 Valentí Rull1*
Valentí Rull1* Encarni Montoya1
Encarni Montoya1 Irantzu Seco1
Irantzu Seco1 Núria Cañellas-Boltà1
Núria Cañellas-Boltà1 Santiago Giralt1
Santiago Giralt1 Olga Margalef2
Olga Margalef2 Sergi Pla-Rabes2William D'Andrea3Raymond Bradley4
Sergi Pla-Rabes2William D'Andrea3Raymond Bradley4 Alberto Sáez5
Alberto Sáez5- 1Institute of Earth Sciences Jaume Almera, ICTJA, CSIC, Barcelona, Spain
- 2Ecological Research Center and Forestry Applications, Cerdanyola del Vallès, Spain
- 3Lamont-Doherty Earth Observatory, Columbia University, Palisades, NY, United States
- 4Climate System Research Center, University of Massachusetts, Amherst, MA, United States
- 5Department of Earth and Ocean Dynamics, Universitat de Barcelona, Barcelona, Spain
This paper reviews the existing hypotheses concerning the cultural shift from the Ancient Cult (AC) to the Birdman Cult (BC) that occurred on Easter Island (Rapa Nui) during the last millennium and introduces a holistic new hypothesis called CLAFS (Climate-Landscape-Anthropogenic Feedbacks and Synergies), which considers a variety of potential drivers of cultural change and their interactions. The CLAFS hypothesis can be tested with future paleoecological studies on new sedimentary sequences such as the new continuous and coherent record encompassing the last millennium from Rano Kao (KAO08-03) using a combination of pollen, non-pollen palynomorphs (NPP), charcoal, and fecal lipid analyses, at decadal to multidecadal resolution. The Kao record should be compared with other continuous records of the last millennium available for the two other freshwater bodies of the island, Rano Aroi and Rano Raraku, to obtain an island-wide perspective of spatio-temporal deforestation patterns in relation to climatic shifts and human activities. The CLAFS hypothesis predicts that the shift from the AC to the BC was associated with the drying out and deforestation of Rano Raraku (the center of the AC) by ~1,570 CE, followed by human migration to Rano Kao (the social center of the BC), where freshwater and forests were still available. Under the CLAFS scenario, this migration would have occurred by ~1,600 CE. Findings to the contrary would require modification and refinement, or outright rejection, of the CLAFS hypothesis and the consideration of alternate hypotheses compatible with new paleoecological evidence. Regardless the final results, archeological evidence will be required to link climatic and ecological events with cultural developments.
Introduction
Easter Island (Rapa Nui) is a small and remote Pacific island of triangular shape (Figure 1) formed by the coalescence of three submarine volcanoes that emerged from the sea ~780 kyrs ago and whose latest major eruption occurred 200–110 kyrs ago (González-Ferrán et al., 2004; Vezzolli and Acocella, 2009). Easter Island has been considered a microcosm to illustrate the potential consequences of the worldwide overexploitation of natural resources (Diamond, 2005). According to the ecocide hypothesis, the original Rapanui society caused their own destruction by resource exhaustion, as manifested in the total removal of forests that had covered the island for millennia before human settlement (Flenley and Bahn, 2003). This hypothesis has dominated the Easter island narrative for decades since its formulation based on pollen analysis of lake sediments, which showed an abrupt change from palm to grass pollen, interpreted as the abrupt replacement of palm-dominated forests by grass meadows similar to those that cover the island today (Flenley and King, 1984; Flenley et al., 1991).
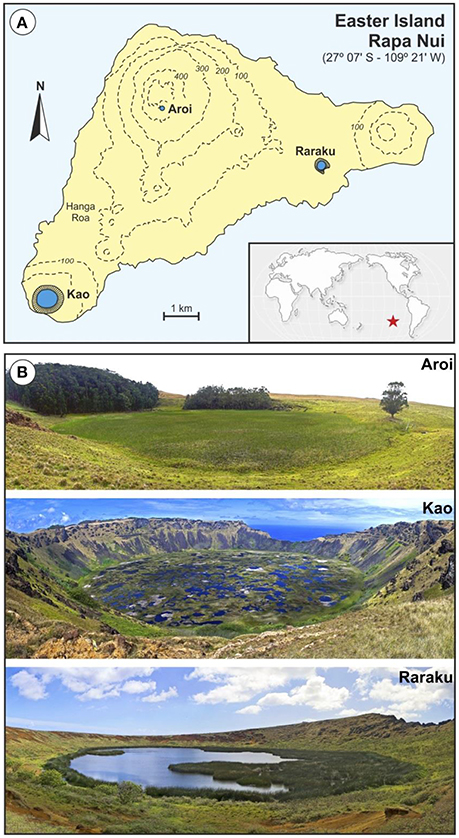
Figure 1. Location map and cored lakes. (A) Sketch map of Easter Island showing the location of the three freshwater bodies cored for paleoecological studies (Aroi, Kao and Raraku). The position of the island in the Pacific Ocean is indicated by a red star. (B) Permanent freshwater lakes and mires from Easter Island (Photos: V. Rull).
Paleoecological reconstructions (notably palynological analyses to examine vegetation and landscape dynamics and charcoal analyses to document fire history) have been instrumental in the study of ecological and cultural change on Easter Island (Rull et al., 2010). Most paleoecological records have been obtained using sediment cores from the three natural freshwater bodies of the island: a mire (Aroi) and two lakes (Kao and Raraku), all of which are located inside volcanic craters; a combination that is locally called “rano” (Herrera and Custodio, 2008) (Figure 1). The sedimentary sequences underlying these water bodies have provided long records extending back to the last glaciation (Flenley and King, 1984; Flenley et al., 1991; Sáez et al., 2009; Margalef et al., 2013). However, detailed records of the last millennia, when the most relevant ecological and cultural shifts seem to have occurred, are rare, which can be attributed to intrinsic sedimentary challenges (Rull, 2016b). Since the earlier studies (Flenley and King, 1984; Flenley et al., 1991), a major obstacle for paleoecological reconstruction has been the difficulty in obtaining continuous and chronologically coherent records, free from sedimentary gaps, and age inversions (Butler et al., 2004). This has been especially problematic for the last millennia, where extensive sedimentary gaps have impeded continuous paleoecological reconstructions (Figure 2). The apparent abruptness of the island's deforestation, cited as supporting evidence for the ecocide hypothesis, is considered to be an artifact resulting from a sedimentary hiatus that obscured vegetation shifts occurred between about 4.0–4.2 and 0.8 cal kyr BP (Mann et al., 2008; Sáez et al., 2009) and created the incorrect impression that replacement of palm forests by grass meadows occurred almost instantaneously (Rull et al., 2013).
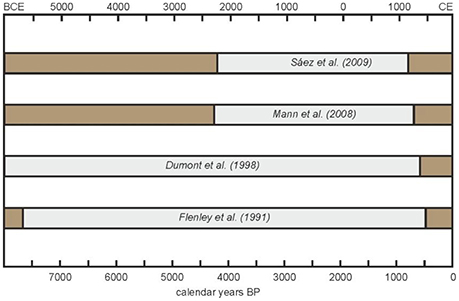
Figure 2. Sedimentary gaps (gray bars) from previous paleoecological analyses on Rano Raraku sediments, from where most paleoecological inferences have been obtained. Redrawn from Rull et al. (2013).
During the last decade, new coring efforts have provided more complete records for the last millennia. Noteworthy are two nearly continuous and chronologically coherent sequences from Aroi and Raraku embracing roughly the last three millennia, which have allowed the reconstruction of deforestation patterns across the island and revealed significant spatial heterogeneities (Cañellas-Boltà et al., 2013; Rull et al., 2015). Such recent studies have also provided evidence for the potential role of climatic shifts and climate-human synergies on the island's ecological and cultural developments, which has challenged former paradigms (Rull et al., 2013). Similar paleoecological reconstructions are still unavailable for Rano Kao, where the attainment of reliable age-depth models for the last millennia has been hampered primarily by the occurrence of frequent age inversions (Flenley and King, 1984; Flenley et al., 1991; Butler et al., 2004; Butler and Flenley, 2010; Horrocks et al., 2012b, 2013). Gossen (2007, 2011) obtained a coherent sequence for the last millennia but paleoecological analyses similar to those of Aroi and Raraku, useful to unravel deforestation trends, are unavailable.
