 Daniel D. Kneeshaw1*
Daniel D. Kneeshaw1* Louis De Grandpré2
Louis De Grandpré2 Loïc D’Orangeville3
Loïc D’Orangeville3 Maryse Marchand2
Maryse Marchand2 Jeanne Moisan-Perrier1
Jeanne Moisan-Perrier1 Louis-Etienne Robert1Mathieu Bouchard4
Louis-Etienne Robert1Mathieu Bouchard4
- 1Center for Forest Research, Université du Québec à Montréal, Montréal, QC, Canada
- 2Natural Resources Canada, Canadian Forest Service, Sainte-Foy, QC, Canada
- 3Faculty of Forestry and Environmental Management, University of New Brunswick, Fredericton, NB, Canada
- 4Faculté de Foresterie et Géomatique, Université de Laval, Sainte-Foy, QC, Canada
Balsam fir (Abies balsamea) is the most vulnerable species to the spruce budworm (Choristoneura fumiferana), one of the most devastating defoliators in the world. For decades, pest managers have advocated for reducing its abundance in the landscape to minimize losses to the spruce budworm (SBW). Although reduction of fir occurred during the endemic phase of the SBW cycle, there is little information about the extent to which this general principle (reduction of fir) was applied during an outbreak and whether it occurs at both stand and landscape levels. The objective of this paper is to compare the effect of insect and harvest disturbances on forest structure during the 1970–80s outbreak in Québec. We evaluate whether, (i) forest management activities targeted fir forests and whether patch size of host species influences management or SBW disturbance, (ii) SBW outbreaks and logging have similar or divergent effects on forest composition. Although data are from an earlier outbreak, they are at a scale rarely studied and will be useful in guiding decisions made at larger scales in the current and future outbreaks. Our results show that spruce was targeted preferentially by harvesting (up to 69% of plots) during the outbreak period, while it represented less than one third of plots defoliated by the SBW. On the other hand, fir stands represented up to 75% of plots that were defoliated by the SBW but less than 35% of plots that underwent harvesting. Harvesting targeted large blocks of spruce forest more than large blocks of fir-dominated forest while the opposite was observed for the SBW. In terms of regeneration, SBW tends to reduce fir and favor spruce recruitment, along with non-host species, whereas the opposite tendency was observed following harvesting. In terms of spatial organization of stands, our results support the suggestion that small stands of fir and large stands of spruce undergo the least SBW damage. Thus, in order to attenuate SBW impacts in the future, efforts should be made to ensure that spruce recruitment is favored and that its abundance increases at both the stand and landscape scale.
Introduction
Managing forests regulated by large-scale insect outbreaks, such as the spruce budworm (SBW) [Choristoneura fumiferana (Clem.)], comes with a unique set of challenges. SBW is one of the most damaging outbreaking insects in the boreal and sub-boreal forests of eastern North America (Blais, 1983; Kneeshaw and Bergeron, 2015). SBW populations reach outbreak levels synchronously over large spatial scales, with an average periodicity of 30–40 years (Boulanger and Arseneault, 2004; Kneeshaw and Bergeron, 2015). Whereas at endemic levels no visible damage is caused, during outbreak periods, repeated defoliation over 7–15 years results in major mortality events in areas dominated by balsam fir (Abies balsamea) or spruces (Picea spp) (MacLean, 1980). One of the most severe SBW outbreaks of the last century, which occurred between 1966 and 1992, affected more than 55 million ha of the eastern boreal forest and caused the loss of over 200 million m3 of balsam fir and spruce (Boulet et al., 1996). Annual losses in forest productivity of up to 91% were estimated during peak outbreak years (Sterner and Davidson, 1982; Volney and Fleming, 2007). This outbreak caused more than ten consecutive years of defoliation in some regions, resulting in moderate to high mortality (Kneeshaw and Bergeron, 2015; Sainte-Marie et al., 2015).
SBW defoliate some tree species more severely than others, and thus forest composition influences the severity of the disturbance (Bognounou et al., 2017). Detailed work at the stand scale shows that balsam fir, the primary host of the SBW, is almost twice as vulnerable as black spruce (Picea mariana) (a secondary host) due to the synchronization of budburst phenology with emergence of the SBW in the spring (Hennigar et al., 2008; Pureswaran et al., 2019). Others have demonstrated that stands where host species (fir and spruces) are mixed with hardwoods (non-hosts) are less vulnerable to defoliation and mortality caused by the SBW (Su et al., 1996; Cappuccino et al., 1998). Similar work shows that although site factors influence outbreak initiation, composition is the main variable determining mortality (Dupont et al., 1991; Bergeron et al., 1995; Bouchard and Auger, 2014). Based on this stand level information MacLean (2004) suggested that it would make ecological sense to promote the management of complex mixed-species stands that would otherwise be rendered homogenous by clearcutting and subsequent tending (Belle-Isle and Kneeshaw, 2007). Although stand-level influences are better understood than landscape-level ones, recent work from outside the main range of the insect suggests that landscapes dominated by the primary host species undergo more severe and synchronized outbreaks (Robert et al., 2012, 2018).
While it remains challenging to clearly identify and quantify all processes that influence tree mortality during insect outbreaks, such as climate or parasitoid-prey interaction (De Grandpré et al., 2019), outbreak severity has been suggested to increase with the availability of large, well-connected areas of susceptible host trees (Baskerville, 1975; Bentz et al., 2020; Kneeshaw et al., 2021). The role of host abundance, spatial extent, and connectivity have been consistently suggested to drive outbreaks in both the SBW-forest system (Robert et al., 2018) and other insect-forest systems in North America and Europe (Hansen et al., 2016; Robert et al., 2020). The abundance and proportion of contiguous tracts of host forest is directly influenced by human forest management. Forest management techniques over the past century have in fact been implicated in the observed increase in outbreak severity (Blais, 1983; Kneeshaw et al., 2021). For example, total harvests that protects advance regeneration (i.e., a clear cut with restrictions), the favored large-scale forestry technique, increases the abundance of the primary host species of the SBW (Kneeshaw and Bergeron, 2015). In North America, many commercially managed forests are more homogeneous in terms of patch size, stand age and species composition than in forests recruited following natural processes (Gauthier et al., 2009). Nonetheless, despite comparisons of harvesting to fire effects on forest structure (McRae et al., 2001; Moussaoui et al., 2016; Jean et al., 2019), few studies have directly compared the influence of insect pest outbreaks and harvesting on ensuing forest structure. Our work thus attempts to fill this void. An analysis of landscape level effects of forest management will be critical to lower forest vulnerability especially at a forest management scale where agglomerated effects, that have rarely if ever been evaluated, can have unintended consequences.
Over recent decades, several authors have proposed that decreasing the abundance of large tracts of the primary host, balsam fir, is a tool that could be used in forest pest management to increase forest resistance to insect pests (MacLean, 2004; Kneeshaw et al., 2021). Sainte-Marie et al. (2015) went even further and suggested that forest management techniques that increase the abundance of the secondary host black spruce would reduce timber losses due to the SBW. Decision support tools have also been developed to identify forest stands at risk for which silvicultural interventions against the SBW should be targeted during endemic periods and reactive measures (spraying and salvage logging) during outbreaks (MacLean et al., 2001; Liu et al., 2019). However, there is a dearth of information on how silvicultural interventions used during outbreaks follow or deviate from interventions to reduce the dominance of the primary host balsam fir during outbreaks and whether increasing a secondary host species would effectively reduce SBW outbreak severity.
Following SBW outbreaks, natural forest dynamics lead to balsam fir recruitment of pure fir stands in maritime regions (Baskerville, 1975), but are a key disturbance leading to recruitment of other species in many other boreal forest regions (Batzer and Popp, 1985; Kneeshaw and Bergeron, 1998; Bouchard et al., 2006; Leduc et al., 2021). On the contrary, forest management techniques practices have been shown to increase the proportion of balsam fir throughout much of the eastern boreal forest. This increase is due to balsam fir’s well-developed understory seedling bank being released following harvesting (Kneeshaw and Bergeron, 1996; Belle-Isle and Kneeshaw, 2007; Barrette et al., 2019). These bodies of work suggest that recruitment may differ after SBW outbreaks and forest harvesting, but we still lack sufficient data.
