 J. Morgan Varner
J. Morgan Varner Jeffrey M. Kane
Jeffrey M. Kane Jesse K. Kreye3
Jesse K. Kreye3 Timothy M. Shearman
Timothy M. Shearman- 1Tall Timbers Research Station, Tallahassee, FL, United States
- 2Wildland Fire Laboratory, Department of Forestry and Wildland Resources, Humboldt State University, Arcata, CA, United States
- 3Department of Ecosystem Science and Management, The Pennsylvania State University, University Park, PA, United States
Widespread fire exclusion and land-use activities across many southeastern United States forested ecosystems have resulted in altered species composition and structure. These changes in composition and structure have been implicated in positive fire-vegetation feedbacks termed “mesophication” where fire spread and intensity are diminished. In forests and woodlands, inherent flammability of different species is the mechanistic driver of mesophication. To date, there has been limited work on documenting the high diversity of flammability among species in the region, limiting the ability to differentiate among species to restore fuels that sustain fire regimes. Here, we coalesce disparate flammability data and add missing species across the spectrum from species that facilitate fire (so called “pyrophytes”) to those that dampen fire (so called “mesophytes”). We present data on 50 important tree species from across the southeast, all burned using identical laboratory methods. We divide our results for four dominant ecosystems: Coastal Plain uplands, oak-hickory woodlands, Appalachian forests, and bottomland forests. Across ecosystems, the most flammable species were American chestnut (Castanea dentata), a suite of pines (Pinus palustris, P. elliottii, P. serotina, and P. rigida), several oaks (Q. laevis, Q. falcata, Q. margaretta, and Q. alba), and sourwood (Oxydendrum arboreum). At the mesophytic end, the least flammable species were Tsuga canadensis, Acer rubrum, and several other hardwoods previously implicated in mesophication. Each of the four ecosystems we studied contained species that spanned the pyrophytic to mesophytic gradient. These data fill in some key holes in our understanding of southeastern fire adaptations, but also provide context for restoration decisions and fire management prioritization efforts to restore and sustain fire-prone ecosystems of the region.
Introduction
Widespread fire exclusion and land-use change have resulted in dramatic impacts on species composition, structure, and ecological processes of many fire-prone terrestrial ecosystems. Many fire-adapted forested landscapes in North America, e.g., pine forests and oak woodlands, were cut over in the late 19th and early 20th century and fires were actively suppressed during recovery (MacCleery, 1993; Frost, 1998). The removal of fire from these ecosystems led to the establishment of fire-sensitive tree species into formerly fire-prone landscapes, altering species composition dramatically (Fei and Steiner, 2007; Nowacki and Abrams, 2008; Knott et al., 2019). The recruitment of fire-sensitive mesophytes coincided with increasing tree density, the formation of heavily shaded, multi-canopied forests, and development of moist forest floor conditions that resist ignition (Kreye et al., 2018a, b). This positive feedback phenomenon has been widely observed and is commonly referred to as “mesophication” (Nowacki and Abrams, 2008; Hanberry et al., 2020; Alexander et al., 2021).
Fires were very frequent across much of the southeastern United States (Stambaugh et al., 2011, 2015; Lafon et al., 2017) as reflected in the extent and diversity of fire-tolerant plant communities (Frost, 1998; Platt, 1999). In spite of the clear evidence of frequent fire and numerous species that are enhanced by fire, substantial gaps remain in the understanding of fire-related traits across tree species in the region. Protective traits such as thick bark and rapid wound closure allow fire-tolerant species to persist in fire-prone landscapes (Landers, 1991; Jackson et al., 1999; Romero et al., 2009; Varner et al., 2016). Flammability of forest litter is also a key component in fire-prone landscapes as it supports the fire regimes under which fire-tolerant or pyrophytic (hereafter simply pyrophyte) species persist (Kane et al., 2008). Recent studies have highlighted how flammable litter often corresponds with fire tolerance at the species level and may itself be a fire-adapted trait (Varner et al., 2016; Stevens et al., 2020; Varner et al., in review). Field measurements in forests, woodlands, and savannas validate the relative importance of senesced litter as the primary mechanistic driver of surface fire behavior (Wenk et al., 2011; Whelan et al., 2021).
Several studies have evaluated litter flammability of southeastern United States tree species. The majority of these studies follow standard methods as in Fonda (2001), burning single species in controlled environments (Fonda, 2001; Fonda and Varner, 2004; Kane et al., 2008, 2019, 2021; Mola et al., 2014; Varner et al., 2015a; Varner et al., in review). From these, the diverse southeastern tree flora is represented by only ca. 30 species, primarily pines and oaks. Several important tree species implicated in mesophication (e.g., Acer rubrum, Nyssa sylvatica, Tilia americana, and Ostrya virginiana) and other species thought to represent the pyrophytic end of the flammability spectrum (Carya spp. and other Quercus spp.) have yet to be evaluated. Beyond the missing species, there has been no synthetic compilation of these species.
In order to fill some of the gaps in our understanding of litter flammability of southeastern United States tree species, we conducted litter flammability experiments on a suite of species not yet studied and compiled them with published data from species examined using the same methods. The objectives of this study were to (1) synthesize litter flammability of ca. 50 southeastern tree species and (2) examine how litter flammability differs across the mesophyte-to-pyrophyte spectrum. We hypothesized that litter of pyrophytic species would be more flammable than mesophytic species. We tested this global hypothesis for each of four ecosystems where these species are important (Coastal Plain uplands, oak-hickory woodlands, Appalachian forests, and bottomland forests; Figure 1). Results are expected to elucidate flammability as a potential fire-adapted trait among pyrophytic species whereas dampened flammability among mesophytic species acts as a mechanism for the hypothesized mesophication of southeastern oak and pine forests resulting from decades of fire exclusion.
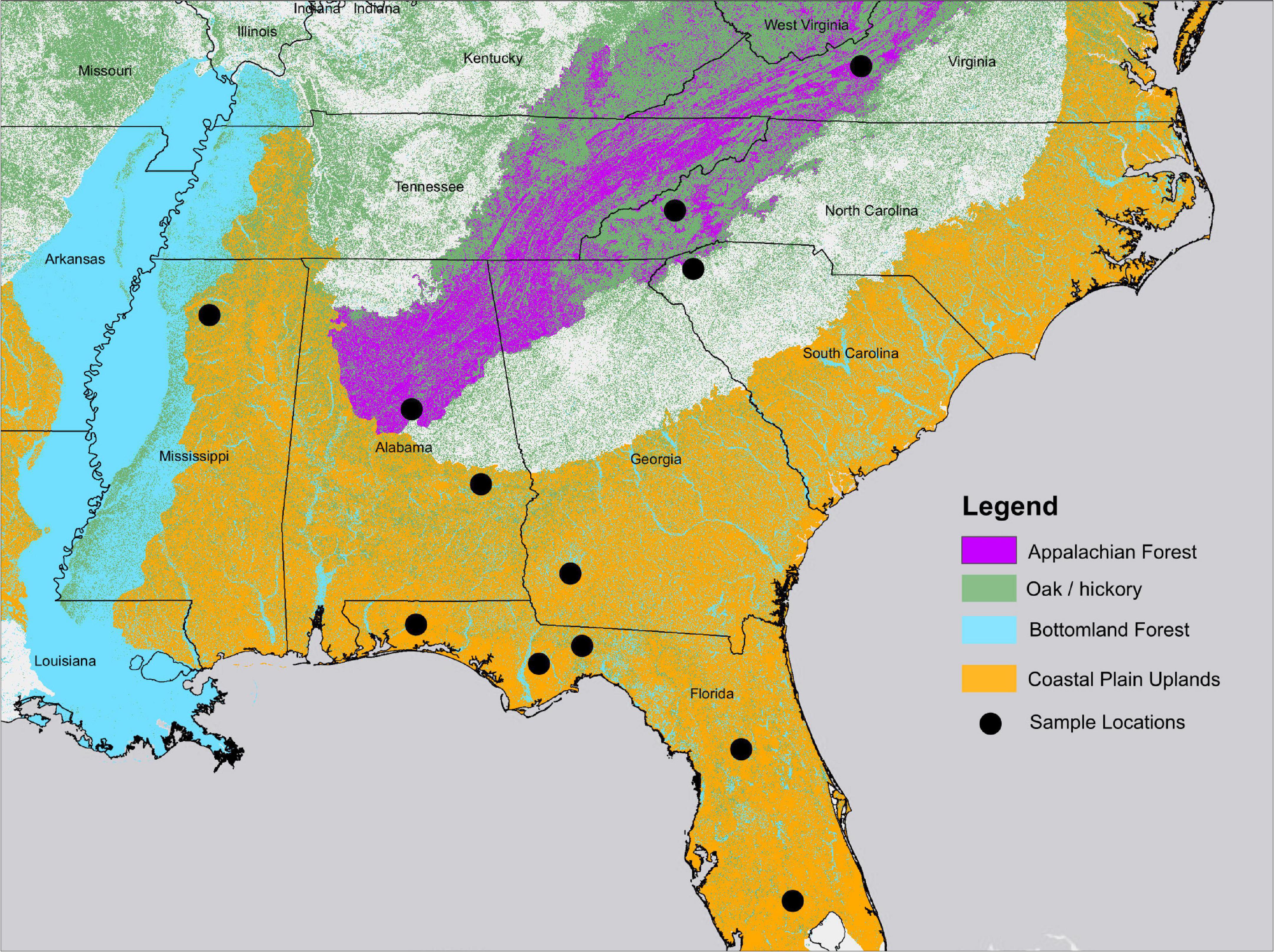
Figure 1. Geographic range of forest types and sample locations used in flammability analysis. Forest types were adapted from United States Forest Inventory and Analysis Forest Types and EPA Ecoregions (USEPA, 2013).
