 Norbert Móricz1*
Norbert Móricz1* Ilona Mészáros2
Ilona Mészáros2 Zoltán Kern3Gábor Zoltán Illés1Balázs Garamszegi4
Zoltán Kern3Gábor Zoltán Illés1Balázs Garamszegi4 Csaba Béla Eötvös1Imre Berki5
Csaba Béla Eötvös1Imre Berki5 Tamás Márton Németh1
Tamás Márton Németh1- 1Department of Ecology and Forest Management, Forest Research Institute, University of Sopron, Sopron, Hungary
- 2Department of Botany, Faculty of Science and Technology, University of Debrecen, Debrecen, Hungary
- 3Institute for Geological and Geochemical Research, HUN-REN Research Centre for Astronomy and Earth Sciences, Budapest, Hungary
- 4Department of Ecosystem Management, Climate and Biodiversity, Institute of Forest Ecology, BOKU University, Vienna, Austria
- 5Independent Researcher, Sopron, Hungary
Climate change is placing stress on forests, making silvicultural planning more challenging in many temperate regions. Since some major tree species are prone to drought, there is an increasing interest in selecting native broadleaf minor species, which are believed to have greater drought tolerance. Therefore, it is crucial to evaluate the growth performance and vitality of these tree species in the light of the changing climate. Based on tree-ring width data, we analysed the drought response of four minor tree species Acer campestre L., Fraxinus ornus L., Quercus pubescens Willd. and Tilia tomentosa Moench. in relation with a major tree species Quercus cerris L. in different mixed stands along a precipitation gradient in Hungary with the aim to compare the species’ drought tolerance. All the species analysed showed a high drought tolerance, with only minor differences among them. A. campestre, F. ornus and T. tomentosa showed an overall greater climate sensitivity than the oak species, but drought resilience was high in all species. Furthermore, none of the species showed signs of dieback on the study sites with recent climate warming, suggesting that all studied native tree species are also well suited as admixed species in future forest stands.
1 Introduction
Droughts could trigger a long-term decrease in forest productivity (Peltier et al., 2016; Camarero et al., 2018; D’Orangeville et al., 2018; Mátyás et al., 2018), reduce carbon sequestration or, ultimately lead to forest decline and mortality (Allen et al., 2015; Móricz et al., 2018; Schuldt et al., 2020; Senf et al., 2020; Hammond et al., 2022). A decrease in tree growth due to prolonged drought periods might diminish the future economic value of currently productive forests (Hanewinkel et al., 2012), which also requires the study also of tree physiological adaptations (Andreu-Hayles et al., 2011; Gagen et al., 2011).
In Central Europe, over the past three decades, the frequency and severity of droughts have increased (Spinoni et al., 2017; Ionita and Nagavciuc, 2021). These changes pose significant challenges to silvicultural concepts. Various forest management strategies have been proposed to alleviate drought stress, including reducing stand density (Schmitt et al., 2020; Steckel et al., 2020), replacing monocultures with mixtures (Pretzsch et al., 2013; Pretzsch et al., 2020), transferring propagating material from drought-adapted populations (Mátyás, 2021) and introducing more drought-tolerant native or non-native temperate tree species (Zimmermann et al., 2015; Kunz et al., 2018; Latte et al., 2020; Fuchs et al., 2021b; Schmucker et al., 2023; Bouwman et al., 2025). Among these strategies, species mixing and stand thinning are favored silvicultural practices to address challenges posed by climate change, particularly in maintaining stable forest cover in drought-prone regions. However, the choice of suitable drought-tolerant native tree species which are currently secondary tree species, seems more convincing and gaining growing interest. Native trees are generally preferable to non-native species because they present fewer uncertainties regarding pests, wildlife interactions, wood utilization, and their overall impact on ecosystem services (Castro-Díez et al., 2019; Sapsford et al., 2020; Matevski and Schuldt, 2021). Therefore, it is crucial to evaluate the potential of minor domestic tree species by examining their general growth patterns, economic viability, and drought tolerance even though these tree species are of lower economic importance. Nevertheless, only a limited number of dendroecological studies have included minor tree species from older age classes that are assumed to be more drought-tolerant in Central Europe (Hemery et al., 2010; de Jaegere et al., 2016; Kunz et al., 2018; Leuschner et al., 2019; Kasper et al., 2022; Schmucker et al., 2023; Leuschner et al., 2024). In Hungary, tree species from the genera Acer, Fraxinus and Tilia are commonly found as admixed species in forest communities. Their geographical distribution extends into the more continental climate of eastern Central Europe, suggesting that they may be more drought-tolerant than the major native tree species (de Rigo et al., 2016). Among native species, field maple (Acer campestre L.), manna ash (Fraxinus ornus L.), pubescent oak (Quercus pubescens Willd.) and silver lime (Tilia tomentosa Moench.) seem to be promising options for admixture in Central European forests.
The diffuse-porous species A. campestre has a broad ecological range, covering most of Europe, though it is most commonly found in mesophile stands, especially in deciduous oak forests (Zecchin et al., 2016). It has moderate water demand and avoids waterlogging, preferring calcareous soils (Nagy and Ducci, 2004). A combined analysis of species distribution models and local growth responses revealed that A. campestre displayed a relatively high drought tolerance (Walentowski et al., 2014). An extensive study of the drought tolerance of minor tree species in southwest Germany suggested that A. campestre exhibits similar drought tolerance to that of sessile oak (Kunz et al., 2018). F. ornus has the smallest distribution range among the ashes, primarily found from southeast France through Central Europe (Hungary) and the Balkan peninsula to western Turkey (Caudullo and de Rigo, 2016). This ring-porous species requires a minimum mean annual precipitation of at least 500 mm but demonstrates good drought resistance due to ability to store water in its densely branched roots and its low transpiration rate (Italiano et al., 2024). The ring-porous species Q. pubescens has a wide distribution, covering nearly all of central and southern Europe (Pasta et al., 2016) and able to survive in drought-prone environments (Damesin and Rambal, 1995; Nardini and Pitt, 1999; Pasta et al., 2016). It is known for its conservative water usage, largely due to to the anatomy of its conducting tissues (Eilmann et al., 2006; Vodnik et al., 2019). T. tomentosa is a fast-growing tree species with diffuse-porous tree-rings, predominantly found in the Balkans and Hungary. It behaves more like a water-spending and drought-avoiding tree species with high leaf tissue elasticity and the ability to tolerate temporary leaf wilting (Eaton et al., 2016; Leuschner et al., 2019). However, research indicates that T. tomentosa shows relatively high drought sensitivity in western Romania, showing negative growth trends and lower resilience to drought conditions compared to oak species (Kasper et al., 2022). The sites chosen in Hungary for A. campestre are located within its main distribution area in Europe. In contrast, the sample areas for the other tree species included in this study are situated near the northern limit of their distribution.
To enhance the applicability of our findings concerning minor tree species for forest managers and scientists, we compare them with Quercus cerris. As this oak species is a dominant tree species with economic importance in most Southeast European countries, and its growth and drought reactions are already well studied (Ciceu et al., 2020; Móricz et al., 2021; Mészáros et al., 2022). Generally, Q. cerris exhibits a high drought tolerance and is expected to maintain stable growth even under drier and hotter conditions (Mazza et al., 2021; Kasper et al., 2022).
The main goal of the study is to retrospectively evaluate the growth response of four minor tree species - A. campestre, F. ornus, Q. pubescens and T. tomentosa - compared to the major tree species Q. cerris, particularly in the context of changing climatic conditions in drought-prone forest areas of Hungary. Our research aims to address the following questions: (1) Which climate variables most significantly influence the radial growth of the species analysed? (2) Is there a variation in drought tolerance among the different tree species?
2 Data and methods
2.1 Study sites and tree species
Five study sites were designated in Hungary (Figure 1, Table 1). These sites exhibit annual precipitation ranging from 562 mm to 705 mm and annual air temperatures between 9.3°C and 10.3°C. The southwestern regions typically have moister climates, while the northeastern areas are characterized by drier conditions. None of the selected sites have access to groundwater and they are situated at elevations between 215 m and 370 m above mean sea level (Table 1). Since 1971, annual precipitation at the sampling sites has not shown a significant trend, however, the drier sites showed greater interannual variability (Figure 2). In contrast, air temperature has exhibited a clearer overall trend and significant interdecadal variability. A notable cooler period occurred during the 1980s at all sites, but since 1990, warmer years have been recorded. Over the study period, all sites have experienced a warming trend, with particularly steep increases in annual mean temperature of 0.05°C to 0.06°C per year in the past 30 years (Figure 2). For this study, we designated one species-rich mixed forest stand at each selected site, featuring varying species composition and age, ranging from 42 to 82 years. Each tree species was represented in at least two of the mixed forest stands (Table 2). The five tree species investigated - Acer campestre L., Fraxinus ornus L., Quercus pubescens Willd., Tilia tomentosa Moench. and Quercus cerris L.—are commonly found in Central European broadleaf mixed forests. They typically occur in communities classified under the phytosociological alliances Carpinion betuli (oak-hornbeam forests), Quercetum petraeae-cerris (sessile-Turkey oak forests), Aceri campestri and tatarici-Quercetum (maple-oak forests) (Leuschner and Ellenberg, 2017). To minimize the impact of forest management practices—such as selective thinning and regeneration cutting - on growth-climate relationships, we selected forest stands with low management intensity over the past few decades. Additionally, to reduce competition, we avoided selecting suppressed trees, as they exhibit larger growth responses to various interventions compared to trees in the upper canopy (Nowacki and Abrams, 1997). The selected trees were primarily grown from seed, with a smaller portion being coppice regeneration.
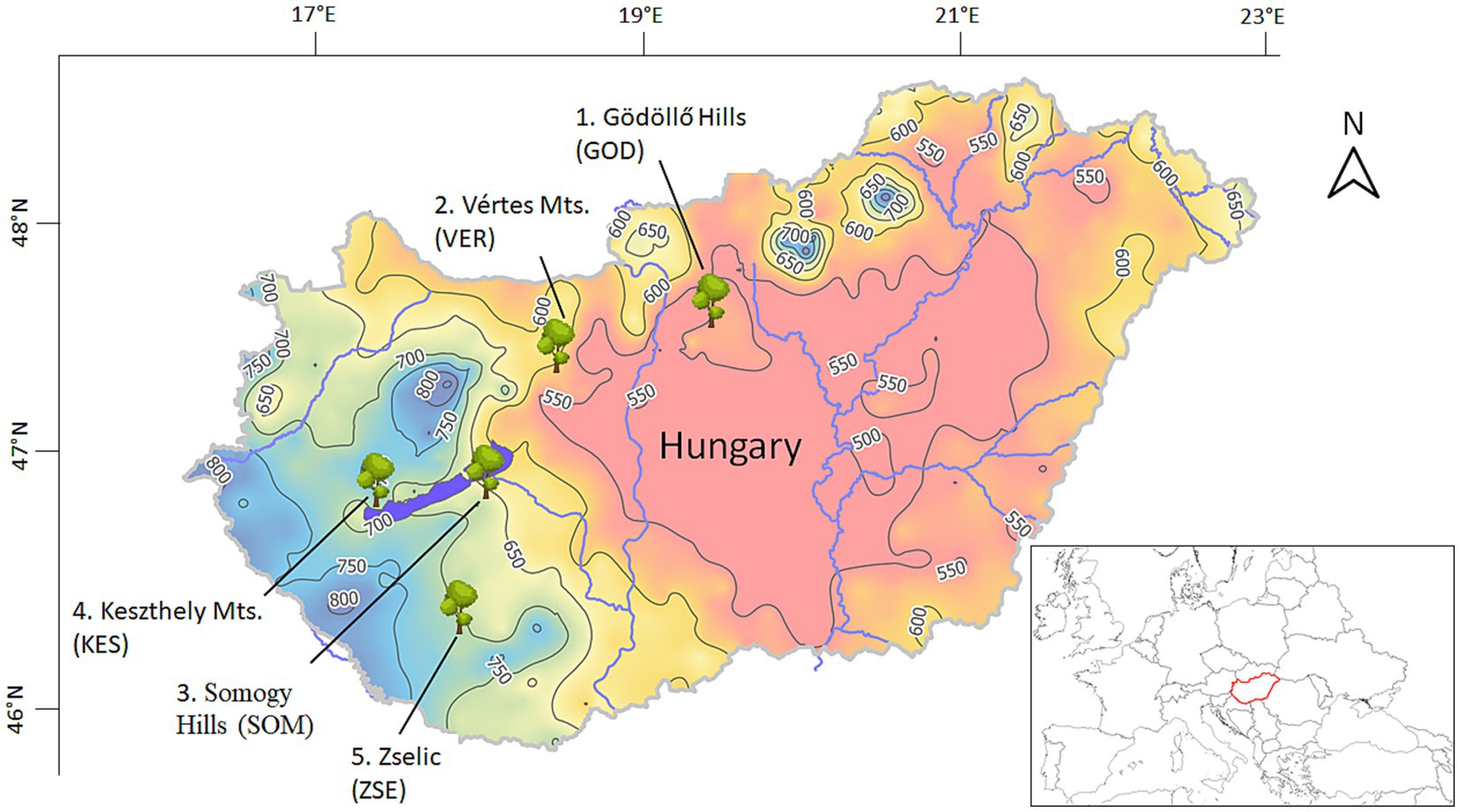
Figure 1. Study sites in Hungary (1. Gödöllő Hills—GOD, 2. Vértes Mts.—VER, 3. Somogy Hills—SOM, 4. Keszthely Mts.—KES, 5. Zselic—ZSE), the color shaded map shows the annual mean precipitation sum (mm) for the period 1971–2022 (HMS, 2022).
