 Paola Musella
Paola Musella Ismahen Lalaymia
Ismahen Lalaymia Rik Verdonck3
Rik Verdonck3 Jan Colpaert
Jan Colpaert Quentin Ponette
Quentin Ponette Stéphane Declerck
Stéphane Declerck- 1Université Catholique de Louvain, Earth and Life Institute, Mycology, Louvain-la-Neuve, Belgium
- 2Université Catholique de Louvain, Earth and Life Institute, Forest Sciences, Louvain-la-Neuve, Belgium
- 3University of Hasselt, Centre for Environmental Sciences (CMK), Environmental Biology Group, Diepenbeek, Belgium
Introduction: Scots pine (Pinus sylvestris) is an important tree species that is often regenerated through seedling establishment. The resistance of these seedlings to drought treatments and their recovery are essential to forest regeneration. The association with ectomycorrhizal (EcM) fungi such as Paxillus involutus has the potential to improve drought resistance and possibly recovery. Here, we conducted two experiments to evaluate the impact of P. involutus inoculation on growth and water use of pine seedlings under contrasting drought intensities.
Methods: In the first experiment, EcM fungal-inoculated and non-inoculated seedlings were subjected to well-watered (100% field capacity—control treatment) or low-watered (10% field capacity for 1 week—drought treatment) conditions, followed by a recovery phase in a semi-hydroponic cultivation system. In the second experiment, seedlings were initially subjected to well-watered or moderate water conditions (100 and 30% field capacity, respectively), followed by prolonged low-watered conditions (10% field capacity for 1 month), each followed by a recovery phase.
Results: In the first experiment, EcM fungal-colonized pine seedlings showed increased aboveground biomass, higher photosynthetic rates, water-use efficiency, and elevated nitrogen and phosphorus contents under low-water conditions. In the second experiment, a prolonged low-water regime reduced root colonization by the EcM fungus. During recovery, P. involutus-inoculated seedlings had an increase in ammonium assimilation under well-watered conditions and appeared to recover more efficiently, in particular from prolonged low-water conditions, with higher chlorophyll production, possibly due to increased nutrient availability and/or fungal demand.
Discussion: Our results suggest that the benefits of P. involutus depend on both water regime and drought duration: a short-term low-water regime (10% field capacity for 1 week) enhances EcM benefits, while under a moderate water regime (30% field capacity) the fungal effect was not evident. We conclude that P. involutus can confer significant benefits to pine seedlings, making it a valuable strategy for forest regeneration in temperate climates facing drought episodes.
1 Introduction
Global surface temperatures have risen faster since 1970 than in any other 50-year period, at least over the last 2000 years, and several regions in Europe are projected to experience an increase in the frequency and/or severity of droughts over the coming decades (IPCC, 2023). These extreme droughts combined with rising temperatures are weakening trees, threatening forest ecosystems and the services they provide. Extensive tree dieback and mortality related to drought have already been detected across the globe. For instance, over the period 2000–2022, the average annual area of European forests affected by drought was 56.000 km2 (ca. 4% of forests) (Medium Resolution Vegetation Phenology and Productivity, Oct. 2022, European Environment Agency).
Drought severity is typically viewed as the primary factor leading to a decline in forest growth, although the functional traits of different tree species can also provide varying degrees of drought resistance (Greenwood et al., 2017; Peltier et al., 2016; Yin and Bauerle, 2017). Severity, duration, and frequency of drought can cause a gradual rise in tree mortality rates and even sudden die-off events. Numerous recent events of tree mortality related to drought and heat from various regions globally indicate that no forest type or climate zone is immune to the consequences of climate change, even in areas typically not considered water-limited (Allen et al., 2010). Repeated drought events cause cumulative damage to tree vascular systems, leading to increased mortality (Anderegg et al., 2015). On the other hand, severe drought may kill plants, but a mild drought or a drought for a short duration can trigger short-term memory, usually established by transcriptional training or metabolic reprogramming, leading to survival under a subsequent drought (Sadhukhan et al., 2022).
Successful seedling establishment is a prerequisite for the long-term persistence of a tree species (Lloret et al., 2009). However, seedlings are generally more sensitive to drought and less likely to recover than mature trees due to their lower carbohydrate reserves, smaller rooting volume, and shallower rooting depths than older trees (Seidel and Menzel, 2016).
P. sylvestris (Scots pine) is an ideal candidate for investigating the impacts of drought stress on seedling establishment because it is a pioneer species (Krakau et al., 2013; Vacek et al., 2021), with ecological importance and wide distribution (Pyhäjärvi et al., 2007). Research on Scots pine can provide critical insights into the mechanisms of drought tolerance, inform conservation strategies, and improve reforestation success in the face of increasing drought frequency and intensity.
Scots pine, as most temperate region forest trees, is associated with ectomycorrhizal (EcM) fungi. These root fungal symbionts may facilitate the access of seedlings to water and nutrients (Smith and Read, 1997; Lehto and Zwiazek, 2011). They are able to penetrate rock and soil pores as small as 2 μm diam thanks to the microscopic size of their hyphae (Allen, 2007), therefore accessing water and nutrients otherwise not available to roots (Lehto and Zwiazek, 2011).
Various studies have highlighted the beneficial effects of EcM fungi on tree seedlings. For example, Wang et al. (2021) showed that Suillus variegatus reduced the mortality rate and increased the trunk height, root and leaf biomass, photosynthetic rate, stomatal conductance and transpiration of P. tabulaeformis subjected to moderate to severe drought stress. Kipfer et al. (2012) also observed that S. granulatus can increase aboveground biomass of P. sylvestris seedling under drought stress conditions. Muhsin and Zwiazek (2002) found that Hebeloma crustuliniforme can increase root hydraulic conductivity in white spruce (Picea glauca) seedlings under salt treatments. Garbaye and Churin (1997) showed that P. involutus significantly improved the phosphorus status of young oaks (Quercus petraea and Q. robur) during summer drought, while Danielsen and Polle (2014) showed that this EcM fungus increased potassium, magnesium and phosphorus concentrations in Populus canescens leaves under drought conditions.
Previous studies have shown that drought can negatively affect EcM symbioses by reducing fungal colonization and activity, particularly under prolonged or severe water stress (Davies et al., 1996; Morte et al., 2001). However, the extent of this decline is highly variable and depends on plant genotype and environmental conditions, highlighting the context-dependent nature of EcM responses to drought (Gehring et al., 2006).
While a number of studies have been conducted on the effect of EcM fungi on seedling nutrition (Colpaert et al., 1996, 1999; Zhang and George, 2010; Pohjanen et al., 2014; Nehls and Plassard, 2018), and drought stress alleviation (Bogeat-Triboulot et al., 2004; Kipfer et al., 2012; De Quesada et al., 2024; Himanen et al., 2024) fewer studies have focused specifically on the post-drought recovery phase, particularly in seedlings (Cregg and Zhang, 2001). However, recent work (Carter and Dickman, 2022; Castaño et al., 2023; Liu et al., 2024) has begun to address this gap. Understanding the effect of EcM fungi on tree seedling recovery after drought is crucial for promoting forest resilience in the face of increasing climate change. This knowledge can enhance reforestation efforts by improving seedling survival and growth in drought-prone environments.
In the present study, we investigated the effects of the EcM fungus P. involutus on P. sylvestris (Scots pine) seedlings growth under normal, moderate or low water regime and on tree recovery after drought treatments. P. involutus was used because it has a broad-host-range (Molina and Trappe, 1982), is very common in temperate ecosystems (Hahn, 1996) and is one of the most widely used EcM fungus in experimental laboratory work, thanks to its ease of cultivation (Wallander and Söderström, 1999). In particular, in this study, we hypothesized that P. involutus (1) enhances the resistance of pine seedlings to severe, moderate, and prolonged severe drought treatments (2) and improves the recovery of pine seedlings after exposure to severe, moderate, and prolonged severe drought treatments.
2 Materials and methods
2.1 Biological material
Scots pine (P. sylvestris L.) seeds collected in Hanau (France) were supplied by the Comptoir Wallon des Matériels forestiers de Reproduction. P. involutus (Batsch.) Fr., was supplied by the Centre for Environmental Sciences (CMK) at Hasselt University. It was obtained from basidiocarps collected in 2021 from a stand of P. sylvestris in Lommel (Belgium) (sampling location: 51°14′27.9564”N, 5°15′27.1188″E) and grown on Fries medium on cellophane sheets.
2.2 Experimental procedure
2.2.1 Inoculation of seedlings with P. involutus
Scots pine seeds were washed with Tween 20 and surface disinfected with H2O2 (30%). They were then sown in a substrate made of a mixture of perlite and vermiculite (vol 2:1), previously sieved (200 μm pore size), washed with demineralized water to remove impurities and oven-dried for 48 h at 70°C. The substrate was moistened with Ingestad nutrient solution (Ingestad and Kähr, 1985) modified according to the optimum macronutrient weight proportions for P. sylvestris (100 N/15 P/60 K/6 Ca/6 Mg/9 S) (Nylund and Wallander, 1989). The plants were maintained in a growth chamber at 22/18°C (day/night), 65% relative humidity (RH) with a photoperiod of 12 h day−1, under a photosynthetic photon flux density (PPFD) of 200 μmol m−2 s−1. Seventy-two days after sowing, the plants reached a height of 6.0 ± 0.3 cm. The sandwich technique detailed in Colpaert et al. (1996) was used to inoculate the plants with the EcM fungus to produce mycorrhizal (M) plants. Briefly, it involved gently placing the plant roots in intimate contact with the mycelium of the EcM fungus grown on a sheet of cellophane. This arrangement was carefully nested between two layers of moist filter paper in sealed Petri dishes. Non-mycorrhizal (NM) plants followed the same procedure without adding the fungal inoculum.
2.2.2 Plant growth system
After 4 days of contact with the mycelium (for the M treatment) or without it (for NM treatment), 130 plants were transferred to inverted plastic bottles (500 mL - VWR, United States—named containers afterward) cut at the base and covered with aluminum foil to prevent the development of algae. A microporous membrane (41 μm pore size) was glued to the inside of each container at the neck to ensure the free passage of water, while preventing the passage of substrate and roots. Each container was filled with 42 g of perlite (DCM, Belgium), sieved and washed as described above. The containers were then transferred in the growth chamber (see conditions described above) on tables. On each table, five M plants, five NM plants, and two non-vegetated (NV) containers were placed and moved randomly every week. All containers were watered twice a week with Ingestad nutrient solution at field capacity.
2.2.3 Drought phases and experimental design
Eleven months after transfer to containers, 10 P. sylvestris seedlings—five mycorrhizal (M) and five non-mycorrhizal (NM)—were randomly selected and harvested to assess initial biomass and root colonization. The M plants showed 81.3 ± 18.3% root colonization, while no colonization was observed in NM plants. There was no significant difference in dry biomass between treatments, with M and NM seedlings averaging 1.8 ± 0.6 g and 1.9 ± 0.5 g, respectively (p = 0.7133). The remaining 120 plants (60 M and 60 NM) were randomly assigned to two separate drought experiments.
Experiment 1 (Figure 1A) involved 40 plants—20 M and 20 NM—divided into two water regimes. The water regimes were established by refraining from watering the containers until the soil moisture reached the desired relative water content (RWC) based on field capacity: a severe drought treatment (LWR; 10% RWC, −1,500 kPa) and a control treatment (NWR; 90–100% RWC, 0 kPa) (Lim et al., 2021). Once the LWR treatment reached the target RWC and was maintained for 1 week, 20 plants were harvested. The remaining 20 plants were then transferred to a semi-hydroponic recovery system for 6 days before final harvest. Once the LWR treatment reached the target RWC and was maintained for 1 week, 20 plants were harvested. The remaining 20 plants were transferred to a semi-hydroponic recovery system for 6 days before being harvested.
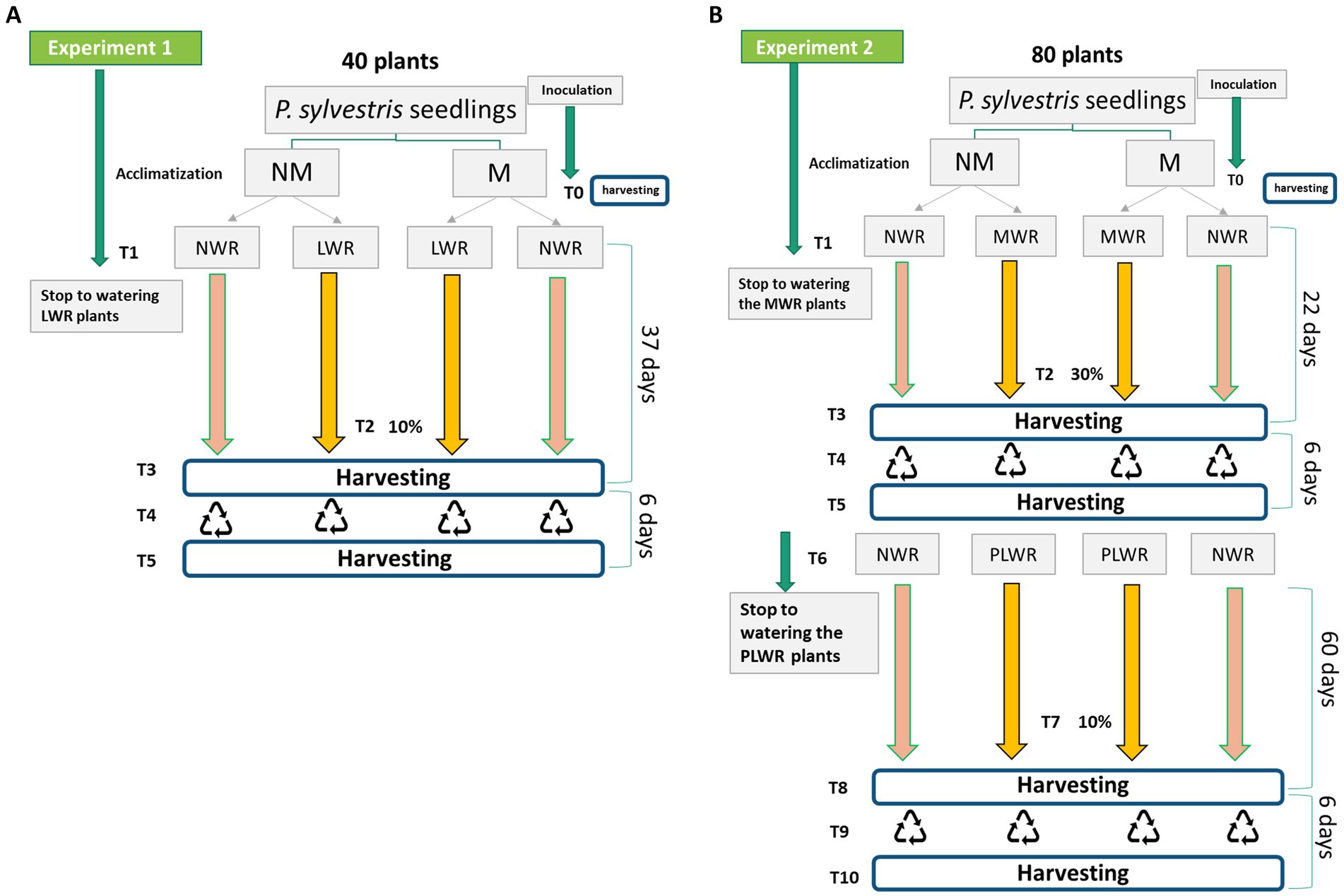
Figure 1. (A) Schematic representation of the experimental design for Experiment 1 and (B) for Experiment 2. NM = non-mycorrhizal plants; M = mycorrhizal plants; NWR = normal water regime (i.e., the control); LWR = low water regime (watering reduced and maintained to 10–15% RWC for 1 week); MWR = moderate water regime (watering reduced to 30–35% RWC); PLWR: prolonged low water regime (watering reduced during 30 days and maintained to 10–15% RWC for one more month). Experiment 1: T0 = Fungal inoculation; T1 = interruption of watering the LWR treatments; T2 = Containers of the LWR treatments reaching 10–15% RWC; T3 = harvesting of 20 plants (5 per treatment); T4 = Start of water circulation for 20 containers for 1 week; T5 = harvesting of 20 plants (5 per treatment). Experiment 2: T0 = Fungal inoculation; T1 = Interruption of watering the MWR treatments; T2 = Containers reaching 30–35% RWC; T3 = harvesting of 20 plants (5 per treatment); T4 = Start of water circulation for 60 containers for 1 week; T5 = harvesting of 20 plants (5 per treatment); T6 = Interruption of watering the PLWR treatments; T7 = Containers reaching 10% RWC, kept for 1 month; T8 = harvesting of 20 plants (5 per treatment); T9 = Start of water circulation for 20 containers (5 per treatment) for 1 week; T10 = harvesting of 20 plants.
