 Leandro Schlemmer Brasil1,2*
Leandro Schlemmer Brasil1,2* Victor Rennan Santos Ferreira1,2
Victor Rennan Santos Ferreira1,2 Bethânia Oliveira de Resende1,2Leandro Juen1,2
Bethânia Oliveira de Resende1,2Leandro Juen1,2 Joana Darc Batista3
Joana Darc Batista3 Lourivaldo Amâncio de Castro3
Lourivaldo Amâncio de Castro3 Nubia França da Silva Giehl3,4
Nubia França da Silva Giehl3,4- 1Laboratório de Ecologia e Conservação, Instituto de Ciências Biológicas, Universidade Federal do Pará, Belém, Brazil
- 2Programa de Pós-Graduação em Ecologia, Universidade Federal do Pará, Belém, Brazil
- 3Laboratório de Entomologia, Universidade do Estado de Mato Grosso, Nova Xavantina, Brazil
- 4Laboratório de Ecologia de Bentos, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
The veredas are wetland ecosystems responsible for supplying most of the water for rivers and streams in the Cerrado. The veredas’ hydromorfic soils retain a large amount of rainwater, releasing it slowly during drier periods. Therefore, these habitats are often used to build dams for cattle raising. Here we assessed the environmental conditions and beta-diversity of Odonata and Heteroptera on veredas impacted by dams in the Brazilian Cerrado. We sampled biological communities and a set of environmental variables in 13 veredas, six with dams and seven without dams. One limnological variable [oxidation–reduction potential (ORP)] and one landscape metric (% of the altered area) differed among veredas with and without dams. These variables were important predictors of the beta-diversity of both Odonata (R2 = 0.650; p < 0.001) and Heteroptera (R2 = 0.740; p < 0.001). The veredas stand among the most sensitive wetland ecosystems of the Cerrado. In this study, we show, for the first time, that veredas with dams may lose environmental quality resulting in changes in biological communities, especially ecologically unique species. Because the Cerrado naturally goes through approximately 5 months of severe drought, the veredas are critical for the Cerrado’s hydric safety. Therefore, we recommend that cattle-raising activities should rely on artificial water tanks instead of using dams for water storage since it affects ecologically unique species in this poorly known ecosystem.
Introduction
Aquatic ecosystems are strongly affected by the land-use change (Allan and Castillo, 2007). Cattle-raising activities are directly or indirectly responsible for the highest deforestation rates in Brazil (Alencar et al., 2020; Pereira et al., 2020), being considered one of the main sources of anthropogenic impact in aquatic ecosystems. The integration of aquatic and terrestrial environments occurs because rivers, lakes, and streams are affected by rainwater coming from water catchment areas in the landscape, which compose the river basin (Allan and Castillo, 2007). Among the main negative impacts of cattle raising on aquatic ecosystems, we can highlight the increased entry of inorganic sediments, causing siltation (Casatti et al., 2009), the trampling of microhabitats, which happens when cattle drink water from streams, and dam construction to store water for cattle (Brasil et al., 2014a). Dam construction converts natural lotic environments into lentic or semi-lentic ones, seriously threatening the continuous transfer of aloctone and autoctone energy (Vannote et al., 1980) and partially interrupting the migration of organisms (Brasil et al., 2014a) and energy flow. Therefore, the high number of small dams and ponds in areas used for cattle raising changes the biotic diversity and the environmental conditions of aquatic systems (De Marco et al., 2014).
Among tropical ecosystems, the Brazilian Cerrado (i.e., Brazilian savanna) harbors a great diversity of vegetation formations, ranging from forests to grasslands (Ribeiro and Walter, 2008). Within these formations, the “veredas” are characterized by an aggregation of palm trees (Mauritia flexuosa) under a complex set of water flows surrounded by herbaceous, grassy native savanna (Ribeiro and Walter, 2008). Despite this apparent low structural complexity when compared to forest formations, the “veredas” are of great hydrological importance because its many water table outcrops directly contribute to the formation of river and stream springs (Meirelles et al., 2006). Additionally, the “veredas” are important ecosystems for the maintenance of local biodiversity, functioning as refuge and habitat to several species endemic to the Cerrado (Tubelis, 2009; Vilela et al., 2016). These are open habitats with high light incidence when compared to other Cerrado streams, especially those draining forest formations, which result in important differences in the trophic composition of aquatic communities (Brasil et al., 2014b). Despite all that, the veredas’ environmental conditions and aquatic biodiversity have suffered intense anthropogenic pressures, especially from cattle-raising activities (Giehl et al., 2020).
Within the aquatic biodiversity, insects are known as bioindicators of environmental quality and providers of important ecosystem services by shredding organic matter, filtering sediments suspended in water, and even controlling populations of other organisms through predation (Merritt and Cummins, 1996). Among the predatory aquatic insects, the Odonata (dragonflies and damselflies) and Heteroptera (true bugs) orders stand out as sensitive to habitat change (Dias-Silva et al., 2010; Miguel et al., 2017; Brasil et al., 2020a). Even though we know that both Odonata and Heteroptera communities are affected by pond and dam construction (Brasil et al., 2014a; De Marco et al., 2014; Klein et al., 2018), there is no knowledge of how their communities respond to these environmental changes in veredas (Vilela et al., 2016; Giehl et al., 2020). Biological diversity can be measured in different ways, divided into three main components: alpha (α), beta (β), and gamma (γ). Since this division was originally proposed, several advances in measuring biodiversity have been suggested (Baselga, 2010). More recently, Legendre and De Cáceres (2013) proposed a decomposition of the total variation among communities [total beta diversity (BDTotal)] into local contribution to beta diversity (LCBD) and species contribution to beta diversity (SCBD). LCBD represents how ecologically singular communities are (Legendre and De Cáceres, 2013), whereas SCBD shows how much individual species contribute to variation in species composition among local communities (Legendre and De Cáceres, 2013; Heino and Grönroos, 2017).