In this paper, we introduce the first continuous and coherent sequence from Rano Kao encompassing the last millennium and discuss the potential significance of its paleoecological analysis, which is in progress, for testing previous and novel hypotheses about deforestation in relation to eventual climate changes and/or cultural developments (Rull, 2016a). This Kao sequence will complement the aforementioned Aroi and Raraku records of the last millennia to provide an island-wide history, capable of resolving spatio-temporal trends and patterns of deforestation, as well as addressing questions concerning human occupation and eventual intra-island migrations. The paper begins with an account of cultural developments of the last millennium in relation to ecological shifts, with an emphasis on the island's deforestation. The next section discusses the potential role of climate and climate-human interactions on ecological and cultural shifts. Then, Rano Kao and its sediments are briefly introduced and the new continuous record from this lake is presented, showing the age-depth model and describing the analyses that are in progress. Finally, the potential significance of this new record is discussed in light of the hypotheses and theories previously debated and others that can emerge from the new sedimentary evidence.
Cultural Change and Deforestation
Current archeological evidence seems to point to a relatively recent human settlement of Easter Island by 1,100–1,300 CE (Hunt and Lipo, 2006; Wilmshurst et al., 2011), although earlier dates ranging from 800 to 1,000 CE have also been proposed (Flenley and Bahn, 2003; Vargas et al., 2006). Regarding the origin of colonizers, the dominant view is that they proceeded from East Polynesia (Flenley and Bahn, 2003) but potential contributions from Amerindian cultures cannot be dismissed (Thorsby, 2016). Therefore, the main cultural developments of Easter Island seem to have occurred in the last millennium, during which two major cultural transformations have been documented. The first was a profound change in lifestyle, social organization and ritual practices within the Rapanui society symbolized by the shift from the so called Ancient Cult to the Birdman Cult (Edwards and Edwards, 2013). The second cultural change was the consequence of the arrival of the first Europeans in 1722. In Easter Island, this date marks the boundary between prehistoric and historical times.
The Prehistoric Cultural Shift
During the Ancient Cult (AC) phase, the standard worship subjects were the moais, the iconic megalithic and anthropomorphic statues that have become the most popular symbol of Easter Island. The moais represented deified ancestors and were considered to guarantee land and sea fertility and, hence, social prosperity (Edwards and Edwards, 2013). The socio-political center of the AC was Rano Raraku, the quarry were almost all moais were carved from the relatively soft volcanic tuff that forms this crater using basalt tools (metals were not known) obtained from other craters (Gioncada et al., 2010). During those times, the Rapanui culture flourished and its society attained its population maximum, estimated to be approximately 6,000–8,000 inhabitants (range: 3,000–20,000) (Brandt and Merico, 2015; Puleston et al., 2017). The Rapanui society was organized into clans (Figure 3) in a hierarchical pyramidal organization where the maximum authority of the island, the Ariki Mau, was invariably the chief of the Miru clan, considered to directly descend from the first Polynesian settlers. Rapanui society was organized around the moai industry (carving, transport, and emplacement on their final platforms or ahu), and people involved in these tasks, especially sculpture makers, were a privileged class (Flenley and Bahn, 2003). The process of moai building and the related industry has been called ahu moai.
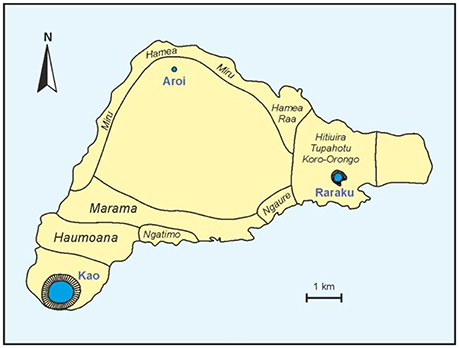
Figure 3. Sketch-map of Easter Island showing the clans of the ancient Rapanui civilization. Redrawn from Routledge (1919).
The shift to the Birdman Cult (BC) represented a cultural revolution (Robinson and Stevenson, 2017). The cult of the moais declined and was eventually abandoned and the religion became monotheistic, with Make Make as the highest deity. The hierarchical dynasty-based political system disappeared and was replaced by a system in which the most powerful people were the clan chiefs and the warrior leaders (matato'a). In the BC, the symbols of fertility and prosperity were migratory birds, notably the sooty tern or manutara (Onychoprion fuscatus) nesting on the surrounding islets, which was considered to be a nexus between the divine and the human worlds (Edwards and Edwards, 2013). The maximum authority (the Birdman or tangata manu) or the representative of Make Make on Earth) was renewed yearly following a competition among representatives of all clans. The Birdman of the year was the chief of the clan whose representative (hopu manu) was able to swim to the Motu Nui islet, obtain the first sooty-tern egg of the season and carry it back, in intact condition, to the ceremonial village of Orongo, situated on the SW rim of the Kao crater (Figures 4, 7). Therefore, the geographical core of the Rapanui culture shifted from Rano Raraku, the moai quarry, to Rano Kao, the headquarters of the BC (Kirch, 2000).

Figure 4. The ceremonial village of Orongo at the SW rim of the Kao crater. See Figure 7 for the precise location. (A) Google Earth view showing the typical ellipsoidal boat-shaped stone houses. (B) Close front view of a typical stone house (Photo: N. Cañellas).
The timing of the AC-BC shift has been debated for a long time, mainly on the basis of oral tradition, which may have been subject to historical biases and distortions (Hunt and Lipo, 2011). A minimum age could be provided by the foundation of the ceremonial village of Orongo, the center of the BC (Robinson and Stevenson, 2017) (Figure 4). The first excavations, performed by the Norwegian expedition led by Thor Heyerdahl gave radiocarbon ages between 1,540 and 1,576 CE for the beginning of room construction in Orongo (Ferdon, 1961). However, these ages have been considered unreliable due to the lack of accurate sample descriptions (Robinson and Stevenson, 2017) and dubious stratigraphic correlations (Golson, 1965). Robinson and Stevenson (2017) summarized all the reliable chronometric information available from radiocarbon and obsidian hydration dating (Orliac and Orliac, 1996; Stevenson et al., 2013) and concluded that the first petroglyph complexes and stone houses of Orongo are not older than 1,600 CE and the activities on the site experienced an intensification in the very late 18th century (Figure 5), well after the European contact (Robinson and Stevenson, 2017).
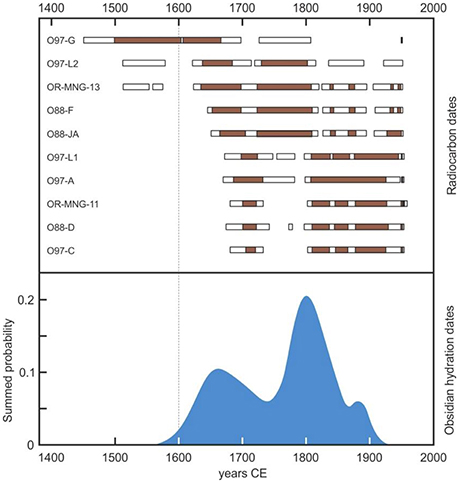
Figure 5. Summary of all reliable radiocarbon and obsydian hydration dates obtained in Orongo. Redrawn from Robinson and Stevenson (2017), after original data from Orliac and Orliac (1996) and Stevenson et al. (2013). Radiocarbon dates are expressed as 1 α (brown bars) and 2 α (white bars) intervals.
Additional evidence of human activity in the Kao catchment has been found in the lake shores just beneath Orongo (Figures 4, 7), where pollen, phytolith, and starch analyses of marginal sediment cores have documented relatively intense horticultural activity based on cultigens of Polynesian origin (Horrocks et al., 2012b, 2013, 2016). Unfortunately, sediments had been mixed by human activity, which has prevented the development of a reliable chronological model. Therefore, it is not possible to know whether agricultural activities on lake terraces predated the foundation of Orongo. In summary, the available evidence points toward a minimum age of 1,600 CE for the AC-BC cultural shift. According to Van Tilburg (1996), however, the AC and the BC would have overlapped for some time and the cultural shift would have not been abrupt but transitional (Edwards and Edwards, 2013).