Materials and Methods
We combined published litter flammability data using the laboratory combustion methods of Fonda (2001) with new observations from recently collected litter from the region (Table 1 and Figure 1). All published and new litter flammability experiments were conducted using identical methods and in the same laboratory. For all species, we collected freshly fallen litter (i.e., no signs of decomposition; Oi horizon) beneath several individual trees, with each sample serving as an independent replicate. All collected litter was shipped to the Humboldt State University Wildland Fire Laboratory for burning.
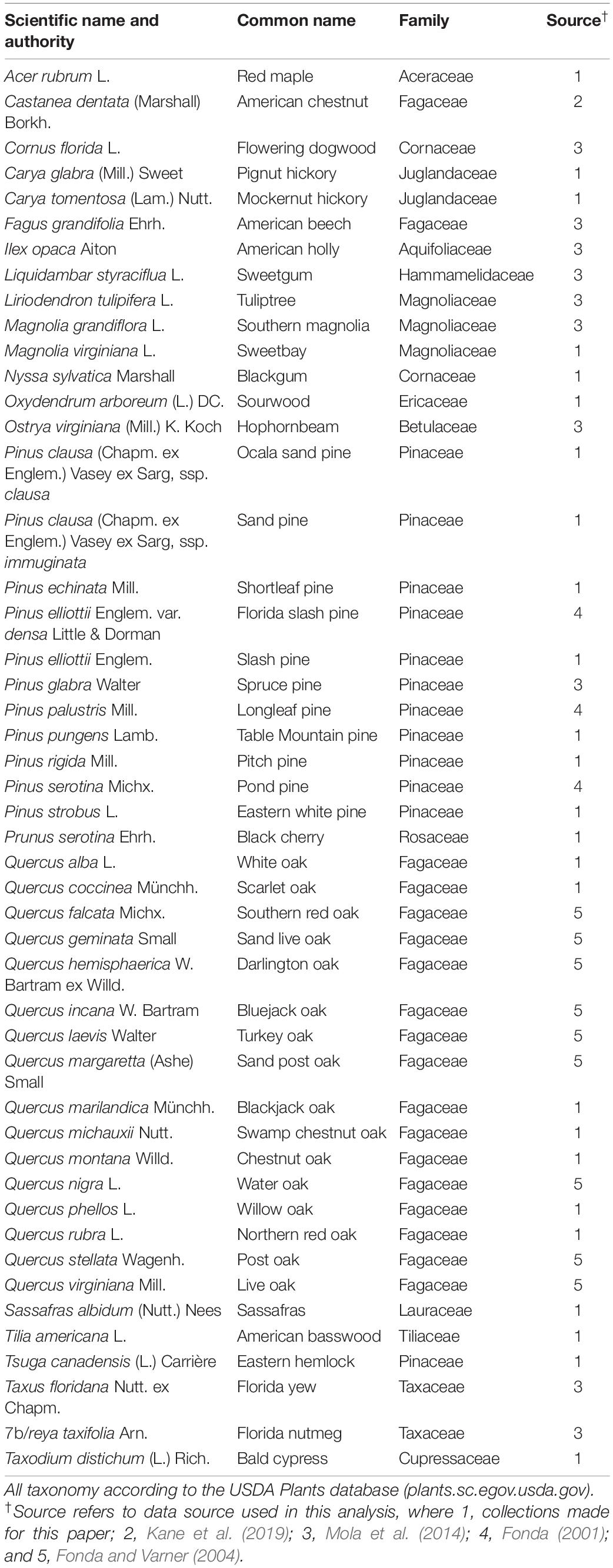
Table 1. List of all 50 southeastern tree species burned in these experiments.
Laboratory Burning
Litter flammability experiments, for each species separately, were conducted using established methods (Fonda, 2001). Litter samples were oven-dried at 50–60°C for at least 24 h until no further weight loss occurred. Each 15 g (dry weight) litter fuelbed was evenly distributed within a 35 cm × 35 cm lattice of xylene-soaked cotton strings on a stainless-steel platform. A 2.75 m × 2.75 m fume hood above the platform generated a constant draw of 15–20°cm s–1, though no airflow was detected at the fuelbed. Burn trials were repeated five to seven times for each species. Strings were ignited from all sides and a timer was started once litter ignited. Maximum flame height was visually estimated to the nearest cm using a vertical ruler placed behind the burning platform. Flaming time was measured as the duration from litter ignition to flaming extinction, and smoldering time was measured as the duration from flaming extinction to smoldering extinction (no visible smoke and no glowing combustion observed under darkened conditions). Both flaming time and smoldering time were recorded to the nearest second. Percent litter consumption was calculated as the proportion of the initial litter mass (15 g) that was consumed, measured following extinction of smoldering (and removing unburned string). We used flame height measurements to calculate fireline intensity (Byram, 1959) using the equation:
where I is fireline intensity (kW m–1) and h is flame height (m).
Data Analysis
Flammability metrics from all published and newly collected species were combined using Principal Components Analysis (PCA). PCA is used because flammability metrics are often highly correlated (e.g., species with rapid flaming times tend to burn with tall flames; Varner et al., 2015b). For all analyses, we first log transformed the four flammability metrics (flame height, flaming time, smoldering time, and percent consumed) to reduce the effect of outliers in the data. We standardized (mean = 0, SD = 1) four flammability metrics: flame height, flaming time, smoldering time, and percent consumed. Outliers in the transformed data were detected by calculating the maximum median absolute deviation for each species and principal component then using the departure from histogram method in the R package “bigutilsr” to identify outliers (Privé, 2021). Each PCA was re-fit after outliers were removed. Principal components that had eigenvalues greater than 1 and where cumulative variance explained was 70% were retained for further evaluation (Affifi and Clark, 1996). We then conducted a k-means cluster analysis to group species clusters of similar flammability. The number of clusters selected in the analyses was determined by plotting the within cluster sum of squares with increasing number of clusters and identifying the “bend” in the curve at which increasing the number of clusters does not markedly decrease the within cluster sum of squares. All analyses were conducted in R version 4.0.3 (R Core Team, 2020).
Given the number of species and the diversity of ecosystems in the region, we performed PCA for each ecosystem. The four ecosystems were: Coastal Plain uplands; oak-hickory woodlands; Appalachian forests; and bottomland forests. Coastal plain uplands span a wide longitudinal and latitudinal gradient in the region, from Texas to Virginia (and beyond along the Atlantic coastal plain) and are characterized by frequent fires (Stambaugh et al., 2011, 2017; Rother et al., 2020) on the order of 1–5 years in woodlands and savannas and by longer intervals (ca. 50–100 years) in pine-oak scrub forests (Menges, 1999). Oak-hickory woodlands are widespread and often linked with the central hardwood region. Fire regimes in these ecosystems are characterized by frequent surface fires ca. 2–10 years and longer (Hart and Buchanan, 2012; Stambaugh et al., 2015). Appalachian forests span from the mountains of northern Alabama and Georgia to the northern limits of the region, have high species diversity, and diverse fire regimes dependent on landscape position (Lafon et al., 2017). Bottomland forests have high overstory species diversity and follow major alluvial valleys and incised floodplains across the region. Fires in these ecosystems are rare, occurring perhaps on the order of 50–100 or more years between fires (Harper, 1911; Platt and Schwartz, 1990).