Due to the limited knowledge of the impact of small dams on the veredas’ aquatic biota, we assessed the environmental conditions and beta diversity of Odonata and Heteroptera communities in veredas impacted by dams in the Brazilian Cerrado. We tested the hypothesis that beta diversity would be negatively affected by environmental changes caused by dam construction. Because both Odonata (Oliveira-Junior and Juen, 2019) and Heteroptera (Dias-Silva et al., 2020) orders harbor taxa sensitivity and taxa are tolerant to environmental change, we expect that dam construction will promote species turnover, where tolerant species will replace sensitive ones.
Materials and Methods
Study Area
We sampled 13 veredas of which six had associated dams and seven had no associated dams. Surveys were carried out in the July of 2015, which is the season with the lowest precipitation in the sampled region. The sampled veredas are located in the municipalities of Barra do Garças and Nova Xavantina, Brazil (Figure 1). The climate in these localities is Köppen’s Aw, that is, tropical savanna with a dry (May to September) and a rainy season (October to April) (Peel et al., 2007).
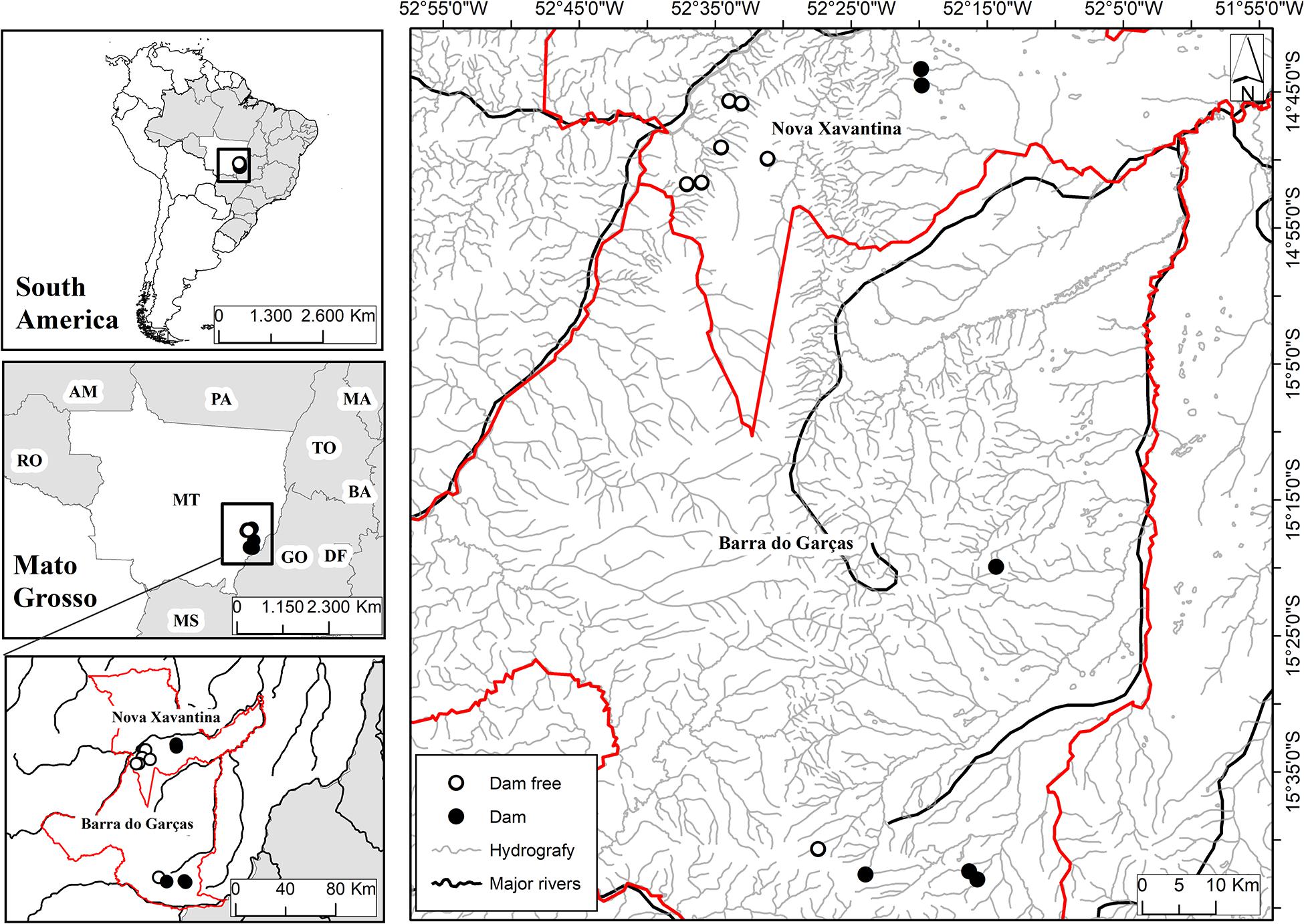
Figure 1. Spatial distribution of the sample points in veredas with dam and dam-free located at the municipalities of Nova Xavantina and Barra do Garças, MT, Brazil.
