 Yiming Wang
Yiming Wang Xiaohua Liu1
Xiaohua Liu1 Gu-ai Xie
Gu-ai Xie- 1Forestry Pest Control Research Institute, Jiangxi Academy of Forestry, Nanchang, China
- 2Jiangxi Institute of Natural Resources and Monitoring, Nanchang, China
Introduction: Pine wilt disease (PWD), caused by Bursaphelenchus xylophilus, has led to widespread logging interventions in subtropical China, especially clearcutting, yet the ecological consequences for insect communities remain poorly understood. Insects play critical roles in ecosystem functioning, making it essential to quantify how different logging strategies shape their diversity, community composition, and functional guilds.
Methods: We established 48 stratified plots across four PWD-affected counties in Jiangxi Province to compare unlogged controls (UC), selection cutting (SC), and clearcutting (CC) treatments. Insect diversity was evaluated using Shannon and Simpson indices, while community structure was analyzed with PERMANOVA and NMDS ordinations. Indicator species were identified via IndVal analysis. Functional guild dynamics were measured across trophic groups, and structural equation modeling (SEM) was used to assess vegetation cover and litter thickness as mediators of diversity responses.
Results: Clearcutting caused significant declines in insect diversity (Shannon index: UC 3.63 ± 0.02 vs. CC 3.14 ± 0.18; F = 19.17, p = 0.00057; Simpson index: UC 0.960 vs. CC 0.919). PERMANOVA attributed 60.7–89.5% of variation in insect community structure to logging treatment (p < 0.05), with NMDS ordinations showing distinct clustering: CC sites were dominated by disturbance-tolerant taxa (e.g., Cicindelidae, IndVal = 0.94), while UC sites harboured specialists (e.g., Libellulidae, IndVal = 0.95). Functional guild analysis revealed strong reductions in predators (−63.7%) and parasitoids (p < 0.001) under CC, whereas decomposers were relatively resilient. SEM demonstrated that vegetation cover (β = 0.69, p < 0.001) and litter thickness (β = 0.25, p = 0.041) acted as key mediators, with logging intensity indirectly depressing insect diversity via microhabitat degradation (model R2 = 0.81).
Discussion and conclusion: Our findings show that intensive clearcutting exacerbates biodiversity loss and trophic collapse in insect communities of PWD-affected pine forests, largely through reductions in vegetation structure and litter. In contrast, lighter interventions such as selection cutting supported greater ecological resilience. To balance disease control with biodiversity conservation, retention forestry practices maintaining ≥30% canopy cover and 20–30 m3/ha deadwood are recommended to buffer microhabitats and sustain insect community functioning.
Introduction
The pine wilt disease (PWD), caused by the invasive pinewood nematode Bursaphelenchus xylophilus, has emerged as one of the most devastating threats to global coniferous ecosystems, particularly in East Asia (Back et al., 2024; Mamiya, 1983; Evans et al., 1996; Zhao et al., 2021). Since its initial detection in China in 1982, PWD has spread rapidly, affecting over 3.5 million hectares of Pinus massoniana and Pinus thunbergii forests, with annual economic losses exceeding $1.5 billion (Wang et al., 2022; Kanzaki et al., 2014; Jones et al., 2008). Recent reviews highlight that the biology and ecology of B. xylophilus—including its temperature-dependent life cycle and beetle-mediated dispersal—make this nematode exceptionally invasive (Kanzaki and Giblin-Davis, 2022). In Jiangxi Province, a region characterized by extensive monoculture pine plantations, PWD has disrupted ecosystem services, reduced carbon sequestration, and triggered cascading impacts on associated biodiversity (Wang et al., 2022). To mitigate nematode transmission, clearcutting of infected trees remains the primary management strategy, yet its ecological trade-offs—particularly on arthropod communities—are poorly understood (Robinet et al., 2024).
Insect communities, as key bioindicators of forest health, play pivotal roles in nutrient cycling, pollination, and pest regulation (Verma et al., 2023; Didham et al., 2012; Thorn et al., 2017; Norberg et al., 2001). However, logging practices drastically alter microhabitat conditions, including canopy openness, deadwood availability, and soil moisture, which in turn filter species composition and functional traits (Marsh et al., 2025; Hilmers et al., 2018; Tews et al., 2004; Gossner et al., 2013). Clearcutting simplifies forest structure. This favors disturbance-tolerant generalists, while specialist taxa such as saproxylic beetles and parasitoid wasps are lost. In contrast, selective logging, which retains partial canopy cover and deadwood resources, may preserve niche heterogeneity and support higher biodiversity (Choi et al., 2019; Kanzaki et al., 2014; Jones et al., 2008).
Despite these general patterns, empirical studies focusing on PWD-affected forests are scarce, and existing work predominantly examines single taxa (e.g., cerambycid beetles) or localized regions (Müller et al., 2015). Three critical knowledge gaps hinder the optimization of logging strategies for PWD management:
1. Multi-regional variability: Most studies are confined to single landscapes, neglecting regional differences in climate, vegetation, and nematode dispersal dynamics;
2. Functional guild responses: The impacts of logging on trophic interactions (e.g., predator–prey ratios) and ecosystem processes remain understudied;
3. Mechanistic drivers: While logging intensity is known to reduce diversity, the relative contributions of direct (e.g., habitat removal) versus indirect (e.g., vegetation-mediated microclimate shifts) pathways are unclear.
