 Hao Li1†
Hao Li1† Zeyu Yang2†Xiao Guo1Hangyu Li1Xing Li3
Zeyu Yang2†Xiao Guo1Hangyu Li1Xing Li3 Yantao Wu4Ying Han5Zhiyong Li1*
Yantao Wu4Ying Han5Zhiyong Li1* Jinghui Zhang1Bailing Miao6
Jinghui Zhang1Bailing Miao6 Cunzhu Liang1
Cunzhu Liang1- 1Key Laboratory of Mongolian Plateau Ecology and Resource Utilization of Ministry of Education, Inner Mongolia Key Laboratory of Grassland Ecology and the Candidate State Key Laboratory of Ministry of Science and Technology, School of Ecology and Environment, Inner Mongolia University, Hohhot, China
- 2Houma North Copper and Copper Industry Co., Ltd., Linfen, China
- 3Inner Mongolia Academy of Forestry Sciences, Hohhot, China
- 4College of Life Sciences, Inner Mongolia University, Hohhot, China
- 5Inner Mongolia Academy of Governance, Hohhot, China
- 6Inner Mongolia Meteorological Institute, Hohhot, China
Soil aggregates are vital to soil structure, influencing grassland ecosystem health and sustainability. As a dominant land use in grasslands, grazing acts as a key driver of ecosystem structure and function, yet the responses of aggregate size distribution and stability to grazing intensity across soil depths remain insufficiently understood. Here, we conducted an eight-year sheep grazing experiment in a semi-arid steppe with four treatments: no grazing (NG), light (LG), moderate (MG), and heavy grazing (HG). Aggregate characteristics were assessed across four soil layers (topsoil: 0–10 cm, midsoil: 10–30 cm, subsoil: 30–40 cm, Whole soil: 0–40 cm) using dry- and wet-sieving. Results showed that dry-sieved aggregate size distribution was more sensitive to grazing than wet-sieved aggregates. HG dry-sieved >2 mm aggregates significantly increased by 4.98% in whole soil, with MG and HG enhancing large aggregates in topsoil (5.80% and 6.31%, respectively), while LG and MG < 0.25 mm aggregates increased by 8.28% and 11.01%, respectively, in the subsoil. HG had no significant effect on subsoil <0.25 mm aggregates. Aggregate stability was highest under HG, particularly in the subsoil, where both mechanical and water stability were significantly improved 0.14 mm and 0.32 mm, respectively. Soil aggregate stability varied notably with soil depth. Mechanical stability consistently increased with depth, irrespective of grazing intensity. In contrast, water stability displayed distinct patterns among treatments: a progressive increase with depth under NG, a topsoil minimum under LG and MG, and relatively constant values across depths under HG. Soil aggregate stability was jointly regulated by grazing intensity coupled with soil depth, significantly influenced by soil bulk density, microbial biomass carbon, and belowground biomass. Although higher grazing intensity can improve aggregate stability, it may also have adverse effects on soil porosity, bulk density, and belowground biomass. Thus, intermediate grazing is critical for maintaining aggregate stability and the long-term sustainability of typical steppe ecosystems.
1 Introduction
Grasslands constitute the most extensively distributed and expansive terrestrial ecosystem, covering approximately 40% of the Earth’s land surface and storing nearly 10% of global soil organic carbon (Dlamini et al., 2016; Zhao et al., 2017; Buisson et al., 2022). As the predominant land-use practice in grasslands, grazing plays a pivotal role in shaping ecosystem structure and function (Wang et al., 2019b; Wan et al., 2020). However, excessive grazing has increasingly contributed to the degradation of soil quality and the decline of associated ecosystem services (Han et al., 2021; Hu et al., 2024b). Existing research on grazing effects in grassland ecosystems has predominantly focused on aboveground plant communities and soil nutrient dynamics (Jiang et al., 2023), while investigations into soil aggregate size distribution and stability remain relatively limited.
Soil aggregates are fundamental structural units and key indicators of soil quality and fertility (Singh and Singh, 1996; Even and Cotrufo, 2024). Particle size and distribution reflect the degree of aggregation and structural stability (Tisdall and Oades, 1982). Aggregate formation is primarily driven by soil organic carbon and microbial activity (Hannula et al., 2023), with microaggregates offering a more stable environment for long-term carbon sequestration (Puget et al., 2000). Aggregate stability is closely tied to soil structure and regulates processes such as water movement and retention (Dou et al., 2020), microbial activity, plant growth, and soil erosion resistance (An et al., 2008). Furthermore, Water-stable aggregates provide better physical protection for SOC than dry-sieved aggregates due to their greater stability and persistent microstructure (Li et al., 2025). Maintaining high soil aggregate stability is essential for improving windbreak and sand fixation, conserving water and soil, and promoting soil carbon sequestration. (An et al., 2013; Li et al., 2023). A deeper understanding of aggregate size distribution and stability under grazing is critical for sustainable grassland management and provides a solid basis for assessing soil health in grazed ecosystems.
Livestock directly and indirectly affect vegetation, soil physicochemical properties, and microbial communities through grazing, trampling, and urination, thereby influencing the formation of soil aggregates (Wiesmeier et al., 2012; Wen et al., 2016). Many studies have shown that grazing leads to the fragmentation of large soil aggregates under wet sieving, increases the proportion of microaggregates, and reduces overall stability (Zhou et al., 2010; Reszkowska et al., 2011). However, some research indicates that light grazing promotes the formation of >1 mm dry-sieved macroaggregates in alpine meadow grasslands, thereby enhancing aggregate stability (Wang et al., 2020; Zhang et al., 2022). Grazing prohibition has been found to improve the water stability of aggregates in semiarid grassland (Liu et al., 2021), while heavy grazing increases the proportion of >2 mm water-stable aggregates in desert steppes (Fan et al., 2021). Therefore, current conclusions about how grazing intensity affects soil aggregate size distribution and stability are still inconsistent. Additionally, some studies suggest that grazing may enhance the mechanical stability of aggregates in typical grasslands while reducing their water stability (Zhang et al., 2022). Research indicates that soil depth is a key driver of aggregate carbon (Pan et al., 2023). In alpine meadows, the influence of root biomass and microbial biomass carbon on aggregate-associated carbon diminishes with increasing soil depth (Pan et al., 2023). However, most previous studies have overlooked two key issues: (1) the divergent responses of dry-sieved aggregates reflecting mechanical stability and wet-sieved aggregates reflecting water stability to grazing; and (2) the depth dependent changes in aggregate composition and stability induced by grazing. This study investigates the effects of eight-year grazing treatments on soil aggregate composition and stability in Xilingol typical steppe. Based on preliminary results, we propose two hypotheses: (H1) Dry-sieved aggregates, whose stability depends primarily on physical binding, will be more resistant to grazing disturbance. (H2) Intermediate grazing will enhance the mechanical stability of dry-sieved aggregates by promoting compaction, while simultaneously reducing the water stability of wet-sieved aggregates. The research addresses two fundamental questions: (1) the differential response mechanisms of dry- versus wet-sieved aggregates to grazing, and (2) whether the contrasting effects of grazing intensity on aggregate composition and stability persist across soil-depth gradients.
