 Hao Wu
Hao Wu Xiaodong Deng2
Xiaodong Deng2 Shaobo Gao
Shaobo Gao- 1Forestry College, Inner Mongolia Agricultural University, Hohhot, Inner Mongolia, China
- 2Inner Mongolia Ecological and Agricultural Meteorological Center, Hohhot, Inner Mongolia, China
- 3Grassland Research Institute of Chinese Academy of Agricultural Science, Hohhot, Inner Mongolia, China
- 4Hohhot Meteorological Bureau, Hohhot, Inner Mongolia, China
Investigating the impact of grazing on soil respiration in artificial Caragana korshinskii is crucial for harmonizing the carbon sequestration potential of ecological restoration with the sustainable development of animal husbandry. This research is directly linked to addressing the challenges of carbon balance regulation in arid regions and mitigating the conflict between forestry and livestock farming. Continuous grazing modifies biomass and soil properties in artificial C. korshinskii shrublands, potentially influencing soil respiration. To assess the effects of grazing on soil respiration in artificial C. korshinskii shrublands, we established three experimental plots (no grazing, light grazing, and heavy grazing) in the northern foothills of the Daqing Mountains in Inner Mongolia in 2022. Soil respiration, physicochemical properties, and biomass were monitored throughout the growing season. The study results indicate that: (1) No significant difference in soil respiration was observed in C. korshinskii shrublands under grazing conditions. (2) The chemical properties of surface soil may serve as the primary regulatory factors influencing soil respiration under grazing conditions. Based on the average level of soil respiration under different grazing intensities, to effectively mitigate soil carbon emissions, we recommend a moderate reduction in the grazing intensity of C. korshinskii shrublands in arid and semi-arid agro-pastoral regions.
1 Introduction
Soil respiration (SR) is the main process that exports carbon dioxide (CO2) from the soil carbon pool to the atmosphere, and this process is an important link in the carbon exchange between terrestrial ecosystems and the atmosphere (Xu and Shang, 2016; Kim et al., 2024). As the largest carbon pool in terrestrial ecosystems, the soil carbon pool is approximately twice the size of the atmospheric carbon pool and three times that of the plant carbon pool (Pang and Xu, 2024). Its CO2 release is the second-largest flux in the global carbon cycle, playing a critical role in ecosystem carbon cycling (Li et al., 2017). Small changes in SR may disrupt the balance of soil carbon pools in terrestrial ecosystems and may even alter the functions of ecosystem carbon sources and sinks, with potential impacts on global climate change (Li et al., 2019; Zheng et al., 2021; Shen H. et al., 2023; Song et al., 2021).
SR is well known to vary in response to changes in vegetation, soil temperature (ST), soil moisture (SM), soil nutrient levels, air temperature, and other environmental factors (Raich et al., 2022; Wang et al., 2020). In arid regions, alterations in soil physical and chemical properties are the predominant factors influencing SR (Raich et al., 2022; Wang et al., 2020). SR is also related to the type of vegetation, which not only directly affects SR but also indirectly affects SR by influencing ST and SM (Post and Kwon, 2000). Different land use practices have varying degrees of influence on SR, with grazing being a traditional and significant land use practice that has garnered substantial research attention. Grazing herbivores can alter vegetation and soil characteristics, as well as nutrient cycling and flow, through selective foraging, trampling, resting, and excreta return. These changes can ultimately influence ecosystem SR and the stability and health of ecosystems (Ju et al., 2024; Zhao et al., 2017; Mekuria et al., 2007; Kölbl et al., 2011; Kelsey et al., 2023). Numerous authors have studied the effects of grazing behavior on SR in different ecosystems (Peri et al., 2015; Gourlez de la Motte et al., 2018; Ondier et al., 2020), but globally, the effects of grazing on SR remain unclear because of different and sometimes contradictory results observed (Raiesi and Asadi, 2006; Chen et al., 2016; Naidu et al., 2022; Li et al., 2024). Previous research has demonstrated that grazing exerts variable and, at times, contrasting effects on soil respiration across diverse ecosystems and plant communities (Li et al., 2024). Specifically, grazing has been found to significantly suppress SR in two typical steppe species, Stipa grandis and Leymus chinensis (Chen et al., 2019). In Inner Mongolia’s typical grasslands, rotational grazing resulted in the highest SR rates, followed by grazing prohibition, whereas continuous grazing led to the lowest SR rates due to reductions in TN and TP (Nie et al., 2019). In the alpine meadows of the Tibetan Plateau, grazing was shown to suppress SR, with soil nitrogen content having a significant influence on SR levels (Du et al., 2022). Conversely, some studies have reported that grazing in the meadow steppes of the Tibetan Plateau accelerated SR (Chen et al., 2016). Research by Sharkhuu et al. (2016) further indicates that light grazing may actually enhance SR. These findings collectively underscore the context-dependent relationship between grazing and SR, which can vary substantially depending on ecosystem type, soil characteristics, and grazing intensity.
Since the 1990s, Caragana korshinskii (cited as C. korshinskii) has been cultivated on abandoned land at the northern foothills of Daqing Mountain in Inner Mongolia. C. korshinskii is a perennial shrub of the genus Caragana, family Leguminosae, that is light-loving, cold-resistant, heat-resistant, and highly adaptable. It thrives in environments with soil pH ranging from 6.5 to 10.5 and average annual precipitation of 100–800 mm. This plant species is crucial in central Inner Mongolia, serving as an excellent forage resource favored by sheep and demonstrating strong branch regeneration capabilities Following grazing, it can produce a substantial number of new branches, forming dense shrubs. This plant acts as a life-saving resource for livestock in grass-deficient areas and harsh conditions, such as winter. It also plays a crucial role in enhancing soil structure, increasing soil fertility, and maintaining ecological balance (De et al., 2024).
Investigating the impact of grazing on SR in artificial C. korshinskii is crucial for harmonizing the carbon sequestration potential of ecological restoration with the sustainable development of animal husbandry. This research is directly linked to addressing the challenges of carbon balance regulation in arid regions and mitigating the conflict between forestry and livestock farming (Bastani et al., 2023). However, no relevant studies have been identified to date. Therefore, a comprehensive understanding of the SR characteristics of C. korshinskii shrublands under grazing and its relationship to soil physicochemical properties and vegetation changes is essential not only for maintaining and enhancing carbon sequestration capacity but also for optimizing the use of existing C. korshinskii forage resources, thereby maximizing ecological and economic benefits in these regions.
Grazing is recognized for its direct and indirect effects on vegetation, soil properties, and nutrient cycling processes. Therefore, we hypothesized that grazing could influence the soil physicochemical properties and vegetation biomass of the C. korshinskii shrublands, consequently affecting SR. This study focuses on the C. korshinskii shrublands located at the northern foothills of the Daqing Mountains in central Inner Mongolia, aiming to clarify two scientific issues: 1. What are the dynamic characteristics of SR in the C. korshinski shrublands under grazing, and are there significant differences in SR across these intensities? 2. Under grazing conditions, which factors primarily regulate SR? The overarching objective of this research is to provide a robust theoretical foundation for establishing appropriate grazing intensity, optimizing land use practices, and enhancing the sustainable management and development of C. korshinskii shrublands in central Inner Mongolia.
2 Materials and methods
2.1 The study area
The experimental site is located at the junction of Damao Banner and Wuchuan County (N 40°74′-41°23′, E 110°30′-111°52′), at the northern foothills of Daqing Mountain Range in the central part of Inner Mongolia (Figure 1). The area is characterized by an elevated terrain (average elevation: 1700 m) and an arid to semi-arid grassland climate. The region experiences extreme temperature variability, with an annual average of 1.2°C, ranging from −14.89°C in January (coldest month) to 18.80°C in July (hottest month). Precipitation is low (343 mm annually), predominantly occurring from May to September, with high solar radiation (2,960 sunshine hours/year) and evaporation (2,200 mm/year). Frequent strong winds (gusts of 6–7 m/s) are observed between April and June. The frost-free period spans 75–105 days. The soil is dominated by chestnut soils, and the formation of vegetation types dominated by degraded hay meadows and secondary forests (Han et al., 2024).
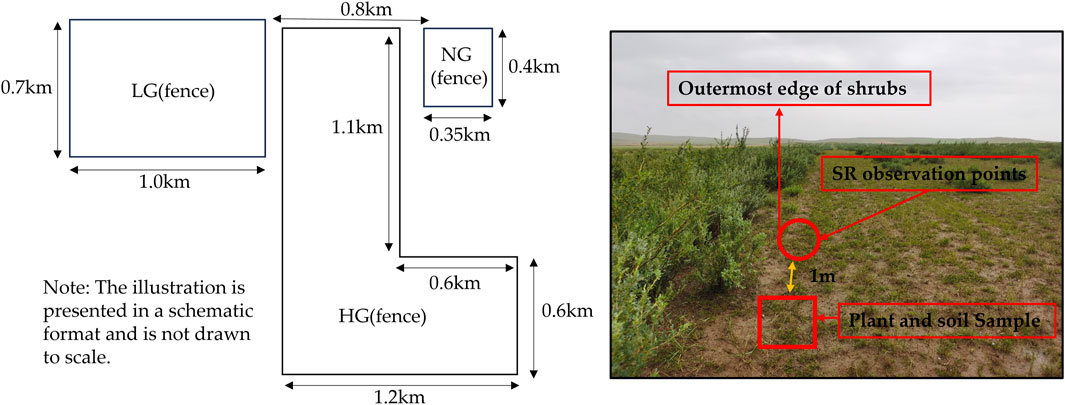
Figure 1. Sampling schematic.
