 Rubén Ortega-Álvarez
Rubén Ortega-Álvarez Alana Pacheco-Flores
Alana Pacheco-Flores Alejandro Casas
Alejandro Casas- Instituto de Investigaciones en Ecosistemas y Sustentabilidad (IIES) – Universidad Nacional Autónoma de México, Morelia, Mexico
Indigenous management of biocultural resources can influence behavior of non-target animals. Hence, identifying shared resources between indigenous communities and fauna is essential to understand the ecological relationships that occur within cultural landscapes and promote sustainable practices. In this ethnoecological research we analyzed the feeding importance of a wild food that is used by both people and birds. We employed unstructured interviews and a survey to learn about food resources used by people of a Nahua community in western Mexico. Trees were frequently mentioned by locals; thus, we determined salience and cultural meaning of arboreal feeding sources for the indigenous community. The “Guamúchil” or Manila tamarind (Pithecellobium dulce) was the most bioculturally salient tree for the feeding and economy of local people, and we evaluated its significance for birds too. We analyzed avian foraging behavior to estimate the number of species that used the tree as a food source, the relevance of Guamúchil as foraging substrate, food types provided by this tree, and the avian feeding use of Guamúchil when it was locally employed as a live fence. We observed that local people tracked fruiting Guamúchil across the landscape and cultivated it in homegardens to facilitate fruit access. Seed pods were locally appreciated for direct consumption by households and commercialization. Guamúchil’s cover was low in the surrounding forest (mean: 0.4, standard deviation: 0.7), but much higher in homegardens (mean: 20.9, standard deviation: 16.3). We estimated that about a fourth of the avian species that foraged across the cultural landscape during our sampling feed on Guamúchil. Birds prey on invertebrates associated with this tree, but Guamúchil was mostly important to avian foraging for the fruit it produces. We determined that live fences of Guamúchil functioned as complementary food sources to birds. Our research shows how humans and birds share key feeding resources within cultural landscapes. Also, we depict how indigenous agroforestry practices positively impact wildlife foraging, which should be promoted as conservation and restoration tools to support sustainability. Identifying key foods and analyzing their use might facilitate the development of activities aimed to benefiting both humans and animal communities.
Introduction
Communities of indigenous people usually have a long tradition of ecological interactions with the environment, using and managing components of ecosystems that occur within their territories (Berkes et al., 2000). As consequence, they have influenced evolutionary and ecological processes on several kinds of organisms, including plants, animals, mushrooms, and microbiota (Luat-Hu’eu et al., 2021; Ojeda-Linares et al., 2021). The domestication of particular species has induced changes in the demography and genetic composition of their populations, leading to their maintenance, propagation, and diversification (Casas et al., 2007; Clement et al., 2021). In addition, these interactions have influenced the composition and structure of biotic communities, therefore shaping landscapes (Casas et al., 1997; Levis et al., 2017; Clement et al., 2021). Besides the direct effects that indigenous management might have on particular species of biocultural interest, the utilization and management of common resources might provoke indirect impacts on populations of other companion organisms (Perfecto and Vandermeer, 2008). For example, indigenous practices might include plants that are also vital to other animals (Turner and Bhattacharyya, 2016). As a result, use and management decisions on shared plants employed by humans might influence the demography and behavior of animal populations (Perfecto et al., 2009).
The long and intimate relationship that prevails between indigenous knowledge, practices, and biodiversity in sites known as “cultural landscapes” is widely recognized (Droste et al., 1995; Farina, 2000; UNESCO, 2003; Gfeller, 2013). Still, further efforts are needed to document the local particularities of biodiversity and ecosystem management and expand our comprehension about the ecological implications of indigenous use on wildlife (Moreno-Calles et al., 2013; Casas et al., 2016). Such information might result valuable to preserve local collective memory (Monroy and Colín, 2004), biocultural diversity conservation (Maffi and Woodley, 2010), and understanding changes in indigenous practices resulting from the influence of forces external to the community (hegemonic economic models, globalization, climate change, communication media, extensionists, among others) (Chinlampianga et al., 2013; Thompson et al., 2020). Additionally, identifying shared resources between indigenous people and animal communities is essential to comprehend the ecological relationships that occur within cultural landscapes, foster biodiversity conservation, promote management practices that might lead to sustainability, and facilitate environmental restoration (Nakashima and Roué, 2002; Jernigan and Dauphiné, 2008; Turner and Bhattacharyya, 2016).
Feeding is vital for both humans and other animals. Its study is fundamental to identify the factors, preferences, and resources that require survival. Research has largely focused on determining the food items that are consumed by indigenous communities within their territories (de Paula et al., 2018; Akinola et al., 2020; Thompson et al., 2020), whereas animal feeding use of cultural landscapes has been poorly assessed (Ortega-Álvarez and Casas, 2022). Moreover, little investigation has been performed to understand how a common food resource might simultaneously affect the fauna and indigenous communities. In particular birds are tightly associated with indigenous life (Tidemann and Gosler, 2010). Still, avian use of indigenous territories has been documented through the study of traditional knowledge on the natural history of birds (Jernigan and Dauphiné, 2008; Turner and Bhattacharyya, 2016), but scarce ecological research has addressed the effects of local management practices on bird foraging (Ortega-Álvarez and Casas, 2022). Investigation on avian ethology might be valuable to increase our understanding about the human-bird ecological relationships that occur within cultural landscapes and determine how people’s use of resources impact animal behavior (Ortega-Álvarez et al., 2021).
The “Guamúchil” or Manila tamarind (Pithecellobium dulce) is a member of the Fabaceae family, native to the American continent. It grows as an evergreen tree that might reach a height up to 20 m, creating a wide canopy. The length of the pod that it produces is about 20 cm, containing a sweet fleshy pulp (the aril) that protects circular black seeds (Pennington and Sarukhán, 2005). People eats its fruit, whereas birds are known to disperse its seeds. The Guamúchil is a fast-growing species that tolerates droughts and is mostly distributed across tropical lands of the Pacific Coast, from Mexico to northern South America. It is usually found from sea level to 1,800 m (Pennington and Sarukhán, 2005).
In this ethnoecological research we analyzed the feeding importance of wild food that is used by both a Nahua community and birds in western Mexico. Two main questions guided our investigation: (1) which is the wild food that is the most relevant to the local indigenous community? and (2) how does such food impact avian foraging? Human relevance was assessed in terms of biocultural salience, use type, and destiny of products (i.e., self-supply, commercialization), whereas impact on birds was determined through an ethological approach. We detected that arboreal plants were frequently mentioned by local people and investigated on the most salient trees. In particular, we identified the “Guamúchil” (P. dulce) having exceptionally high biocultural salience for the feeding and economy of the indigenous community; thus, we evaluated its significance for birds too. We analyzed the number of species that used the tree as a food source, the relevance of Guamúchil as a surface for obtaining food (foraging substrate), the types of food provided by the tree (e.g., fruits, invertebrates), and the avian feeding use of Guamúchil when it was locally employed as a live fence. Other ornithological studies have recorded that the tree provides insects and fruits to several bird species (Torres-González et al., 2014; Alvarez-Alvarez et al., 2018), whereas critical live fences might influence avian foraging (Harvey et al., 2005). Hence, we hypothesized that: a great number of bird species use the Guamúchil for feeding, specially to obtain invertebrates and fruit; foraging activity is higher in the Guamúchil than in other feeding substrates; and birds forage proportionally higher on Guamúchil than in other types of live fences.
Materials and methods
Study site
We carried out the research in the Nahua community of Zacualpan, Colima, western Mexico. Despite modern lifestyle, the traditional use of several wild food resources persists in the site (Grupo Xolocuahuitl Zacualpan, 2020; Mejía, 2021). Five different typologies constitute the cultural landscape: crop fields, Hog plum (Spondias purpurea) orchards, riparian habitats, forested areas, and an urban settlement. Maize (Zea mays) and squash (Cucurbita spp.) are mainly produced within crop fields. Riparian habitats are mostly used as water and food sources, as well as for local tourism. Tropical dry forests are dominant in the area, but they are negatively impacted by extensive cattle ranching. Most of the local people live in the urban settlement (∼2,000 inhabitants), which has an extension of ∼30 ha. Homegardens are common in the town and represent an important food source for the population. Principal productive activities include farming and commerce. Agricultural products are usually sold within the town and in the City of Colima and Villa de Álvarez, the capital of the state, which is about 20 km from Zacualpan.