Description of the Veredas and Dams
The dam-free veredas predominantly consisted of grassy, herbaceous vegetation and clumps of M. flexuosa (buriti) across the channel (Figure 2A). The veredas with dams were inserted in cattle-raising areas, which is the most common land use type within the sampled micro basin (Figure 2B). Therefore, when present, the few M. flexuosa were older and sparser in veredas with dams. Dams were small with surface area varying from 200 to 2,750 m2 (602.30 ± 926.21) and used to store water for cattle. These dams were at least 7 years old.

Figure 2. Dam-free preserved vereda (A) and altered vereda with interrupted water flow and presence of cattle (B) located at the municipalities of Nova Xavantina and Barra do Garças, MT, Brazil.
Odonata and Heteroptera Collection and Identification
We sampled the Odonata adults using an entomological net, between 10:00 and 15:00 h, when the probability of high activity in all ecophysiological groups of both orders is the highest. The sampling method used was a scanning method in fixed areas, where each path delimits a linear transect of 100 m, subdivided into 20 segments of 5 m (Juen and De Marco, 2011; Cezário et al., 2020). The sampling effort was of approximately 1 h (De Marco and Resende, 2002). After being captured, the individuals were stored in entomological envelopes and transported to the laboratory for sorting, guarding, and identification using specialized bibliography (Lencioni, 2005, 2006; Garrison et al., 2006, 2010).
We sampled the Heteroptera at the same transects and time using a sieve with a circumference of 18 cm and a fine mesh for approximately 1 h on each vereda (see Dias-Silva et al., 2010). We stored the sampled individuals in flasks with 8.5% ethanol and took them to the laboratory for identification using specialized bibliography (e.g., Nieser and Melo, 1997; Rúdio and Moreira, 2011; Magalhães et al., 2016). When necessary, we compared the sampled individuals to reference material in the Coleção Zoobotânica “James Alexander Ratter,” Universidade do Estado de Mato Grosso, Nova Xavantina, Brazil, in which all sampled individuals were deposited. The individuals were collected under the Biodiversity Authorization and Information System license, SISBIO n° 45683-1.
Environmental Variables and Landscape Metrics
We measured 12 limnological environmental variables and landscape metrics that are considered important in assessing the level of environmental impact and in predicting Heteroptera and Odonada assemblages (Brasil et al., 2020b; Giehl et al., 2020): (1) air temperature, (2) nitrate, (3) ammonia, (4) turbidity, (5) water temperature, (6) dissolved oxygen (OD), (7) potential hydrogen (pH), (8) electric conductivity, (9) total dissolved solids (TDS), (10) oxidation–reduction potential (ORP), (11) luminosity, and (12) landscape change (Supplementary Table S1). To measure the chemical variables, we sampled 500 ml of water from each vereda in polyethylene bottles. Later, in the laboratory, we used Vacu-vials kits to quantify ammonia (model K-1503) and nitrate (model K-7003). The analysis was performed using a spectrophotometer (model B572A). The luminosity was measured through a quantum sensor (model LI-250A) every 25 m of the collection transect, totaling five measurements, of which we generate an average per vereda. Finally, we quantify the remaining physical variables using a HORIBA multiparametric.
To assess the anthropogenic impact in the veredas (landscape change), we used parameters established by the Brazillian Environmental Legislation (CONAMA, 2012)—Law N° 12.727. This legislation determines that the vereda area consists of the area dominated by “buritizal” (i.e., clumps of M. flexuosa) and the surrounding native grasses. Therefore, the permanent protection area (PPA) must include the area described above plus a 50-m band. We define the PPA and environmental passive through satellite images from the Global Digital Elevation Model (ASTER GDEM) and software Qgis. Thus, we calculated as a passive the percentage of landscape change (evidence of burning, conversion of native vegetation, and deforestation) present in each sampled vereda. We emphasize that the presence of these environmental changes was confirmed through field observations.
Data Analysis
We considered each vereda as a sample unit; therefore, we had five dam-free sample units and seven sample units with dams, totaling 13 sample units. We ran a sensitivity analysis (Supplementary Figure S1) to assess which environmental variables and landscape metrics were altered due to dam construction (Barbour et al., 1996). After that, we performed a Student T-test on each environmental predictor to test the sensitivity analysis results. The premises of homogeneity of variances and normal distribution were tested, and, when it was not achieved, we performed the T-test with separate variances (Supplementary Table S2). Finally, to reduce dimensionality and visualize the environmental variables that differed among veredas with and without dams, we performed a principal component analysis (PCA). Variables were previously standardized. We used the Broken Stick criteria as our stopping criteria. Therefore, the main axis of the PCA was used to represent a gradient of environmental change related to dam construction.
To test the hypothesis that beta diversity would be affected by environmental changes caused by dam construction, we built simple linear regression models for both Odonata and Heteroptera groups with beta diversity as our response variable and the first axis of the PCA as our environmental predictor. Beta diversity was calculated by LCBD, which is a method to estimate how much each locality (i.e., vereda) contributes to BDTotal. In that case, localities with higher values of LCBD are those with a higher number of unique species (Legendre and De Cáceres, 2013). The normality assumption in the regression residuals was assessed by comparing the residuals to estimated values under a normal distribution (Supplementary Figure S2). To minimize spatial autocorrelation bias, we computed Moran’s I for the residuals of each regression model. Those with Moran’s I < 0.2 in all distance classes were considered free of spatial autocorrelation (Supplementary Figures S3, S4). Finally, to quantify how much each species contributes to the BDTotal of Odonata and Heteroptera, we computed SCBD (Legendre and De Cáceres, 2013).
