 Adriano Conte1
Adriano Conte1 Silvano Fares1*
Silvano Fares1* Luca Salvati1Flavia Savi1
Luca Salvati1Flavia Savi1 Giorgio Matteucci2
Giorgio Matteucci2 Francesco Mazzenga3Donatella Spano4,5
Francesco Mazzenga3Donatella Spano4,5 Costantino Sirca4,5
Costantino Sirca4,5 Serena Marras4,5Marta Galvagno6
Serena Marras4,5Marta Galvagno6 Edoardo Cremonese6
Edoardo Cremonese6 Leonardo Montagnani7,8
Leonardo Montagnani7,8- 1Council for Agricultural Research and Economics-Research Centre for Forestry and Wood, Rome, Italy
- 2National Research Council of Italy-Institute for Agricultural and Forestry Systems in the Mediterranean, Ercolano, Italy
- 3National Research Council of Italy-Research Institute on Terrestrial Ecosystems (IRET), Monterotondo Scalo, Italy
- 4Department of Agraria, University of Sassari, Sassari, Italy
- 5IAFES Division, Euro-Mediterranean Centre on Climate Change (CMCC Foundation), Sassari, Italy
- 6Climate Change Unit, Environmental Protection Agency of Aosta Valley, ARPA Valle d'Aosta, Aosta, Italy
- 7Faculty of Science and Technology, Free University of Bolzano, Bolzano, Italy
- 8Forest Services, Autonomous Province of Bolzano, Bolzano, Italy
In the Mediterranean region, ecosystems are severely affected by climate variability. The Italian Peninsula is a hot spot for biodiversity thanks to its heterogeneous landscape and the Mediterranean, Continental, and Alpine climates hosting a broad range of plant functional types along a limited latitudinal range from 40′ to 46′ N. In this study we applied a comparative approach integrating descriptive statistics, time series analysis, and multivariate techniques to answer the following questions: (i) do the climatic variables affect Gross Primary Productivity (GPP), Reco, Water Use Efficiency (WUE), and ET to a similar extent among different sites? (ii) Does a common response pattern exist among ecosystems along a latitudinal gradient in Italy? And, finally (iii) do these ecosystems respond synchronically to meteorological conditions or does a delayed response exist? Six sites along a latitudinal, altitudinal, and vegetational gradient from semi-arid (southern Italy), to a mountainous Mediterranean site (central Italy), and sub-humid wet Alpine sites (northern Italy) were considered. For each site, carbon and water fluxes, and meteorological data collected during two hydrologically-contrasting years (i.e., a dry and a wet year) were analyzed. Principal Component Analysis (PCA) was adopted to identify temporal and spatial variations in GPP, Ecosystem Respiration (Reco), WUE, and Evapotranspiration (ET). The model outlined differences among Mediterranean semi-arid, Mediterranean mountainous, and Alpine sites in response to contrasting precipitation regimes. GPP, Reco, WUE, and ET increased up to 16, 19, 25, and 28%, respectively in semi-arid Mediterranean sites and up to 15, 32, 15, and 11%, respectively in Alpine sites in the wet year compared to the dry year. Air temperature was revealed to be one of the most important variables affecting GPP, Reco, WUE, and ET in all the study sites. While relative air humidity was more important in southern Mediterranean sites, global radiation was more significant in northern Italy. Our work suggests that a realistic prediction of the main responses of Italian forests under climate change should also take in account delayed responses due to acclimation to abiotic stress or changing environmental conditions.
Introduction
Air temperatures and rainfalls are among the major climatic drivers of the terrestrial carbon cycle, and climate change can strongly affect ecosystem processes such as photosynthesis and respiration (Griffin and Prager, 2017). For instance, drought stress induces stomatal closure and consequently decreases photosynthesis (Osakabe et al., 2014). Global warming is also associated with reduced soil respiration due to a reduced microbial activity and root growth (Jensen et al., 2003; Schlesinger et al., 2016). By means of the increase in the occurrence of extreme heat and drought events, a substantial drying and warming of ecosystems in Mediterranean regions is expected (Gouveia et al., 2017; Paeth et al., 2017). Different patterns of these changes are expected for the various microclimates that characterize the Italian peninsula (Chelli et al., 2017). Increasing mean annual temperature and a reduction of precipitation (in particular snowfall) in summer and winter are expected in both the Alpine and in the mountainous temperate climatic zones (Gobiet et al., 2014). Increasing temperature and an inversion of the precipitation annual pattern (lower in winter and higher in summer) followed by a concurrent increase in intensity and frequency of extreme events is expected for the coastal Mediterranean area (Giorgi and Lionello, 2008). Spatio-temporal variability in rainfall regimes affects ecophysiological processes in a diverging way: on one side drought stress can stimulate plant metabolism to acclimatize to water stress, driving to enhanced plant respiration (Flexas et al., 2005; Bhargava and Sawant, 2013). On the other side, precipitation pulses can stimulate photosynthesis (Huxman et al., 2004; Wu et al., 2011) and ecosystem respiration (Inglima et al., 2009). Water Use Efficiency (WUE, the ratio of carbon gain by photosynthesis to water loss by evapotranspiration) represents the link between carbon and water cycles at a local scale and was considered for a long time to be representative of plants' water use strategies (Donovan and Ehleringer, 1991). Earlier studies showed an increase in WUE over the last decades in response to growing CO2 concentrations, due to reduced stomatal conductance to maintain the intercellular CO2 concentration at a constant level (Yu et al., 2008; Pe-uelas et al., 2011; Keenan et al., 2013). WUE for arid ecosystems showed a negative response to drought, whereas WUE for humid ecosystems showed both positive and negative responses to drought (Huang et al., 2017). A recent study by Limousin et al. (2015) partly contradicts this paradigm, demonstrating that a convergence in WUE response to drought exists in plant species with different tolerances to drought. The responses of variables such as WUE and, more generally, Gross Primary Productivity (GPP), Ecosystem respiration (Reco), and Evapotranspiration (ET) to weather variability and changes in climate regimes at ecosystem level can be studied with the eddy covariance technique. This technique provides high-time frequency resolution carbon and water flux measurements, allowing a reliable evaluation, at high temporal resolution, of ecosystem GPP, Reco, ET, and WUE (Aubinet et al., 2012). Earlier studies identified time-lags of months and years in the response of GPP and Reco to environmental changes (Zhang et al., 2014). Relatively few studies have focused on hours to day delayed eco-physiological and ecological responses to weather variability (Bréda et al., 2006; Incerti et al., 2007). The ecological importance of these lagged responses was recently discussed and experimental methods were proposed to fill this knowledge gap (Detto et al., 2013; Montagnani et al., 2018). We took advantage of an important anomaly observed in the Italian peninsula in 2014, marked by a very cool and wet summer, in particular in the northern part of Italy, where monthly precipitations in July were 84% above the mean for the years 1982–2013 (NIMBUS, 2015; Ratna et al., 2017), preceded by a moderately dry year (2013), to test the following hypotheses: (i) Do the climatic variables affect GPP, Reco, WUE, and ET to a similar extent among different sites? (ii) Does a common response pattern exist among ecosystems along a latitudinal gradient in Italy? (iii) Is the statistical prediction of GPP, Reco, WUE, and ET behavior improved taking account of both synchronic and delayed vegetation responses to meteorological variables?
Materials and Methods
Site Description
The study sites were selected as representative of a wide set of ecosystems in the Italian peninsula and the island of Sardinia from Mediterranean coastal to alpine sites, homogeneously distributed from northern Italy to central-southern Italy, ranging from 40° to 46°N latitude, 7° to 13°E longitude, and 20 m to 2160 m at the sea level (Figure 1). This combined latitudinal and altitudinal diversification influences local climate regimes, soil attributes, and the vegetation characteristics (Table 1).