The objectives of this paper are to evaluate whether forest management during SBW outbreaks modified forest structure and composition in the same way as SBW disturbance. We first evaluated whether different stand compositions of host species across a large region of the boreal forest were equally affected by harvesting and SBW outbreaks. Secondly, we assess whether there is a difference between the effects of outbreaks and harvesting on forest composition and thus future forest vulnerability. This information can ultimately be used to plan forest ecosystem management over the entire forest management period.
Materials and Methods
Study Area
The last SBW outbreak affected over 55 million ha of boreal forest in eastern North America. Affected forests are dominated by balsam fir (Abies balsamea L.) and white spruce [Picea glauca (Moench) Voss] mixed with white birch (Betula papyrifera Marsh.). These forests occur in three forest domains the balsam fir–yellow birch (Betula alleghaniensis), balsam fir-white birch and black spruce-feather moss domains (Figure 1; Saucier et al., 2010). Climate becomes colder and growing seasons shorter along the north-south axis that separates these domains and the proportion of black spruce tends to increase in the north. SBW is limited in the south by warm falls and in the north by short growing seasons (Régnière et al., 2012). These domains are divided into eastern and western sub-domains based on moisture differences with eastern sub-domains more maritime and western ones more continental. The more maritime gradients have a longer fire return interval and generally more balsam fir (Saucier et al., 1998). The climate is characterized by an annual average temperature of approximately 0°C and average annual precipitation that varies from 800 mm in the west to over 1,200 mm in the east with 40–45% falling as snow (Saucier et al., 1998; Grondin et al., 2003). During the last outbreak, all stands with host species in eastern Canada underwent some defoliation (Boulet et al., 1996). Trees usually die after 5–7 years of severe defoliation (MacLean, 1980; Pothier et al., 2012). When a stand is lightly defoliated, trees will only suffer some growth reduction and mortality will not affect the stand.
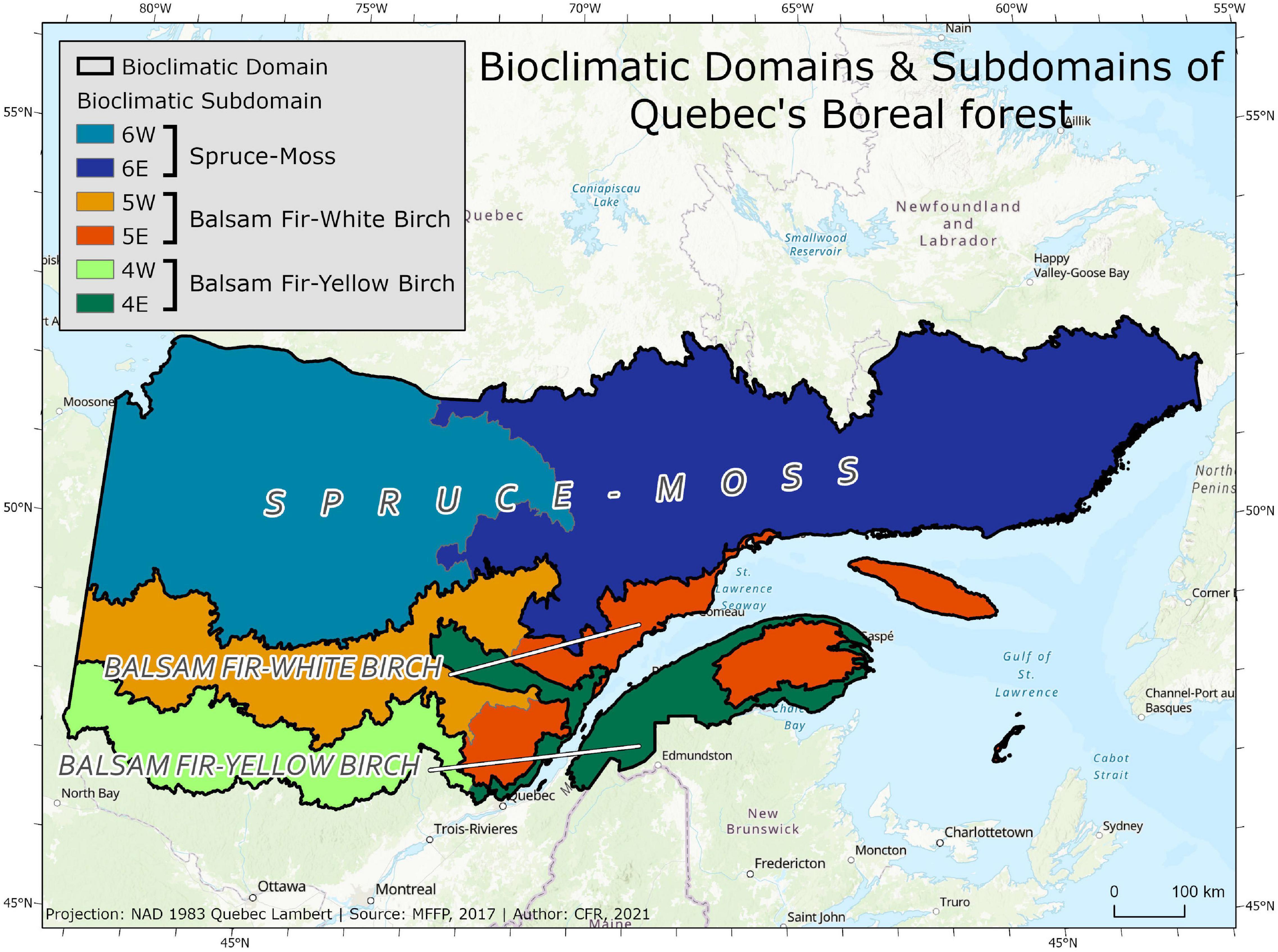
Figure 1. Sub-domains of spruce budworm-susceptible forests with abundant balsam fir and spruce species hosts across Quebec. The subdomains are used in the analyses reported in Table 1.
Approach
We used two distinct forest databases available for the entire commercial forest of Quebec to address our two objectives.1 To answer the first question of this study (described in detail in the following section), we assessed across the entire study area and in each of the sub-domains the proportion of vulnerable stand types that were harvested vs. ones defoliated by the SBW during the outbreak period using simplified forest inventory data derived from aerial photographs. To address the second question, we evaluated composition change data over time from the beginning to the end of the outbreak period in both harvested fir and spruce and SBW affected stands using a network of permanent sample plots (see section “Composition changes due to salvage logging and SBW” for details).
Objective 1 Forest Stand Type and Disturbance Susceptibility to Harvesting vs. Spruce Budworm
To assess the first objective, we used the SIFORT geospatial database (Spatial Information on Forest composition based on Tesserae) (Pelletier et al., 2007). The method, which has successfully been used in other regional forest assessments (Sainte-Marie et al., 2015; Liu et al., 2018, 2019), consists of superimposing a grid of 1,800 14 ha (15″ × 15″) tesserae on forest inventory maps, and assigning forest information to the center of each tesserae (Pelletier et al., 2007). For the purpose of this study, we extracted information related to forest stand (forest cover type, species groups and age class) and disturbance history (including harvest type and SBW outbreak severity). Cover type is classified as softwood, mixedwood or hardwood depending on dominant species. Species groups describes the dominant composition of the stand, i.e., the main species grouping in a stand based on proportion of basal area. SIFORT data is not continuous but rather based on classes. The advantage of this approach is that the same points are sampled through time and the entire forested area of Québec can be covered.
Data from the first inventory (1952–1978), prior to SBW-induced mortality, and from the second forest inventory (1979–1990), once outbreak-induced mortality had begun in most regions in Quebec, were used to assess changes in forest structure. In each of the six sub-domains, we selected SBW-susceptible forest, i.e., tesserae with mature stands where the dominant species were host species (balsam fir, spruce, or both) in the first inventory. To compare the proportion of susceptible stands affected by harvesting and SBW mortality during the 1970–1987 outbreak, we determined the proportion of tesserae (N = 1,453,852) that were affected either by the SBW or harvesting between the first and the second inventory. Disturbances were classified according to Quebec Ministry of Forests, Wildlife and Parks severity classes: partial disturbance (25–75% basal area removed; partial and salvage logging and light/moderate SBW-caused mortality) and total disturbance (≥75% basal area removed; clearcutting and severe SBW-caused mortality).