2 Materials and methods
2.1 Study area
The study area is situated at the Xilinhot National Climate Observatory (44°08′N, 116°19′E, elevation 1,118 m, Figure 1A). The climate is semi-arid continental temperate, with an average annual precipitation of approximately 281 mm, over 85% of which occurs between May and September (Han et al., 2023). The average annual temperature is around 2.4 °C, and the soil is classified as Kastanozems (Wu et al., 2022). The vegetation is characteristic of a semi-arid Stipa grandis steppe, dominated by S. grandis and Leymus chinensis. The community is also composed of perennial herbs such as Cleistogenes squarrosa, Agropyron cristatum, and Anemarrhena asphodeloides, along with annual and biennial herbs such as Salsola collina, Dysphania aristata, and Chenopodium acuminatum.

Figure 1. Study area location and experimental design for grazing manipulation, including a landscape map: (A) shows study area location; (B) shows the randomized block grazing experiment designed with four treatments, namely no grazing (NG), light grazing (LG), moderate grazing (MG) and heavy grazing (HG). Each plot covered an area of 1.44 ha; (C) shows the landscape map of the four different grazing treatments.
2.2 Grazing experiment design and sample collection
The grazing experimental area was established in a relatively flat region with uniform vegetation types. The experiment followed a completely randomized block design with four grazing treatments, each replicated three times: no grazing (NG), light grazing (LG), moderate grazing (MG), and heavy grazing (HG). The stocking rates were 0, 2, 4, and 8 sheep per hectare per day, respectively. Grazing intensity was regulated by adjusting the number of grazing days at a fixed stocking rate. Specifically, 28 two-year-old Ujumqin sheep grazed sequentially on 12 pasture plots for 0, 3, 6, and 12 days, with each grazing cycle lasting 21 days (Figure 1B). The grazing experiment began in 2013 and comprised four grazing periods each year, from early June to late September, between 07:00 and 18:00. Samples were collected in August 2020, the experiment's eighth year.
Based on vegetation characteristics, three sampling points were established per plot to collect soil samples from the 0–10 cm, 10–30 cm, and 30–40 cm depth intervals. Bulk density was determined using 5 cm diameter cutting rings, with samples oven-dried at 105 °C to constant weight. Undisturbed soil samples (10 × 10 × 10 cm aluminum boxes) were collected for aggregate analysis, manually fragmented to approximately 1 cm3 after removing visible roots and gravel, then air-dried. At each aggregate sampling location, duplicate 200 g soil samples were collected, one air-dried for physicochemical analysis (including C/N content) and the other sieved through a 2 mm mesh and stored at −4 °C for microbial biomass carbon determination. Root biomass was sampled using an auger, washed thoroughly, and oven-dried at 65 °C to constant weight. A total of 108 samples were collected for analyses of bulk density, aggregate stability, physicochemical properties, and below-ground biomass.
2.3 Soil aggregate size distribution and analysis
A dry-sieving method was used to quantify mechanically stable aggregates. Dry soil samples from each layer (200 g) were sieved through 2 mm, 1 mm, 0.5 mm, and 0.25 mm meshes at a frequency of 30 revolutions per minute for 2 min (Márquez et al., 2004). After sieving, the aggregates retained on each sieve were collected and weighed. The mass percentage of mechanically stable aggregates for each particle size was then calculated. A wet-sieving method was used to quantify water-stable aggregates. A 50 g subsample of each aggregate size fraction obtained by dry-sieving was immersed in distilled water for 5 min to displace trapped air (Dai et al., 2024). The analyzer was set at 50 cycles per minute with a 3 cm sieve movement. Aggregates were washed into aluminum boxes, dried at 65 °C to constant weight, and the mass percentage of water-stable aggregates was calculated.
This study selected the content of each particle-size aggregate Equation 1 and mean weight diameter (MWD) Equation 2 as evaluation indicators for soil aggregate composition and stability (He et al., 2021). The calculation formulas are as follows:
Content of each particle-size aggregate:
Mean Weight Diameter (MWD):
where Wi denotes the dry weight of the ith level aggregate component (g);
2.4 Measurement of soil physico-chemical properties
In this research, we conducted systematic analyses on physicochemical soil samples. Specifically, soil pH was assessed employing the potentiometric method (soil: water = 1 : 2.5). Total soil carbon and nitrogen were quantified using an elemental analyzer (Elementar Vario MacroCube, Germany), and inorganic carbon content was measured via gas volumetric analysis. The organic carbon content in the soil was derived by subtracting inorganic carbon from the total soil carbon. Microbial carbon and nitrogen contents were determined through the chloroform fumigation-extraction technique.
2.5 Statistical analysis
First, one-way ANOVA and the LSD method were employed in SPSS 23.0 to analyze the effects of varying grazing intensities on belowground biomass, soil physicochemical properties, aggregate size distribution, and stability across different soil depths. Subsequently, Pearson correlation analysis was conducted in R 4.3.3 to examine the relationships between dry- and wet-sieved aggregate characteristics and environmental factors. Two-way ANOVA was applied to assess the effects of grazing intensity, soil depth, and their interaction on aggregate stability. Redundancy analysis (RDA) and hierarchical partitioning (HP) were performed to explore the associations between soil aggregate properties and physicochemical characteristics, followed by the selection of key environmental variables for structural equation modeling (SEM). Principal component analysis (PCA) was used to reduce the dimensionality of soil organic carbon and total nitrogen data, which served as indicators of soil nutrient status. Within the SEM framework, the direct and indirect effects of grazing intensity, soil depth, soil nutrients, microbial biomass carbon, bulk density, and belowground biomass on the mechanical and water stability of aggregates were assessed. All data were standardized prior to analysis.
3 Results
3.1 Characteristics of soil aggregate composition under different grazing intensities
The response of dry-sieved and wet-sieved soil aggregates to grazing intensity follows a similar pattern, with a stronger effect on dry-sieved aggregates (Figure 2). In the whole soil (0–40 cm), heavy grazing (HG) significantly increased dry-sieved aggregates (>2 mm), but had no effect on wet-sieved aggregates (>2 mm). HG resulted in significantly higher dry-sieved and wet-sieved aggregates (>2 mm) compared to light grazing (LG) and moderate grazing (MG). All grazing intensities reduced dry-sieved 1–2 mm aggregates, and LG also decreased the wet-sieved 1–2 mm fraction. Both MG and HG reduced dry-sieved 0.5–1 mm aggregates, while only LG affected wet-sieved 0.5–1 mm aggregates (p < 0.05). MG and HG also reduced dry-sieved 0.25–0.5 mm aggregates, with all grazing intensities decreasing the corresponding wet-sieved fractions (p < 0.05). LG significantly increased dry-sieved <0.25 mm aggregates, but had no effect on wet-sieved <0.25 mm aggregates.