All analyses were carried out in the software R (R Core Team, 2020). PCA was performed using the function “princomp,” the T-tests using function “lm,” and the linear regression models using function “lm,” all from package stats. The analysis of spatial autocorrelation was performed using software SAM (Rangel et al., 2010).
Results
There were changes in one limnological variable and one landscape metric when we compared veredas with and without dams. Both ORP (tvar. sep. = 2.923; df = 9.184; p = 0.016) and landscape change (tvar. sep. = 13.026; df = 5; p > 0.001) were higher in veredas with dams. After ordination, 80% of all environmental variation was synthesized into the first axis of the PCA, which separated veredas with and without dams (Figure 3A).
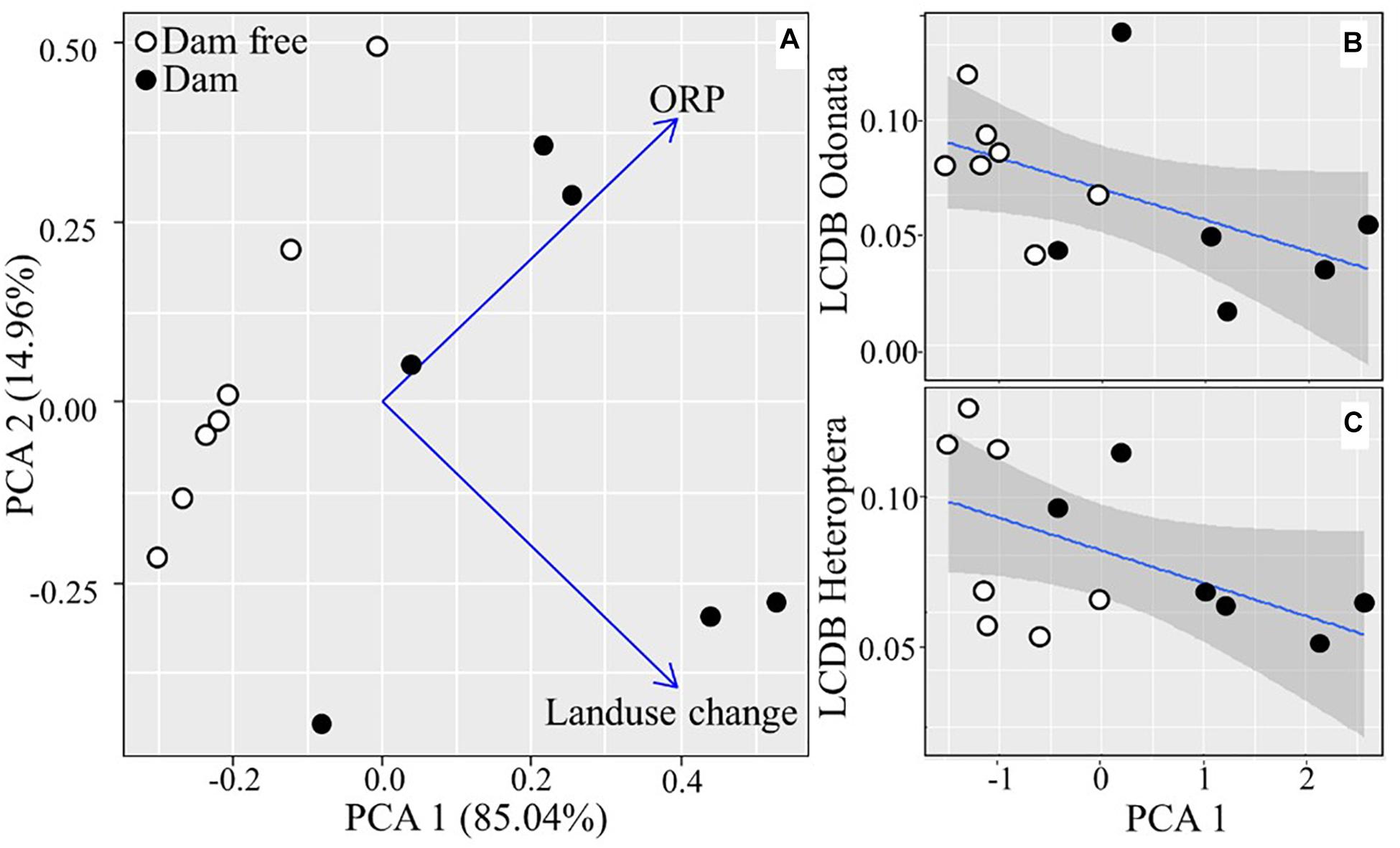
Figure 3. (A) Variable ordination. Regression models between the environmental gradient represented by the first PCA axis and beta diversity of Odonata (B) and Heteroptera (C) in veredas affected by dam construction and dam-free veredas in the Brazilian Cerrado, in the municipalities of Nova Xavantina and Barra do Garças, MT, Brazil.