Experiment 2 (Figure 1B) included 80 plants—40 M and 40 NM—initially exposed to either a moderate drought treatment (MWR; 30% RWC, ~ −500 kPa) or NWR (Lim et al., 2021). After 22 days, 20 plants (five per treatment) were harvested, while the remaining 60 were transferred to the semi-hydroponic system for a six-day recovery. After recovery 20 plants were harvested. The remaining 40 plants (20 M and 20 NM) were then divided again into two groups: one continued under NWR, while the other was subjected to a prolonged severe drought treatment (PLWR; 10% RWC for 30 days, −1,500 kPa) (Lim et al., 2021). At the end of this prolonged drought phase, 20 plants were harvested and the remaining 20 underwent a six-day recovery period in the hydroponic system before final harvesting.
At all phases of both experiments, five biological replicates were maintained per treatment group and time point.
Water regimes were established by withholding irrigation until containers reached target RWC, monitored gravimetrically by weighing six randomly selected containers per treatment every 3 days. Field capacity, defined as the maximum water retention of 40 g of perlite, was 120 mL. Non-vegetated containers were included as evaporation controls. Plants under NWR were fertilized twice weekly with 50 mL of Ingestad nutrient solution. Moderate and severe drought treatments were simulated at 30 and 10% RWC, respectively—thresholds validated by previous studies. Meng et al. (2023) used 40 and 20% field capacity to model drought in P. sylvestris var. mongolica, and Pearson et al. (2013) reported significant drops in photochemical efficiency at 30 and 10% RWC in P. sylvestris seedlings. In mature trees, transpiration rates fell sharply near 12% RWC, and 10% aligns with the species’ estimated permanent wilting point in sandy soils (Kirkham, 2023).
2.2.4 Recovery phase
After each period of MWR or LWR, the plants were subjected to a recovery phase, using the circulatory semi-hydroponic cultivation system developed by Garcés-Ruiz et al. (2017) (Figure 2). In brief, for each treatment, 1 L of Ingestad nutrient solution in glass bottles was circulated via peristaltic pumps through the containers and returned to the bottles in a closed circuit. After a flushing step to saturate the perlite, the inorganic P (i.e., Pi) and NH4+ depletion (%) in the circulating solution was measured over a period of 6 days (see for details Section 2.3.4). Initial nutrient concentration in the Ingestad solution was determined by sampling 25 mL from each bottle and considered as Time 0 (T0). The nutrient solution was then collected as follows: the day one after 2, 4, 6, 8, and 10 h from T0; the day two after 21, 24, 27, and 30 h from T0; the day three after 45, 50, and 55 h from T0; the day four after 70 and 80 h from T0; the day five, 100 h from T0, the day six, 120 h from T0. Finally, the plants were harvested and the solution samples were stored at 4°C in the dark.
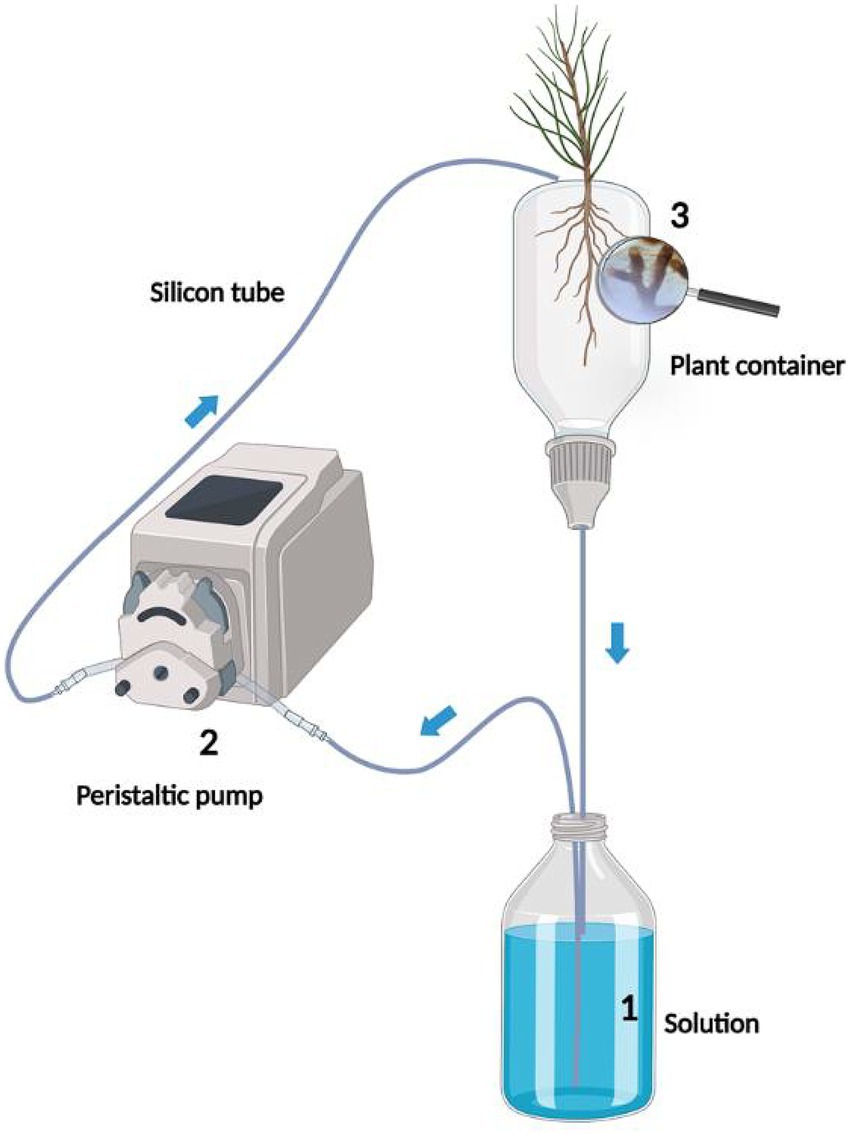
Figure 2. Schematic representation of the circulatory semi-hydroponic cultivation system, used to inorganic P (Pi) and NH4+ depletion in the Ingested solution circulating through the containers. The nutrient solution in the glass bottle (1) is pumped with a peristaltic pump (2) to the upper part of the container (3) via silicon tubes. The solution percolates through the plant container back into the glass bottle. Gray arrows indicate the flow direction of the nutrient solution in the tubing (Garcés-Ruiz et al., 2017; Cartabia et al., 2021).
2.3 Harvest and analysis
At each phase of Experiment 1 and 2 (Figures 1A,B), 5 plants per treatment were harvested. Just before harvesting, physiological indicators such as leaf gas exchange and chlorophyll content were measured. Stems were then cut to determine shoot water potential and plants harvested entirely to assess morphological characteristics.
2.3.1 EcM fungal colonization of root
Root colonization rate (RCR) was estimated on three randomly selected root sub-samples per plant. The RCR was obtained by counting the number of tips colonized by the EcM fungus divided by the total number of tips observed (Brundrett et al., 1996).
2.3.2 Physiological parameters
Just before harvesting, photosynthetic rate, stomatal conductance, and transpiration rates were measured using an integrated fluorometer and gas exchange system (ADC Bioscientific Ltd., Hoddesdon, United Kingdom), with the application of LCpro-SD, a portable conifer-specific photosynthesis measurement chamber. Gas exchange measurements were performed between 9:00 and 18:00 under controlled chamber conditions with constant light intensity of 999 μmol m−2 s−1, chosen based on a preliminary light response curve. CO₂ reference concentrations ranged from 427 to 452 ppm, and humidity inside the needle chamber (H₂O of reference) ranged from 9.9 to 10.8 mmol mol−1. Instantaneous water-use efficiency (WUEt) was calculated using the equation of Bacelar et al. (2012): WUEt = Photosynthetic rate/Transpiration rate.
The chlorophyll content of the needles was measured on four needles per plant before and after recovery phase on the same plants (5 per treatment), using a Chlorophyll Content Meter adapted for very small leaves (CCM-300, Opti-Sciences, Inc., USA), which uses a fluorescence ratio technique for chlorophyll content measurement according to Gitelson et al. (1999). The difference between the values before and after the recovery phase was then calculated.
Shoot water potential (Ψ) was measured at midday (between 12:00 and 14:00) under full light conditions, in order to assess plant water status during peak transpiration. Shoots were excised at the base of the stem immediately before the measurement. Water potential was then determined using an airtight pressure chamber (Model 3,115, Soil Moisture Equipment Corp., USA), in which xylem sap is forced out of the plant tracheids by increasing pressure in the chamber, following the method of Scholander et al. (1965).
2.3.3 Plant growth parameters
The height of the main stem was measured, followed by fresh weight of shoots and roots. The volume of the roots was calculated in a graduated cylinder, measuring the difference between the water volume before and after adding the roots to the cylinder.
Roots and needles of each plant were then stored at −80°C and freeze-dried to obtain dry weights. The water content of the roots and shoots was expressed as g of H2O on g of biomass. After freeze-drying, the roots and shoots samples were ground separately to powder using a mortar and liquid nitrogen, and the concentrations (%) of phosphorus (P), carbon (C), and nitrogen (N) were determined using an inductively coupled plasma atomic emission spectroscopy (ICP-AES), after acid digestion of tissues with HNO3 (ISO 11466:1995) for P and an Elemental Analyzer System (Nelson and Sommers, 1996) for C and N. Phosphorus, C and N content of shoots and roots were calculated by multiplying the nutrient concentration by the dry biomass of shoots and roots.
2.3.4 Measurement of nutrient uptake in the semi-hydroponic cultivation system
The Pi concentration was measured by ICP-AES as described in Garcés-Ruiz et al. (2017). Data were then normalized to the NV controls (Garcés-Ruiz et al., 2017) and converted to % concentrations of Pi in the nutrient solution relative to the initial concentration at T0, using the following formula:
where:
[P] = Pi concentration in the solution.
x = sample.
NV = non-vegetated containers (i.e., perlite control).
[PNV] = mean Pi concentration for the NV containers.
T = time considered
T0 = start of the circulatory system.
For NH4+, the concentration was determined by colorimetric analysis, using the Merck Spectroquant Ammonium-Test (1.14752, Merck - Darmstadt, Germany) which uses the transformation reaction of ammonium into blue indophenol. The same formula as above was applied to ammonium concentration.
2.4 Statistical analysis
Data analysis was performed with JMP Pro 17 with an α-threshold of 5%. Levene and Shapiro–Wilk tests were performed before statistical analysis to confirm the homogeneity of variance and the normality of distribution, respectively. The effects of the factors mycorrhization (Myc) and water regime (WR) were investigated using a general linear model (GLM) followed by a pairwise comparison using the Tukey-HSD test.
3 Results
3.1 Experiment 1
The following sections describe the effects of water regime WR (low water regime, LWR, and normal water regime, NWR) and mycorrhization (Myc) on root colonization, plant growth, physiological parameters, plant nutrient status, and recovery phase of P. sylvestris seedlings.
3.1.1 Root colonization
Whatever the WR, root colonization was observed in each plant, with typical dichotomous tips surrounded by extraradical hyphae (Figure 3). Root colonization rates were unaffected with values of 99.7 ± 0.3% and 92.5 ± 5.9%, for the NWR and LWR treatments, respectively. No root colonization was observed in the plants of the NM treatments.
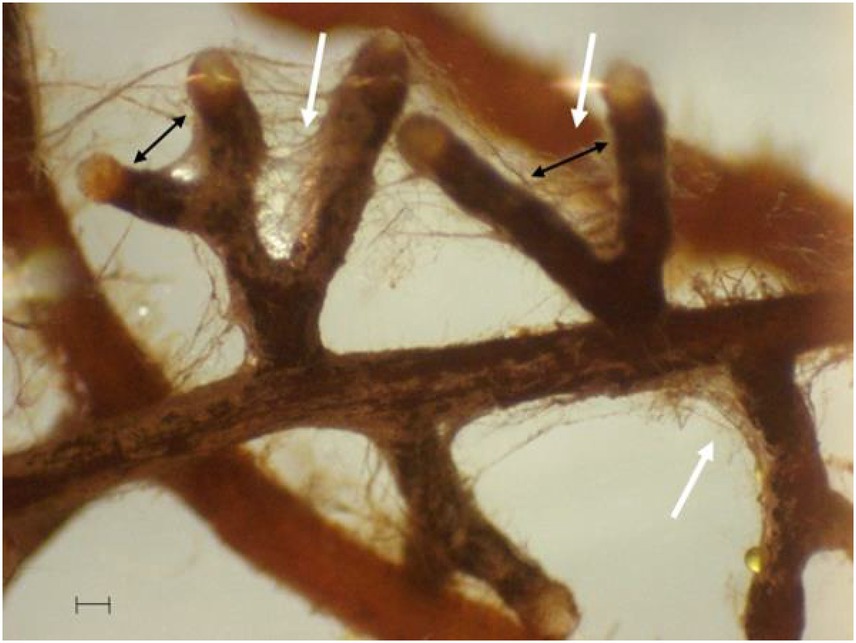
Figure 3. Characteristic dichotomous root tips induced by P. involutus in P. sylvestris seedlings (black double arrows), surrounded by mycelium (white arrows). Scale bar = 1 μm.