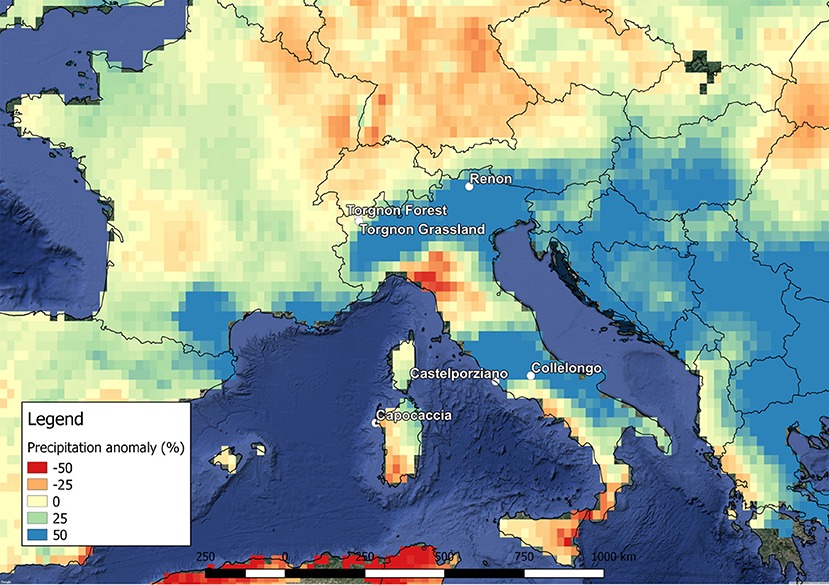
Figure 1. Annual precipitation anomaly (%) observed in 2014 compared with the climatic average 1981–2010 method used for analysis of anomalies by Haylock et al. (2008). The six experimental sites in Italy are reported in the map.
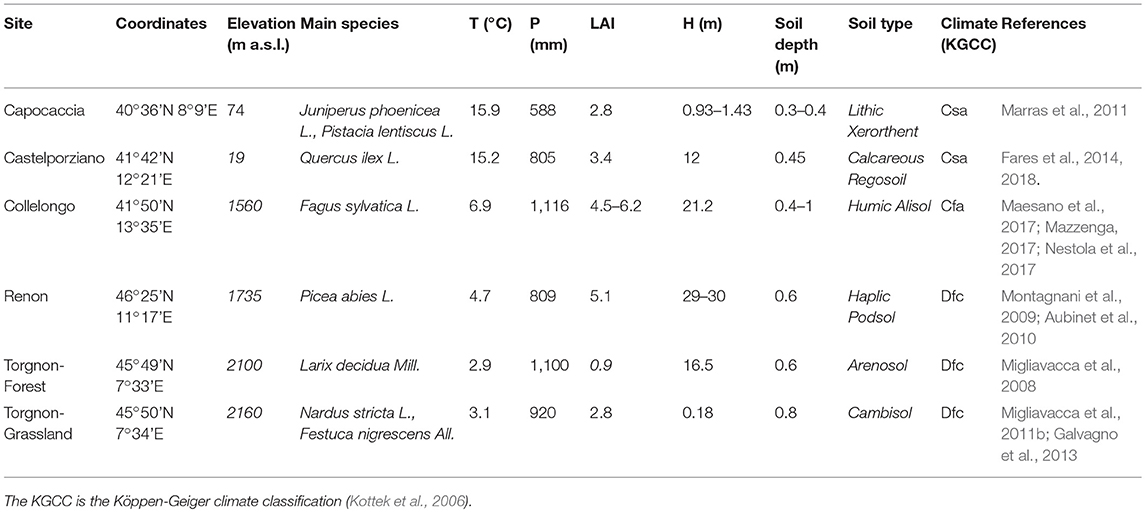
Table 1. Main characteristics of the experimental sites.
The selected sites fall into three main climate classes: Mediterranean semi-arid (Temperate with dry and hot summer, Csa), Mediterranean mountainous (Temperate without dry season, Cfa), and intra-alpine semi-continental (Continental, without a dry season and with a cool summer, Dfc). The vegetation at the sites is represented by maquis (shrubland with densely growing evergreen shrubs), a holm oak forest, a beech forest, a spruce forest, a subalpine grassland, and a larch forest, for Capo Caccia, Castelporziano, Collelongo, Renon, Torgnon grassland and Torgnon forest sites, respectively. A detailed description and instruments used for each site is presented in Supplementary Materials. The sites are equipped with eddy covariance towers measuring CO2, H2O, and energy fluxes between the ecosystems and the atmosphere. All the sites considered in this study are part of the ICOS and/or Fluxnet international networks.
Meteorology and Flux Analysis
Two years (2013 and 2014) of continuous meteorological and flux data (30 min. time resolution) measured at all sites were considered in this study. Air and soil temperature (Tair and Tsoil, °C), precipitation (P, mm), relative air humidity (RH, %), global radiation (RAD, W m-2), and soil water content (SWC, %) were recorded every minute and averaged for 30 min. intervals with meteorological sensors. Flux measurement equipment was installed on scaffold towers. Tri-dimensional sonic anemometers were used to measure instantaneous wind speed and temperature fluctuations. H2O and CO2 concentrations were measured with an infrared gas analyser. Data were recorded at 10 Hz for all trace gases. For information on instrumentation models used in each site and flux calculation routines, please see the references cited in Table 1.
H2O, CO2, and latent and sensible heat fluxes were calculated according to the eddy covariance technique (Aubinet et al., 2012). In order to avoid any discrepancy in flux calculation and partitioning that may arise from the use of different flux processing routines, GPP (GPP, μmol (CO2) m−2 s−1), and Ecosystem Respiration (Reco, μmol (CO2) m−2 s−1) were calculated with the REddyProcWeb flux-partitioning tool based on Reichstein et al. (2005). Ecosystem Water Use Efficiency [WUE, g(CO2) kg−1(H2O)] was calculated as the ratio of carbon gain [GPP, g(CO2)] to water consumption [Evapotranspiration ET, kg(H2O)].
Data Analysis
The investigated meteorological variables were Tair, Tsoil, vapor pressure deficit (VPD), RH, SWC, RAD, and P. A mixed analysis strategy grounded on exploratory multivariate statistics (Principal Component Analysis, PCA), correlation techniques and multiple regression models, was adopted in this study with the aim to identify (and rank the importance of) a relatively large number of contextual variables considered as candidate predictors of the eco-physiological processes investigated here (Colantoni et al., 2016). A comprehensive analysis strategy including different techniques with less stringent normality assumptions (Duvernoy et al., 2018) is particularly appropriate to investigate a vast set of synchronic and delayed relationships among environmental variables reflecting (apparent and latent) ecological processes behind an ecosystem's complexity and eco-physiological response to climate variability. The use of PCA is not a novel approach per sé, but it allows orthogonalization of a number of contextual variables (i.e., predictors) identifying only relevant, non-redundant variables that can be inserted in a subsequent regression model. We preferred to use a two-step procedure (PCA and block regression) instead of single-step approaches, such as stepwise regression or partial least square regression, because of the large number of redundant variables (stepwise regression works better with a relatively low number of redundant predictors) and the broad sample size (partial least square regression is better for smaller sample sizes). In other words, we used PCA as a preparatory analysis (Pili et al., 2017), i.e., to prepare a redundant dataset to be used as a non-redundant predictors' dataset, having investigated the latent, multivariate relationships among the original variables within the dataset. More specifically, to take into account the short-time (i.e., 3-day time window) delayed response of GPP, Reco, and WUE to the investigated variables, partial autocorrelations (PAC), i.e., the conditional correlation between the time series x0 and its own lagged values x0+I, were calculated for each dependent variable, site, and year. It was arbitrarily decided to take account of the first 3 highest (positive and/or negative) PAC coefficients shown by the resulting partial autocorrelation function (Figures S1–S8). Under the hypothesis that the selected PAC represents the response induced by one of the independent variables, they were delayed according to the lags corresponding to the selected PAC. Therefore, each dataset was finally composed by the instantaneous variables and their three delayed replicates (indicated with “_lag1”, “_lag2”, ”lag3”, respectively), ranging from 30 min to 48 h. Prior to multivariate analysis, the standardization of all variables was applied to obtain data with zero mean and variance equal to 1 with the aim to make them comparable, regardless of the original unit of measurement (Pili et al., 2017). Standardization was applied to each observed value calculating z-scores by subtracting the mean (μ) and dividing the result by the standard deviation (σ). To reduce the number of variables, a PCA run, using a Spearman non-parametric rank correlation matrix as input data matrix, was applied to the collected environmental variables (Salvati et al., 2014). A non-parametric correlation PCA matrix instead of the most traditional Pearson's one-moment correlation coefficient was adopted with the aim to analyse together linear and non-linear correlations among variables. Only principal components with absolute eigenvalues higher than 1 were considered in subsequent analysis. The components' loadings matrix (the matrix that contains the correlations between the variables and components) was used to identify which variable(s) was (were) more strongly correlated with each extracted component (Duvernoy et al., 2018). Variables with loadings > |0.4| were considered in the subsequent analysis (Colantoni et al., 2016). The scores of the selected principal components were used as statistically-independent predictors in three models (using both linear and quantile regressions) with standardized GPP, Reco, and WUE as dependent variables, respectively. The resulting models were:
Where βi is the slope of the i-th principal component (based on component scores) and εi is the model's error for each dependent variable. Comparison of the model's outputs between linear and non-linear (quantile) regression techniques allows identifying specific functional forms characterizing the statistical relationship existing between the dependent variable and the predictors, contributing to reduce the impact of variables with (more or less evident) deviations from normality on the model's results. To estimate the overall impact of environmental variables on GPP, Reco, and WUE, a composite index was finally calculated by multiplying the slope of the linear model covariates with the absolute loading of each native variable on the selected principal components, in turn multiplied by the proportion of variance explained by the same component (Salvati and Zitti, 2005)
Where Ii is the index measuring the impact of the i-th variable on the dependent variable; βi is the i-th regression coefficient; ρi is the correlation coefficient between the original variable and the i-th principal component (PCi); σ2 is the proportion of variance explained by the i-th PC.