Surveys on the use of food resources by the indigenous community
During December 2021 and January 2022, we employed unstructured interviews to learn about the food resources that people from Zacualpan gather, hunt, and cultivate within its territory. We directed our interviews to thirteen people who perform farming, trading, and traditional cooking activities within Zacualpan to obtain key information. Nine of them were older than 60 years, whereas the rest aged between 30 and 60 years. Eight of the participants were females. Our conversations focused on the food resources that they obtain within their territory, their origin (i.e., cultivated, gathered, hunted, raised), their use (i.e., self-supply, commercialized), and the landscape typology where they are acquired (i.e., homegarden, orchard, cropfield, forest, river). After compiling the list of foods, we determined the frequency of mention of each resource (Supplementary Appendix 1). In this way, we observed that arboreal plants, headed by the Guamúchil, were frequently mentioned by people. In fact, all participants denoted the food and economic importance of the Guamúchil’s fruit. Based on these results, we designed a survey for rapid interviews to 32 members of the community (i.e., 16 women and 16 men aging between 24 and 87 years) targeted on identifying salient trees as local food. Survey data was used to compute Smith, Sutrop, and B’ score indices with the online, open-source software for free-list analyses FLARES (Wencelius et al., 2017), in order to determine the cultural salience of the trees that were used by the community as food sources. Cultural salience is calculated by the Smith index with the number of items in a list, the rank of items in the list, and the number of lists in the sample (Smith and Borgatti, 1997). The Sutrop index combines two list task parameters (term frequency and its mean position in the lists) for computing cultural salience of a given term (Sutrop, 2001). Finally, B’ score index measures cognitive salience by including list position and frequency, but enabling comparisons irrespective of list length or number of respondents because it varies between 0 and 1 (Robbins et al., 2017).
Bird foraging surveys
We performed avian surveys during the end of the rainy season of 2022 (January), when overwintering species are present in Zacualpan. This is the local fruiting season of Guamúchil, and people actively search for its fruits. We used 30-m fixed-radius point count stations to record birds during a 10 min period per point count (Ruiz Gutiérrez et al., 2020). Observations initiated since daybreak and lasted 4 h to cover avian peak activity. Stations were located in the four typologies of the cultural landscape where people usually obtain food (i.e., homegardens, crop fields, Hog plum orchards, forested areas). We set a minimum distance of 200 m to separate point count stations (Bibby et al., 2000; Ruiz Gutiérrez et al., 2020), which allowed us to establish eight stations that covered all the surface of the smallest typology of the landscape (i.e., homegardens). Thus, we decided to establish the same number of stations for all typologies. We located 32 stations across the cultural landscape, which were visited in eight different occasions for a total of 256 point counts. We alternated the visitation sequence in each station to reduce a temporal bias on our observations.
In each station we recorded all seen bird species and included information about the number of individuals, behavior, substrate where the behavior was displayed, food types consumed by individuals, and use of live fences. Behavior was assessed during the first 5 s after detection, in order to reduce correlation among observations and observers’ effect on bird behavior (Wagner, 1981; Acevedo, 2007; Ortega-Álvarez et al., 2021). We differentiated among two types of behaviors: feeding, when birds actively foraged or ingested food; and other, when birds performed other action than feeding. We considered different types of substrates, including Guamúchil, Hog plum, trees other than the two mentioned before, shrubs, herbs, ground, pavement, cable, posts, buildings, and air. Food resources included nectar, fruits, invertebrates, vertebrates, seeds, and vegetation material other than seeds (e.g., leaves, roots, sprouts). Additionally, we recorded if the substrates that birds used were live fences, which consisted of living plants employed to set boundaries among orchards, homegardens, or crop fields.
Variation of Guamúchil cover across the cultural landscape
We assessed the variation of Guamúchil cover among the typologies of the cultural landscape. First, a single observer visually estimated the percentage of each point count station that was covered by the tree. Then, we used an ANOVA to determine if the mean cover of Guamúchil differed between Hog plum orchards, crop fields, homegardens, and the tropical dry forest.
Avian species richness that used Guamúchil as a food source
We employed the observed abundance of each species that foraged on Guamúchil to estimate the avian species richness that used such tree as a food source. For comparison purposes, we also utilized the observed abundances of all foraging birds to estimate the number of species that fed across the study site. Estimations were performed by computing sample-size-based rarefaction (interpolated estimation) and extrapolation (predictive estimation) sampling curves for bird species richness with the “iNext” package (Hsieh et al., 2016). This approach enabled us to quantify and compare foraging species richness between Guamúchil and the whole bird community with a unified standardization method (Hsieh et al., 2016). Finally, to identify the bird species that relied the most on Guamúchil as a food source, we calculated the percentage of the feeding events that occurred in the Guamúchil for each species, in relation to the total number of foraging records for the same species.
Relevance of Guamúchil for avian foraging
We analyzed the foraging relevance of Guamúchil to birds from three different perspectives: (1) as a foraging substrate, (2) in relation to the types of food that it provides, and (3) its feeding use when locally employed as a live fence. First, we determined the relevance of Guamúchil as a foraging substrate to birds by comparing the proportion of feeding events for different types of substrates. We used a hierarchical binomial model (Kruschke, 2015) to estimate the foraging proportion for each substrate in relation to the total number of feeding events for all substrates. Substrates included ground, herbs, shrubs, Hog plum, Guamúchil, tree species other than the two mentioned before, and air. Hog plums were included in this analysis because we observed that they were particularly attractive to foraging birds. We did not model foraging proportions for cables, posts, pavement, and buildings given that we observed that birds neglected them (<3 feeding records for each substrate).
Then, we compared the proportion of food types consumed by birds for distinct kinds of feeding sources. We estimated the proportion of consumed food items for each source in relation to the total number of feeding events on each food item for all feeding sources by using a hierarchical binomial model (Kruschke, 2015). Modeling was restricted to fruits and invertebrates, as both were the only food types that birds obtained from the Guamúchil. Feeding sources included shrubs, Guamúchil, other tree species, and air. We selected these sources because they had enough associated data to perform our analyses.
Finally, we investigated the avian foraging relevance of Guamúchil when locally used as a live fence by contrasting the proportion of feeding occasions for different types of live fences. Again, a hierarchical binomial model (Kruschke, 2015) was used to determine the foraging proportion for each live fence in relation to the total number of observed behaviors for the same type of fence. We considered two kinds of live fences: Guamúchil and other live fences; the latter included both shrubs and tree species other than Guamúchil.
All the aforementioned hierarchical binomial models (Kruschke, 2015) were analyzed by using a Bayesian proportion test with the “BayesianFirstAid” R package (Baath, 2014), which was implemented with JAGS (Plummer, 2003). We used an uninformative beta prior with α = β = 1 to model all proportions. We employed Markov Chain Monte Carlo with three Markov chains and 5,000 iterations to perform Bayesian inference. Convergence was determined with the R-hat statistic (Gelman and Rubin, 1992). Differences in proportions were assessed when 95% highest density intervals of the posterior distributions of the estimated differences excluded 0.
Results
The survey confirmed the biocultural salience of the Guamúchil for local human feeding. Among the 35 tree species that were used by the community as food sources, the Guamúchil exhibited the highest values for all computed indices (Table 1). People track fruiting Guamúchil trees across the landscape, gathering the pods from the trees that are dispersed within the forest. Moreover, they have practiced its cultivation in homegardens and propagation in agricultural lands to facilitate the access and increase the abundance of the fruit. Similarly to central Mexico (Monroy and Colín, 2004), the community uses Guamúchil as a live fence for defining the limits of plots while promoting its cultivation. The seed pods of this tree are locally appreciated for self-supply; their pulp might be eaten raw, roasted, or ground with corn to prepare “sopes” (a thick, small maize tortilla with sauce and other ingredients). Also, the fruit is valued for its commercialization in the streets of the town, the local market, and the city of Colima. Thus, Guamúchil simultaneously feeds and provides incomes to the families of Zacualpan. In addition, we have observed that the tree might be locally utilized as medicine, firewood, construction material (e.g., poles), for elaborating handcrafts, and providing shade for the cattle, which enhances even more the biocultural relevance of this plant.