3.1.2 Plant growth
Total plant biomass (i.e., shoot + roots) was significantly affected by the factor Myc (F₁, ₁₆ = 3.23, p < 0.01) and the factor WR (F₁, ₁₆ = 2.90, p < 0.05). No interaction effect was observed between both factors. Pairwise comparisons showed that the total plant biomass in the NM-LWR treatment was significantly lower than that of the plants in the M-NWR treatment. No other differences were observed among the treatments (Figure 4A). A major impact of the factors Myc and WR was observed on the aboveground biomass of the plants of the NM-LWR treatment (F₁, ₁₆ = 10.4, p < 0.001, F₁, ₁₆ = 8.42, p < 0.005, respectively), with values significantly lower than that of all other treatments (Figure 4B). The LWR impacted aboveground biomass but not belowground biomass (Figure 4C). The factor Myc significantly impacted the root/shoot ratio (F₁, ₁₆ = 6.11, p < 0.05), while the factor WR had no significant effect on this parameter. Pairwise comparisons showed that the plants grown in the M-NWR and M-LWR had a significantly lower root/shoot ratio than those grown in the NM-LWR treatment (Figure 4D). An ANCOVA was performed to assess the effect of treatment on the root/shoot ratio with total biomass as covariate. The analysis showed that treatment had a significant effect (p < 0.05), while total biomass and its interaction with treatment were not significant (p > 0.05), indicating that treatment differences in root/shoot ratio were independent of plant size.
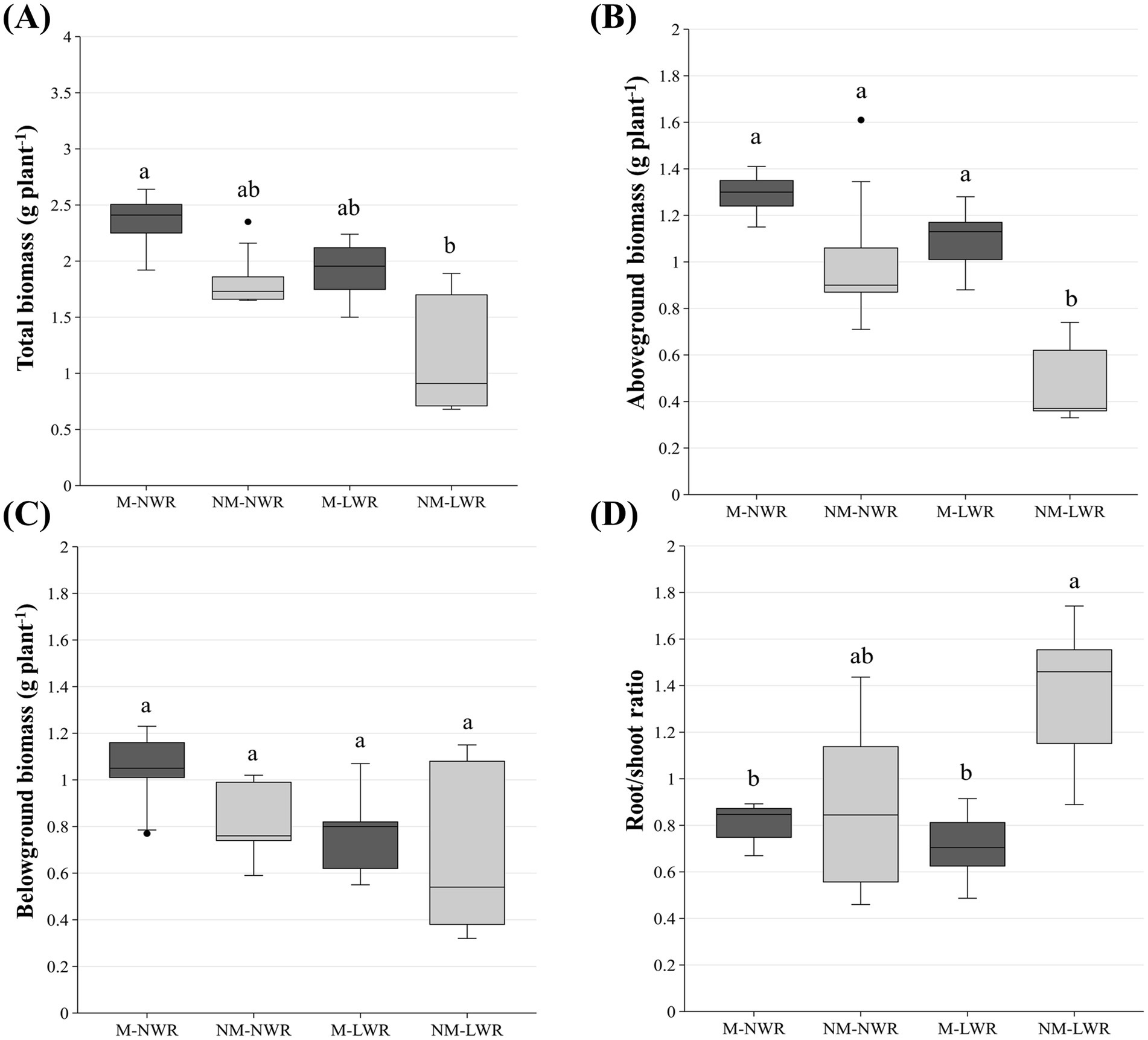
Figure 4. Exp 1: impact of water regime and root colonization of P. sylvestris by P. involutus on (A) total plant biomass (i.e., shoot + roots); (B) on aboveground biomass, (C) on belowground biomass and (D) on root/shoot ratio. The box plot displays the interquartile range (IQR) of the scores (central boxes), with the lower edge of the box indicating the first quartile and the upper edge marking the third quartile. The horizontal line inside each box represents the median score. The whiskers extend to the smallest and largest values within 1.5 * IQR from the first and third quartiles. Scores outside this range are considered outliers and are depicted as individual points. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. LWR: low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
3.1.3 Plant physiology
The photosynthetic rate was significantly affected by the factor Myc (F₁, ₁₆ = 27.2, p < 0.0001) and the factor WR (F₁, ₁₆ = 6.20, p < 0.05). No significant effect was observed for the interaction Myc x WR. Pairwise comparison showed that the plants of the NM-LWR treatment had a significantly lower photosynthetic rate than the plants of the M treatments (Figure 5A) but did not differ from the plants in the NM-NWR treatment. Similarly, transpiration rate was significantly affected by the factor Myc (F₁, ₁₆ = 5.90, p < 0.05) and the factor WR (F₁, ₁₆ = 6.00, p < 0.05). Pairwise comparison showed that the plants of the NM-LWR treatment had a significantly lower transpiration rate than the plants of the M-NWR treatment (Figure 5B). Water use efficiency was significantly impacted by the factor Myc (F₁, ₁₆ = 30.0, p < 0.0001) but not by the factor WR. The interaction between the two factors was also significant (F₁, ₁₆ = 6.20, p < 0.05). Pairwise comparison showed that the plants of the NM-LWR treatment had a significantly lower water use efficiency compared to those in the M-LWR and M-NWR treatments, but did not differ from the plants in the NM-NWR treatment. In addition, plants in the M-LWR treatment had significantly higher water use efficiency than both the NM treatments (Figure 5C). The other physiological parameters (i.e., stomatal conductance, water potential) did not differ significantly between the treatments (Supplementary material S1).
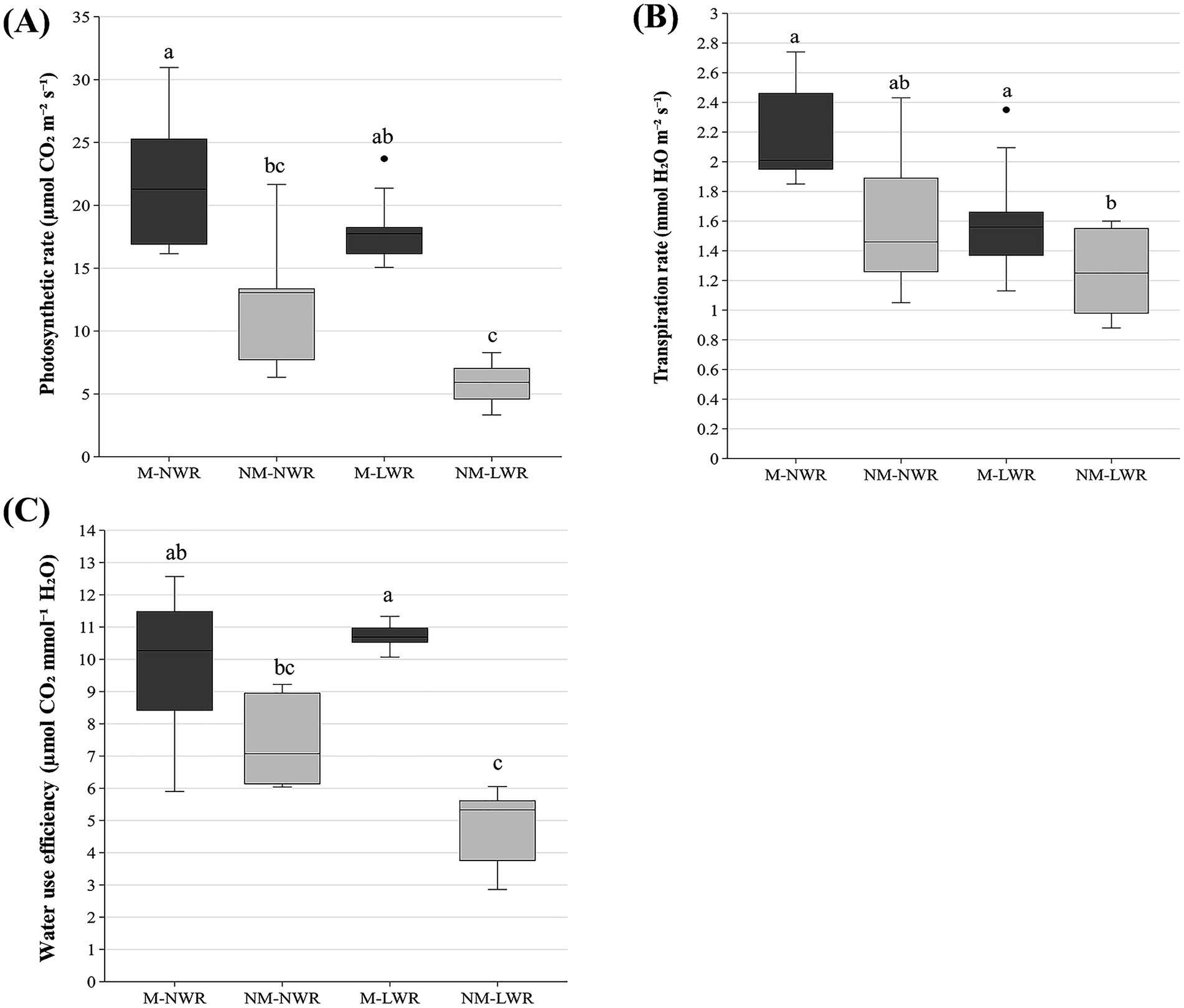
Figure 5. Exp 1: impact of water regime and root colonization of P. sylvestris by P. involutus on (A) photosynthetic rate (B) transpiration rate and (C) water use efficiency. The box plot displays the IQR of the scores, with the lower edge of the box indicating the first quartile and the upper edge marking the third quartile. The horizontal line inside each box represents the median score. The whiskers extend to the smallest and largest values within 1.5 * IQR from the first and third quartiles. Scores outside this range are considered outliers and are depicted as individual points. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. LWR: low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
3.1.4 Plant nutrient status
Nitrogen, P and C concentrations of plants were not significantly impacted by the factors Myc and WR (Figure 6). Conversely, the N content of plants was significantly impacted by the factor Myc (F₁, ₁₆ = 4.80, p < 0.05), the factor WR (F₁, ₁₆ = 5.00, p < 0.05) and their interaction (F₁, ₁₆ = 6.90, p < 0.05). Pairwise comparisons showed that N content was significantly lower in the plants of the NM-LWR treatment compared to the plants in the other treatments (Figure 7A). Plant P content was also affected by the factor Myc (F₁,₁₆ = 18.2, p < 0.001) and WR (F₁,₁₆ = 4.70, p < 0.05) but not by their interaction. The total P content in the plants of the NM-LWR treatment was significantly lower compared to that of the plants in the M treatments but did not differ from those in the NM-NWR treatment (Figure 7B). The total C content of the plants was also significantly impacted by the factor Myc (F₁,₁₆ = 10.7, p < 0.01) and the factor WR (F₁,₁₆ = 7.30, p < 0.05). No effect of the interaction between factors Myc x WR was found for this parameter. Pairwise comparison showed that the total C content of the plants of the NM-LWR treatment was significantly lower compared to that of the plants in the M-NWR treatment, but did not differ from the two other treatments (Figure 7C).