Results
Dynamics of Climatic Conditions Among Sites
The 2 years of study (2013 and 2014) were characterized by markedly different rainfall regimes. Cumulative precipitation registered in 2013 and 2014 were, respectively: 613 and 522 mm for Capo Caccia; 665 and 900 mm for Castelporziano; 1,287 and 1,319 mm for Collelongo; 834 and 1,400 mm for Renon; 1,561 and 1,716 mm for Torgnon-forest; and 1,044 and 1,085 mm for Torgnon-grassland. The year 2013 was drier than 2014 except for at Capo Caccia. A similar rainfall distribution was observed in the precipitation anomaly map of 2014 shown in Figure 1. Four study sites fell in an area characterized by evident precipitation anomalies, indicating an increase of 9–39% rainfalls except for Capo Caccia, where an opposite trend was observed. At Collelongo, Torgnon-grassland, and Torgnon-forest the precipitation anomaly was very limited, with 3, 4, and 9%, respectively. However, these three sites experienced higher precipitations during the summer months in the wet year, with an increase of 14, 31, and 30%, respectively. In accordance with the observations described above, the year 2013 was considered the “wet year” for Capo Caccia and the “dry year” for all the other study sites; consequently, 2014 is the “dry year” for Capo Caccia and the “wet year” for all the other sites.
Response of GPP, Reco, WUE, and ET to Varying Precipitation Regimes
For each site an increase in GPP was observed during the wet year (Table 2). A similar pattern was found in the Reco for all the study sites with an exception made for Collelongo, where a slight (4%) reduction in Reco was observed in the wet year. Collelongo experienced however a very limited precipitation anomaly, therefore we expected no or very minor changes in all ecophysiological variables. Finally, WUE increased in the sites of Capo Caccia, Renon, Torgnon forest, and Torgon grassland in the wet year (Table 2), while lower values were observed in Castelporziano and Collelongo sites. Considering the magnitude of the differences in precipitation between the 2 years of study, the most responsive sites to rainfall increase concerning GPP and WUE were Collelongo forest and Torgnon-grassland, while Torgnon-forest was the most responsive site for Reco.
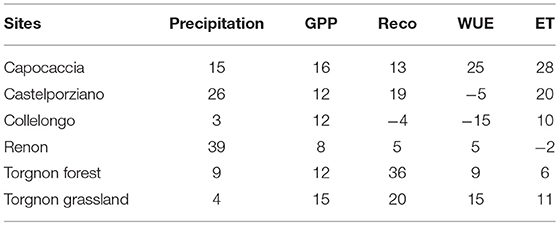
Table 2. Percent differences in Precipitation, GPP, Reco, WUE, and ET between the dry and the wet year for each site.
For the whole dataset, the highest partial autocorrelation coefficients were found at lags of 0.5, 3.5, and 24 h for GPP, 0.5, 1, and 12 h for Reco, and 0.5, 9, and 24 h for WUE, respectively (Figures S1–S8). The biplot resulting from the PCA (Figure 2) shows that the two first components specifically outline Tair (PC1) and RH (PC2) gradients. For each site, we show differences between the first two components explaining together approximately half of the total variance. Collelongo (COLL), Torgnon grassland (TOR_G), Torgnon forest (TOR), and Capo Caccia (CAP) showed similar patterns between the 2 years. The biplot showed low or no discrimination within each site, with the exception of Castelporziano (CPZ) and Renon where differences in precipitation between the 2 years of study were higher (around 500 and 240 mm, respectively) compared with the other sites. Interestingly, Northern Italian sites were concentrated on the top left of the panel, while Southern Italian sites were clustered on the bottom right.
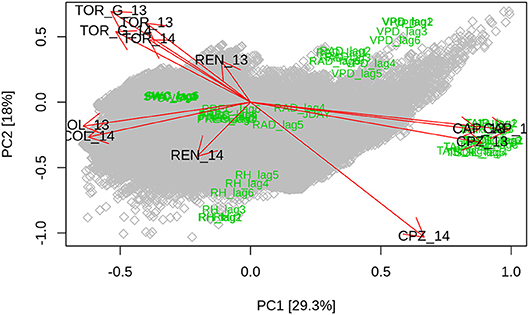
Figure 2. Biplot (arrows indicate average scores for different sites; gray squares are scores; green labels are loadings) of the two principal components showing differences in the position of the investigated sites between the 2 years of study (indicated with”_13” and “_14” for 2013 and 2014, respectively). Variables clustering together indicate similar environmental conditions. Divergence between variables' position in the biplot indicates different underlying conditions.
Linear regression models (Equations 1, 2, 3, Table 3) helped us to describe the relationship between each of the three dependent variables and the environmental factors summarized into statistically-independent variables (i.e., principal components). For GPP and Reco, the site with the highest goodness of fit (adjusted R2) was Collelongo (0.70 and 0.75, respectively for 2013 and 2014), although it was the only site with the highest R2 observed for the dry year. The site with the lowest R2 was Capo Caccia (0.40 and 0.15, respectively). Regression models on WUE showed lower R2 when compared to GPP and Reco (Table 3). WUE displayed the lowest R2 for the Northern Italy sites and the highest R2 for the Southern Italy sites. Furthermore, adjusted R2 of the WUE models in the Southern part of Italy were higher in the dry year (Collelongo and Capo Caccia) or similar between the 2 years (Castelporziano). Comparable results were obtained using quantile regressions (Tables S1–S4).
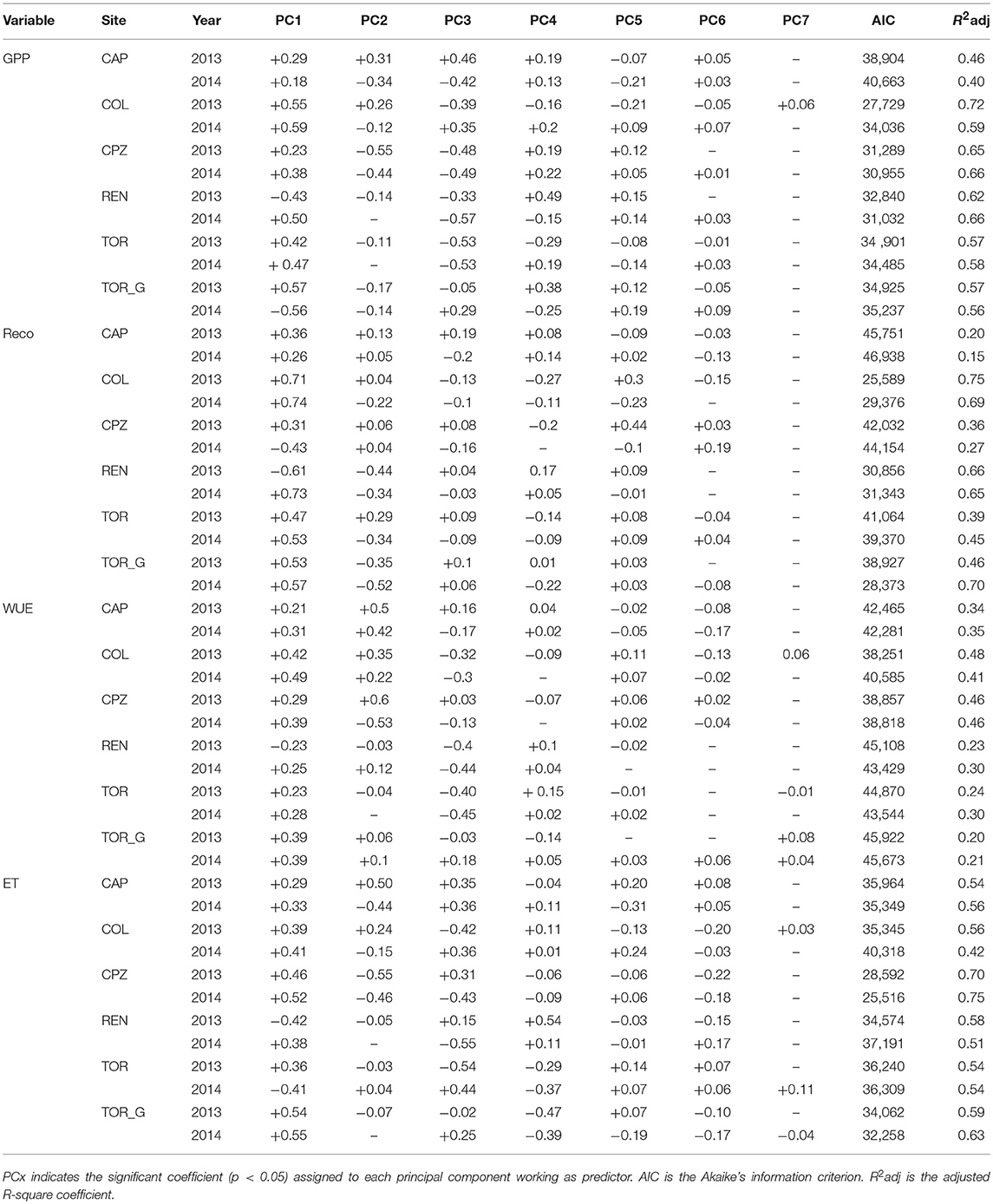
Table 3. Linear regression models for GPP, Reco, and WUE as dependent variables.