As a first step to evaluate if different stand types are equally affected by harvesting and SBW outbreaks, we used the Gallagher index, a “least squares index.” This index allows us to compare the impact of different disturbances in different stand types without being biased by unbalanced raw data. For each disturbance intensity (partial disturbance and total disturbance), the Gallagher index (LSq) was used to assess whether the proportion of tesserae that were harvested is similar to the proportion of tesserae affected by SBW disturbance (Gallagher, 1991; Gallagher and Mitchell, 2005).
Where V is the proportion of forests in a given composition type defoliated by the SBW and S is the proportion of forests of a given composition type that were harvested. The index weighs the deviations between harvested and SBW defoliated stand compositions such that the larger the differences between the observed disturbances (harvesting and SBW outbreak), the larger the Gallagher index. When based on percent data the index is bounded from 0 to 100. We used this index to compare the two severities of natural disturbance with the two severities of harvest disturbance. We performed this analysis for all susceptible forest types in eastern and western subdomains of the balsam fir-yellow birch, balsam fir-white birch and black spruce-feather moss domains and globally for the boreal forest i.e., all six subdomains.
We also assessed the occurrence of harvesting and SBW mortality by stand size and forest composition. We grouped tesserae having the same dominant cover and that were direct neighbors, i.e., that shared at least one of the four borders of a tesserae (from the first inventory, generally before the outbreak) and summed these neighboring tesserae to determine the patch size of a given cover type (balsam fir or spruce). We grouped stands into patch sizes of less than 100 ha, from 101 to 500 ha, from 501 to 1,000 ha and greater than 1,000 ha. The size of the first bin was based on maximum clearcut size and then followed principles in Freedman et al. (2007) with the largest size approximating a small forest management grouping unit in Québec (the Unité Territoiral de Référence). Then, we determined the proportion of harvesting (clearcutting) and SBW disturbance (SBW-caused mortality) in each patch and used this to determine the proportion per patch size class.
Objective 2: Composition Changes Due to Salvage Logging and Spruce Budworm
To compare changes in forest composition following logging and SBW we used a different data set that permitted us to evaluate species level changes. We used Quebec’s Ministry of Forests, Wildlife and Parks permanent sample plot network (Ministère des Forêts de la Faune et des Parcs, 2016) to characterize stand response to logging and SBW disturbance. We evaluated the proportion of stems by species or species class (balsam fir; black spruce and shade-intolerant hardwood) before and after each type of disturbance. We used the number of stems rather than basal area as slower growing species such as black spruce would not have reached a sufficient size to be evaluated by basal area and thus stem counts may provide a better indicator of future composition. However, we recognize that stem number measurements are not a perfect proxy of future composition. We compared unmanaged stands that sustained light defoliation by the SBW and managed stands that underwent salvage logging due to severe defoliation. We used 143 SBW host-dominated plots reported to have experienced logging during the last outbreak. Live merchantable (>9 cm DBH) stems and harvested stems were selected and grouped into three species categories: balsam fir, spruce [white, red (Picea rubens) and black spruce], and shade-intolerant hardwoods. To characterize impacts of light to moderate SBW disturbance, we selected 207 plots in host-dominated stands (>75% fir or spruce) that were not managed and that were first measured before or early in the course of the outbreak (i.e., at most 5 years after first reported defoliation and the second measurement period occurred after the outbreak had finished) (Ministère des Forêts de la Faune et des Parcs, 2019). For the first measurement period, plots not defoliated or defoliated for less than 5 years were used as mortality only begins to occur after five or more years of severe defoliation (Blais, 1958; MacLean, 1980) whereas peak mortality occurs 9–12 years after defoliation begins (Ministère des Forêts de la Faune et des Parcs, 2014). We selected and grouped live merchantable stems according to the categories described earlier. The proportion of dead or cut stems was then determined for each species group and disturbance type, allowing us to compare pre- and post-disturbance species composition. If the proportion of a given species at a site was the same before and after the disturbance (harvest or SBW) then the ratio would be 1. For example, if 40% of the total number of stems before a disturbance were balsam fir and 40% of the stems that recruited or survived after the disturbance (harvesting or SBW) were fir then the ratio was one and stand composition was not changed by the disturbance. Deviations from this condition (i.e., a ratio of one) are illustrated in box plots (Figure 2) to allow the reader to better assess the data and better evaluate the interpretations. This graphical approach, in which the reader sees the data spread associated with each of the treatment groups, was used given recent calls from statisticians to avoid the use of arbitrary significance based statistical tests and provide readers with results that permit an understanding of the robustness of interpretations (Wasserstein et al., 2019). These figures allow us to illustrate the proportion of stands for which a species increased, decreased or remained the same after the disturbance. We further evaluated how the distributions of species changed from a normal distribution (i.e., representing no change) and report on the degree of skewness as a complementary measure of change. Skewness between 0 and 0.5, represent relatively symmetrical data and thus little change, while moderately skewed data have skewness value between 0.5 and 1, and stands undergoing large changes in composition have skewness values greater than 1. We also fitted binomial regressions of the post-disturbance stand recruitment data to the pre-disturbance composition and compared the resulting slopes to a 1:1 slope where the post and pre-disturbance composition did not change. As these results in which both dependent and independent data are binomial are complementary to the skewness data and are less intuitive, we thus present them in supplementary material as support for the robustness of our interpretations (Supplementary Figure 1).
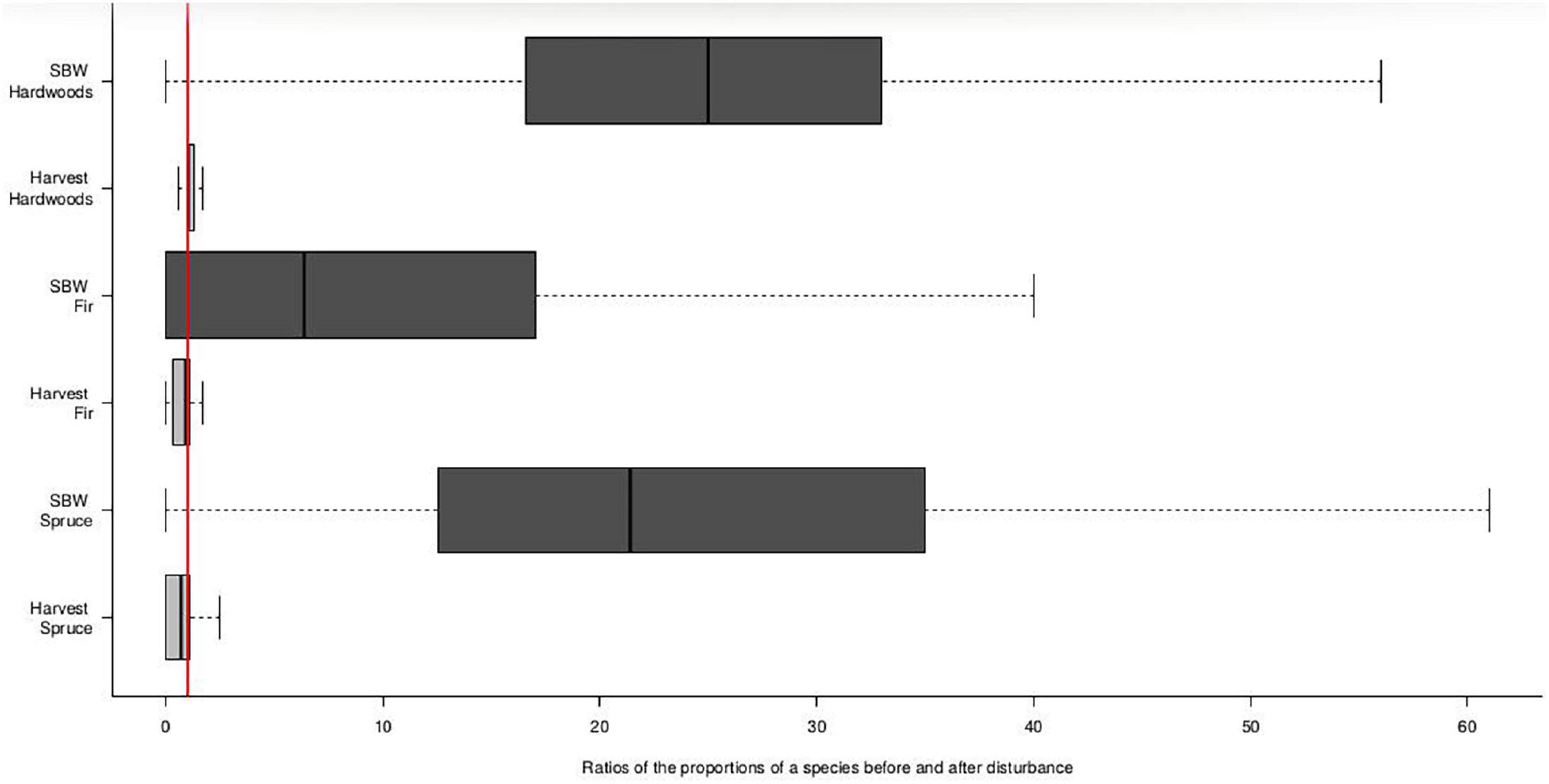
Figure 2. Proportion of composition change for three species groups from before to after a disturbance (light-moderate or severe SBW, and partial or total harvest). The red line indicates the 1:1 ratio where there is no change in composition before and after the disturbance. Proportions lower than the 1:1 line show a reduction in a species after the disturbance relative to before and higher values indicate an increased proportion. For example, hardwoods tend to increase following disturbance. Boxes represent the 25th and 75th percentiles with the middle line representing the median, bars show the 95th percentiles and points show extreme values.