The environmental gradient of landscape and limnological change between veredas with and without dams represented by the first PCA axis explained 65% of the beta diversity of Odonata (R2 = 0.650; p < 0.001; y = 0.076–0.013 ∗ x) (Figure 3B). The species that contributed the most to the BDTotal of Odonata were as follows: Argia lilacina (Selys, 1865) (SCBD = 0.178), Acanthagrion apicale (Selys, 1876) (SCBD = 0.128), Acanthagrion truncatum (Selys, 1876) (SCBD = 0.118), and Argia chapadae (Calvert, 1909) (SCBD = 0.108). This environmental gradient also explained 74% of the beta diversity of Heteroptera (R2 = 0.740; p < 0.001; y = 0.076–0.019 ∗ x) (Figure 3C). The species that mostly contributed to this pattern were: Rhagovelia whitei (Breddin, 1898) (SCBD = 0.303), Limnogonus recurvus (Drake and Harris, 1930) (SCBD = 0.145), Neogerris lotus (White, 1879) (SCBD = 0.132), and Rhagovelia robusta (Gould, 1931) (SCBD = 0.118).
Discussion
Dam construction changes the environmental, limnological, and landscape conditions of veredas, affecting the beta diversity of both Heteroptera and Odonata communities, confirming our hypothesis. There are already indications that small dams can change Odonata and Heteroptera assemblages (Brasil et al., 2014a; De Marco et al., 2014) in Cerrado streams. Additionally, Odonata (Carvalho et al., 2013; Calvão et al., 2018) and Heteroptera (Vieira et al., 2015; Dias-Silva et al., 2020) assemblages can be affected by changes in land use when they are embedded in areas used for cattle raising, even in streams without dams. It happens mostly because cattle change the riparian vegetation by trampling on it when they drink water. Our results demonstrate that even small dams in veredas may promote both biotic and abiotic changes.
Only dams without livestock may be less impactful, but it is impossible to disassociate these impacts completely. Cattle raising is the main driver of the increase in Brazilian deforestation rates (Pereira et al., 2020). Although most rural properties in Brazil comply with environmental legislation, when we look at absolute values, 17% of beef exported to the European Union is produced in violation of the legislation (Rajão et al., 2020). This work shows that the highest rates of landscape change happen in veredas subject to both damming and cattle-raising activities. This increase in landscape conversion results in changes in the limnological conditions and the aquatic biota of streams in the Cerrado (Calvão et al., 2018), in the Amazon forest (Montag et al., 2019), and the transition between these two formations (Vieira et al., 2015). Those landscape changes are also taking place in the veredas, even though they are areas of permanent protection by the Brazilian legislation (Giehl et al., 2020).
Among water quality parameters, the ORP of aquatic environments is related to the integrity of these environments. Even though the Conselho Nacional do Meio Ambiente (CONAMA) resolution (N° 357/2005) does not establish maximum standard values of ORP, positive values are considered indicative of oxidation of chemical elements in aquatic systems (Hinchman and Kuczynski, 2019). In our study, the highest values of ORP in the veredas with dams may have been caused by the nitrification process, which consists of the biological oxidation of ammonia-producing nitrate as a subproduct (Grunditz and Dalhammar, 2001). Therefore, although we do not see differences in nitrate values, ORP values indicate nitrate inputs in the recent past but now altered by biological oxidation processes and indicated by ORP values. This process probably happens due to the input of cattle urine and nitrogen fertilizers from monocultures in the uppermost areas of the river basins (Giehl et al., 2020). All the above cited processes change the limnological parameters likely affecting the majority of aquatic organisms (Esteves, 2011), including Odonata and Heteroptera insects.
The species that mostly contributed to Odonata beta diversity were the following: A. lilacina, A. apicale, A. truncatum, and A. chapadae, respectively. These taxa belong to the same family (Coenagrionidae) and exhibit a wide range of occurrence in the Cerrado’s aquatic systems, including the veredas (Cortês et al., 2011; Vilela et al., 2016; Borges et al., 2019). Species such as A. lilacina and A. truncatum have similar environmental requirements and generally occur in open and lotic environments (Lencioni, 2005). However, they are also frequently found in habitats impacted by agriculture (Ferreira-Peruquetti and Fonseca-Gessner, 2003) or cattle-raising activities (Juen et al., 2014; Calvão et al., 2018). This wide distribution shows these species’ higher tolerance to environmental change, which may explain why they occur in veredas both with and without dams.
The species that contributed the most to Heteroptera beta diversity were R. whitei (SCBD = 0.303), L. recurvus (SCBD = 0.145), Nymphaea lotus (SCBD = 0.132), and R. robusta (SCBD = 0.118). R. whitei was considered one of the most abundant species in streams from the uppermost regions of the Pantanal (Heckman, 1998). It is also commonly found and widely distributed in streams of the eastern portion of the Mato Grosso State, with a higher frequency in habitats with gallery forests (Dias-Silva et al., 2013, 2020) and in veredas (Giehl et al., 2020). L. recurvus is also widely distributed and has been found in altered habitats with high solar incidence (Dias-Silva et al., 2013), mostly occurring in altered veredas (Giehl et al., 2020). N. lotus is also common in streams of the eastern portion of the Mato Grosso State, occurring in both shaded and open habitats in the Cerrado (Dias-Silva et al., 2013). It is considered a generalist in veredas (Giehl et al., 2020). R. robusta has been found in environments that range from streams with well-preserved riparian vegetation to streams embedded in pastures (Dias-Silva et al., 2013). However, in veredas, it mostly occurs in preserved areas (Giehl et al., 2020).