The Ecocide Hypothesis
According to the ecocide hypothesis, the AC-BC shift was a direct consequence of the island's deforestation. Based primarily on paleoecological evidence, the proponents of this hypothesis argue that deforestation commenced by 1,200–1,300 CE and was complete by 1,400 CE or, at the latest, by 1,600–1,650 CE (Orliac, 2000; Flenley and Bahn, 2003; Mann et al., 2008; Mieth and Bork, 2010). Mieth and Bork (2010) estimated that, prior to deforestation, ~16 million palm trees would have dominated the forests that covered ~70% of the island. The botanical identity of this palm is still a matter of controversy but the dominant view is that the species was endemic to Easter Island and is now extinct (Dransfield et al., 1994). In the ecocide scenario, deforestation would have been part of a general process of natural resource exhaustion by uncontrolled exploitation leading to an ecological crash and a profound crisis of the Rapanui culture (Flenley and Bahn, 2003; Diamond, 2005). Such ecological and cultural collapse would have instigated a general degradation of Rapanui society characterized by dramatic demographic reduction due to starvation, social conflicts and internal warfare, as well as a general and profound reorganization characterized by the shift to the BC. It has also been suggested that the abandonment of the moai cult included the intentional toppling of these statues, a practice called huri moai. According to some model-based estimates, the demographic reduction of Rapanui would have been >80%, from ~14,000 to ~2,000 inhabitants (Brandt and Merico, 2015), who lived in precarious conditions, mostly inhabiting lava-tube caves for protection. Some proponents of the ecocide hypothesis interpret the geographical shift of the Rapanui cultural center from Raraku to Kao and the onset of the BC as a manifestation of newly acquired supremacy of western clans over eastern ones (Figure 3), which had dominated the scene until then, following internal conflicts (McCall, 1980; Flenley and Bahn, 2003).
Cultural Resilience and the Historical Genocide
An alternative hypothesis is that deforestation was not caused directly by humans but by massive consumption of palm fruits by rats, thus preventing forest regeneration. This idea is based on the frequent occurrence in soils and archeological sites of endocarps of palm fruits with rat gnaw-markings (Hunt, 2006, 2007; Hunt and Lipo, 2011). Rats would have been of Polynesian origin, likely transported by the first settlers (West et al., 2017). In this scenario, the timing of deforestation does not differ considerably from that of the ecocide hypothesis. Regardless of the ultimate cause of deforestation, a number of researchers dismiss the occurrence of social crisis following forest clearing and argue that the Rapanui were resilient to forest removal and remained healthy until European contact (Hunt and Lipo, 2011; Boersema, 2015; Jarman et al., 2017). Analysis of widespread land-use evidence and radiocarbon dates from archeological sites across the island support this hypothesis and suggest that the ancient Rapanui developed sustainable agricultural practices and did not experience a social crisis prior to Europen arrival (Stevenson and Haoa-Cardinali, 2008; Mulrooney, 2013; Stevenson et al., 2015). According to this view, the cultural collapse of Rapanui society was in fact a genocide, which started with the European arrival and was the outcome of direct violence, the introduction of unknown epidemic diseases (smallpox, syphilis, tuberculosis), and slave trading (Hunt and Lipo, 2011). Such post-contact cultural and demographic degradation of the Rapanui society has been confirmed by historical documents (Routledge, 1919; Métraux, 1940) and seems to have been especially intense during the 19th century (McCall, 1980; Fischer, 2005; Boersema, 2015). In this scenario, the AC-BC shift, the deforestation and the collapse of the ancient Rapanui culture are separate events that are not causally linked. A pre-contact population reduction is not dismissed but is attributed to land-use heterogeneities leading to an eventual decline in food production, rather than to a hypothetical island-wide pre-contact collapse, as proposed by the ecocidal theory (Stevenson et al., 2015; Puleston et al., 2017).
Other Hypotheses
Other hypotheses have been proposed to explain cultural changes on the Rapanui during the last millennium. For example, a theory combining elements from both the ecocide and genocide hypotheses was developed by Brandt and Merico (2015), who modeled the demographic evolution of Rapanui society since 1,200 CE and suggested an initial population decline around 1,400 CE, likely linked to deforestation, and a second decline by 1,800 CE, as a consequence of European contact. Others contend that the collapse of Rapanui society would have occurred in a short time period between European contact (1,722 CE) and the visit of Captain Cook (1,774 CE). This is based on the respective navigation logs, the first reporting a healthy society still practicing moai worship and the second documenting a depauperate island with toppled moais. Some speculate that the change must have been due to a catastrophic event between the times of the two visits, such as an internal war (Fischer, 2005; Hunt and Lipo, 2011).
Climatic and Ecological Shifts
The hypotheses discussed above attributed ecological and cultural changes on Easter Island to anthropogenic causes and this has become the prevalent narrative. McCall (1993) speculated that climatic changes, especially eventual Little Ice Age (LIA) droughts, should also have had a role. However, this proposal was ignored or explicitly dismissed owing to the lack of empirical support (Flenley and Bahn, 2003). However, the intensification of coring campaigns and paleoecological studies during the past decade have challenged this paradigm (Rull et al., 2013, 2016). For example, Mann et al. (2008) and Sáez et al. (2009) interpreted the sedimentary gap recorded in the Raraku peaty sediments between about 4.0–4.2 and 0.8 cal kyr BP (Figure 2) as evidence for drought and lowered lake levels, which would have caused subaerial exposure and erosion of former wetland sediments. The fairly continuous Raraku and Aroi records obtained later closed the sedimentary gap and allowed for refined paleoecological interpretation, with authors proposing the existence of two episodes of drought at about 880–1,170 CE and 1,570–1,720 CE, respectively (Cañellas-Boltà et al., 2013). The first encompassed most of the Medieval Climate Anomaly (MCA) and the second occurred during the LIA (Rull et al., 2016), supporting the initial proposal of McCall (1993), that there is a connection between climatic severity and Rapanui societal changes. However, the link between environmental and cultural shifts remained unclear (Rull, 2016a). Palynological analysis of the Raraku and Aroi continuous sequences provided more information in this regard.
In the Raraku catchment, a significant forest reduction coinciding with the continuous presence of charcoal was recorded shortly after the MCA drought (880–1,170 CE) (Figure 6) and was interpreted as anthropogenic forest clearing using fire (Cañellas-Boltà et al., 2013). The timing is consistent with early deforestation hypotheses and the evidence for burning points toward clearance by humans, rather than deforestation by rats. However, forest removal was not complete and open palm woodlands remained around the lake. It is possible that drought would have increased vegetation flammability by increasing the accumulation of dead biomass or by decreasing the moisture content of live biomass (Bond and Keane, 2017), thus facilitating burning practices. The anthropogenic hypothesis is reinforced by the fact that climatic dryness did not affect the open palm woodlands that covered Rano Aroi area, which was likely inhabited by those times (Horrocks et al., 2015; Rull et al., 2015).
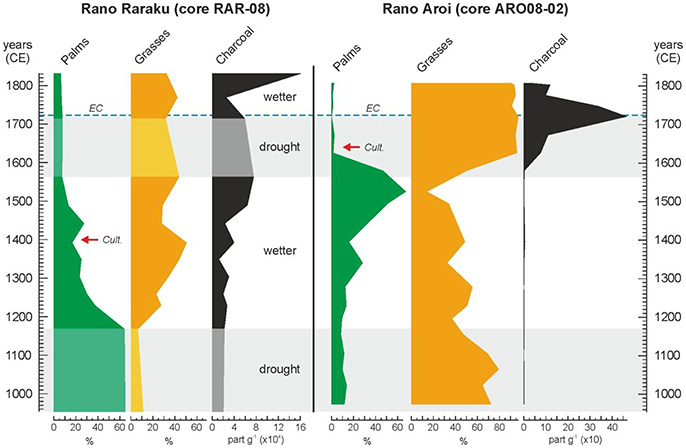
Figure 6. Summary pollen diagrams of Raraku and Aroi continuous sequences for the last millennia. Redrawn from Cañellas-Boltà et al. (2013) and Rull et al. (2015). In Raraku, light-colored areas indicate the sedimentary gaps with no pollen data. Red arrows indicate the first evidence of cultivation according to Horrocks et al. (2012a, 2015). The European contact (EC) is indicated by a dotted blue line.