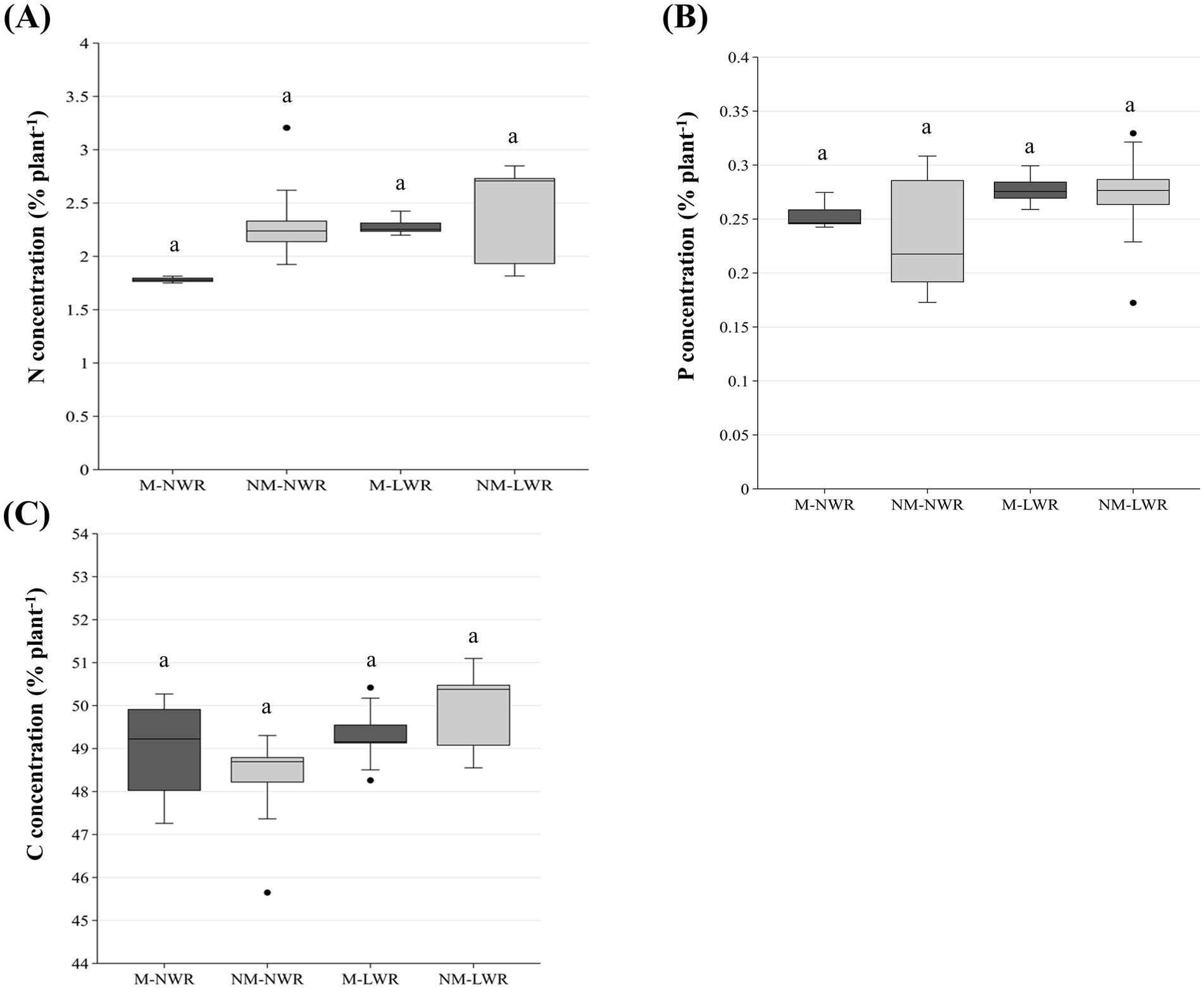
Figure 6. Exp 1: impact of water regime and root colonization of P. sylvestris by P. involutus on concentrations of (A) nitrogen (N), (B) phosphorus (P) and (C) carbon (C). The height of each bar indicates the average concentration of the respective element for that treatment. Error bars on top of each bar represent the standard deviation. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. LWR: low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
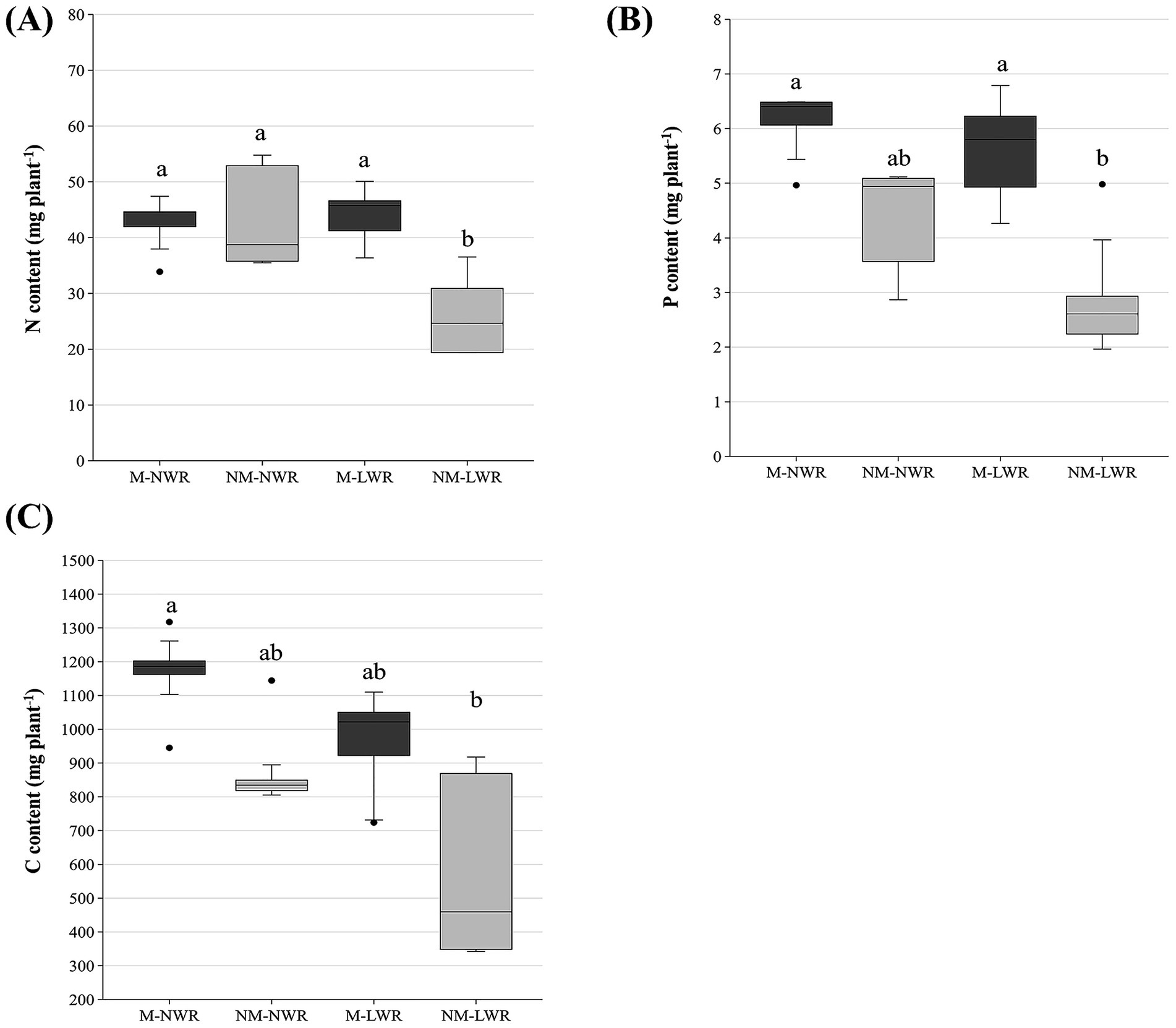
Figure 7. Exp 1: impact of water regime and root colonization of P. sylvestris by P. involutus on contents of (A) nitrogen (N), (B) phosphorus (P) and (C) carbon (C). The height of each bar indicates the average content of the respective element for that treatment. Error bars on top of each bar represent the standard deviation. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. LWR: low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
3.1.5 Plant recovery
At the end of the recovery phase, the colonization rate did not differ significantly between the plants in the M-LWR treatment and those in the M-NWR treatment. The percentage of mycorrhization was 78.9 ± 8.3% and 91.2 ± 5.4%, respectively. No root colonization was observed in the plants of the NM-NWR and NM-LWR treatments.
Plant growth and physiological parameters were not affected by the factor Myc, while the factor WR affected significantly the photosynthetic rate (F₁,₁₆ = 4.60, p < 0.05), the transpiration rate (F₁,₁₆ = 5.30, p < 0.05), stomatal conductance (F₁,₁₆ = 4.70, p < 0.05) with a decrease of 24.1, 17.5 and 19.8%, respectively, in plants of the LWR treatment compared to those in the NWR treatment. Water use efficiency was not affected by neither factor (Supplementary material S2). In plants subjected to LWR, the difference in chlorophyll content before and after recovery was not significantly impacted by the factors Myc and WR (Supplementary material S2).
3.2 Experiment 2
3.2.1 First cycle of drought treatment
The following sections describe the effects of water regime WR (moderate water regime, MWR, and normal water regime, NWR) and mycorrhization (Myc) on root colonization, plant growth, physiological parameters, and recovery phase of P. sylvestris seedlings.
3.2.1.1 Root colonization
Root colonization rates were not affected by the WR. For the mycorrhizal plants grown under NWR and MWR, the rate of root colonization was 64.2 ± 18.1% and 54.3 ± 15.0%, respectively. No root colonization was observed for the plants in the NM treatments.
3.2.1.2 Plant growth
The factor Myc and WR and their interaction did not impact any of the growth parameters (Supplementary material S3).
3.2.1.3 Physiological parameters
The photosynthetic rate was not significantly impacted by the factor WR and the factor Myc. Transpiration rate was significantly affected by the factor WR (F₁,₁₆ = 24.5, p < 0.0001) and by the factor Myc (F₁,₁₆ = 8.13, p < 0.01) but not by their interaction. Pairwise comparison showed that plants of the MWR treatments had a significantly lower transpiration rate compared to those in the M-NWR treatment, but did not differ from the plants in the NM-NWR treatment. Stomatal conductance was also significantly affected by the factor WR (F₁,₁₆ = 19.7, p < 0.0005), with the plants in the MWR treatments showing a significant difference with the plants in M-NWR treatment. Pairwise comparison did not show any significant effects on water use efficiency and water potential (Supplementary material S3).
3.2.1.4 Plant recovery
The colonization rate was not impacted by the factor WR. Root colonization of the plants in the M-NWR and M-MWR treatments was 75.9 ± 18.9% and 62.5 ± 21.7%, respectively. No root colonization was observed in the plants of the NM treatments.
The factor Myc had a significant effect on photosynthetic rate, transpiration rate, and water use efficiency (F₁,₁₆ = 15.2, p < 0.001; F₁,₁₆ = 8.10, p < 0.01; F₁,₁₆ = 6.70, p < 0.05, respectively), while stomatal conductance was not affected. The factor WR significantly affected the photosynthetic rate and transpiration rate (F₁,₁₆ = 7.70, p < 0.05; F₁,₁₆ = 24.5, p < 0.0001). Pairwise comparison showed that photosynthetic rate was significantly higher in the plants of M-NWR treatment compared to the other treatments, while transpiration rate was significantly higher compared to MWR treatments but not compared to the NM-NWR treatment (Figure 8).
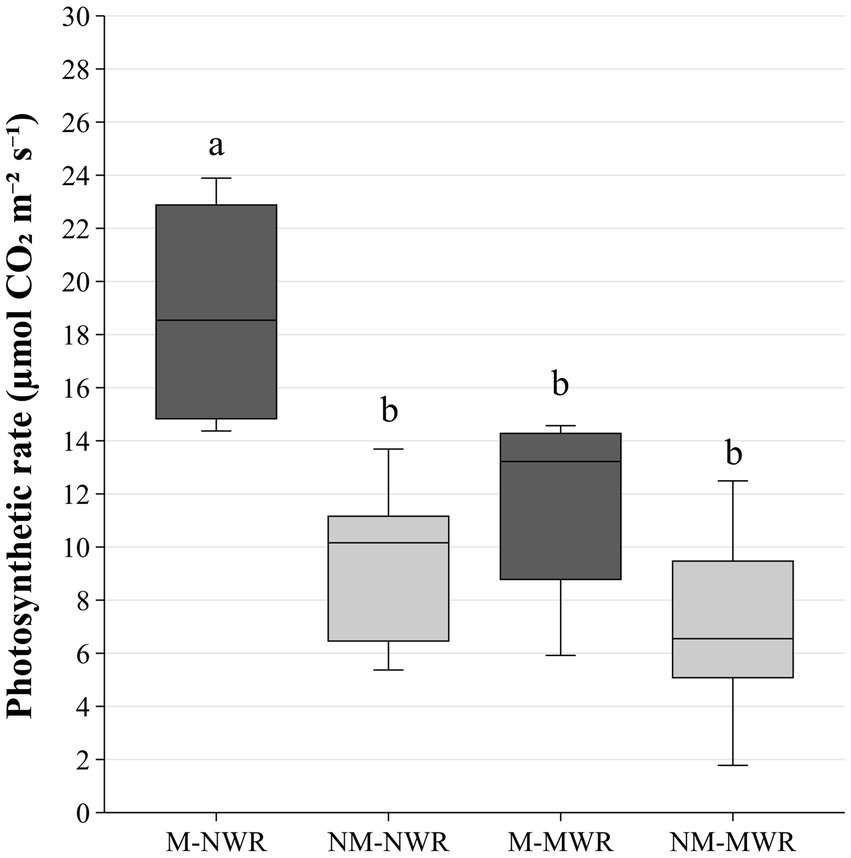
Figure 8. Exp 2: impact of water regime and root colonization of P. sylvestris by P. involutus on photosynthetic rate after recovery phase. The box plot displays the IQR of the scores, with the lower edge of the box indicating the first quartile and the upper edge marking the third quartile. The horizontal line inside each box represents the median score. The whiskers extend to the smallest and largest values within 1.5 * IQR from the first and third quartiles. Scores outside this range are considered outliers and are depicted as individual points. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. MWR: moderate water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
Pairwise comparison did not show any significant effects on the growth and the other physiological parameters (Supplementary material S4).
During the recovery phase, no significant difference was observed between the treatments in the depletion of Pi (Supplementary material S4). Conversely, The factor Myc (F₁,₁₆ = 31.7, p < 0.0001), WR (F₁,₁₆ = 21.5, p < 0.0001), and their interaction (F₁,₁₆ = 29.3, p < 0.0001) had a significant effect on the depletion of NH4+ at D3T3. Pairwise comparisons showed a significative lower value of NH4+ concentration in the solution in plants of M-NWR treatment at D3T3 compared to the other treatments (Figure 9).