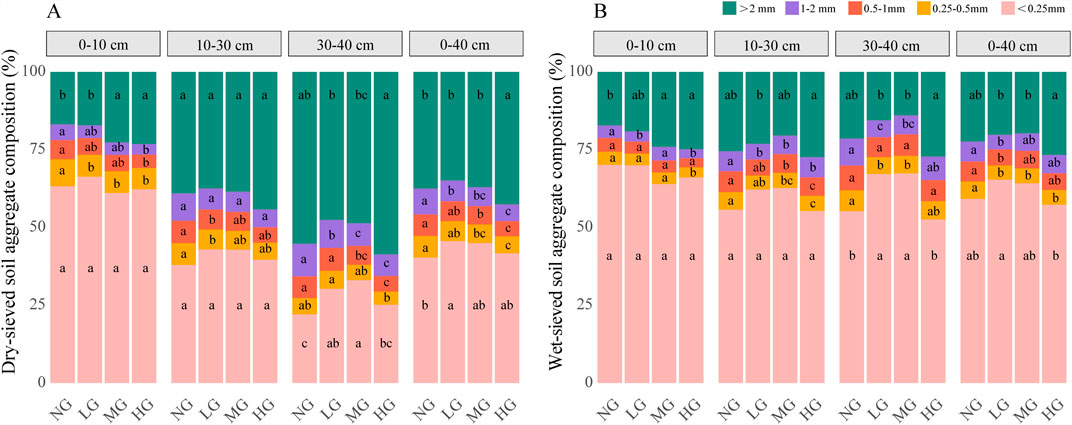
Figure 2. Responses of soil aggregates composition to grazing intensity in different soil layers: (A) dry-sieved soil aggregate composition, (B) wet-sieved soil aggregate composition. Note: NG, no grazing; LG, light grazing; MG, moderate grazing; HG, heavy grazing. Different letters indicate significant differences between different grazing intensities (p < 0.05).
Differential responses of dry- and wet-sieved soil aggregate size distribution to grazing intensity across soil depths (Figure 2). In the topsoil (0–10 cm), both MG and HG significantly increased the proportion of >2 mm aggregates under both dry and wet sieving. All grazing intensities significantly reduced the proportion of 0.25–0.5 mm aggregates under dry sieving, with a significant reduction under wet sieving observed only under HG. No significant differences were found for <0.25 mm aggregates under either sieving method. The proportions of 1–2 mm and 0.5–1 mm aggregates showed no consistent response to grazing intensity under both sieving methods. In the midsoil (10–30 cm), there was no discernible pattern in the variation of aggregates of different particle sizes under varying grazing intensities. In the subsoil (30–40 cm), the proportion of >2 mm aggregates was significantly higher under HG than LG and MG, but not different from NG. All grazing intensities significantly reduced 1–2 mm dry-sieved aggregates, while only LG and MG reduced wet-sieved aggregates. MG and HG significantly decreased 0.5–1 mm dry-sieved aggregates, with no significant differences for wet-sieved aggregates. LG and MG significantly reduced 0.25–0.5 mm wet-sieved aggregates, but no significant differences were observed for dry-sieved aggregates. LG and MG significantly increased the proportion of <0.25 mm aggregates, while HG had no significant effect.
3.2 Characteristics of soil aggregate stability under different grazing intensities
The mechanical and water stability of soil aggregates exhibited generally consistent responses to grazing intensity. However, their stability characteristics differed significantly with changes in soil depth. Within the whole soil, HG exhibited higher mechanical and water stability of aggregates compared to other treatments (p > 0.05), with mechanical stability being significantly greater than under LG (Figures 3D,H). In the topsoil, both mechanical and water stability of soil aggregates showed an increasing trend with grazing intensity (p > 0.05; Figures 3A,E). Across all soil layers, HG enhanced the mechanical and water stability of soil aggregates compared to LG and MG (Figure 3). Specifically, In the subsoil, both mechanical and water stability under HG were significantly higher than LG (Figures 3A–C,E–G). Mechanical stability of soil aggregates consistently increased with increasing soil depth irrespective of grazing intensity (Figures 3A–C). In contrast, water stability varied distinctly with depth: it increased progressively under no grazing (NG), reached subsurface minima under light (LG) and moderate (MG) grazing, and remained largely invariant across depths under heavy grazing (HG) (Figures 3E–G).
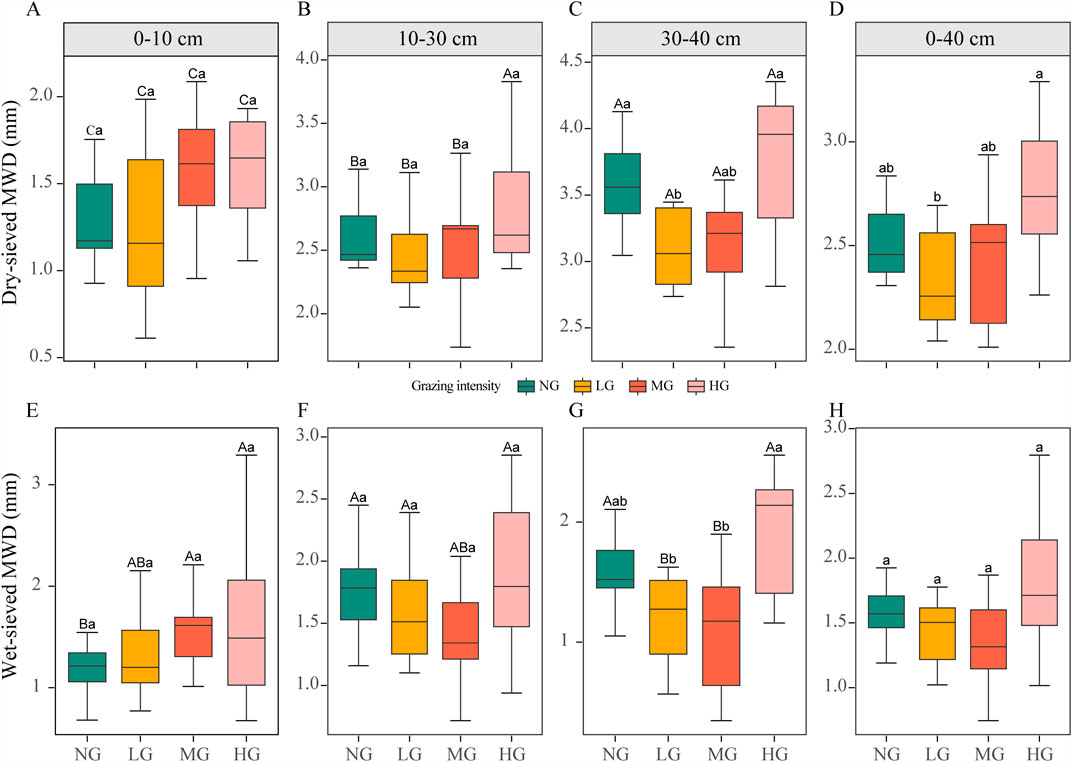
Figure 3. Responses of the soil aggregates stability to grazing intensity in different soil layers: (A–D) dry-sieved MWD, (E–H) wet-sieved MWD. Note: NG, no grazing; LG, light grazing; MG, moderate grazing; HG, heavy grazing; MWD, mean weight diameter. Different uppercase letters indicate significant differences (p < 0.05) between soil depths within the same grazing intensity (p < 0.05). Different letters indicate significant differences between different grazing intensities (p < 0.05).