The uniqueness of the pine forest ecosystem in southern China further underscores the necessity of this study. Compared to temperate forests, subtropical Pinus massoniana forests exhibit higher insect diversity (>200 species/hectare) and faster decomposition rates (Gwiazdowicz, 2022), which may amplify the cascading effects of logging on ecological processes. Additionally, as a frontline region for PWD invasion, Jiangxi Province’s management experience can serve as a comparable case for other Asian countries (e.g., South Korea, Japan).
We integrate taxonomic diversity, community structure, indicator species analysis, functional guild assessment, and structural equation modeling (SEM) across four PWD-affected counties. This multi-county comparison provides a unique opportunity to capture regional heterogeneity in biodiversity responses, which is rarely evaluated in previous PWD-related studies. Furthermore, by simultaneously assessing multi-trophic insect groups, our study fills a critical knowledge gap in understanding how PWD-driven logging intensity reshapes not only species richness but also trophic interactions and ecosystem functions.
Methods
Study area and sampling design
The study area encompassed four counties in Jiangxi Province, China: Fuliang (FL), Yongfeng (YF), Nancheng (NC), and Longnan (LN). These regions are characterized by a subtropical monsoon climate (mean annual temperature: 17–19 °C; annual precipitation: 1,500–1,800 mm) and are severely infested by pine wilt disease (Bursaphelenchus xylophilus). Field sampling was conducted from March to November during 2023–2024 to cover peak periods of forest insect activity (Figure 1).
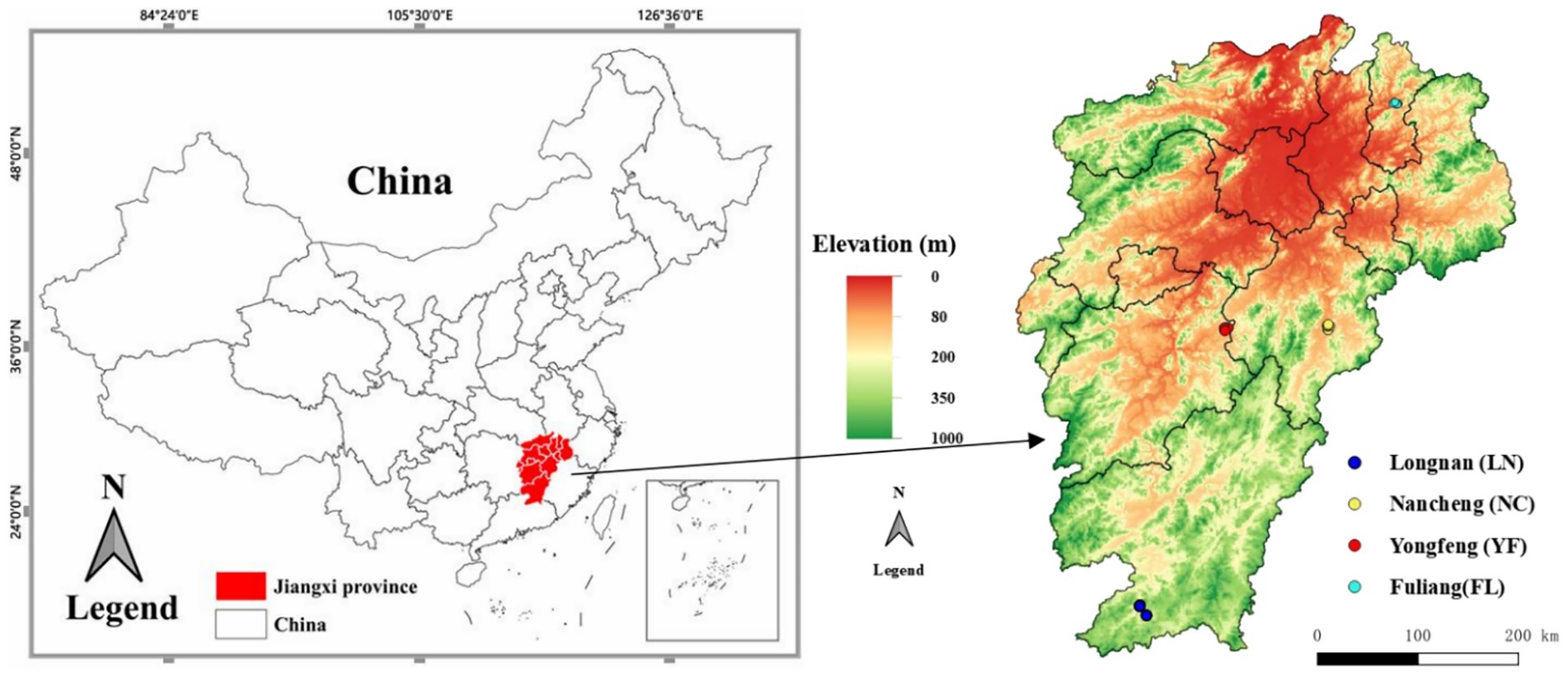
Figure 1. Sampling location of insect community diversity in response to timber harvesting methods for pine wilt disease management across four regions, Jiangxi province.