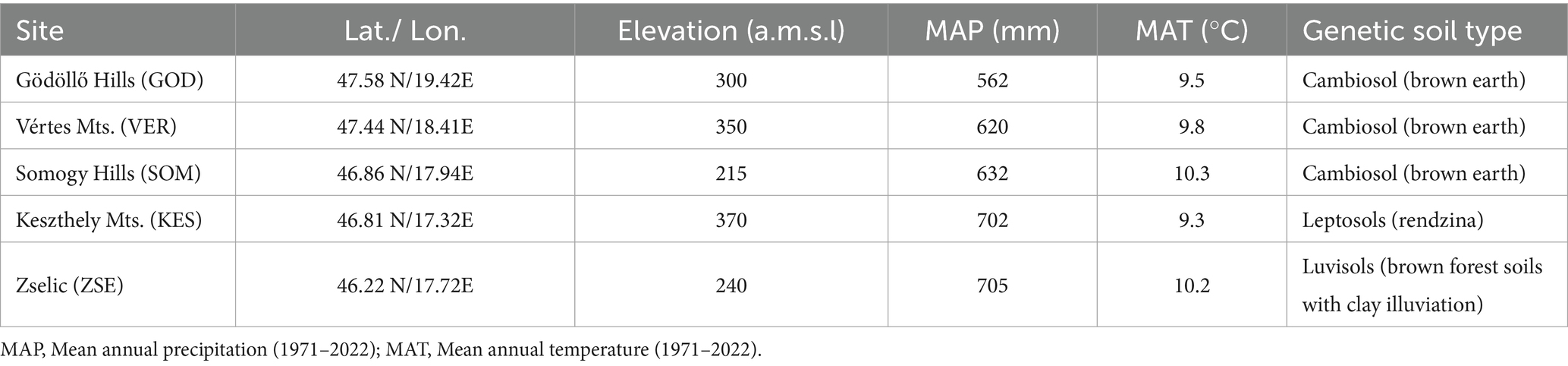
Table 1. Main site characteristics.
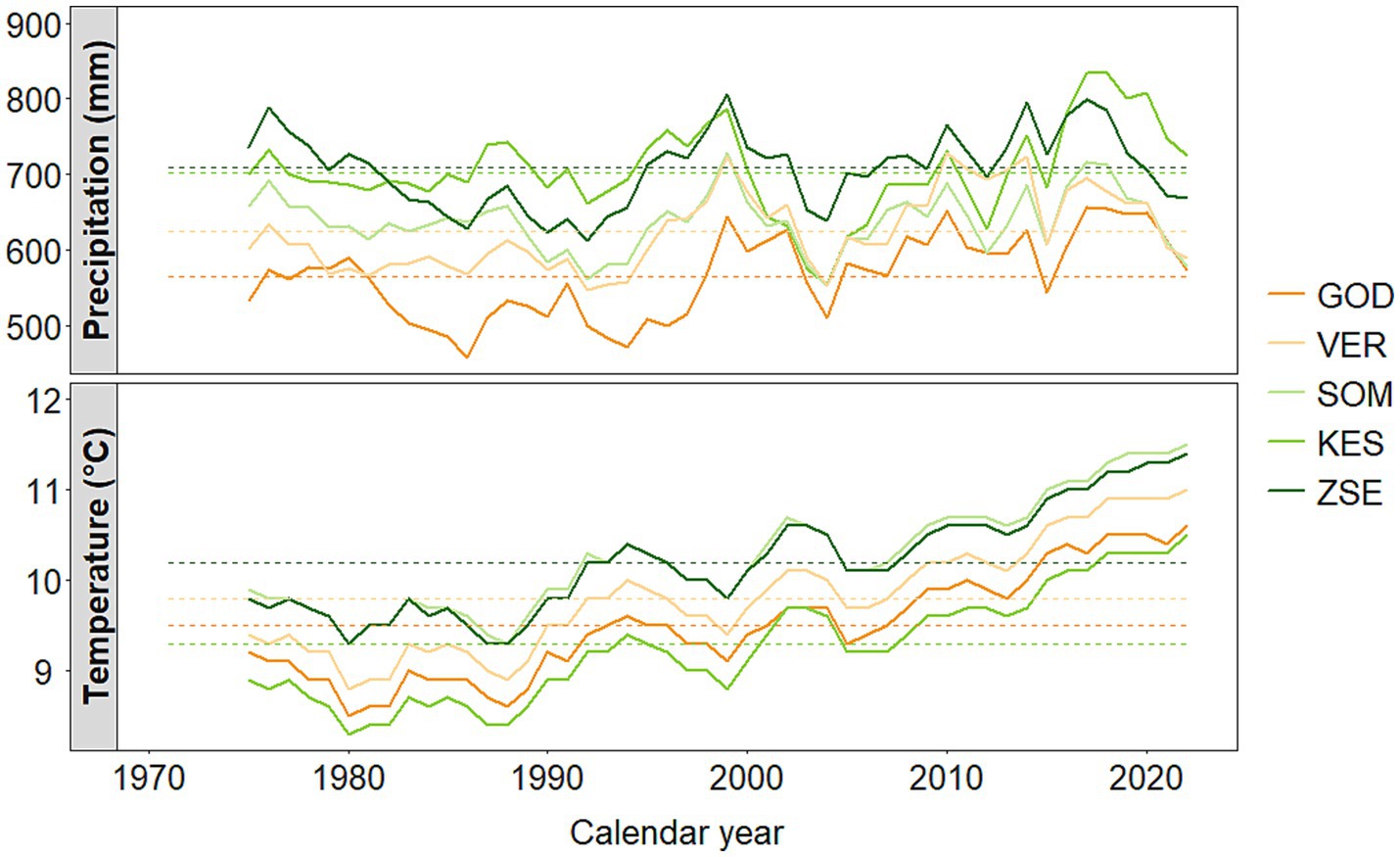
Figure 2. Trends of annual precipitation sum (mm) and annual mean temperature (°C) at the study sites during the period 1971–2022 (HMS, 2022). The dashed horizontal lines represent the average values of the reference period 1971–2000; the curves are smoothed using a 5-year trailing moving average for better visualization of climatic trends. The color code for site labels indicates the gradient of mean annual precipitation (orange: low, green: high).
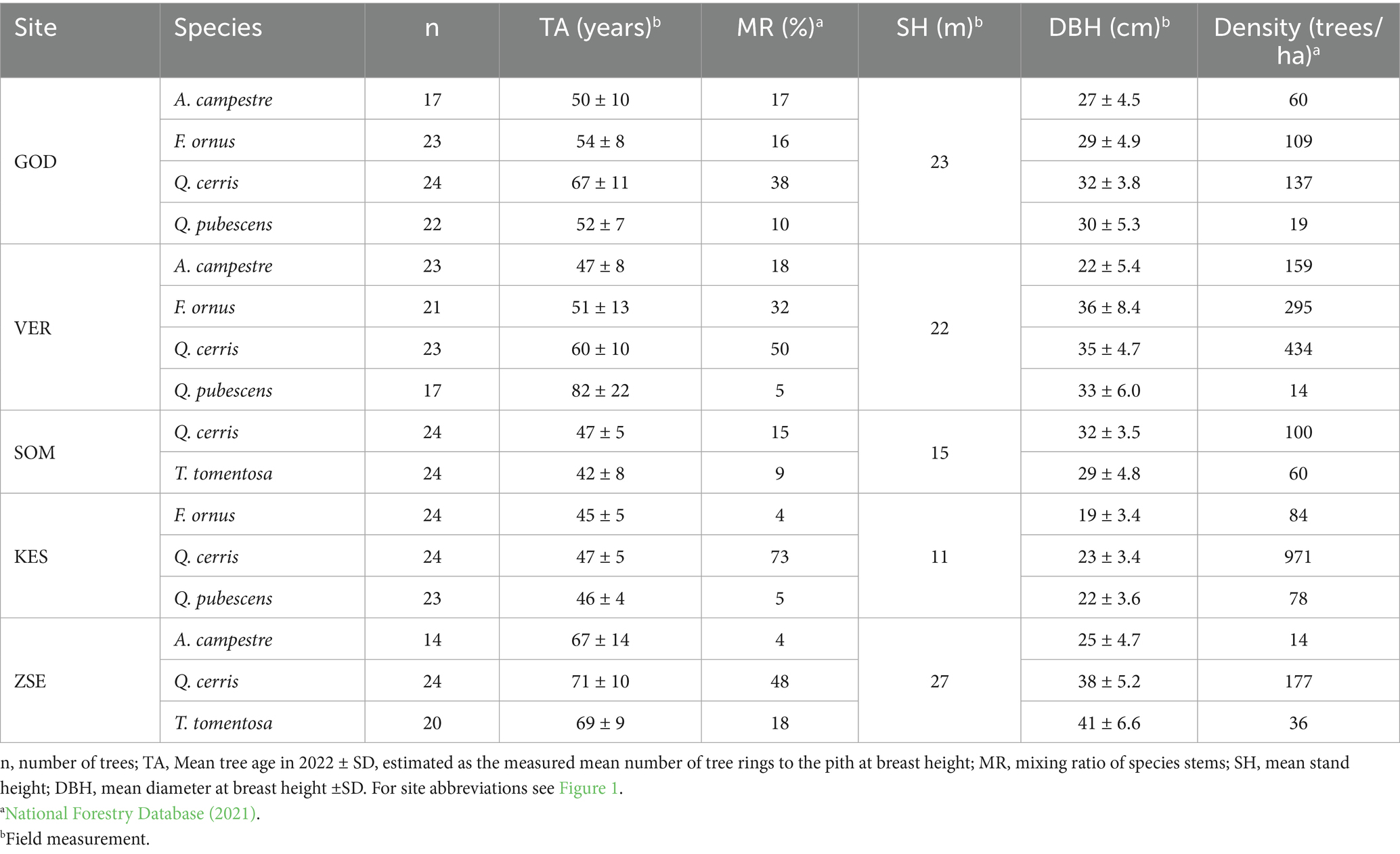
Table 2. Main stand characteristics.
2.2 Meteorological data and soil water budget calculations
Meteorological data were obtained from the HUCLIM daily gridded climate dataset of the Hungarian Meteorological Service (HMS, https://odp.met.hu/). This dataset has an approximate spatial resolution of 10 km and covers the period from 1971 to 2022. We assigned the nearest grid points to the study sites and aggregated daily mean temperatures and total precipitation into monthly averages. To account for the altitude differences between the study sites and the corresponding grid points, we adjusted the mean monthly temperature data using monthly elevation gradients (Péczely, 1979).
The monthly water balance (WB) was calculated as the difference between precipitation and potential evapotranspiration, following the method, described by McCabe and Markstrom (2007). WB serves as an ecologically relevant indicator of water availability for tree growth and has a strong correlation with the radial growth of various tree species (Stojanović et al., 2018; Vitasse et al., 2019). WB effectively reflects the negative impacts of increased temperatures on water availability (Vitasse et al., 2019). We calculated WB for different periods ranging from 1 to 12 months, starting in August of the current year of growth. In addition to WB, we used the Forestry Aridity Index (FAI) (Führer et al., 2011) which is defined as the ratio of the mean temperature of July and August and the precipitation sums of May to July plus the precipitation sum recorded from July to August. This index is particularly relevant for assessing tree growth (Führer et al., 2011).
For monthly soil water balance modelling, we used the Thornthwaite-type model (Thornthwaite, 1948) covering the period from 1971 to 2022. The input variables for the model included monthly mean air temperature, precipitation sum, the latitude of the sites and the plant-available soil-water storage capacity (AWC). To estimate the AWC of the soil, we collected 4–5 soil samples from individual soil horizons down to a depth of 100 cm. These samples were analysed for various chemical and physical parameters (Table 3). Field capacity and wilting point water content of the soil samples were estimated from the particle composition and base rock fraction content using pedo-transfer functions and the Rosetta3 model within the “soilDB” package of R software (Zhang and Schaap, 2017). We summed the differences in water content between field capacity (pF = 2.5) and permanent wilting point (pF = 4.2) over the entire rooting depth, which exceeded 1 meter at all sites, as indicated by visual observations of fine roots in the soil samples. Finally, we calculated the summer water stress index (Is) by dividing soil water deficit and the maximum extractable water for 120 cm soil depth or up to the bedrock depth, assuming uniform soil texture below 100 cm depth, as described by Granier et al. (1999).
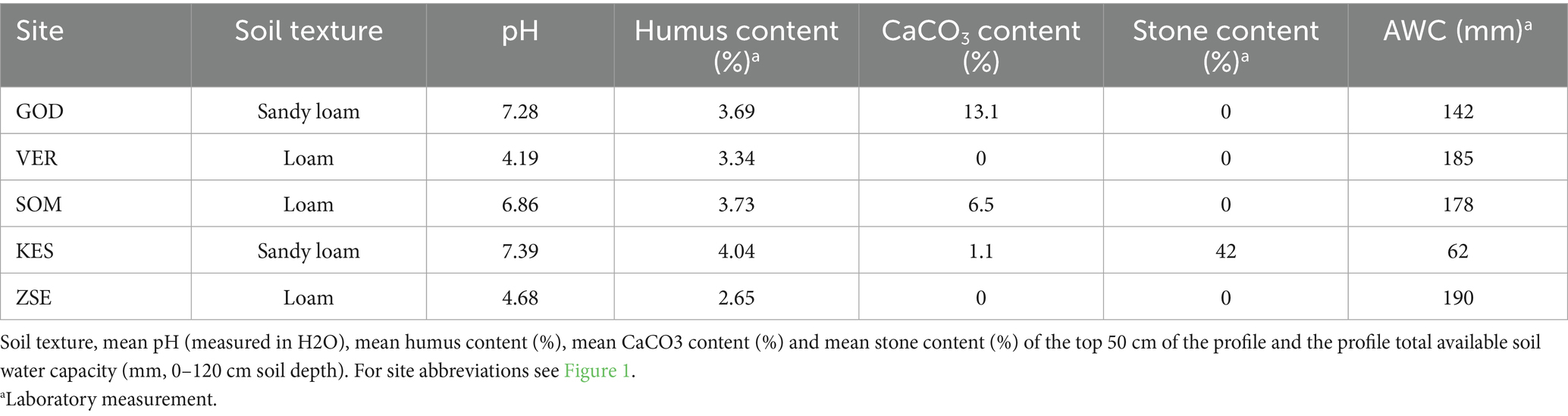
Table 3. Soil characteristics at the studied sites.