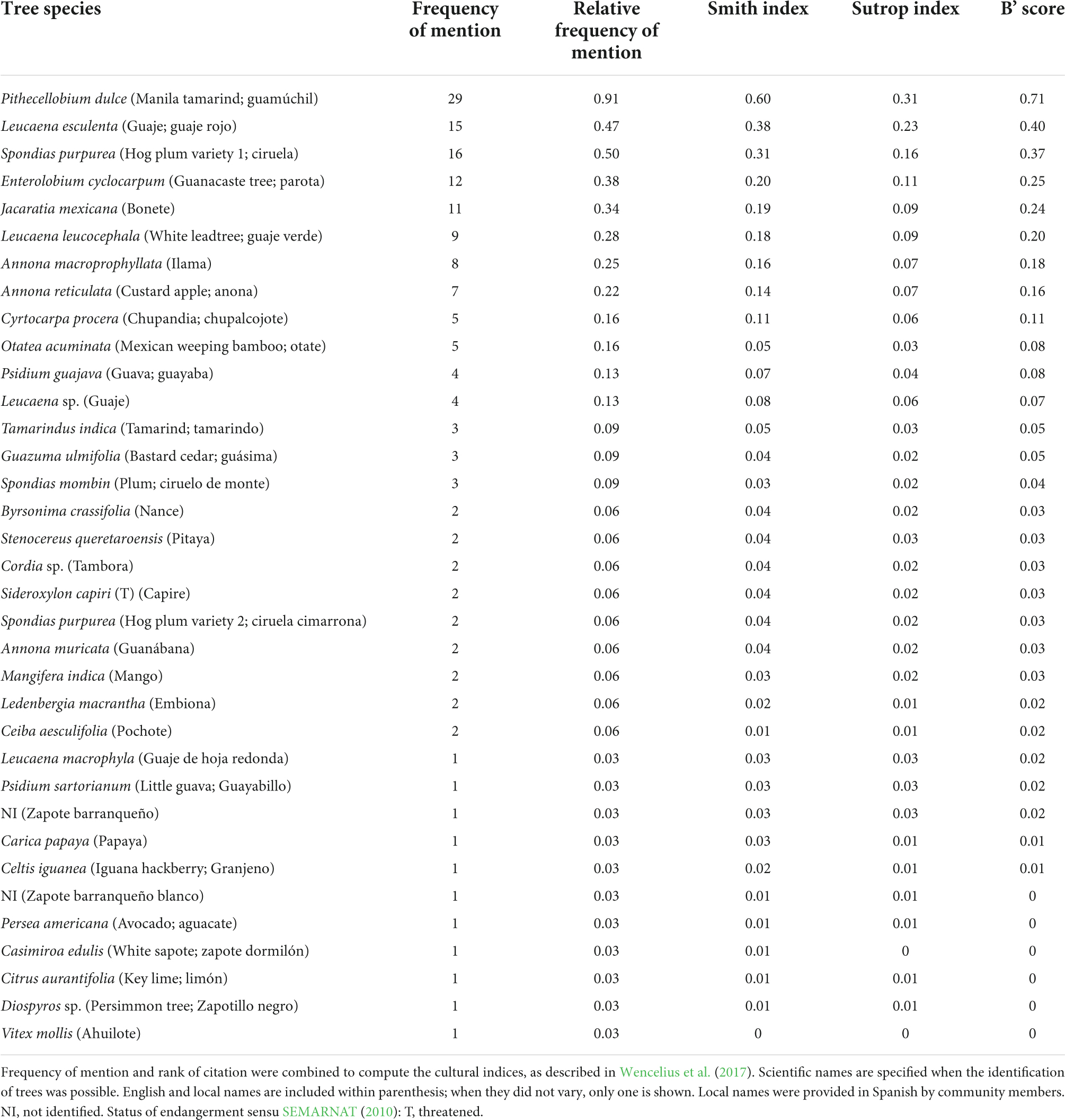
Table 1. Frequency of mention and cultural salience indices of the trees that are used as feeding sources in Zacualpan, Colima, Mexico.
We observed that Guamúchil cover differed among landscape typologies (Figure 1). In particular, it was high in homegardens (mean: 20.9, standard deviation: 16.3), but low in the tropical dry forest (mean: 0.4, standard deviation: 0.7) (Figure 1). Of the 69 bird species that we observed feeding across the cultural landscape, 19 (27.5%) used Guamúchil to obtain food (Supplementary Appendix 2). According to our estimations, up to 82 species were expected to be foraging in the study site (Table 2), whereas 22 were estimated to be using Guamúchil as a food source (Table 3). Species such as the White-throated Magpie-Jay (Calocitta formosa), the Orange-fronted Parakeet (Eupsittula canicularis), the Yellow-rumped Warbler (Setophaga coronata), the Rufous-backed Robin (Turdus rufopalliatus), and the Western Tanager (Piranga ludoviciana) heavily used Guamúchil for foraging (Supplementary Appendix 2).
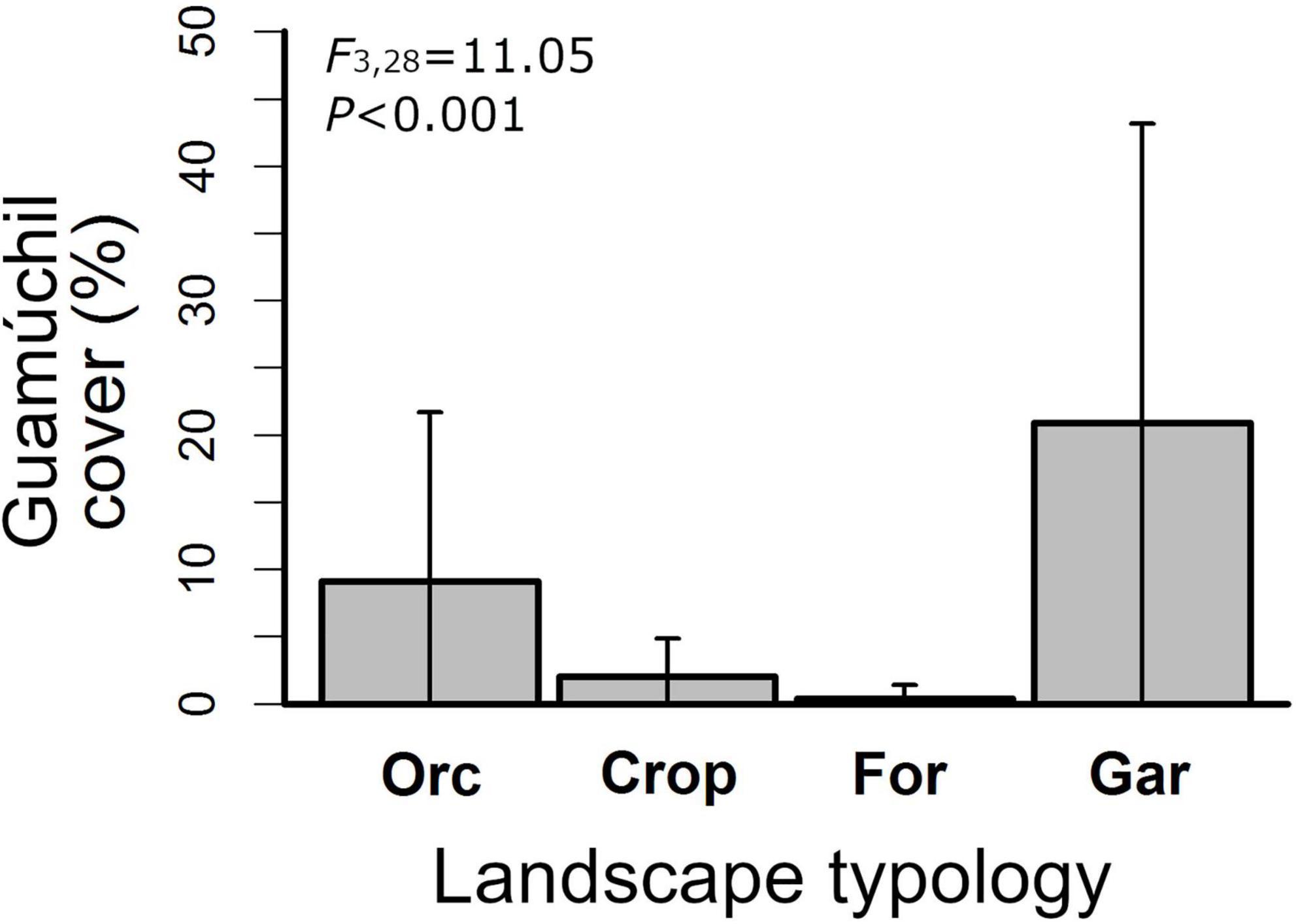
Figure 1. Variation of Guamúchil cover among the typologies of the cultural landscape of Zacualpan, Colima, Mexico. Orc, Hog plum orchards; Crop, cropfields; For, tropical dry forest; Gar, homegardens.