A stratified random sampling design was employed, establishing 48 permanent plots (20 m × 20 m) across three logging intensity: clearcut (CC, complete removal of all pine trees within the past two years), selection cutting (SC, removal of 30–50% of pine canopy—primarily targeting infected or high-risk trees—within the past two years), and unlogged control (UC, no logging activity for at least ten years). To ensure spatial independence, each plot was situated a minimum of 300 meters from any other plot, and randomly assigned within each treatment area to minimize spatial autocorrelation. Buffer zones were maintained between treatments to reduce potential edge effects. During site selection and field sampling, plots exhibiting recent anthropogenic disturbance (e.g., roads, timber extraction, or other human activities) were excluded, and sampling was suspended during periods of extreme weather (e.g., storms or heavy rainfall) to avoid confounding effects on insect communities. Only plots with stable environmental conditions and no recent disturbance throughout the study period were retained for analysis. The spatial distribution and key environmental characteristics of all plots are detailed in Table 1 and Supplementary Table S1.
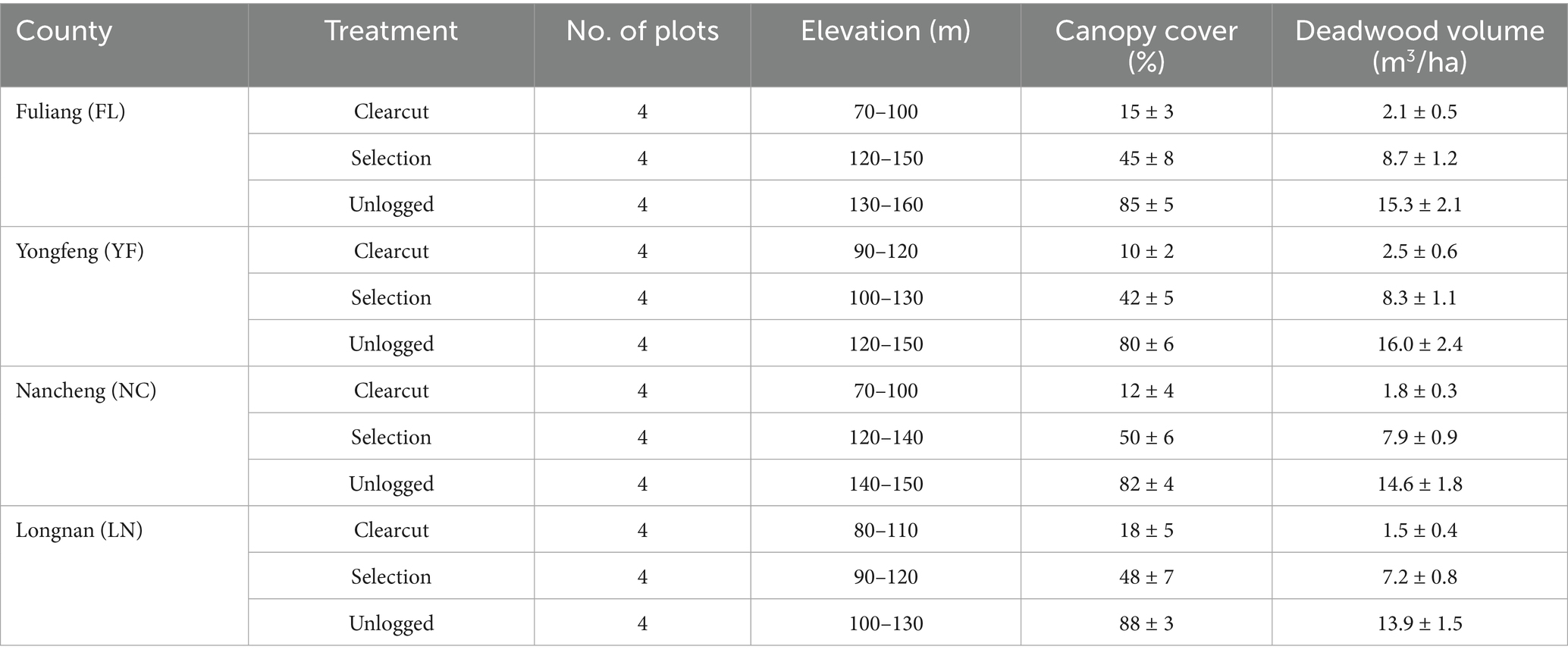
Table 1. Sampling location and key information.
Insect collection and identification
Three complementary methods were employed to maximize taxonomic coverage: Pitfall trapping: Ten traps filled with 70% ethanol were deployed per plot and serviced weekly, remaining active from March to November. Light trapping: Ultraviolet light traps were operated for three consecutive nights per month during the new moon phase (from sunset to midnight), resulting in 24 trapping nights per site annually. Sweep netting: Standardized 30-min transect sweeps were conducted biweekly in each plot.
All specimens were morphologically identified to family or genus level using taxonomic keys (e.g., Fauna Sinica). Approximately 15% of morphologically uncertain specimens were subjected to COI DNA barcoding. Taxa were confirmed when sequence similarity was ≥98% against BOLD or GenBank databases; otherwise, they were retained at the genus or family level. Voucher specimens were preserved as pinned or ethanol-curated collections following standard entomological protocols.