Our results show that veredas impacted by dams are altered environments, which causes pressure upon the distribution of regionally exclusive species here shown by the LCBD. Because the veredas are areas of permanent protection by the Brazilian legislation and ecosystems that are critical for the Cerrado hydric maintenance, we emphasize the necessity of greater supervision of the veredas, especially of those embedded in areas used for cattle raising, where veredas have been commonly used for water storage by damming. In addition, there is another important issue that we do not address here. Is the 50-m strip efficient to mitigate anthropogenic activities on aquatic ecosystems, represented here by the veredas? We suggest that other studies research this issue question.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Author Contributions
LB, NG, LJ, BR, and VF developed the concept and contributed to method development. JB, LC, NG, BR, and VF contributed to field testing. JB and LC contributed to obtaining funding. All authors contributed to method critique and sections of text for the manuscript.
Funding
We are grateful to the “Fundação de Amparo à Pesquisa do Estado de Mato Grosso” for funding (#300321/2013). We would also like to thank CAPES and CNPq for supporting this research through individual research grants for the authors. BR and VF are grateful to the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (Capes) for the doctoral scholarship (88887.493682/2020-00 and 88887.469089/2019). NG thanks the “Programa de Pesquisa e Desenvolvimento Tecnológico do Setor de Energia Elétrica-Companhia Energética de Minas Gerais” (P&D Aneel-Cemig GT-599) for a Ph.D. scholarship. LJ is grateful for the productivity grant (304710/2019-9) of Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the hydroecology laboratory of Nova Xavantina for measuring nitrate and ammonia in water samples. Finally, we would also like to thank the “Laboratório de Entomologia de Nova Xavantina” for both logistical and structural support.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fevo.2021.612642/full#supplementary-material
References
Alencar, A., Shimbo, J. Z., Lenti, F., Balzani Marques, C., Zimbres, B., Rosa, M., et al. (2020). Mapping three decades of changes in the Brazilian savanna native vegetation using landsat data processed in the google earth engine platform. Remote Sens. 12:924. doi: 10.3390/rs12060924
Allan, J. D., and Castillo, M. M. (2007). Stream Ecology: Structure and Function of Running Waters. Berlin: Springer.
Barbour, M. T., Gerritsen, J., Griffith, G. E., Frydenbourg, R., Mc Carron, E., White, J. S., et al. (1996). A framework for biological criteria for Florida streams using benthic macroinvertbrates. J. N. Am. Benthol. Soc. 15, 185–211. doi: 10.2307/1467948
Baselga, A. (2010). Partitioning the turnover and nestedness com- ponents of beta diversity. Glob. Ecol. Biogeogr. 19, 134–143. doi: 10.1111/j.1466-8238.2009.00490.x
Borges, L. R., Barbosa, M. S., Carneiro, M. A. A., Vilela, D. S., and Santos, J. C. (2019). Dragonflies and damselflies (Insecta: Odonata) from a Cerrado area at Triângulo Mineiro, Minas Gerais, Brazil. Biota Neotrop. 19, 1–10.
Brasil, L. S., Giehl, N. F. S., Almeida, S. M., Valadão, M. B. X., Santos, J. O., Pinto, N. S., et al. (2014a). Does the damming of streams in the southern Amazon basin affect dragonfly and damselfly assemblages (Odonata: Insecta)? A preliminary study. Int. J. Odonatol. 17, 187–197. doi: 10.1080/13887890.2014.963712
Brasil, L. S., Juen, L., Batista, J. D., Pavan, M. G., and Cabette, H. S. R. (2014b). Longitudinal distribution of the functional feeding groups of aquatic insects in streams of the Brazilian Cerrado Savanna. Neotrop. Entomol. 43, 421–428. doi: 10.1007/s13744-014-0234-9
Brasil, L. S., Lima, E. L., Spigoloni, Z. A., Ribeiro-Brasil, D. R. G., and Juen, L. (2020a). The habitat integrity index and aquatic insect communities in tropical streams: a meta-analysis. Ecol. Indic. 116:106495. doi: 10.1016/j.ecolind.2020.106495
Brasil, L. S., Luiza-Andrade, A., Calvão, L. B., Dias-Silva, K., Faria, A. P. J., Shimano, Y., et al. (2020b). Aquatic insects and their environmental predictors: a scientometric study focused on environmental monitoring in lotic environmental. Environ. Monit. Assess. 192:194.
Calvão, L. B., Juen, L., Oliveira Junior, J. M. B., Batista, J. D., and De Marco Júnior, P. (2018). Land use modifies Odonata diversity in streams of the Brazilian Cerrado. J. Insect Conserv. 22, 675–685. doi: 10.1007/s10841-018-0093-5
Carvalho, F. G., Pinto, N. S., Oliveira-Júnior, J. M. B., and Juen, L. (2013). Effects of marginal vegetation removal on Odonata communities. Acta Limnol. Bras. 25, 10–18. doi: 10.1590/s2179-975x2013005000013
Casatti, L., Paula-Ferreira, C., and Carvalho, F. R. (2009). Grass-dominated stream sites exhibit low fish species diversity and dominance by guppies: an assessment of two tropical pasture river basins. Hydrobiologia 632, 273–283. doi: 10.1007/s10750-009-9849-y
Cezário, R. R., Firme, P. P., Pestana, G. C., Vilela, D. S., Juen, L., Cordero-Rivera, A., et al. (2020). “Sampling methods for dragonflies and damselflies,” in Measuring Arthropod Biodiversity: A Handbook of Sampling Methods, eds J. C. Santos and G. W. Fernandes (Cham: Springer).