Results
Disturbance Susceptibility per Forest Cover Type
The relative proportion of stand types affected by harvesting during an outbreak was found to differ from those affected by SBW (Table 1). We acknowledge that, since this is a retrospective analysis, the numbers should be interpreted with caution as defoliated stands that were already scheduled to be harvested would not be considered as salvage cuts, and that increased salvage harvesting will have modified the original harvest schedule. Keeping this caveat in mind the comparison to pre-disturbance (pre-SBW) proportions helps provide a gauge of the overall changes in targeted stands. Globally in the boreal forest, for partial disturbances, a larger fraction of fir stands was affected by SBW (52%) than by partial cuts (27%), consistent with the interpretation that harvesting targeted spruce stands more than light to moderate SBW outbreak (Gallagher Index = 23.5 for the global analysis). In the case of total disturbance, the disproportionality between clearcutting and severe SBW outbreak was even larger (Gallagher LSq = 45.4). Over 64% of the forest area that underwent severe SBW disturbance was fir-dominated, while only 29% was dominated by spruce. The results also show that the proportion of stands harvested by composition type tended to more closely follow the pre-disturbance stand proportions of host stands than did the SBW.
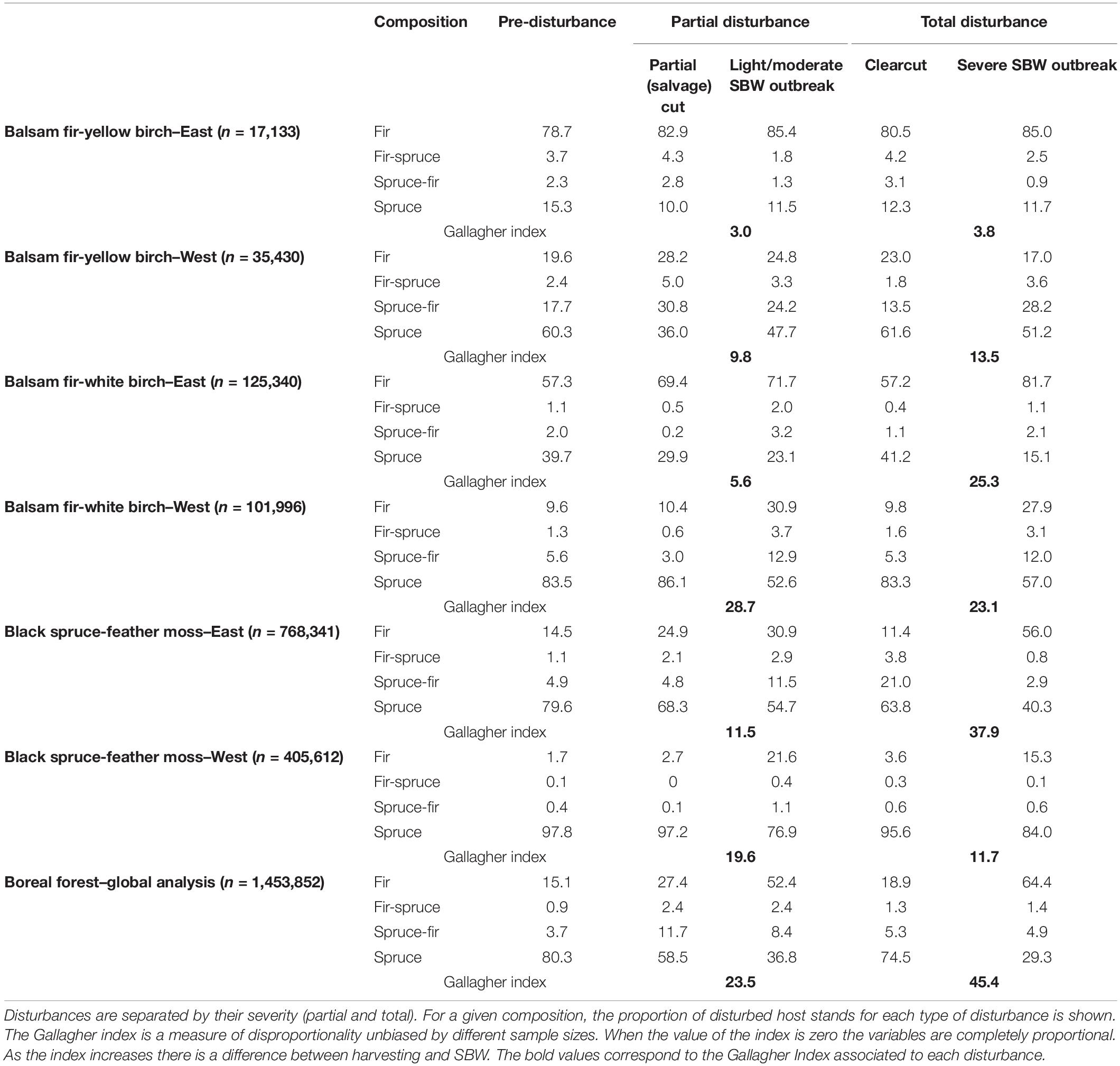
Table 1. Proportion of area per disturbance type for stands dominated by host species of the SBW.
However, it should be highlighted there are great differences in composition and harvesting and SBW across the different host dominated forest regions of Québec. The analysis by subdomain shows that the differences between severe SBW and total harvesting are most pronounced in the balsam fir-white birch domain (both east and western subdomains) and in the eastern black spruce-feather moss subdomain where the Gallagher index is 25.3, 23.1, and 37.9. Differences between harvesting and SBW are much less pronounced in the meridional balsam fir--yellow birch domain. The variation in the Gallagher index is also more pronounced at a finer spatial scale, the ecological region (approximate 1,000 km2 landscape with homogeneous composition, climate, topography, altitude etc.) (Supplementary Table 1). On a region-by-region basis, the Gallagher index ranges from a low of 0 in northern regions beyond where populations of the SBW are able to successfully complete its life cycle (Régnière et al., 2012) to 70 and higher in other regions. SBW systematically affects fir forests more than other types especially when this is compared to the pre-disturbance proportion of fir in a subdomain (Table 1). In contrast, spruce forests were less affected by SBW than their relative proportion in the pre-disturbance landscape.
Effect of Patch Size on Disturbance Susceptibility
The analyses revealed that SBW tended to disturb larger blocks of fir more than smaller blocks as more than 40% of balsam fir stands over 1,000 ha in size were defoliated while 30% of stands of fir blocks less than 100 ha were severely defoliated (Figure 3). In contrast, large blocks of spruce were significantly less (approximately half) vulnerable to the SBW as smaller ones. Harvesting on the other hand was concentrated in blocks larger than 100 ha for both spruce and fir. Although SBW caused almost four times the mortality in large blocks of fir compared to large blocks of spruce, harvesting during the last outbreak tended to target large blocks of spruce equivalently to large blocks of fir. Our data thus show that smaller blocks of fir are less affected by the SBW than large blocks of fir whereas large blocks of spruce are less vulnerable than smaller blocks of spruce.