Results
Litter from the 50 southeastern tree species varied widely in their flammability. Notable among the most flammable species were the fire-tolerant pines (Pinus palustris, P. serotina, P. echinata, P. taeda, and P. rigida) and several oaks (Quercus laevis, Q. falcata, and Q. alba), American chestnut (Castanea dentata), and sourwood (Oxydendrum arboreum). The species with the least flammable litter were two endemic conifers (Torreya taxifolia and Taxus floridana), Tsuga canadensis, Pinus clausa var. clausa, and two evergreen oaks (Q. virginiana and Q. geminata). The classic mesophytes Acer rubrum, Tilia americana, Liquidambar styraciflua, and Nyssa sylvatica tended to burn with low flammability. We detail community-specific flammability below and list all species and their mean flammability values in Supplementary Table 1.
Coastal Plain Uplands
The 23 tree species in the coastal plain upland community spanned a wide flammability gradient (Figure 2). The first two principal components accounted for 82.8% of the variance, with axis 1 and 2 having eigenvalues of 2.28 and 1.03, respectively. Axis 1 was correlated with flame height (−0.53), and percent consumption (−0.63) as well as flaming time (0.53). Axis 2 was mostly correlated with smoldering time (−0.92). P. clausa var. clausa (Ocala sand pine) appeared to be an outlier with short smoldering time but was left in the analysis because it did not meet our criteria for removal. Fireline intensity ranged from 53 kW m–1 (Q. virginiana) to 419 kW m–1 (Pinus serotina). The k-means cluster analysis suggested four groups which explained approximately 80.3% of the variance of the ordination. A tall flaming, long smoldering group consisted of pines (P. palustris, P. elliottii var. elliottii and var. densa), oaks (Q. alba, Q. laevis, and Q. falcata), as well as Liquidambar styraciflua, Sassafras albidum, and Prunus serotina. A tall flaming, brief smoldering group also consisted of pines (P. echinata, P. serotina, and P. taeda), as well as Q. stellata, Cornus florida, and Carya tomentosa. An intermediate flaming, protracted smoldering group consisted mostly of oaks (Q. margaretta, Q. incana, Q. nigra, and Q. hemisphaerica) along with P. clausa var. immuginata. Lastly, a low fire intensity group consisted of P. clausa var. clausa, Q. virginiana, and Q. geminata (Figure 2).
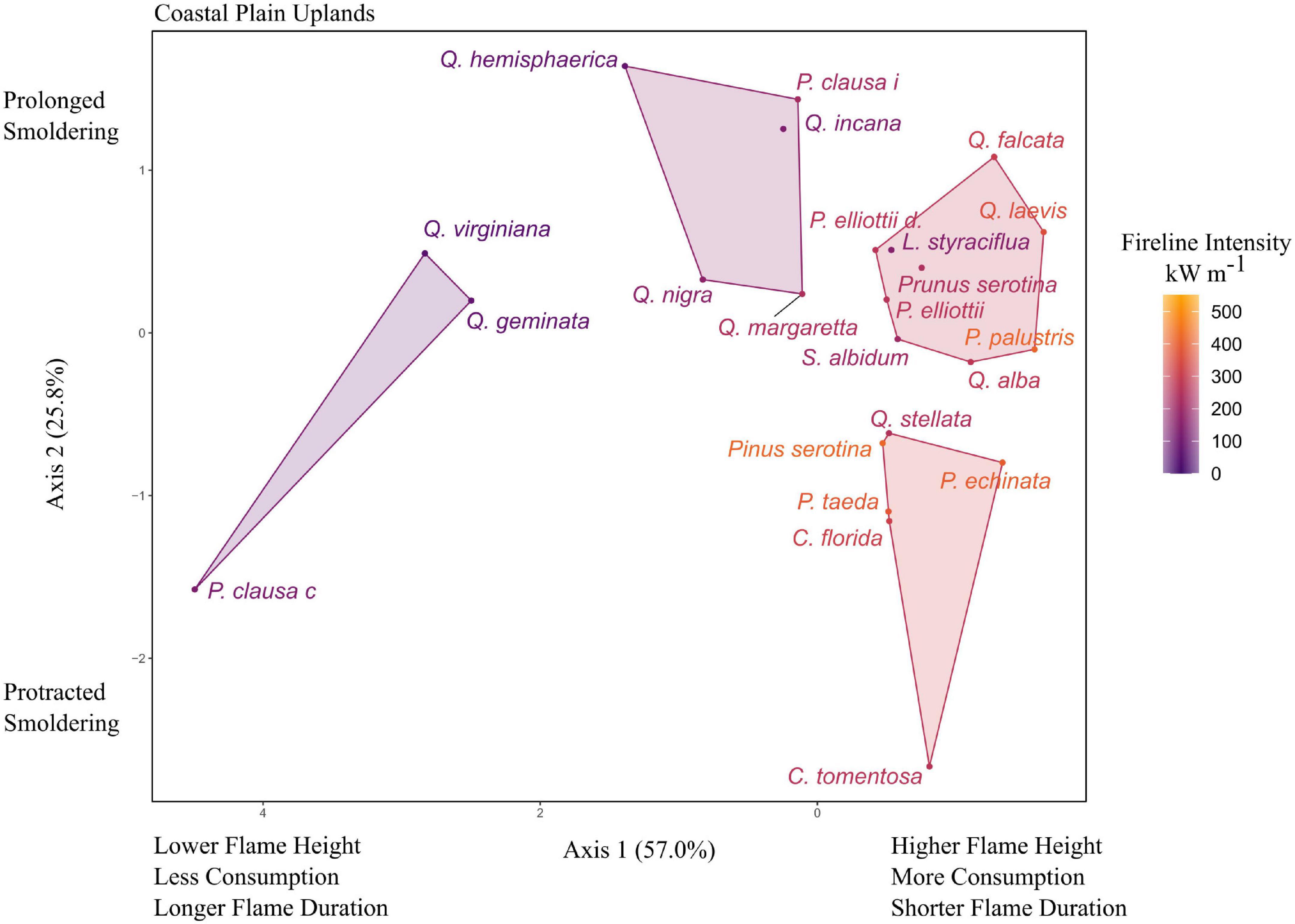
Figure 2. Principal components analysis (PCA) for flammability of tree litter in coastal plain uplands of the southeastern United States. Axis 1 was inverted to be consistent with the other figures (increased flame height and consumption on the right). Species codes and mean values listed in Table 1 and Supplementary Table 1.
Oak-Hickory Woodlands
The 21 tree species in the oak-hickory community also varied widely in flammability (Figure 3). The first two principal components accounted for 83.3% of the variance, with axes 1 and 2 having eigenvalues of 2.33 and 1.01, respectively. Axis 1 was correlated with flame height (0.59), flame duration (−0.55), and percent consumption (0.53). Axis 2 was primarily correlated with smoldering duration (−0.89). Fireline intensity ranged from 91 kW m–1 (O. virginiana) to 551 kW m–1 for Castanea dentata. The k-means cluster analysis again suggested four groups which explained approximately 67.5% of the variance of the ordination. The highest flaming group consisted of only two species: Castanea dentata and Oxydendrum arboreum. A slightly lower flaming, brief smoldering group contained P. echinata, L. tulipifera, F. grandifolia, Cornus florida, and two hickories (C. glabra and C. tomentosa). A second group with similar flaming characteristics as the previous but longer smoldering was made up of mostly oaks (Q. alba, Q. coccinea, Q. rubra, Q. montana, and Q. falcata) as well as Prunus serotina. Lastly, a lower flaming group consisted of O. virginiana, T. americana, A. rubrum, L. styraciflua, N. sylvatica, S. albidum, and Q. stellata (Figure 3).