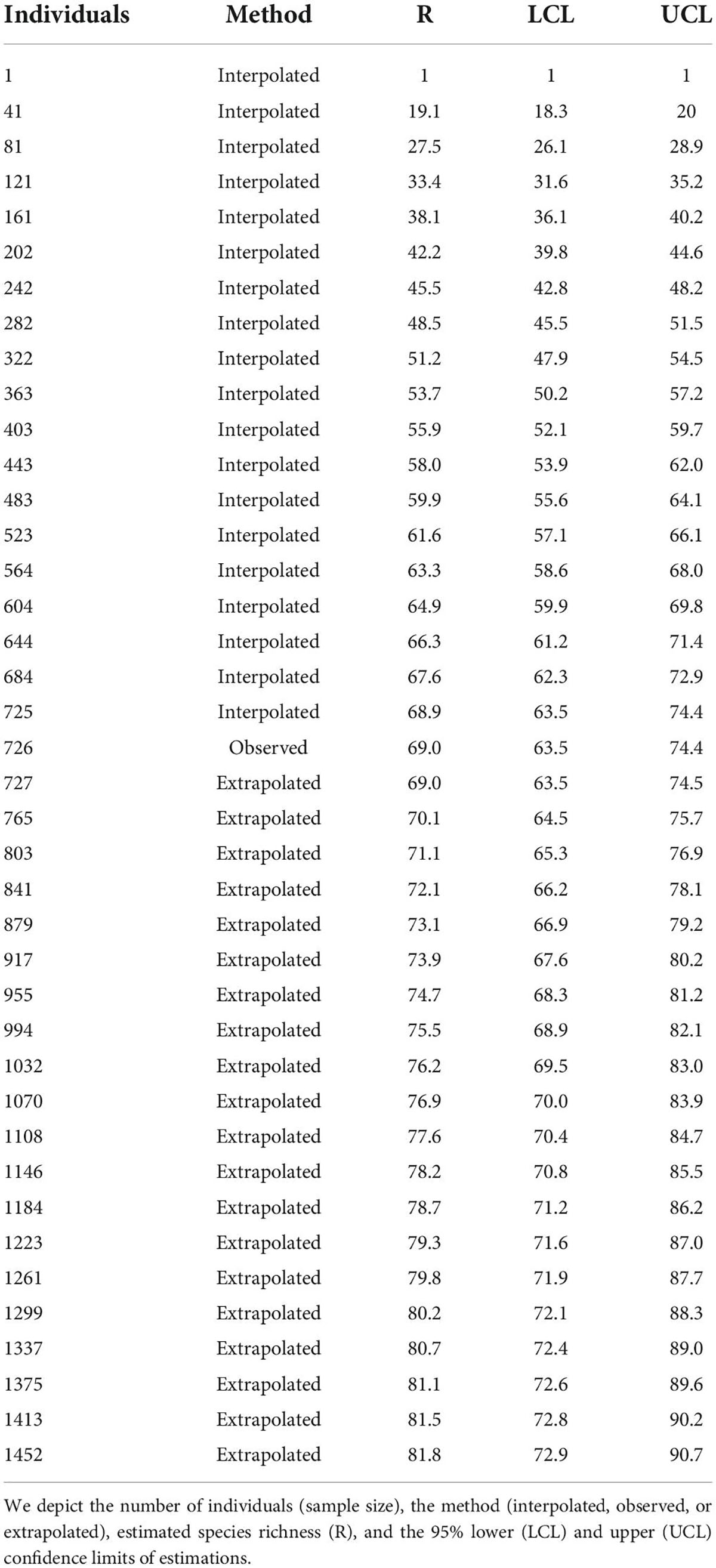
Table 2. Estimated richness of bird species that forage across Zacualpan, Colima, Mexico.
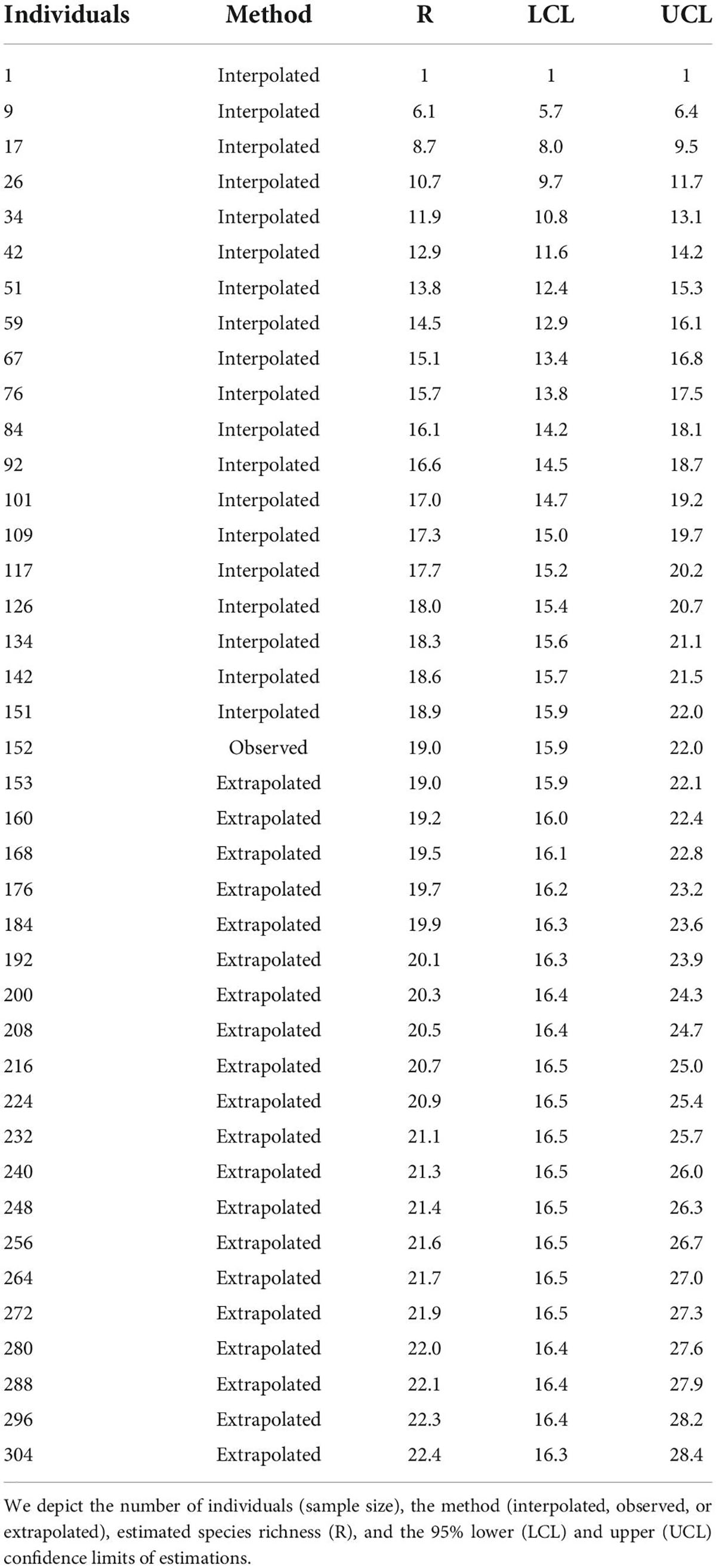
Table 3. Estimated richness of bird species that forage on Guamúchil across Zacualpan, Colima, Mexico.
Birds used different substrates to feed across the cultural landscape. We determined that the proportion of avian feeding events peaked on tree species other that Hog plums and Guamúchil trees, followed by the use of aerial and Guamúchil substrates (Figure 2). Food types consumed by birds were obtained from distinct sources. Guamúchil served as the main source of fruit to birds, whereas such animals caught invertebrates mostly in the air and on trees other than Guamúchil (Figure 3). When locally employed as a live fence, the Guamúchil was visited by a higher number of birds than any other type of live fence (Figure 4). Bird species that most frequently foraged on Guamúchil live fences included Blue-gray Gnatcatcher (Polioptila caerulea), Western Tanager, and Rufous-backed Robin.