The relationship between adjusted R2 and precipitation (top) or latitude (bottom) is illustrated in Figure S9. Considering precipitation, a positive correlation was observed for Reco (with a Spearman correlation coefficient (rs) = 0.51 while a negative correlation was observed for ET (rs = −0.5), and no relationship was found for both GPP and WUE. Considering latitude, a positive correlation was observed for Reco (with a Spearman correlation coefficient (rs) = 0.61) while a negative correlation was observed for WUE (rs = −0.66), while no significant correlations were observed for GPP and ET.
To better compare environmental variables affecting GPP, Reco, and WUE between the dry and the wet year, we classified environmental variables by their coefficient of impact (Equation 4, Figure 3 and in Figures S10–S13). We observed that although the independent variables were the same for each site, there were differences in their coefficients of impact between the dry and the wet year. Tair was the variable with the highest coefficient of impact on GPP, Reco, WUE, and ET with values ranging between 50 and 88%, respectively of the total impact. The highest values were observed for Reco in the sites of Collelongo and Castelporziano with coefficient values of 87 and 88% in the wet and in the dry year, respectively. Among the others, RAD and RH were the variables with the highest coefficient. In particular, RAD showed its maximum coefficients of impact for the WUE, with values increasing from 10% (Torgnon grassland) in the northern sites to 42% (Capo Caccia) in the southern sites. RH showed different patterns among sites and dependent variables, with maximum coefficients of impact for GPP ranging from −35 to 10% for Colleongo and Torgnon grassland, respectively. Interestingly, both Capo Caccia and Colleongo showed concurrent opposite coefficients of impact during the same year. In fact, GPP resulted in a negative correlation with RH (from −7 to −35%) and a positive correlation with its 3.5 h delayed variable RH lag2 (11 to 15%), while only positive coefficients of impact were observed for the northern sites Renon and Torgnon.
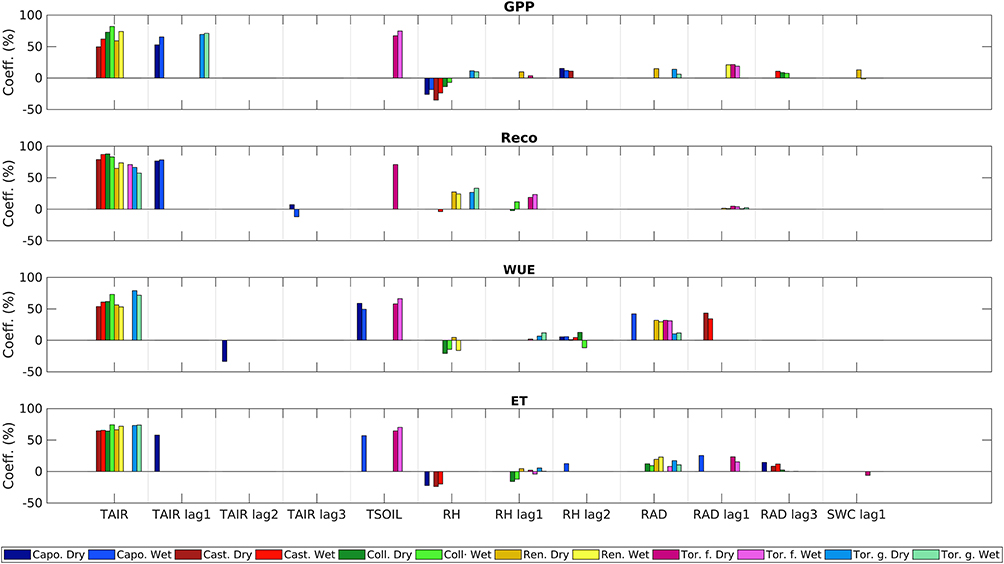
Figure 3. Comparison of the environmental variables affecting Gross Primary Productivity (GPP), Reco, Water Use Efficiency (WUE), and ET. On the X axes are reported the variables with the highest correlation coefficient with the selected principal component. On the Y axis is reported the normalized coefficient of impact with GPP, Reco and WUE. Variables with a coefficient of impact <10% are not shown.
Discussions
Inter-site Comparison
Marked climatic differences among the study sites were observed across the latitudinal gradient studied in this work. PCA analysis showed that the sites converge into three distinct groups (Figure 2). These results suggested that mountain sites such as Renon and Torgnon grassland and forest could have similar response patterns, and were strongly distinguished from Mediterranean sites such as Castelporziano and Capo Caccia. Located in central Italy and representing a mountain area with intermediate climate conditions between southern and northern Italy, Collelongo site resulted in an intermediate position between the two aforementioned groups. GPP, Reco, WUE, and ET increased in almost all the experimental sites during the wet year (the only exception was the WUE in Collelongo). These results confirm the hypothesis that changes in water availability can effectively impact plant eco-physiological processes. Marked differences between sites were observed in the goodness of fit of different regression models run in this study (Table 3). Focusing on Reco, we observed that goodness of fit of the models increased with precipitation, a statistically significant negative correlation was observed with ET (Figure S9). Collelongo, one of the sites with the highest average annual precipitation regime, showed the highest R2 while Capo Caccia, the site with the lowest precipitation, showed the lowest R2. High R2 coefficients may indicate that the selected variables were suitable to describe this process in sites characterized by high (or medium-high) water availability, while they became poor predictors in ecosystems with “water-saving” strategy where plants respond less promptly to water increase/scarcity in the soil. This strategy consists of an efficient stomatal control and other several morpho-functional traits such as roots and wood density, and leaves persistence (Canadell and Zedler, 1995; Hacke et al., 2001; Vilagrosa et al., 2003). Therefore, we found the lowest R2 in regression models predicting both Reco and GPP, in Capo Caccia, which, similarly to the holm oak ecosystem in Castelporziano, is a coastal Mediterranean schlerophyllous maquis i.e., a “water saving” ecosystem well-adapted to semi-arid climate (Gratani and Varone, 2004).
Interestingly, the sites for which the model was more efficient to predict Reco and GPP showed the lowest predictive capability for WUE (Table 3, Figure S9). Geographically, we found decreasing WUE moving from southern, Mediterranean sites to northern, mountain sites, together with the goodness of fit of our model (Table 3). This may be explained by the degree of adaptation of the vegetation to average higher condition of dryness as observed at the Mediterranean sites, with the evolution of water-stress-avoiding morphological features, like the thickening of the leaf cuticle characterizing sclerophyll leaves. WUE displayed the lowest R2 in the alpine sites, while southern sites showed the highest R2. These results confirm our hypothesis that WUE is an important indicator of plants' adaptation to drought stress, with “water saving” ecosystems (especially maquis and Holm Oak forests) able to sustain their physiological activity unaltered under drought conditions. The R2 of the models in the southern part of Italy were generally higher in the dry year (Collelongo and Capo Caccia) or comparable between the 2 years (Castelporziano). Collelongo did not show water limitations, the reason for its high R2 could lie in the vegetation characteristics. In fact, Collelongo forest is dominated by Fagus sylvatica and it was previously demonstrated that this species has the capability to recover after drought periods, maintaining also a low stomatal conductance, thus increasing the WUE (Gall and Feller, 2007).