2.2 Experimental design
The experimental plots were established on flat terrain with uniform climatic conditions. The study subjects consisted of artificial C. korshinskii shrublands planted in 2014, arranged in two-row bands. Three grazing treatments were implemented based on regional practices: no grazing (NG), light grazing (LG), and heavy grazing (HG). The NG plots (initiated in 2014) were permanently fenced to exclude grazing. The LG treatment (70 hm2) was stocked with 72 sheep (1.0 sheep/hm2/year), while the HG treatment (140 hm2) contained 350 sheep (2.5 sheep/hm2/year). Both LG and HG grazing throughout the year, grazing during the day, and sheep returning to the pen at night, grazing started in 2016.
SR was measured in 2022. The specific design of setting up SR observation points in the above shrubland experimental plots is as follows: 3 SR observation points were set up in each experimental plot along the outermost edge of the canopy of the shrub with an interval of about 50 m between the observation points within the same plot, total of nine observation points were established. Measurement, 1 month in advance, the circular steel circle (30 cm diameter, 8 cm height) buried at each observation point, vertical pressure into the soil 5 cm, to ensure that the steel circle and the surrounding soil seamless fit. The steel ring will remain stationary until the year-end test. To minimize interference from above-ground plant respiration, we carefully removed herbaceous above-ground portions within the steel rings using scissors and cleared away dead leaves and branches. We also minimized soil disturbance and root damage to reduce their potential impact on the measurement results.
The ACE soil carbon flux measurement system (Manufactured by ADC United Kingdom) was installed, and SM and ST probes were inserted into the soil at a depth of 10 cm to measure SM and ST, at the conclusion of the measurement process, the respiratory chamber is opened to perform a zero-point measurement, and the ambient CO2 concentration is recalibrated to calculate the net carbon exchange rate accurately. The SR of C. korshinskii shrublands under the three treatments was measured from 08:00 to 18:00 on the measurement day. Each observation points was measured for 10 min every 2 h. The test dates for the entire growing season in 2022 were as follows: May 8, May 28, June 7, June 25, July 6, July 23, August 3, August 29, September 10, and September 24. To ensure representativeness, the measurement days were selected to coincide with clear, cloudless, and either windless or breezy weather conditions.
To avoid disturbance to SR, three 1 m × 1 m herbaceous sample plots were set up 1 m away from the SR monitoring steel ring to investigate the basic condition of herbaceous plants, the above-ground plant materials were harvested, placed in paper bags, and transported to the laboratory where they were dried to a constant weight to determine the above-ground biomass (AGB).
After the investigation, an 8 cm diameter electric root drill was used in the sample plots to extract underground roots at a depth of 20 cm (the root systems of herbaceous plants are mainly distributed between 0 and 20 cm), repeated five times. Roots from the same soil layer were carefully collected, washed with deionized water to remove adhering soil, then dried and weighed to determine belowground biomass (BGB).
The soil bulk density sample was collected using a ring knife, transported to the laboratory, and subsequently dried in an oven set at 105°C until it reached a constant weight.
A soil drill with an inner diameter of 5 cm was used to collect soil from the sample plots at a depth of 20 cm for chemical analysis, repeated 3 times. The soil samples were placed in sealed plastic bags, transported to the laboratory, and air-dried. Subsequently, they were sieved through a 2 mm sieve to remove non - soil substances. After that, the samples were ground using a lapping body and then passed through a 0.25 mm sieve. The sieved soil samples were placed in sample bottles to be measured for soil organic carbon (SOC), total nitrogen (TN), alkali-hydrolyzable nitrogen (ASN), total phosphorus (TP), available phosphorus (AP), available potassium (AK), soil acidity and alkalinity (pH), soil C/N ratio (C/N), Soil pH (pH). SOC was measured using the dichromate oxidation with external heating method. TN was determined via the diffusion method. For TP, samples were first digested with concentrated sulfuric acid and perchloric acid, and then analyzed using a continuous flow analyzer. ASN was measured by the alkali - hydrolysis diffusion method. AP was extracted with sodium hydroxide and analyzed by the molybdenum - antimony - potassium antimony tartrate colorimetric method. AK was extracted with ammonium acetate and measured using a flame photometer. The pH was directly measured with a pH meter (model PHS - 2F, produced by Shanghai INESA Scientific Instrument Co., Ltd.). Soil sample collection date: May 8, June 7, July 6, August 3, September 10 (Bao, 2007).
2.3 Statistical analysis
Data were organized using Excel 2021 (Microsoft Corporation, Redmond, WA, United States) Figures and correlation analyses were generated using Origin 2021 (Origin Lab Corporation, Northampton, MA, United States). Two-way and three-way factorial ANOVA was conducted to evaluate the effects of grazing intensity, month, and soil depth, as well as their interaction effects. Post hoc multiple comparisons were performed for each factor across different levels of grazing intensity. Additionally, a structural equation model (SEM) was constructed to explore the relationships among variables. All statistical analyses were performed using SPSSPRO software (www.spsspro.com).
2.4 Basic information of the sample plot
In 2022, the total rainfall from May to September, covering the plant growing season, amounted to 333 mm. The monthly distribution of precipitation was highly uneven, with a significant concentration of rainfall in July and August, while May, June, and September experienced relatively lower precipitation levels. The average temperature for this period was approximately 11.75°C, but daily temperature variations were considerable (Figure 2).
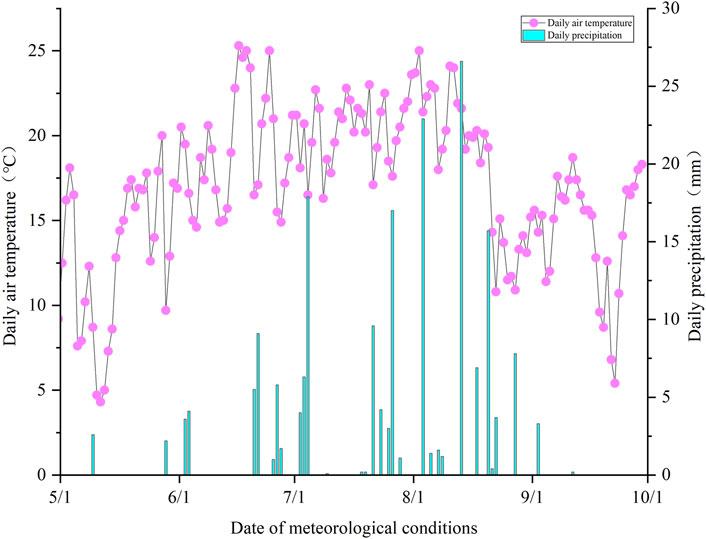
Figure 2. Daily meteorological conditions in 2022.
The mechanical composition of the three plots exhibits a high degree of similarity, with no statistically significant differences. The particle size distribution is primarily concentrated within the range of 2–50 μm, predominantly consisting of silt (Table 1).
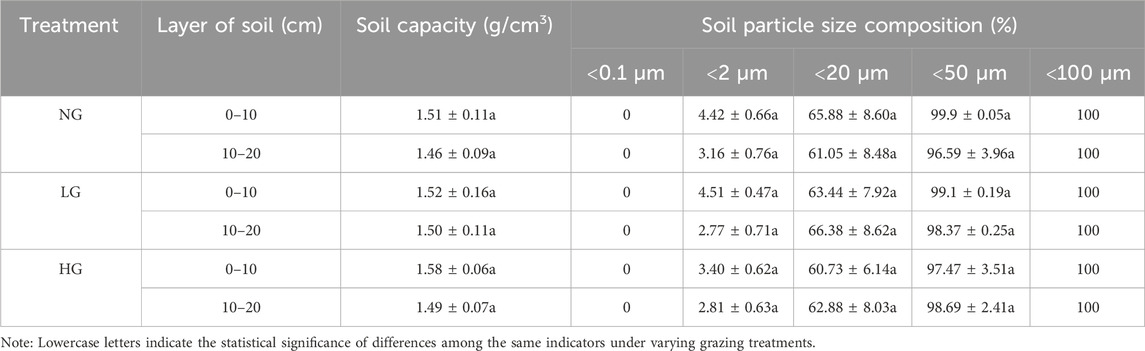
Table 1. Soil bulk weight and soil mechanical composition.