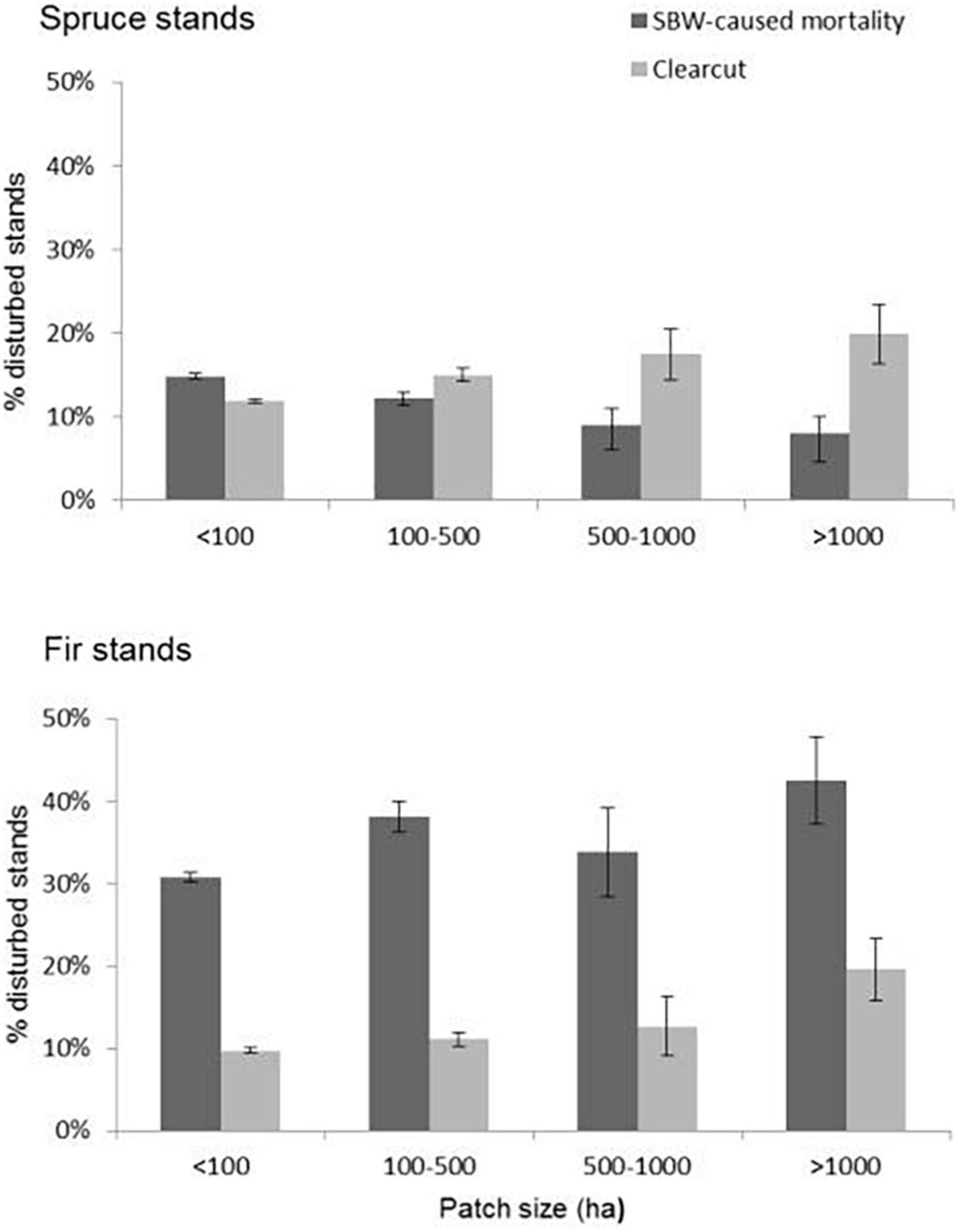
Figure 3. Preponderance of a disturbance type (harvesting vs. SBW) to occur in fir and spruce stands according to stand size. The y-axis represents the % of a given size-class of host stands disturbed by SBW or harvesting, e.g., 42% of all fir stands greater than 1,000 ha in size underwent mortality. Error bars represent 95% confidence intervals.
Differential Impacts of Logging and Spruce Budworm on Forest Composition
Analysis of pre- and post-disturbance species composition in permanent sample plots shows that both SBW and harvesting disturbances lead to an increase in hardwoods. This can be seen in Figure 2 where quartile values are greater than a ratio of one. Similarly, skewness values of 6.5 and 3.3, respectively, were calculated. The proportion of balsam fir on the other hand decreased after SBW but tended to remain similar, or decrease slightly, following harvesting (Figure 2). Light/moderate SBW disturbance reduced the proportion of balsam fir in 78% of the plots. Although spruce did not systematically change there was a strong difference in patterns occurring before and after SBW vs. before and after harvest. After harvest, there was less spruce on the majority of sites evaluated than before the disturbance whereas after the SBW outbreak most sites had a higher proportion of spruce than before (Figure 2).
Our results also highlight different impacts of both disturbances on the recruitment of shade-intolerant hardwood species [aspen (Populus tremuloides) and white birch]. Hardwoods tend to be less abundant after SBW outbreaks, whereas their post-harvest response tends to depend on their pre-harvest abundance. Sites with a high pre-harvest proportion of hardwoods tend to have an even greater proportion of hardwoods whereas the opposite is true at low pre-abundance levels of hardwoods (Supplementary Figure 1).
Discussion
Despite working with coarse landscape data that may have been affected by changes in harvest scheduling during the outbreak, we provide a rare quantification of the differences between logging and SBW outbreaks compared to pre-disturbance forest conditions. At a regional scale, our results are consistent with the SBW being more destructive in the central part of its range than in the southern part of its range where warmer fall temperatures may limit population growth (Régnière et al., 2012). Regions that are close to industrial centers where harvesting is logistically easier have more extreme Gallagher indices and thus greater differences between harvesting and SBW. A direct evaluation of the influence of harvest scheduling, proximity to mills, and ease of access in future studies will be useful to refine the results presented here Our work also shows that compared to SBW, logging led to divergent changes in forest composition (i.e., globally fir decreased in almost 80% of plots affected by SBW while the proportion of spruces decreased following harvesting on the majority of sites). The observed changes in forest composition between SBW and harvesting are thus consistent with the changes in tree composition and size patterns reported between post-fire and post-harvest forests (McRae et al., 2001; Dragotescu and Kneeshaw, 2012; Taylor et al., 2013).
From an ecosystem management point of view, forest management could focus on increasing the resistance of forests subject to the SBW by decreasing the proportion of the primary host across the landscape (Sánchez-Pinillos et al., 2019). Our work showing the vulnerability of large blocks of fir, and the resistance of large blocks of spruce to defoliation thus validates the stand level research that suggested that large blocks of fir should be broken-up to reduce outbreak severity (Blum and MacLean, 1984; Cotton-Gagnon et al., 2018; Kneeshaw et al., 2018). Similarly, our results support work arguing that large blocks of spruce would be an effective option to reduce forest vulnerability to the SBW (Sainte-Marie et al., 2015; Fuentealba et al., 2019). However, past forest management, instead of maintaining large blocks of spruce instead reduced their abundance (Figure 3).
It may be that smaller blocks of spruce are more vulnerable to the SBW because they are more subject to spill-over from adjacent stands of fir (sensu Bognounou et al., 2017) than larger contiguous blocks. This is supported by a regional study where black spruce forests were highly resistant to SBW outbreaks except when there was spill-over as outbreak duration increased (Sánchez-Pinillos et al., 2019). Planning at the landscape scale also suggests consideration of landscape connectivity (Robert et al., 2012, 2018, 2020) and SBW dispersal through the landscape (Boulanger et al., 2017), factors which are only beginning to be teased apart by genetic analysis of dispersing moths (Larroque et al., 2020). However, in the interim, we shows that as well as breaking up large blocks of fir to reduce vulnerability, foresters should ensure that large blocks of spruce are maintained or restored to minimize the impact of future outbreaks (Sainte-Marie et al., 2015).