Multiple effects of environmental variables on GPP, Reco, WUE, and ET were evident in Figure 3 where the same variables appeared both instantaneously and with delays. In fact, by using only instantaneous variables, the models showed low R2 suggesting that a delayed effect on these biological processes was missing (data not shown). Instead, the implementation of the dataset with lagged variables contributed to the increase of the adjusted R2. Such evidence gave us the opportunity to assess which environmental factor affected—instantaneously or delayed—the vegetation ecophysiological processes. The importance of these variables was first observed in the factor loadings of the PCA (see Tables S5–S8). These variables, such as the daily-lagged precipitation showed high correlation (up to r = 0.89) with one of the selected Principal Components being representative of the whole component. In some cases, the PC representative of these lagged variables resulted to affect the dependent variable more than the other predictors, as in Capo Caccia, where the lagged air temperature affected GPP more than the other variables in both 2013 and 2014. The same was observed for Collelongo, where a 24 h-delayed variable affected GPP more than the other predictors in 2013.
Site Specific Differences Between the Dry and the Wet Year
GPP
At all our sites, both in dry and wet years, we observed that Tair was the most important factor affecting GPP, with a coefficient of impact up to 70% with few exceptions (Figure 3). As expected favorable growing temperatures promote higher GPP level as amply demonstrated in the last decades for the northern hemisphere (Niu et al., 2011; Wu et al., 2016). Negative impacts on GPP down to 25% were revealed by RH especially for drought-prone sites such as Capocaccia and Castelporziano. Such sites also displayed a positive delayed effect of RH. This behavior could be related to rainy events that even if they instantaneously reduce photosynthesis, they recharge the water table which in turn may sustain the photosynthetic process in a later period (Huxman et al., 2004; Ogle and Reynolds, 2004). By comparing dry and wet years in Castelporziano, a holm oak forest, which is a successional evolution of the maquis forest present in Capo Caccia, there were marked differences between the third most important variables which are RH in the dry year, and 24-h delayed RAD in the wet year, were observed. This pattern can be easily explained, since small changes in relative humidity lead to a fast response in GPP from Quercus ilex under dry climate conditions as already reported for this site by Savi and Fares (2014). Conversely, under wet conditions one of the most important limiting factors for carbon assimilation is solar radiation. This delayed behavior observed in drought-prone ecosystems is interpreted as the delayed response of the respective biochemical and physio-chemical processes in leaves and in the soil thus affecting carbon assimilation. Several authors indeed found delayed physiological responses by plants after rainfall events. Sala and Lauenroth (1982) observed an increase in leaf water potential and conductance within 12 h after the simulation of a small rainfall in a semi-arid grassland ecosystem. Schwinning et al. (2003) observed a 3-days delayed response by Hilaria jamesii after watering. These delays were associated with physiological acclimation and the growth of new roots and leaves (Huxman et al., 2004). Such delays should not be interpreted as a response to an isolated event, but could also be caused by prolonged exposure to changing conditions. For this reason, delayed variables assessing relevant factors and processes allow a more complete view of the ecosystem responses to environmental changes.
Concerning ecosystems with “water spending” strategies at higher latitudes, Collelongo showed differences in the third variable affecting GPP: while RH became more important than the RAD in the dry year, the latter was more relevant in the wet year, when RH is no more a limiting factor. A similar pattern was observed in Renon, where Tair and RAD had a larger impact on GPP in both years while only in the dry year variables related to water availability such as RH lag1 and SWC had a significant impact to GPP. Finally, both Torgnon forest and Torgnon grassland did not show strong differences between the 2 years perhaps because in the dry year, even if precipitation were lower than 2014, these are not enough to represent limiting conditions for GPP in these environments, where ecosystems benefit from the high water input derived from snowmelt at the beginning of the growing season. Moreover, European larch (Torgnon forest) has been considered from different studies as a anisohydric species (Anfodillo et al., 1998; Leo et al., 2014), especially when growing at high (i.e., higher than 2,000 m asl) altitudes, where a severe reduction of soil water content is less probable (Obojes et al., 2018).
Reco
The ecosystems present along the latitudinal gradient showed an increasing sensitivity of Reco to temperature, moving from the plain to mountain top, confirming the findings from Rodeghiero and Cescatti (2005). To different extents, all these ecosystems depend on concurrent carbon assimilation and soil water availability and, more specifically on the presence of rain pulses, the effect possibly combined with that of temperature (He and Callaway, 2009). When soils are relatively dry, metabolic activity was shown to increase strongly with water availability, while there is a broad range of near optimum soil water content where changes in soil moisture only have little effect, if any, on soil respiration (Migliavacca et al., 2011a).
Concerning temporal variability in Reco (Figure 3), temperatures (Tair and Tsoil) played a dominant role in all the sites, in both the dry and wet years, since temperature is one of the main drivers for soil microbial emissions (Shao et al., 2013). Precipitation and soil water content affected Reco more in the dry year than in the wet year, as shown by Renon, Torgnon forest, Castelporziano and Capo Caccia. These variables are important for soil respiration because they control microbial activity and all the related processes (Yan et al., 2015). The pivotal role of precipitation in the dry year could be connected to the Birch effect (Birch, 1958) that identifies pulse in soil respiration caused by the wetting of dry soils. This effect is particularly relevant in Mediterranean sites, as observed by Jarvis et al. (2007).
Interestingly, we observed that in the southern sites, Tair had an impact 5-times higher than the other variables on Reco. Instead in the northern sites, the RH played an important role, with an impact 0.5 times lower than Tair. We hypothesize that this difference could be explained by the observable gap in average water availability between the Northern and the Southern sites (wetter and drier, respectively). In support to our finding, (Gunti-as et al., 2013) observed that the sensitivity of soil respiration to temperature is lower at field capacity than under drier conditions. They have formulated two hypotheses to explain this behavior: (1) the labile component of organic matter is more sensitive to temperature than the recalcitrant one; (2) the rewetting of soil combined with high temperatures can lead to higher CO2 emissions.
Lagged responses were previously observed by Chen et al. (2009) for both GPP and Reco. Furthermore, (Unger et al., 2012) observed a 3-day delayed response after rain events in a Mediterranean ecosystem and associated this delay to the second phase of the Birch effect. Finally, Ruehr et al. (2010) observed a strong correlation between the soil respiration and the soil temperatures delayed by 2–4 h. In our study however, lagged responses in Reco were less pronounced compared to GPP, and only appreciated in the sites of Collelongo and Torgnon.
WUE
Consistent among sites, we observed an increase in WUE moving from the dry to the wet year, in agreement with Limousin et al. (2015) who found convergence in WUE responses to drought between drought-tolerant and less drought-tolerant plants. This can be partly counterintuitive, since plant trait plasticity would move in the opposite direction. A similar increase in WUE at the ecosystem scale with increasing precipitation, in contrast with findings from leaf-physiological studies, was found also by Sun et al. (2016). Two arguments can be raised to explain these features. Firstly, in the wet year the average VPD is lower. Wang et al. (2018) observed that the VPD, in addition to the increasing CO2 concentration in the air, can partly explain the increasing trend in WUE as observed in several other studies (Keenan et al., 2013; Giammarchi et al., 2017). Additionally, an often neglected effect in the same direction is given by the different ratio between direct and diffuse radiation that decreases during rainy periods. Diffuse light typically favors assimilation, and to a minor extent evapotranspiration in the presence of atmospheric aerosol (Steiner et al., 2013), therefore, we suggest that higher diffuse/direct radiation fraction may have favored WUE in the wet year as experimentally found by Brodersen et al. (2008). Previous research in alpine sites, including Renon, also demonstrated that the increased WUE in the wet years is a clear effect of the reduction in radiation and VPD. These sites are evidently switching from 1 year to the next according to radiation condition change (Ryu et al., 2008; Teuling et al., 2009). In agreement with these studies, radiation had a higher effect on ET in wet years at the Alpine sites (Figure 3).
In Torgnon forest and grassland sites, WUE did not change markedly between the 2 years of study. This behavior suggests that water availability in the dry year could have been enough to satisfy the vegetation requirements at the “water spending” ecosystems of northern latitudes. Interestingly, a delayed Tair had a negative impact on WUE in Capo Caccia, thus suggesting that extended exposure to higher growing temperature may have led to an increase of evapotranspiration (ET) with unbalanced water loss with respect to carbon assimilation (Trepekli et al., 2016). In general, lagged variables did not play a significant impact in our predictive models, except for at Castelporziano where lagged radiation effect had a positive impact on WUE, as also previously found (Boese et al., 2017), thus suggesting that prolonged periods of high solar radiation, or even episodes of cloud cover and diffuse radiation have a delayed positive impact on WUE in both the wet and dry year. This may represent evidence of an adaptive capacity of this drought-prone ecosystem.