3 Results
3.1 Dynamic characteristics of soil respiration
The temporal dynamics of SR in C. korshinskii shrublands were studied under different grazing treatments from May to September. SR showed a clear seasonal pattern, with an initial increase from May, peaking in July, and then declining until the end of the study in September. SM in C. korshinskii shrublands fluctuated during the study period, with the highest recorded on June 7 and the lowest on June 25. ST beneath C. korshinskii shrublands showed complex variations. From May onwards, ST increased consistently across all grazing treatments, maintaining a range of 30°C–40°C from June through August. A rapid decline in ST was observed by late September (Figure 3).
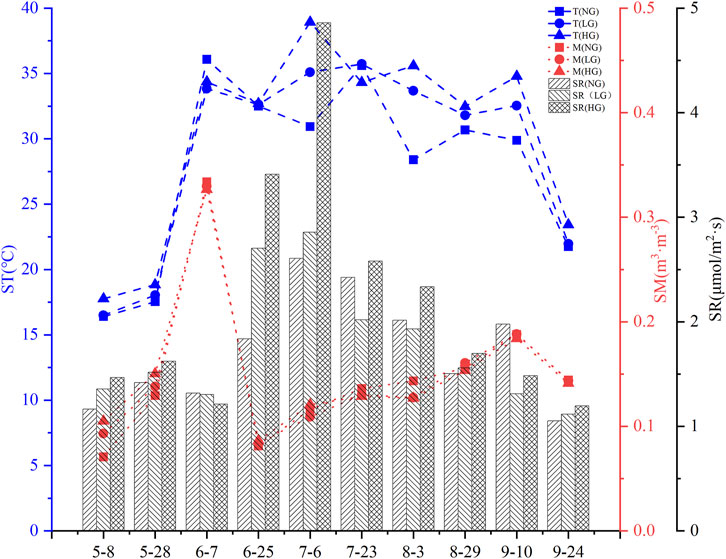
Figure 3. Dynamic characteristics of soil respiration, soil temperature, soil moisture.
3.2 Responses of soil respiration, soil physical and chemical properties and biomass to grazing
G has a main effect on SOC, TN, ASN, C/N, pH, AGB and BGB. D has a main effect on all the physical and chemical properties of the soil. M has a major effect on SR and all soil physical and chemical properties and biomass except TP. G × D has an interaction on SOC, TN, C/N, TP, AP and pH. G × M has an interaction effect on all soil physicochemical properties and biomass. D × M has an interaction effect on all soil physical and chemical properties; G × D × M has an interaction with SOC, TN, TP, AP, AK and pH (Table 2).
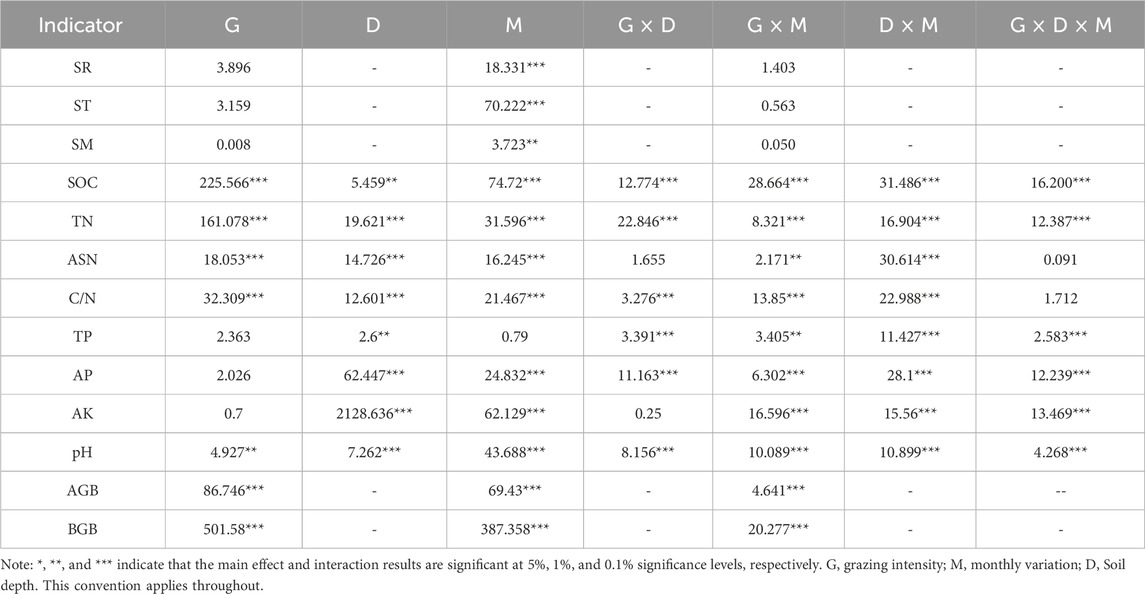
Table 2. Results of variance analysis of grazing intensity, month and soil depth on three/two factors of soil respiration, soil chemical properties and biomass.
In this study, no significant differences in SR, ST, SM, TP, AP, and AK were observed among various grazing treatments. SOC and TN increased significantly with rising grazing intensity, and differences among all grazing treatments were significant. ASN also showed an increasing trend, with a significant difference between NG and HG treatments, while no significant differences were found between other treatments. Grazing enhanced the C/N ratio, and the C/N under NG was significantly different from that under LG and HG. Notably, HG significantly reduced soil pH, with significant differences found between HG and both NG and LG treatments. Furthermore, AGB and BGB decreased significantly with increasing grazing intensity, and significant differences were observed among all treatments (Table 3).
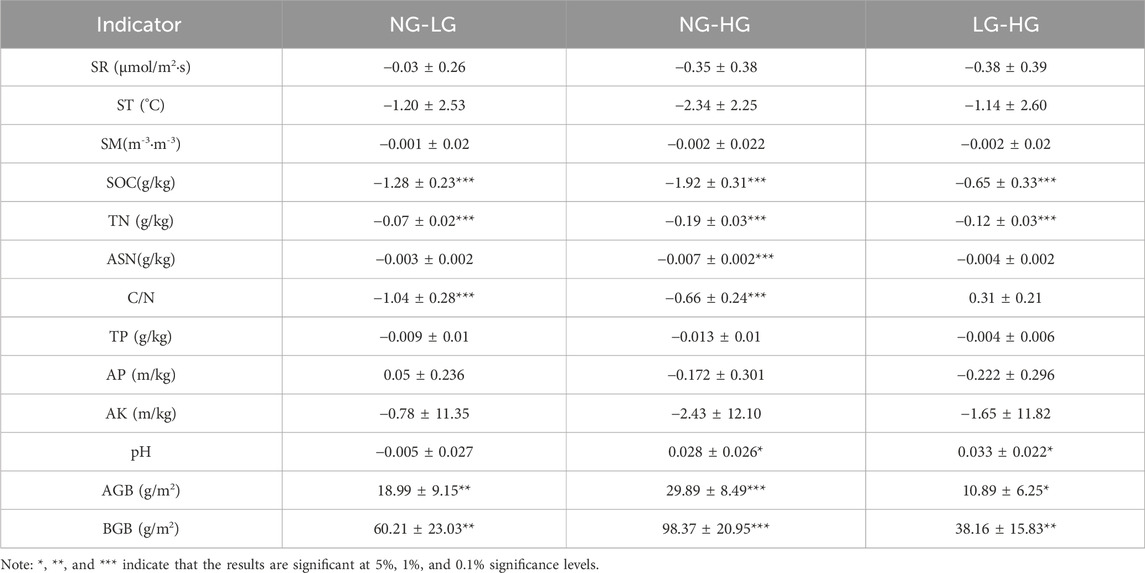
Table 3. Post-event multiple comparisons (grazing intensity).
3.3 Control factors of soil respiration
SR showed significant positive correlations with ST, SOC10, TN10, AHN10, TP10, AP10, AK10, AK20, and C/N10. SR showed non-significant positive correlations with SOC20, TN20, ASN20, C/N20, aboveground biomass and below-ground biomass; and SR showed non-significant negative correlations with SM, TP20, pH10, and pH20 (Figure 4). Grazing influences SR by altering the chemistry of the topsoil, both directly and indirectly. Soil physical properties, deep soil chemistry, and biomass do not have a significant direct impact on SR (Figure 5).