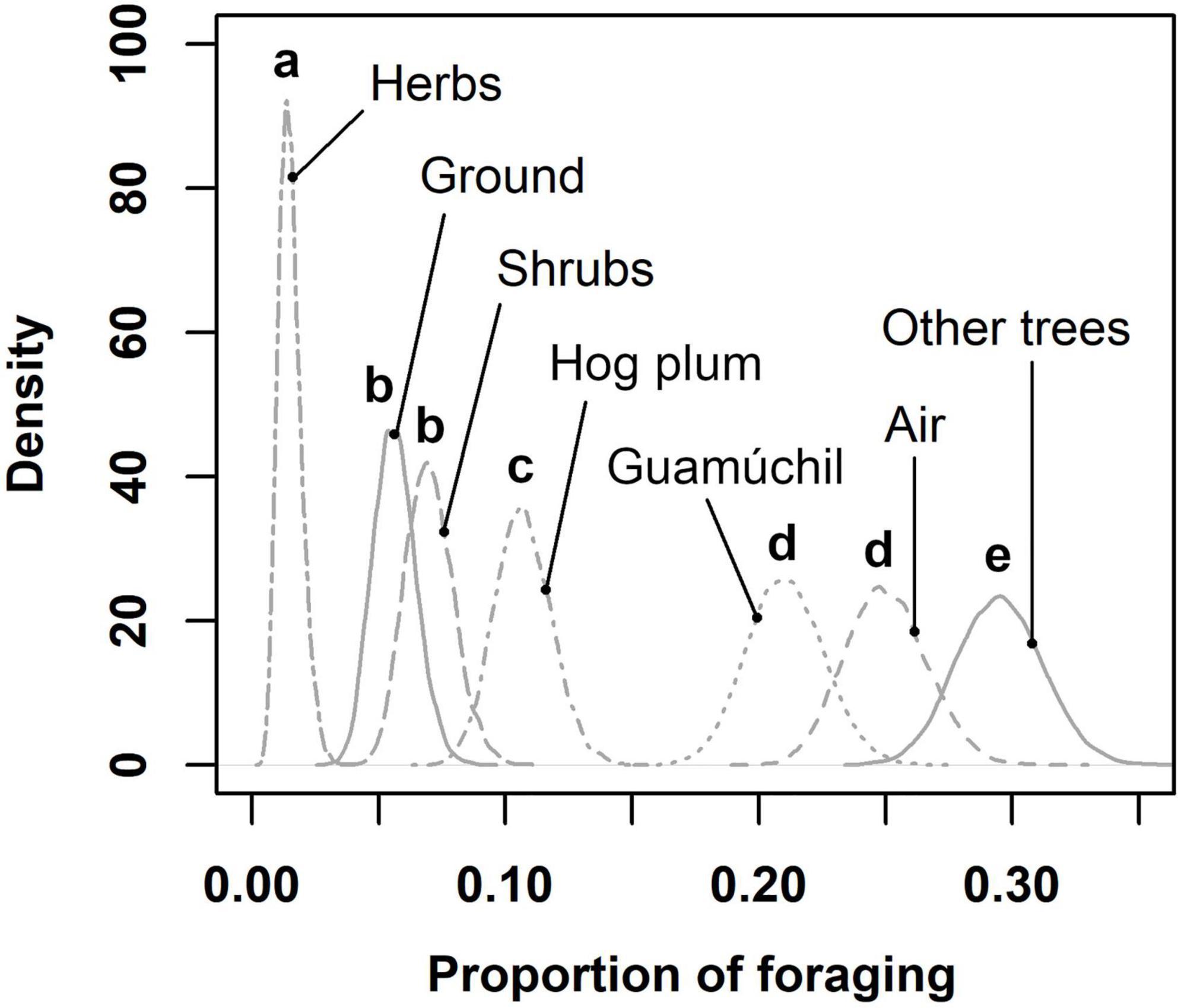
Figure 2. Estimated proportion of bird foraging for different types of substrates in Zacualpan, Colima, Mexico. Letters above curves depict statistical differences among estimations.
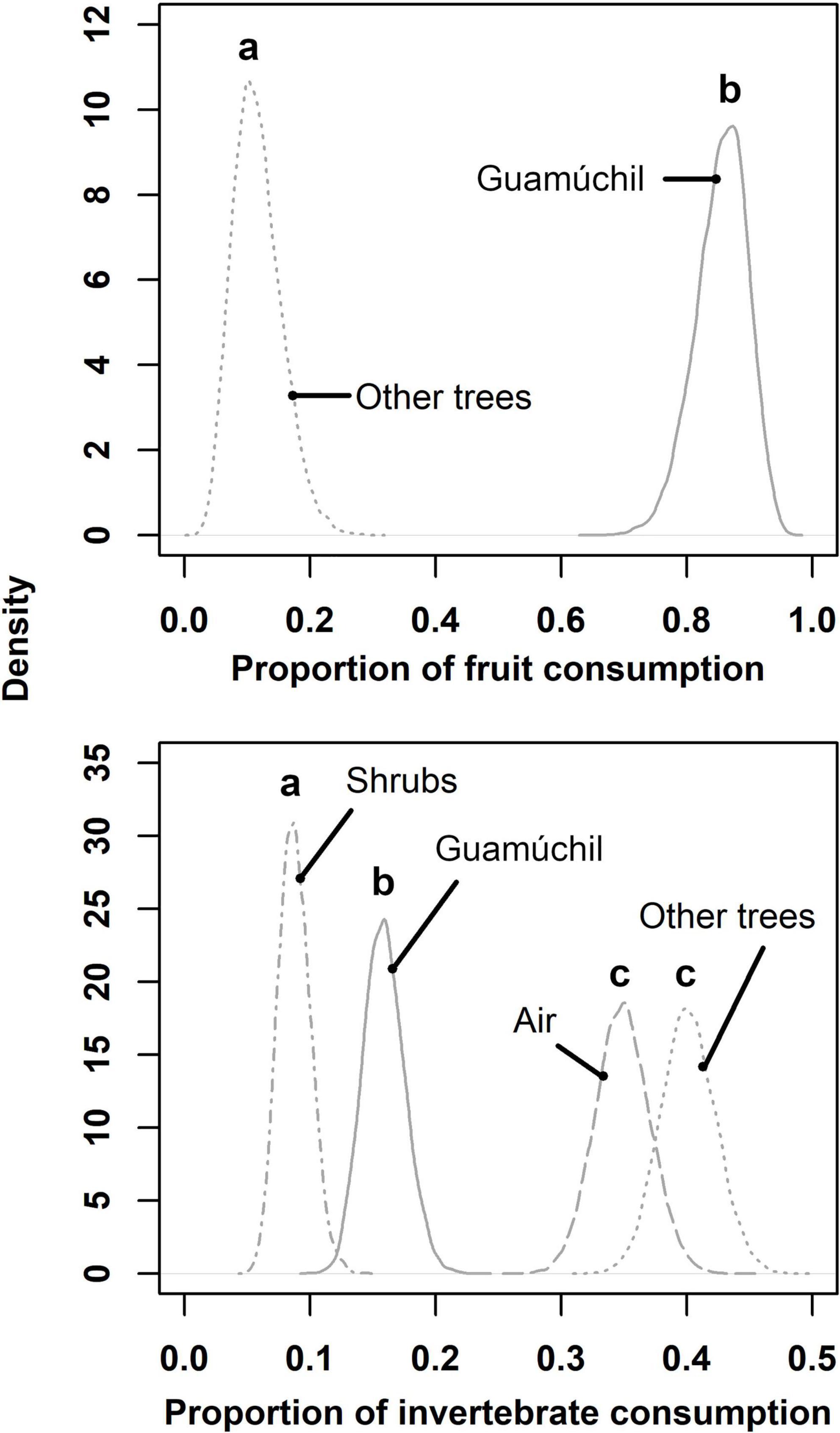
Figure 3. Estimated proportions of fruits and invertebrates consumed by birds for distinct kinds of feeding sources in Zacualpan, Colima, Mexico. Letters above curves depict statistical differences among estimations.