Conclusions
In this study, we presented results of a mixed statistical analysis applied to eddy covariance data from six experimental sites in order to evaluate how changes in precipitation could affect could affect ecosystem processes. Data showed different responses to precipitation regimes, with marked differences along the latitude gradient (i.e., comparing results from northern and southern sites) with general increases of GPP, Reco, WUE and ET during wet years (hypothesis i).
Results of the multivariate analysis also indicated that changes in the limiting factors with latitude and among wet and dry years exist (hypothesis ii). Southern Mediterranean sites are indeed more adapted to water stress occurring regularly during the dry and hot summer months. For that reason, and also for a reduced meteorological variability between the considered years, we found minor differences in ecophysiological responses between a dry and a wet year in these sites compared with northern sites. This behavior was accurately described by predictive models that produced similar R2 between the 2 years of study in southern sites. The best performance for prediction of WUE was observed in southern sites under the assumption of a more resilient response to meteorological variables changing between wet and dry years, in accordance with the hypothesis (ii). By contrast, predictive models for GPP and Reco performed best in sites characterized by high precipitation regimes.
Multivariate analysis also identified joint changes in temperature regimes and rapid increases in VPD as a major challenge for Mediterranean Ecosystems. Net differences were observed between coastal and mountain sites, with GPP in coastal sites being mainly affected by relative humidity, while the mountain sites are affected more by global radiation. The inclusion of lagged variables in the dataset contributed significantly to the increase of the goodness-of-fit of regression models. The identification of time lags, mainly for GPP and Reco, indicated that environmental changes exert not only an instantaneous effect on these variables, but they can have delayed effects shaping vegetation response (hypothesis iii). This evidence suggests that delayed effects, even on short-time windows, should be taken into account in order to better understand the multifaceted relationships between environmental variables and biological processes.
Author Contributions
SF coordinated the work and contributed to drafting. AC is the first author and primarily worked on drafting and editing of Figures and Tables. LS responsible for statistical analysis, all other coauthors equally contributed with their field data and with manuscript drafting.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We acknowledge the General Secretariat of the Presidency of Italian Republic for financing the CASTEL4 project, and the Directorate of Castelporziano Estate. We also acknowledge the E-OBS climate indices dataset from the EU-FP7 project EUPORIAS (http://www.euporias.eu) and the data providers in the ECA&D project (http://www.ecad.eu). We also want to acknowledge Dr. Andrew Francis Speak for his careful correction of the English form.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2019.00016/full#supplementary-material
References
Anfodillo, T., Rento, S., Carraro, V., Furlanetto, L., Urbinati, C., and Carrer, M. (1998). Tree water relations and climatic variations at the alpine timberline: seasonal changes of sap flux and xylem water potential in Larix decidua Miller, Picea abies (L.) Karst. and Pinus cembra L. Annales des Sci. Forest. 55, 159–172. doi: 10.1051/forest:19980110
Aubinet, M., Feigenwinter, C., Heinesch, B., Bernhofer, C., Canepa, E., Lindroth, A., et al. (2010). Direct advection measurements do not help to solve the night-time CO2 closure problem: evidence from three different forests. Agric. For. Meteorol. 150, 655–664. doi: 10.1016/j.agrformet.2010.01.016
Aubinet, M., Vesala, T., and Papale, D. (eds) (2012). Eddy Covariance: A Practical Guide to Measurement and Data Analysis. Springer. doi: 10.1007/978-94-007-2351-1
Bhargava, S., and Sawant, K. (2013). Drought stress adaptation: metabolic adjustment and regulation of gene expression. Plant Breed. 132, 21–32. doi: 10.1111/pbr.12004
Birch, H. F. (1958). The effect of soil drying on humus decomposition and nitrogen availability. Plant Soil 10, 9–31. doi: 10.1007/BF01343734
Boese, S., Jung, M., Carvalhais, N., and Reichstein, M. (2017). The importance of radiation for semiempirical water-use efficiency models. Biogeosciences 14, 3015–3026. doi: 10.5194/bg-14-3015-2017
Bréda, N., Huc, R., Granier, A., and Dreyer, E. (2006). Temperate forest trees and stands under severe drought: a review of ecophysiological responses, adaptation processes and long-term consequences. Ann. For. Sci. 63, 625–644. doi: 10.1051/forest:2006042
Brodersen, C. R., Vogelmann, T. C., Williams, W. E., and Gorton, H. L. (2008). A new paradigm in leaf-level photosynthesis: direct and diffuse lights are not equal. Plant Cell Environ. 31, 159–164.
Canadell, J., and Zedler, P. H. (1995). “Underground structures of woody plants in Mediterranean ecosystems of Australia, California, and Chile,” in Ecology and Biogeography of Mediterranean Ecosystems in Chile, California, and Australia. Ecological Studies (Analysis and Synthesis), Vol. 108, eds M. T. K. Arroyo, P. H. Zedler, M. D. Fox (New York, NY: Springer), 177–210. doi: 10.1007/978-1-4612-2490-7_8
Chelli, S., Wellstein, C., Campetella, G., Canullo, R., Tonin, R., Gerdol, S., et al. (2017). Climate change response of vegetation across climatic zones in Italy. Clim. Res. 71, 249–262. doi: 10.3354/cr01443
Chen, S., Lin, G., Huang, J., and Jenerette, G. D. (2009). Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe. Glob. Chang. Biol. 15, 2450–2461. doi: 10.1111/j.1365-2486.2009.01879.x
Colantoni, A., Grigoriadis, E., Sateriano, A., Venanzoni, G., and Salvati, L. (2016). Cities as selective land predators? A lesson on urban growth, deregulated planning and sprawl containment. Sci. Total Environ. 545, 329–339. doi: 10.1016/j.scitotenv.2015.11.170
Detto, M., Bohrer, G., Nietz, J. G., Maurer, K. D., Vogel, C. S., Gough, C. M., et al. (2013). Multivariate conditional Granger causality analysis for lagged response of soil respiration in a temperate forest. Entropy 15, 4266–4284. doi: 10.3390/e15104266
Donovan, L. A., and Ehleringer, J. R. (1991). Ecophysiological differences among juvenile and reproductive plants of several woody species. Oecologia 86, 594–597. doi: 10.1007/BF00318327
Duvernoy, I., Zambon, I., Sateriano, A., and Salvati, L. (2018). Pictures from the other side of the fringe: urban growth and peri-urban agriculture in a post-industrial city (Toulouse, France). J. Rural Stud. 57, 25–35. doi: 10.1016/j.jrurstud.2017.10.007
Fares, S., Conte, A., and Chabbi, A. (2018). Ozone flux in plant ecosystems: new opportunities for long-term monitoring networks to deliver ozone-risk assessments. Environm. Sci. Pollution Res. 25, 8240–8248. doi: 10.1007/s11356-017-0352-0
Fares, S., Savi, F., Muller, J., Matteucci, G., and Paoletti, E. (2014). Simultaneous measurements of above and below canopy ozone fluxes help partitioning ozone deposition between its various sinks in a Mediterranean Oak Forest. Agric. Forest Meteorol. 198, 181–191. doi: 10.1016/j.agrformet.2014.08.014
Flexas, J., Galmes, J., Ribas-Carbo, M., and Medrano, H. (2005). “The effects of water stress on plant respiration,” in Plant Respiration. Advances in Photosynthesis and Respiration, Vol 18, eds H. Lambers and M. Ribas-Carbo (Dordrecht: Springer), 85–94. doi: 10.1007/1-4020-3589-6_6