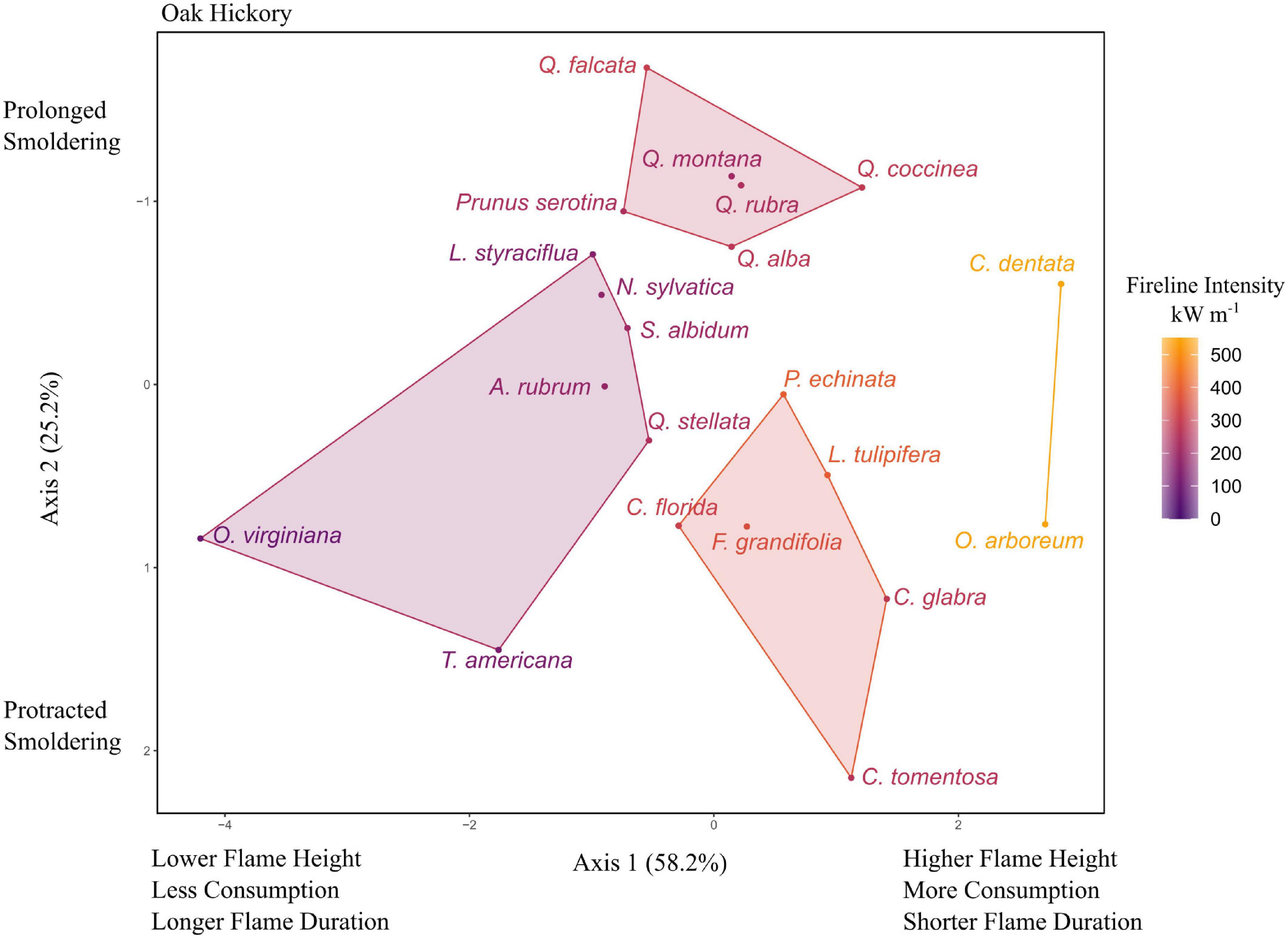
Figure 3. Principal components analysis (PCA) for flammability of tree litter in oak-hickory uplands of the southeastern United States. Axis 2 was inverted to be consistent with the other figures (increased smoldering on the top). Species codes and mean values listed in Table 1 and Supplementary Table 1.
Appalachian Forest
The 16 Appalachian forest trees had the widest range of flammability of the four ecosystems, including the least flammable Tsuga canadensis and the most flammable Castanea dentata. Tsuga canadensis was identified as an outlier even after the log transformation (likely due to having the lowest flame height and only 2.3% consumption) and was removed from the PCA. The first two principal components accounted for 86.6% of the variance, with axes 1 and 2 having eigenvalues of 2.19 and 1.27, respectively. Axis 1 was correlated with flame height (−0.50), flame duration (0.59) and percent consumption (−0.63). Axis 2 was again mostly correlated with smoldering time (0.84). Fireline intensity ranged from 3 kW m–1 (T. canadensis) to 546 kW m–1 (C. dentata). The k-means cluster analysis suggested three groups which explained 59.8% of the variance of the ordination. The highest flammability cluster contained two pines (P. echinata and P. rigida), C. dentata, O. arboreum, Q. coccinea, Q. alba, and Cornus florida. A lower flammability group with longer smoldering contained Q. montana, Q. rubra, Prunus serotina, Pinus pungens, Pinus virginiana, Sassafras albidum, A. rubrum, N. sylvatica, and T. americana. Pinus strobus was a singleton in a cluster with low flames and brief smoldering (Figure 4).
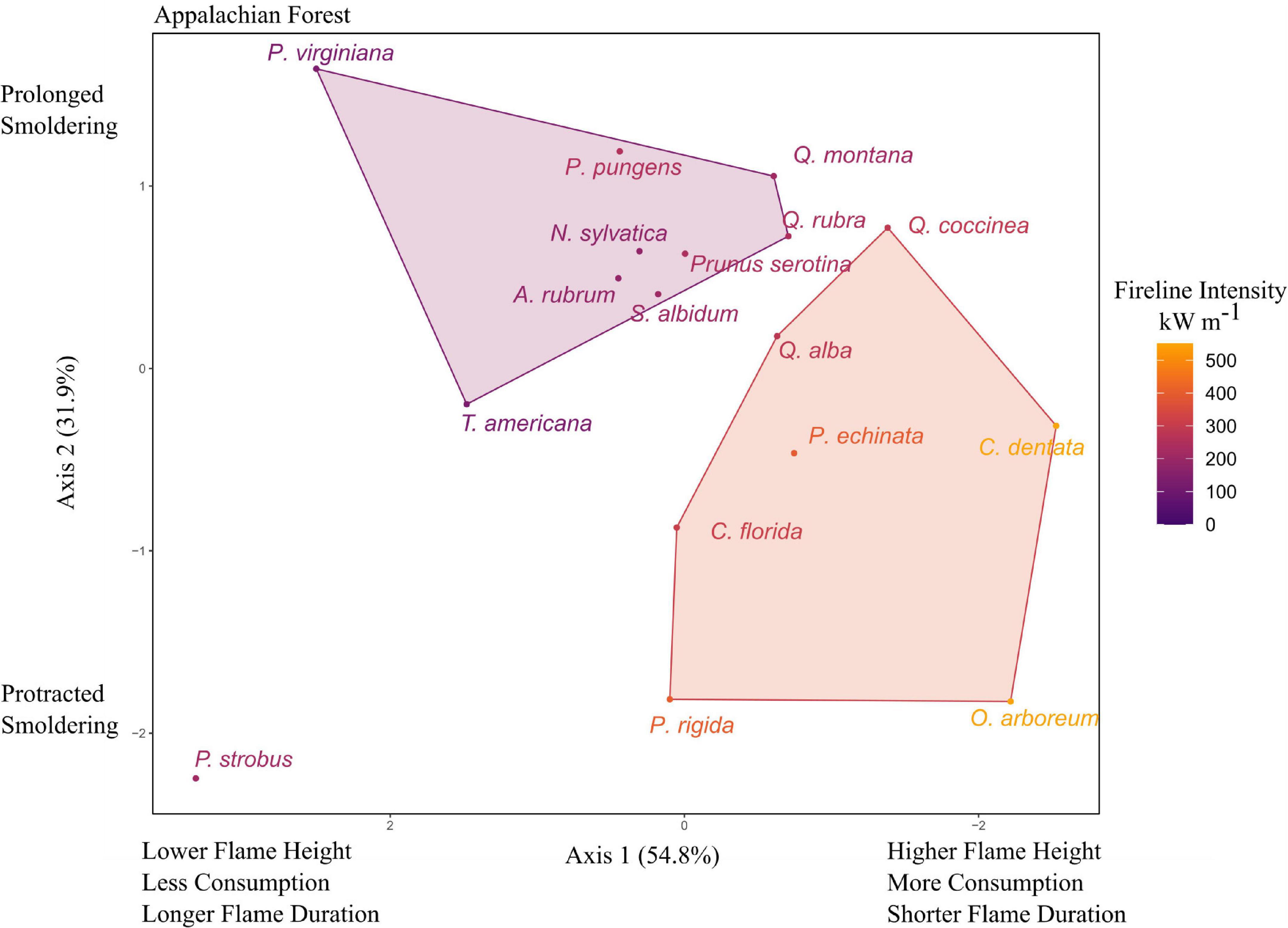
Figure 4. Principal components analysis (PCA) for flammability of tree litter in Appalachian forests of the southeastern United States. Axis 1 was inverted to be consistent with the other figures (increased flame height and consumption on the right). Species codes and mean values listed in Table 1 and Supplementary Table 1.