A subsequent humid phase, between about 1,200 and 1,550 CE, coincided with a significant increase of palm forests in Rano Aroi but not in Rano Raraku (Figure 6), perhaps due to the continued human pressure (Rull et al., 2016). Indeed, fires persisted in Raraku and the first signs of cultivation appeared around 1,400 CE (Horrocks et al., 2012a). A second deforestation event, linked to an increase in evidence for burning, took place in Raraku by 1,450 CE and palm forests disappeared from the catchment by 1,500 CE (Cañellas-Boltà et al., 2013). In Aroi, wet climatic conditions and the absence of human settlements, would have favored palm forest expansion and/or densification.
During the LIA drought (1,570–1,720 CE), the Raraku catchment was likely devoid of forests and freshwater due to the combination of former human deforestation practices and the drought-induced desiccation of the lake (Cañellas-Boltà et al., 2013). The Aroi forests still remained but were totally removed by 1,620 CE, also with the aid of fire, roughly a century after the deforestation around Raraku (Rull et al., 2015) (Figure 6). During this dry phase, by 1,640 CE, Rano Aroi started to be cultivated (Horrocks et al., 2015), which suggests the presence of humans and enough freshwater availability for agriculture. A subsequent humid phase did not change the situation in Raraku, where forests did not recover likely due to the continued fires. A similar situation was recorded in Aroi, where fires peaked at about 1,720 CE and strongly receded later (Figure 6).
In addition to climate change, other natural hazards such as regional volcanic eruptions or tsunamis have been considered potential drivers for ecological and cultural change across the Pacific archipelagos, including Easter Island (Margalef et al., in press). Evidence for the influence of these environmental shifts on Easter Island is still to be identified in sedimentary records but the interest on verifying this possibility is increasing due to geomorphological and documentary evidence for the impact of historical tsunamis on some coastal moai complexes (Cortez et al., 2009).
An Alternative Scenario
The combination of the available paleoecological evidence and the known cultural developments of the ancient Rapanui society described above may provide a new holistic scenario that considers climatic changes and human activities, as well as their corresponding feedbacks and synergies, as the main drivers of Easter Island's ecological and cultural change. This proposal should be considered an integrative hypothesis to be tested with future studies from the different disciplines involved. The overall framework for this integrative hypothesis, tentatively called CLAFS (Climate-Landscape-Anthropogenic Feedbacks and Synergies), is provided hereafter.
The MCA drought (880–1,170 CE) roughly coincided with the time range proposed for initial human settlement. If settlement was early (i.e., between 800 and 1,000 CE), then significant deforestation began 200 to 400 years after Polynesian arrival. This may suggest that resource exploitation had a low impact on vegetation until 1,200 CE when land-use practices became unsustainable (Orliac, 2000; Mann et al., 2008; Mieth and Bork, 2010). If, on the contrary, Polynesian settlement took place later, between 1,100 and 1,300 CE, deforestation practices would have started almost immediately in Rano Raraku. The humid phase between the MCA and the LIA (1,200–1,550 CE) coincided with the ahu moai practices characteristic of the AC and the florescence of the ancient Rapanui society. It is possible that favorable weather conditions promoted human population growth thus increasing the demand for food and raw materials, which would have led to the above-mentioned unsustainable land-use practices and exacerbated deforestation likely due to positive feedbacks.
The transition from Ancient Cult to Birdman Cult occurred shortly after the onset of the LIA drought (1,570–1,720 CE), suggesting a novel hypothesis for the cultural change and the corresponding geographical displacement of the Rapanui cultural core (Rull, 2016a). During this drought, Rano Raraku was a very inhospitable place devoid of forests and freshwater due to the synergistic action of climatic drought and unsustainable human practices. As a consequence, lifestyles and cultural developments characteristic of the ancient Rapanui society that practiced the AC would have become unworkable. The Rapanuis might have been forced to abandon the catchment in the search for freshwater and other natural resources. It is likely that part of the Rapanui population migrated to Rano Aroi by 1,500–1,600 CE, where forests and freshwater were still available, but the small size of this catchment would have led to quick resource exhaustion after 1,620 CE. The other option for relocation was Rano Kao, which contained a large and permanent lake that could provide freshwater and possibly more extensive forests to exploit. According to the available chronological evidence, Orongo, the center of the BC, was founded by 1,600 CE (Robinson and Stevenson, 2017), shortly after the beginning of the LIA drought, which is consistent with the idea that the Rapanui migrated in search of more favorable living conditions. It is also possible that the environmental deterioration of Rano Raraku contributed to an eventual superiority of western clans (Figure 3) due to their permanent freshwater supply from Lake Kao.
The walls of the Rano Kao vent are made of basalt, which is too hard for moai carving with the rocks available on the island (the Rapanui was a Neolithic society that did not know metals). Indeed, stone tools used during the ahu moai phase of statue carving were obtained in Rano Kao and other quarries, where basalt was hard enough to sculpt the softer Raraku tuff (Gioncada et al., 2010; Simpson and Dussubieux, 2018). This could explain, in part, why the relocation of the Rapanui cultural center to Rano Kao coincided with the decline of moai carving and the shift to different ritual practices. The shift to a radically different lifestyle and social organization requires a more complex explanation. As we mentioned before, the flourishing of the Rapanui civilization occurred during a phase of wet climate and the oncoming LIA drought, along with the deforestation of the craters containing freshwater, would have been unfavorable for population growth. In this context, it has been speculated that a more flexible socio-political organization would have increased human adaptability to less predictable environmental and probably unprecedented conditions. The shift from the AC rigid, hierarchical and dynasty-based political system to the more dynamic BC system, based on the continuous political renovation has been interpreted in terms of cultural adaptation to more unpredictable environmental conditions (Rull, 2016a).
Some archeological evidence for intra-island cultivation shifts is consistent with this CLAFS view. Stevenson et al. (2015) suggest that the AC-BC cultural transition was associated with reductions in land-use intensity in the drier sectors of the island and in infertile soils from upland areas, which increased land pressure in other parts of the island. Similar intra-island migrations, notably from coastal to inland sites, have been reported for many other Pacific islands by the same LIA times (Nunn, 2003, 2007; Nunn et al., 2007). From a paleoecological perspective, a continuous and coherent Kao record similar to those of Raraku and Aroi is essential to resolve deforestation trends and patterns over time and across space, in relation to cultural developments.
Lake Kao and the New Record
The Lake and Its Sediments
Rano Kao is the largest freshwater body of Easter Island with about 1,250 m diameter and >10 m of maximum water depth (Figure 7). A notable feature of this lake is the presence of a discontinuous floating mat typically up to ~3 m depth that forms a mosaic with open water spots all around the lake surface. The floating mat is dominated by Scirpus californicus (Cyperaceae) and Polygonum acuminatum (Polygonaceae) a common association in the island's freshwater bodies (Zizka, 1991). According to the available information, this floating mat formed during the last 900–1,000 years by progressive accumulation of plant remains; therefore, during roughly the last millennium, sediments (and therefore proxies for ecological change) have been accumulating on both the floating mat (developing a peat-like deposit) and at the lake bottom (as lacustrine sediments) (Rull, 2016b). It has been suggested that these two archives have been partially mixed by the eventual sinking of mat fragments to the lake bottom, causing the observed chronological anomalies in the Kao sedimentary sequence (Butler et al., 2004). This idea is supported by the morphological patterns of the floating mat mosaic, where the mostly sharp and straight edges of vegetation patches (Figure 7) suggest disaggregation and sinking of mat fragments. Therefore, it seems reasonable to assume that best likelihood for a continuous and coherent paleoecological record of the last millennium would be found within the floating mat, rather than in the lake-bottom sediments.