Gall,é, A., and Feller, U. (2007). Changes of photosynthetic traits in beech saplings (Fagus sylvatica) under severe drought stress and during recovery. Physiol. Plant. 131, 412–421. doi: 10.1111/j.1399-3054.2007.00972.x
Galvagno, M., Wohlfahrt, G., Cremonese, E., Rossini, M., Colombo, R., Filippa, G., et al. (2013). Phenology and carbon dioxide source/sink strength of a subalpine grassland in response to an exceptionally short snow season. Environ. Res. Lett. 8:025008. doi: 10.1088/1748-9326/8/2/025008
Giammarchi, F., Cherubini, P., Pretzsch, H., and Tonon, G. (2017). The increase of atmospheric CO 2 affects growth potential and intrinsic water-use efficiency of Norway spruce forests: insights from a multi-stable isotope analysis in tree rings of two Alpine chronosequences. Trees 31, 503–515. doi: 10.1007/s00468-016-1478-2
Giorgi, F., and Lionello, P. (2008). Climate change projections for the mediterranean region. Glob. Planet. Change 63, 90–104. doi: 10.1016/j.gloplacha.2007.09.005
Gobiet, A., Kotlarski, S., Beniston, M., Heinrich, G., Rajczak, J., and Stoffel, M. (2014). 21st century climate change in the European Alps-a review. Sci. Total Environ. 493, 1138–1151. doi: 10.1016/j.scitotenv.2013.07.050
Gouveia, C. M., Trigo, R. M., Beguería, S., and Vicente-Serrano, S. M. (2017). Drought impacts on vegetation activity in the Mediterranean region: an assessment using remote sensing data and multi-scale drought indicators. Glob. Planet. Change 151, 15–27. doi: 10.1016/j.gloplacha.2016.06.011
Gratani, L., and Varone, L. (2004). Adaptive photosynthetic strategies of the Mediterranean maquis species according to their origin. Photosynthetica 42, 551–558. doi: 10.1007/S11099-005-0012-3
Griffin, K. L., and Prager, C. M. (2017). Where does the carbon go? Thermal acclimation of respiration and increased photosynthesis in trees at the temperate-boreal ecotone. Tree Physiol. 37, 281–284. doi: 10.1093/treephys/tpw133
Gunti-as, M. E., Gil-Sotres, F., Leirós, M. C., and Trasar-Cepeda, C. (2013). Sensitivity of soil respiration to moisture and temperature. J. Soil Sci. Plant Nutri. 13, 445–461. doi: 10.4067/S0718-95162013005000035
Hacke, U. G., Sperry, J. S., Pockman, W. T., Davis, S. D., and McCulloh, K. A. (2001). Trends in wood density and structure are linked to prevention of xylem implosion by negative pressure. Oecologia 126, 457–461. doi: 10.1007/s004420100628
Haylock, M. R., Hofstra, N., Klein Tank, A. M. G., Klok, E. J., Jones, P. D., and New, M. (2008). A European daily high-resolution gridded data set of surface temperature and precipitation for 1950-2006. J. Geophys. Res. 113:D20119. doi: 10.1029/2008JD010201
He, W.-M., and Callaway, R. M., (2009). The potential for misleading correlations in single-factor analysis of complex gradients. Web Ecol. 9, 77–81. doi: 10.5194/we-9-77-2009
Huang, L., He, B., Han, L., Liu, J., Wang, H., and Chen, Z. (2017). A global examination of the response of ecosystem water-use efficiency to drought based on MODIS data. Sci. Total Environ. 601, 1097–1107. doi: 10.1016/j.scitotenv.2017.05.084
Huxman, T. E., Snyder, K. A., Tissue, D., Leffler, A. J., Ogle, K., Pockman, W. T., et al. (2004). Precipitation pulses and carbon fluxes in semiarid and arid ecosystems. Oecologia 141, 254–268. doi: 10.1007/s00442-004-1682-4
Incerti, G., Feoli, E., Salvati, L., Brunetti, A., and Giovacchini, A. (2007). Analysis of bioclimatic time series and their neural network-based classification to characterise drought risk patterns in South Italy. Int. J. Biometeorol. 51, 253–263. doi: 10.1007/s00484-006-0071-6
Inglima, I., Alberti, G., Bertolini, T., Vaccari, F. P., Gioli, B., Miglietta, F., et al. (2009). Precipitation pulses enhance respiration of Mediterranean ecosystems: the balance between organic and inorganic components of increased soil CO2 efflux. Glob. Chang. Biol. 15, 1289–1301. doi: 10.1111/j.1365-2486.2008.01793.x
Jarvis, P., Rey, A., Petsikos, C., Wingate, L., Rayment, M., Pereira, J., et al. (2007). Drying and wetting of Mediterranean soils stimulates decomposition and carbon dioxide emission: the “Birch effect”. Tree Physiol. 27, 929–940. doi: 10.1093/treephys/27.7.929
Jensen, K. D., Beier, C., Michelsen, A., and Emmett, B. A. (2003). Effects of experimental drought on microbial processes in two temperate heathlands at contrasting water conditions. Appl. Soil Ecol. 24, 165–176. doi: 10.1016/S0929-1393(03)00091-X
Keenan, T. F., Hollinger, D. Y., Bohrer, G., Dragoni, D., Munger, J. W., Richardson, A. D., et al. (2013). Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 499, 324–327. doi: 10.1038/nature12291
Kottek, M., Grieser, J., Beck, C., Rudolf, B., and Rubel, F. (2006). World map of the Köppen-Geiger climate classification updated. Meteorol. Zeitsch. 15, 259–263. doi: 10.1127/0941-2948/2006/0130
Leo, M., Oberhuber, W., Schuster, R., Grams, T. E., Matyssek, R., and Wieser, G. (2014). Evaluating the effect of plant water availability on inner alpine coniferous trees based on sap flow measurements. Eur. J. For. Res. 133, 691–698. doi: 10.1007/s10342-013-0697-y
Limousin, J. M., Yepez, E. A., McDowell, N. G., and Pockman, W. T. (2015). Convergence in resource use efficiency across trees with differing hydraulic strategies in response to ecosystem precipitation manipulation. Funct. Ecol. 29, 1125–1136. doi: 10.1111/1365-2435.12426
Maesano, M., Matteucci, G., Corona, P., and Scarascia Mugnozza, G. (2017). Ecosystem Services in Mountain Forests: The Interaction of Carbon and Water Balance in a Beech Stand in Central Italy. Mountain Watersheds and Ecosystem Services: Balancing multiple demands of forest management in head-watersheds. 53.
Marras, S., Pyles, R. D., Sirca, C., Snyder, R. L., Duce, P., and Spano, D. (2011). Evaluation of the advanced canopy-atmosphere-soil algorithm (ACASA) model performance over Mediterranean maquis ecosystem. Agric. Forest Meteorol. 151, 730–745. doi: 10.1016/j.agrformet.2011.02.004
Mazzenga, F. (2017). Analisi di Lungo Termine sui fattori di Controllo Dello Scambio di Carbonio in una Faggeta dell'Italia Centromeridionale. Doctoral dissertation, PhD Thesis, DIBAF, University of Tuscia, Viterbo, Italy.
Migliavacca, M., Cremonese, E., Colombo, R., Busetto, L., Galvagno, M., Ganis, L., et al. (2008). European larch phenology in the Alps: can we grasp the role of ecological factors by combining field observations and inverse modelling? Int. J. Biometeorol. 52, 587–605. doi: 10.1007/s00484-008-0152-9
Migliavacca, M., Galvagno, M., Cremonese, E., Rossini, M., Meroni, M., Sonnentag, O., et al. (2011b). Using digital repeat photography and eddy covariance data to model grassland phenology and photosynthetic CO2 uptake. Agric. Forest Meteorol. 151, 1325–1337. doi: 10.1016/j.agrformet.2011.05.012
Migliavacca, M., Reichstein, M., Richardson, A. D., Colombo, R., Sutton, M. A., Michiel, G., et al. (2011a). Semiempirical modeling of abiotic and biotic factors controlling ecosystem respiration across eddy covariance sites. Glob. Chang. Biol. 17, 390–409. doi: 10.1111/j.1365-2486.2010.02243.x
Montagnani, L., Manca, G., Canepa, E., Georgieva, E., Acosta, M., Feigenwinter, C., et al. (2009). A new mass conservation approach to the study of CO2 advection in an alpine forest. J. Geophys. Res. 114:D07306. doi: 10.1029/2008JD010650
Montagnani, L., Zanotelli, D., Tagliavini, M., and Tomelleri, E. (2018). Timescale effects on the environmental control of carbon and water fluxes of an apple orchard. Ecol. Evol. 8, 416–434. doi: 10.1002/ece3.3633
Nestola, E., Sánchez-Zapero, J., Latorre, C., Mazzenga, F., Matteucci, G., Calfapietra, C., et al. (2017). Validation of PROBA-V GEOV1 and MODIS C5 and C6 fAPAR Products in a deciduous beech forest site in Italy. Remote Sens. 9:126. doi: 10.3390/rs9020126
NIMBUS (2015). Available online at: http://www.nimbus.it/clima/2015/150114clima2014.htm. (accessed September 26, 2017).