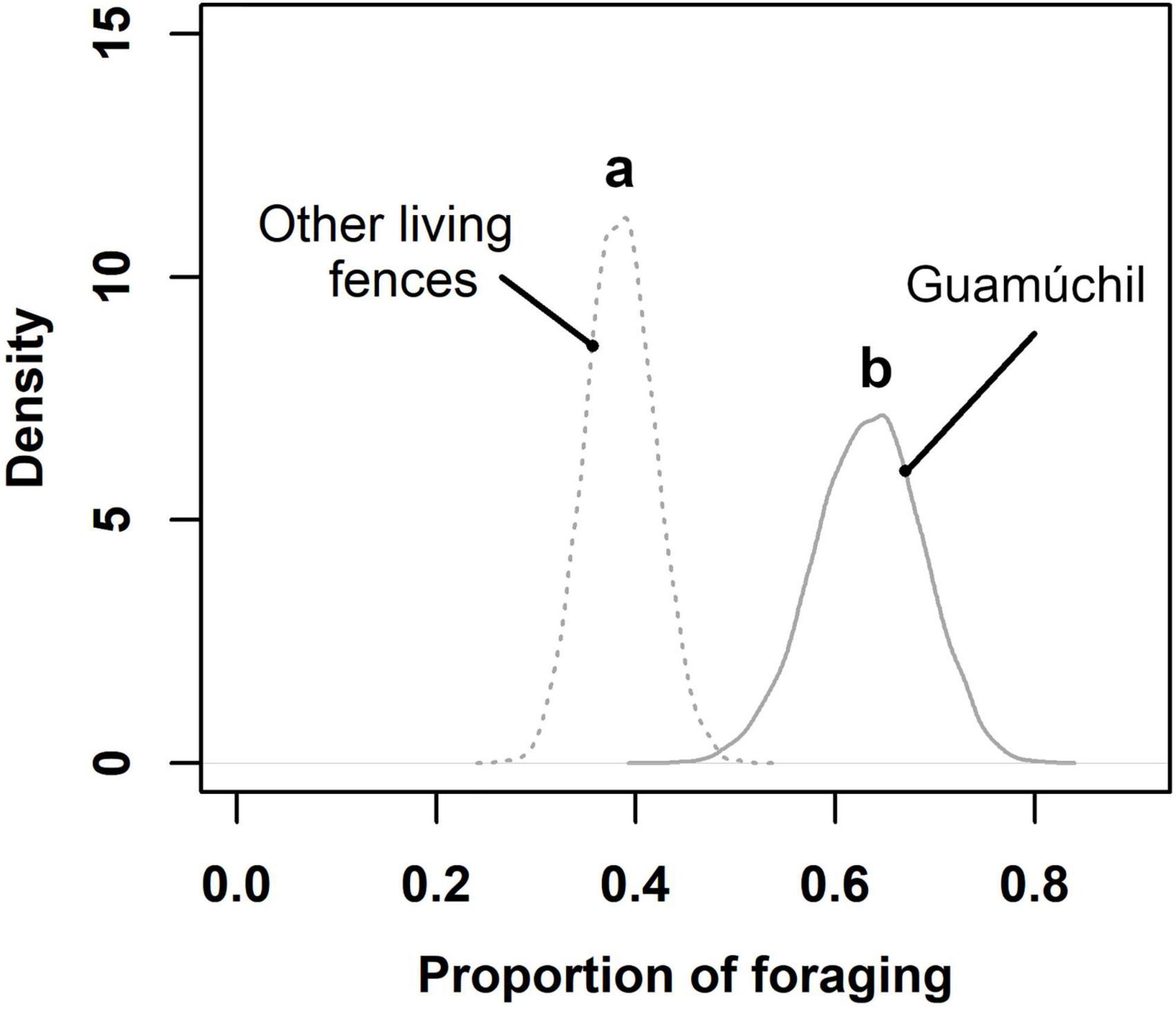
Figure 4. Estimated proportion of bird foraging for different types of live fences in Zacualpan, Colima, Mexico. Letters above curves depict statistical differences among estimations. Other live fences included both shrubs and non-Guamúchil trees.
Discussion
The Guamúchil is regularly present in the urban settlement of the indigenous community. We observed that homegardens could be covered with such tree, benefiting not only humans, but birds that feed on it. A previous study in the site has shown that about 49 bird species specifically forage within homegardens (Ortega-Álvarez and Casas, under review), which might be heavily attracted by fruiting trees. Given that we determined that Guamúchil cover is naturally low in the surrounding tropical dry forest, homegardens might represent an important source of such tree to birds. In this way, birds might benefit by the local use and management of a salient biocultural food resource.
According to our estimations, about a fourth of the bird species that foraged across the cultural landscape might feed on Guamúchil. As suggested by other studies (Torres-González et al., 2014), this highlights the relevance of the tree to the avian community, given that a single arboreal species might provide food to several bird species. Such result is consistent with our foraging substrate analysis, in which we determined that local birds mostly relied on trees as feeding substrates. In particular, our results suggested that Guamúchils were less used than other trees as avian foraging substrates (0.21 and 0.29 foraging proportion, respectively). However, other trees might include about 82 species that are potentially present in the region (Padilla-Velarde et al., 2006). Consequently, the Guamúchil alone might harbor a bird foraging activity that is almost similar to that associated with multiple trees. Hence, Guamúchil must be considered as a key substrate to the avian foraging activity in the study site.
We identified that Guamúchil provided two types of food to birds: invertebrates and fruit. Although birds might prey on the invertebrates associated with this tree (Torres-González et al., 2014), we showed that Guamúchil was mostly important to local avian foraging for the seed pods that it produces. Fruit is a valuable but a spatial-temporal unpredictable food to birds in natural habitats (Tellería et al., 2005). However, local indigenous practices might foster its availability by cultivating Guamúchil, especially in homegardens. In tropical disturbed forests, frugivore birds are less abundant than in intact mature forest (Gray et al., 2007; Almazán-Núñez et al., 2015). Still, increased cover of urban Guamúchils in Zacualpan might enhance fruit abundance and attract frugivores (Alvarez-Alvarez et al., 2018), even to anthropogenic habitats where human related disturbances reach its highest point.
The fruit of Guamúchil trees is an essential source of vitamins, amino acids, fiber, and minerals (Murugesan et al., 2019), which might result attractive to different types of birds. For instance, we observed that species associated to mature forests, such as Orange-fronted Parakeets (Almazán-Núñez et al., 2015, 2021), visited homegardens to feed on the fruit. Additionally, Western Tanager, a boreal overwintering species, frequently fed on the fruit of Guamúchil, possibly because ubiquitous fruit decreases foraging energetic costs and might provide energy for migration (McWilliams and Karasov, 2001). Other generalist species, like Golden-cheeked Woodpeckers and White-throated Magpie-Jays, also regularly consumed this fruit, with potential implications for seed dispersal and forest succession (Almazán-Núñez et al., 2021).
There is evidence that live fences might function as corridors that connect forest fragments, enhancing avian movements across the landscape (Chacón León and Harvey, 2006; Francesconi et al., 2011; Pulido-Santacruz and Renjifo, 2011). Also, these agroforestry practices increase the habitat, resources, and diversity of bird communities in human dominated systems (Estrada et al., 2000; Harvey et al., 2005; Maglianesi, 2021). Our results add evidence to the benefits associated with live fences to wildlife because we observed that they have positive impacts on bird access to food. Besides acting as steppingstones, live fences of Guamúchil might be functioning as complementary food sources to birds in the fragmented landscape of Zacualpan (Harvey et al., 2005). Hence, using fruiting trees as live fences might have relevant implications on bird use and survival within cultural landscapes.
Conclusion
Our research shows how humans and birds share key feeding resources within cultural landscapes. Moreover, we depict how indigenous practices positively impact wildlife foraging. Identifying key foods and analyzing their management might facilitate the development of both in-situ (e.g., reforestation activities, use of live fences, home gardening) and ex-situ activities (e.g., propagation of tree species in nurseries, creation of seed banks) aimed to benefiting both humans and bird communities. We demonstrated that using fruiting trees as live fences provide different human benefits and aid avian foraging. Such agroforestry practices should be promoted as land management tools to foster the conservation and restoration of nature and cultural identity, under an agroforestry sustainability scope (Farina, 2018). Further research should be performed to determine the relevance of Guamúchil to birds during its non-fruiting season.
Data availability statement
The original contributions presented in this study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author/s.
Ethics statement
Ethical review and approval was not required for the study on human participants in accordance with the local legislation and institutional requirements. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements. Ethical review and approval was not required for the animal study because we did not manipulate animals. Our research was not experimental, but observational.
Author contributions
RO-Á and AC conceived the study, interpreted the data, and wrote the manuscript. RO-Á and AP-F collected the data. RO-Á analyzed the data. All authors approved the final version of the manuscript.
Funding
This work was funded with the projects PAPIIT, DGAPA, UNAM (IN206520) and CONACYT (A1-S-14306). We acknowledged the Posgrado en Ciencias Biológicas, UNAM, and the Consejo Nacional de Ciencia y Tecnología (CONACYT) for supporting AP-F Ph.D. scholarship. Authors recognize the aid provided to RO-Á by the Programa de Becas Posdoctorales DGAPA-UNAM 2021-2022.