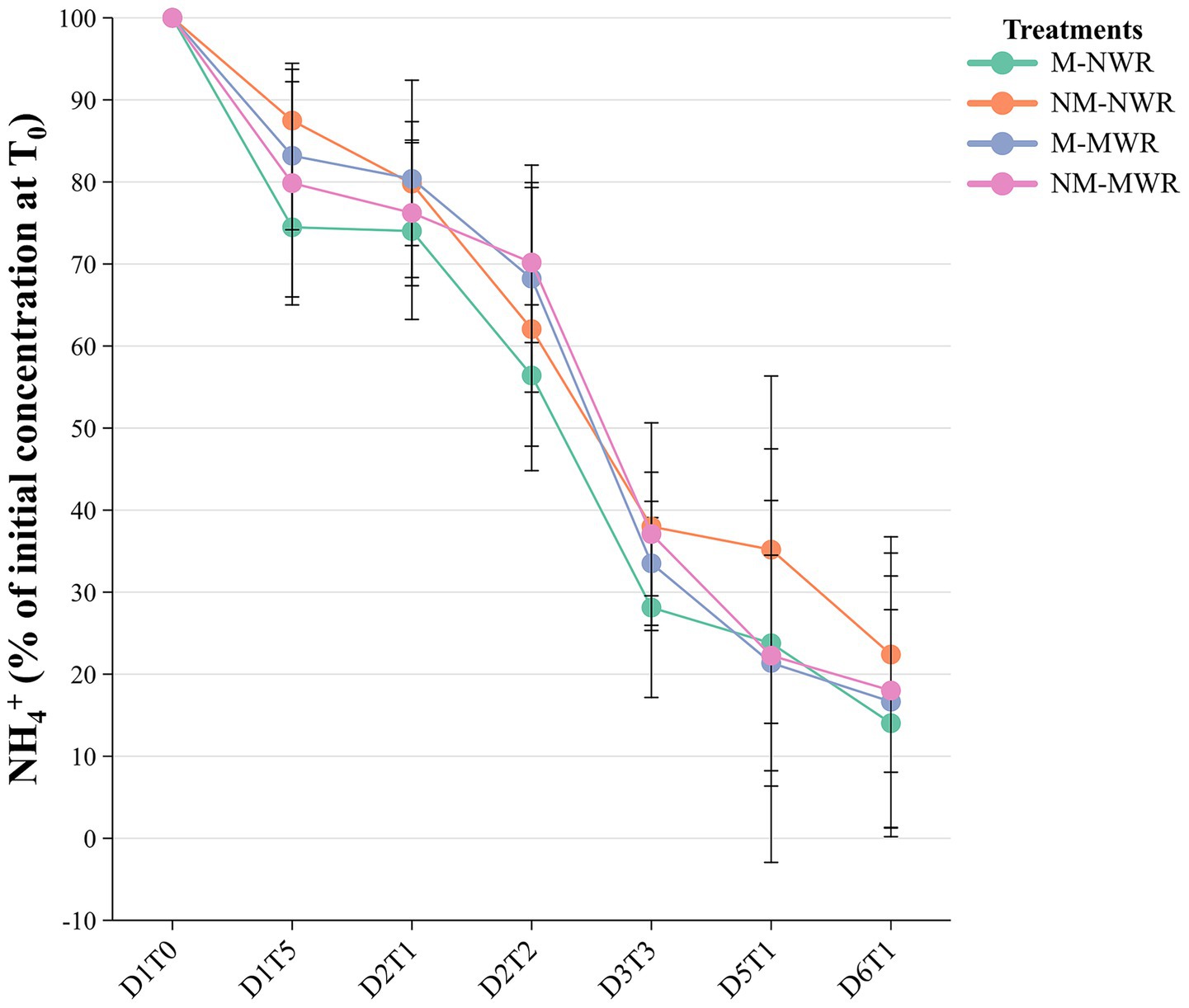
Figure 9. Exp 2: impact of water regime and root colonization of P. sylvestris by P. involutus on the depletion of ammonium during recovery phase. Data (n = 8/12) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05) per each day and timing. MWR: moderate water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants; D = day of sampling; T = time of sampling during the day.
In plants subjected to MWR, the difference in chlorophyll content before and after recovery was not significantly affected by the factors Myc and WR (Supplementary material S4).
3.2.2 Second cycle of drought treatment
The following sections describe the effects of water regime WR (prolonged low water regime, PLWR, and normal water regime, NWR) and mycorrhization (Myc) on root colonization, plant growth, physiological parameters, and recovery phase of P. sylvestris seedlings.
3.2.2.1 Root colonization
Root colonization was significantly affected by WR (p = 0.0316). It was significantly higher for the plants in the M-NWR treatment (i.e., 94.2 ± 9.5%) compared to the plants in the M-PLWR treatment (i.e., 63.7 ± 22.1%). No root colonization was observed for the plants in the NM treatments.
3.2.2.2 Plant growth
Total biomass was significantly impacted by the factor Myc (F₁,₁₆ = 6.70, p < 0.05) but not by the factor WR. No interaction effect between both factors was observed. Pairwise comparison showed that the biomass of plants in the NM-LWR treatment was significantly lower than in those of the M-NWR treatment (Figure 10). No other growth parameters (i.e., fresh weight, root volume, shoot height) differed between the treatments (Supplementary material S5).
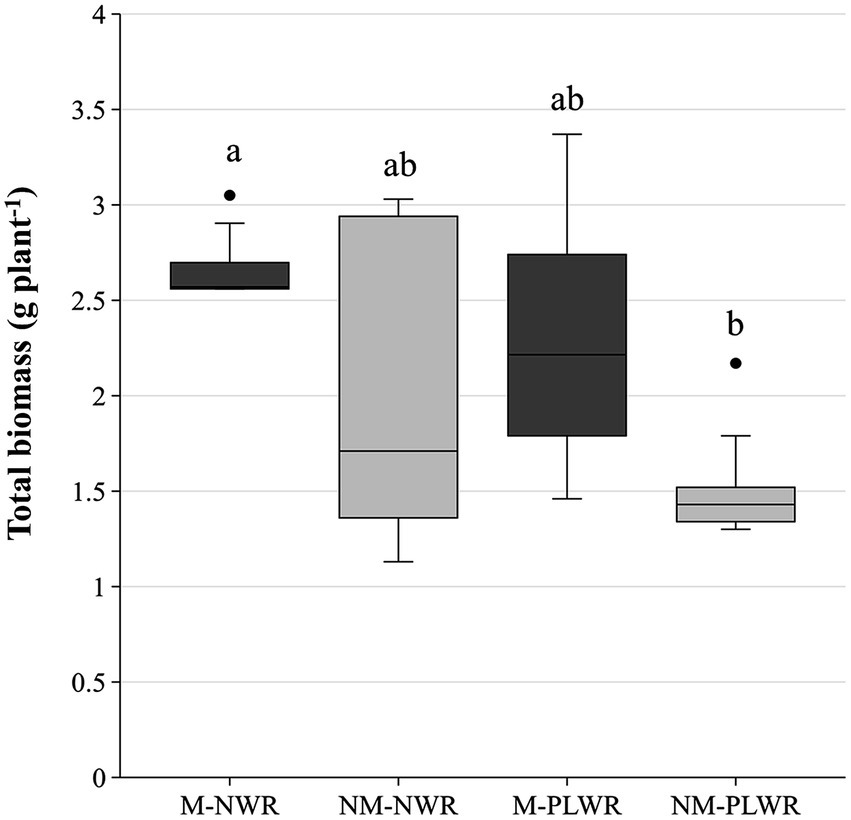
Figure 10. Exp 2: impact of water regime and root colonization of P. sylvestris by P. involutus on plant total biomass. The box plot displays the IQR of the scores, with the lower edge of the box indicating the first quartile and the upper edge marking the third quartile. The horizontal line inside each box represents the median score. The whiskers extend to the smallest and largest values within 1.5 * IQR from the first and third quartiles. Scores outside this range are considered outliers and are depicted as individual points. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. PLWR: prolonged low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
3.2.2.3 Physiological parameters
Physiological parameters photosynthetic rate, transpiration rate and stomatal conductance were significantly affected by the factor WR (F₁,₁₆ = 10.2, p < 0.01), (F₁,₁₆ = 30.2, p < 0.0001), (F₁,₁₆ = 19.4, p < 0.001), but water use efficiency was not affected by WR. The factor Myc did not significantly affect these parameters. Pairwise comparisons showed that plants in the NM-PLWR had a significantly lower photosynthetic rate compared to the M-NWR treatment and significantly lower transpiration rate and stomatal conductance compared to the NWR treatments (Figures 11A–C).
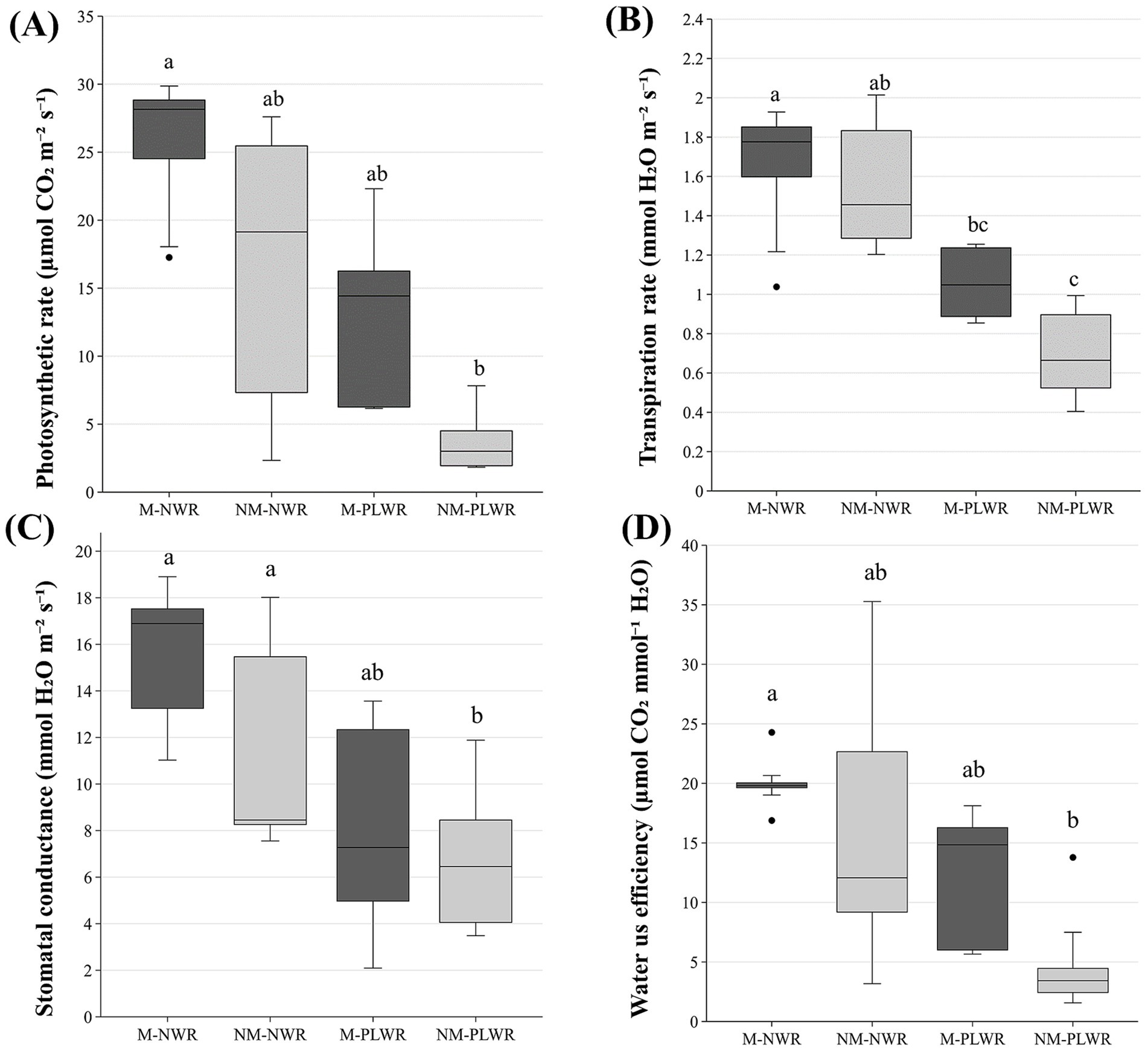
Figure 11. Exp 2: impact of water regime and root colonization of P. sylvestris by P. involutus on (A) photosynthetic rate, (B) transpiration rate, (C) stomatal conductance and (D) water use efficiency. The box plot displays the IQR of the scores, with the lower edge of the box indicating the first quartile and the upper edge marking the third quartile. The horizontal line inside each box represents the median score. The whiskers extend to the smallest and largest values within 1.5 * IQR from the first and third quartiles. Scores outside this range are considered outliers and are depicted as individual points. Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. PLWR: prolonged low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
3.2.2.4 Plant recovery
The colonization rate was significantly impacted by the factor WR (p < 0.01) with values significantly higher for the plants in the M-NWR treatment (90.8 ± 13.8%) compared to the plants in the M-PLWR treatments (40.0 ± 14.7%). No root colonization was observed for the plants in the NM treatments.
Plant growth parameters were not affected by the factors Myc, WR or their interaction, during recovery phase and did not show significant differences between the treatments (Supplementary material S6).
The photosynthetic rate was significantly affected by the factor Myc (F₁,₁₆ = 8.50, p < 0.01) and by the factor WR (F₁,₁₆ = 15.5, p < 0.001), but not by their interaction. Pairwise comparison showed that the photosynthetic rate in plants of the NM-PLWR treatment was significantly lower than those in the M-NWR treatment, while no other differences were observed. Similarly, stomatal conductance and water use efficiency were significantly affected by the factor WR (F₁,₁₆ = 10.6, p < 0.01 and F₁,₁₆ = 8.20, p < 0.05, respectively) but not by the factor Myc. No interaction between both factors was observed. The other physiological parameters (i.e., transpiration rate, and water potential), did not show a significant effect (Supplementary material S6).
The chlorophyll content was significantly influenced by the factor Myc (F₁,₁₆ = 32.5, p < 0.0001), but not by the factor WR, although their interaction was significant (F₁,₁₆ = 32.2, p < 0.0001). Pairwise comparison showed a significant decrease in chlorophyll content in plants of the NM-PLWR treatment compared to those in the NWR treatment, while the plants in the M-PLWR treatment showed a significant increase (Figure 12).
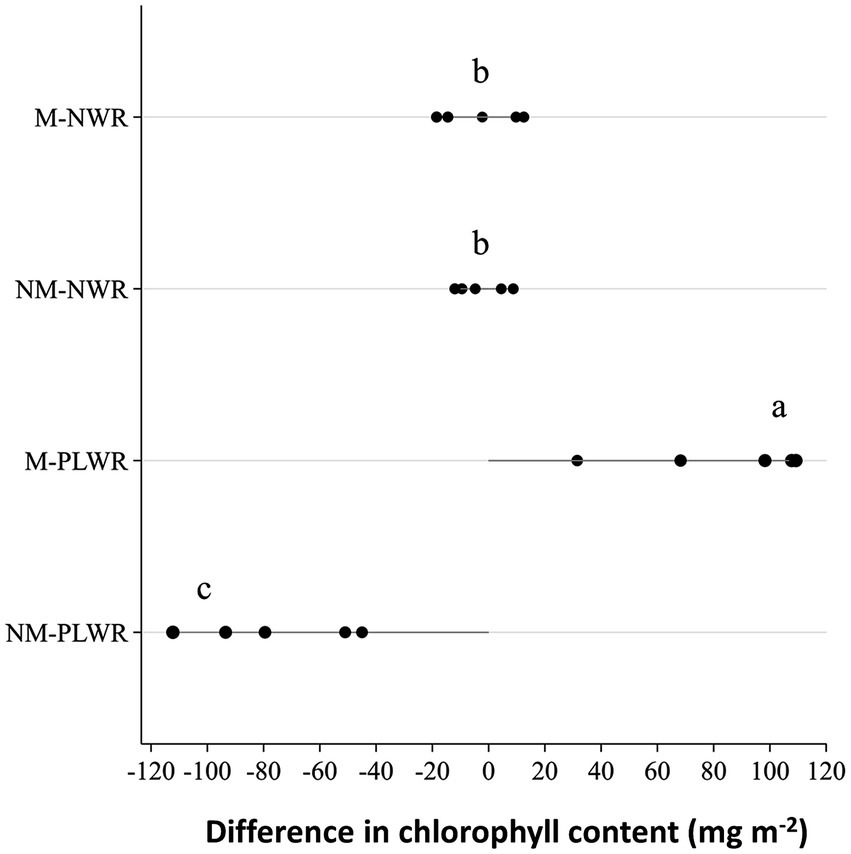
Figure 12. Exp 2: impact of water regime and root colonization of P. sylvestris by P. involutus on the difference in chlorophyll content before and after the recovery phase. Each point represents the actual value of the data point being plotted, each line extends from the baseline (0). Data (n = 5) were analyzed by a GLM followed by a Tukey post-hoc test (p ≤ 0.05). Different lower-case letters indicate significant differences between the treatments. PLWR: prolonged low water regime; NWR: normal water regime; M: mycorrhizal plants; NM: non-mycorrhizal plants.