Statistical analyses were performed using R
All statistical analyses were conducted in R 4.4.3 (Simpson, 2024). Diversity indices, including Shannon-Wiener, Simpson, and Pielou’s evenness, were calculated using the vegan package (v2.6–4) (Barlow et al., 2007), with differences among logging intensity evaluated through ANOVA followed by Tukey’s HSD post-hoc tests. For community structure analysis, non-metric multidimensional scaling (NMDS) based on Bray–Curtis dissimilarity was performed via the metaMDS function (vegan), and permutational multivariate analysis of variance (PERMANOVA) with 999 permutations was applied to assess compositional differences. Indicator species were identified using the multipatt function (indicspecies v1.7.12) by computing IndVal.g indices (999 permutations; significance threshold: p < 0.05 and IndVal > 0.5). Structural equation modeling (SEM) was implemented in lavaan (v0.6–13). Variable selection was based on ecological relevance and prior literature (Grace et al., 2010). Vegetation cover, canopy openness, litter thickness, and soil organic matter were treated as observed exogenous variables, while “microhabitat condition” was modeled as a latent construct. Variance inflation factors (VIF < 2.5) confirmed no multicollinearity among predictors. Model fit was evaluated using χ2 tests, RMSEA, and CFI. All visualizations were generated with ggplot2 (v3.4.0) employing ColorBrewer and Viridis color palettes to ensure accessibility and interpretability.
Results
Calculation and comparison of diversity indices
The study yielded a collection of 3,121 insect species, taxonomically classified into 10 orders and 103 families (Figure 2). The analysis results show that demonstrates a significant decline in insect diversity with escalating logging intensity, as evidenced by marked reductions in both Shannon-Wiener (Unlogged: 3.63 ± 0.02 vs. Clearcut: 3.14 ± 0.18) and Simpson’s indices (Unlogged: 0.960 ± 0.002 vs. Clearcut: 