Our work highlights the need to focus on reducing fir content and ensure the recruitment of companion species such as spruce during outbreaks (Bouchard et al., 2006; Kneeshaw et al., 2016; Sánchez-Pinillos et al., 2019). Using a multi-decade analysis in one region of the boreal forest, Sánchez-Pinillos et al. (2019) showed that although black spruce forests are the most resistant to SBW outbreaks, this resistance can break down if long-term severe defoliation exceeds a threshold. Fir forests were, however, shown to be ecologically resilient and despite large initial shifts in biomass and composition they eventually recover (Sánchez-Pinillos et al., 2019; Leduc et al., 2021). Our work highlights the importance of considering not only the stand scales but also the additive effects at the landscape scale and the importance of longterm objectives (Kneeshaw et al., 2021). We show that fir increases following harvesting whereas spruce tends to decrease after harvesting but increase following SBW outbreaks. This supports research showing that salvage logging can increase the risk of defoliation and mortality of understory advance black spruce regeneration, which would then further compromise long-term recruitment of black spruce (Cotton-Gagnon et al., 2018).
As noted in the literature (Table 2), our results also show that hardwood species are recruited equally following harvesting and outbreaks but that spruce, which is favored after outbreaks, was negatively impacted by harvesting during outbreaks (Figure 2). Since spruce is both the preferred species by the forest industry and is less vulnerable to the SBW than fir (Hennigar et al., 2008), it is paramount to ensure spruce recruitment during outbreak periods. Otherwise, the risk is that forest management by increasing the proportion of fir will lead to more severe outbreaks (Blais, 1983). We acknowledge that due to climate change spruce vulnerability to SBW may increase, as the insect will be able to complete its life cycle in northern regions, where spruce is dominant (Pureswaran et al., 2015). Although our analyses concentrate on the last outbreak recent evidence suggests, that for at least one region, the same pattern of harvesting leading to increasing fir recruitment is also occurring in the current outbreak (Sonntag, 2016; Cotton-Gagnon et al., 2018). Our work is thus in-line with research that shows that spruce should be maintained as it is not at risk of much defoliation or mortality due to the SBW (Sainte-Marie et al., 2015; Fuentealba et al., 2019).

Table 2. Differential impacts of spruce budworm outbreak (SBW) and harvesting on forest characteristics.
Conclusion
Spruce budworm outbreaks favor spruce recruitment (Bouchard et al., 2006) while spruce forests are both more valuable and less affected by SBW outbreaks (Sainte-Marie et al., 2015; Bognounou et al., 2017). Thus, from a traditional economic based forest perspective, efforts should be made to ensure that spruce recruitment is favored and that its abundance increased. This is generally a goal during endemic periods, but our work shows that is should be pursued during outbreaks.
Effective forest ecosystem management for SBW will need to consider both the emulation of ecological processes driven by outbreaks (Bouchard et al., 2008; Table 2) and management practices that don’t exacerbate SBW outbreaks (Kneeshaw et al., 2016, 2021). Our work helps quantify the differences in effects of harvesting and spruce outbreaks on forest composition and highlights that this occurs at the landscape as well as the stand scale. For ecosystem management the message is relatively clear, SBW outbreaks occur over long periods and multiple spatial scales and thus managers must have long-term strategies at both stand and landscape scales to not only reduce fir occurrence, but also to increase large blocks of spruce forest.
Data Availability Statement
All data are available from the Ministère des Forêts, Faune et Parcs du Québec (MFFP) (Ministry of Forests, Wildlife and Parks) at https://www.donneesquebec.ca/recherche/dataset/placettes-echantillons-permanentes-1970-a-aujourd-hui and https://www.donneesquebec.ca/recherche/dataset/systeme-d-information-forestiere-par-tesselle-sifort.
Author Contributions
JM-P, L-ER, MM, and LDG conducted the analysis. All authors contributed to the writing, and read and approved the final manuscript.
Funding
Financial support for this research was provided by the Spray Efficacy Research Group (SERG-International) through the SOPFIM, NSDNR. AlbertaAAF. The NSERC Discovery Program and FRQNT Team program also contributed to support this work. UQAM also contributed to support this work. Data was provided as in-kind support by the Quebec Ministry of Forests, Wildlife and Parks.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
We thank Guillaume Sainte-Marie, Mathieu Neau, Dominique Tardif and Ariane Breault for their work structuring the data and performing analyses. We also thank Patti Sonntag for discussions that enriched this work.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2022.680262/full#supplementary-material
Abbreviations
SBW, Spruce budworm; SIFORT, Spatial Information on Forest composition based on Tesserae; LSq, Gallagher Index; DBH, Diameter at breast height.
Footnotes
- ^ https://www.donneesquebec.ca/recherche/dataset/placettes-echantillons-permanentes-1970-a-aujourd-hui
- ^ https://www.donneesquebec.ca/recherche/dataset/systeme-d-information-forestiere-par-tesselle-sifort
References
Barrette, M., Thiffault, N., Tremblay, J. P., and Auger, I. (2019). Balsam fir stands of northeastern North America are resilient to spruce plantation. For. Ecol. Manag. 450:117504. doi: 10.1016/j.foreco.2019.117504
Baskerville, G. L. (1975). Spruce budworm: super silviculturist. For. Chronicle 51, 138–140. doi: 10.5558/tfc51138-4
Batzer, H. O., and Popp, M. P. (1985). Forest succession following a spruce budworm outbreak in Minnesota. For. Chronicle 61, 75–80. doi: 10.5558/tfc61075-2
Belle-Isle, J., and Kneeshaw, D. (2007). A stand and landscape comparison of the effects of a spruce bud-worm (Choristoneura Fumiferana (Clem.)) outbreak to the combined effects of harvesting and thinning on forest structure. For. Ecol. Manag. 246, 163–174. doi: 10.1016/j.foreco.2007.03.038
Bentz, B., Pierluigi, P., Delb, H., Fettig, C., Poland, T., Pureswaran, D., et al. (2020). “Advances in understanding and managing insect pests of forest trees,” in Achieving Sustainable Management Of Boreal And Temperate Forests, ed. J. A. Stanturf (Cam-bridge: Burleigh Dodds Science Publishing), 2019. doi: 10.3390/insects11070440
Bergeron, Y., Leduc, A., Joyal, C., and Morin, H. (1995). Balsam fir mortality following the last spruce budworm outbreak in northwestern Quebec. Can. J. For. Res. 25, 1375–1384.
Blais, J. R. (1958). The vulnerability of balsam fir to spruce budworm attack in northwestern ontario, with special reference to the physiological age of the tree. For. Chronicle 34, 405–422. doi: 10.5558/tfc34405-4
Blais, J. R. (1983). Trends in the frequency, extent, and severity of spruce budworm outbreaks in eastern Canada. Can. J. For. Res. 13, 539–547. doi: 10.1139/x83-079
Blum, B. M., and MacLean, D. A. (1984). Silviculture, forest management, and the spruce budworm.” man-aging the spruce budworm in eastern north america. edited by dm schmitt, dg grimble, and jl searcy. US Dep. Agric. Agric. Handb. 620, 83–102.
Bognounou, F., De Grandpré, L., Pureswaran, D. S., and Kneeshaw, D. (2017). Temporal variation in plant neighborhood effects on the defoliation of primary and secondary hosts by an insect pest. Ecosphere 8:e01759. doi: 10.1002/ecs2.1759
Bouchard, M., and Auger, I. (2014). Influence of environmental factors and spatio-temporal covariates during the initial development of a spruce budworm outbreak. Landsc. Ecol. 29, 111–126. doi: 10.1007/s10980-013-9966-x
Bouchard, M., Kneeshaw, D., and Bergeron, Y. (2006). Tree recruitment pulses and long-term species coexistence in mixed forests of western québec. Ecoscience 13, 82–88. doi: 10.2980/1195-6860(2006)13[82:trpals]2.0.co;2
Bouchard, M., Kneeshaw, D., and Bergeron, Y. (2008). Ecosystem management based on large-scale disturbance pulses: a case study from sub-boreal forests of western quebec (Canada). For. Ecol. Manag. 256, 1734–1742. doi: 10.1016/j.foreco.2008.05.044
Boulanger, Y., and Arseneault, D. (2004). Spruce budworm outbreaks in eastern quebec over the last 450 years. Can. J. For. Res. 34, 1035–1043. doi: 10.1139/x03-269
Boulanger, Y., Fabry, F., Kilambi, A., Pureswaran, D. S., Sturtevant, B. R., and Saint-Amant, R. (2017). The use of weather surveillance radar and high-resolution three dimensional weather data to monitor a spruce budworm mass exodus flight. Agric. For. Meteorol. 234, 127–135. doi: 10.1016/j.agrformet.2016.12.018
Boulet, B., Chabot, M., Dorais, L., Dupont, A., Gagnon, R., and Morneau, L. (1996). Entomologie Forest-Ière. Manuel De Foresterie, ed. Ordre des Ingénieurs Forestiers (Québec, QC: Les Presses de l’Université Laval), 1008–1043.