2.3 Tree-ring data and chronology building
We selected up to 24 trees of each species based on the available number of trees in each stand and extracted one core at breast height (1.3 m) from each tree (Fritts, 1976). A total of 375 cores were collected, using a Pressler increment borer (Haglöf, Långsele, Sweden), where the influence of tension wood and other anomalies were assumed to be smallest. The cores were then air-dried and glued to grooved wooden mounting boards (Speer, 2010). Afterwards, we sanded and scanned the cores at a resolution of 1,200 dpi (EPSON Expression 11000XL Model: J331A) (Supplementary Figure S1). We measured tree-ring widths (TRW) on the digital images with an accuracy of 0.01 mm using WinDENDRO software ver. 2014a (Regent Instruments Inc., Canada). The resulting TRW series were visually checked for characteristic rings and cross-dated using the software COFECHA with 50 years segments lagged successively by 25 years (Holmes, 1983). Cores that did not meet the default cross-dating correlation threshold were excluded, leaving a total of 346 cores available for further analysis. Tree age was estimated based on the number of tree rings counted from the bark to the pith (Table 2). The number of missing rings to the pith was approximated using the diameter and core length of each tree with the radius-length method (Norton et al., 1987).
To account for age and size-dependent trends, we applied rigorous detrending techniques since the stands varied in age and potential management intensity. As the focus was on single-year climate-growth interactions and short-term responses to drought events, flexible cubic smoothing splines with a 50% frequency cut-off at 25 years were used to detrend and standardize the raw ring-width series (Cook and Peters, 1981; Speer, 2010). We did not remove the first-order autocorrelation as it could significantly affect drought legacies (Yue et al., 2011). Detrended chronologies were built for all populations via Tukey’s biweight robust mean (Mosteller and Tukey, 1977) and truncated to the period of 1972–2021 to exclude younger tree life stages (Supplementary Figure S2). The signal strength of the final chronologies was assessed by the expressed population signal (EPS) and mean inter-series correlation (Rbar).
To assess long-term growth trends, we used the regional curve standardization (RCS) method (Briffa et al., 1983; Biondi and Qeadan, 2008) to age-detrend the ring-width series. We estimated the regional age trend for all investigated species by aligning the tree-ring series for each species based on cambial age. The estimated average growth curve was then used to detrend individual series. After this, we constructed RCS chronologies using Tukey’s bi-weight robust mean. We evaluated the significance of growth trends over the period from 1972 to 2021 using the Mann-Kendall trend test. All detrending procedures and the statistical analyses of the chronologies were performed using the software R (version 4.0, R Core Team, Vienna) with the “dplr” package (Bunn, 2008).
2.4 Climate sensitivity analysis
Large-scale pest outbreaks can significantly impact tree growth and confound the climate sensitivity analysis. Considering the occurrence of such events (Hirka, 2022), we assessed any potential biotic effects on radial growth by utilizing Cook’s distance in the linear regression between detrended growth and water balance (calculated from the previous September to the current August) for each population (Cook and Weisberg, 1982). This approach helped us identify any outlier years in the datasets that aligned with reports of biotic damage from local forest managers.
We assessed the relationship between climate variables (monthly temperature and precipitation, and derived indices FAI, WB, and Is) and standard chronologies using Pearson correlation coefficients. Monthly temperature and precipitation were examined throughthe response-function analysis taking into account the inter-correlations among the climatic variables using the “treeclim” package (Zang and Biondi, 2015). We analysed monthly meteorological data over the preceding 16 months, from June of the previous year until September of the actual year of ring formation. For FAI and Is, the correlation coefficient was computed on an annual basis, while for WB, we considered all seasons (March–May, June–August, September–November and December–February). Additionally, the correlation for WB was calculated for a 3-to-12-month window from August of the current year until September of the previous year. The significance of the correlations was tested using bootstrap resampling (Zang and Biondi, 2015). We also explored whether there were significant differences in the climate-growth correlations among different tree species within a specific stand. The Pearson r values were transformed to Fisher’s z to normalize the variance, followed by a pairwise t-test. This way we were able to determine if the differences among correlation coefficients were statistically significant. After identifying the climatic parameter with the highest correlations, we calculated the correlation coefficients for this parameter using a moving window approach (window size: 20 years, window offset: 1 year). This allowed us to search for temporal changes in growth sensitivity to this climatic driver over a period of 50 years.
We evaluated growth synchrony by calculating the mean inter-series correlation (Rbar), which reflects the average Pearson correlation among all tree-ring series within a specific chronology (Wigley et al., 1984). To obtain running synchrony values (Rbar), we used a 20-year moving window. Additionally, we assessed the significance of trends for the period from 1972 to 2021 using the Mann-Kendall trend test.
2.5 Growth response to droughts
A certain year was considered a drought year when the standardized 12-month water balance (from August of the current year until September of the previous year) was lower than −0.84 (Fuchs et al., 2021b), regardless of any observed growth reductions, as suggested by Schwarz et al. (2020). The range of the standardized 12-month water balance varied from −2.2 to 3.4. We selected all drought events for conducting the superposed epoch analysis (SEA) of growth depressions. For multi-year droughts, we selected the year with the most negative water balance. An epoch of 11 years was chosen, encompassing five years before and after each drought year. SEA calculates the mean departure in growth performance for each year within the epoch from the mean of all analysed epochs per chronology (Lough and Fritts, 1987). To define 95% confidence intervals of the departures, we employed bootstrapping with 5,000 random draws from the respective chronology. SEAs were conducted using the “dplR” package in R (Bunn, 2008).
We calculated three indices of drought response for each population based on the detrended radial growth (Lloret et al., 2011). These indices assess how the trees withstand drought (resistance), recover from growth reduction during the drought (recovery) and their capacity to reach pre-drought growth levels (resilience). We analysed the effects of drought on these indices using reference periods of varying lengths: 1 year, 3 years and 5 years of mean radial growth before and after the drought event. This approach helped limit the influence of other factors, such as defoliation caused by insects (Schwarz et al., 2020). For multi-year droughts, the resistance and recovery indices were calculated by averaging the radial growth across successive drought years.
To test for differences in drought indices - resistance, recovery and resilience - among species and sites, a simple one-way ANOVA and for pair-wise comparison Tukey HSD test was used (Abdi and Williams, 2010). We also analysed the relationship between recovery and resistance for each tree species across all sites and available drought events using linear regressions by examining the ANOVA p-value from the interaction of resistance by species and comparing the slopes in the R package “lsmeans” (Lenth, 2016).
3 Results
3.1 General characteristics of the chronologies
The mean annual tree-ring width (MRW) of the analysed species ranged from 1.3 to 3.0 mm, with no clear trend observed as we moved from moister to drier sites (Table 4). The standard deviation of ring widths for the two diffuse-porous species A. campestre and T. tomentosa was approximately 25% higher (ranging from 1.21 to 1.66 mm) compared to the ring-porous species Q. cerris, Q. pubescens and F. ornus, which ranged from 1.07 to 1.22 mm (Table 4). Among the sites, the lowest annual radial growth was observed in the Keszthely Mts. (1.88 mm) and the highest in the Somogy Hills (2.81 mm). The MRW did not show any significant relationship with tree age, suggesting that the populations experienced a stable growth rate during the period analyzed. Instead, the MRW of the tree populations was more closely associated with site conditions. Growth rates were lower in the Keszthely Mountains, which have low water-holding capacity, while areas with greater soil water capacity, such as Zselic, Vértes, and Szántód, exhibited higher growth rates (Tables 2, 4). The expressed population signal (EPS) was remarkably high, ranging from 0.85 to 0.99 for all species, consistently reaching or exceeding the generally accepted threshold of 0.85 (Wigley et al., 1984) (Table 4).
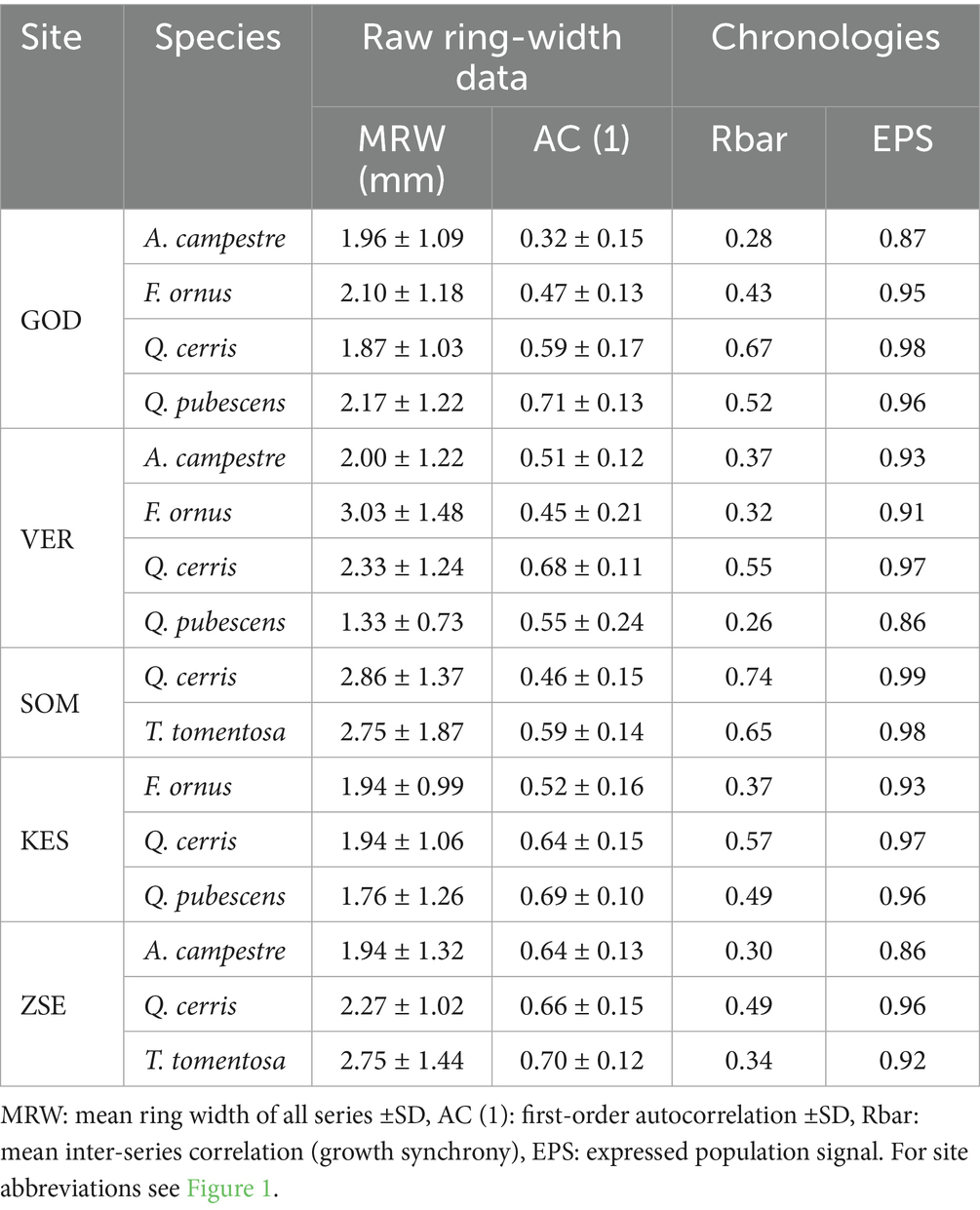
Table 4. Dendrochronological statistics for the ring-width series (raw data) and index-based chronologies (all truncated to the period 1972–2021) of the populations.
We observed only a few significant growth trends during the studied period. Specifically, the growth of A. campestre in Gödöllő Hills and Q. pubescens in Vértes Mts. has increased, while Q. cerris in Keszthely Mts. and T. tomentosa in the Zselic have shown a decline (Supplementary Figure S3).
3.2 Climate sensitivity of growth
The outlier analysis using Cook’s distance revealed that the growth of Q. cerris and Q. pubescens was significantly affected by the spongy moth outbreak (Lymantria dispar L.) in the Keszthely Mts. and the Somogy Hills sites in 2005 (Csóka and Hirka, 2009). This outbreak had a considerable effect on radial growth, obscuring the influence of climate during that year, leading us to exclude that year from further analysis in the affected populations.
Pearson’s correlation coefficients between monthly precipitation and ring widths were highest in May, June, and July of the current year, as well as in September of the previous year with r values reaching up to 0.52 (Figure 3A). Significant negative correlations with temperature were mainly observed from May to August of the current year with r values dropping to −0.48 (Figure 3B). Considering the intercorrelations among climatic variables considerably reduced the number of significant climatic variables (Supplementary Figure S4). Correlations of precipitation mostly remained significant during the May–July period and for September of the previous year (Supplementary Figure S4A). Furthermore, the temperature signals of May and June continued to show significance, particularly for A. campestre (Supplementary Figure S4B).