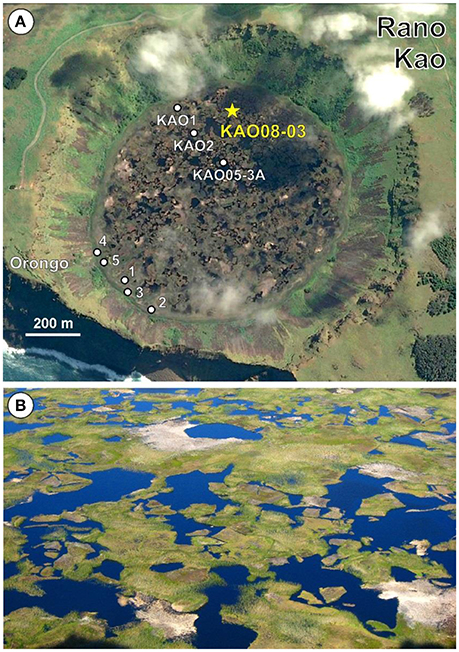
Figure 7. Rano Kao. (A) Google Earth view of Rano Kao indicating the coring sites from previous works (white dots) and the locality described in this paper (yellow star). The ceremonial village of Orongo is also indicated. Redrawn from Rull (2016b). (B) Close up of the floating mats around the center of Lake Kao. Note the sharp and angular nature of most mat/water contacts (Photo: O. Margalef).
Eight coring localities within the lake basin have been documented: six near the lake shores, one close to the lake center, and another in an intermediate position (Figure 7). The more marginal sites have provided composite sequences where the floating mat overlies the bottom sediments, whereas in more central localities, the floating mat and the bottom sediment records are separated by a water column of variable depth, up to ~10 m. The longest sequences retrieved from the lake are of ~10 to 20 m depth, and have maximum ages between ~15 and ~34 cal kyr BP, respectively (Gossen, 2011; Horrocks et al., 2013). These cores did not reach the bedrock and the total depth of Kao sediments remains unknown.
The KAO08-03 Core Record
The new sedimentary record reported in this paper (KAO08-03; 27° 10′ 57.526″ S - 109° 26′ 7.591″ W; 109 m elevation) was taken in October 2008 with a Russian corer, on a floating mat close to the northernmost lake shore (Figure 8). The uppermost part of the core was water and the sedimentary record started at 80 cm, extending downward until 300 cm, which was the depth of the floating mat at the coring point. The sediment consisted of fibrous peat, light to dark brown in color. The core (consisting of five drives) was sealed and transported to the Institute of Earth Science Jaume Almera (ICTJA, Spain), where it was preserved in a cool chamber at 4°C.

Figure 8. Coring the Rano Kao floating mat. (A) Two members of the 2008 expedition (V. Rull and S. Pla) coring the floating mat of Rano Kao with a Russian corer at site KAO08-03. (B) Drive 2 (131-181 cm) of core KAO08-03 showing the fibrous brown peat.
Nine samples were obtained for radiocarbon dating, which was conducted at the Radiocarbon Laboratory of the Université Laval (Canada). We used pollen extracts obtained by acid/base digestion (KOH, HCl and HF) to remove the rejuvenating effect of eventual incorporation of younger plant material and/or humic acids percolating through the peat profile (Vandergoes, 2003). All samples yielded ages corresponding to the last millennium except one (Table 1), which was considered anomalous and was excluded from the age-depth model. This type of anomalies have been attributed to an increase of the reservoir effect due to (i) punctual increases of aquatic plants, (ii) reductions in CO2 exchange with the atmosphere due to increase of depth-to-surface ratio, (iii) incorporation of older oxidized organic matter from former shore-lake lacustrine deposits, and (iv) inflows of CO2-rich waters due to volcanic degassing or eventual inputs from nearby aquifers (Philippsen, 2013). In our case, the more probable causes were (i) and (iii) but more detailed studies are needed for a sound assessment. Radiocarbon calibration was carried out with CALIB 7.1 (calib.org/calib) using the SHcal13 database for the Southern Hemisphere (Hogg et al., 2013). Age-depth modeling was done using the smooth-spline option (smoothness 0.4) in the Clam.R 2.2 software (Blaauw, 2010). The model obtained shows strong chronological coherence and continuity, indicating that the record can provide a detailed and continuous paleoecological reconstruction for the last millennium, and can provide comparison with the main cultural and ecological events discussed above (Figure 9). We also tried the Bayesian age-depth model called Bacon (using R programming) for comparison (Blaauw and Christen, 2011). Results are shown in Figures S1, S2 of the Supplementary Material, where it can be realized that differences in estimated ages are small and non-significant.
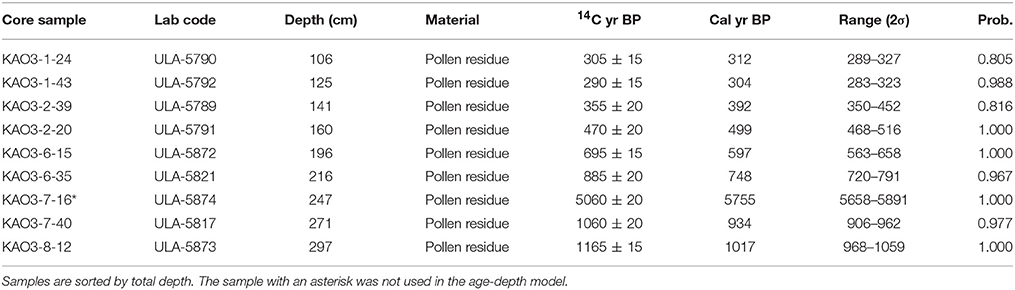
Table 1. Results of radiocarbon dating carried out at the Radiochronology Lab of the Université Laval (Canada).
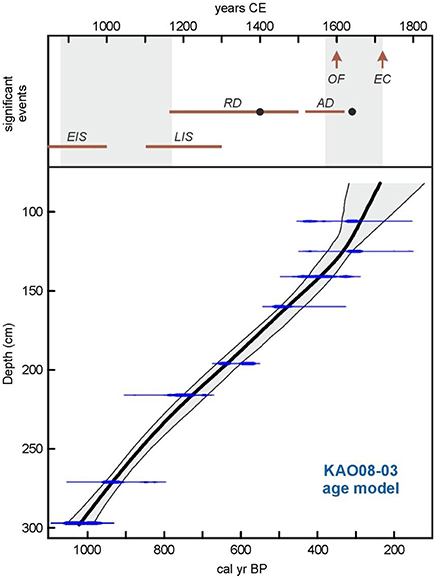
Figure 9. Age-depth model of core KAO08-03 using the smooth-spline option. See text and Table 1 for more details. The upper part of the figure shows some significant events discussed in the text. Gray bands are the droughts recorded at Rano Raraku (Cañellas-Boltà et al., 2013). EIS, Proposed range for early island settlement (Flenley and Bahn, 2003; Vargas et al., 2006); LIS, proposed range for late island settlement (Hunt and Lipo, 2006; Wilmshurst et al., 2011); RD, Rano Raraku deforestation (Cañellas-Boltà et al., 2013), the black dot indicates the first evidence of cultivation (Horrocks et al., 2012a); AD, Rano Aroi deforestation (Rull et al., 2015), the black dot indicates the first evidence of cultivation) (Horrocks et al., 2015); OF, Foundation of the ceremonial village of Orongo (Robinson and Stevenson, 2017); EC, European contact.
A total of 107 samples were collected from the core, 55 for palynological analysis and 52 for lipid analysis, providing an average resolution of <20 years between adjacent samples. Palynological analysis is in progress at the Laboratory of Paleoecology of the (ICTJA), and consists of analysis of pollen, spores, microcharcoal and non-pollen palynomorphs (NPP), notably the spores of coprophilous fungi. These proxies will provide information on deforestation, fires, and the eventual presence of introduced plants and domestic animals (Bennett and Willis, 2001; Van Geel, 2001; Whitlock and Larsen, 2001). These samples will also be used to conduct macrocharcoal analyses for a better record of local fires and also to try to distinguish fuel types (Aleman et al., 2013), notably palms and grasses. Lipid analysis is being carried out at the Lamont-Doherty Earth Observatory (LDEO) of Columbia University (USA). This analysis is aimed at identifying specific fecal lipids, including 5α-stanols and bile acids, biomarkers for the presence of humans and/or ruminant livestock (Leeming et al., 1996; Bull et al., 2002; D'Anjou et al., 2012). Together, this suite of analyses is expected to provide direct in situ evidence for the level of human activity in and around the catchment.