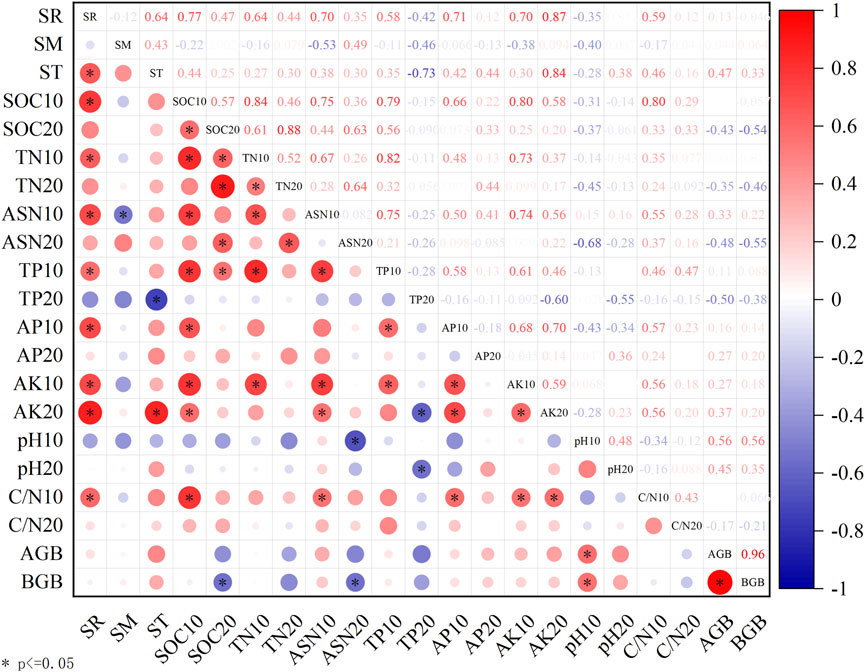
Figure 4. The heat map illustrates the correlation between soil respiration and soil physicochemical properties. Note: SR: soil respiration rate; SM: soil moisture; ST: soil temperature; SOC10: soil organic carbon in 0–10 cm soil layer; SOC20: organic carbon in 10–20 cm soil layer; TN10: soil total nitrogen in 0–10 cm soil layer; TN20: soil total nitrogen in 10–20 cm soil layer; ASN10: soil alkali-hydrolyzed nitrogen in 0–10 cm soil layer; ASN20: soil alkali-hydrolyzed nitrogen in 10–20 cm soil layer; TP10: total phosphorus in 0–10 cm soil layer; TP20: total phosphorus in 10–20 cm soil layer; AP10: soil available phosphorus in 0–10 cm soil layer; AP20: soil available phosphorus in 10–20 cm soil layer; AK10: soil available potassium in 0–10 cm soil layer; AK20: soil available potassium in 10–20 cm soil layer; pH10: pH value in 0–10 cm soil layer; pH: pH value in 10–20 cm soil layer; C/N10: soil layer carbon nitrogen ratio in 0–10 cm soil layer; C/N20: soil layer carbon nitrogen ratio in 10–20 cm soil layer; AGB: aboveground biomass; BGB: below-ground biomass.
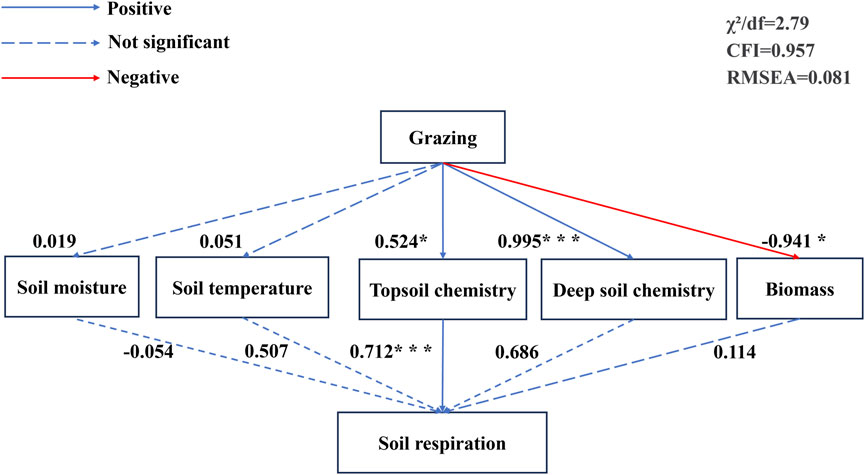
Figure 5. Structural equation modeling. Note: Structural equation modelling of factors on soil respiration, grazing intensity including NG, LG, HG; surface soil chemical properties including SOC10, TN10, ASN10, C/N10, TP10, AP10, AK10, pH10; deep soil properties including SOC20, TN20, ASN20, C/N20, TP20, AP20, AK20, pH20; biomass including aboveground biomass, BGB: below-ground biomass. ***, **, * are significant at 5%, 1%, and 0.1% significance levels.
4 Discussion
4.1 The regulation of abiotic factors on soil respiration under grazing effect
Currently, there is no definitive conclusion regarding the impact of grazing on SM (Odriozola et al., 2014; Gao et al., 2004). The findings of this study, which indicate an insignificant effect of grazing on SM, align with those of Qu et al. (2024). ST is closely associated with SM and plant shading. no significant differences in ST are observed across different treatments. The reduced water holding capacity and limited deep infiltration potential of sandy soils may contribute to a mitigating effect on both SM and ST (Vermeire et al., 2005).
The research findings on the effect of grazing intensity on soil chemical properties have not yet reached a consistent conclusion (Jalilpour et al., 2022). Bolan posited that the chemical attributes of pasture soils are significantly shaped by the deposition of manure and urine and their subsequent transformations within the soil (Bolan et al., 2024). As grazing intensity increases, livestock food intake rises, potentially reducing the decomposition of dead branches, leaves, and ground vegetation, thereby decreasing soil nutrient levels. In contrast, high-intensity grazing can enhance soil nutrient content through increased livestock excretion of manure and urine (Bastani et al., 2023).
Some studies suggest that grazing can enhance SOC, primarily due to the trampling by animals breaking down litter and facilitating its intimate contact with the soil, thereby accelerating decomposition and the rapid return of organic carbon to the soil, or altering the root-to-shoot ratio of plants and increasing the allocation of carbon to below ground biomass (Reeder and Schuman, 2002; Pan et al., 2023; Shen Y. et al., 2023; Wei et al., 2023; Wienhold et al., 2001). Other studies indicate that grazing has either no significant effect or a negligible impact on SOC depending on whether soil erosion occurs following grazing activities (Robertson et al., 2016; Derner et al., 2019; Nüsse et al., 2017; Wang G. et al., 2022; Hancock and Vallely, 2020; Zhang et al., 2023). Furthermore, some research indicates that the relationship between SOC and grazing lacks a consistent pattern, showing both positive and negative correlations at different times (Derner et al., 2019). In this study, grazing was found to increase SOC in Caragana korshinskii shrublands. The current grazing intensity or duration does not cause soil erosion, and animal trampling may facilitate the decomposition of litter.
Soil nitrogen content is influenced by the rates of accumulation and decomposition of soil organic matter. Higher organic matter content generally leads to higher soil nitrogen levels (Calazans et al., 2018; Yao et al., 2023). In this study, both TN and ASN showed significant positive correlations with SOC, which supports the aforementioned findings.
In this study grazing did not have a direct effect on TP, AP, or AK, and the differences among various grazing treatments were not statistically significant. This result aligns with the findings of Navasardyan et al. (2024). The lack of immediate impact may be attributed to the fact that most phosphorus and potassium inputs originate from animal manure, whose decomposition is a prolonged process lasting 5–10 years and is heavily influenced by climatic and soil conditions (Vinograd et al., 2019). The significant interaction between grazing and month or soil depth observed in this study suggests that although grazing does not directly alter TP, AP, or AK levels, it may influence these nutrients indirectly through modifications in environmental conditions or other ecological processes. This also explains the varying results reported by Yao et al. (2019) across different sampling times and soil layers under diverse grazing regimes.
Grazing exerts a direct influence on soil pH; however, the pH value within the C. korshinskii shrublands remains stable between 8.25 and 8.55, preserving the soil’s weakly alkaline condition. This stability can be attributed to the loose soil structure and the rapid evaporation of surface moisture in the experimental area.
In arid regions, soil physical and chemical properties account for 65%–70% of the variability in SR, with soil chemical properties being the predominant factor influencing SR in shrubland ecosystems (Wen et al., 2018). In this study, SR exhibited significant positive correlations with ST, SOC10, TN10, ASN10, TP10, AP10, AK10, and AK20. The chemical properties of surface soil may be the primary determinants of SR.
SM is the primary determinant of dissolved organic matter availability and mobility in soil, serving as a respiratory substrate and energy source for soil microorganisms (Chowdhury et al., 2011; Lloyd et al., 2016). Additionally, SM significantly influences plant root growth and survival, particularly in water-scarce regions (Lloyd et al., 2016). Numerous studies have demonstrated a positive correlation between SM and SR during the growing season, with the relationship potentially being linear, logarithmic, quadratic, or parabolic (Ondier et al., 2020; Luo et al., 2012; Zhang et al., 2017; Xu and Wan, 2008; Hanpattanakit et al., 2015; Luo et al., 2012; Rubio and Detto, 2017). Some research indicates that SM affects SR within an optimal range; excessive or insufficient SM can either inhibit or have no significant effect on SR (Knowles et al., 2015). However, this study found no significant relationship between SM and SR. The study area, characterized by arid conditions, low precipitation, loose soil texture, and poor water retention, suggests that water availability becomes a non-limiting factor. This finding aligns with previous research conducted in similar arid environments (Wang et al., 2023; Lin et al., 2020).
ST constitutes a critical determinant of SR, particularly when SM is not a limiting factor, thereby becoming the predominant environmental influence on SR. Extensive research has demonstrated that SR exhibits significant or highly significant positive correlations with both air and ST, which can be mathematically represented by linear or exponential functions. The findings of this study align with those of previous researchers, indicating a positive correlation between SR and ST (Guo et al., 2018).