Bottomland Forest
The diverse species of the bottomland forest ecosystem were equally diverse in their litter flammability. Fireline intensity in bottomland forest species ranged from 31 kW m–1 (T. taxifolia) to 419 kW m–1 (Pinus serotina). The narrow endemic conifers Torreya taxifolia and Taxus floridana were identified as outliers having the second and third lowest flame heights in the dataset and were removed from the PCA. For the remaining 19 species, the first two principal components accounted for 75.5% of the variance, with axes 1 and 2 having eigenvalues of 2.01 and 1.01, respectively. Axis 1 was correlated with flame height (−0.61) and percent consumption (−0.60). Axis 2 was mostly correlated with smoldering time (−0.76) but was also correlated with flame time (0.54). The bottomland forest trees clustered into four groups that explained approximately 83.7% of the variance of the ordination. In this community, a high flaming, long smoldering time group was a mix of species from oaks (Q. alba and Q. michauxii), other hardwoods (L. tulipifera, F. grandifolia, and C. florida), and the bottomland pine, P. glabra. A high flame, brief smoldering group was made up of Pinus serotina, P. taeda, M. grandiflora, M. virginiana, and I. opaca. The intermediate flaming, long smoldering cluster contained two oaks (Q. phellos and Q. nigra), as well as A. rubrum, L. styraciflua, and T. americana. The lowest flammability cluster contained two species, Q. virginiana and Taxodium distichum (Figure 5).
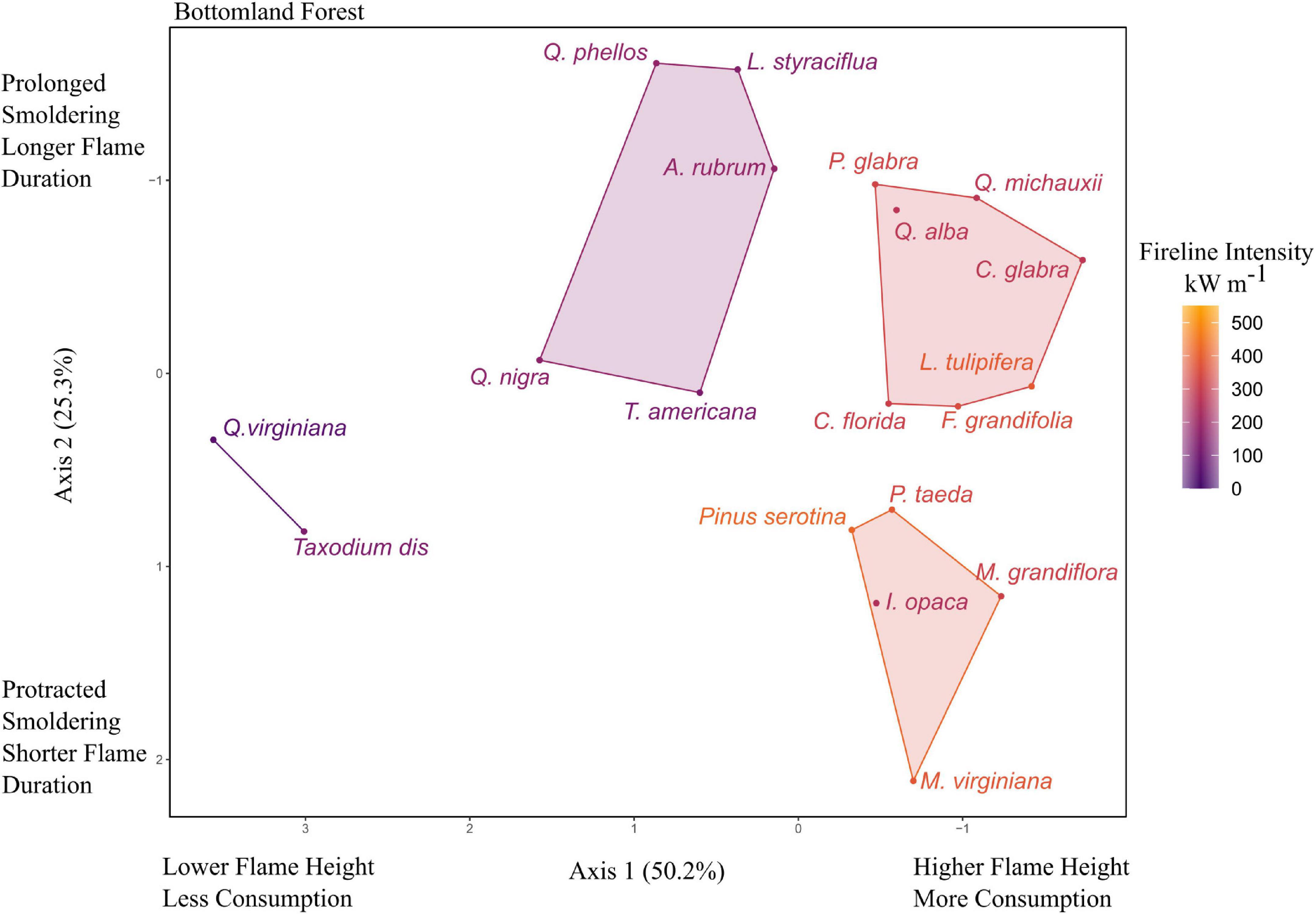
Figure 5. Principal components analysis (PCA) for flammability of litter from of trees in bottomland forests of the southeastern United States. Axes 1 and 2 were inverted to be consistent with the other figures (increased flames etc., on right and increased smoldering on the top). Species codes and mean values listed in Table 1 and Supplementary Table 1.
Discussion
The diverse southeastern tree species we evaluated here varied widely in their litter flammability metrics. Many of the species implicated in mesophication (e.g., A. rubrum, Tilia americana, Liquidambar styraciflua, and Nyssa sylvatica; Nowacki and Abrams, 2008; Kreye et al., 2018b; Babl et al., 2020) had lower flammability that diminishes fire intensity and spread, enabling these species to persist in the presence of low intensity fires or continued fire exclusion. Many of the ecosystem dominants (particularly many Pinus, Quercus, Carya, and the formerly dominant Castanea dentata) occupied the pyrophytic end of the ordinations with their ability to burn with great intensity that prevents establishment or causes mortality of invading mesophytes. These patterns were emergent across the four ecosystems we evaluated.
Flammability Across Ecosystems: Evidence for Mesophication
In the coastal plain uplands, the most flammable species were Pinus serotina, P. echinata, and P. palustris, with Q. laevis almost as flammable. All of these species are very fire tolerant and dominate frequently burned sites (Landers, 1991; Platt, 1999). P. serotina occurs in wetter flatwoods sites that can burn very intensely as surface fires or stand-replacing fires during dry windy periods (Frost, 1998). P. serotina also has serotinous cones that require intense heat to open and individual trees can persist through intense fire because of its inherent ability to basally and epicormically resprout. P. taeda, Q. alba, and Q. falcata persist in sites that are burned regularly and each of these species burned with relatively high flammability. The least flammable species included three oaks known to be fire sensitive in the coastal plain, Q. virginiana, Q. geminata, and Q. hemisphaerica. Q. virginiana and Q. geminata are evergreen oaks with massive crowns that can extend horizontally for several meters, enabling persistence in fire-prone uplands within fire shadows that are presumably formed as a result of their heavy shading and lack of flammable fuels beneath their crowns (Varner et al., 2016). Q. hemisphaerica is a thin-barked (Graves et al., 2014) fire sensitive species that can invade upland pines in the absence of fire. These poorly flammable oaks highlight an important finding that all oaks are not flammable, despite the widely held belief (Nowacki and Abrams, 2008).
Flammability results for oak-hickory forests were less consistent with our expectations. Supportive of our hypothesis, the mesophytes A. rubrum, L. styraciflua, O. virginiana, T. americana, and N. sylvatica had the least flammability. However, a few other species unexpectedly had higher flammability, e.g., F. grandifolia, Liriodendron tulipifera, Oxydendrum arboreum, that are more commonly identified as mesophytes. These mismatches likely reflect the need to incorporate differences in moisture dynamics (e.g., absorption capacity and drying rates; Kreye et al., 2013, 2018b), which we discuss in more detail below. The inclusion of these mesophytes in the high flammability cluster likely contributed to some of the more classic pyrophytes (e.g., Q. alba, Q. stellata) being included in the intermediate flammability cluster. Regardless, those species with high flammability and rapid bark accumulation (Jackson et al., 1999; Hammond et al., 2015) and/or rapid wound responses (Smith and Sutherland, 1999; Romero et al., 2009) are capable of persisting in fire-prone environments. Those species without thick bark (the thin barked mesophytes) suffer fire-caused stress and mortality in intense surface fires (Jackson et al., 1999; Hammond et al., 2015).