Some Testable Predictions
Hypothesis tests that can be conducted with the new Kao record concern the timing of human occupation and deforestation of the Kao catchment, whether these events were simultaneous or not, and the rates at which they occurred. Comparison of the new Kao records with those from Raraku and Aroi could provide the pan-island, continuous, and coherent paleoecological reconstruction of the last millennium. Regardless the results obtained, further archeological evidence will be necessary to examine their cultural implications. Among the predictions of the CLAFS scenario, the following could be highlighted:
The CLAFS hypothesis predicts that evidence for full human occupation of the Kao catchment should be found close to the date of the foundation of Orongo (~1,600 CE). An earlier occupation cannot be dismissed but, under the CLAFS hypothesis, this would have been minimal with a significant increase ca. 1,600 CE. If the Kao catchment was fully occupied before the LIA drought (1,570–1,720 CE) and there is no evidence supporting a population increase at Kao by 1,600 CE, the CLAFS hypothesis should be reconsidered in favor of alternative hypotheses, such as the ecocide hypothesis. If a population increase is found to have occurred later than 1,600 CE, the migration to Kao, and possibly the onset of the BC, could conceivably have been influenced by European contact (1,722 CE), as has been suggested by some researchers (Robinson and Stevenson, 2017).
Regarding deforestation, the CLAFS scenario is compatible only with spatio-temporal heterogeneities; that is, with different deforestation timing among Aroi, Raraku and Kao. Evidence for this already exists for the first two sites, for which the onset of deforestation appears to have been separated by several centuries (Figure 6). The CLAFS hypothesis predicts that deforestation of the Kao catchment coincided with the intensification of human activity at the site ca. 1,600 CE, similar to the timing of forest clearing at Aroi. Again, an earlier deforestation, coeval with that of Raraku, would instead favor ecocide hypothesis, whereas a later forest clearing, after the LIA drought, would support the influence of post-contact events. Under the CLAFS scenario, it is expected that deforestation and fire occurrence, as inferred from sedimentary charcoal counts, are strongly related, as has been observed for Raraku and Aroi (Figure 6). However, while the coincidence of proxies for fire, deforestation, and human presence support anthropogenic forest clearing practices, such a co-occurrence cannot distinguish between the CLAFS and ecocide hypotheses.
It is important to note that there are no unique predictions under the CLAFS hypothesis concerning rates of Kao deforestation and human settlement. Possibilities include (1) fast and synchronous events, as inferred for Aroi (Figure 6), (2) early settlement and gradual deforestation, as inferred for Raraku, and (3) gradual settlement and abrupt deforestation.
Alternative Scenarios
There also remains the possibility of obtaining unexpected results. Some of these eventual findings could support the CLAFS hypothesis while others do not, in which case the working hypothesis should be reconsidered or reformulated to become more general. We are not particularly attached to any specific hypothesis, but rather in the explanation that best fits the entire body of available evidence. We use the multiple working hypotheses approach that seeks to explore any reasonable explanation for observed phenomena and develop every testable hypothesis (Chamberlin, 1965). In this framework, the CLAFS model should be viewed as a research tool to be subjected to scrutiny through hypothesis testing, and improvement or rejection, just as any other hypothesis. It is possible that paleoecological analysis of the new Kao record will suggest a different framework for understanding the complexities of the Rapanui, thereby promoting new unexpected ideas, in which case, the results will inspire new opportunities for a better understanding of Easter Island's prehistory.
Contribution to Global Databases
The results obtained in this and future research on core KAO08-03 will be contributed to the EIRA database (https://www.ncdc.noaa.gov/paleo-search/study/19805) (Rull, 2016b) in the case of radiocarbon dates, Global Charcoal Database (http://paleofire.org) for charcoal data and Neotoma (http://neotomadb.org) for pollen counts.
Author Contributions
VR: Conceived and wrote the paper; EM: Did the age-depth model; IS and WD: Processed and analyzed samples; VR, NC-B, OM, SP-R, and SG: Obtained the core. All authors contributed to the manuscript with ideas, comments, edits and references, and approved submission.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was funded by the Spanish Ministry of Science and Education (project CGL2007-60932/BTE) and the Catalan Agency for Management of University and Research Grants (grant 2014 SGR 1207).
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2018.00032/full#supplementary-material
References
Aleman, J. C., Blarquez, O., Bentaleb, I., Bonté, P., Brossier, B., Carcaillet, C., et al. (2013). Tracking land-cover changes with sedimentary charcoal in the Afrotropics. Holocene 23, 1853–1862. doi: 10.1177/0959683613508159
Bennett, K. D., and Willis, K. J. (2001). “Pollen,” in Tracking Environmental Change Using Lake Sediments, Terrestrial, Algal and Siliceous Indicators, Vol. 3, eds J. P. Smol, H. J. B. Birks, and W. M. Last (Dordrecht: Kluwer), 5–32.
Blaauw, M. (2010). Methods and code for classical age-modelling of radiocarbon sequences. Quat. Geochronol. 5, 512–518. doi: 10.1016/j.quageo.2010.01.002
Blaauw, M., and Christen, A. J. (2011). Flexible paleoclimate age-depth models using autoregressive gamma process. Bayesian Anal. 6, 457–474. doi: 10.1214/11-BA618
Boersema, J. J. (2015). The Survival of Easter Island: Dwindling Resources and Cultural Resilience. Cambridge: Cambridge University.
Bond, W. J., and Keane, R. E. (2017). Ecological Effects of Fire. US Forest Service Reference Module in Life Sciences. Fort Collins: Rocky Mountain Research Station. doi: 10.1016/B978-0-12-809633-8.02098-7
Brandt, G., and Merico, A. (2015). The slow demise of Easter Island: insights from a modelling investigation. Front. Ecol. Evol. 3:13. doi: 10.3389/fevo.2015.00013
Bull, I. D., Lockheart, M. J., Elhmmali, M. M., Roberts, D. J., and Evershed, R. P. (2002). The origin of faeces by means of biomarker detection. Environ. Int. 27, 647–654. doi: 10.1016/S0160-4120(01)00124-6
Butler, K. R., Prior, C. A., and Flenley, J. R. (2004). Anomalous radiocarbon dates from Easter Island. Radiocarbon 46, 395–405. doi: 10.1017/S0033822200039709
Butler, K., and Flenley, J. R. (2010). The Rano Kau 2 pollen diagram: paleoecology revealed. Rapa Nui J. 24, 5–10.
Cañellas-Boltà, N., Rull, V., Sáez, A., Margalef, O., Bao, R., Pla-Rabes, S., et al. (2013). Vegetation changes and human settlement of Easter Island during the last millennia: a multiproxy study of the lake Raraku sediments. Quat. Sci. Rev. 72, 36–48. doi: 10.1016/j.quascirev.2013.04.004
Chamberlin, T. C. (1965). The method of multiple working hypotheses. Science 148, 754–759. doi: 10.1126/science.148.3671.754
Cortez, C., Zárate, E., and González-Ferrán, O. (2009). Isla de Pascua: Impactos y evidencias del Tsunami del 22 de mayo de 1960. XII Cong. Geol. Chileno S2, 1–4.
D'Anjou, R. M., Bradley, R. S., Balascio, N. L., and Finkelstein, D. B. (2012). Climate impacts on human settlement and agricultural activities in northern Norway revealed through sediment biogeochemistry. Proc. Natl. Acad. Sci. U.S.A. 109, 20332–20337. doi: 10.1073/pnas.1212730109
Dransfield, J., Flenley, J. R., King, S. M., Harkness, D. D., and Rapu, S. (1994). A recently extinct palm of Easter Island. Nature 312, 350–352.