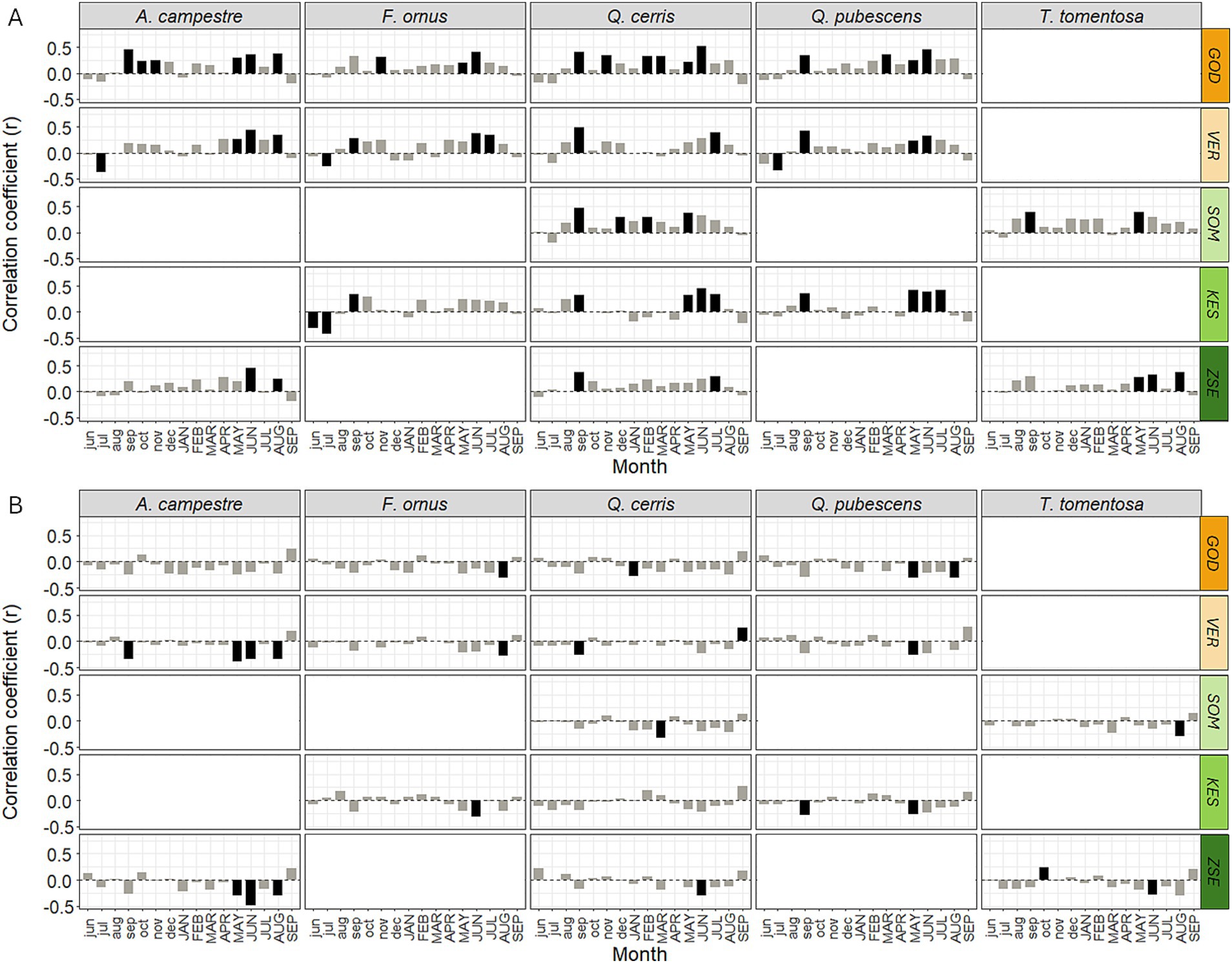
Figure 3. Correlation coefficients between monthly precipitation (A) and temperature (B) for the five tree species at the five sites. Months abbreviated with lower case letters refer to the previous year, months with capital letters to the current year. The color code indicates the gradient of mean annual precipitation (orange: low, green: high). Significant departures (p < 0.05) are shown with black bars. For site abbreviations see Figure 1.
The derived indices demonstrated a strong correlation with radial growth. The mean correlation coefficient for FAI was r = 0.45, while the mean for Is was r = 0.55 when considering all populations. The water balance displayed the highest positive correlation (ranging from r = 0.36 to r = 0.63) with radial growth during the current summer across different populations. The water balances in the spring of the current year and the autumn of the prior year also positively influenced tree growth. The radial growth of A. campestre, Q. cerris and T. tomentosa showed a moderate correlation (ranging from r = 0.38–0.52) with summer water balance, but only Q. cerris and T. tomentosa showed a higher correlation with winter water balance (r = 0.29 for both species) compared to the other species.
The 12-month water balance between previous year September and current year August (WBsep-aug) exhibited the highest correlation among the monthly aggregated water balance variables with radial growth, achieving an average correlation coefficient of r = 0.6 across most populations (Figure 4).
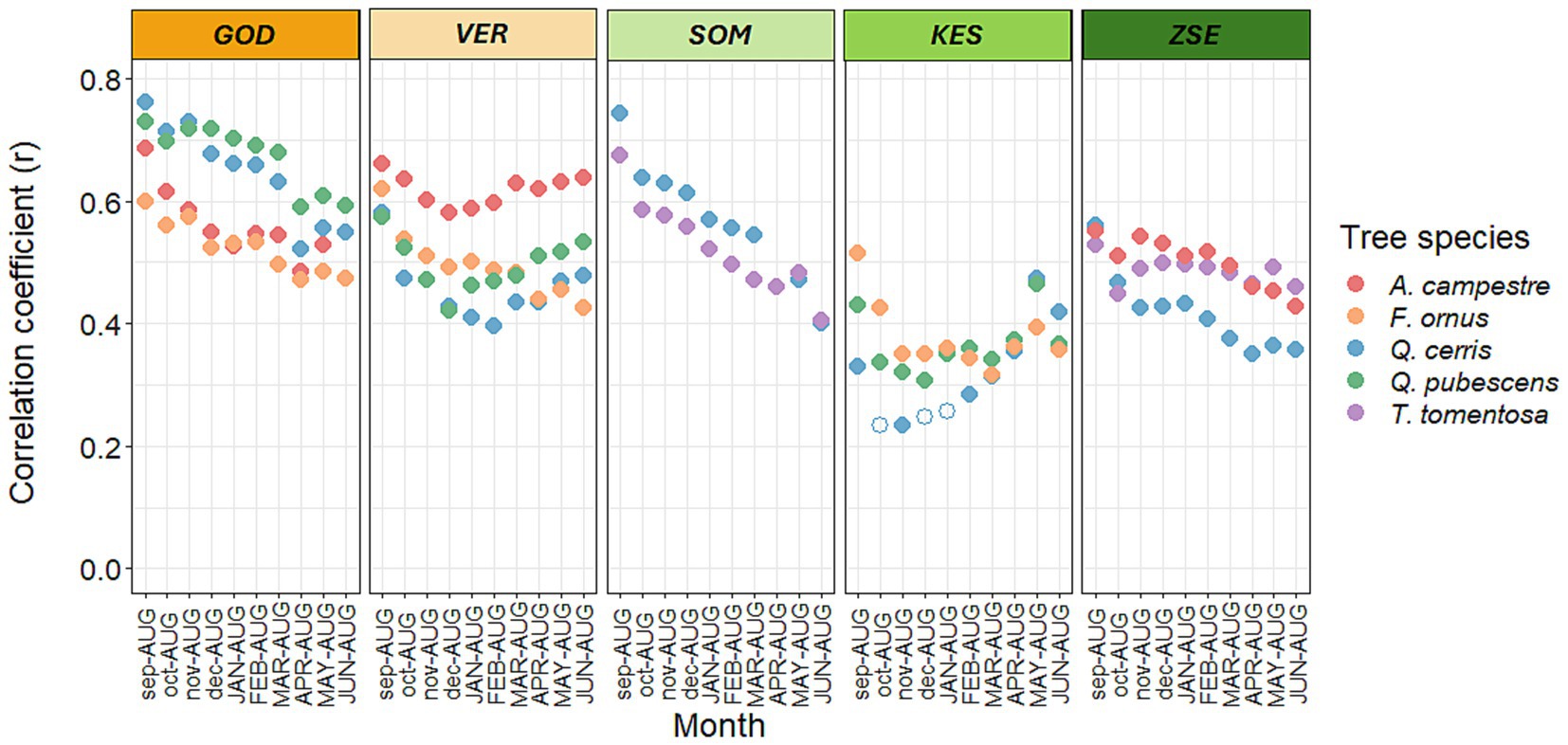
Figure 4. Correlation between radial growth (ring width index values) and the water balance (WB) for 3–12 months-window from the August of the current year until September of the previous year for the tree species at the five sites. Months abbreviated with lower case letters refer to the previous year, months with capital letters to the current year. The color spectrum from orange to green indicates the position of the site in the gradient of mean annual precipitation. Significant correlations are denoted with filled circles (p < 0.05).
This correlation was notably high for the climatically drier sites, with average coefficient values of r > =0.6, while it was comparatively lower for wetter sites (ranging from r = 0.42 to r = 0.55) (Figure 4). In the Keszthely Mts, the radial growth of Q. cerris and Q. pubescens showed the highest correlation with the four-month water balance from May to August of the current year. In contrast, A. campestre and T. tomentosa had stronger correlations with the summer water stress index (Is) in the Zselic (Figure 4).
The temporal change in correlations between the detrended chronologies and the seasonal water balance revealed the increasing (positive) effect of winter water balance on most sites and tree species, particularly over the past two decades (Figure 5). However, this trend was not apparent for A. campestre and T. tomentosa. Generally, the summer water balance played a crucial role for all populations, showing only a slight decline in the strength of its positive correlation in recent years. For the drier sites—GOD, VER, and SOM - the effect of spring water balance was significant, while it had a lesser impact on the wetter sites, KES and ZSE. Finally, the autumn water balance typically exhibited a negative correlation with radial growth, although some sites showed an increase in correlation (Figure 5).
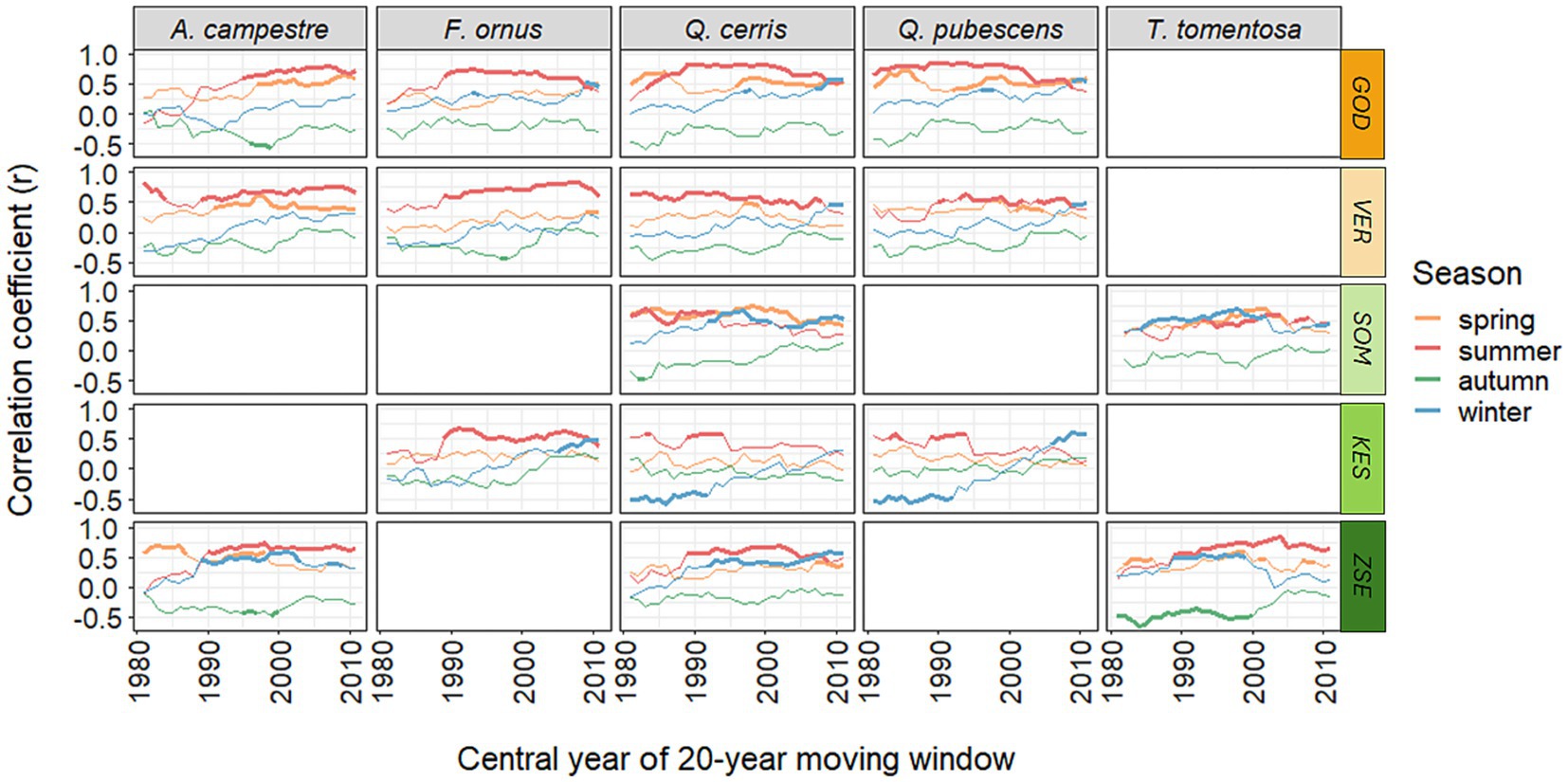
Figure 5. Moving window correlation analysis of climate-growth relationships in the five species at five sites for the period 1972–2021. Shown are the central years of the respective 20-year periods, testing for the correlation between the seasonal water balance variables (current spring: March–May, current summer: June–August, current autumn: September–November and previous winter: December–February) and annual ring width indices. The color code indicates the gradient of mean annual precipitation (orange: low, green: high). Significant periods are denoted with bold lines (p < 0.05). For site abbreviations see Figure 1.
Growth synchrony (Rbar) of the detrended chronologies was higher for Q. cerris, T. tomentosa and Q. pubescens than for A. campestre and F. ornus (Table 4). Generally, growth synchrony was lowest at the wettest site (Zselic) and highest at the driest site (Gödöllő) for most species. The lower growth synchrony of A. campestre at certain sites was due to the limited number of sampled trees and difficulties in accurately dating its tree rings, which often resulted from a high occurrence of uncertain ring boundaries. Notably, two-thirds of the populations exhibited a significant change in growth synchrony during the analysed period, most of which were increases (Figure 6). Recently, there has been a decrease in growth synchrony for most of populations except in Somogy Hills, where it has increased and remained at high levels for both Q. cerris and T. tomentosa over the past few decades (Figure 6).