4 Discussion
4.1 Effect of P. involutus on seedlings growth and physiological parameters under contrasting water regimes
Ectomycorrhizal fungi have been widely studied for their contributions to forest regeneration, mainly because of their potential to enhance the growth and survival of trees under adverse conditions (Brundrett, 2009). However, studies of how different drought treatments affect tree seedlings inoculated with EcM fungi have produced mixed results, often showing variability depending on fungal and tree species, plant’s age and environmental context.
Here, we compared the growth and physiology of P. sylvestris seedlings, associated or not to the EcM fungus P. involutus, under different drought treatments (i.e., normal water regime (NWR), moderate water regime (MWR), low water regime (LWR), and prolonged low water regime (PLWR)), and during recovery from drought treatments. We have shown that mycorrhizal plants exhibit increased aboveground biomass, photosynthetic rate and water use efficiency, particularly under LWR.
4.1.1 Low water regime
Under LWR conditions, the total plant biomass (shoot + roots) and shoot biomass of seedlings associated with the EcM fungus was significantly higher compared to the non-colonized ones, while no difference was observed for root biomass (Figures 4A–C). This increase in shoot biomass suggests that the plants associated to the EcM fungus allocated more resources to aboveground parts. This result corroborates the study of Yin et al. (2018) who explored how P. sylvestris var. mongolica seedlings inoculated with S. luteus responded to water stress. They found that EcM fungal-inoculated seedlings exhibited significantly greater growth than their non-inoculated counterparts. Similarly, Li et al. (2023) demonstrated that P. massoniana seedlings inoculated with Suillus luteus had significantly higher aboveground biomass, but in contrast to our study, also higher root biomass, under conditions of moderate and severe drought treatments. This was also reported by Wang et al. (2021), who observed that the inoculation of P. tabulaeformis seedlings with S. variegatus resulted in a marked increase in leaf biomass and as well root biomass, all under conditions of moderate and severe drought treatments. This suggests that EcM fungal inoculation can provide a buffer against drought, helping seedlings to maintain growth and development under limited water conditions. Results may also vary according to the EcM fungus as shown by Kipfer et al. (2012). These authors discovered that among eight EcM fungal species tested, only S. granulatus had a significant positive effect on the shoot growth of P. sylvestris seedlings because it was the fungus with the highest activity of secreted extracellular enzymes. This beneficial effect was more pronounced under well-watered conditions compared to dry conditions, suggesting that EcM fungi may not always provide additional support during periods of drought. This variability in EcM fungal effectiveness could be attributed to the fungal species, the extent of colonization, and the environmental context in which the symbiosis occurs.
When comparing the root-to-shoot ratio, we noticed that it was significantly higher in the non-colonized seedlings under LWR (Figure 4D), suggesting that these plants allocated more resources to root growth, potentially as a strategy to access limited water resources. However, this allocation came at the expense of aboveground parts, indicating a trade-off between root and shoot growth under drought conditions. Moser et al. (2015) also emphasized the importance of root architecture plasticity in P. sylvestris seedlings during early establishment under drought conditions. Their results suggested that the structural adaptability of the root system played a more crucial role in the seedlings’ drought resistance than colonization by EcM fungi. This supports the hypothesis that while EcM symbiosis can be beneficial, other factors such as the inherent plasticity in biomass allocation patterns may also play an important role in plant survival under stress.
The resource allocation pattern in NM-LWR seedlings (where resources are allocated in a way that prioritizes root biomass over aboveground growth) could also be hypothetically explained by the reduced photosynthetic rate observed. Indeed, the NM-LWR seedlings showed a 63.8% decrease in photosynthesis compared with the M-LWR seedlings (Figure 5A). Colpaert et al. (1996) also found that EcM fungi stimulated net photosynthesis in P. sylvestris seedlings colonized with S. bovinus, but in this case this increase did not translate into greater shoot growth and failed to compensate for the costs of maintaining the symbiotic relationship. Kipfer et al. (2012), did not investigate photosynthetic activity but in contrast reported that EcM fungal inoculation increased aboveground biomass in P. sylvestris seedlings, but only when inoculated with S. granulatus. In presence of P. involutus, Cenococcum geophilum or Rhizopogon roseolus, no significant increase in aboveground biomass was observed under either dry or wet conditions. This suggests that the benefits of EcM fungal inoculation is highly species-specific.
Under LWR, the photosynthetic rate of non-EcM fungus-associated seedlings was significantly negatively affected. In contrast, in presence of the EcM fungus, the seedlings exhibited no significant differences in photosynthesis compared to the well-watered treatments. This indicated that the association with the EcM fungus helped the seedlings to maintain photosynthetic efficiency under drought treatment conditions. Furthermore, we observed that the transpiration rate and WUEt differed significantly between the M and NM seedlings under LWR (Figures 5B,C). However, neither group exhibited differences from their respective well-watered controls, suggesting that the EcM fungus colonized seedlings maintained a higher WUEt across different water regimes and M-LWR seedlings maintained an even higher WUEt compared to NM seedlings in the control group (Figure 5C). This could be explained by the demand of EcM fungi for carbon, which in turn promotes photosynthesis (Lamhamedi et al., 1994). As a consequence of the increased carbohydrate demand in symbiosis, trees increase their photosynthetic capacity (Nehls et al., 2010). Similarly, Li et al. (2023) also highlighted the role of S. luteus in improving the plant’s hydraulic regulation, particularly by increasing WUE and promoting a more balanced water status in the plant.
On the other hand, the fact that the non-inoculated seedlings showed a reduced photosynthetic rate and aboveground biomass, could be hypothetically attributed to the isohydric behavior of P. sylvestris, characterized by strict stomatal control that limits transpiration when soil moisture falls below a certain threshold (Irvine et al., 1998; Poyatos et al., 2007; Martínez-Vilalta et al., 2009), even if data of stomatal conductance and water potential in the present work do not truly validate this hypothesis. It has also to be considered that this isohydric strategy, while helping to conserve water, also limits carbon assimilation (Martín-Gómez et al., 2017).
When analyzing the nutrient status of the seedlings subjected to NWR and LWR, we found that the concentrations of N and P did not differ significantly between the seedlings associated or not to the EcM fungus (Figures 6A,B). However, the N content in NM-LWR seedlings was 39.5% lower compared to M-LWR seedlings (Figure 7A). This difference in N content is directly linked to the increased shoot biomass in the M-LWR seedlings. Similar differences were observed in P content, with NM-LWR seedlings exhibiting 46.3% lower P content compared to M-LWR seedlings, while no significant differences were observed between NM-LWR and NM-NWR seedlings (Figure 7B). The inoculated plants in the LW treatment received less water and nutrients than those in the NW treatment. Nevertheless, they absorbed similar amounts of nutrients, highlighting the importance of mycorrhizae for N and P uptake when both water and nutrients are limited. Since nutrient concentrations did not change, this also indicates that the absorbed nutrients were effectively used to sustain growth. This finding is particularly noteworthy, as water deficit typically leads to reduced growth as one of its earliest effects.
Carbon concentration (Figure 6C) and content (Figure 7C) remained stable across treatments, with the exception of a substantial variation between NM-LWR and M-NWR seedlings. This disparity is most likely due to the different physiological and allocation strategies used during drought stress and in response to ectomycorrhizal symbiosis. Non-mycorrhizal seedlings exposed to drought typically exhibit reduced stomatal conductance and photosynthesis, thereby limiting carbon assimilation (Dosskey et al., 1991; Davies et al., 1996). In response, biomass allocation often shifts toward root development to enhance water uptake, resulting in increased root: shoot ratios in non-mycorrhizal individuals (Davies et al., 1996). In contrast, mycorrhizal seedlings grown under ideal watering circumstances benefit from increased nutrient absorption and development (Koele and Hildebrand, 2011), but must allocate a significant amount of their photosynthates to sustain the fungal symbiont (Langley et al., 2006). The carbon drain caused by ectomycorrhizal colonisation is estimated to account for 10–20% of the host’s photosynthetic production (Langley et al., 2006; Smith and Read, 2008).
The allocation of nutrients to shoots and roots also showed interesting patterns. In all treatments except NM-LWR, N, P, and C were preferentially allocated to the shoots, which aligns with the increased shoot biomass observed in these treatments. However, in NM-LWR seedlings, the higher root-to-shoot ratio suggests a greater allocation of resources to belowground parts. This could be a response to drought treatment, as the seedlings attempt to maximize water uptake through increased root growth and metabolic activity, even at the expense of shoot biomass. Such resource allocation strategies are commonly observed in plants under water-limited conditions, where prioritizing root growth can enhance access to water and improve drought tolerance. Carbon allocation, in particular, is generally prioritized to tissues increasing the uptake of limiting resources (Freschet et al., 2018).
4.1.2 Moderate and prolonged low water regimes
Under MWR conditions, we did not observe the same beneficial effects of EcM fungus inoculation as seen under LWR. One possible explanation is that MWR did not induce sufficient stress in P. sylvestris, which is known to be relatively drought-tolerant (Bose et al., 2020), to affect growth. Seedlings maintained stable water potential through stomatal regulation under MWR, avoiding growth limitation despite reduced photosynthesis. The absence of a reduction in biomass under water deficit in Experiment 2, unlike in Experiment 1, may suggest physiological acclimation. In this experiment, only the well-watered control plants (NWR) received regular nutrient supplementation, while the drought-treated plants (LWR, MWR, PLWR) did not, in order to simulate the reduction in nutrient availability typically associated with water deficit.
Under PLWR conditions, a significantly reduced level of root colonization was observed (from 94.3% under NWR to 63.8%), indicating that the mycelium of P. involutus was adversely affected by prolonged drought treatment, or that the symbiosis was downregulated by the plant when the cost–benefit balance was no longer favorable. This reduction in colonization likely decreased the ability of the EcM fungus to support the photosynthetic rate and growth of the plants, as observed under LWR. These results are consistent with those of Wang et al. (2021), who found that EcM fungal colonization in P. tabulaeformis seedlings was lowest under severe drought conditions and highest under moderate drought. Kennedy and Peay (2007) showed that the benefits of ectomycorrhizal symbiosis between P. muricata and Rhizopogon species are highly dependent on soil moisture. Under moderate moisture, mycorrhizal plants outperformed non-mycorrhizal ones, but under severe drought, reduced fungal colonization led to the collapse of mutualistic benefits. This suggests that EcM fungi are themselves sensitive to the severity of drought and that their ability to confer drought tolerance to seedlings may depend on maintaining an adequate level of root colonization. Hagenbo et al. (2021) found that ectomycorrhizal mycelial production and turnover in P. pinaster and P. sylvestris forests were highly responsive to short-term variations in precipitation. Mycorrhizal biomass showed strong seasonal fluctuations, with increased growth during wetter periods and sharp declines during drought, indicating a close association between fungal activity and water availability. Gehring et al. (2020) showed that chronic drought significantly reduced ectomycorrhizal abundance and diversity in P. edulis. Similarly, Swaty et al. (2004) discovered that drought stress decreased ectomycorrhizal colonization and fungal community variability, with severely drought-affected areas having fewer active root tips and a different fungal community structure, but, interestingly, trees under moderate drought stress showed the largest EcM colonization, indicating a nonlinear response of the symbiosis to water availability. Inoculated seedlings subjected to PLWR that were earlier exposed to MWR and recovery phase showed no significant differences in leaf gas exchange parameters compared to the NWR controls, except for transpiration rate that was significantly lower (Figures 11A–D). On the other hand, NM-PLWR seedlings showed significantly lower transpiration rate and stomatal conductance compared to the seedlings in well-watered conditions, but not photosynthetic rate or water use efficiency. This finding suggests the possibility of drought-induced plant memory or a priming effect, wherein plants that have experienced a stress event “remember” it and adjust their responses to future stress (Bruce et al., 2007). Priming can be defined as the potential of plants to memorize environmental stress events and to strengthen their response to repeated stress (Harris et al., 2023).
During recovery, these stored stress imprints allow the plant to respond more quickly or effectively to subsequent stress events, thanks to the accumulation of metabolites and epigenetic modifications (Jacques et al., 2021). In our case, the seedlings’ ability to recover rapidly after PLWR suggests that some form of drought memory may be involved. However, more in-depth investigations, including genetic and molecular analyses, would be required to confirm whether such a priming effect is really present in P. sylvestris. Although some studies have demonstrated a priming effect to drought induced by EcM fungi in poplar trees (Luo et al., 2009) and pedunculate oak (Kebert et al., 2023), it is not possible to affirm the EcM fungus was involved in a possible priming effect in our study. Remarkably, Erlandson et al. (2022) demonstrated that long-term drought causes the EcM fungus S. pungens to exhibit a transcriptional acclimation response. Specifically, the fungus, showed no significant change in regulation of gene expression activity, when exposed to chronic drought. This finding suggests that some EcM fungi may undergo a molecular shift towards drought tolerance, potentially contributing to the host plant’s increased resilience. Although it is unknown if P. involutus exhibits comparable behavior, these fungal acclimation processes could significantly influence how the host plant responds to drought over time.
On the other hand, P. involutus root colonization was significantly reduced under PLWR conditions even after recovery phase, indicating that the EcM fungus was also strongly impacted by prolonged drought treatment. This decrease in P. involutus root colonisation is consistent with prior research demonstrating the harmful effects of drought on EcM fungi. For instance, Parke et al. (1983) found that mycorrhizal colonization in Pseudotsuga menziesii seedlings declined with water stress, with non-watered plants showing fewer surviving mycorrhizal root tips than well-watered controls. Coleman et al. (1989) also showed that the growth of EcM fungi, including Paxillus species, was severely inhibited under low water conditions, indicating a direct susceptibility of fungal hyphae to water stress in pure culture. Supporting these controlled-environment results, Nickel et al. (2018) discovered that, in a temperate forest, 3 years of experimental summer drought resulted in a quantitative decrease in ectomycorrhizal colonization of fine roots, including in conifer species, while maintaining a relatively stable fungal community composition. Collectively, these investigations offer strong evidence that extended drought conditions impair both the colonization of roots and the development of the EcM fungi.
This reduction in colonization may have decreased the potential role of the EcM fungus in helping plants to recover from drought. Based on our data, it is difficult to definitively conclude whether the EcM fungus played a role in the potential priming effect observed in the seedlings. However, the reduced colonization levels and the decreased physiological benefits under PLWR suggests that the mycorrhizal symbiosis may have been compromised under more extreme drought conditions.