The litter flammability of Appalachian tree species largely conformed to expectations with a few minor exceptions. P. echinata. P. rigida, Q. alba, and C. glabra all were in the highly flammable category and are known to have relatively thick bark capable of withstanding frequent low intensity fires (Landers, 1991). American chestnut burned with high flammability as was discovered recently (Kane et al., 2019, 2020). The overstory tree species that replaced American chestnut varied widely: where flammable oaks replaced chestnut, the effects of its functional extinction may have been minor; where mesophytic species replaced chestnut, its functional extinction may have exacerbated changes to community flammability and local fire regimes (Kane et al., 2020). The surprisingly flammable sourwood is noteworthy. Sourwood is often lumped as a mesophyte, but its thick bark as an adult and its flammable litter provoke a need to learn more about its role in these fire-prone landscapes. It may be that its litter decomposes rapidly, making its flammability contribution ephemeral or that it indeed remains and contributes to intense surface fires. This species clearly deserves more attention. Mesophytic species, such as A. rubrum, N. sylvatica, and S. albidum, were less flammable as expected, however, several species were grouped with them as “intermediate.” P. pungens was located within the intermediate flammability category as well, a result that is consistent with other species that have serotinous cones and that typically burn as a high severity crown fire (Fonda, 2001; Varner et al., in review). Although T. canadensis was removed from the PCA as an outlier, it is a very strong mesophyte.
The diverse bottomland forest ecosystem had species spanning a wide variety of flammability. Aside from the two rare conifer outliers noted before (Mola et al., 2014), the species in this ecosystem had four main groups: a poorly flammable pair associated with riparian and lacustrine margins (bald-cypress and live oak); a cluster of typical mesophytes; and two clusters with moderate flammability and varying smoldering durations (pines, oaks, and a diverse suite of bottomland dominants; Figure 4). Many bottomlands are fire-protected due to their landscape position in riparian or lacustrine margins or because they are natural fire shadows where fire struggles to spread (Harper, 1911; Platt and Schwartz, 1990). We have observed fire spread into these ecosystems in a number of scenarios, and this likely occurred during droughts historically. At present, we have a poor understanding of the role of infrequent fire in regulating species composition in these ecosystems where the study of fire has been ignored or overlooked.
Flammability Research: What Next?
Over the last two decades, few southeastern species have been evaluated for their flammability, despite strong suggestions that many of ecosystems are highly flammable (Platt et al., 2016) and the widespread acknowledgment that mesophication is a much more common phenomenon than previously thought (Nowacki and Abrams, 2008; Alexander et al., 2021; Kane et al., 2021). This compilation provides data for 50 tree species, which is a fraction of the high diversity of overstory dominants in the eastern United States. The discoveries of past frequent fire regimes across eastern North America (Guyette et al., 2012; Stambaugh et al., 2015; Lafon et al., 2017; Rother et al., 2020) and the widespread interest in developing restoration and management to improve resilience and restore fire regimes begs for additional evaluation of species. Focusing research on other eastern North American species, particularly those that have debated fire regimes, should be prioritized.
Our results echo previous limitations of evaluating flammability of dry litter for species in humid environments (Varner et al., 2015b). In contrast to the western United States and many fire-prone landscapes worldwide, the southeastern United States is humid and for the most part lacks extended dry periods (Schroeder and Buck, 1970; Chen and Gerber, 1990; Stambaugh et al., 2017). Frequent rainfall and rapid overnight relative humidity recovery result in moist litter punctuated by diel drying (Kreye et al., 2018a). Flammability methods that include moisture gain and loss (as in Kreye et al., 2013; Mola et al., 2014; Kreye et al., 2020a, b) offer more potential to differentiate species that may burn well when dry, but in humid settings their water absorption capacity and slow moisture loss dampen their ignitability and diminishes their intensity once they do ignite. Understanding which leaf or fuelbed traits slow or hasten moisture loss (e.g., leaf curling or cuticle retention) may enable analyses of past flammability data to elucidate these patterns. The interaction of shading from overstory crowns or continuous canopies highlight these needs (Kreye et al., 2018a; Alexander et al., 2021). More work on including moisture retention as an interacting factor with flammability is clearly warranted (Kreye et al., 2013, 2018b).
Since we have collected litter when freshly fallen, our litter is a “best case” for a species’ flammability (i.e., before decomposition degrades litter fuel quality). Only limited research has been conducted on how decomposition may affect flammability following leaf fall (Weir and Limb, 2013). The diversity of the southeastern tree species and their differential mass loss rates and perhaps inherent fuel chemistry (Baker et al., 2001; Alexander and Arthur, 2014; Babl, 2018) should be a focus of future work. Fires in the southeastern United States can burn almost year-round, either soon after autumn litterfall or long after (months or years following senescence), affecting the fuel quality in ways that deserve attention (Weir and Limb, 2013). The humid climate and diversity of species begs for incorporating litter decomposition or time since senescence into future flammability experiments to better capture the variable contribution of litter fuels to surface fire behavior and the cascade of ecological processes these incite.
Litter is a primary fuel in woodlands and forests, but the contribution of other fuels should be better integrated. In studies of fire behavior drivers in eastern woodlands and forests, litter has been repeatedly shown to drive intensity and spread (Dickinson et al., 2016; Whelan et al., 2021). Where herbaceous fuels are proportionally more dominant, speculation exists over the importance of comparatively rare tree litter. Fill et al. (2016) measured flammability of the dominant bunchgrass Aristida stricta in longleaf pine savannas and woodlands and found that senesced tree litter diminished intensity and prolonged flaming and smoldering. Beyond herbaceous fuels, Emery and Hart (2020) found diverse flammability of woody, bark, and vine fuels in these ecosystems. These tree litter, herbaceous, and other fuel interactions should be prioritized in both laboratory and field research moving forward.
Flammability as a Fire-Adapted Trait
Litter flammability is a single fire-adapted trait, but one that is increasingly linked to other important traits that enable survival in fire-prone environments. Typical trait analyses focus on defensive traits, most classically bark thickness or the rate at which species accrue bark as juveniles (Keeley and Zedler, 1998; Jackson et al., 1999; Hammond et al., 2015). Other defensive traits include the ability to respond quickly to wounding and resist secondary effects following injury, e.g., decay (Romero et al., 2009). Other typical fire-adapted traits include pruning rates, juvenile height growth rates, resprouting or suckering capacity, and reproductive traits including fire-triggered flowering or cone serotiny (Landers, 1991; Rodríguez-Trejo and Fulé, 2003; Climent et al., 2004). Combining the multitude of fire-adapted traits including flammability into synthetic analyses that illustrate the diversity of ways that perennial woody plants persist in fire-prone environments is warranted (Schwilk and Ackerly, 2001; Varner et al., 2016; Pausas et al., 2017; Stevens et al., 2020).
Management Implications
Restoring landscapes that historically burned regularly may be challenged by fire-sensitive mesophytic species that establish following long-term fire-exclusion (Nowacki and Abrams, 2008). The dampening effect of these species on flammability increase the difficulty of killing invading mesophytes or establishing pyrophytes (Stambaugh et al., 2015). Our results highlight how variability in flammability might explain the persistence of fire-tolerant tree species through their influence on fire behavior, but quantitative research on the relative impact of mesophytes on flammability in mixed forests deserves more attention. Mesophyte litter may have minimal effect on flammability unless it dominates the surface fuels (Kreye et al., 2018b). For example, determining the mesophication impact of different species may enable managers to prioritize removal of species in a more targeted way. These removals over time will lead to the reduction of residual mesophytic litter and the canopy shade that often facilitates the growth and expansion of mesophytes while also diminishing future fires (Alexander et al., 2021). Research should be prioritized to determine the relative effectiveness of individual mesophytic species across the region.