Cappuccino, N., Lavertu, D., Bergeron, Y., and Régnière, J. (1998). Spruce budworm impact, abundance and parasitism rate in a patchy landscape. Oecologia 114, 236–242. doi: 10.1007/s004420050441
Cotton-Gagnon, A., Simard, M., De Grandpré, L., and Kneeshaw, D. (2018). Salvage logging during spruce budworm outbreaks increases defoliation of black spruce regeneration. For. Ecol. Manag. 430, 421–430. doi: 10.1016/j.foreco.2018.08.011
Danneyrolles, V., Arseneault, D., and Bergeron, Y. (2016). Long-term compositional changes following partial disturbance revealed by the resurvey of logging concession limits in the northern tem-perate forest of eastern canada. Can. J. For. Res. 46, 943–949. doi: 10.1139/cjfr-2016-0047
De Grandpré, L., Kneeshaw, D. D., Périgon, S., Boucher, D., Marchand, M., Pureswaran, D., et al. (2019). Adverse climatic periods precede and amplify defoliators-induced tree mortality in eastern boreal North-America. J. Ecol. 107, 452–467. doi: 10.1111/1365-2745.13012
Dragotescu, I., and Kneeshaw, D. D. (2012). A comparison of residual forest following fires and harvesting in boreal forests in Quebec, Canada. Silva Fennica 46, 365–376.
Dupont, A., Bélanger, L., and Bousquet, J. (1991). Relationship between balsam fir vulnerability to spruce budworm and ecological site conditions of fir stands in central Quebec. Can. J. For. Res. 21, 1752–1759. doi: 10.1139/x91-242
Fuentealba, A., Dupont, A., Hébert, C., Berthiaume, R., Quezada-García, R., and Bauce, É (2019). Comparing the efficacy of various aerial spraying scenarios using bacillus thuringiensis to protect trees from spruce budworm defoliation. For. Ecol. Manag. 432:10131021. doi: 10.1016/j.foreco.2018.10.034
Gallagher, M. (1991). Proportionality, disproportionality and electoral systems. Electoral Stud. 10, 33–51. doi: 10.1016/0261-3794(91)90004-C
Gallagher, M., and Mitchell, P. (2005). The Politics of Electoral Systems. Oxford: Oxford University Press, 1–688. doi: 10.1093/0199257566.001.0001
Gauthier, S., Vaillancourt, M.-A., Leduc, A., De Grandpré, L., Kneeshaw, D., Morin, H., et al. (2009). Ecosystem Management in the Boreal Forest. Quebec City, QC: Presse de l’Université du Québec, 539.
Gendreau-Berthiaume, B., Kneeshaw, D., and Harvey, B. D. (2012). Effects of partial cutting and partial disturbance by wind and insects on stand composition, structure and growth in boreal mixed-woods. Forestry 85, 551–565. doi: 10.1093/forestry/cps051
Grondin, P., Saucier, J.-P., Blouin, J., Gosselin, J., and Robitaille, A. (2003). Information Écologique et Pla-nification Forestière Au Québec, Canada. Québec, QC: Ressources Naturelles, Faune et Parcs.
Hansen, W. D., Chapin, F. S., Naughton, H. T., Rupp, T. S., and Verbyla, D. (2016). Forest-landscape structure mediates effects of a spruce bark beetle (dendroctonus rufipennis) outbreak on subse-quent likelihood of burning in alaskan boreal forest. For. Ecol. Manag. 369, 38–46. doi: 10.1016/j.foreco.2016.03.036
Hennigar, C. R., and MacLean, D. A. (2010). Spruce budworm and management effects on forest and wood product carbon for an intensively managed forest. Can. J. For. Res. 40, 1736–1750. doi: 10.1139/x10-104
Hennigar, C. R., MacLean, D. A., Quiring, D. T., and Kershaw, J. A. (2008). Differences in spruce bud-worm defoliation among balsam fir and white, red, and black spruce. For. Sci. 54, 158–166. doi: 10.1093/forestscience/54.2.158
Jean, M., Lafleur, B., Fenton, N. J., Paré, D., and Bergeron, Y. (2019). Influence of fire and harvest severity on understory plant communities. For. Ecol. Manag. 436, 88–104. doi: 10.1016/j.foreco.2019.01.004
Kemball, K. J., Wang, G. G., and Dang, Q. L. (2005). Response of understory plant community of boreal mixedwood stands to fire, logging, and spruce budworm outbreak. Can. J. Bot. 83, 1550–1560. doi: 10.1139/b05-134
Kneeshaw, D. D., and Bergeron, Y. (1996). Ecological factors affecting the abondance of advance regen-eration in Quebec’s southwestern boreal forest. Can. J. For. Res. 26, 888–898. doi: 10.1139/x26-097
Kneeshaw, D., and Bergeron, Y. (1998). Canopy gap characteristics and tree replacement in the southeast-ern boreal forest. Ecology 79, 783–794. doi: 10.1890/0012-9658(1998)079[0783:cgcatr]2.0.co;2
Kneeshaw, D., and Bergeron, Y. (2015). “Applying knowledge of natural disturbance regimes to develop an ecosystem management approach in forestry,” in Ecological Forest Management Handbook, ed. G. R. Larocque (Boca Raton, FL: CRC Press), 3–31. doi: 10.1201/b19150-3
Kneeshaw, D., Burton, P. J., De Grandpré, L., Gauthier, S., and Boulanger, Y. (2018). “Is Management or Conservation of Old Growth Possible in North American Boreal Forests?” Eastern Old-Growth. Washington, DC: Is-land Press.
Kneeshaw, D., Harvey, B. D., Reyes, G. P., Caron, M. N., and Barlow, S. (2011). Spruce budworm, wind-throw and partial cutting: do different partial disturbances produce different forest structures? For. Ecol. Manag. 262, 482–490. doi: 10.1016/j.foreco.2011.04.014
Kneeshaw, D., Sturtevant, B. R., Cooke, B., Work, T., Pureswaran, D., De Grandpre, L., et al. (2016). “Insect disturbances in forest ecosystems,” in Routledge Handbook of Forest Ecology, eds K. Peh, R. Corlett, and Y. Bergeron (Abingdon: Routledge), 93–113. doi: 10.4324/9781315818290
Kneeshaw, D., Sturtevant, B., De Grandpé, L., Doblas-Miranda, E., James, P. M. A., Tardif, D., et al. (2021). The vision of managing for pest-resistant landscapes: realistic or utopic? Curr. For. Rep. 7, 97–113. doi: 10.1007/s40725-021-00140-z
Larroque, J., Johns, R., Canape, J., Morin, B., and James, P. M. (2020). Spatial genetic structure at the lead-ing edge of a spruce budworm outbreak: the role of dispersal in outbreak spread. For. Ecol. Manag. 461:117965. doi: 10.1016/j.foreco.2020.117965
Leduc, A., Leduc, A., Kneeshaw, D., and Bergeron, Y. (2021). Advancing and reversing succession as a function of time since fire and insect outbreaks: an 18 year in situ remeasurement of changes in forest com-position. J. Vegetation Sci. 32:e12974. doi: 10.1111/jvs.12974
Liu, Z., Peng, C., De Grandpré, L., Candau, J. N., Work, T., Huang, C., et al. (2019). Simula-tion and analysis of the effect of a spruce budworm outbreak on carbon dynamics in boreal forests of Québec. Ecosystems 22, 1838–1851. doi: 10.1007/s10021-019-00377-7
Liu, Z., Peng, C., De Grandpré, L., Candau, J. N., Zhou, X., and Kneeshaw, D. (2018). Development of a new TRIPLEX-Insect model for simulating the effect of spruce budworm on forest carbon dynamics. Forests 9:513. doi: 10.3390/f9090513
MacLean, D. A. (1980). Vulnerability of fir-spruce stands during uncontrolled spruce budworm out-breaks: a review and discussion. For. Chronicle 56, 213–221. doi: 10.5558/tfc56213-5
MacLean, D. A. (2004). “Chap. 6: Predicting natural forest insect disturbance regimes,” in Emulating Natural Forest Landscape Disturbances: Concepts And Applications, eds A. H. Perera, L. J. Buse, and M. G. Weber (New York, NY: Columbia University Press), 69–82. doi: 10.7312/pere12916-009
MacLean, D. A., Erdle, T. A., MacKinnon, W. E., Porter, K. B., Beaton, K. P., Cormier, G., et al. (2001). The spruce budworm decision support system: forest protection planning to sustain long- term wood supply. Can. J. For. Res. 31, 1742–1757. doi: 10.1139/x01-102
McRae, D. J., Duchesne, L. C., Freedman, B., Lynham, T. J., and Woodley, S. (2001). Comparisons between wildfire and forest harvesting and their implications in forest management. Environ. Rev. 9, 223–260. doi: 10.2134/jeq2018.04.0169
Ministère des Forêts de la Faune et des Parcs (2014). L’aménagement Écosystémique Dans Un Contexte D’épidémie De La Tordeuse Des Bourgeons De L’épinette – Guide De Référence Pour Moduler Les Activités D’aménagement Dans Les Forêts Publiques. Québec, QC: Gouvernement Du Québec, 127.