4.2 Effect of P. involutus and water regime on seedlings’ recovery
Regardless of the water regime applied, all seedlings, whether mycorrhized or not, exhibited similar levels of physiological parameters after 6 days. No significant differences were observed compared to the control groups under NWR. This suggests that the seedlings were able to re-establish adequate photosynthetic rates and WUE after the period of drought treatment. However, nutrient uptake during the recovery phase revealed that seedlings in the M-NWR treatment had a higher ability to take up NH4+, suggesting that mycorrhizal colonization may facilitate more efficient nutrient uptake during the recovery phase (Figure 9), allowing them to maintain a higher photosynthetic rate compared to the other treatments (Figure 8), a phenomenon that is consistent with field observations of EcM mycelium production tracking moisture availability (Hagenbo et al., 2021; Querejeta et al., 2009). The functional rebound of EcM mycelium with improved moisture, as suggested by Hagenbo et al. (2021), could support both nutrient availability and possibly plant recovery. This is further corroborated by León-Sánchez et al. (2018), who showed in a 4-year manipulative study in a semi-arid shrubland that the decline in EcM presence, under rainfall reduction and warming, was correlated with reductions in foliar nitrogen, phosphorus, and dry matter content, reflecting impaired nutrient status and declining plant tissue quality. Nonetheless, by the end of the recovery period, seedlings from all treatments had depleted the NH4+ present in the solution, indicating that even non-mycorrhized seedlings were able to resume normal nutrient uptake after drought treatment. These results imply that, while EcM fungi may confer an advantage during the initial stages of recovery, the plants themselves can restore their nutrient uptake capacity.
During the recovery phase, an increase in chlorophyll content was observed in the M seedlings, while a reduction in chlorophyll was seen in the NM seedlings that had been exposed to PLWR (Figure 12). This discrepancy could be due to the damage to Photosystem II or nutrient limitations in NM seedlings, particularly those in the PLWR treatment, which may have hindered the rapid production of chlorophyll when water became available again. In contrast, M seedlings appeared to recover more efficiently, likely due to the increased nutrient availability provided by the EcM fungal symbiosis. Yin et al. (2018) found that inoculation with S. luteus inhibited the reduction of chlorophyll content in P. sylvestris var. mongolica seedlings under drought conditions, highlighting the potential role of EcM fungi in preserving chlorophyll content and protecting the photosynthetic machinery during drought. Our findings suggest that a similar mechanism may be at play in the recovery of P. sylvestris seedlings inoculated with P. involutus, particularly under prolonged drought conditions.
In conclusion, the results of our study demonstrate that the EcM fungus P. involutus plays a crucial role in growth and physiology of P. sylvestris seedlings under drought-simulated conditions. Mycorrhized seedlings exhibited enhanced shoot biomass, improved photosynthetic rates, and increased nutrient uptake compared to non-mycorrhized seedlings, particularly under low water conditions. Under a moderate water regime, the stress was not sufficient to affect the growth of the plants and also the effect of the EcM fungus could not be evidenced. Also, the benefits of EcM symbiosis were no longer visible under prolonged low water conditions, likely due to a reduction in root colonization by the fungus. The potential for drought memory or priming effects in P. sylvestris seedlings warrants further investigation, particularly in relation to the role of EcM fungi in mediating these processes.
Overall, our findings highlight the importance of considering both plant and fungal responses to drought when assessing the resilience of forest ecosystems in the face of climate change. Field-based studies have shown that EcM fungi are highly sensitive to changes in water availability (Bell and Adams, 2004). For instance, Fernandez et al. (2023) demonstrated that climate-induced drought disrupts EcM interaction networks in boreal–temperate ecotones, leading to a breakdown in symbiotic connectivity that may compromise forest stability. Similarly, Hagenbo et al. (2021) reported that in Mediterranean forests dominated by P. sylvestris, P. pinaster, and Quercus ilex, the production and turnover of EcM mycelium were strongly influenced by drought intensity, with lower mycelial output observed under dry conditions. These field observations underscore that the functional capacity of both partners in the symbiosis can be impaired under climate stress, affecting nutrient dynamics, carbon exchange, and ultimately forest ecosystem resilience. Our findings reinforce the broader understanding that the ability of EcM fungi to tolerate environmental stress—such as drought—plays a pivotal role not only in their own survival but also in shaping plant physiological responses. A recent review highlights the stress tolerance mechanisms of EcM fungi directly influence the outcome of the symbiosis, with cascading effects on forest ecosystem functioning and resilience (Branco et al., 2022). In the face of accelerating climate change, accounting for the dual responses of both host plants and their fungal symbionts is thus essential for predicting ecosystem stability and guiding conservation strategies.
It is also important to acknowledge that the semi-hydroponic cultivation system used in these experiments simplifies the complex interactions occurring in natural soil environments and may not fully replicate field conditions. However, our findings align well with those from several field studies, showing a positive role of EcM fungi in supporting plant nutrient uptake and physiological performance under water-limited conditions (e.g., Querejeta et al., 2009; Fernandez et al., 2023). They also document reduced ectomycorrhizal colonization and function under prolonged drought stress (e.g., Bell and Adams, 2004; León-Sánchez et al., 2018; Gehring et al., 2020; Hagenbo et al., 2021). These parallels suggest that, despite the controlled nature of our experimental system, the mechanisms we observed are ecologically relevant and reinforce broader patterns documented in natural ecosystems.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
PM: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. IL: Formal analysis, Investigation, Supervision, Validation, Writing – review & editing. RV: Data curation, Formal analysis, Writing – review & editing. JC: Data curation, Formal analysis, Writing – review & editing. QP: Conceptualization, Formal analysis, Funding acquisition, Methodology, Project administration, Supervision, Writing – review & editing. SD: Conceptualization, Formal analysis, Funding acquisition, Methodology, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was financially supported by the Belgian Science Policy Office (BELSPO) – Belgium, in the frame of the BiodivERsA research program, under the Horizon 2020 ERA-NET COFUND scheme (project MixForChange).
Acknowledgments
We thank the Support en Méthodologie et Calcul Statistique (SMCS) of UCLouvain team, particularly Dr. Catherine Rasse, for the statistical advice and consultation. We thank the MOCA (Mineral & Organic Chemical Analysis) platform of the Earth & Life Institute of UCLouvain for the chemical analyses. We thank Mevrouw Nasrin Esteki for her help in the set-up of the experiment. We would also like to thank Joannès Guillemot and Nicolas Martin-StPaul for their advices and for sharing their knowledge.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1574237/full#supplementary-material
References
Allen, C. D. (2007). Mycorrhizal fungi: highways for water and nutrients in arid soils. Vadose Zone J. 6, 291–297. doi: 10.2136/vzj2006.0068
Allen, C. D., Macalady, A. K., Chenchouni, H., Bachelet, D., McDowell, N., Vennetier, M., et al. (2010). A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For. Ecol. Manag. 259, 660–684. doi: 10.1016/j.foreco.2009.09.001
Anderegg, W. R. L., Hicke, J. A., Fisher, R. A., Allen, C. D., Aukema, B., Bentz, B., et al. (2015). Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 208, 674–683. doi: 10.1111/nph.13477
Bacelar, E. L. V. A., Moutinho-Pereira, J. M., Gonçalves, B. M. C., Brito, C. V. Q., Gomes-Laranjo, J., Ferreira, H. M. F., et al. (2012). “Water use strategies of plants under drought conditions” in Plant responses to drought stress: From morphological to molecular features. ed. R. Aroca (Berlin, Heidelberg: Springer), 145–170.
Bell, T. L., and Adams, M. A. (2004). Ecophysiology of ectomycorrhizal fungi associated with Pinus spp. in low rainfall areas of Western Australia. Plant Ecol. 171, 35–52. doi: 10.1023/B:VEGE.0000029375.05683.40
Bogeat-Triboulot, M.-B., Bartoli, F., Garbaye, J., Marmeisse, R., and Tagu, D. (2004). Fungal ectomycorrhizal community and drought affect root hydraulic properties and soil adherence to roots of Pinus pinaster seedlings. Plant Soil 267, 213–223. doi: 10.1007/s11104-005-5349-7
Bose, A. K., Gessler, A., Bolte, A., Bottero, A., Buras, A., Cailleret, M., et al. (2020). Growth and resilience responses of scots pine to extreme droughts across Europe depend on predrought growth conditions. Glob. Chang. Biol. 26, 4521–4537. doi: 10.1111/gcb.15153
Branco, S., Schauster, A., Liao, H.-L., and Ruytinx, J. (2022). Mechanisms of stress tolerance and their effects on the ecology and evolution of mycorrhizal fungi. New Phytol. 235, 2158–2175. doi: 10.1111/nph.18308
Bruce, T. J. A., Matthes, M. C., Napier, J. A., and Pickett, J. A. (2007). Stressful “memories” of plants: evidence and possible mechanisms. Plant Sci. 173, 603–608. doi: 10.1016/j.plantsci.2007.09.002
Brundrett, M. C. (2009). Mycorrhizal associations and other means of nutrition of vascular plants: understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 320, 37–77. doi: 10.1007/s11104-008-9877-9
Brundrett, M., Bougher, N., Dell, B., Grove, T., and Malajczuk, N. (1996). Working with mycorrhizas in forestry and agriculture ACIAR Monograph 32. Canberra: Australian Centre for International Agricultural Research.
Cartabia, A., Tsiokanos, E., Tsafantakis, N., Lalaymia, I., Termentzi, A., Miguel, M., et al. (2021). The arbuscular mycorrhizal fungus Rhizophagus irregularis MUCL 41833 modulates metabolites production of Anchusa officinalis L. under semi-hydroponic cultivation. Front. Plant Sci. 12:352. doi: 10.3389/fpls.2021.724352
Carter, K. R., and Dickman, L. T. (2022). Recovery of seedling carbon balance despite hydraulic impairment following hot drought. Tree Physiol. 42, 1527–1531. doi: 10.1093/treephys/tpac045
Castaño, C., Suarez-Vidal, E., Zas, R., Bonet, J. A., Oliva, J., and Sampedro, L. (2023). Ectomycorrhizal fungi with hydrophobic mycelia and rhizomorphs dominate in young pine trees surviving experimental drought stress. Soil Biol. Biochem. 178:108932. doi: 10.1016/j.soilbio.2022.108932
Coleman, M. D., Bledsoe, C. S., and Lopushinsky, W. (1989). Pure culture response of ectomycorrhizal fungi to imposed water stress. Can. J. Bot. 67, 29–39. doi: 10.1139/b89-005
Colpaert, J. V., van Laere, A., and van Assche, J. A. (1996). Carbon and nitrogen allocation in ectomycorrhizal and non-mycorrhizal Pinus sylvestris L. seedlings. Tree Physiol. 16, 787–793. doi: 10.1093/treephys/16.9.787
Colpaert, J. V., Van Tichelen, K. K., Van Assche, J. A., and Van Laere, A. (1999). Short-term phosphorus uptake rates in mycorrhizal and non-mycorrhizal roots of intact Pinus sylvestris seedlings. New Phytol. 143, 589–597. doi: 10.1046/j.1469-8137.1999.00471.x
Cregg, B. M., and Zhang, J. W. (2001). Physiology and morphology of Pinus sylvestris seedlings from diverse sources under cyclic drought stress. For. Ecol. Manag. 154, 131–139. doi: 10.1016/S0378-1127(00)00626-5
Danielsen, L., and Polle, A. (2014). Poplar nutrition under drought as affected by ectomycorrhizal colonization. Environ. Exp. Bot. 108, 89–98. doi: 10.1016/j.envexpbot.2014.01.006
Davies, F. T. Jr., Potter, J. R., and Linderman, R. G. (1996). Non-nutritional stress acclimation of mycorrhizal woody plants exposed to drought. Tree Physiol. 16, 985–993. doi: 10.1093/treephys/16.11-12.985
De Quesada, G., Xu, J., Salmon, Y., Lintunen, A., Poque, S., Himanen, K., et al. (2024). The effect of ectomycorrhizal fungal exposure on nursery-raised Pinus sylvestris seedlings: plant transpiration under short-term drought, root morphology and plant biomass. Tree Physiol. 44:29. doi: 10.1093/treephys/tpae029
Dosskey, M. G., Linderman, R. G., and Boersma, L. (1991). Role for the photosynthate demand of ectomycorrhizas in the response of Douglas fir seedlings to drying soil. New Phytol. 117, 327–334. doi: 10.1111/j.1469-8137.1991.tb00001.x
Erlandson, S. R., Koziol, L., Bever, J. D., Lekberg, Y., and Hart, M. M. (2022). Transcriptional acclimation and spatial differentiation characterize drought response by the ectomycorrhizal fungus Suillus pungens. New Phytol. 234, 1910–1913. doi: 10.1111/nph.18052
European Environment Agency (EEA). (2022) Medium resolution vegetation phenology and productivity. European Environment Agency. Available online at: https://www.eea.europa.eu/en/datahub/datahubitem-view/fc62ff38-d77b-452e-9878-acdd14f91b3b?activeAccordion=1083802. (Accessed October 10, 2022).
Fernandez, C. W., Bhatnagar, J. M., Koide, R. T., and Kennedy, P. G. (2023). Climate change–induced stress disrupts ectomycorrhizal interaction networks at the boreal–temperate ecotone. Proc. Natl. Acad. Sci. USA 120:e2221619120. doi: 10.1073/pnas.2221619120
Freschet, G. T., Violle, C., Bourget, M. Y., Scherer-Lorenzen, M., and Fort, F. (2018). Allocation, morphology, physiology, architecture: the multiple facets of plant above- and below-ground responses to resource stress. New Phytol. 219, 1338–1352. doi: 10.1111/nph.15225
Garbaye, J., and Churin, J. L. (1997). Growth stimulation of young oak plantations inoculated with the ectomycorrhizal fungus Paxillus involutus with special reference to summer drought. For. Ecol. Manag. 98, 221–228. doi: 10.1016/S0378-1127(97)00105-9
Garcés-Ruiz, M., Calonne-Salmon, M., Plouznikoff, K., Misson, C., Navarrete-Mier, M., Cranenbrouck, S., et al. (2017). Dynamics of short-term phosphorus uptake by intact mycorrhizal and non-mycorrhizal maize plants grown in a circulatory semi-hydroponic cultivation system. Front. Plant Sci. 8:1471. doi: 10.3389/fpls.2017.01471
Gehring, C. A., Sthultz, C. M., Flores-Rentería, L., Whipple, A. V., and Whitham, T. G. (2020). Ectomycorrhizal and dark septate fungal associations of pinyon pine are differentially affected by experimental drought and warming. Front. Plant Sci. 11:582574. doi: 10.3389/fpls.2020.582574
Gehring, C. A., Wolf, J. E., and Theimer, T. C. (2006). Environmental and genetic effects on the formation of ectomycorrhizal and arbuscular mycorrhizal associations in cottonwoods. Oecologia 149, 158–164. doi: 10.1007/s00442-006-0435-5
Gitelson, A. A., Buschmann, C., and Lichtenthaler, H. K. (1999). The chlorophyll fluorescence ratio F735/F700 as an accurate measure of the chlorophyll content in plants. Remote Sens. Environ. 69, 296–302. doi: 10.1016/S0034-4257(99)00023-1
Greenwood, S., Ruiz-Benito, P., Martínez-Vilalta, J., Lloret, F., Kitzberger, T., Allen, C. D., et al. (2017). Tree mortality across biomes is promoted by drought intensity, lower wood density and higher specific leaf area. Ecol. Lett. 20, 539–553. doi: 10.1111/ele.12748
Hagenbo, A., Lindahl, B. D., and Fransson, P. (2021). Production and turnover of mycorrhizal soil mycelium relate to variation in drought conditions in Mediterranean Pinus pinaster, Pinus sylvestris and Quercus ilex forests. New Phytol. 230, 1609–1622. doi: 10.1111/nph.17237
Hahn, C. (1996). Studies in the genus Paxillus I: Paxillus gymnopus – a new Paxillus from the Pacific rainforests of Colombia. Zeitschrift für Mykologie. 62, 43–60.
