
Zhang, X., Flato, G., Kirchmeier-Young, M., Vincent, L., Wan, H., Wang, X., et al. (2019). “Changes in Temperature and Precipitation across Canada; chapter 4,” in Canada’s changing climate report. Editors E. Bush, and D. S. Lemmen (Ottawa, Ontario: Government of Canada), 112–193.
Zhao, J., Yue, C., Wang, J., Hantson, S., Wang, X., He, B., et al. (2024). Forest fire size amplifies postfire land surface warming. Nature 633, 828–834. doi:10.1038/s41586-024-07918-8
Keywords: wildlife, novel disturbance regime, boreal wildfire, ecological resilience, state changes, habitat changes
Citation: Baltzer JL, Haché S, Hodson J, Ahmed MR, Carrière S, Coyle M, McIntire EJB, McLaren A, Riordan-Short E, Turetsky MR, van der Sluijs J, Wilson J and Woodworth BK (2025) Impacts of novel wildfire disturbance on landcover and wildlife in boreal North America. Front. Environ. Sci. 13:1504568. doi: 10.3389/fenvs.2025.1504568
Received: 01 October 2024; Accepted: 30 September 2025;
Published: 14 November 2025.
Edited by:
Torben Røjle Christensen, Aarhus University, DenmarkReviewed by:
Michael Pollock, NOAA Fisheries, United StatesJohan Eckdahl, University of California, Berkeley, United States
Copyright © 2025 Baltzer, Haché, Hodson, Ahmed, Carrière, Coyle, McIntire, McLaren, Riordan-Short, Turetsky, van der Sluijs, Wilson and Woodworth. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jennifer L. Baltzer, amJhbHR6ZXJAd2x1LmNh
†These authors share first authorship