Dumont, H. J., Cocquyt, C., Fontugne, M., Arnold, M., Reyss, J.-L., Bloemendal, J., et al. (1998). The end of moai quarrying and its effects on Lake Rano Raraku, Easter Island. J. Paleolimnol. 20, 409–422. doi: 10.1023/A:1008012720960
Edwards, E., and Edwards, A. (2013). When the Universe was an Island. Exploring the Cultural and Spiritual Cosmos of the Ancient Rapa Nui. Hangaroa: Hangaroa Press.
Ferdon, E. N. (1961). “The ceremonial site of Orongo,” in Archaeology of Easter Island, Volume 1: Reports of the Norwegian Archaeological Expedition to Easter Island and the East Pacific, eds T. Heyerdahl and E. N. Ferdon (London: Allen & Unwin), 221–255.
Fischer, S. R. (2005). Island at the End of the World. The Turbulent History of Easter Island. London: Reaction Books.
Flenley, J. R., and Bahn, P. G. (2003). The Enigmas of Easter Island. Oxford: Oxford University Press.
Flenley, J. R., and King, S. M. (1984). Late quaternary pollen records from Easter Island. Nature 307, 47–50. doi: 10.1038/307047a0
Flenley, J. R., King, S. M., Jackson, J., Chew, C., Teller, J. T., and Prentice, M. E. (1991). The late quaternary vegetation and climatic history of Easter Island. J. Quat. Sci. 6, 85–115. doi: 10.1002/jqs.3390060202
Gioncada, A., González-Ferran, O., Lezzerini, M., Mazzuoli, R., Bisson, M., and Rapu, S. A. (2010). The volcanic rocks of Easter Island (Chile) and their use for the moai sculptures. Eur. J. Mineral. 22, 855–867. doi: 10.1127/0935-1221/2010/0022-2057
Golson, J. (1965). Thor Heyerdahl and the prehistory of Easter Island. Oceania 36, 38–83. doi: 10.1002/j.1834-4461.1965.tb00277.x
González-Ferrán, O., Mazzuoli, R., and Lahsen, A. (2004). Geologia del complejo volcanico Isla de Pascua Rapa Nui, Chile. (1:30.000). Santiago: Centro de Estudios Volcanologicos de Santiago.
Gossen, C. (2011). Deforestation, Droughtand Humans: New Discoveries of the Late Quaternary Paleoenvironment of Rapa Nui (Easter Island). Ph. D. Dissertation, Portland State University, Portland.
Herrera, C., and Custodio, E. (2008). Conceptual hydrogeological model of volcanic Easter Island (Chile) after chemical and isotopic surveys. Hydrogeol. J. 16, 1329–1348. doi: 10.1007/s10040-008-0316-z
Hogg, A. G., Hua, Q., Blackwell, P. G., Niu, M., Buck, C. E., Guilderson, T. P., et al. (2013). SHcal13 Southern Hemisphere calibration, 0-50,000 years cal BP. Radiocarbon 55, 1889–1903. doi: 10.2458/azu_js_rc.55.16783
Horrocks, M., Baisden, T., Flenley, J., Feek, D., Love, C., Haoa-Cardinali, S., et al. (2016). Pollen, phytolith and starch analyses of dryland soils from Easter Island (Rapa Nui) show widespread vegetation clearance and Polynesian-introduced crops. Palynology 41, 339–350. doi: 10.1080/01916122.2016.1204566
Horrocks, M., Baisden, W. T., Flenley, J., Feek, D., González-Nualart, L., Haoa- Cardinali, S., et al. (2012a). Fossil plant remains at Rano Raraku, Easter Island's statue quarry: evidence for past elevated lake level and ancient Polynesian agriculture. J. Paleolimnol. 46, 767–783. doi: 10.1007/s10933-012-9643-0
Horrocks, M., Baisden, W. T., Harper, M. A., Marra, M., Flenley, J., Feek, D., et al. (2015). A plant microfossil record of late quaternary environments and human activity from Rano Aroi and surroundings, Easter Island. J. Paleolimnol. 54, 279–303. doi: 10.1007/s10933-015-9852-4
Horrocks, M., Baisden, W. T., Nieuwoudt, W. T., Flenley, J., Feek, D., González-Nualart, L., et al. (2012b). Microfossils of Polynesian cultigen sin lake sediment cores from Rano Kau, Easter Island. J. Paleolimnol. 47, 185–204. doi: 10.1007/s10933-011-9570-5
Horrocks, M., Marra, M., Baisden, W. T., Flenley, J., Feek, D., González-Nualart, L., et al. (2013). Pollen, phytoliths, arthropods and high-resolution 14C sampling from Rano Kau, Easter Island: evidence for late quaternary environments, ant (formicidae) distributions and human activity. J. Paleolimnol. 50, 417–432. doi: 10.1007/s10933-013-9735-5
Hunt, T. L. (2006). Rethinking the fall of Easter Island: new evidence points to an alternative explanation for a civilization's collapse. Am. Sci. 94, 412–419. doi: 10.1511/2006.61.1002
Hunt, T. L. (2007). Rethinking Easter Island's ecological catastrophe. J. Archaeol. Sci. 34, 485–502. doi: 10.1016/j.jas.2006.10.003
Hunt, T. L., and Lipo, C. P. (2006). Late colonization of Easter Island. Science 311, 1603–1606. doi: 10.1126/science.1121879
Jarman, C. L., Larsen, T., Hunt, T., Lipo, C., Solsvik, R., Wallsgrove, N., et al. (2017). Diet of the prehistoric population of Rapa Nui (Easter Island) shows environmental adpatation and resilience. Am. J. Phys. Anthropol. 164, 343–361. doi: 10.1002/ajpa.23273
Kirch, P. V. (2000). On the Road of the Winds. An Archaeological History of the Pacific Islands before European Contact. Berkeley: University of California Press.
Leeming, R., Ball, A., Ashbolt, N., and Nichols, P. (1996). Using faecal sterols from humans and animals to distinguish faecal pollution in receiving wàters. Water Res. 30, 2893–2900. doi: 10.1016/S0043-1354(96)00011-5
Mann, D., Edwards, J., Chase, J., Beck, W., Reanier, R., Mass, M., et al. (2008). Drought, vegetation change, and human history on RapaNui (Isla de Pascua, Easter Island). Quat. Res. 69, 16–28. doi: 10.1016/j.yqres.2007.10.009
Margalef, O., Álvarez-Gómez, J. A., Pla-Rabes, S., Ca-ellas-Bolt,à, N., Rull, V., Sáez, A., et al. (in press). Revisiting the role of high-energy Pacific events on the environmental cultural history of Easter Island (Rapa Nui). Geogr. J. doi: 10.1111/geoj.12253
Margalef, O., Cañellas-Boltà, N., Pla-Rabes, S., Giralt, S., Pueyo, J. J., Joosten, H., et al. (2013). A 70,000 year multiproxy record of climatic and environmental change from Rano Aroi peatland (Easter Island). Glob. Planet. Change 108, 72–84. doi: 10.1016/j.gloplacha.2013.05.016
McCall, G. (1980). Rapanui. Tradition and Survival on Easter Island. Honolulu: University Press of Hawaii.
Métraux, A. (1940). Ethnology of Easter Island. Honolulu: Bernice P. Bishop Museum Bulletin, 160 (reprinted 1971).
Mieth, A., and Bork, H. (2010). Humans, climate or introduced rats - which is to blame for the woodland destruction on prehistoric Rapa Nui (Easter Island)? J. Archaeol. Sci. 37, 417–426. doi: 10.1016/j.jas.2009.10.006
Mulrooney, M. (2013). An island-wide assessment of the chronology of settlement and land use on RapaNui (Easter Island) based on radiocarbon data. J. Archaeol. Sci. 40, 4377–4399. doi: 10.1016/j.jas.2013.06.020
Nunn, P. D. (2003). Nature-society interactions in the Pacific Islands. Geogr. Ann. 85, 219–229. doi: 10.1111/j.0435-3684.2003.00144.x
Nunn, P. D. (2007). Climate, Environment and Society in the Pacific During the Last Millennium. Amsterdam: Elsevier.