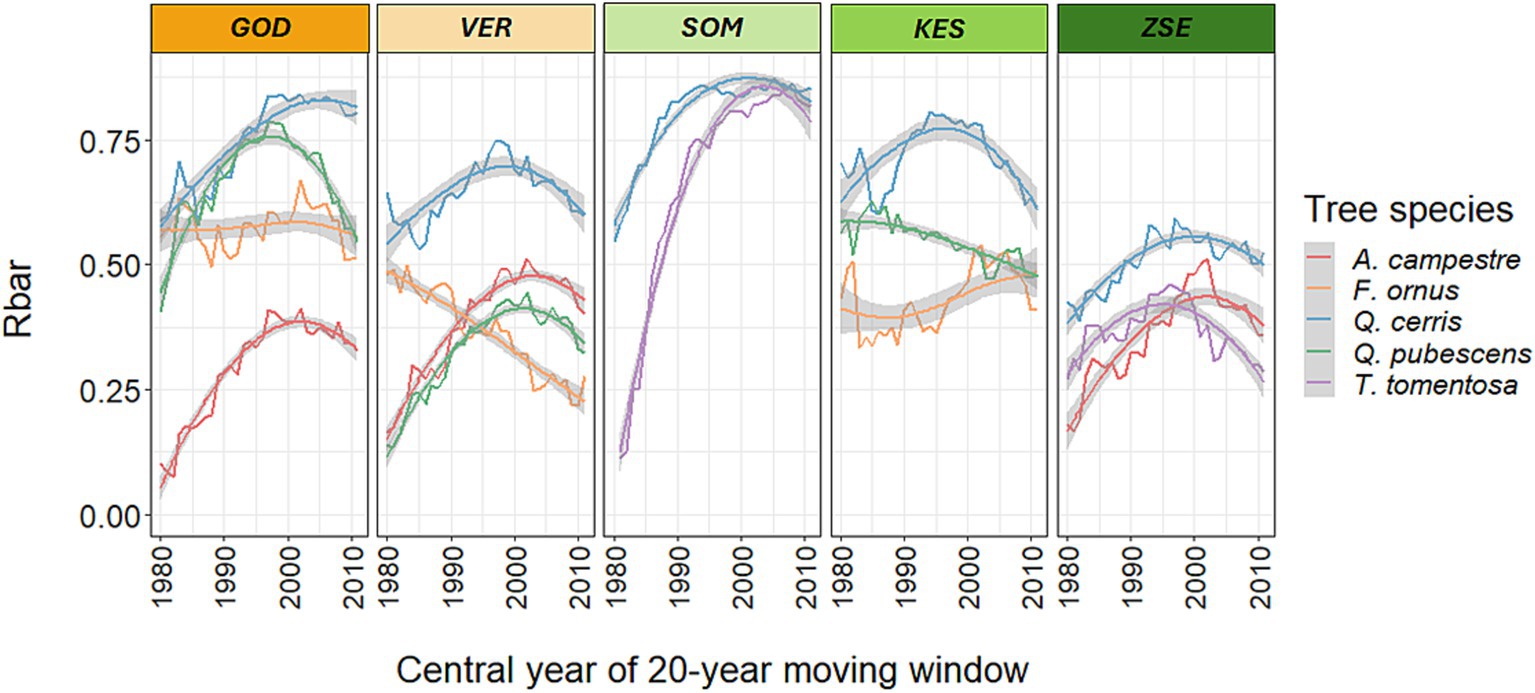
Figure 6. Moving averages of growth synchrony (Rbar: the mean correlation between all tree-ring index series in a chronology) of the five species at the five sites from 1972 to 2021 with loess regressions and 95% confidence intervals (span value: 1). Shown are the central years of the respective 20-year periods. The color code for site labels indicates the gradient of mean annual precipitation (orange: low, green: high). For site abbreviations see Figure 1.
3.3 Growth response to drought years
Drought years were rare events between 1971 and 1990 but their frequency and severity increased significantly afterward (Supplementary Figure S5). We identified 5–7 drought years across different sites (1983, 1990, 1992–1993, 2000–2003, 2007, 2011–2012 and 2017). Superposed epoch analysis (SEA) revealed a significant (p < 0.05) reduction in growth for all populations in the drought years (Figure 7). Among the analysed tree species, Q. pubescens, Q. cerris and A. campestre experienced the strongest growth reductions. Most sites showed no significant differences between ring-porous and diffuse-porous species. However, T. tomentosa demonstrated a lag effect after drought years at both study sites, showing reduced growth in subsequent years (Figure 7). Moving further away from the drought year, the differences become increasingly difficult to interpret, particularly since droughts have occurred every 2–3 years in recent decades.
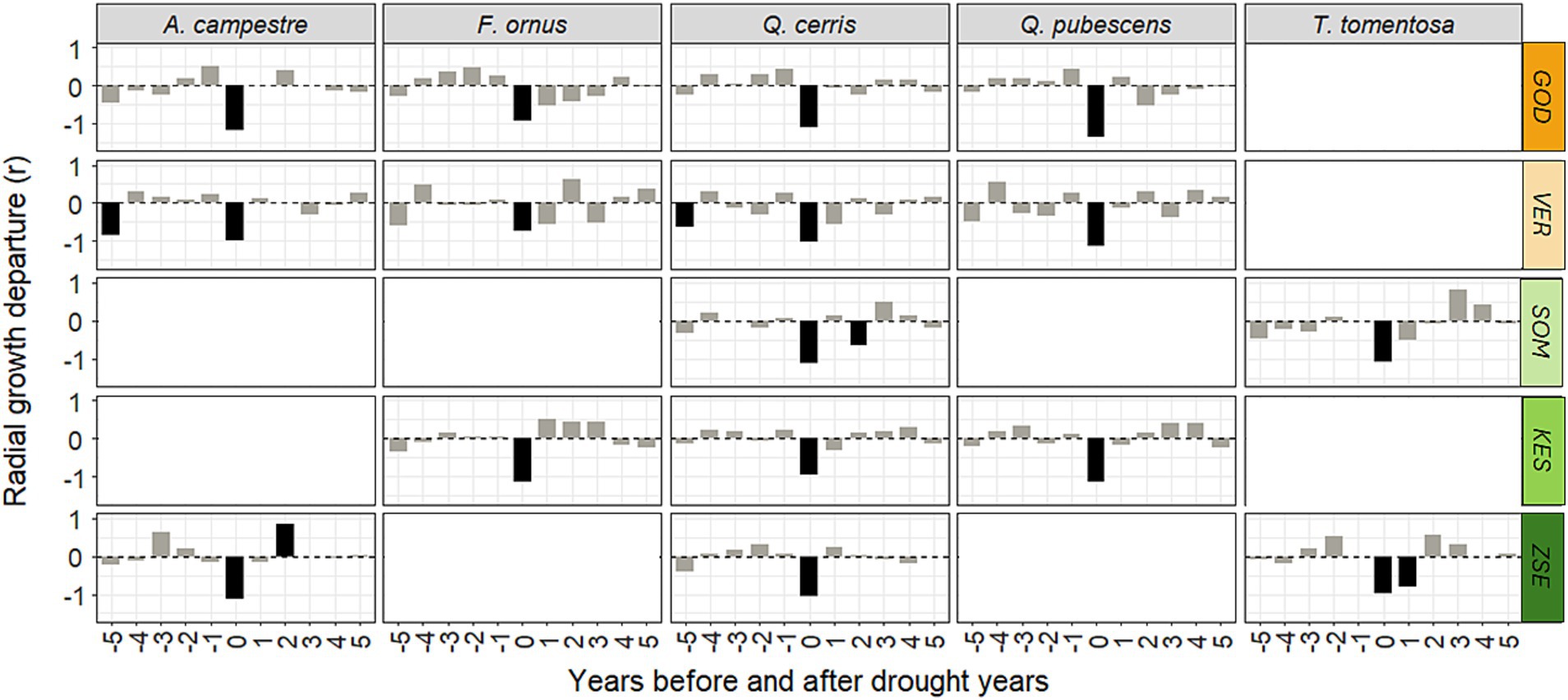
Figure 7. Results of superposed epoch analyses on the response of radial growth of the five species at the five sites to all drought events. Shown is the deviation of mean ring width in a 11-year period with the drought years in the center from the mean ring widths in all analysed 11-year epochs of the chronology. The color code for site labels indicates the gradient of mean annual precipitation (orange: low, green: high). Significant departures (p < 0.05) are displayed in black. For site abbreviations see Figure 1.
The combined data from all sites and drought events indicated comparable resistance levels among the five tree species (Figure 8). In terms of the recovery index, Q. cerris demonstrated the highest recovery value following drought, while F. ornus exhibited the least recovery in the 5 years after the drought event. Over the one-year reference period, a tendency of lower recovery can be seen for T. tomentosa (Supplementary Figure S6). The variability in recovery data was extensive for the two diffuse-porous species but more concentrated for the three ring-porous species. The growth resilience among the species was balanced (close to 1) with no significant differences noted during the five-year reference period, although a tendency for lower resilience in T. tomentosa was apparent in the distribution of index values (Figure 8).
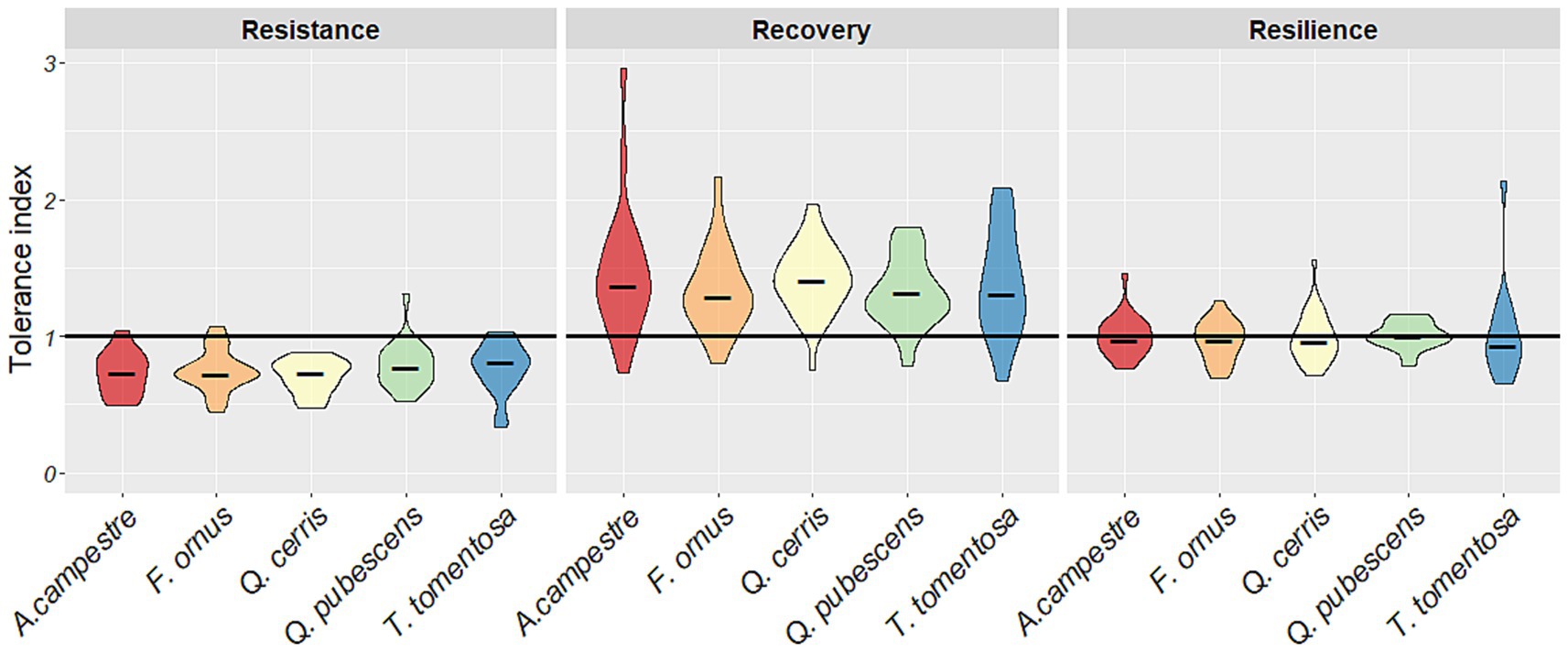
Figure 8. Resistance, recovery and resilience of radial growth of the five species from all sites and drought events using the five-year reference period. The violin plots depict the distributions of drought indices data using density curves. The width of each curve corresponds with the approximate frequency of data points in each region. Horizontal black lines of the violins are the medians.
Regression analysis of resistance and recovery for all species indicated that A. campestre, F. ornus and T. tomentosa generally have higher recovery values for lower resistance levels, than the two oak species (Figure 9). For the one-year reference period, we found no significant differences among the tree species, although the slope of regression was more gradual for T. tomentosa and F. ornus (Supplementary Figure S8). The slope of the linear regression between resistance and recovery values was significantly steeper for A. campestre compared to Q. cerris and Q. pubescens for both the three- and five-year reference periods (Supplementary Figure S9, Figure 9).