SOC serves as the material foundation for CO2 emissions resulting from microbial decomposition activities and constitutes the largest carbon pool within the ecosystem, significantly influencing SR (Wang et al., 2017). Previous studies have demonstrated a positive correlation between SOC and SR, findings that are consistent with our study.
Soil nitrogen serves as the primary source of plant nitrogen nutrition. The majority of soil nitrogen exists in organic forms, while only a minor portion is inorganic. Organic nitrogen undergoes mineralization to transform into ammonium, which can then be absorbed and utilized by crops, leading to the release of CO2 (Wang et al., 2017). Several studies have demonstrated that higher TN significantly increases the CO2 emission flux in grassland communities (Graham et al., 2014), and Zhou also observed a significant positive correlation between SR and TN (Zhou et al., 2021). Some studies have also shown that excessive nitrogen inhibits SR, possibly because the affinity between nitrogen and carbon reduces the availability of carbon, thus hindering the metabolic activities of microorganisms and slowing down CO2 emission (Zhou et al., 2009). In this study, SR showed a significant positive correlation with TN in 0–10 cm soil layer, based on the research findings of other authors, it is hypothesized that the SR of C. korshinskii shrublands exhibits a strong positive correlation with the denitrification rate of surface soil (Van Kessel et al., 1993; Christensen et al., 1990; Zhao et al., 2015). ASN is a major component affecting plant roots and soil microbial metabolic activities, and generally has a significant positive correlation with SR, and the results of this study are consistent with this inference (Zhao et al., 2015).
C/N serves as an indicator of the mineralization efficiency of soil organic matter during decomposition. Studies have demonstrated that C/N is inversely related to the rate of organic matter decomposition. A lower C/N facilitates soil mineralization, leading to faster decomposition of soil organic carbon and increased CO2 release from the soil, whereas a higher C/N can limit this process (Sinsabaugh et al., 2002; Mande et al., 2014; Xu et al., 2016; Tian et al., 2019). Other researchers have observed that low C/N can restrict microbial activity and decomposition due to the presence of recalcitrant carbon sources, suggesting that low C/N indicates more decomposed SOC by microorganisms (Tian et al., 2019; Ngao et al., 2012; Webster et al., 2018; Klimek et al., 2021). During organic matter mineralization, organotrophic microorganisms require carbon sources for cellular components and nitrogen for nutrient supply to maintain C/N balance within their cell composition. The C/N conditions thus constrain microbial growth and activity, influencing the decomposition rate of organic matter, which in turn affects SR. This suggests that microbial decomposition activity requires an optimal C/N (Zhao et al., 2015; Sinsabaugh et al., 2002). Our study reveals a positive correlation between SR and C/N in the 0–10 cm soil layer, differing from some studies but aligning with others (Tian et al., 2019; Klimek et al., 2021). Our findings may indicate enhanced carbon consumption with increased C/N under non-nitrogen-limited conditions (Tian et al., 2019; Ngao et al., 2012; Darenova et al., 2024).
The observed low soil C/N ratios are likely driven by the nitrogen-fixing capacity of C. korshinskii. Although biological nitrogen fixation (BNF) was not directly measured in this study, C. korshinskii, as a nitrogen-fixing plant, enriches the soil nitrogen pool. This increase in nitrogen availability provides a mechanistic explanation for the lower C/N ratios observed in its presence. This finding aligns with the established hypothesis that nitrogen-fixing species can significantly alter soil C/N dynamics by enhancing nitrogen inputs, thereby potentially influencing soil organic matter decomposition and stabilization rates (SR). Future studies directly comparing soils with and without C. korshinskii would further elucidate its specific impact on soil nitrogen cycling and SR.
Plant growth requires phosphorus (P) and potassium (K) elements. When P and K in soil become limiting factors, even minor changes in their concentrations can significantly affect SR (Li et al., 2021). P can enhance the amount of easily decomposable organic matter, thereby increasing the substrate available for SR. Additionally, it can boost root biomass, thus promoting microbial decomposition activities and root respiration. Studies have demonstrated a significant or highly significant positive correlation between TP and SR, with increased TP stimulating higher soil CO2 emissions. However, some studies indicate that excessive TP may inhibit SR (Wang et al., 2017). In this study, SR in C. korshinskii shrublands was positively correlated with TP10 and AP10, suggesting that topsoil P content within an optimal range influenced SR. AK refers to potassium ions in soil that plants can directly absorb and utilize. It is widely accepted that its concentration directly impacts crop growth, development, and root respiration, which constitutes a crucial component of overall SR. Therefore, the supply of AK directly affects root respiration and consequently the overall intensity of SR. In this study, AK10 and AK20 showed a positive correlation with SR, aligning with the aforementioned hypothesis.
In this study, the SR values across different grazing treatments did not exhibit statistically significant differences. The results align with findings from investigations conducted in the Stipa krylovii desert steppe and sandy grasslands of Inner Mongolia, where grazing did not significantly affect SR (Wang et al., 2023; Lin et al., 2020). The authors of those studies proposed that this pattern might be due to the arid and semi-arid conditions of the regions, which are characterized by consistently low and stable SM levels. Furthermore, the absence of a significant relationship between SM and SR can be attributed to the asynchrony between fluctuations in SM and variations in ST, suggesting that SM does not act as a limiting factor for SR under these environmental conditions. Our findings further support the conclusions drawn by these previous studies.
Our study demonstrates that grazing exerts both direct and indirect effects on soil chemical properties, and surface soil chemistry shows a significant correlation with SR. This supports Li’s findings that in arid and semi-arid shrubland ecosystems, variations in soil chemical properties are the dominant drivers of SR (Wen et al., 2018). However, no significant differences were observed in annual average SR across different grazing treatments. This outcome may be explained by several factors: First, the alterations in soil chemical composition under varying grazing intensities may not have reached a threshold sufficient to elicit measurable changes in SR. Although statistically significant, these changes might not have been physiologically or biochemically meaningful enough to influence in SR. Secondly, the influence of soil chemical properties on soil respiration may exhibit a lagging effect, which necessitates continued monitoring in future studies. Last, other factors such as microbial community structure and activity could have buffered the effects of soil chemical changes on SR. Microbial adaptation to grazing-induced shifts in soil nutrients may have mitigated potential impacts on respiration rates, an area that will be explored in future research.
4.2 The regulation of biotic factors on soil respiration under grazing effect
Most studies have demonstrated that grazing reduces the AGB of plants, primarily through nibbling and trampling by animals. These activities inhibit plant photosynthesis and significantly hinder their recovery. Other studies suggest that grazing selectively decreases palatable plant species while creating favorable conditions for less palatable weeds. While the AGB of weeds increased, this increment was insufficient to offset the reduction in total AGB. (Chen et al., 2024; Seabloom et al., 2020). A few studies support the “moderate disturbance hypothesis,” indicating that moderate grazing intensity initially increases AGB before decreasing it. Moderate grazing can promote the growth of forage grasses, enhance AGB, and increase species diversity, thereby boosting overall biomass. Studies also indicate that grazing reduces BGB by compacting soil through animal trampling, creating an anaerobic environment that inhibits root growth (Wang J. et al., 2022). Some authors argue that moderate grazing promotes BGB as plants allocate more resources below ground to recover from reduced AGB, thus obtaining more nutrients and water (Frank et al., 2002). In this study, both the AGB and BGB of herbal in LG and NG were significantly reduced, consistent with previous findings (Chen et al., 2024; Wang G. et al., 2022), highlighting the significant impact of grazing on the vegetation structure of C. korshinskii shrublands.
It is widely accepted that SR is closely linked to biomass. An increase in AGB enhances photosynthetic capacity, leading to greater carbon fixation. A portion of this fixed carbon is transported to the roots as photosynthetic products, providing additional carbon sources for root respiration and soil microorganisms, thereby stimulating SR. Roots serve as the primary interface between plants and soil, and their respiration constitutes a significant component of overall SR. Typically, higher root biomass results in more vigorous root respiration, which directly contributes to an increased SR (Carrillo et al., 2011). However, existing research presents divergent conclusions. Hu posited that the relationship between SR and biomass varies depending on vegetation type (Hu et al., 2024). Previous studies have demonstrated that in arid regions, soil heterotrophic respiration (Rh), which microorganisms decompose SOC to release CO2, constitutes the primary contribution to SR, as opposed to soil autotrophic respiration (Ra), which involves CO2 release from root activities (Balogh et al., 2016). In arid and water-scarce regions, SM is the predominant factor influencing SR, as opposed to biomass, plant roots require water to maintain active metabolism and respiration. Drought stress induces stomatal closure in plants, thereby reducing root respiration. Secondly, soil microorganisms are more sensitive to moisture fluctuations and can exhibit rapid bursts of respiratory activity. Finally, microorganisms are spatially closer to carbon sources. Most organic matter is concentrated in the upper layers of the soil, where it can be directly utilized by microorganisms. In contrast, roots must transport photosynthetic products over long distances before they can be used. (Han et al., 2019). Our experiment was also conducted in an arid region, where no significant correlation was observed between biomass and SR, a finding consistent with previous studies carried out in similar environments. A previous global meta-analysis of the relationship between grazing and soil respiration (SR), which included 69 studies, revealed that grazing significantly inhibits soil respiration, encompassing both autotrophic and heterotrophic respiration. The negative impact on soil respiration becomes more pronounced with increased grazing intensity, prolonged grazing duration (>10 years), and higher temperatures (>5°C). Grazing-induced changes in belowground biomass primarily drive variations in soil respiration. The discrepancy between our findings and those of the meta-analysis may stem from several factors: the grazing duration at our experimental site was less than 10 years, the growing season temperature at our site was relatively low, and the meta-analysis did not differentiate study regions based on water availability (Li et al., 2024).