Acknowledgments
We thank the community of Zacualpan for supporting the development of this study. In particular, we acknowledge J. Santos, A. Santos, D. Zamora, and F. Domínguez, as well as all the community members who participated in the interviews. We are grateful to F. Estañol Tecuatl, S. Rangel, and D. Paz for their fieldwork assistance, and E. Ruiz for his aid on plant identification. We acknowledge the comments and suggestions provided by the reviewers, as they enhanced the quality of our manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2022.1020207/full#supplementary-material
References
Acevedo, M. A. (2007). Bird feeding behavior as a measure of restoration success in a Caribbean forested wetland. Ornitol. Neotrop. 18, 305–310. doi: 10.3390/su12083493
Akinola, R., Pereira, L. M., Mabhaudhi, T., de Bruin, F. M., and Rusch, L. (2020). A review of indigenous food crops in Africa and the implications for more sustainable and healthy food systems. Sustainability 12, 1–30. doi: 10.3390/SU12083493
Almazán-Núñez, R. C., Alvarez-Alvarez, E. A., Sierra-Morales, P., and Rodríguez-Godínez, R. (2021). Fruit size and structure of zoochorous trees: identifying drivers for the foraging preferences of fruit-eating birds in a mexican successional dry forest. Animals 11:3343. doi: 10.3390/ani11123343
Almazán-Núñez, R. C., Arizmendi, M., del, C., Eguiarte, L. E., and Corcuera, P. (2015). Distribution of the community of frugivorous birds along a successional gradient in a tropical dry forest in south-western Mexico. J. Trop. Ecol. 31, 57–68. doi: 10.1017/S0266467414000601
Alvarez-Alvarez, E. A., Corcuera, P., and Almazán-Núñez, R. C. (2018). Spatiotemporal variation in the structure and diet types of bird assemblages in tropical dry forest in southwestern Mexico. Wilson J. Ornithol. 130, 457–469.
Baath, R. (2014). “Bayesian first aid: A package that implements Bayesian alternatives to the classical test functions in R,” in Proceedings of the UseR! 2014-the international r user conference, Los Angeles, CA.
Berkes, F., Colding, J., and Folke, C. (2000). Rediscovery of traditional ecological knowledge as adaptive management. Ecol. Appl. 10, 1251–1262. doi: 10.1890/1051-0761(2000)010[1251:ROTEKA]2.0.CO;2
Bibby, C., Burgess, N., Hill, D., and Mustoe, S. (2000). Bird Census Techniques. Great Britain: Academic Press.
Casas, A., Caballero, J., Mapes, C., and Zárate, S. (1997). Manejo de la vegetación, domesticación de plantas y origen de la agricultura en Mesoamérica. Bot. Sci. 61, 31–47. doi: 10.17129/botsci.1537
Casas, A., Otero-Arnaiz, A., Pérez-Negrón, E., and Valiente-Banuet, A. (2007). In situ management and domestication of plants in Mesoamerica. Ann. Bot. 100, 1101–1115. doi: 10.1093/aob/mcm126
Casas, A., Parra, F., Blancas, J., Rangel-Landa, S., Vallejo, M., Figueredo, C. J., et al. (2016). “Origen de la domesticación y la agricultura: cómo y por qué,” in Domesticación en el Continente Americano. Manejo de biodiversidad y evolución dirigida por las culturas del Nuevo Mundo, eds A. Casas, J. Torres-Guevara, and F. Parra (Mexico: UNAM-UNALM), 189–224. doi: 10.22201/iies.0000001p.2016
Chacón León, M., and Harvey, C. A. (2006). Live fences and landscape connectivity in a neotropical agricultural landscape. Agroforest. Syst. 68, 15–26. doi: 10.1007/s10457-005-5831-5
Chinlampianga, M., Singh, R. K., and Shukla, A. C. (2013). Ethnozoological diversity of Northeast India: empirical learning with traditional knowledge holders of Mizoram and Arunachal Pradesh. Indian J. Tradit. Knowl. 12, 18–30.
Clement, C. R., Casas, A., Parra-Rondinel, F. A., Levis, C., Peroni, N., Hanazaki, N., et al. (2021). Disentangling domestication from food production systems in the Neotropics. Quaternay 4:4. doi: 10.3390/quat4010004
de Paula, M. J., Xerente, V. S., Silva, A. A. F., Godoy, B. S., and Pezzuti, J. C. B. (2018). Collaborative research and the hunting in the Brazilian Cerrado: the case of Xerente Indigenous Land. Biota Neotrop. 18:e20180556. doi: 10.1590/1676-0611-bn-2018-2556
Droste, B. V., Plachter, H., and Rossler, M. (1995). Cultural Landscapes of Universal Value. New York, NY: UNESCO.
Estrada, A., Cammarano, P. L., and Coates-Estrada, R. (2000). Bird species richness in vegetation fences and in strips of residual rain forest vegetation at Los Tuxtlas, Mexico. Biodivers. Conserv. 9, 1399–1416. doi: 10.1023/A:1008935016046
Farina, A. (2000). The cultural landscape as a model for the integration of ecology and economics. BioScience 50, 313–320. doi: 10.1641/0006-3568(2000)050[0313:TCLAAM]2.3.CO;2
Farina, A. (2018). Rural sanctuary: an ecosemiotic agency to preserve human cultural heritage and biodiversity. Biosemiotics 11, 139–158. doi: 10.1007/s12304-018-9319-x
Francesconi, W., Montagnini, F., and Ibrahim, M. (2011). “Using bird distribution to evaluate the potential of living fences to restore landscape connectivity in pasturelands,” in Agroforestry as a Tool for Landscape Restoration, eds F. Montagnini, W. Francesconi, and E. Rossi (New York, NY: Nova Science Publishers), 133–142. doi: 10.1057/978-1-349-96042-2_1048
Gelman, A., and Rubin, D. B. (1992). Inference from iterative simulation using multiple sequences. Stat. Sci. 7, 457–511. doi: 10.1214/ss/1177011136
Gfeller, A. E. (2013). Negotiating the meaning of global heritage:‘cultural landscapes’ in the UNESCO World Heritage Convention, 1972-92. J. Glob. Hist. 8, 483–503. doi: 10.1017/S1740022813000387
Gray, M. A., Baldauf, S. L., Mayhew, P. J., and Hill, J. K. (2007). The response of avian feeding guilds to tropical forest disturbance. Conserv. Biol. 21, 133–141. doi: 10.1111/j.1523-1739.2006.00557.x
Grupo Xolocuahuitl Zacualpan (2020). Comida Tradicional Rural de la Comunidad Indígena de Zacualpan, Comala, Colima. Colima: Gobierno del Estado, PACMYC.
Harvey, C. A., Villanueva, C., Villacís, J., Chacón, M., Muñoz, D., López, M., et al. (2005). Contribution of live fences to the ecological integrity of agricultural landscapes. Agric. Ecosyst. Environ. 111, 200–230. doi: 10.1016/j.agee.2005.06.011
Hsieh, T. C., Ma, K. H., and Chao, A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 7, 1451–1456. doi: 10.1111/2041-210X.12613
Jernigan, K., and Dauphiné, N. (2008). Aguaruna knowledge of bird foraging ecology: a comparison with scientific data. Ethnobot. Res. Appl. 6, 93–106. doi: 10.17348/era.6.0.93-106
Kruschke, J. K. (2015). Doing Bayesian Data Analysis: A Tutorial with R, JAGS, and STAN, 2nd Edn. Cambridge, MA: Academic Press. doi: 10.1016/B978-0-12-405888-0.00008-8
Levis, C., Costa, F. R. C., Bongers, F., Peña-Claros, M., Clement, C. R., Junqueira, A. B., et al. (2017). Persistent effects of pre-Columbian plant domestication on Amazonian forest composition. Science 355, 925–931. doi: 10.1126/science.aal0157
Luat-Hu’eu, K. K., Winter, K. B., Vaughan, M. B., Barca, N., and Price, M. R. (2021). Understanding the co-evolutionary relationships between Indigenous cultures and non-native species can inform more effective approaches to conservation: the example of pigs (pua’a; Sus scrofa) in Hawai’i. Pacific Conserv. Biol. 27, 442–450. doi: 10.1071/PC20086
Maffi, L., and Woodley, E. (2010). Biocultural Diversity Conservation. A Global Sourcebook. London: Earthscan.