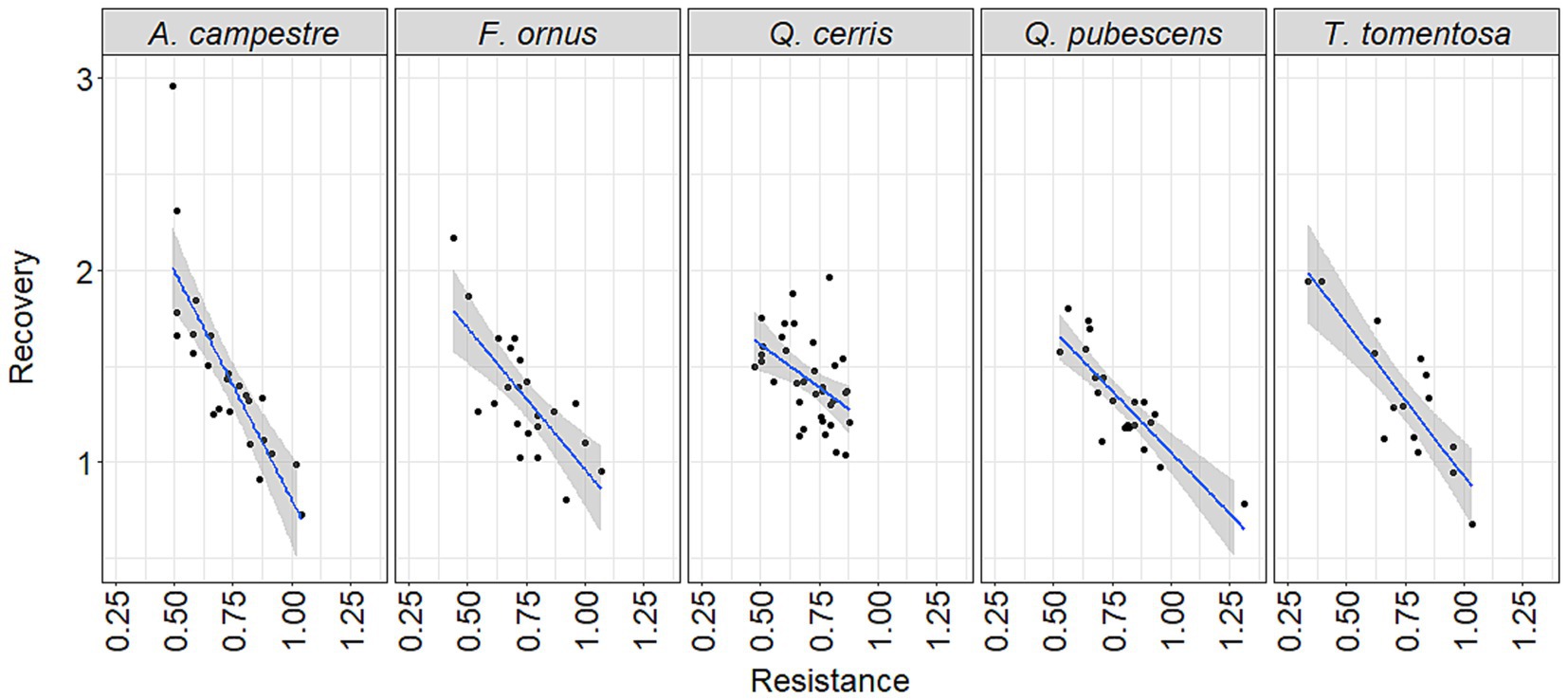
Figure 9. Relationship between the resistance and recovery values of the five species from all sites and available drought events using the five-year reference period. Solid blue lines denote linear regressions with confidence intervals (97.5%) are shown as shaded areas around the fit for the datapoints.
4 Discussion
4.1 Climate sensitivity of growth
Under stressful climatic conditions, particularly at the distribution margins of tree species, strong environmental drivers such as drought would commonly affect entire populations, leading to high growth synchrony among individuals (Shestakova et al., 2016; del Río et al., 2021). In contrast, in optimal climatic conditions, individual competitive abilities can develop better, resulting in more diverse growth patterns. In our study sites, we observed a constantly rising summer water deficit over the study period (Supplementary Figure S10). However, most populations have shown a bell-shaped growth synchrony curve with decreasing tendencies in recent decades (Figure 6). Although the number of stands examined was small, this may suggest an increased within-population diversity in responses to drought, i.e., that some individuals have a better capacity to adapt to the stressor (Muffler et al., 2020). Therefore, decreasing growth synchrony could signify conditions conducive for natural selection leading to local adaptation under changing climatic conditions, whereas high synchrony may reflect a lower adaptive potential, even in the face of significant stress. Factors such as microenvironmental heterogeneity, differing availability of deep soil water pools, and variable intrinsic tree characteristics—like tree height—could also explain the observed decrease in growth synchrony (Vilà-Cabrera et al., 2019; Muffler et al., 2020; Ripullone et al., 2020; González de Andrés et al., 2021).
The water balance was more important for the radial growth of all species than precipitation or temperature alone. This indicates that soil water availability, in conjunction with atmospheric evaporative demand regulates the water status of leaves, stems, and roots, ultimately affecting cambial growth activity in the investigated sites (Trotsiuk et al., 2021). It is worth mentioning that beyond the available soil water, extreme heat events can impact a wide variety of tree functions. At the leaf level, photosynthesis may be reduced, photooxidative stress can increase, leaves may abscise and at the whole plant level the growth rates can decrease (Teskey et al., 2015). Of the seasons analysed, the summer water balance had the most substantial impact on growth, a finding that aligns with several studies conducted in Central Europe (Fuchs et al., 2021b; Kasper et al., 2022; Mészáros et al., 2022). In the preceding year, only the precipitation in September was notable for all tree species and there was no significant carry-over effect from the past summer, as observed with F. sylvatica (Di Filippo et al., 2007; Müller-Haubold et al., 2015). Most populations showed the strongest correlations between growth and the water balance of the preceding 12 months, between September of the previous year and August of the growth year, similar to the observations on Quercus species in Serbia (Stojanović et al., 2018). However, in the Keszthely Mts (KES) the growth of oaks (Q. cerris and Q. pubescens) showed the strongest correlation with the water balance between May and July. A very similar growth response was found for oak stands studied ~30–40 km westward from the KES site (Kern et al., 2009). This trend is likely due to the low soil water capacity at that site (Table 3), which might contribute significantly to the limiting summer conditions for growth.
We found that the dry populations were more sensitive to climate than those in wetter locations (Figures 4, 5). This aligns with regional and European-wide studies that reported stronger growth responses in areas with low water availability compared to wetter sites (Scharnweber et al., 2011; Bose et al., 2021; Bouwman et al., 2025). Following the year 2000, we observed an increasing positive correlation between radial growth and winter water balance (Figure 5). This upward trend was more pronounced for the studied deep-rooting oak species than for the other investigated species with a more horizontal root system (Crow, 2005; Taneda and Sperry, 2008). The likely reason for this change is the shift in the overall water balance in our study sites over recent decades, driven by increased evapotranspiration pressure from rising temperatures, while precipitation levels have largely remained unchanged (Figure 2) (Trotsiuk et al., 2021). Consequently, the tree species in the examined sites may increasingly rely on deep soil water sources due to worsening drought conditions during the growing season, as reported in other studies involving various tree species (Mészáros et al., 2022).
4.2 Growth response to drought years
Lloret indices have revealed only a few significant differences in the drought response among the species. However, it was evident that A. campestre, F. ornus and T. tomentosa exhibited higher variability in their drought indices, particularly when compared to the two oak species studied (Figure 8, Supplementary Figure S6, S7). This observation aligns with previous studies, such as those by Kunz et al. (2018) and Schmucker et al. (2023), regarding A. campestre and Italiano et al. (2024) for F. ornus. In our study, A. campestre demonstrated a great capacity for recovery across all three reference periods, particularly during extreme droughts. In contrast, Q. pubescens showed the lowest variability of the drought indices and therefore the most balanced growth among the analysed tree species. This phenomenon is well-documented for temperate tree species that generally display lower resistance but tend to have higher recovery rates than their more resistant counterparts (Gazol et al., 2017; Schwarz et al., 2020). All examined tree species showed high resilience, with values around one, indicating their ability to maintain vitality and growth even after experiencing extreme droughts. Nonetheless, both the superposed epoch analysis and the Lloret indices revealed that T. tomentosa took longer to return to its pre-drought growth rate compared to the other species studied (Figure 7, Supplementary Figure S6). This finding is consistent with low resilience values for T. tomentosa in two- and five-year assessments following drought events in western Romania (Kasper et al., 2022). Leuschner et al. (2024) also found that Tilia (T. cordata Mill.) exhibits moderate drought resistance hardly withstanding extreme droughts, as noted by de Jaegere et al. (2016).
The seasonality and duration of drought events impact species resistance and resilience (D’Orangeville et al., 2018). However, due to the limited number of available drought events (ranging from 5 to 7 events per site), we could not take this into account. This is important because different tree species exhibit varying growth dynamics and wood anatomy, leading to different vulnerability to spring or summer droughts (Michelot et al., 2012). Additionally, in ring-porous species, the formation of earlywood is heavily influenced by the remobilization of stored carbon, making it less reflective of the actual weather conditions during that time (Michelot et al., 2012). Furthermore, we did not sample dead trees, as only a few were encountered during our study. Consequently, we were unable to evaluate any climatic factors that might have a fatal impact on tree growth.
In the absence of long-term mortality records for unmanaged stands of the studied species, it remains debatable whether tree species that are less responsive to droughts are more successful than those that are highly sensitive. Recently, Gessler et al. (2020) proposed that the delayed recovery of trees following a disturbance is not necessarily indicative of vitality loss due to the negative impacts of drought rather it may signify physiological acclimation processes. Drought-induced growth legacies and wood anatomical adjustments can lead to improved resistance to recurrent droughts (Tomasella et al., 2019). In this context, a reduction in growth following a drought can be viewed as a positive adjustment aimed at enhancing a tree’s long-term survival (Galiano et al., 2017). In contrast, fast-growing tree species often adopt a riskier strategy. After experiencing drought, damaged xylem conduits need to be rebuilt, which may decrease pest defence capacity and make these species more vulnerable to subsequent droughts (Beloiu et al., 2022).
The properties of the hydraulic system are also of central importance for the drought tolerance of tree species. The water-conducting channels in trees can withstand varying degrees of tension before embolisms—air bubbles forming in the xylem—occur. Hydraulic safety margins refer to the relationship between resistance at the xylem level and stomatal control (Meinzer et al., 2009). Specifically, these margins are defined as the difference between the minimum leaf water potential and a measure of xylem embolism resistance (e.g., P50). This represents a tree’s hydraulic strategy conservatism (Choat et al., 2012). Generally, Quercus species exhibit wider hydraulic safety margins and greater drought resistance (Cochard et al., 1992; Nardini and Pitt, 1999; Lobo et al., 2018). In contrast, Acer and Fraxinus species show variable but often moderate resistance (Schumann et al., 2019). Tilia, on the other hand, has narrower margins, making it more vulnerable to hydraulic dysfunction during severe drought conditions (Fuchs et al., 2021a). Conversely, the tree species, which display higher responsiveness to drought and narrower hydraulic safety margins may face considerable vulnerability to future climate change. In the current study, this is particularly true for A. campestre, F. ornus and T. tomentosa, consistent with findings by Kunz et al. (2018) in Germany. Additionally, other factors must be considered, such as the trunk’s water storage capacity, the rooting depth, and the competition among crowns and roots within a stand. Understanding the differences among tree species is essential for predicting how they will respond to climate change. This necessitates a more comprehensive approach to assessing the resilience of individual species and selecting the most appropriate species for planting in various environments.
5 Conclusion
All analysed species demonstrate high resilience to drought, with only slight differences among them. The tree species A. campestre, F. ornus and T. tomentosa show higher variability in their drought indices, particularly in terms of recovery and resilience, compared to the two oak species studied. A. campestre exhibits the greatest sensitivity to changing climatic conditions, as indicated by a significant increase in growth synchrony during the study period. In contrast, the age-independent growth of F. ornus remained stable and showed no clear changes in growth synchrony. Although it is slightly less sensitive to drought than A. campestre, the impact of winter water balance on its growth has increased. T. tomentosa displays similar sensitivity to drought and growth patterns as A. campestre and F. ornus; however, it shows a pronounced drought legacy in the first year following drought events. The decreasing trend in its growth and higher growth synchrony observed at the drier site suggest this species’ vulnerability. On the other hand, both Q. cerris and Q. pubescens exhibit a high degree of resilience to climate change. Their stable growth over recent decades, combined with lower variability in drought resistance and resilience, positions them as solid foundations for future forest stands even in drier climate conditions. Despite the differences, all analysed species are valuable options for enriching forest stands, as they demonstrate high resilience to drought and have not shown signs of dieback at the study sites.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
NM: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Writing – original draft, Writing – review & editing. IM: Conceptualization, Funding acquisition, Methodology, Supervision, Writing – original draft, Writing – review & editing. ZK: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Supervision, Writing – original draft, Writing – review & editing. GI: Data curation, Investigation, Supervision, Writing – original draft, Writing – review & editing. BG: Data curation, Investigation, Methodology, Writing – original draft, Writing – review & editing. CE: Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft, Writing – review & editing. IB: Conceptualization, Investigation, Supervision, Writing – original draft, Writing – review & editing. TN: Conceptualization, Data curation, Formal analysis, Funding acquisition, Investigation, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Project no. FK 142468 has been implemented with the support provided by the Ministry of Culture and Innovation of Hungary from the National Research, Development and Innovation Fund, financed under the FK_22 “OTKA” young researchers’ excellence programme funding scheme. This work has been supported by the János Bolyai Research Scholarship of the Hungarian Academy of Sciences (grant no. BO/00291/22/4).
Acknowledgments
The authors express their gratitude to the local state forest companies (SEFAG Ltd., branch office Szántód and Zselic; Bakonyerdő Ltd., branch office Keszthely; Vérteserdő Ltd., branch office Dél-Vértes; Pilisi Parkerdő Ltd., branch office Gödöllő) for their valuable support in the field. The laboratory of the Forest Research Institute is also acknowledged for the soil analysis. In addition, our colleagues are thanked for their help with the fieldwork and assistance. Lastly, we also grateful to the reviewers for their valuable and constructive criticism.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1625371/full#supplementary-material
References
Abdi, H., and Williams, L. J. (2010). “Tukey’s honestly significant difference (HSD) test” in Encyclopedia of research design. ed. N. J. Salkind (California: SAGE Publications), 1566–1570.