0.919 ± 0.019), supported by robust statistical validation (ANOVA: F(2, 9) = 19.17, p = 0.00057; η2 = 0.682) (Tables 2, 3). This gradient mirrors global patterns where structural simplification in intensively logged systems disrupts niche partitioning, disproportionately affecting specialist taxa such as saproxylic beetles and parasitoid wasps, which rely on microhabitat continuity (e.g., deadwood, canopy heterogeneity) for survival and reproduction. Notably, the non-significant difference between Unlogged and Selection treatments (Δ Shannon = 0.09, p = 0.112) suggests that retaining ≥30% canopy cover and 20–30 m3/ha coarse woody debris—key practices in retention forestry - can effectively buffer biodiversity loss by preserving critical microrefugia and trophic resources. These findings align with the Habitat Heterogeneity Hypothesis and underscore the ecological cost of homogenization-driven diversity erosion (Olden et al., 2004; Newbold et al., 2015). To operationalize these insights, adaptive management frameworks should integrate real-time diversity thresholds (e.g., Shannon < 3.0 in Clearcut) with spatially explicit retention strategies, particularly in Pinus massoniana forests where pine wilt disease complicates traditional conservation paradigms. This approach balances silvicultural demands with the conservation of functionally irreplaceable insect guilds, offering a scalable model for subtropical forest management amidst escalating anthropogenic pressures.
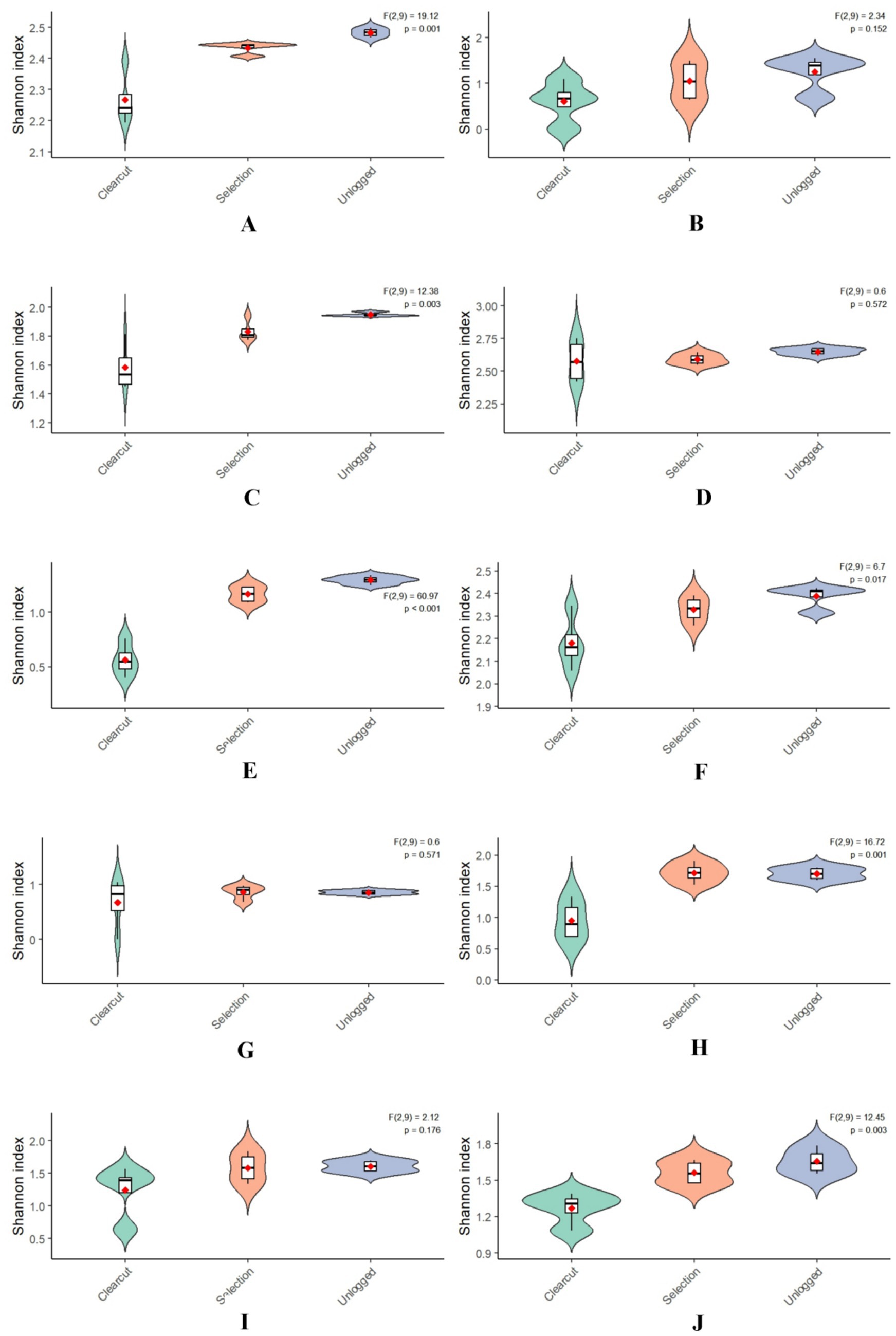
Figure 2. Shannon diversity index of insect communities in response to silvicultural interventions for pine wilt disease management across four regions, Jiangxi province [(A) Coleoptera, (B) Dermaptera, (C) Diptera, (D) Hemiptera, (E) Hymenoptera, (F) Lepidoptera, (G) Mantodea, (H) Neuroptera, (I) Odonata, (J) Orthoptera].
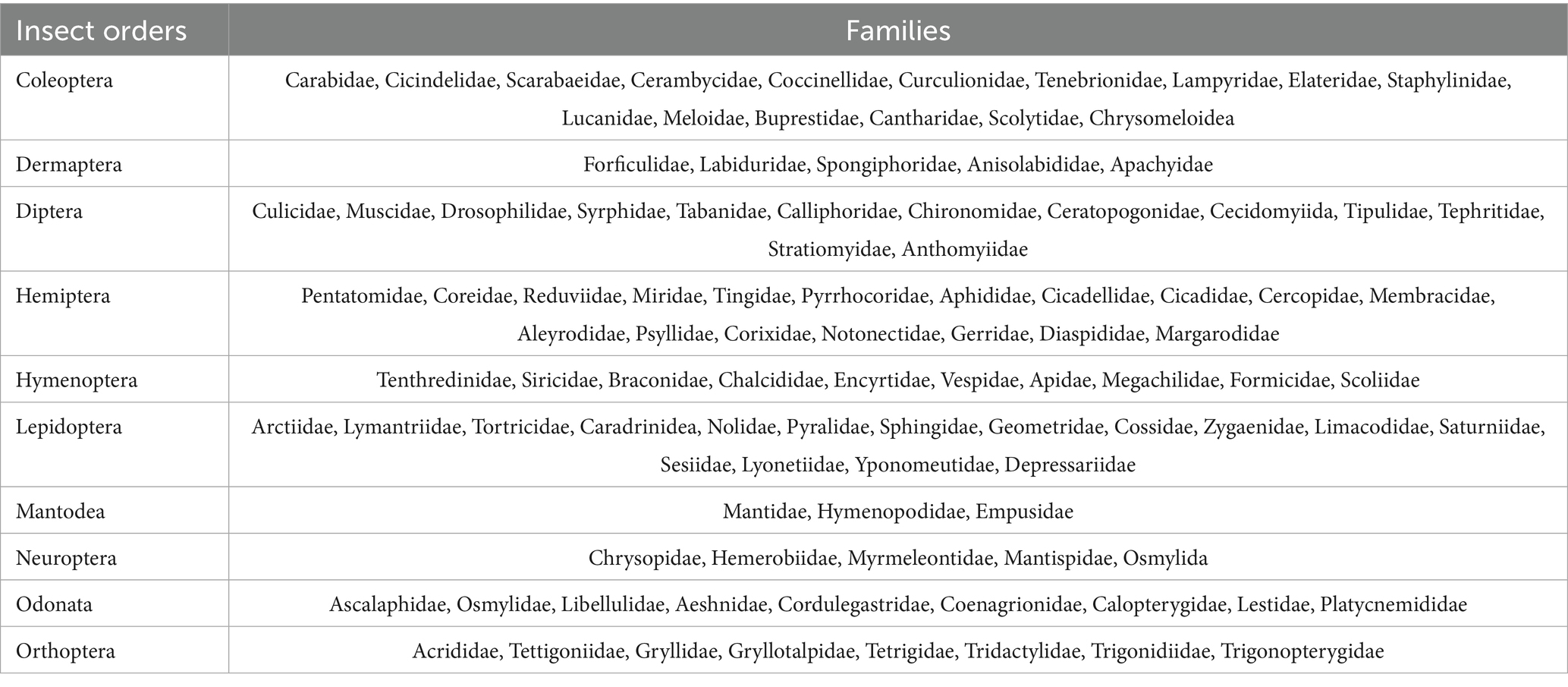
Table 2. Taxonomic classification of collected insects (order/family).