3.3 Drivers of soil aggregate composition and stability under different grazing intensities
Both mechanical and water stability were strongly, negatively correlated with the <0.25 mm aggregate fraction and strongly, positively correlated with the >2 mm fraction. In dry-sieving, 1–2 mm aggregates are significantly associated with mechanical stability in topsoil and midsoil, while wet-sieving reveals significant positive correlations across midsoil, subsoil, and whole soil. Dry-sieved 0.25–0.5 mm aggregates show a significant negative correlation with mechanical stability, except in topsoil. Other size fractions exhibit weak or inconsistent associations with stability (Supplementary Figure S1).
Soil bulk density (BD) was positively associated with the content of wet-sieved macroaggregates (>2 mm) and their water stability (Figures 4A,B; Supplementary Figure S2). In contrast, aboveground biomass (AGB) and below-ground biomass (BGB) had a negative impact on aggregate size distribution and stability, showing inverse correlations with >2 mm fractions from both dry and wet sieving, as well as overall aggregate stability (Figures 5A,B; Supplementary Figure S2). Similarly, microbial biomass carbon (MBC) was negatively correlated with both mechanical and water stability, while also influencing wet-sieved aggregate size distribution (Figures 4A,B,D) In typical steppe ecosystems, grazing intensity indirectly influenced soil aggregate stability by significantly altering BD and BGB (Figure 5). However, BD was not significantly associated with mechanical stability (Figure 5A). Soil depth exerted direct but opposing effects on aggregate stability - enhancing mechanical stability while reducing water stability. Additionally, it indirectly influenced aggregate stability by mediating soil nutrient availability, bulk density (BD), belowground biomass (BGB), and microbial biomass carbon (MBC) (Figure 5; Supplementary Table S1).

Figure 4. Redundancy analysis (RDA) and hierarchical partitioning (HP) of soil aggregate characteristics in relation to soil properties in the 0–40 cm Soil Layer: (A) redundancy analysis of dry-sieved soil aggregate characteristics in relation to soil properties, (B) redundancy analysis of wet-sieved soil aggregate characteristics in relation to soil properties, (C) hierarchical partitioning of dry-sieved soil aggregate characteristics in relation to soil properties (D) hierarchical partitioning of wet-sieved soil aggregate characteristics in relation to soil properties Note: BGB, below-ground biomass; pH, soil acidity; BD, soil bulk density; TC, soil total carbon; SOC, soil organic carbon; MBC, microbial biomass carbon; TN, soil total nitrogen; MBN, microbial biomass nitrogen; MWD, mean weight diameter.
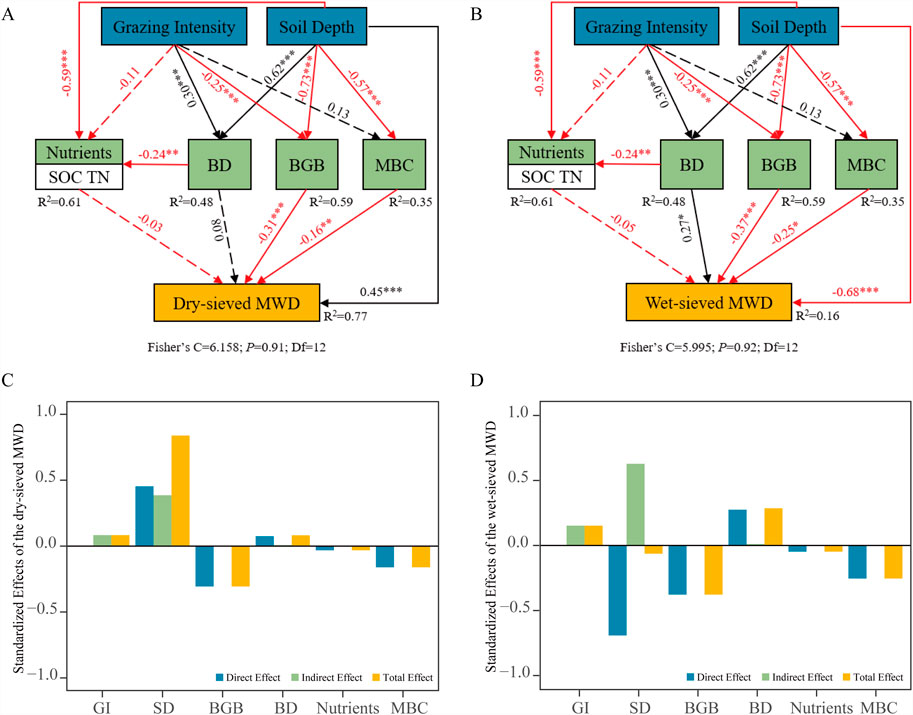
Figure 5. The structural equation model (SEM) depicts the direct and indirect effects of grazing intensity and soil depth on soil aggregate stability: (A) the direct and indirect effects of grazing intensity and soil depth on dry-sieved MWD, (B) the direct and indirect effects of grazing intensity and soil depth on wet-sieved MWD, (C) standardized effects of the dry-sieved MWD, (D) standardized effects of the wet-sieved MWD. Note: GI, grazing intensity; SD, soil depth; BGB, below-ground biomass; BD, soil bulk density; SOC, soil organic carbon; TN, soil total nitrogen; MBC, microbial biomass carbon.