Allen, C. D., Breshears, D. D., and McDowell, N. G. (2015). On underestimation of global vulnerability to tree mortality and forest die-off from hotter drought in the Anthropocene. Ecosphere 6, 1–55. doi: 10.1890/ES15-00203.1
Andreu-Hayles, L., Planells, O., Gutierrez, E., Muntan, E., Helle, G., Anchkaitis, K. J., et al. (2011). Long tree-ring chronologies reveal 20th century increases in water-use efficiency but no enhancement of tree growth at five Iberian pine forests. Glob. Change Biol. 17, 2095–2112. doi: 10.1111/j.1365-2486.2010.02373.x
Beloiu, M., Stahlmann, R., and Beierkuhnlein, C. (2022). Drought impacts in forest canopy and deciduous tree saplings in central European forests. For. Ecol. Manag. 509:120075. doi: 10.1016/j.foreco.2022.120075
Biondi, F., and Qeadan, F. (2008). A theory-driven approach to tree-ring standardization: defining the biological trend from expected basal area increment. Tree-Ring Res. 64, 81–96. doi: 10.3959/2008-6.1
Bose, A., Scherrer, D., Camarero, J., Ziche, D., Babst, F., Bigler, C., et al. (2021). Climate sensitivity and drought seasonality determine post-drought growth recovery of Quercus petraea and Quercus robur in Europe. Sci. Total Environ. 784:147222. doi: 10.1016/j.scitotenv.2021.147222
Bouwman, M., Akhmetzyanov, L., Mohren, F., den Ouden, J., Sass-Klaassen, U., and Copini, P. (2025). Tree growth responses to severe droughts for assessment of forest growth potential under future climate. For. Ecol. Manag. 578:122423. doi: 10.1016/j.foreco.2024.122423
Briffa, K. R., Jones, P. D., Wigley, T. M. L., Pilcher, J. R., and Baillie, M. G. L. (1983). Climate reconstruction from tree rings: part 1, basic methodology and preliminary results for England. Int. J. Climatol. 3, 233–242. doi: 10.1002/joc.3370030303
Bunn, A. G. (2008). A dendrochronology program library in R (dplR). Dendrochronologia 26, 115–124. doi: 10.1016/j.dendro.2008.01.002
Cabrera, A., Premoli, A. C., and Jump, A. S. (2019). Refining predictions of population decline at species' rear edges. Glob. Change Biol. 25, 1549–1560. doi: 10.1111/gcb.14597
Camarero, J. J., Gazol, A., Sangüesa-Barreda, G., Cantero, A., Sánchez-Salguero, R., Sánchez-Miranda, A., et al. (2018). Forest growth responses to drought at short- and long-term scales in Spain: squeezing the stress memory from tree rings. Front. Ecol. Evol. 6, 1–11. doi: 10.3389/fevo.2018.00009
Castro-Díez, P., Vaz, A. S., Silva, J. S., van Loo, M., Alonso, Á., Aponte, C., et al. (2019). Global effects of non-native tree species on multiple ecosystem services. Biol. Rev. 94, 1477–1501. doi: 10.1111/brv.12511
Caudullo, G., and de Rigo, D. (2016). “Fraxinus ornus in Europe: distribution, habitat, usage and threats” in European atlas of Forest tree species. eds. J. San-Miguel-Ayanz, D. de Rigo, G. Caudullo, T. Houston Durrant, and A. Mauri (Luxembourg: Publications Office of the European Union), 100–101.
Choat, B., Jansen, S., Brodribb, T., Cochard, H., Delzon, S., Bhaskar, R., et al. (2012). Global convergence in the vulnerability of forests to drought. Nature 491, 752–755. doi: 10.1038/nature11688
Ciceu, A., Popa, I., Leca, S., Pitar, D., Chivulescu, S., and Badea, O. (2020). Climate change effects on tree growth from Romanian forest monitoring level II plots. Sci. Total Environ. 698:134129. doi: 10.1016/j.scitotenv.2019.134129
Cochard, H., Bréda, N., Granier, A., and Aussenac, G. (1992). Vulnerability to air embolism of three European oak species (Quercus petraea (Matt) Liebl, Q pubescens Willd, Q robur L). Ann. For. Sci. 49, 225–233. doi: 10.1051/forest:19920302
Cook, E. R., and Peters, K. (1981). The smoothing spline: a new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree-Ring Bull. 41, 45–53.
Cook, R. D., and Weisberg, S. (1982). Residuals and influence in regression. New York: Chapman and Hall.
Csóka, G. Y., and Hirka, A. (2009). A gyapjaslepke (Lymantria dispar L.) legutóbbi tömegszaporodása Magyarországon. Növényvédelem 45, 196–201.
D’Orangeville, L., Maxwell, J., Kneeshaw, D., Pederson, N., Duchesne, L., Logan, T., et al. (2018). Drought timing and local climate determine the sensitivity of eastern temperate forests to drought. Glob. Change Biol. 24, 2339–2351. doi: 10.1111/gcb.14096
Damesin, C., and Rambal, S. (1995). Field study of leaf photosynthetic performance by a Mediterranean deciduous oak tree (Quercus pubescens) during a severe summer drought. New Phytol. 131, 159–167. doi: 10.1111/j.1469-8137.1995.tb05717.xde
de Jaegere, T., Hein, S., and Claessens, H. (2016). A review of the characteristics of small-leaved lime (Tilia cordata mill.) and their implications for silviculture in a changing climate. Forests 7:56. doi: 10.3390/f7030056
de Rigo, D., Caudullo, G., Houston Durrant, T., and San-Miguel-Ayanz, J. (2016) The European atlas of Forest tree species: modelling, data and information on forest tree species J. San-Miguel-Ayanz, D. Rigode, G. Caudullo, T. Houston Durrant, and A. Mauri European atlas of Forest tree species Publications Office of the European Union Luxembourg 40–45.
del Río, M., Vergarechea, M., Hilmers, T., Alday, J. G., Avdagi’c, A., Binderh, F., et al. (2021). Effects of elevation-dependent climate warming on intra-and inter-specific growth synchrony in mixed mountain forests. For. Ecol. Manag. 479:118587. doi: 10.1016/j.foreco.2020.118587
Di Filippo, A., Biondi, F., Čufar, K., De Luis, M., Grabner, M., Maugeri, M., et al. (2007). Bioclimatology of beech (Fagus sylvatica L.) in the eastern Alps: spatial and altitudinal climatic signals identified through a tree-ring network. J. Biogeogr. 34, 1873–1892. doi: 10.1111/j.1365-2699.2007.01747.x
Eaton, E., Caudullo, G., and de Rigo, D. (2016) Quercus petraea in Europe: distribution, habitat, usage and threats J. San-Miguel-Ayanz, D. Rigode, G. Caudullo, T. Houston Durrant, and A. Mauri European atlas of Forest tree species Publications Office of the European Union Luxembourg 160–163
Eilmann, B., Weber, P., Rigling, A., and Eckstein, D. (2006). Growth reactions of Pinus sylvestris L. and Quercus pubescens Willd. To drought years at a xeric site in Valais, Switzerland. Dendrochronologia 23, 121–132. doi: 10.1016/j.dendro.2005.10.002
Fuchs, S., Leuschner, C., Mathias Link, R., and Schuldt, B. (2021a). Hydraulic variability of three temperate broadleaf tree species along a water availability gradient in Central Europe. New Phytol. 231, 1387–1400. doi: 10.1111/nph.17448
Fuchs, S., Schuldt, B., and Leuschner, C. (2021b). Identification of drought-tolerant tree species through climate sensitivity analysis of radial growth in central European mixed broadleaf forests. For. Ecol. Manag. 494:119287. doi: 10.1016/j.foreco.2021.119287
Führer, E., Horváth, L., Jagodics, A., Machon, A., and Szabados, I. (2011). Application of a new aridity index in Hungarian forestry practice. Időjárás 115, 205–216.
Gagen, M., Finsinger, W., Wagner-Cremer, F., McCarroll, D., Loader, N. J., Robertson, I., et al. (2011). Evidence of changing intrinsic water-use efficiency under rising atmospheric CO2 concentrations in boreal Fennoscandia from subfossil leaves and tree ring d13C ratios. Glob. Change Biol. 17, 1064–1072. doi: 10.1111/j.1365-2486.2010.02273.x
Galiano, L., Timofeeva, G., Saurer, M., Siegwolf, R., Martínez-Vilalta, J., Hommel, R., et al. (2017). The fate of recently fixed carbon after drought release: towards unravelling C storage regulation in Tilia platyphyllos and Pinus sylvestris. Plant Cell Environ. 40, 1711–1724. doi: 10.1111/pce.12972
Gazol, A., Camarero, J. J., Anderegg, W. R. L., and Vicente-Serrano, S. M. (2017). Impacts of droughts on the growth resilience of northern hemisphere forests. Glob. Ecol. Biogeogr. 26, 166–176. doi: 10.1111/geb.12526
Gessler, A., Bottero, A., Marshall, J., and Arend, M. (2020). The way back: recovery of trees from drought and its implication for acclimation. New Phytol. 228, 1704–1709. doi: 10.1111/nph.16703
González de Andrés, E., Rosas, T., Camarero, J. J., and Martinez Vilalta, J. (2021). The intraspecific variation of functional traits modulates drought resilience of European beech and Q. Pubescens. J. Ecol. 109, 3652–3669. doi: 10.1111/1365-2745.13743
Granier, A., Bréda, N., Biron, P., and Villette, S. (1999). A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands. Ecol. Model. 116, 269–283. doi: 10.1016/S0304-3800(98)00205-1
Hammond, W. M., Williams, A. P., Abatzoglou, J. T., Adams, H. D., Klein, T., López, R., et al. (2022). Global field observations of tree die-off reveal hotter-drought fingerprint for earth’s forests. Nat. Commun. 13:1761. doi: 10.1038/s41467-022-29289-2
Hanewinkel, M., Cullmann, D. A., Schelhaas, M. J., Nabuurs, G. J., and Zimmermann, N. E. (2012). Climate change may cause severe loss in the economic value of European forest land. Nat. Clim. Chang. 3, 203–207. doi: 10.1038/nclimate1687
Hemery, G. E., Clark, J. R., Aldinger, E., Claessens, H., Malvolti, M. E., O'connor, E., et al. (2010). Growing scattered broadleaved tree species in Europe in a changing climate: a review of risks and opportunities. Forestry 83, 65–81. doi: 10.1093/FORESTRY/CPP034
Hirka, A. (2022) Biotic and abiotic damage in the Hungarian forests in 2021, and the damage forecast for 2022. Report of the Hungarian Forest research institute, Budapest (in Hungarian)
HMS (2022). Meteorological Database. Available online at: https://odp.met.hu/climate/homogenized_data/gridded_data_series/ (Accessed 12 April 2022).
Holmes, R. L. (1983). Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 43, 69–78.
Ionita, M., and Nagavciuc, V. (2021). Changes in drought features at the European level over the last 120 years. Nat. Hazards Earth Syst. Sci. 21, 1685–1701. doi: 10.5194/nhess-21-1685-2021
Italiano, S. S. P., Camarero, J. J., Borghetti, M., Colangelo, M., Rita, A., and Ripullone, F. (2024). Drought legacies in mixed Mediterranean forests: Analysing the effects of structural overshoot, functional traits and site factors. Sci.Total Environ. 927:172166. doi: 10.1016/j.scitotenv.2024.172166
Kasper, J., Leuschner, C., Walentowski, H., Petritan, A. M., and Weigel, R. (2022). Winners and losers of climate warming: declining growth in Fagus and Tilia vs. stable growth in three Quercus species in the natural beech–oak forest ecotone (western Romania). For. Ecol. Manag. 506:119892. doi: 10.1016/j.foreco.2021.119892
Kern, Z., Grynaeus, A., and Morgós, A. (2009). Reconstructed precipitation for southern Bakony Mountains (Transdanubia, Hungary) back to AD 1746 based on ring widths of oak trees. Időjárás 113, 299–314.
Kunz, J., Loffler, G., and Bauhus, J. (2018). Minor European broadleaved tree species are more drought-tolerant than Fagus sylvatica but not more tolerant than Quercus petraea. For. Ecol. Manag. 414, 15–27. doi: 10.1016/j.foreco.2018.02.016
Latte, N., Taverniers, P., de Jaegere, T., and Claessens, H. (2020). Dendroecological assessment of climate resilience of the rare and scattered forest tree species Tilia cordata mill. In northwestern Europe. Forestry 93, 675–684. doi: 10.1093/forestry/cpaa011
Lenth, R. V. (2016). Least-squares means: the R package lsmeans. J. Stat. Softw. 69, 1–33. doi: 10.18637/jss.v069.i01
Leuschner, C., and Ellenberg, H. (2017). Ecology of central European non-forest vegetation: Coastal to alpine, natural to man-made habitats (Vegetation Ecology of Central Europe Volume II). Switzerland: Springer International Publishing.