Our working hypothesis was that species linked to fire exclusion and the feedback phenomenon termed mesophication would burn poorly and support the mechanism for regional dampened flammability. Our results, with few exceptions, confirmed our hypothesis for the four ecosystems we investigated. These results are further evidence that the mesophication feedback is widespread, beyond the eastern United States oak forests where the phenomenon was first detailed by Nowacki and Abrams (2008). Future work across other ecosystems may show similar patterns where excluding fire leads to reduced flammability, counter to expectations in many Mediterranean and dry western North American ecosystems. Understanding how shifting species composition and structure alters ecosystem processes will be particularly necessary as restoration efforts move forward and fire is reconsidered as a natural process in ecosystems more broadly.
Data Availability Statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author/s.
Author Contributions
JMV, JMK, JKK, and TS conceptualized the study. TS and JMV performed the statistical analyses. JMV wrote the first version. All authors contributed to writing and editing and approved the submitted version.
Funding
We acknowledged funding from the Joint Fire Science Program under project JFSP 13-1-04-49 and the USDA National Institute of Food and Agriculture McIntire-Stennis Program (PEN04685 and CALZ-100).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
This manuscript benefited from the collections and interest of several natural historians and ecologists across the southeastern United States. In particular, we thank R. Birkhead, A. Varner, K. Hiers, J. Mola, and T. Spector. Rich Fonda’s early work on this topic stimulated our interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2021.727042/full#supplementary-material
Supplementary Table 1 | Litter flammability metrics for 50 species in the southeastern United States.
References
Affifi, A. A., and Clark, V. (1996). Computer Aided Multivariate Analysis. New York, NY: Chapman and Hall.
Alexander, H. D., and Arthur, M. A. (2014). Increasing red maple leaf litter alters decomposition rates and nitrogen cycling in historically oak-dominated forests of the eastern U.S. Ecosystems 17, 1371–1383. doi: 10.1007/s10021-014-9802-4
Alexander, H. D., Siegert, C., Brewer, J. S., Kreye, J. K., Lashley, M. A., McDaniel, J. K., et al. (2021). Mesophication and oak landscapes: evidence, knowledge gaps, and future research. BioScience 71, 531–542. doi: 10.1093/biosci/biaa169
Babl, E., Alexander, H. D., Siegert, C. M., and Willis, J. L. (2020). Could canopy, bark, and leaf litter traits of encroaching non-oak species influence future flammability of upland oak forests? For. Ecol. Manag. 458:117731. doi: 10.1016/j.foreco.2019.117731
Babl, E. K. (2018). Could Mesophyte Canopy, Bark, and Leaf Litter Traits Drive Future Flammability of Upland Oak Forests?. Thesis. Starkville, MS: Mississippi State University, 65.
Baker, T. T., Lockaby, B. G., Conner, W. H., Meier, C. E., Stanturf, J. A., and Burke, M. K. (2001). Leaf litter decomposition and nutrient dynamics in four southern forested floodplain communities. Soil Sci. Soc. Am. J. 65, 1334–1347. doi: 10.2136/sssaj2001.6541334x
Byram, G. F. (1959). “Combustion of forest fuels,” in Forest Fire: Control and Use. ed K. P. Davis (New York, NY: McGraw-Hill). 61–89
Chen, E., and Gerber, J. F. (1990). “Climate,” in Ecosystems of Florida, eds R. L. Myers and J. J. Ewel (Gainesville: University of Florida Presses), 11–34.
Climent, J., Tapias, R., Pardos, J. A., and Gil, L. (2004). Fire adaptations in the Canary Islands pine (Pinus canariensis). Plant Ecol. 171, 185–196. doi: 10.1023/b:vege.0000029374.64778.68
Dickinson, M. B., Hutchinson, T. F., Dietenberger, M., Matt, F., and Peters, M. P. (2016). Litter species composition and topographic effects on fuels and modeled fire behavior in an oak-hickory forest in the Eastern USA. PLoS One 11:e0159997. doi: 10.1371/journal.pone.0159997
Emery, R. K., and Hart, J. L. (2020). Flammability characteristics of surface fuels in a longleaf pine (Pinus palustris Mill.) woodland. Fire 3:39.
Fei, S., and Steiner, K. C. (2007). Evidence for increasing red maple abundance in the eastern United States. For. Sci. 53, 473–477.
Fill, J. M., Moule, B. M., Varner, J. M., and Mousseau, T. A. (2016). Flammability of the keystone savanna bunchgrass Aristida stricta. Plant Ecol. 217, 331–342. doi: 10.1007/s11258-016-0574-0
Fonda, R. W. (2001). Burning characteristics of needles from eight pine species. For. Sci. 47, 390–396.
Fonda, R. W., and Varner, J. M. (2004). Burning characteristics of cones from eight pine species. Northw. Sci. 78, 322–333.
Frost, C. C. (1998). Presettlement fire frequency regimes of the United States: a first approximation. Tall Timbers Fire Ecol. Conf. Proc. 20, 70–81.
Graves, S. J., Rifai, S. W., and Putz, F. E. (2014). Outer bark thickness decreases more with height on stems of fire-resistant than fire-sensitive Floridian oaks (Quercus spp. Fagaceae). Am. J. Bot. 101, 2183–2188. doi: 10.3732/ajb.1400412
Guyette, R. P., Stambaugh, M. C., Dey, D. C., and Muzika, R. M. (2012). Predicting fire frequency with chemistry and climate. Ecosystems 15, 322–335. doi: 10.1007/s10021-011-9512-0
Hammond, D. H., Varner, J. M., Kush, J. S., and Fan, Z. (2015). Contrasting sapling bark allocation of five southeastern USA hardwood tree species in a fire-prone ecosystem. Ecosphere 6:art112. doi: 10.1890/ES15-00065.1
Hanberry, B. B., Abrams, M. D., Arthur, M. A., and Varner, J. M. (2020). Reviewing fire, climate, and multiple factors as drivers of historically open oak and pine forests and transition to closed forests. Front. For. Glob. Change 3:art56. doi: 10.3389/ffgc.2020.00056
Harper, R. M. (1911). The relation of climax vegetation to islands and peninsulas. Bull. Torrey Bot. Club 38, 515–525. doi: 10.2307/2479382
Hart, J. L., and Buchanan, M. L. (2012). “History of fire in eastern oak forests and implications for restoration,” in Proceedings of the 4th Fire in Eastern Oak Forests Conference; Gen. Tech. Rep, eds D. C. Dey, M. C. Stambaugh, S. L. Clark, and C. J. Schweitzer (Newtown Square, PA: US Department of Agriculture,), 35–44. NRS-P-102.
Jackson, J. F., Adams, D. C., and Jackson, U. B. (1999). Allometry of constitutive defense: a model and a comparative test with tree bark and fire regime. Am. Nat. 153, 614–632. doi: 10.1086/303201
Kane, J. M., Kreye, J. K., Barajas-Ramirez, R., and Varner, J. M. (2021). Litter trait driven dampening of flammability following deciduous forest community shifts in eastern North America. For. Ecol. Manag. 489:119100. doi: 10.1016/j.foreco.2021.119100
Kane, J. M., Varner, J. M., and Hiers, J. K. (2008). The burning characteristics in southeastern oaks: discriminating fire facilitators from fire impeders. For. Ecol. Manag. 256, 2039–2045. doi: 10.1016/j.foreco.2008.07.039
Kane, J. M., Varner, J. M., and Sauders, M. R. (2019). Resurrecting the lost flames of American chestnut. Ecosystems 5, 995–1006. doi: 10.1007/s10021-018-0318-1
Kane, J. M., Varner, J. M., Stambaugh, M. C., and Saunders, M. (2020). Reconsidering the fire ecology of the iconic American chestnut. Ecosphere 11:e03267. doi: 10.1002/ecs2.3267
Keeley, J. E., and Zedler, P. H. (1998). “Evolution of life histories in Pinus,” in Ecology and Biogeography of Pinus, ed. D. M. Richardson (Cambridge: Cambridge University Press), 219–249.
Knott, J. A., Desprez, J. M., Oswalt, C. M., and Fei, S. (2019). Shifts in forest composition in the eastern United States. For. Ecol. Manag. 433, 176–183. doi: 10.1016/j.foreco.2018.10.061
Kreye, J. K., Kane, J. M., Varner, J. M., and Hiers, J. K. (2020a). Radiant heating increases flammability of pine and oak litter via altered moisture dynamics. Fire Ecol. 16:art8. doi: 10.1186/s42408-020-0067-3
Kreye, J. K., Kane, J. M., Varner, J. M., and Hiers, J. K. (2020b). Radiant heating rapidly increases litter flammability through impacts on fuel moisture. Fire Ecol. 16, 1–10.