5 Conclusion
Our study revealed that: (1) No significant difference in SR was observed in C. korshinskii shrublands under grazing conditions. (2) The chemical properties of surface soil may serve as the primary regulatory factors influencing SR under grazing conditions. Based on the average level of SR under different grazing intensities, to effectively mitigate soil carbon emissions, we recommend a moderate reduction in the grazing intensity of C. korshinskii shrublands in arid and semi-arid agro-pastoral regions.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
HW: Investigation, Visualization, Writing – original draft, Writing – review and editing. XD: Formal Analysis, Resources, Supervision, Writing – review and editing. XL: Formal Analysis, Resources, Supervision, Writing – review and editing. SG: Formal Analysis, Validation, Writing – review and editing. RY: Data curation, Formal Analysis, Visualization, Writing – review and editing. JL: Data curation, Investigation, Visualization, Writing – review and editing. HM: Writing – review and editing, Data curation, Investigation, Visualization. XM: Conceptualization, Funding acquisition, Methodology, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by Key Research and Development and Achievement Transformation Program Projects of Inner Mongolia Autonomous Region (2025YFHH0219), Inner Mongolia Natural Science Foundation (2025LHMS03032), Science and the Technology Innovation Project of the Inner Mongolia Meteorological Bureau (nmqxkjcx202583).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Balogh, J., Papp, M., Pinter, K., Fóti, S., Posta, K., Eugster, W., et al. (2016). Autotrophic component of soil respiration is repressed by drought more than the heterotrophic one in dry grasslands. Biogeosciences 13, 5171–5182. doi:10.5194/bg-13-5171-2016
Bao, S. D. (2007). Soil and agricultural chemistry analysis. 3rd ed. Beijing, China: China Agriculture Press.
Bastani, M., Sadeghipour, A., Kamali, N., Zarafshar, M., and Bazot, S. (2023). How does livestock graze management affect woodland soil health? Front. Vet. Sci. 6, 1028149. doi:10.3389/ffgc.2023.1028149
Bolan, N. S., Saggar, S., Luo, J., Bhandral, R., and Singh, J. (2024). Gaseous emissions of nitrogen from grazed pastures: processes, measurements and modelling, environmental implications, and mitigation. Adv. Agron. 84, 37–120. doi:10.1016/S0065-2113(04)84002-1
Calazans, S. O. L., Morais, V. A., Scolforo, J. R., Zinn, Y. L., Mello, J. M., Mancini, L. T., et al. (2018). Soil organic carbon as a key predictor of N in forest soils of Brazil. J. Soils Sediments 18, 1242–1251. doi:10.1007/s11368-016-1557-4
Carrillo, Y., Pendall, E., Dijkstra, F. A., Morgan, J. A., and Newcomb, J. M. (2011). Response of soil organic matter pools to elevated CO2 and warming in A semi-arid grassland. Plant Soil 347, 339–350. doi:10.1007/s11104-011-0853-4
Chen, J., Zhou, X., Wang, J., Hruska, T., Shi, W., Cao, J., et al. (2016). Grazing exclusion reduced soil respiration but increased its temperature sensitivity in a meadow grassland on the Tibetan Plateau. Ecol. Evol. 6, 675–687. doi:10.1002/ece3.1867
Chen, W., Ye, M., Pan, X., Li, M., Zeng, G., Zhang, X., et al. (2024). Relationships and changes in grassland community diversity and biomass in the pastoral areas of the two Rivers under grazing disturbance. Agronomy 14, 1336. doi:10.3390/agronomy14061336
Chen, W., Zheng, X., Wolf, B., Yao, Z., Liu, C., Butterbach-Bahl, K., et al. (2019). Long-term grazing effects on soil-atmosphere exchanges of CO2, CH4 and N2O at different grasslands in Inner Mongolia: a soil core study. Ecol. Indic. 105, 316–328. doi:10.1016/j.ecolind.2017.09.035
Chowdhury, N., Marschner, P., and Burns, R. (2011). Response of microbial activity and community structure to decreasing soil osmotic and matric potential. Plant Soil 344, 241–254. doi:10.1007/s11104-011-0743-9
Christensen, S., Simkins, S., and Tiedje, J. M. (1990). Temporal patterns of soil denitrification: their stability and causes. Soil Sci. Soc. Am. J. 54, 1614–1618. doi:10.2136/sssaj1990.03615995005400060017x
Darenova, E., Adamič, P. C., and Čater, M. (2024). Effect of temperature, water availability, and soil properties on soil CO2 efflux in beech-fir forests along the carpathian mts. Catena 240, 107974. doi:10.1016/j.catena.2024.107974
De, X., Li, H., Kang, J., and Yang, J. (2024). Experimental study on the cutting and crushing performance of Caragana korshinskii strips. Agriculture 14, 1584. doi:10.3390/agriculture14091584
Derner, J. D., Augustine, D. J., and Frank, D. A. (2019). Does grazing matter for soil organic carbon sequestration in the Western north American great plains? Ecosystems 22, 1088–1094. doi:10.1007/s10021-018-0324-3
Du, C., Zhou, G., and Gao, Y. (2022). Grazing exclusion alters carbon flux of alpine meadow in the Tibetan Plateau. Agric. For. Meteorol. 314, 108774. doi:10.1016/j.agrformet.2021.108774
Frank, D. A., Kuns, M. M., and Guido, D. R. (2002). Consumer control of grassland plant production. Ecology 83, 602–606. doi:10.1890/0012-9658(2002)083[0602:CCOGPP]2.0.CO,2
Gao, Y. Z., Han, X. G., and Wang, S. P. (2004). The effects of grazing on grassland soils. Acta Ecol. Sin. 24, 790–797. doi:10.3321/j.issn:1000-0933.2004.04.021
Gourlez de la Motte, L., Mamadou, O., Beckers, Y., Bodson, B., Heinesch, B., and Aubinet, M. (2018). Rotational and continuous grazing does not affect the total net ecosystem exchange of a pasture grazed by cattle but modifies CO2 exchange dynamics. Agric. Ecosyst. Environ. 253, 157–165. doi:10.1016/j.agee.2017.11.011
Graham, S. L., Hunt, J. E., Millard, P., McSeveny, T., Tylianakis, J. M., and Whitehead, D. (2014). Effects of soil warming and nitrogen addition on soil respiration in a New Zealand tussock grassland. PLOS ONE 9, e91204. doi:10.1371/journal.pone.0091204
Guo, N., Wang, A., Allan Degen, A., Deng, B., Shang, Z., Ding, L., et al. (2018). Grazing exclusion increases soil CO2 emission during the growing season in alpine meadows on the Tibetan Plateau. Atmos. Environ. 174, 92–98. doi:10.1016/j.atmosenv.2017.11.053
Han, C. X., Yu, R. H., Lu, X. X., Duan, L., Singh, V. P., and Liu, T. (2019). Interactive effects of hydrological conditions on soil respiration in China's horqin sandy land: an example of dune-meadow Cascade ecosystem. Sci. Total Environ. 651, 3053–3063. doi:10.1016/j.scitotenv.2018.10.198
Han, M., Liang, Y., Gao, Y., Yang, W., and Guo, Y. (2024). Relationship between root system-soil C:N:P and soil microbial diversity at different evolutionary stages of Caragana tibetica scrub in arid desert grassland, northern China. Front. Plant Sci. 15, 1423536. doi:10.3389/fpls.2024.1423536
Hancock, G. R., and Vallely, M. (2020). Effects of grazing exclusion on soil organic carbon: Hillslope and soil profile results (an Australian example). Sci. Total Environ. 705, 135844. doi:10.1016/j.scitotenv.2019.135844
Hanpattanakit, P., Leclerc, M. Y., Mcmillan, A. M. S., Limtong, P., Maeght, J.-L., Panuthai, S., et al. (2015). Multiple timescale variations and controls of soil respiration in a tropical dry dipterocarp forest, Western Thailand. Plant Soil 390, 167–181. doi:10.1007/s11104-015-2386-8
Hu, C., Lei, J.-P., and Wan, J.-Z. (2024). The relationships between biomass and soil respiration across different forest management practices. Forests 15, 712. doi:10.3390/f15040712
Jalilpour, M., Chavoshi, E., and Jalalian, A. (2022). Soil chemico-biological properties affected by the interactions of long-term non-cultivation practices and grazing. J. Soil Sci. Plant Nutr. 22, 4551–4565. doi:10.1007/s42729-022-00844-6