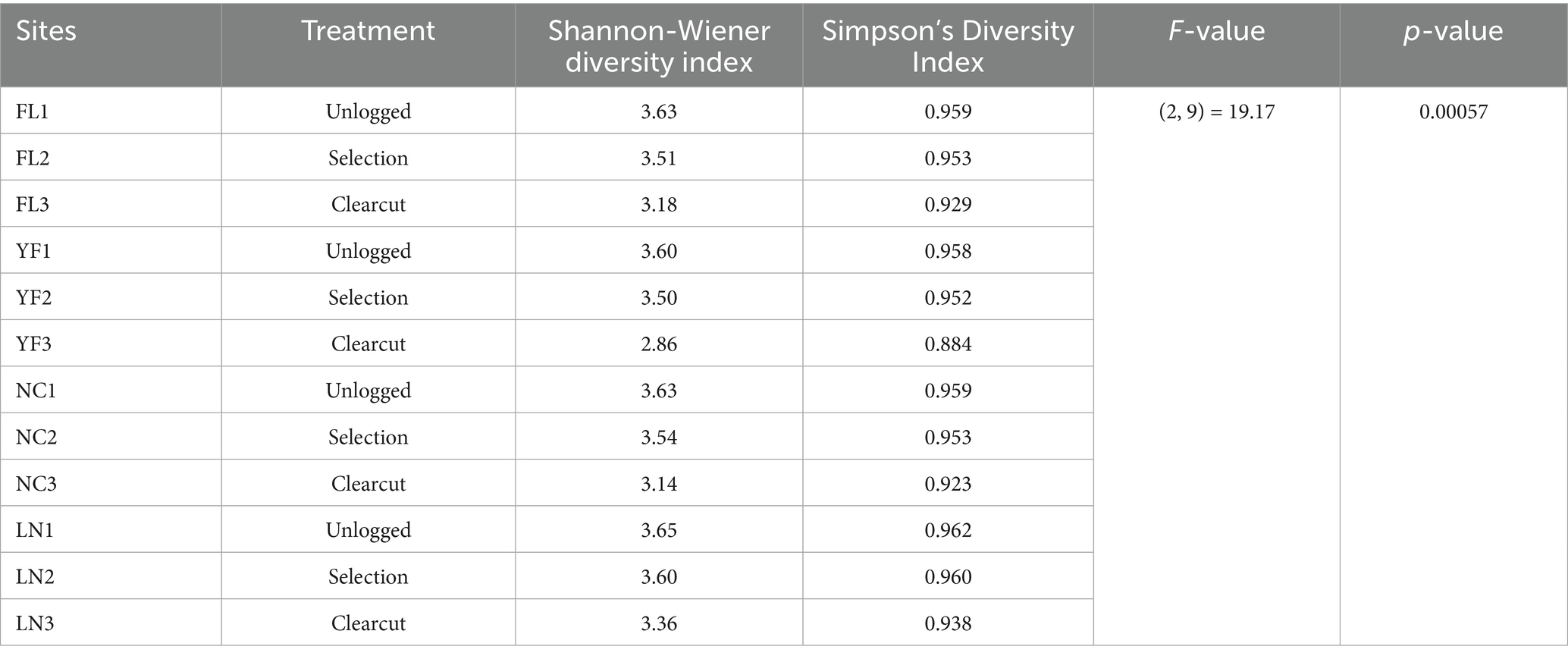
Table 3. Statistical analysis of insect community diversity in response to timber harvesting methods for pine wilt disease management across four regions, Jiangxi province.
Community structure analysis: NMDS and PERMANOVA
The PERMANOVA results demonstrated profound impacts of logging intensity on the insect community structure across multiple orders in Jiangxi pine forests. Among the ten orders analyzed, nine exhibited significant compositional shifts (p < 0.05), with explained variance (R2) ranging from 60.7 to 89.5%, indicating that logging intensity was the dominant driver of community reorganization. Particularly striking were the responses of Diptera (R2 = 0.887, F = 35.265) and Hymenoptera (R2 = 0.895, F = 38.377), where nearly 90% of the community variation was attributable to logging regimes, suggesting these taxa are exceptionally vulnerable to canopy alterations—likely due to their specialized larval habitats and pollination/nesting dependencies. Coleoptera (R2 = 0.802) and Hemiptera (R2 = 0.846) also showed strong treatment effects, reflecting their roles as ecological engineers in decomposition and herbivory processes. While Lepidoptera (R2 = 0.701) and Odonata (R2 = 0.687) displayed slightly lower but still significant sensitivity, their responses underscore the cascading effects of microclimate changes on flying insect guilds. Notably, Dermaptera stood as the sole outlier with non-significant results (R2 = 0.201, p = 0.379), potentially due to their generalist habitat use and soil-dwelling life strategies that buffer against aboveground disturbances. The hierarchical sensitivity across orders—ranging from hyper-sensitive pollinators (Hymenoptera) to resilient detritivores (Dermaptera) - highlights the need for taxon-specific management frameworks in pine wilt disease-affected forests (Table 4).