Harris, C. J., Amtmann, A., and Ton, J. (2023). Epigenetic processes in plant stress priming: open questions and new approaches. Curr. Opin. Plant Biol. 75:102432. doi: 10.1016/j.pbi.2023.102432
Himanen, K., Nygren, M., and Pennanen, T. (2024). Mycelial inoculation of containerized Norway spruce seedlings with ectomycorrhizal fungi. New For. 55, 47–61. doi: 10.1007/s11056-023-09964-y
Ingestad, T., and Kähr, M. (1985). Nutrition and growth of coniferous seedlings at varied relative nitrogen addition rate. Physiol. Plant. 65, 109–116. doi: 10.1111/j.1399-3054.1985.tb02368.x
IPCC (2023). Climate change 2021: The physical science basis. Contribution of working group I to the sixth assessment report of the intergovernmental panel on climate change. Cambridge: Cambridge University Press.
Irvine, J., Perks, M. P., Magnani, F., and Grace, J. (1998). The response of Pinus sylvestris to drought: stomatal control of transpiration and hydraulic conductance. Tree Physiol. 18, 393–402. doi: 10.1093/treephys/18.6.393
Jacques, C., Salon, C., Barnard, R. L., Vernoud, V., and Prudent, M. (2021). Drought stress memory at the plant cycle level: a review. Plants 10:1873. doi: 10.3390/plants10091873
Kebert, M., Kostić, S., Stojnić, S., Čapelja, E., Gavranović Markić, A., Zorić, M., et al. (2023). A fine-tuning of the plant hormones, polyamines and osmolytes by ectomycorrhizal fungi enhances drought tolerance in pedunculate oak. Int. J. Mol. Sci. 24:7510. doi: 10.3390/ijms24087510
Kennedy, P. G., and Peay, K. G. (2007). Different soil moisture conditions change the outcome of the ectomycorrhizal symbiosis between Rhizopogon species and Pinus muricata. Plant Soil 291, 155–165. doi: 10.1007/s11104-006-9182-1
Kipfer, T., Wohlgemuth, T., van der Heijden, M. G. A., Ghazoul, J., and Egli, S. (2012). Growth response of drought-stressed Pinus sylvestris seedlings to single- and multi-species inoculation with ectomycorrhizal fungi. PLoS One 7:e35275. doi: 10.1371/journal.pone.0035275
Kirkham, M. B. (2023). Principles of soil and plant water relations. 3rd Edn. London: Academic Press.
Koele, N., and Hildebrand, E. E. (2011). Differences in growth and nutrient uptake from a coarse-soil substrate by ectomycorrhizal- and fungicide-treated Picea abies seedlings. Eur. J. Forest Res. 130, 317–324. doi: 10.1007/s10342-010-0435-7
Krakau, U.-K., Liesebach, M., Aronen, T., Lelu-Walter, M.-A., and Schneck, V. (2013). “Scots pine (Pinus sylvestris L.)” in Forest tree breeding in Europe: Current state-of-the-art and perspectives. ed. L. E. Pâques (Dordrecht: Springer Netherlands), 267–323.
Lamhamedi, M. S., Godbout, C., and Fortin, J. A. (1994). Dependence of Laccaria bicolor basidiome development on current photosynthesis of Pinus strobus seedlings. Can. J. For. Res. 24, 1797–1804. doi: 10.1139/x94-232
Langley, J. A., Chapman, S. K., and Hungate, B. A. (2006). Ectomycorrhizal colonization slows root decomposition: the post-mortem fungal legacy. Ecology Letters. 9, 955–959. doi: 10.1111/j.1461-0248.2006.00948.x
Lehto, T., and Zwiazek, J. J. (2011). Ectomycorrhizas and water relations of trees: a review. Mycorrhiza 21, 71–90. doi: 10.1007/s00572-010-0348-9
León-Sánchez, L., Nicolás, E., Querejeta, J. I., Prieto, I., and Arrieta, A. (2018). Poor plant performance under simulated climate change is linked to mycorrhizal responses in a semi-arid shrubland. J. Ecol. 106, 960–976. doi: 10.1111/1365-2745.12977
Li, M., Wang, H., Zhao, X., Feng, W., Ding, G., and Quan, W. (2023). Effect of ectomycorrhizal fungi on the drought resistance of Pinus massoniana seedlings. J. Fungi 9:471. doi: 10.3390/jof9040471
Lim, C. S., Kim, K. W., Lee, S. B., Ryu, J., Seo, J., and Lee, S. J. (2021). Estimation of water retention characteristics depending on the particle sizes of perlite using van Genuchten equation with retention curve program. Korean J. Soil Sci. Fertilizer 54, 276–288. doi: 10.7745/KJSSF.2021.54.3.276
Liu, C., Wu, J., Gu, J., and Duan, H. (2024). Growth and C: N: P stoichiometry of Pinus yunnanensis seedlings in response to drought and rewatering. Forests 15:2175. doi: 10.3390/f15122175
Lloret, F., Peñuelas, J., Prieto, P., Llorens, L., and Estiarte, M. (2009). Plant community changes induced by experimental climate change: seedling and adult species composition. Perspect. Plant Ecol. Evol. Syst. 11, 53–63. doi: 10.1016/j.ppees.2008.09.001
Luo, Z. B., Janz, D., Jiang, X., Göbel, C., Wildhagen, H., Tan, Y., et al. (2009). Upgrading root physiology for stress tolerance by ectomycorrhizas: insights from metabolite and transcriptional profiling into reprogramming for stress anticipation. Plant Physiol. 151, 1902–1917. doi: 10.1104/pp.109.145706
Martínez-Vilalta, J., Cochard, H., Mencuccini, M., Sterck, F., Herrero, A., Korhonen, J. F. J., et al. (2009). Hydraulic adjustment of scots pine across Europe. New Phytol. 184, 353–364. doi: 10.1111/j.1469-8137.2009.02954.x
Martín-Gómez, P., Aguilera, M., Pemán, J., Gil-Pelegrín, E., and Ferrio, J. P. (2017). Contrasting ecophysiological strategies related to drought: the case of a mixed stand of scots pine (Pinus sylvestris) and a submediterranean oak (Quercus subpyrenaica). Tree Physiol. 37, 1478–1492. doi: 10.1093/treephys/tpx101
Meng, F., Zhang, T., and Yin, D. (2023). The effects of soil drought stress on growth characteristics, root system, and tissue anatomy of Pinus sylvestris var. mongolica. PeerJ 11:e14578. doi: 10.7717/peerj.14578
Molina, R., and Trappe, J. M. (1982). Patterns of ectomycorrhizal host specificity and potential among Pacific northwest conifers and fungi. For. Sci. 28, 423–458. doi: 10.1093/forestscience/28.3.423
Morte, A., Lovisolo, C., and Schubert, A. (2001). Growth and water relations in mycorrhizal and nonmycorrhizal Pinus halepensis plants in response to drought. Biol. Plant. 44, 263–267. doi: 10.1023/A:1010246429255
Moser, B., Kipfer, T., Richter, S., Egli, S., and Wohlgemuth, T. (2015). Drought resistance of Pinus sylvestris seedlings conferred by plastic root architecture rather than ectomycorrhizal colonisation. Ann. For. Sci. 72, 303–309. doi: 10.1007/s13595-014-0380-6
Muhsin, T. M., and Zwiazek, J. J. (2002). Colonization with Hebeloma crustuliniforme increases water conductance and limits shoot sodium uptake in white spruce (Picea glauca) seedlings. Plant Soil 238, 217–225. doi: 10.1023/A:1014435407735
Nehls, U., Göhringer, F., Wittulsky, S., and Dietz, S. (2010). Fungal carbohydrate support in the ectomycorrhizal symbiosis: a review. Plant Biol. 12, 292–301. doi: 10.1111/j.1438-8677.2009.00312.x
Nehls, U., and Plassard, C. (2018). Nitrogen and phosphate metabolism in ectomycorrhizas. New Phytol. 220, 1047–1058. doi: 10.1111/nph.15257
Nelson, D. W., and Sommers, L. E. (1996). Total carbon, organic carbon, and organic matter. Methods of Soil Analysis: Part 3, Chemical Methods, SSSA Book Series No. 5 ed. D. L. Sparks. Madison, WI, USA: Soil Science Society of America, 961–1010.
Nickel, U. T., Weikl, F., Kerner, R., Schäfer, C., Kallenbach, C., Munch, J. C., et al. (2018). Quantitative losses vs. qualitative stability of ectomycorrhizal community responses to 3 years of experimental summer drought in a beech–spruce forest. Glob. Change Biol. 24, e560–e576. doi: 10.1111/gcb.13967
Nylund, J. E., and Wallander, H. (1989). Effects of ectomycorrhiza on host growth and carbon balance in a semi-hydroponic cultivation system. New Phytol. 112, 389–398. doi: 10.1111/j.1469-8137.1989.tb00328.x
Parke, J. L., Linderman, R. G., and Black, C. H. (1983). The role of ectomycorrhizas in drought tolerance of Douglas-fir seedlings. New Phytol. 95, 83–95. doi: 10.1111/j.1469-8137.1983.tb03474.x
Pearson, M., Saarinen, M., Heiskanen, J., Sarjala, T., and Laine, J. (2013). High and dry: consequences of drought exposure in scots pine seedlings grown in authentic peat soil. Tree Physiol. 33, 363–376. doi: 10.1093/treephys/tpt016
Peltier, D. M. P., Fell, M., and Ogle, K. (2016). Legacy effects of drought in the southwestern United States: a multi-species synthesis. Ecol. Monogr. 86, 312–326. doi: 10.1002/ecm.1219
Pohjanen, J., Koskimäki, J. J., Sutela, S., Ardanov, P., Suorsa, M., Niemi, K., et al. (2014). Interaction with ectomycorrhizal fungi and endophytic Methylobacterium affects nutrient uptake and growth of pine seedlings in vitro. Tree Physiol. 34, 993–1005. doi: 10.1093/treephys/tpu062
Poyatos, R., Martínez-Vilalta, J., Čermák, J., Ceulemans, R., Granier, A., Irvine, J., et al. (2007). Plasticity in hydraulic architecture of scots pine across Eurasia. Oecologia 153, 245–259. doi: 10.1007/s00442-007-0740-0
Pyhäjärvi, T., Garciá-Gil, M. R., Knürr, T., Mikkonen, M., Wachowiak, W., and Savolainen, O. (2007). Demographic history has influenced nucleotide diversity in European Pinus sylvestris populations. Genetics 177, 1713–1724. doi: 10.1534/genetics.107.077099
Querejeta, J. I., Egerton-Warburton, L. M., and Allen, M. F. (2009). Topographic position modulates the mycorrhizal response of oak trees to interannual rainfall variability. Ecology 90, 649–662. doi: 10.1890/08-0644.1
Sadhukhan, A., Prasad, S. S., Mitra, J., Siddiqui, N., Sahoo, L., Kobayashi, Y., et al. (2022). How do plants remember drought? Planta 256:7. doi: 10.1007/s00425-022-03924-0
Scholander, P. F., Hemmingsen, E., and Garey, W. (1965). Sap pressure in vascular plants. Science 148, 339–346. doi: 10.1126/science.148.3668.339
Seidel, H., and Menzel, A. (2016). Above-ground dimensions and acclimation explain variation in drought mortality of scots pine seedlings from various provenances. Front. Plant Sci. 7:1014. doi: 10.3389/fpls.2016.01014
Smith, S. E., and Read, D. J. (1997). Mycorrhizal Symbiosis. 2nd ed. San Diego: Academic Press, ix + 605.
Swaty, R. L., Gehring, C. A., Van Ert, M., Theimer, T. C., Keim, P., and Whitham, T. G. (2004). Ectomycorrhizal abundance and community composition shifts with drought: predictions from tree rings. Ecology 85, 1072–1084. doi: 10.1890/02-0753
Vacek, Z., Linda, R., Cukor, J., Vacek, S., Šimůnek, V., Gallo, J., et al. (2021). Scots pine (Pinus sylvestris L.), the suitable pioneer species for afforestation of reclamation sites? For. Ecol. Manag. 485:118951. doi: 10.1016/j.foreco.2021.118951
Wallander, H., and Söderström, B. (1999). “Paxillus” in Ectomycorrhizal fungi key genera in profile. eds. J. W. G. Cairney and S. M. Chambers (Berlin, Heidelberg: Springer), 231–252.
Wang, J., Zhang, H., Gao, J., Zhang, Y., Liu, Y., and Tang, M. (2021). Effects of ectomycorrhizal fungi (Suillus variegatus) on the growth, hydraulic function, and non-structural carbohydrates of Pinus tabulaeformis under drought stress. BMC Plant Biol. 21:171. doi: 10.1186/s12870-021-02945-3
Yin, J., and Bauerle, T. L. (2017). A global analysis of plant recovery performance from water stress. Oikos 126, 1377–1388. doi: 10.1111/oik.04534
Yin, D., Song, R., Qi, J., and Deng, X. (2018). Ectomycorrhizal fungus enhances drought tolerance of Pinus sylvestris var. mongolica seedlings and improves soil condition. J. For. Res. 29, 1775–1788. doi: 10.1007/s11676-017-0583-4
Keywords: ectomycorrhizal fungi, drought, seedlings, recovery, forests, climate change, drought resistance, Scots pine
Citation: Musella P, Lalaymia I, Verdonck R, Colpaert J, Ponette Q and Declerck S (2025) Paxillus involutus enhances drought resistance of Pinus sylvestris seedlings by improving photosynthetic rate and water use efficiency. Front. For. Glob. Change. 8:1574237. doi: 10.3389/ffgc.2025.1574237
Edited by:
Aaron S. Weed, National Park Service, United StatesReviewed by:
José Ignacio Querejeta, Spanish National Research Council (CSIC), SpainTingting Mei, Zhejiang A&F University, China
Copyright © 2025 Musella, Lalaymia, Verdonck, Colpaert, Ponette and Declerck. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stéphane Declerck, U3RlcGhhbi5kZWNsZXJja0B1Y2xvdXZhaW4uYmU=