4 Discussion
4.1 Grazing effects on soil aggregate composition and stability
This study found that MG and HG significantly increased >2 mm aggregates in both dry- and wet-sieving within the topsoil (Figure 2). Comparable studies have shown that moderate grazing boosts large aggregate content in soils of the Tibetan Plateau (Zhang et al., 2019). The composition, distribution, and stability of soil aggregates are crucial in forming optimal soil structure (Six et al., 2000). The size distribution of aggregates influences biogeochemical cycling and energy flow, while their stability dictates the soil’s resistance to erosion (Zhang et al., 2022; Peng et al., 2023). Higher macroaggregate (>0.25 mm) content and a larger mean weight diameter are positively associated with improved aggregate stability (Yang and Wander, 1998; Zhao et al., 2022). In this study, the content of >2 mm aggregates and the mechanical stability of aggregates under HG were significantly higher than those under LG in whole soil (Figures 2A, 3D). Heavy grazing, by promoting controlled soil compaction, improves soil aeration and permeability, thereby enhancing the content and stability of >2 mm soil aggregates (Reinhart et al., 2021). Across the whole soil profile, the 1–2, 0.5–1 and 0.25–0.5 mm aggregate fractions attained their highest contents under no grazing (NG) (Figure 2). This is likely due to the greater accumulation of surface litter, which enhanced organic binding agents and promoted the formation of larger aggregates (>0.25 mm) (Wang et al., 2020).
In this study, the content of wet-sieved aggregates <0.25 mm was consistently lower than that of dry-sieved aggregates in whole soil, with the disparity increasing significantly with soil depth (Figure 2). This pattern reflects the disintegration of unstable aggregates into <0.25 mm fractions during wet-sieving treatment, resulting in substantially higher proportions of smaller aggregates in the wet-sieved samples (Elliott, 1986; Sodhi et al., 2009). Our findings demonstrate that grazing intensity similarly influences both mechanical and water stability of soil aggregates in typical steppe ecosystems, with mechanical stability exhibiting greater sensitivity (Figure 3). This contrasts with previous studies reporting increased mechanical stability but decreased water stability under grazing (Zhang et al., 2022), suggesting these responses may be ecosystem-dependent, potentially due to differences in grassland types (Pérès et al., 2013). Our results revealed distinct depth-dependent patterns for the two aggregate stability metrics: mechanical stability consistently increased with soil depth (Figures 3A–C; Supplementary Table S1), while water stability followed a unimodal trend, initially increasing and then decreasing in deeper layers (Figures 3E–G). The decline in water stability with depth reflects the increasing proportion of non-water-stable mechanical aggregates, which disintegrate into smaller fractions during wet-sieving (Xiao et al., 2021), consistent with established pedogenic processes.
4.2 Factors affecting the stability of soil aggregates under grazing
Grazing influences soil aggregates by affecting soil pore volume, pore size, and connectivity (Parwada and Van Tol, 2019; Levi et al., 2020). Overgrazing reduces aboveground vegetation cover (Reszkowska et al., 2011), leading to soil compaction and a decrease in aggregate stability (Zhang et al., 2018; Zhang et al., 2022). In contrast, moderate grazing can enhance aggregate stability (Ju et al., 2023). In this study, grazing indirectly influenced soil aggregate stability by significantly affecting BD and BGB (Figure 5; Supplementary Figure S2; Supplementary Table S2). Moderate grazing increases soil bulk density and reduces erodible sediment content (Reinhart et al., 2021), thereby affecting aggregate stability. Soil aggregate stability is highly correlated with root activity (Obia et al., 2016), as dense root systems disrupt aggregates, leading to an increase in microaggregates (<0.25 mm) and a subsequent decrease in stability (Zhou et al., 2022). These findings are consistent with our results, such as the significant positive correlation between BD and the content and stability of >2 mm wet-sieved aggregates (Figures 5A,B; Supplementary Figure S2B). Additionally, in agroecosystems, soil aggregate stability has also been found to exhibit a significant positive correlation with soil bulk density (Dai et al., 2024). Conversely, BGB negatively correlates with the content and stability of >2 mm dry- and wet-sieved aggregates (Figures 4, 5A,B). The mean weight diameter (MWD) of water-stable aggregates was positively, though not significantly, correlated with soil carbon stock (Supplementary Figure S2B). While heavy grazing did not significantly enhance carbon stock, it consistently maintained higher levels across all soil depths compared to light and moderate grazing (Supplementary Table S2). Despite reduced carbon inputs from litter under heavy grazing (Wang et al., 2023; Wang et al., 2025), the concentrated deposition of organic matter via livestock excreta may have transiently enhanced the soil organic carbon pool (Fan et al., 2021), facilitating organic binding within aggregates and modestly improving their stability. Contrary to our findings, some studies have reported that high grazing intensity reduces soil carbon stocks by accelerating decomposition and disrupting carbon cycling (Wang et al., 2023). This discrepancy may stem from the relatively short grazing duration in our study, during which soils had not yet entered a degraded state. In this study, Microbial biomass carbon showed an significant negative correlation with soil aggregate stability (Figures 5A,B). It has been found that microaggregates sup-port more diverse microbial communities than macroaggregates (Bach et al., 2018; Hu et al., 2023). The increase in aggregate stability was mainly driven by > 2 mm aggregates (Supplementary Figures S1, S2), leading to a negative correlation between microbial biomass carbon and stability. Notably, a similarly significant negative correlation has been observed between enhanced macroaggregate stability and microbial carbon use efficiency in calcareous karst soils (Hu et al., 2024a).
Although numerous studies have shown that overgrazing reduces vegetation cover and disrupts biological soil crusts (Thomas, 2012), leading to a significant decrease in large aggregates and an increase in microaggregates, thereby lowering aggregate stability (Wang et al., 2019a; Zhang et al., 2022), our study found that in whole soil layer, both dry- and wet-sieved >2 mm aggregates were highest under HG, while <0.25 mm aggregates (microaggregates) were highest under LG (Figure 2). Other studies have also reported that 15 years of heavy grazing in desert grasslands increased >2 mm soil aggregates (Fan et al., 2021). Our results showed no significant differences in the mechanical and water stability of soil aggregates across treatments in the 0–30 cm layer. However, both mechanical and water stability were significantly higher under HG compared to LG in subsoil (Figures 3C, G, p < 0.05). This indicates a notable variation in the response of aggregate stability to grazing intensity across soil depths. Similarly, a previous study found that soil organic carbon in deeper layers (30–50 cm) responded more significantly to light grazing than in surface and midsoil layers (Wu et al., 2022). Our study shows that after 8 years of grazing, aggregate size is more sensitive to grazing intensity than stability. Higher grazing intensity significantly increases the content of large aggregates and shows a trend toward enhanced stability, whereas lower grazing intensity increases the content of microaggregates in deeper soil layers and tends to reduce stability. Heavy grazing is generally associated with reduced soil aggregate stability due to nutrient depletion (Zhou et al., 2017; Hu et al., 2023). However, in this study, 8 years of heavy grazing did not significantly reduce key soil nutrients such as organic carbon and total nitrogen (Supplementary Table S2). Although vegetation cover declined and biomass decreased, the lack of appreciable nutrient losses indicates that the soil had not become degraded. This may explain the slight but positive short-term effect of heavy grazing on aggregate stability, likely reflecting a temporary response rather than long-term improvement.