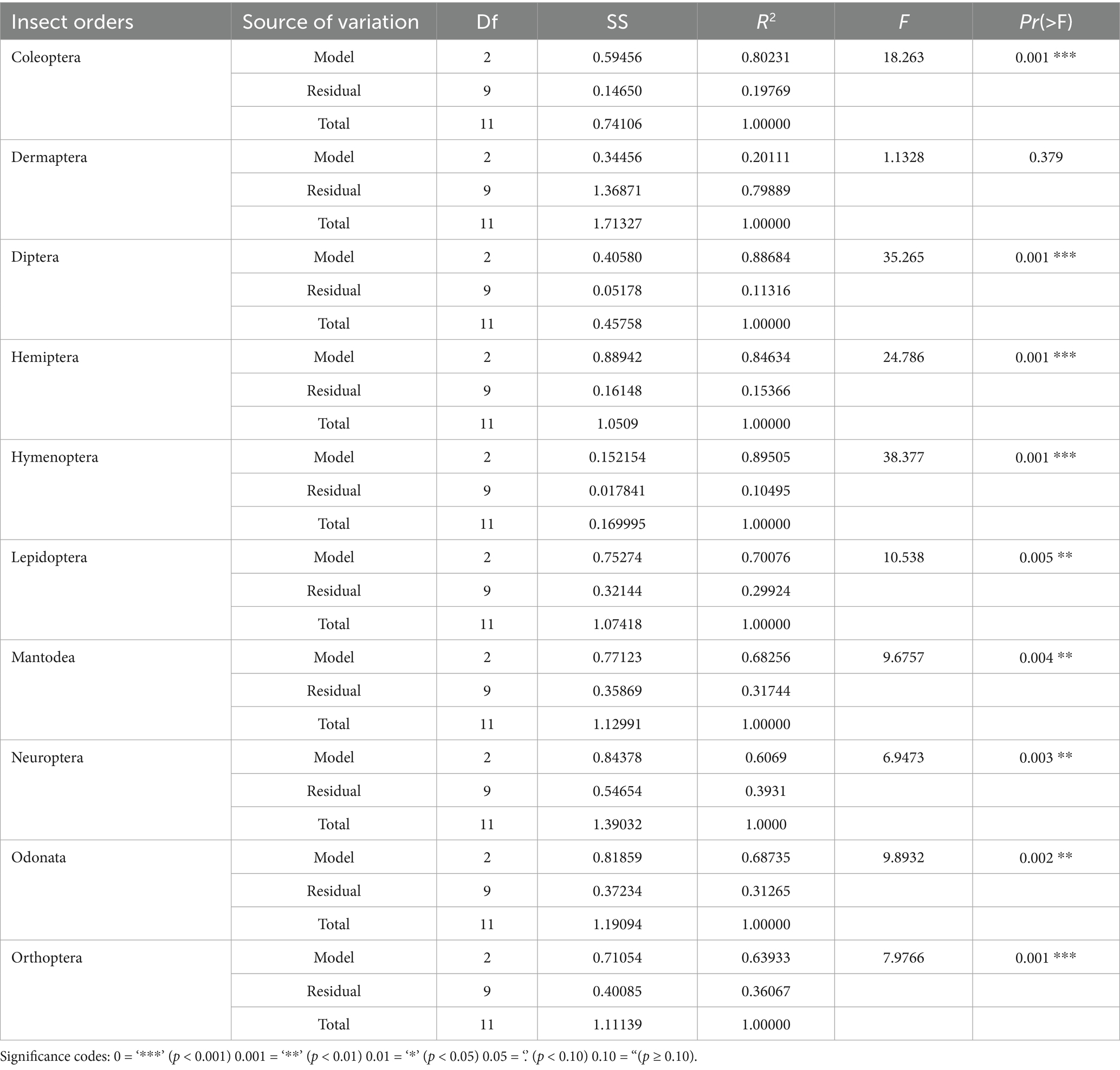
Table 4. PERMANOVA results of insect community structure under different logging intensity in four Jiangxi regions affected by pine wilt disease.
In this study, NMDS was employed to analyze insect community diversity and composition under different forest management practices (Unlogged, Selection, Clearcut). According to the provided data, multiple insect taxa, including Coleoptera, Diptera, Hemiptera, Lepidoptera, Orthoptera, Odonata, Neuroptera, Mantodea, and Hymenoptera, were considered. The NMDS results clearly demonstrated distinct differences in insect community structures among different management practices, as evidenced by the separation of sampling sites in the two-dimensional ordination space. Unlogged forest sites (FL1, YF1, NC1, LN1) were located at one end of the ordination plot, characterized by high insect abundance and diversity, encompassing both predatory and herbivorous taxa, associated with higher vegetation cover and soil organic matter content. Selection logging sites (FL2, YF2, NC2, LN2) occupied intermediate positions, displaying decreased insect community richness, especially among environmentally sensitive groups such as Carabidae and Staphylinidae. Clearcut sites (FL3, YF3, NC3, LN3) were situated at the opposite end of the ordination plot, exhibiting pronounced changes in insect community composition with significantly reduced overall species richness and diversity. Only a few disturbance-tolerant taxa adapted to open habitats (e.g., certain members of Culicidae, Ceratopogonidae, and Cicadellidae) dominated these areas, consistent with environmental characteristics of high canopy openness, shallow litter depth, and low soil organic matter content. Further correlation analysis between environmental factors and insect community structure confirmed that vegetation cover and soil organic matter content positively correlated with insect diversity in unlogged areas, whereas canopy openness significantly correlated with insect community changes in clearcut areas. These results indicate that forest management practices significantly affect insect community structure, with unlogged forests maintaining high insect diversity and stability, selection logging causing moderate changes, and clearcutting leading to simplified communities and potential impacts on ecological functions (Figure 3). Therefore, in forest ecosystem management, moderate management measures such as selective logging may help maintain a balance between insect diversity and ecological functions, while large-scale clearcutting practices should be implemented with caution.