Ju, X., Wang, B., Wu, L., Zhang, X., Wu, Q., and Han, G. (2024). Grazing decreases net ecosystem carbon exchange by decreasing shrub and semi-shrub biomass in a desert steppe. Ecol. Evol. 14, e11528. doi:10.1002/ece3.11528
Kelsey, M. B., Mariana, M. A., Ivan, M., Krista, N. M., and Amélie, C. M. G. (2023). Long-term integrated crop-livestock grazing stimulates soil ecosystem carbon flux, increasing subsoil carbon storage in California perennial agroecosystems. Geoderma 438, 116598. doi:10.1016/j.geoderma.2023.116598
Kim, M., Lopez-Canfin, C., Lázaro, R., Sánchez-Cañete, E. P., and Weber, B. (2024). Unravelling the main mechanism responsible for nocturnal CO2 uptake by dryland soils. Sci. Total Environ. 926, 171751. doi:10.1016/j.scitotenv.2024.171751
Klimek, B., Chodak, M., and Niklińska, M. (2021). Soil respiration in seven types of temperate forests exhibits similar temperature sensitivity. J. Soils and Sediments 21, 338–345. doi:10.1007/s11368-020-02785-y
Knowles, J. F., Blanken, P. D., and Williams, M. W. (2015). Soil respiration variability across a soil moisture and vegetation community gradient within a snow-scoured alpine meadow. Biogeochemistry 125, 185–202. doi:10.1007/s10533-015-0122-3
Kölbl, A., Steffens, M., Wiesmeier, M., Hoffmann, C., Funk, R., Krümmelbein, J., et al. (2011). Grazing changes topography-controlled topsoil properties and their interaction on different spatial scales in a semi-arid grassland of Inner Mongolia, P.R. China. Plant soil 340, 35–58. doi:10.1007/s11104-010-0473-4
Li, G., Kim, S., Han, S. H., Chang, H., and Son, Y. (2017). Effect of soil moisture on the response of soil respiration to open-field experimental warming and precipitation manipulation. Forests 8, 56. doi:10.3390/f8030056
Li, J., Liu, X. M., Yao, C. S., Gong, H. H., Wu, C. Z., and Fan, H. L. (2021). Dynamie change of soil respiration and its effecting factors in different forestlands of wuyishan national park. Acta Ecol. Sin. 41, 3588–3602. doi:10.5846/stxb201905130981
Li, Q., Song, X., Chang, S. X., Peng, C., Xiao, W., Zhang, J., et al. (2019). Nitrogen depositions increase soil respiration and decrease temperature sensitivity in a moso bamboo forest. Agric. For. Meteorol. 268, 48–54. doi:10.1016/j.agrformet.2019.01.012
Li, S., Xing, T., Sa, R., Zhang, Y., Chen, H., Jin, K., et al. (2024). Effects of grazing on soil respiration in global grassland ecosystems. Soil Tillage Res. 238, 106033. doi:10.1016/j.still.2024.106033
Lin, L., Sun, X., Yu, Z., Huang, Y., and Zeng, D. (2020). Responses of soil respiration to N fertilization and grazing in a keerqin sandy grassland in northeast China. Arid. Land Res. Manag. 35, 230–245. doi:10.1080/15324982.2020.1827464
Lloyd, D. A., Ritz, K., Paterson, E., and Kirk, G. J. D. (2016). Effects of soil type and composition of rhizodeposits on rhizosphere priming phenomena. Soil Biol. biochem. 103, 512–521. doi:10.1016/j.soilbio.2016.10.002
Luo, J., Chen, Y., Wu, Y., Shi, P., She, J., and Zhou, P. (2012). Temporal-spatial variation and controls of soil respiration in different primary succession stages on glacier forehead in gongga Mountain, China. PLOS ONE 7, e42354. doi:10.1371/journal.pone.0042354
Mande, H. K., Abdullah, A. M., Aris, A. Z., and Ainuddin, A. N. (2014). Factors responsible for spatial and temporal variation of soil CO2 efflux in a 50 year recovering tropical forest, peninsular Malaysia. Environ. Earth Sci. 73, 5559–5569. doi:10.1007/s12665-014-3810-8
Mekuria, W., Veldkamp, E., Haile, M., Nyssen, J., Muys, B., and Gebrehiwot, K. (2007). Effectiveness of exclosures to restore degraded soils as a result of overgrazing in Tigray, Ethiopia. J. Arid. Environ. 69, 270–284. doi:10.1016/j.jaridenv.2006.10.009
Naidu, D. G. T., Roy, S., and Bagchi, S. (2022). Loss of grazing by large mammalian herbivores can destabilize the soil carbon pool. Proc. Natl. Acad. Sci. U.S.A. 43, e2211317119. doi:10.1073/pnas.2211317119
Navasardyan, M., Sargsyan, T., Daveyan, H., Mezhunts, B., and Abraham, E. M. (2024). Effect of grazing on plant and soil parameters of steppe pastures on Mount Aragats, Armenia. Armen. Land 13 (9), 1430. doi:10.3390/land13091430
Ngao, J., Epron, D., Delpierre, N., Bréda, N., Granier, A., and Longdoz, B. (2012). Spatial variability of soil CO2 efflux linked to soil parameters and ecosystem characteristics in a temperate beech forest. Agric. For. Meteorol. 154, 136–146. doi:10.1016/j.agrformet.2011.11.003
Nie, C., Li, Y., Niu, L., Liu, Y., Shao, R., Xu, X., et al. (2019). Soil respiration and its Q10 response to various grazing systems of a typical steppe in Inner Mongolia, China. PeerJ 7, e7112. doi:10.7717/peerj.7112
Nüsse, A., Linsler, D., Kaiser, M., Ebeling, D., Tonn, B., Isselstein, J., et al. (2017). Effect of grazing intensity and soil characteristics on soil organic carbon and nitrogen stocks in a temperate long-term grassland. Arch. Agron. Soil Sci. 63, 1776–1783. doi:10.1080/03650340.2017.1305107
Odriozola, I., García-Baquero, G., Laskurain, N. A., and Aldezabal, A. (2014). Livestock grazing modifies the effect of environmental factors on soil temperature and water content in A temperate grassland. Geoderma 235–236, 347–354. doi:10.1016/j.geoderma.2014.08.002
Ondier, J., Okach, D. O., John, O. C., and Otieno, D. O. (2020). Influence of rainfall amount and livestock grazing on soil respiration in a moist Kenyan Savannah. Afr. J. Ecol. 58, 92–99. doi:10.1111/aje.12670
Pan, S., Shi, J., Peng, Y., Wang, Z., and Wang, X. (2023). Soil organic carbon pool distribution and stability with grazing and topography in a Mongolian grassland. Agric. Ecosyst. Environ. 348, 108431. doi:10.1016/j.agee.2023.108431
Pang, D., and Xu, H. (2024). Carbon sequestration and stability and soil erosion in forest ecosystems. Forests 15, 1961. doi:10.3390/f15111961
Peri, P. L., Bahamonde, H., and Christiansen, R. (2015). Soil respiration in patagonian semiarid grasslands under contrasting environmental and use conditions. J. Arid. Environ. 119, 1–8. doi:10.1016/j.jaridenv.2015.03.008
Post, W. M., and Kwon, K. C. (2000). Soil carbon sequestration and land-use change: processes and potential. Glob. Change Biol. 6, 317–327. doi:10.1046/j.1365-2486.2000.00308.x
Qu, Y., Wang, D., Jin, S., Zheng, Z., Diao, Z., and Rong, Y. (2024). Flooding length mediates fencing and grazing effects on soil respiration in meadow steppe. Plants 13, 666. doi:10.3390/plants13050666
Raich, J. W., Potter, C. S., and Bhagawati, D. (2022). Interannual variability in global soil respiration, 1980-94. Glob. Change Biol. 8, 800–812. doi:10.1046/j.1365-2486.2002.00511.x
Raiesi, F., and Asadi, E. (2006). Soil microbial activity and litter turnover in native grazed and ungrazed rangelands in a semiarid ecosystem. Biol. Fertil. Soils. 43, 76–82. doi:10.1007/s00374-005-0066-1
Reeder, J. D., and Schuman, G. E. (2002). Influence of livestock grazing on C sequestration in semi-arid mixed-grass and short-grass rangelands. Environ. Pollut. 116, 457–463. doi:10.1016/s0269-7491(01)00223-8
Robertson, F., Crawford, D., Partington, D., Oliver, I., Rees, D., Aumann, C., et al. (2016). Soil organic carbon in cropping and pasture systems of Victoria, Australia. Soil Res. 54, 64–77. doi:10.1071/sr15008
Rubio, V. E., and Detto, M. (2017). Spatiotemporal variability of soil respiration in a seasonal tropical forest. Ecol. Evol. 7, 7104–7116. doi:10.1002/ece3.3267
Seabloom, E. W., Borer, E. T., and Tilman, D. (2020). Grassland ecosystem recovery after soil disturbance depends on nutrient supply rate. Ecol. Lett. 23, 1756–1765. doi:10.1111/ele.13591