CONAMA (2012). Lei n° 12.727. Conselho Nacional do Meio Ambiente, Brasília, Brasil. Available online at: https://www.planalto.gov.br/ccivil_03/_ato2011-2014/2012/lei/L12727.htm (accessed August 14, 2020).
Cortês, L. G., Almeida, M. C., Pinto, N. S., and De Marco Júnior, P. (2011). Fogo em Veredas: avaliação de Impactos sobre Comunidades de Odonata (Insecta). Biodivers. Bras. 2, 128–145.
De Marco, P., Nogueira, D. S., Correa, C. C., Vieira, T. B., Silva, K. D., Pinto, N. S., et al. (2014). Patterns in the organization of Cerrado pond biodiversity in Brazilian pasture landscapes. Hydrobiologia 723, 87–101. doi: 10.1007/s10750-013-1695-2
De Marco, P., and Resende, D. C. (2002). Activity patterns and thermoregulation in a tropical dragonfly assemblage. Odonatologica 31, 129–138.
Dias-Silva, K., Brasil, L. S., Veloso, G. K. O., Cabette, H. S. R., and Juen, L. (2020). Land use change causes environmental homogeneity and low beta-diversity in Heteroptera of streams. Ann. Limnol. Int. J. Lim. 56, 1–9.
Dias-Silva, K., Cabette, H. R. S., Juen, L., and De Marco Júnior, P. (2010). The of habitat integrity and physical-chemical water variables on the structure of aquatic and semi-aquatic Heteroptera. Rev. Bras. Zool. 27, 918–930. doi: 10.1590/s1984-46702010000600013
Dias-Silva, K., Moreira, F. F. F., Giehl, N. F. S., Nóbrega, C. C., and Cabette, H. S. R. (2013). Gerromorpha (Hemiptera: Heteroptera) of eastern Mato Grosso State, Brazil: checklist, new records, and species distribution modeling. Zootaxa 3, 201–235. doi: 10.11646/zootaxa.3736.3.1
Ferreira-Peruquetti, P. S., and Fonseca-Gessner, A. A. (2003). Comunidade de Odonata (Insecta) em áreas naturais de Cerrado e monocultura no nordeste do Estado de São Paulo, Brasil: relação entre o uso do solo e a riqueza faunística. Rev. Bras. Zool. 20, 219–224. doi: 10.1590/s0101-81752003000200008
Garrison, R. W., von Ellenrieder, N., and Louton, J. A. (2006). Dragonfly Genera of the New World: An Illustrated and Annotated Key to the Anisoptera. Baltimore, MD: The Johns Hopkins University Press.
Garrison, R. W., von Ellenrieder, N., and Louton, J. A. (2010). Damselfly Genera of the New World: An Illustrated and Annotated Key to the Zygoptera. Baltimore, MD: The Johns Hopkins University Press.
Giehl, N. F. S., Cabette, H. S. R., Dias-Silva, K., Juen, L., Moreira, F. F. F., Castro, L. A., et al. (2020). Variation in the diversity of semiaquatic bugs (Insecta: Heteroptera: Gerromorpha) in altered and preserved veredas. Hydrobiologia 847, 3497–3510. doi: 10.1007/s10750-020-04364-1
Grunditz, C., and Dalhammar, G. (2001). Development of nitrification inhibition assays using pure cultures of nitrosomonas and nitrobacter. Water Res. 3, 433–440. doi: 10.1016/s0043-1354(00)00312-2
Heckman, C. W. (1998). The seasonal succession of biotic communities in wetlands of the tropical wet-and-dry climatic zone: V. Aquatic Invertebrate Communities in the Pantanal of Mato Grosso, Brazil. Int. Rev. Hydrobiol. 83, 31–63. doi: 10.1002/iroh.19980830105
Heino, J., and Grönroos, M. (2017). Exploring species and site contributions to beta diversity in stream insect assemblages. Oecologia 183, 151–160. doi: 10.1007/s00442-016-3754-7
Hinchman, A., and Kuczynski, T. L. (2019). Determination of Fluid Quality Using pH and Conductivity/ORP. U.S. Patent No 2019/0041376 A1. Washington, DC: U.S. Patent and Trademark Office.
Juen, L., and De Marco, P. Jr. (2011). Odonate biodiversity in terra-firme forest streamlets in Central Amazonia: on the relative effects of neutral and niche drivers at small geographical extents. Insect Conserv. Divers. 4, 265–274. doi: 10.1111/j.1752-4598.2010.00130.x
Juen, L., Oliveira-Junior, J. M. B., Shimano, Y., Mendes, T. P., and Cabette, H. S. R. (2014). Composição e riqueza de Odonata (Insecta) em riachos com diferentes níveis de conservação em um ecótone Cerrado-Floresta Amazônica. Acta Amaz. 44, 223–233. doi: 10.1590/s0044-59672014000200008
Klein, C. E., Pinto, N. S., Spigoloni, Z. A. V., Bergamini, F. M., De Melo, F. R., De Marco, J. P., et al. (2018). The influence of small hydroelectric power plants on the richness and composition of Odonata species in the Brazilian Savanna. Int. J. Odonatol. 21, 33–44. doi: 10.1080/13887890.2017.1419884
Legendre, P., and De Cáceres, M. (2013). Beta diversity as the variance of community data: dissimilarity coefficients and partitioning. Ecol. Lett. 16, 951–963. doi: 10.1111/ele.12141
Lencioni, F. A. A. (2006). Damselflies of Brazil: An Illustrated Identification Guide 2 – Coenagrionidae Families. São Paulo: All Print Editora.