Despite the observed trends, this study has two main limitations. First, the single 8-year evaluation lacks temporal resolution to assess the progressive impacts of grazing on aggregate formation. Second, the absence of particle-size-specific analyses of organic carbon and microbial communities within aggregates limits our understanding of underlying mechanisms. Future research should adopt chronosequence designs and incorporate size-specific biogeochemical and microbial analyses to better reveal how grazing intensity shapes soil aggregate dynamics over time.
5 Conclusion
Investigations of soil aggregate characteristics in a typical Inner Mongolian steppe under varying grazing intensities revealed distinct depth-dependent responses. HG significantly increased dry-sieved aggregates >2 mm (whole soil) but showed no effect on wet-sieved counterparts. Grazing consistently reduced 1–0.25 mm aggregates across all treatments. Notably, MG and HG enhanced >2 mm aggregates in topsoil, while LG and MG increased <0.25 mm aggregates in subsoil. Aggregate mechanical stability increased with soil depth, whereas water stability followed a unimodal pattern. HG consistently improved both stability metrics, particularly showing significant enhancement in subsoil compared to LG. These findings demonstrate the dual regulation of aggregate stability by grazing intensity and soil depth. In the typical steppe, 8 years of heavy grazing degraded plant community structure and reduced biomass, yet the soil itself had not entered a degraded state; as a result, heavy grazing produced a modest, short-lived increase in soil aggregate stability. We recommend layer-specific assessments of aggregate properties under different grazing regimes, with particular consideration given to the temporal dynamics and cumulative effects of grazing management.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
The animal studies were approved by the Bioethics Committee of Inner Mongolia University, and the requirement for ethical approval was waived for this animal study. The studies were conducted in accordance with local legislation and institutional requirements. Written informed consent was obtained from the owners for the participation of their animals in this study.
Author contributions
HL: Conceptualization, Methodology, Writing – original draft, Writing – review and editing. ZY: Conceptualization, Validation, Writing – original draft. XG: Formal Analysis, Validation, Writing – original draft. HL: Validation, Writing – original draft. XL: Conceptualization, Writing – original draft. YW: Formal Analysis, Writing – original draft. YH: Formal Analysis, Writing – original draft. ZL: Conceptualization, Formal Analysis, Writing – original draft. JZ: Conceptualization, Writing – original draft. BM: Conceptualization, Writing – original draft. CL: Conceptualization, Methodology, Writing – original draft.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the National Natural Science Foundation of China, grant number 32271656, the National Key Research and Development Program of China, grant number 2022YFF1300601, the Natural Science Foundation of Inner Mongolia Autonomous Region of China, grant number 2024FX27.
Acknowledgments
We appreciate fieldwork assistance from Yingshun Wang, Wen Wang and Zheng Wang.
Conflict of interest
Author ZY was employed by Houma North Copper and Copper Industry Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2025.1610919/full#supplementary-material
References
An, S. S., Huang, Y. M., Zheng, F. L., and Yang, J. G. (2008). Aggregate characteristics during natural revegetation on the loess Plateau. Pedosphere 18 (6), 809–816. doi:10.1016/s1002-0160(08)60077-6
An, S. S., Darboux, F., and Cheng, M. (2013). Revegetation as an efficient means of increasing soil aggregate stability on the loess Plateau (China). Geoderma 209, 75–85. doi:10.1016/j.geoderma.2013.05.020
Bach, E. M., Williams, R. J., Hargreaves, S. K., Yang, F., and Hofmockel, K. S. (2018). Greatest soil microbial diversity found in micro-habitats. Soil Biol. Biochem. 118, 217–226. doi:10.1016/j.soilbio.2017.12.018
Buisson, E., Archibald, S., Fidelis, A., and Suding, K. N. (2022). Ancient grasslands guide ambitious goals in grassland restoration. Science 377 (6606), 594–598. doi:10.1126/science.abo4605
Dai, W., Feng, G. Y., Huang, Y. B., Adeli, A., and Jenkins, J. N. (2024). Influence of cover crops on soil aggregate stability, size distribution and related factors in a no-till field. Soil. Tillage Res. 244, 106197. doi:10.1016/j.still.2024.106197
Dlamini, P., Chivenge, P., and Chaplot, V. (2016). Overgrazing decreases soil organic carbon stocks the Most under dry climates and low soil pH: a meta-analysis shows. Agric. Ecosyst. Environ. 221, 258–269. doi:10.1016/j.agee.2016.01.026
Dou, Y. X., Yang, Y., An, S. S., and Zhu, Z. L. (2020). Effects of different vegetation restoration measures on soil aggregate stability and erodibility on the Loess Plateau, China. Catena 185, 104294. doi:10.1016/j.catena.2019.104294
Elliott, E. T. (1986). Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci. Soc. Am. J. 50 (3), 627–633. doi:10.2136/sssaj1986.03615995005000030017x
Even, R. J., and Cotrufo, M. F. (2024). The ability of soils to aggregate, more than the state of aggregation, promotes protected soil organic matter formation. Geoderma 442, 116760. doi:10.1016/j.geoderma.2023.116760
Fan, J. L., Jin, H., Zhang, C. H., Zheng, J. J., Zhang, J., and Han, G. D. (2021). Grazing intensity induced alternations of soil microbial community composition in aggregates drive soil organic carbon turnover in a desert steppe. Agric. Ecosyst. Environ. 313, 107387. doi:10.1016/j.agee.2021.107387
Han, S., Delgado-Baquerizo, M., Luo, X. S., Liu, Y. R., Van Nostrand, J. D., Chen, W. L., et al. (2021). Soil aggregate size-dependent relationships between microbial functional diversity and multifunctionality. Soil Biol. Biochem. 154, 108143. doi:10.1016/j.soilbio.2021.108143