Nunn, P. D., Hunter-Anderson, R., Carson, M. T., Thomas, F., Ulm, S., and Rowland, M. J. (2007). Times of plenty, times of less: last-millennium societal disruption in the Pacific Basin. Hum. Ecol. 35, 385–401. doi: 10.1007/s10745-006-9090-5
Orliac, C. (2000). “The woody vegetation of Easter Island between the early 14th and the mid-17th centuries AD,” in Research on Early Rapanui Culture, eds C. M. Stevenson and W. S. Ayres (Los Osos: Easter Island Foundation), 211–220.
Orliac, C., and Orliac, M. (1996). Arbres et arbrustes de l'Ile de Paques: Composition et evolution de la flore depuis l'arrivee des Polynesiens. Rapport intermediaire. Compte-Rendu de la Mission Archeologique a l'Ile de Paques. Ministere des Affaires Etrangeres/CNRS/Consejo de Monumentos/CONAF.
Philippsen, B. (2013). The freshwater reservoir effect in radiocarbon dating. Heritage Sci. 1, 24. doi: 10.1186/2050-7445-1-24
Puleston, C. O., Ladefoged, T. N., Haoa, S., Chadwick, O. A., Vitousek, P. M., and Stevenson, C. M. (2017). Rain, sun, soil, and sweat: a consideration of population limits on Rapa Nui (Easter Island) before European Contact. Front. Ecol. Evol. 5:69. doi: 10.3389/fevo.2017.00069
Robinson, T., and Stevenson, C. M. (2017). The cult of the Birdman: religious change at 'Orongo, Rapa Nui (Easter Island). J. Pacific Archaeol. 8, 88–102.
Rull, V. (2016a). Natural and anthropogenic drivers of cultural change on Easter Island: review and new insights. Quat. Sci. Rev. 150, 31–41. doi: 10.1016/j.quascirev.2016.08.015
Rull, V. (2016b). The EIRA database: last glacial and holocene radiocarbon ages from Easter Island's sedimentary records. Front. Ecol. Evol. 4:44. doi: 10.3389/fevo.2016.00044
Rull, V., Cañellas-Boltà, N., Margalef, O., Pla-Rabes, S., Sáez, A., and Giralt, S. (2016). Three millennia of climatic, ecological and cultural change on Easter Island: an integrative overview. Front. Ecol. Evol. 4:29. doi: 10.3389/fevo.2016.00029
Rull, V., Cañellas-Boltà, N., Margalef, O., Sáez, A., Pla-Rabes, S., and Giralt, S. (2015). Late Holocene vegetation dynamics and deforestation in Rano Aroi: implications for Easter Island's ecological and cultural history. Quat. Sci. Rev. 126, 219–226. doi: 10.1016/j.quascirev.2015.09.008
Rull, V., Ca-ellas-Bolt,à, N., Sáez, A., Giralt, S., Pla, S., and Margalef, O. (2010). Paleoecology of Easter Island: evidence and uncertainties. Earth-Sci. Rev. 99, 50–60. doi: 10.1016/j.earscirev.2010.02.003
Rull, V., Ca-ellas-Bolt,à, N., Sáez, A., Margalef, O., Bao, R., Pla-Rabes, S., et al. (2013). Challenging Easter Island's collapse: the need for interdisciplinary synergies. Front. Ecol. Evol. 2:3. doi: 10.3389/fevo.2013.00003
Sáez, A., Valero-Garcés, B., Giralt, S., Moreno, A., Bao, R., Pueyo, J. J., et al. (2009). Glacial to Holocene climate changes in the SE Pacific. the Raraku Lake sedimentary record (Easter Island, 27?S). Quat. Sci. Rev. 28, 2743–2759. doi: 10.1016/j.quascirev.2009.06.018
Simpson, D. F., and Dussubieux, L. (2018). A collapsed narrative? geochemistry and spatial distribution of basalt quarries and fine-grained artifacts reveal communal use of sstone on Rapa Nui (Easter Island). J. Archaeol. Sci. Rep. 18, 370–385. doi: 10.1016/j.jasrep.2018.01.038
Stevenson, C. M., and Haoa-Cardinali, S. (2008). Prehistoric Rapa Nui: Landscape and Settlement Archaeology at Hanga Ho'onu. Los osos: Easter Island Foundation.
Stevenson, C. M., Ladefoged, T. N., and Novak, S. (2013). Prehistoric settlement on Rapa Nui, Chile: an evaluation of obsidian hydration dating using infrared photoacoustic spectroscopy. J. Archaeol. Sci. 40, 3021–3030. doi: 10.1016/j.jas.2013.03.019
Stevenson, C. M., Puleston, C. O., Vitousek, P. M., Chadwick, O. A., Haoa-Cardinali, S., and Ladefoged, T. N. (2015). Variation in Rapa Nui (Easter Island) land use indicates production and population peaks prior to European contact. Proc. Natl. Acad. Sci. U.S.A. 112, 1025–1030. doi: 10.1073/pnas.1420712112
Thorsby, E. (2016). Genetic evidence for a contribution of native Americans to the early settlement of Rapa Nui (Easter Island). Front. Ecol. Evol. 4:118. doi: 10.3389/fevo.2016.00118
Van Geel, B. (2001). “Non-pollen palynomorphs,” in Tracking Environmental Change using Lake Sediments. Terrestrial, Algal and Siliceous Indicators, eds J. P. Smol, H. J. B. Birks, and W. M. Last (Dordrecht: Kluwer), Vol. 3, 99–119.
Van Tilburg, J. A. (1996). “Easter Island (Rapa Nui) archaeology since 1955: some thoughts on progress, problems and potential,” in Oceanic Culture History: Essays in Honour of Roger Green, eds J. M. Davidson, G. Irwin, B. F. Leach, A. Pawley, and D. Brown (Dunedin: N. Zealand J. Archaeol. Spec. Publ.), 555–577.
Vandergoes, M. J. (2003). AMS dating of pollen concentrates-a methodological study of Late Quaternary sediments from South Westland, New Zealand. Radiocarbon 45, 479–491. doi: 10.1017/S0033822200032823
Vargas, P., Cristino, C., and Izaurieta, R. (2006). 1000 a-os en Rapa Nui. Arqueología delasentamiento. Santiago: Editorial Universitaria, Universidad de Chile.
Vezzolli, L., and Acocella, V. (2009). Easter Island, SE Pacific: an end-member type of hotspot volcanism. Geol. Soc. Am. Bull. 121, 869–886. doi: 10.1130/B26470.1
West, K., Collins, C., Kardailsky, O., Kahn, J., Hunt, T. L., Burley, D. V., et al. (2017). The Pacific rat race to Easter Island: tracking the prehistoric dispersal of rattus exulansusing ancient mitochondrial genomes. Front. Ecol. Evol. 5:52. doi: 10.3389/fevo.2017.00052
Whitlock, C., and Larsen, C. (2001). “Charcoal as a fire proxy,” in Tracking Environmental Change using Lake Sediments. Terrestrial, Algal and Siliceous Indicators, Vol. 3, eds J. P. Smol, H. J. B. Birks, and W. M. Last (Dordrecht: Kluwer), 75–98.
Keywords: Easter Island, Rapa Nui, radiocarbon dating, age-depth model, last millennium, deforestation, cultural change
Citation: Rull V, Montoya E, Seco I, Cañellas-Boltà N, Giralt S, Margalef O, Pla-Rabes S, D'Andrea W, Bradley R and Sáez A (2018) CLAFS, a Holistic Climatic-Ecological-Anthropogenic Hypothesis on Easter Island's Deforestation and Cultural Change: Proposals and Testing Prospects. Front. Ecol. Evol. 6:32. doi: 10.3389/fevo.2018.00032
Received: 07 February 2018; Accepted: 19 March 2018;
Published: 04 April 2018.
Edited by:
Joel Guiot, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Olivier Blarquez, Université de Montréal, CanadaMark Bush, Florida Institute of Technology, United States
Copyright © 2018 Rull, Montoya, Seco, Cañellas-Boltà, Giralt, Margalef, Pla-Rabes, D'Andrea, Bradley and Sáez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Valentí Rull, dnJ1bGxAaWN0amEuY3NpYy5lcw==