Leuschner, C., Fuchs, S., Wedde, P., Rüther, E., and Schuldt, B. (2024). A multi-criteria drought resistance assessment of temperate Acer, Carpinus, Fraxinus, Quercus and Tilia species. Perspect. Plant Ecol. Evol. Syst. 62:125777. doi: 10.1016/j.ppees.2023.125777
Leuschner, C., Wedde, P., and Lübbe, T. (2019). The relation between pressure–volume curve traits and stomatal regulation of water potential in five temperate broadleaf tree species. Ann. For. Sci. 76:93. doi: 10.1007/s13595-019-0838-7
Lloret, F., Keeling, E. G., and Sala, A. (2011). Components of tree resilience: effects of successive low-growth episodes in old ponderosa pine forests. Oikos 120, 1909–1920. doi: 10.1111/j.1600-0706.2011.19372.x
Lobo, A., Torres-Ruiz, J. M., Burlett, R., Lemaire, C., Parise, C., Francioni, C., et al. (2018). Assessing inter- and intraspecific variability of xylem vulnerability to embolism in oaks. For. Ecol. Manag. 424, 53–61. doi: 10.1016/j.foreco.2018.04.031
Lough, J. M., and Fritts, H. C. (1987). An assessment of the possible effects of volcanic eruptions on north American climate using tree-ring data, 1602 to 1900 a.D. Clim. Chang. 10, 219–239. doi: 10.1007/BF00143903
Matevski, D., and Schuldt, A. (2021). Tree species richness, tree identity and non-native tree proportion affect arboreal spider diversity, abundance and biomass. For. Ecol. Manag. 483:118775. doi: 10.1016/j.foreco.2020.118775
Mátyás, C. (2021). Adaptive pattern of phenotypic plasticity and inherent growth reveal the potential for assisted transfer in sessile oak (Quercus petraea L.). For. Ecol. Manag. 482:118832. doi: 10.1016/j.foreco.2020.118832
Mátyás, C., Berki, I., Bidló, A., Csóka, G., Czimber, K., Führer, E., et al. (2018). Sustainability of forest cover under climate change on the temperate-continental xeric limits. Forests 9:489. doi: 10.3390/f9080489
Mazza, G., Markou, L., and Sarris, D. (2021). Species-specific growth dynamics and vulnerability to drought at the single tree level in a Mediterranean reforestation. Trees 35, 1697–1710. doi: 10.1007/s00468-021-02151-6
McCabe, G. J., and Markstrom, S. L. (2007). A monthly water-balance model driven by a graphical user interface. U. S. Geol. Surv. Open-File Rep. 1008:12. doi: 10.3133/ofr20071088
Meinzer, F. C., Johnson, D. M., Lachenbruch, B., McCulloh, K. A., and Woodruff, D. R. (2009). Xylem hydraulic safety margins in woody plants: coordination of stomatal control of xylem tension with hydraulic capacitance. Funct. Ecol. 23, 922–930. doi: 10.1111/j.1365-2435.2009.01577.x
Mészáros, I., Adorján, B., Nyitrai, B., Kanalas, P., Oláh, V., and Levanič, T. (2022). Long-term radial growth and climate-growth relationships of Quercus petraea (Matt.) Liebl. And Quercus cerris L. in a xeric low elevation site from Hungary. Dendrochronologia 76:126014. doi: 10.1016/j.dendro.2022.126014
Michelot, A., Simard, S., Rathgeber, C., Dufrêne, E., and Damesin, C. (2012). Comparing the intra annual wood formation of three European species (Fagus sylvatica, Quercus petraea and Pinus sylvestris) as related to leaf phenology and non-structural carbohydrate dynamics. Tree Physiol. 32, 1033–1045. doi: 10.1093/treephys/tps052
Móricz, N., Garamszegi, B., Rasztovits, E., Bidló, A., Horváth, A., Jagicza, A., et al. (2018). Recent drought-induced vitality decline of black pine (Pinus nigra Arn.) in south-West Hungary–is this drought-resistant species under threat by climate change? Forests 9:414. doi: 10.3390/f9070414
Móricz, N., Illés, G., Mészáros, I., Garamszegi, B., Berki, I., Bakacsi, Z., et al. (2021). Different drought sensitivity traits of young sessile oak (Quercus petraea (Matt.) Liebl.) and Turkey oak (Quercus cerris L.) stands along a precipitation gradient in Hungary. For. Ecol. Manag. 492:119165. doi: 10.1016/j.foreco.2021.119165
Muffler, L., Weigel, R., Hacket-Pain, A. J., Klisz, M., Maaten, E., and Wilmking, M. (2020). Lowest drought sensitivity and decreasing growth synchrony towards the dry distribution margin of European beech. J. Biogeogr. 47, 1910–1921. doi: 10.1111/jbi.13884
Müller-Haubold, H., Hertel, D., and Leuschner, C. (2015). Climatic drivers of mast fruiting in European beech and resulting C and N allocation shifts. Ecosystems 18, 1083–1100. doi: 10.1007/s10021-015-9885-6
Nagy, L., and Ducci, F. (2004). EUFORGEN technical guidelines for genetic conservation and use for A. campestre. Italy: International Plant Genetic Resources Institute.
Nardini, A., and Pitt, F. (1999). Drought resistance of Quercus pubescens as a function of root hydraulic conductance, xylem embolism and hydraulic architecture. New Phytol. 143, 485–493. doi: 10.1046/j.1469-8137.1999.00476.x
National Forestry Database (2021) Stand-based inventory. Available at: https://erdoleltar.nfk.gov.hu
Norton, D., Palmer, J., and Ogden, J. (1987). Dendroecological studies in New Zealand 1. An evaluation of tree age estimates based on increment cores. N. Z. J. Bot. 25, 373–383. doi: 10.1080/0028825X.1987.10413355
Nowacki, G. J., and Abrams, M. D. (1997). Radial-growth averaging criteria for reconstruction disturbance histories from presettlement-origin oaks. Ecol. Monogr. 67, 225–249. doi: 10.2307/2963514
Pasta, S., de Rigo, D., and Caudullo, G. (2016). “Quercus pubescens in Europe: distribution, habitat, usage and threats” in European atlas of Forest tree species. eds. J. San-Miguel-Ayanz, D. de Rigo, G. Caudullo, T. Houston Durrant, and A. Mauri (Luxembourg: Publications Office of the European Union), 156–157.
Peltier, D. M. P., Fell, M., and Ogle, K. (2016). Legacy effects of drought in the southwestern United States: a multi-species synthesis. Ecol. Monogr. 86, 312–326. doi: 10.1002/ecm.1219
Pretzsch, H., Grams, T., Häberle, K. H., Pritsch, K., Bauerle, T., and Rätzer, T. (2020). Growth and mortality of Norway spruce and European beech in monospecific and mixed species stands under natural episodic and experimentally extended drought. Results of the KROOF throughfall exclusion experiment. Trees 34, 957–970. doi: 10.1007/s00468-020-01973-0
Pretzsch, H., Schütze, G., and Uhl, E. (2013). Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation. Plant Biol. 15, 483–495. doi: 10.1111/j.1438-8677.2012.00670.x
Ripullone, F., Camarero, J. J., Colangelo, M., and Voltas, J. (2020). Variation in the access to deep soil water pools explains tree-to-tree differences in drought-triggered dieback of Mediterranean oaks. Tree Physiol. 40, 591–604. doi: 10.1093/treephys/tpaa026
Sapsford, S. J., Brandt, A. J., Davis, K. T., Peralta, G., Dickie, I. A., Gibson, R. D., et al. (2020). Towards a framework for understanding the context dependence of impacts of non-native tree species. Funct. Ecol. 34, 944–955. doi: 10.1111/1365-2435.13544
Scharnweber, T., Manthey, M., Criegee, C., Bauwe, A., Schröder, C., and Wilmking, M. (2011). Drought matters – declining precipitation influences growth of Fagus sylvatica L. and Quercus robur L. in North-Eastern Germany. For. Ecol. Manag. 262, 947–961. doi: 10.1016/j.foreco.2011.05.026
Schmitt, A., Trouvé, R., Seynave, I., and Lebourgeois, F. (2020). Decreasing stand density favors resistance, resilience, and recovery of Quercus petraea trees to a severe drought, particularly on dry sites. Ann. For. Sci. 77:52. doi: 10.1007/s13595-020-00959-9
Schmucker, J., Uhl, E., Schmied, G., and Pretzsch, H. (2023). Growth and drought reaction of European hornbeam, European white elm, A. Campestre and wild service tree. Trees 37, 1515–1536. doi: 10.1007/s00468-023-02441-1
Schuldt, B., Buras, A., Arend, M., Vitasse, Y., Beierkuhnlein, C., Damm, A., et al. (2020). A first assessment of the impact of the extreme 2018 summer drought on central European forests. Basic Appl. Ecol. 45, 86–103. doi: 10.1016/j.baae.2020.04.003
Schumann, K., Leuschner, C., and Schuldt, B. (2019). Xylem hydraulic safety and efficiency in relation to leaf and wood traits in three temperate Acer species differing in habitat preferences. Trees 33, 1475–1490. doi: 10.1007/s00468-019-01874-x
Schwarz, J., Skiadaresis, G., Kohler, M., Kunz, J., Schnabel, F., and Vitali, V. (2020). Quantifying growth responses of trees to drought–a critique of commonly used resilience indices and recommendations for future studies. Curr. For. Rep. 6, 185–200. doi: 10.1007/s40725-020-00119-2
Senf, C., Buras, A., Zang, C. S., Rammig, A., and Seidl, R. (2020). Excess forest mortality is consistently linked to drought across Europe. Nat. Commun. 11:6200. doi: 10.1038/s41467-020-19924-1
Shestakova, T. A., Gutiérrez, E., Kirdyanov, A. V., Camarero, J. J., Génova, M., Knorre, A. A., et al. (2016). Forests synchronize their growth in contrasting Eurasian regions in response to climate warming. Proc. Natl. Acad. Sci. USA 113, 662–667. doi: 10.1073/pnas.1514717113
Spinoni, J., Naumann, G., and Vogt, J. V. (2017). Pan-european seasonal trends and recent changes of drought frequency and severity. Glob. Planet. Change 148, 113–130. doi: 10.1016/j.gloplacha.2016.11.013
Steckel, M., Moser, W. K., del Río, M., and Pretzsch, H. (2020). Implications of reduced stand density on tree growth and drought susceptibility: a study of three species under varying climate. Forests 11:627. doi: 10.3390/f11060627
Stojanović, D. B., Levanič, T., Matović, B., Stjepanović, S., and Orlović, S. (2018). Growth response of different tree species (oaks, beech and pine) from SE Europe to precipitation over time. Dendrobiology 79, 97–110. doi: 10.12657/denbio.079.009
Taneda, H., and Sperry, J. S. (2008). A case-study of water transport in co-occurring ring- versus diffuse-porous trees: contrasts in water-status, conducting capacity, cavitation and vessel refilling. Tree Physiol. 28, 1641–1651. doi: 10.1093/treephys/28.11.1641
Teskey, R., Wertin, T., Bauweraerts, I., Ameye, M., Mcguire, M. A., and Steppe, K. (2015). Tree response to extreme heat. Plant Cell Environ. 38, 1699–1712. doi: 10.1111/pce.12417
Thornthwaite, C. W. (1948). An approach toward a rational classification of climate. Geogr. Rev. 38, 55–94. doi: 10.2307/210739
Tomasella, M., Nardini, A., Hesse, B. D., Machlet, A., Matyssek, R., and Häberle, K. H. (2019). Close to the edge: effects of repeated severe drought on stem hydraulics and nonstructural carbohydrates in European beech saplings. Tree Physiol. 39, 717–728. doi: 10.1093/treephys/tpy142
Trotsiuk, V., Babst, F., Grossiord, C., Gessler, A., Forrester, D. I., Buchmann, N., et al. (2021). Tree growth in Switzerland is increasingly constrained by rising evaporative demand. J. Ecol. 109, 2981–2990. doi: 10.1111/1365-2745.13712
Vitasse, Y., Bottero, A., Cailleret, M., Bigler, C., Fonti, P., Gessler, A., et al. (2019). Contrasting resistance and resilience to extreme drought and late spring frost in five major European tree species. Glob. Change Biol. 25, 3781–3792. doi: 10.1111/gcb.14803
Vodnik, D., Gričar, J., Lavrič, M., Ferlan, M., Hafner, P., and Eler, K. (2019). Anatomical and physiological adjustments of Q. Pubescens Willd. From two adjacent sub-Mediterranean ecosites. Environ. Exp. Bot. 165, 208–218. doi: 10.1016/j.envexpbot.2019.06.010
Walentowski, H., Falk, W., Mette, T., Kunz, J., Bräuning, A., Meinardus, C., et al. (2014). Assessing future suitability of tree species under climate change by multiple methods: a case study in southern Germany. Ann. For. Res. 60, 101–126. doi: 10.15287/afr.2016.789
Wigley, T. M. L., Briffa, K. R., and Jones, P. D. (1984). On the Average Value of Correlated Time Series, with Applications in Dendroclimatology and Hydrometeorology. J. Appl. Meteor. Climatol. 23, 201–213. doi: 10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2
Yue, C., Kohnle, U., Hanewinkel, M., and Klädtke, J. (2011). Extracting environmentally driven growth trends from diameter increment series based on a multiplicative decomposition model. Can. J. For. Res. 41, 1577–1589. doi: 10.1139/x11-056
Zang, C., and Biondi, F. (2015). Treeclim: an R package for the numerical calibration of proxy-climate relationships. Ecography 38, 431–436. doi: 10.1111/ecog.01335
Zecchin, B., Caudullo, G., and de Rigo, D. (2016) Acer campestre in Europe: distribution, habitat, usage and threats J. San-Miguel-Ayanz, D. Rigode, G. Caudullo, T. Houston Durrant, and A Mauri European atlas of Forest tree species Publications Office of the European Union Luxembourg 52–53.
Zhang, Y., and Schaap, M. G. (2017). Weighted recalibration of the Rosetta pedotransfer model with improved estimates of hydraulic parameter distributions and summary statistics (Rosetta3). J. Hydrol. 547, 39–53. doi: 10.1016/j.jhydrol.2017.01.004
Keywords: minor native tree species, drought tolerance, growth, tree rings, Quercus, dendrochronology
Citation: Móricz N, Mészáros I, Kern Z, Illés GZ, Garamszegi B, Eötvös CB, Berki I and Németh TM (2025) Evaluating the drought tolerance of five native broadleaf tree species using dendroecological analysis in East Central Europe. Front. For. Glob. Change. 8:1625371. doi: 10.3389/ffgc.2025.1625371
Edited by:
Gabriel Sangüesa-Barreda, University of Valladolid, SpainReviewed by:
Cristina Valeriano, Spanish National Research Council (CSIC), SpainEnrico Tonelli, Marche Polytechnic University, Italy
Copyright © 2025 Móricz, Mészáros, Kern, Illés, Garamszegi, Eötvös, Berki and Németh. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Norbert Móricz, bW9yaWN6Lm5vcmJlcnRAdW5pLXNvcHJvbi5odQ==