Kreye, J. K., Varner, J. M., Hamby, G., and Kane, J. M. (2018b). Mesophytic litter dampens flammability in fire-excluded pyrophytic oak-hickory woodlands. Ecosphere 9:e02078. doi: 10.1002/ecs2.2078
Kreye, J. K., Hiers, J. K., Varner, J. M., Hornsby, B., Drukker, S., and O’Brien, J. J. (2018a). Effects of solar heating on the moisture dynamics of forest floor litter in humid environments: composition, structure, and position matter. Can. J. For. Res. 48, 1331–1342. doi: 10.1139/cjfr-2018-0147
Kreye, J. K., Varner, J. M., Hiers, J. K., and Mola, J. (2013). Toward a mechanism for eastern North American forest mesophication: the role of litter drying. Ecol. Appl. 23, 1976–1986. doi: 10.1890/13-0503.1
Lafon, C. W., Naito, A. T., Grissino-Mayer, H. D., Horn, S. P., and Waldrop, T. A. (2017). Fire History of the Appalachian Region: A Review and Synthesis. Asheville, NC: US Department of Agriculture, 97. Gen. Tech. Rep. SRS-219.
Landers, J. L. (1991). Disturbance influences on pine traits in the southeastern United States. Proc. Tall Timbers Fire Ecol. Conf. 17, 61–98.
MacCleery, D. W. (1993). American Forests: A History of Resiliency and recOvery. Asheville, NC: US Department of Agriculture. FS-540.
Menges, E. S. (1999). “Florida Scrub,” in The Savanna, Barren, and Rock Outcrop Plant Communities of North America, eds R. C. Anderson, J. S. Fralish, and J. M. Baskin (Cambridge: Cambridge University Press), 7–22.
Mola, J. M., Varner, J. M., Jules, E. S., and Spector, T. (2014). Altered community flammability in Florida’s Apalachicola ravines and implications for the persistence of the endangered conifer Torreya taxifolia. PLoS One 9:e103933. doi: 10.1371/journal.pone.0103933
Nowacki, G. J., and Abrams, M. D. (2008). The demise of fire and “mesophication” of forests in the eastern United States. BioScience 58, 123–138. doi: 10.1641/B580207
Pausas, J. G., Keeley, J. E., and Schwilk, D. W. (2017). Flammability as an ecological and evolutionary driver. J. Ecol. 105, 289–297. doi: 10.1111/1365-2745.12691
Platt, W. J. (1999). “Southeastern pine savannas,” in Savannas, Barrens, and Rock Outcrop Plant Communities of North America, eds R. C. Anderson, J. S. Fralish, and J. M. Baskin (Cambridge: Cambridge Press), 23–51.
Platt, W. J., Ellair, D. P., Huffman, J. M., Potts, S. E., and Beckage, B. (2016). Pyrogenic fuels produced by savanna trees can engineer humid savannas. Ecol. Monogr. 86, 352–372. doi: 10.1002/ecm.1224
Platt, W. J., and Schwartz, M. W. (1990). “Temperate hardwood forests,” in Ecosystems of Florida, eds R. L. Myers and J. J. Ewel (Gainesville: University of Florida Presses), 194–229.
Privé, F. (2021). Bigutilsr: Utility Functions for Large-Scale Data. R Package Version 0.3.4. Avaliable online at: https://CRAN.R-project.org/package=bigutilsr (accessed June 11, 2021).
R Core Team (2020). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rodríguez-Trejo, D. A., and Fulé, P. Z. (2003). Fire ecology of Mexican pines and a fire management proposal. Int. J. Wildland Fire 12, 23–37. doi: 10.1071/WF02040
Romero, C., Bolker, B. M., and Edwards, C. E. (2009). Stem responses to damage: the evolutionary ecology of Quercus species in contrasting fire regimes. New Phytol. 182, 261–271. doi: 10.1111/j.1469-8137.2008.02733.x
Rother, M. T., Huffman, J. M., Guiterman, C. H., Robertson, K. M., and Jones, N. (2020). A history of recurrent, low-severity fire without fire exclusion in southeastern pine savannas. USA. For. Ecol. Manag. 475:118406. doi: 10.1016/j.foreco.2020.118406
Schroeder, M. J., and Buck, C. C. (eds) (1970). “Fire weather–: a guide for application of meteorological information to forest fire control operations,” in USDA Forest Service Agriculture Handbook, Vol. 360, (Washington, D.C.: U.S. Dept. of Agriculture), 229.
Schwilk, D. W., and Ackerly, D. D. (2001). Flammability and serotiny as strategies: correlated evolution in pines. Oikos 94, 326–336. doi: 10.1034/j.1600-0706.2001.940213.x
Smith, K. T., and Sutherland, E. K. (1999). Fire-scar formation and compartmentalization in oak. Can. J. For. Res. 29, 166–171.
Stambaugh, M. C., Guyette, R. P., and Marschall, J. M. (2011). Longleaf pine (Pinus palustris Mill.) fire scars reveal new details of a frequent fire regime. J. Veg. Sci. 22, 1094–1104. doi: 10.1111/j.1654-1103.2011.01322.x
Stambaugh, M. C., Varner, J. M., and Jackson, S. T. (2017). “Biogeography: an interweave of climate, fire, and humans,” in Ecological Restoration and Management of Longleaf Pine Forests, eds L. K. Kirkman and S. B. Jack (Boca Raton, FL: CRC Press), 17–37. doi: 10.1201/9781315152141-2
Stambaugh, M. C., Varner, J. M., Noss, R. F., Dey, D. C., Christensen, N., Baldwin, R. F., et al. (2015). Clarifying the role of fire in the Eastern deciduous forest: reply to matlack. Conserv. Biol. 29, 942–946. doi: 10.1111/cobi.12473
Stevens, J. T., Kling, M. M., Schwilk, D. W., Varner, J. M., and Kane, J. M. (2020). Biogeography of fire regimes in western U.S. conifer forests: a trait-based approach. Glob. Ecol. Biogeogr. 29, 944–955. doi: 10.1111/geb.13079
U.S. Environmental Protection Agency. (2013). Level III and IV ecoregions of the continental United States. U.S. EPA, National Health and Environmental Effects Research Laboratory, Corvallis, Oregon. Available online at: https://www.epa.gov/eco-research/level-iii-and-iv-ecoregions-continental-united-states (accessed 28 September 2021).
Varner, J. M., Kane, J. M., Kreye, J. K., and Veldman, J. W. (2016). Suites of fire-adapted traits in the southeastern USA oaks: multiple strategies for persistence. Fire Ecol. 12, 48–64. doi: 10.4996/fireecology.1202048
Varner, J. M., Kane, K. M., Banwell, E. M., and Kreye, J. K. (2015a). “Flammability of litter from southeastern trees: a preliminary assessment,” in Proceedings of the 17th Southern Silvicultural Research Conference, eds G. A. Holley, K. F. Connor, and J. D. Haywood (Asheville, NC: USDA Forest Service), 183–187. eGTR-SRS-203.
Varner, J. M., Kane, J. M., Kreye, J. K., and Engber, E. (2015b). The flammability of forest and woodland litter: a synthesis. Curr. For. Rep. 1, 91–99. doi: 10.1007/s40725-015-0012-x
Weir, J. R., and Limb, R. F. (2013). Seasonal variation in flammability characteristics of Quercus marilandica and Quercus stellata leaf litter burned in the laboratory. Fire Ecol. 9, 80–88. doi: 10.4996/fireecology.0903080
Wenk, E. S., Wang, G. G., and Walker, J. L. (2011). Within-stand variation in understorey vegetation affects fire behaviour in longleaf pine xeric sandhills. Int. J. Wildland Fire 20, 866–875. doi: 10.1071/WF10087
Keywords: Acer rubrum, fire-adapted traits, fire regimes, fuels, mesophication, Pinus, Quercus
Citation: Varner JM, Kane JM, Kreye JK and Shearman TM (2021) Litter Flammability of 50 Southeastern North American Tree Species: Evidence for Mesophication Gradients Across Multiple Ecosystems. Front. For. Glob. Change 4:727042. doi: 10.3389/ffgc.2021.727042
Received: 17 June 2021; Accepted: 21 September 2021;
Published: 12 October 2021.
Edited by:
Thomas J. Dean, Louisiana State University, United StatesReviewed by:
Gregory Jay Nowacki, United States Forest Service (USDA), United StatesLauren Pile Knapp, US Forest Service Research and Development, United States
Copyright © 2021 Varner, Kane, Kreye and Shearman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: J. Morgan Varner, bXZhcm5lckB0YWxsdGltYmVycy5vcmc=