Maglianesi, M. A. (2021). Live fences have greater diversity of bird assemblages than gallery forests in human-modified ecosystems. Ornitol. Neotrop. 32, 68–74.
McWilliams, S. R., and Karasov, W. H. (2001). Phenotypic flexibility in digestive system structure and function in migratory birds and its ecological significance. Comp. Biochem. Physiol. 128, 579–593. doi: 10.1016/s1095-6433(00)00336-336
Mejía, N. (2021). Montañas de Colima, Una Cocina de Altura. Colima. Una gran Travesía Gastronómica. Tomo III. México: Nicolás Mejía.
Monroy, R., and Colín, H. (2004). El guamúchil Pithecellobium dulce (Roxb.) Benth, un ejemplo de uso múltiple. Madera Bosques 10, 35–53. doi: 10.21829/myb.2004.1011278
Moreno-Calles, A. I., Toledo, V. M., and Casas, A. (2013). Los sistemas agroforestales tradicionales de México: una aproximación biocultural. Bot. Sci. 91, 375–398. doi: 10.17129/botsci.419
Murugesan, S., Lakshmanan, D. K., Arumugam, V., and Alexander, R. A. (2019). Nutritional and therapeutic benefits of medicinal plant Pithecellobium dulce (Fabaceae): a review. J. Appl. Pharm. Sci. 9, 130–139. doi: 10.7324/JAPS.2019.90718
Nakashima, D., and Roué, M. (2002). “Indigenous knowledge, peoples and sustainable practice,” in Encyclopedia of Global Environmental Change, ed. P. Timmerman (Chichester: John Wiley & Sons, Ltd), 314–324.
Ojeda-Linares, C., Álvarez-Ríos, G. D., Figueredo-Urbina, C. J., Islas, L. A., Lappe-Oliveras, P., Nabhan, G. P., et al. (2021). Traditional fermented beverages of Mexico: a biocultural unseen foodscape. Foods 10:2390. doi: 10.3390/foods10102390
Ortega-Álvarez, R., and Casas, A. (under review). “Las aves están en el monte, no en el pueblo”: Percepción comunitaria sobre la riqueza aviar asociada a los traspatios de zacualpan, colima, (México). El Hornero
Ortega-Álvarez, R., and Casas, A. (2022). The feeding landscape: bird and human use of food resources across a biocultural landscape of the Colombian Andes. Sustainability 14:4789.
Ortega-Álvarez, R., Ruiz-Gutiérrez, V., Robinson, O. J., Berrones Benítez, E., Medina Mena, I., and Zúñiga-Vega, J. J. (2021). Beyond incidence data: assessing bird habitat use in indigenous working landscapes through the analysis of behavioral variation among land uses. Landsc. Urban Plan. 211:104100. doi: 10.1016/j.landurbplan.2021.104100
Padilla-Velarde, E., Cuevas-Guzmán, R., Ibarra-Manríquez, G., and Moreno-Gómez, S. (2006). Riqueza y biogeografía de la flora arbórea del estado de Colima. México. Rev. Mex. Biodivers. 77, 271–295. doi: 10.22201/ib.20078706e.2006.002.337
Pennington, T. D., and Sarukhán, J. (2005). Árboles Tropicales de México. Manual Para la Identificación de las Principales Especies. Ciudad de México: Universidad Nacional Autónoma de México, Fondo de Cultura Económica.
Perfecto, I., and Vandermeer, J. (2008). Biodiversity conservation in tropical agroecosystems. a new conservation paradigm. Ann. N. Y. Acad. Sci. 1134, 173–200. doi: 10.1196/annals.1439.011
Perfecto, I., Vandermeer, J., and Wright, A. (2009). Nature’s Matrix: Linking Agriculture, Conservation and Food Sovereignty. London: Earthscan. doi: 10.4324/9781849770132
Plummer, M. (2003). “JAGS: a program for analysis of Bayesian graphical models using Gibbs sampling,” in Proceedings of the 3rd International Workshop on Distributed Statistical Computing, eds K. Hornik, F. Leisch, and Z. Achim (Vienna).
Pulido-Santacruz, P., and Renjifo, L. M. (2011). Live fences as tools for biodiversity conservation: a study case with birds and plants. Agroforest. Syst. 81, 15–30. doi: 10.1007/s10457-010-9331-x
Robbins, M. C., Nolan, J. M., and Chen, D. (2017). An improved measure of cognitive salience in free listing tasks: a Marshallese example. Field Methods 29, 395–403. doi: 10.1177/1525822X17726726
Ruiz Gutiérrez, V., Berlanga García, H. A., Calderón-Parra, R., Savarino Drago, A., Aguilar-Gómez, M. Á, and Rodríguez-Contreras, V. (2020). Manual Ilustrado Para el Monitoreo de Aves. Proalas: Programa de América Latina para las Aves Silvestres. Ciudad de México: Comisión Nacional para el Conocimiento y Uso de la Biodiversidad/Iniciativa para la Conservación de las Aves de Norte América, Laboratorio de Ornitología de Cornell.
SEMARNAT (2010). Norma Oficial Mexicana NOM- 059- ECOL- 2010, Protección Ambiental- Especies Nativas de México de Flora y Fauna Silvestres- Categorías de Riesgo y Especificaciones Para su Inclusión, Exclusión o Cambio- Lista de Especies en Riesgo. México: Diario Oficial de la Federación, Secretaría de Medio Ambiente y Recursos Naturales.
Smith, J. J., and Borgatti, S. P. (1997). Salience counts—and so does accuracy: correcting and updating a measure for free-list-item salience. J. Linguist. Anthropol. 7, 208–209. doi: 10.1525/jlin.1997.7.2.208
Sutrop, U. (2001). List task and a cognitive salience index. Field Methods 13, 263–276. doi: 10.1177/1525822X0101300303
Tellería, J. L., Ramírez, Á, and Pérez-Tris, J. (2005). Conservation of seed-dispersing migrant birds in Mediterranean habitats: shedding light on patterns to preserve processes. Biol. Conserv. 124, 493–502. doi: 10.1016/j.biocon.2005.02.011
Thompson, K. L., Hill, C., Ojeda, J., Ban, N. C., and Picard, C. R. (2020). Indigenous food harvesting as social-ecological monitoring: a case study with the Gitga’at First Nation. People Nat. 2, 1085–1099. doi: 10.1002/pan3.10135
Tidemann, S., and Gosler, A. G. (2010). Ethno-ornithology: Birds, Indigenous Peoples, Culture and Society. London: Earthscan.
Torres-González, A. M., Vargas-Figueroa, J. A., Guevara-Ibarra, L., Llano-Almario, M., Orrego-Pineda, J. O., Duque-Palacio, O. L., et al. (2014). Use of Samanea saman and Pithecellobium dulce (Fabaceae: Mimosoideae) by birds in the University Botanical Garden, Cali, Colombia. Rev. Ciencias 18, 63–78.
Turner, N. J., and Bhattacharyya, J. (2016). Salmonberry bird and goose woman: birds, plants, and people in indigenous peoples lifeways in Northwestern North America. J. Ethnobiol. 36, 717–745. doi: 10.2993/0278-0771-36.4.717
UNESCO (2003). Cultural Landscapes: the Challenges of Conservation. Ferrara: UNESCO World Heritage Centre.
Wagner, J. L. (1981). Visibility and bias in avian foraging data. Condor 83, 263–264. doi: 10.2307/1367320
Wencelius, J., Garine, E., and Raimond, C. (2017). FLARES. Free List Analysis Under R Environment Using Shiny. Available online at: http://www.anthrocogs.com/shiny/flares/ (accessed August 1, 2022).
Keywords: ethnoecology, feeding, homegardens, indigenous community, live fence, Pithecellobium dulce, tropical dry forest, behavior
Citation: Ortega-Álvarez R, Pacheco-Flores A and Casas A (2022) The “Guamúchil” cultivation in a Mexican cultural landscape: A wild food source for people and birds. Front. For. Glob. Change 5:1020207. doi: 10.3389/ffgc.2022.1020207
Received: 15 August 2022; Accepted: 31 August 2022;
Published: 16 September 2022.
Edited by:
Renata Sõukand, Ca’ Foscari University of Venice, ItalyReviewed by:
Almo Farina, University of Urbino Carlo Bo, ItalyRaivo Kalle, University of Gastronomic Sciences, Italy
Copyright © 2022 Ortega-Álvarez, Pacheco-Flores and Casas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rubén Ortega-Álvarez, cnViZW5vcnRlZ2EuYWxAZ21haWwuY29t