Niu, S., Luo, Y., Fei, S., Montagnani, L., Bohrer, G., Janssens, I. A., et al. (2011). Seasonal hysteresis of net ecosystem exchange in response to temperature change: patterns and causes. Glob. Chang. Biol. 17, 3102–3114. doi: 10.1111/j.1365-2486.2011.02459.x
Obojes, N., Meurer, A., Newesely, C., Tasser, E., Oberhuber, W., Mayr, S., et al. (2018). Water stress limits transpiration and growth of European larch up to the lower subalpine belt in an inner-alpine dry valley. N. Phytol. 220, 460–475. doi: 10.1111/nph.15348
Ogle, K., and Reynolds, J. F. (2004). Plant responses to precipitation in desert ecosystems: integrating functional types, pulses, thresholds, and delays. Oecologia 141, 282–294. doi: 10.1007/s00442-004-1507-5
Osakabe, Y., Osakabe, K., Shinozaki, K., and Tran, L. S. P. (2014). Response of plants to water stress. Front. Plant Sci. 5:86. doi: 10.3389/fpls.2014.00086
Paeth, H., Vogt, G., Paxian, A., Hertig, E., Seubert, S., and Jacobeit, J. (2017). Quantifying the evidence of climate change in the light of uncertainty exemplified by the Mediterranean hot spot region. Glob. Planet. Change 151, 144–151. doi: 10.1016/j.gloplacha.2016.03.003
Pe-uelas, J., Canadell, J. G., and Ogaya, R. (2011). Increased water-use efficiency during the 20th century did not translate into enhanced tree growth. Glob. Ecol. Biogeogr. 20, 597–608. doi: 10.1111/j.1466-8238.2010.00608.x
Pili, S., Grigoriadis, E., Carlucci, M., Clemente, M., and Salvati, L. (2017). Towards sustainable growth? A multi-criteria assessment of (changing) urban forms. Ecol. Indicat. 76, 71–80. doi: 10.1016/j.ecolind.2017.01.008
Ratna, S. B., Ratnam, J. V., Behera, S. K., Cherchi, A., Wang, W., and Yamagata, T. (2017). The unusual wet summer (July) of 2014 in Southern Europe. atmosphere. Res. 189, 61–68. doi: 10.1016/j.atmosres.2017.01.017
Reichstein, M., Falge, E., Baldocchi, D., Papale, D., Aubinet, M., Berbigier, P., et al. (2005). On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Glob. Chang. Biol. 11, 1424–1439. doi: 10.1111/j.1365-2486.2005.001002.x
Rodeghiero, M., and Cescatti, A. (2005). Main determinants of forest soil respiration along an elevation/temperature gradient in the Italian Alps. Glob. Change Biol. 11, 1024–1041. doi: 10.1111/j.1365-2486.2005.00963.x
Ruehr, N. K., Knohl, A., and Buchmann, N. (2010). Environmental variables controlling soil respiration on diurnal, seasonal and annual time-scales in a mixed mountain forest in Switzerland. Biogeochemistry 98, 153–170. doi: 10.1007/s10533-009-9383-z
Ryu, Y., Baldocchi, D. D., Ma, S., and Hehn, T. (2008). Interannual variability of evapotranspiration and energy exchange over an annual grassland in California. J. Geophys. Res. 113:D09104. doi: 10.1029/2007JD009263
Sala, O. E., and Lauenroth, W. K. (1982). Small rainfall events: an ecological role in semiarid regions. Oecologia 53, 301–304. doi: 10.1007/BF00389004
Salvati, L., Sabbi, A., and Carlucci, M. (2014). Exploring complex relationships using non-parametric principal components analysis: a case study with land-use data. Int. J. Ecol. Econ. Stat. 33, 90–97.
Salvati, L., and Zitti, M. (2005). Land degradation in the Mediterranean basin: linking bio-physical and economic factors into an ecological perspective. Biota 5, 67–77.
Savi, F., and Fares, S. (2014). Ozone dynamics in a Mediterranean Holm oak forest: comparison among transition periods characterized by different amounts of precipitation. Ann. Silvicul. Res. 38, 1–6. doi: 10.12899/asr-801 Available online at: https://journals-crea.4science.it/index.php/asr/article/view/801 (accessed April 25, 2019)
Schlesinger, W. H., Dietze, M. C., Jackson, R. B., Phillips, R. P., Rhoades, C. C., Rustad, L. E., et al. (2016). Forest biogeochemistry in response to drought. Glob. Chang. Biol. 22, 2318–2328. doi: 10.1111/gcb.13105
Schwinning, S., Starr, B. I., and Ehleringer, J. R. (2003). Dominant cold desert plants do not partition warm season precipitation by event size. Oecologia 136, 252–260. doi: 10.1007/s00442-003-1255-y
Shao, P., Zeng, X., Moore, D. J., and Zeng, X. (2013). Soil microbial respiration from observations and Earth System Models. Environ. Res. Lett. 8:034034. doi: 10.1088/1748-9326/8/3/034034
Steiner, A. L., Mermelstein, D., Cheng, S. J., Twine, T. E., and Oliphant, A. (2013). Observed impact of atmospheric aerosols on the surface energy budget. Earth Interac. 17, 1–22. doi: 10.1175/2013EI000523.1
Sun, Y. S., Piao, M., Huang, P., Ciais, Z., Zeng, L., Cheng, X., et al. (2016). Global patterns and climate drivers of water-use efficiency in terrestrial ecosystems deduced from satellite-based datasets and carbon cycle models. Glob. Ecol. Biogeogr. 25, 311–323. doi: 10.1111/geb.12411
Teuling, A. J., Hirschi, M., Ohmura, A., Wild, M., Reichstein, M., Seneviratne, P., et al. (2009). Regional radiation impacts on evapotranspiration trends. Geophys. Res. Lett. 36:L02404. doi: 10.1029/2008GL036584
Trepekli, A., Loupa, G., and Rapsomanikis, S. (2016). Seasonal evapotranspiration, energy fluxes and turbulence variance characteristics of a Mediterranean coastal grassland. Agric. Forest Meteorol. 226, 13–27. doi: 10.1016/j.agrformet.2016.05.006
Unger, S., Máguas, C., Pereira, J. S., David, T. S., and Werner, C. (2012). Interpreting post-drought rewetting effects on soil and ecosystem carbon dynamics in a Mediterranean oak savannah. Agric. Forest Meteorol. 154, 9–18. doi: 10.1016/j.agrformet.2011.10.007
Vilagrosa, A., Cortina, J., Gil-Pelegrín, E., and Bellot, J. (2003). Suitability of drought-preconditioning techniques in Mediterranean climate. Restor. Ecol. 11, 208–216. doi: 10.1046/j.1526-100X.2003.00172.x
Wang, M., Chen, Y., Wu, X., and Bai, Y. (2018). Forest-type-dependent water use efficiency trends across the northern hemisphere. Geophys. Res. Lett. 45, 8283–8293. doi: 10.1029/2018GL079093
Wu, X., Liu, H., Li, X., Liang, E., Beck, P. S., and Huang, Y. (2016). Seasonal divergence in the interannual responses of Northern Hemisphere vegetation activity to variations in diurnal climate. Sci. Rep. 6:19000. doi: 10.1038/srep19000
Wu, Z., Dijkstra, P., Koch, G. W., Pe-uelas, J., and Hungate, B. A. (2011). Responses of terrestrial ecosystems to temperature and precipitation change: a meta-analysis of experimental manipulation. Glob. Chang. Biol. 17, 927–942. doi: 10.1111/j.1365-2486.2010.02302.x
Yan, N., Marschner, P., Cao, W., Zuo, C., and Qin, W. (2015). Influence of salinity and water content on soil microorganisms. Int. Soil Water Conserv. Res. 3, 316–323. doi: 10.1016/j.iswcr.2015.11.003
Yu, G., Song, X., Wang, Q., Liu, Y., Guan, D., Yan, J., et al. (2008). Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables. N. Phytol. 177, 927–937. doi: 10.1111/j.1469-8137.2007.02316.x
Keywords: forest ecophysiology, GPP, WUE (water use efficiency), rainfall variability, carbon flux and partitioning, RECo2
Citation: Conte A, Fares S, Salvati L, Savi F, Matteucci G, Mazzenga F, Spano D, Sirca C, Marras S, Galvagno M, Cremonese E and Montagnani L (2019) Ecophysiological Responses to Rainfall Variability in Grassland and Forests Along a Latitudinal Gradient in Italy. Front. For. Glob. Change 2:16. doi: 10.3389/ffgc.2019.00016
Received: 23 November 2018; Accepted: 16 April 2019;
Published: 07 May 2019.
Edited by:
Iolanda Filella, Spanish National Research Council (CSIC), SpainReviewed by:
Paul Christopher Stoy, Montana State University, United StatesPenélope Serrano Ortiz, University of Granada, Spain
Copyright © 2019 Conte, Fares, Salvati, Savi, Matteucci, Mazzenga, Spano, Sirca, Marras, Galvagno, Cremonese and Montagnani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Silvano Fares, c2lsdmFuby5mYXJlc0BjcmVhLmdvdi5pdC4=