Han, Y., Wu, Y., Cui, J., Li, H., Li, H., Zhang, J., et al. (2023). Temporal stability of grazed grassland ecosystems alters response to climate variability, while resistance stability remains unchanged. Agronomy-Basel 13 (8), 2030. doi:10.3390/agronomy13082030
Hannula, S. E., Jongen, R., and Morriën, E. (2023). Grazing by collembola controls fungal induced soil aggregation. Fungal Ecol. 65, 101284. doi:10.1016/j.funeco.2023.101284
He, X. X., Huang, Y. Z., Zhang, Q. C., Ye, S. M., and Wang, S. Q. (2021). Distribution of organic carbon fractions in soil aggregates in Chinese fir plantations with different stand ages. Ecol. Process. 10 (1), 49. doi:10.1186/s13717-021-00321-5
Hu, Y., Yu, G. L., Zhou, J. Q., Li, K. H., Chen, M., Abulaizi, M., et al. (2023). Grazing and reclamation-induced microbiome alterations drive organic carbon stability within soil aggregates in alpine steppes. Catena 231, 107306. doi:10.1016/j.catena.2023.107306
Hu, P. L., Zhang, W., Nottingham, A. T., Xiao, D., Kuzyakov, Y., Xu, L., et al. (2024a). Lithological controls on soil aggregates and minerals regulate microbial carbon use efficiency and necromass stability. Environ. Sci. Technol. 58 (48), 21186–21199. doi:10.1021/acs.est.4c07264
Hu, Y., Zhang, H., Sun, X., Zhang, B., Wang, Y., Rafiq, A., et al. (2024b). Impact of grassland degradation on soil multifunctionality: linking to protozoan network complexity and stability. Sci. total Environ. 929, 172724. doi:10.1016/j.scitotenv.2024.172724
Jiang, S., Zhang, J. H., Tang, Y. W., Li, Z. Y., Liu, H. M., Wang, L. X., et al. (2023). Plant functional traits and biodiversity can reveal the response of ecosystem functions to grazing. Sci. Total Environ. 899, 165636. doi:10.1016/j.scitotenv.2023.165636
Ju, W. L., Moorhead, D. L., Shen, G. T., Cui, Y. X., and Fang, L. C. (2023). Soil aggregate development and associated microbial metabolic limitations alter grassland carbon storage following livestock removal. Soil Biol. Biochem. 177, 108907. doi:10.1016/j.soilbio.2022.108907
Levi, E. M., Archer, S. R., Throop, H. L., and Rasmussen, C. (2020). Soil-litter mixing promotes decomposition and soil aggregate formation on contrasting geomorphic surfaces in a shrub-invaded sonoran Desert grassland. Plant Soil 450 (1-2), 397–415. doi:10.1007/s11104-020-04508-1
Li, J. Y., Chen, P., Li, Z. G., Li, L. Y., Zhang, R. Q., Hu, W., et al. (2023). Soil aggregate-associated organic carbon mineralization and its driving factors in rhizosphere soil. Soil Biol. Biochem. 186, 109182. doi:10.1016/j.soilbio.2023.109182
Li, J., Wang, Q., Zhu, B., and Cui, M. (2025). Influence of Plateau pika disturbance on soil organic carbon in Zoige alpine grasslands — based on the role of soil aggregate. J. Mt. Sci. 22 (7), 2555–2573. doi:10.1007/s11629-024-9403-4
Liu, D. D., Ju, W. L., Jin, X. L., Li, M. D., Shen, G. T., Duan, C. J., et al. (2021). Associated soil aggregate nutrients and controlling factors on aggregate stability in semiarid grassland under different grazing prohibition timeframes. Sci. Total Environ. 777, 146104. doi:10.1016/j.scitotenv.2021.146104
Márquez, C. O., Garcia, V. J., Cambardella, C. A., Schultz, R. C., and Isenhart, T. M. (2004). Aggregate-size stability distribution and soil stability. Soil Sci. Soc. Am. J. 68 (3), 725–735. doi:10.2136/sssaj2004.7250
Obia, A., Mulder, J., Martinsen, V., Cornelissen, G., and Borresen, T. (2016). In situ effects of biochar on aggregation, water retention and porosity in light-textured tropical soils. Soil. Tillage Res. 155, 35–44. doi:10.1016/j.still.2015.08.002
Pan, J. X., Shi, J. W., Tian, D. S., Zhang, R. Y., Li, Y., He, Y. L., et al. (2023). Depth-dependent drivers of soil aggregate carbon across Tibetan alpine grasslands. Sci. Total Environ. 867, 161428. doi:10.1016/j.scitotenv.2023.161428
Parwada, C., and Van Tol, J. (2019). Effects of litter quality on macroaggregates reformation and soil stability in different soil Horizons. Environ. Dev. Sustain. 21 (3), 1321–1339. doi:10.1007/s10668-018-0089-z
Peng, J., Yang, Q. S., Zhang, C. Y., Ni, S. M., Wang, J. G., and Cai, C. F. (2023). Aggregate pore structure, stability characteristics, and biochemical properties induced by different cultivation durations in the Mollisol region of northeast China. Soil. Tillage Res. 233, 105797. doi:10.1016/j.still.2023.105797
Pérès, G., Cluzeau, D., Menasseri, S., Soussana, J., Bessler, H., Engels, C., et al. (2013). Mechanisms linking plant community properties to soil aggregate stability in an experimental grassland plant diversity gradient. Plant Soil 373 (1-2), 285–299. doi:10.1007/s11104-013-1791-0
Puget, P., Chenu, C., and Balesdent, J. (2000). Dynamics of soil organic matter associated with particle-size fractions of water-stable aggregates. Eur. J. Soil Sci. 51 (4), 595–605. doi:10.1046/j.1365-2389.2000.00353.x
Reinhart, K. O., Worogo, H. S. S., Rinella, M. J., and Vermeire, L. T. (2021). Livestock increase soil organic carbon in the Northern great Plains. Rangel. Ecol. Manag. 79 (1), 22–27. doi:10.1016/j.rama.2021.07.006
Reszkowska, A., Krümmelbein, J., Peth, S., Horn, R., Zhao, Y., and Gan, L. (2011). Influence of grazing on hydraulic and mechanical properties of semiarid steppe soils under different vegetation type in Inner Mongolia, China. Plant Soil 340 (1-2), 59–72. doi:10.1007/s11104-010-0405-3
Singh, S., and Singh, J. S. (1996). Water-stable aggregates and associated organic matter in forest, savanna, and cropland soils of a seasonally dry tropical region, India. Biol. Fertil. Soils 22 (1-2), 76–82. doi:10.1007/s003740050079
Six, J., Elliott, E. T., and Paustian, K. (2000). Soil macroaggregate turnover and microaggregate formation: a mechanism for C sequestration under no-tillage agriculture. Soil Biol. Biochem. 32 (14), 2099–2103. doi:10.1016/s0038-0717(00)00179-6