Magalhães, O. M., Moreira, F. F. F., and Galvão, C. (2016). A new species of Rhagovelia Mayr, 1865 (Hemiptera: Heteroptera: Veliidae) from Pará state, with an updated key to Brazilian species of the robusta group. Zootaxa 4171, 586–594. doi: 10.11646/zootaxa.4171.3.12
Meirelles, M. L., Ferreira, A. B., and Franco, A. C. (2006). Dinâmica Sazonal do Carbono em campo Úmido do Cerrado. Planaltina: Embrapa Cerrados.
Miguel, T. B., Calvão, L. B., Vital, M. V. C., and Juen, L. (2017). A scientometric study of the order Odonata with special attention to Brazil. Int. J. Odonatol. 20, 27–42. doi: 10.1080/13887890.2017.1286267
Montag, L. F., Winemiller, K. O., Keppeler, F. W., Leão, H., Benone, N. L., Torres, N. R., et al. (2019). Land cover, riparian zones and instream habitat influence stream fish assemblages in the eastern Amazon. Ecol. Freshw. Fish. 28, 317–329. doi: 10.1111/eff.12455
Nieser, M., and Melo, A. L. (1997). Os Heterópteros Aquáticos de Minas Gerais. Belo Horizonte: UFMG.
Oliveira-Junior, J. M. B., and Juen, L. (2019). The Zygoptera/Anisoptera ratio (Insecta: Odonata): a new tool for habitat alterations assessment in Amazonian streams. Neotrop. Entomol. 48, 552–560. doi: 10.1007/s13744-019-00672-x
Peel, M. C., Finlayson, B. L., and McMahon, T. A. (2007). Updated world map of the Köppen–Geiger climate classification. Hydrol. Earth Syst. Sci. 4, 439–473.
Pereira, E. J. A. L., Santana Ribeiro, L. C., Silva Freitas, L. F., and Barros Pereira, H. B. (2020). Brazilian policy and agribusiness damage the Amazon rainforest. Land Use Policy 92:104491. doi: 10.1016/j.landusepol.2020.104491
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rajão, R., Soares-Filho, B., Nunes, F., Börner, J., Machado, L., Assis, D., et al. (2020). The rotten apples of Brazil’s agribusiness. Science 369, 246–248.
Rangel, T. F., Diniz-Filho, J. A. F., and Bini, L. M. (2010). SAM: a comprehensive application for spatial analysis in macroecology. Ecography 33, 46–50. doi: 10.1111/j.1600-0587.2009.06299.x
Ribeiro, J. F., and Walter, B. M. T. (2008). “As Principais Fitofisionomias do bioma Cerrado,” in Cerrado: Ecologia E Flora, eds J. F. Ribeiro, S. P. Almeida, and S. M. Sano (Planaltina: Embrapa Cerrados), 153–212.
Rúdio, J. A., and Moreira, F. F. F. (2011). A new species of Microvelia (Insecta: Hemiptera: Heteroptera: Gerromorpha) from coastal Espírito Santo State, with a key to the species of the genus recorded from southeastern Brazil. Zootaxa 3004, 63–68. doi: 10.11646/zootaxa.3004.1.6
Tubelis, D. P. (2009). Veredas and their use by birds in the Cerrado, South America: a review. Biota Neotrop. 9, 363–374. doi: 10.1590/s1676-06032009000300031
Vannote, R. L., Minshall, G. W., Cummins, K. W., Sedell, J. R., and Cushing, C. E. (1980). The river continuum concept. Can. J. Fish. Aquat. Sci. 37, 130–137.
Vieira, T. B., Dias-Silva, K., and Pacífico, E. D. S. (2015). Effects of riparian vegetation integrity on fish and heteroptera communities. Appl. Ecol. Environ. Res. 13, 53–65.
Keywords: aquatic insects, Brazilian savanna, livestock, cattle raising, metacommunities, water security, palm swamp
Citation: Brasil LS, Ferreira VRS, Resende BO, Juen L, Batista JD, Castro LA and Giehl NFS (2021) Dams Change Beta Diversity of Aquatic Communities in the Veredas of the Brazilian Cerrado. Front. Ecol. Evol. 9:612642. doi: 10.3389/fevo.2021.612642
Received: 30 September 2020; Accepted: 18 January 2021;
Published: 19 February 2021.
Edited by:
Haipeng Wu, Changsha University of Science and Technology, ChinaReviewed by:
Rogério Pereira Bastos, Universidade Federal de Goiás, BrazilEduardo Périco, Universidade do Vale do Taquari - Univates, Brazil
Cássia Munhoz, University of Brasilia, Brazil
Copyright © 2021 Brasil, Ferreira, Resende, Juen, Batista, Castro and Giehl. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Leandro Schlemmer Brasil, bGVhbmRyb2JyYXNpbEB1ZnBhLmJy; YnJhc2lsX2Jpb2xvZ2lhQGhvdG1haWwuY29t