Ministère des Forêts de la Faune et des Parcs (2016). Norme D’inventaire Écoforestier Placettes-Échantillons Permanentes, Québec. Québec City, QC: Gouvernement du Québec, 254.
Ministère des Forêts de la Faune et des Parcs (2019). Aires Infestées Par La Tordeuse Des Bourgeons de l’épinette Au Québec En 2019. Québec, QC: Gouvernement Du Québec, 32.
Moussaoui, L., Fenton, N. J., Leduc, A., and Bergeron, Y. (2016). Can retention harvest maintain natural structural complexity? A comparison of post-harvest and post-fire residual patches in boreal for-est. Forests 7:243. doi: 10.3390/f7100243
Pelletier, G., Dumont, Y., and Bédard, M. (2007). SIFORT: Système d’Information Forestière Par Tesselle. Manuel de l’usager. Québec, QC: Ressources Naturelles et Faune.
Pothier, D., Elie, J. G., Auger, I., Mailly, D., and Gaudreault, M. (2012). Spruce budworm-caused mortality to balsam fir and black spruce in pure and mixed conifer stands. For. Sci. 58, 24–33. doi: 10.5849/forsci.10-110
Pureswaran, D. S., De Grandpré, L., Paré, D., Taylor, A., Barrette, M., Morin, H., et al. (2015). Climate-induced changes in host tree–insect phenology may drive ecological state-shift in boreal forests. Ecology 96, 1480–1491. doi: 10.1890/13-2366.1
Pureswaran, D. S., Neau, M., Marchand, M., De Grandpré, L., and Kneeshaw, D. (2019). Phenological syn-chrony between eastern spruce budworm and its host trees increases with warmer temperatures in the boreal forest. Ecol. Evol. 9, 576–586. doi: 10.1002/ece3.4779
Régnière, J., St-Amant, R., and Duval, P. (2012). Predicting insect distributions under climate change from physiological responses: spruce budworm as an example. Biol. Invasions 14, 1571–1586. doi: 10.1007/s10530-010-9918-1
Robert, L. E., Kneeshaw, D., and Sturtevant, B. R. (2012). Effects of forest management legacies on spruce budworm (choristoneura fumiferana) outbreaks. Can. J. For. Res. 42, 463–475. doi: 10.1139/X2012-005
Robert, L. E., Sturtevant, B. R., Cooke, B. J., James, P. M. A., Fortin, M. J., Townsend, P. A., et al. (2018). Landscape host abundance and configuration regulate periodic outbreak behavior in spruce budworm choristoneura fumiferana. Ecography 41, 1556–1571. doi: 10.1111/ecog.03553
Robert, L. E., Sturtevant, B. R., Kneeshaw, D., James, P. M., Fortin, M. J., Wolter, P. T., et al. (2020). Forest landscape structure influences the cyclic-eruptive spatial dynamics of forest tent caterpillar outbreaks. Ecosphere 11:e03096. doi: 10.1002/ecs2.3096
Sainte-Marie, G. B., Kneeshaw, D., MacLean, D. A., and Hennigar, C. R. (2015). Estimating forest Vul-nerability to the next spruce budworm outbreak: will past silvicultural efforts pay dividends? Can. J. For. Res. 45, 314–324. doi: 10.1139/cjfr-2014-0344
Sánchez-Pinillos, M., Leduc, A., Ameztegui, A., Kneeshaw, D., Lloret, F., and Coll, L. (2019). Resistance, resilience or change: post-disturbance dynamics of boreal forests after insect outbreaks. Ecosystems 22, 1886–1901. doi: 10.1007/s10021-019-00378-6
Saucier, J. P., Gosselin, J., Morneau, C., and Grondin, P. (2010). Utilisation de la classification de la végétation dans l’aménagement forestier au Québec. Rev. For. Française 62, 428–438.
Saucier, J.-P., Bergeron, J.-F., Grondin, P., and Robitaille, A. (1998). Les régions écologiques du québec méridional: un des éléments du systeme hiérarchique de classification écologique du territoire mis au point par le ministere des ressources naturelles. L’Aubelle 1–12.
Schmalholz, M., Hylander, K., and Frego, K. (2011). Bryophyte species richness and composition in young forests regenerated after clear-cut logging versus after wildfire and spruce budworm outbreak. Biodivers. Conserv. 20, 2575–2596. doi: 10.1007/s10531-011-0092-2
Sonntag, P. (2016). Attack of the Budworms: The Current Infestation Threatens Canadian Forests.” 2016. Available online at: https://thewalrus.ca/attack-of-the-budworms/ (accessed December 6, 2021).
Sterner, T. E., and Davidson, A. G. (1982). Forest Insect And Disease Conditions In Canada. Ottawa, ON: Canada Forest Service, Forest Insect and Disease Survey, 46.
Su, Q., Needham, T. D., and MacLean, D. A. (1996). The influence of hardwood content on balsam fir de-foliation by spruce budworm. Can. J. For. Res. 26, 1620–1628. doi: 10.1139/x26-182
Taylor, A. R., Hart, T., and Chen, H. Y. (2013). Tree community structural development in young boreal for-ests: a comparison of fire and harvesting disturbance. For. Ecol. Manag. 310, 19–26. doi: 10.1016/j.foreco.2013.08.017
Virgin, G. V., and MacLean, D. A. (2017). Five decades of balsam fir stand development after spruce bud-worm-related mortality. For. Ecol. Manag. 400, 129–138. doi: 10.1016/j.foreco.2017.05.057
Volney, W. J. A., and Fleming, R. A. (2007). Spruce budworm (Choristoneura spp.) biotype reactions to for-est and climate characteristics. Glob. Change Biol. 13, 1630–1643. doi: 10.1111/j.1365-2486.2007.01402.x
Keywords: spruce budworm, forest ecosystem management, long-term management, outbreaks vs. endemics, composition
Citation: Kneeshaw DD, De Grandpré L, D’Orangeville L, Marchand M, Moisan-Perrier J, Robert L-E and Bouchard M (2022) Forest Structure and Composition Diverge Following Harvesting Compared to a Spruce Budworm Choristoneura fumiferana (Clem.) Outbreak. Front. For. Glob. Change 5:680262. doi: 10.3389/ffgc.2022.680262
Received: 13 March 2021; Accepted: 02 March 2022;
Published: 11 April 2022.
Edited by:
Barry Alan Gardiner, Institut Européen de la Forêt Cultivée (IEFC), FranceReviewed by:
Aaron Weiskittel, University of Maine, United StatesOlivier Bouriaud, Ştefan cel Mare University of Suceava, Romania
Copyright © 2022 Kneeshaw, De Grandpré, D’Orangeville, Marchand, Moisan-Perrier, Robert and Bouchard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Daniel D. Kneeshaw, a25lZXNoYXcuZGFuaWVsQHVxYW0uY2E=