Sodhi, G. P. S., Beri, V., and Benbi, D. K. (2009). Soil aggregation and distribution of carbon and nitrogen in different fractions under long-term application of compost in rice–wheat system. Soil. Tillage Res. 103 (2), 412–418. doi:10.1016/j.still.2008.12.005
Thomas, A. D. (2012). Impact of grazing intensity on seasonal variations in soil organic carbon and soil CO2 efflux in two semiarid grasslands in southern Botswana. Philosophical Trans. R. Soc. B-Biological Sci. 367 (1606), 3076–3086. doi:10.1098/rstb.2012.0102
Tisdall, J. M., and Oades, J. M. (1982). Organic matter and water-stable aggregates in soils. J. Soil Sci. 33, 141–163. doi:10.1111/j.1365-2389.1982.tb01755.x
Wan, N. F., Zheng, X. R., Fu, L. W., Kiær, L. P., Zhang, Z. J., Chaplin-Kramer, R., et al. (2020). Global synthesis of effects of plant species diversity on trophic groups and interactions. Nat. Plants 6 (5), 503–510. doi:10.1038/s41477-020-0654-y
Wang, K., Jiao, Z., Tian, X., Fan, H., Zhang, F., and Li, Y. (2019a). The influence of land use and management measures on the quality of grassland soil. Appl. Ecol. Environ. Res. 17 (4), 7511–7522. doi:10.15666/aeer/1704_75117522
Wang, L., Delgado-Baquerizo, M., Wang, D. L., Isbell, F., Liu, J., Feng, C., et al. (2019b). Diversifying livestock promotes multidiversity and multifunctionality in managed grasslands. Proc. Natl. Acad. Sci. U. S. A. 116 (13), 6187–6192. doi:10.1073/pnas.1807354116
Wang, J. W., Zhao, C. Z., Zhao, L. C., Wen, J., and Li, Q. (2020). Effects of grazing on the allocation of mass of soil aggregates and aggregate-associated organic carbon in an alpine meadow. Plos One 15 (6), e0234477. doi:10.1371/journal.pone.0234477
Wang, Z., Tang, K., Struik, P. C., Ashraf, M. N., Zhang, T. R., Zhao, Y. N., et al. (2023). Alteration of microbial carbon and nitrogen metabolism within the soil metagenome with grazing intensity at semiarid steppe. J. Environ. Manag. 347, 119078. doi:10.1016/j.jenvman.2023.119078
Wang, Z. J. Y., Chen, M., Song, Z. B., Zhang, X. X., Qiao, J. J., Li, J. Y., et al. (2025). Effects of different grazing intensities on the plant reproductive traits and heterogeneous distributions of soil properties. Catena 250, 108785. doi:10.1016/j.catena.2025.108785
Wen, D., He, N. P., and Zhang, J. J. (2016). Dynamics of soil organic carbon and aggregate stability with grazing exclusion in the inner Mongolian grasslands. Plos One 11 (1), e0146757. doi:10.1371/journal.pone.0146757
Wiesmeier, M., Steffens, M., Mueller, C. W., Kölbl, A., Reszkowska, A., Peth, S., et al. (2012). Aggregate stability and physical protection of soil organic carbon in semi-arid steppe soils. Eur. J. Soil Sci. 63 (1), 22–31. doi:10.1111/j.1365-2389.2011.01418.x
Wu, Y. T., Guo, Z. W., Li, Z. Y., Liang, M. W., Tang, Y. K., Zhang, J. H., et al. (2022). The main driver of soil organic carbon differs greatly between topsoil and subsoil in a grazing steppe. Ecol. Evol. 12 (8), e9182. doi:10.1002/ece3.9182
Xiao, L., Huang, Y. M., Zhao, J. F., Zhou, J. Y., and Abbas, F. (2021). Effects of planting structure on soil water-stable aggregates, microbial biomass and enzyme activity in a catchment of loess Plateau terraces, China. Appl. Soil Ecol. 159, 103819. doi:10.1016/j.apsoil.2020.103819
Yang, X. M., and Wander, M. M. (1998). Temporal changes in dry aggregate size and stability: tillage and crop effects on a silty loam Mollisol in Illinois. Soil. Tillage Res. 49 (3), 173–183. doi:10.1016/s0167-1987(98)00170-6
Zhang, Q. Y., Shao, M. A., Jia, X. X., and Zhang, C. C. (2018). Understory vegetation and drought effects on soil aggregate stability and aggregate-associated carbon on the loess Plateau in China. Soil Sci. Soc. Am. J. 82 (1), 106–114. doi:10.2136/sssaj2017.05.0145
Zhang, N. N., Sun, G., Zhong, B., Wang, E. T., Zhao, C. Z., Wang, Y. J., et al. (2019). Impacts of wise grazing on physicochemical and biological features of soil in a sandy grassland on the Tibetan Plateau. Land Degrad. Dev. 30 (7), 719–729. doi:10.1002/ldr.3252
Zhang, X. R., Zhang, W. Q., Sai, X., Chun, F., Li, X. J., Lu, X. X., et al. (2022). Grazing altered soil aggregates, nutrients and enzyme activities in a Stipa Kirschnii steppe of Inner Mongolia. Soil and Tillage Res. 219, 105327. doi:10.1016/j.still.2022.105327
Zhao, F. Z., Ren, C. J., Shelton, S., Wang, Z. T., Pang, G. W., Chen, J., et al. (2017). Grazing intensity influence soil microbial communities and their implications for soil respiration. Agric. Ecosyst. Environ. 249, 50–56. doi:10.1016/j.agee.2017.08.007
Zhao, Y. K., Wang, H., Chen, X. W., and Fu, Y. (2022). Effect of rainfall on soil aggregate breakdown and transportation on cultivated land in the Black soil region of northeast China. Sustainability 14 (17), 11028. doi:10.3390/su141711028
Zhou, Z. C., Gan, Z. T., Shangguan, Z. P., and Dong, Z. B. (2010). Effects of grazing on soil physical properties and soil erodibility in semiarid grassland of the Northern loess Plateau (China). Catena 82 (2), 87–91. doi:10.1016/j.catena.2010.05.005
Zhou, G. Y., Zhou, X. H., He, Y. H., Shao, J. J., Hu, Z. H., Liu, R. Q., et al. (2017). Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis. Glob. Change Biol. 23 (3), 1167–1179. doi:10.1111/gcb.13431
Keywords: soil aggregate stability, grazing intensity, soil depth, dry-sieving, wet-sieving
Citation: Li H, Yang Z, Guo X, Li H, Li X, Wu Y, Han Y, Li Z, Zhang J, Miao B and Liang C (2025) Responses of soil aggregate characteristics to grazing intensity differ between topsoil and subsoil in a typical steppe. Front. Environ. Sci. 13:1610919. doi: 10.3389/fenvs.2025.1610919
Received: 14 April 2025; Accepted: 05 August 2025;
Published: 03 September 2025.
Edited by:
Ilan Stavi, Dead Sea and Arava Science Center, IsraelCopyright © 2025 Li, Yang, Guo, Li, Li, Wu, Han, Li, Zhang, Miao and Liang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Zhiyong Li, enlsZWUwMDdAaW11LmVkdS5jbg==
†These authors have contributed equally to this work and share first authorship.