Sharkhuu, A., Plante, A. F., Enkhmandal, O., Gonneau, C., Casper, B. B., Boldgiv, B., et al. (2016). Soil and ecosystem respiration responses to grazing, watering and experimental warming chamber treatments across topographical gradients in northern Mongolia. Geoderma 269, 91–98. doi:10.1016/j.geoderma.2016.01.041
Shen, H., Zhang, T., Zhao, Y., Wu, A., Zheng, Z., and Cao, J. (2023). Effects of precipitation variation on annual and winter soil respiration in a semiarid Mountain shrubland in northern China. Sustainability 15, 7461. doi:10.3390/su15097461
Shen, Y., Fang, Y., Chen, H., Ma, Z., Huang, C., Wu, X., et al. (2023). New insights into the relationships between livestock grazing behaviors and soil organic carbon stock in an alpine grassland. Agric. Ecosyst. Environ. 355, 108602. doi:10.1016/j.agee.2023.108602
Sinsabaugh, R. L., Carreiro, M. M., and Repert, D. A. (2002). Allocation of extracellular enzymatic activity in relation to litter composition, N deposition, and mass loss. Biogeochemistry 60, 1–24. doi:10.1023/A:1016541114786
Song, X., Zhu, Y., and Chen, W. (2021). Dynamics of the soil respiration response to soil reclamation in a coastal wetland. Sci. Rep. . 11, 2911. doi:10.1038/s41598-021-82376-0
Tian, Q., Wang, D., Tang, Y., Li, Y., Wang, M., Liao, C., et al. (2019). Topographic controls on the variability of soil respiration in a humid subtropical forest. Biogeochemistry 145, 177–192. doi:10.1007/s10533-019-00598-x
Van Kessel, C., Pennock, D. J., and Farrell, R. E. (1993). Seasonal variations in denitrification and nitrous oxide evolution at the landscape scale. Soil Sci. Soc. Am. J. 57, 988–995. doi:10.2136/sssaj1993.03615995005700040018x
Vermeire, L. T., Wester, D. B., Mitchell, R. B., and Fuhlendorf, S. D. (2005). Fire and grazing effects on wind erosion, soil water content, and soil temperature. J. Environ. Qual. 34, 1559–1565. doi:10.2134/jeq2005.0006
Vinograd, A., Zaady, E., and Kigel, J. (2019). Dynamics of soil nutrients in abandoned sheep corrals in semi-arid mediterranean planted forests under grazing. J. Arid Environ. 164 (MAY), 38–45. doi:10.1016/j.jaridenv.2019.02.007
Wang, G., Mao, J., Fan, L., Ma, X., and Li, Y. (2022). Effects of climate and grazing on the soil organic carbon dynamics of the grasslands in northern Xinjiang during the past twenty years. Glob. Ecol. Conserv. 34, e02039. doi:10.1016/j.gecco.2022.e02039
Wang, H., Wang, S., Li, J., and Alimira Yerken, A. (2017). Characteristics and the influencing factors of forest soil respiration: a rview. J. Northwest For. Univ. 32, 92–97. doi:10.3969/j.issn,1001-7461.2017.01.15
Wang, J., Li, W., Cao, W., and Wang, S. (2022). Effects of different intensities of long-term grazing on plant diversity, biomass and carbon stock in alpine shrubland on the Qinghai-Tibetan Plateau. PeerJ 10, e12771. doi:10.7717/peerj.12771
Wang, Y., Wang, Z., Li, H., Shen, T., Zhang, X., Li, J., et al. (2023). Grazing decreased soil organic carbon by decreasing aboveground biomass in a desert steppe in Inner Mongolia. J. Environ. Manag. 347, 119112. doi:10.1016/j.jenvman.2023.119112
Wang, Z., Wan, X., Tian, M., Wang, X., Chen, J., Chen, X., et al. (2020). Grazing season alters soil respiration in a semiarid grassland on the loess Plateau. Nutr. Cycl. Agroecosyst 118, 177–191. doi:10.1007/s10705-020-10092-8
Webster, K. L., Creed, I. F., Bourbonnière, R. A., and Beall, F. D. (2018). Sensitivity of catchment-aggregated estimates of soil carbon dioxide efflux to topography under different climatic conditions. J. Geophys. Res. 113, G03018. doi:10.1029/2008jg000707
Wei, Y., Wei, B., Ryo, M., Bi, Y., Sun, X., Zhang, Y., et al. (2023). Grazing facilitates litter-derived soil organic carbon formation in grasslands by fostering microbial involvement through microenvironment modification. Catena 232, 107389. doi:10.1016/j.catena.2023.107389
Wen, L., Jinlan, W., Xiaojiao, Z., Shangli, S., and Wenxia, C. (2018). Effect of degradation and rebuilding of artificial grasslands on soil respiration and carbon and nitrogen pools on an alpine meadow of the Qinghai-Tibetan Plateau. Ecol. Eng. 111, 134–142. doi:10.1016/j.ecoleng.2017.10.013
Wienhold, B. J., Hendrickson, J. R., and Karn, J. F. (2001). Pasture management influences on soil properties in the northern great plains. J. Soil Water Conserv. 56, 27–31. doi:10.1080/00224561.2001.12457351
Xu, M., and Shang, H. (2016). Contribution of soil respiration to the global carbon equation. J. Plant Physiol. 203, 16–28. doi:10.1016/j.jplph.2016.08.007
Xu, W., and Wan, S. (2008). Water- and plant-mediated responses of soil respiration to topography, fire, and nitrogen fertilization in a semiarid grassland in northern China. Soil Biol. biochem. 40, 679–687. doi:10.1016/j.soilbio.2007.10.003
Xu, X., Shi, Z., Li, D., Rey, A., Ruan, H., Craine, J. M., et al. (2016). Soil properties control decomposition of soil organic carbon: results from data-assimilation analysis. Geoderma 262, 235–242. doi:10.1016/j.geoderma.2015.08.038
Yao, W., Nan, F., Li, Y., Li, Y., Liang, P., and Zhao, C. (2023). Effects of different afforestation years on soil properties and quality. Forests 14, 329. doi:10.3390/f14020329
Yao, X., Wu, J., Gong, X., Lang, X., and Wang, C. (2019). Grazing exclosures solely are not the best methods for sustaining alpine grasslands. PeerJ 7, e6462. doi:10.7717/peerj.6462
Zhang, B., Qu, Z., Lv, S., Han, G., Gao, C., Wilkes, A., et al. (2023). Grazing effects on total carbon and nitrogen content of wind-eroded soils in desert steppe. Land Degrad. Dev. 34, 5328–5342. doi:10.1002/ldr.4847
Zhang, X., Tan, Y., Zhang, B., Li, A., Daryanto, S., Wang, L., et al. (2017). The impacts of precipitation increase and nitrogen addition on soil respiration in a semiarid temperate steppe. Ecosphere 8, e01655. doi:10.1002/ecs2.1655
Zhao, F., Ren, C., Shelton, S., Wang, Z., Pang, G., Chen, J., et al. (2017). Grazing intensity influence soil microbial communities and their implications for soil respiration. Agric. Ecosyst. Environ. 249, 50–56. doi:10.1016/j.agee.2017.08.007
Zhao, J. X., Wang, S. J., Chen, Q. B., Chen, M. Q., Shu, J. J., and Li, Y. J. (2015). Soil respiration and its affecting factors of pinus Yunnanensisin the middle regions of Yunnan. J. Northwest For. Univ. 30, 8–13+20. doi:10.3969/j.issn.1001-7461.2015.03.02
Zheng, P., Wang, D., Yu, X., Jia, G., Liu, Z., Wang, Y., et al. (2021). Effects of drought and rainfall events on soil autotrophic respiration and heterotrophic respiration. Agric. Ecosyst. Environ. 308, 107267. doi:10.1016/j.agee.2020.107267
Zhou, J., Chen, Z., Yang, Q., Jian, C., Lai, S., Chen, Y., et al. (2021). N and P addition increase soil respiration but decrease contribution of heterotrophic respiration in semiarid grassland. Agric. Ecosyst. Environ. 318, 107493. doi:10.1016/j.agee.2021.107493
Keywords: Caragana korshinskii, grazing, soil respiration, soil physicochemical properties, biomass
Citation: Wu H, Deng X, Li X, Gao S, Yuan R, Li J, Min H and Ma X (2025) Effects of grazing on soil respiration and associated factors in artificial Caragana korshinskii shrublands in Inner Mongolia, China. Front. Environ. Sci. 13:1639769. doi: 10.3389/fenvs.2025.1639769
Received: 03 June 2025; Accepted: 22 July 2025;
Published: 31 July 2025.
Edited by:
Dima Chen, Inner Mongolia University, ChinaReviewed by:
Bing Wang, China Three Gorges University, ChinaJianjun Li, Jiangxi Agricultural University, China
Copyright © 2025 Wu, Deng, Li, Gao, Yuan, Li, Min and Ma. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiuzhi Ma, bHVja214eUBpbWF1LmVkdS5jbg==