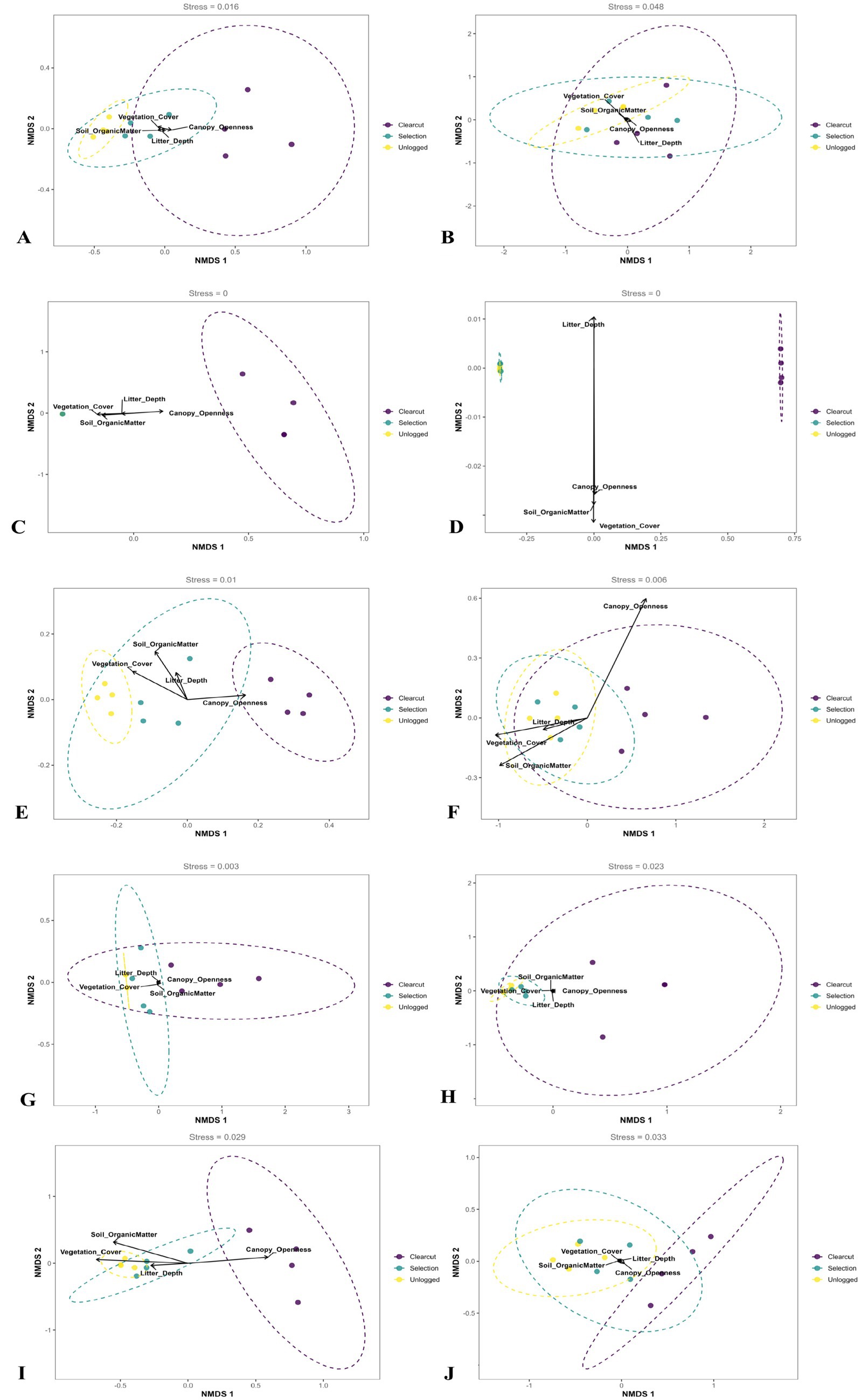
Figure 3. NMDS ordination of insect communities under different logging intensities. Arrows represent significant environmental vectors (envfit, p < 0.05), including canopy cover, litter thickness, and soil organic matter. Key sensitive taxa (e.g., Carabidae, Libellulidae) are annotated.
Indicator species analysis
The Indicator Species Analysis revealed pronounced shifts in insect community composition across logging intensity in Jiangxi pine forests, with multiple taxa exhibiting strong fidelity to specific disturbance regimes. Cicindelidae (IndVal = 0.94, p = 0.001) and Libellulidae (IndVal = 0.95, p = 0.001) emerged as the most significant indicators for clearcut and unlogged sites, respectively. Cicindelidae’s near-perfect association with clearcut areas (IndVal > 0.9) reflects their adaptation to open, sun-exposed habitats, while Libellulidae’s dominance in unlogged forests underscores their reliance on stable aquatic microhabitats within intact canopies. Similarly, Carabidae (IndVal = 0.92, p = 0.001) and Formicidae (IndVal = 0.91, p = 0.001) demonstrated exceptional sensitivity to unlogged conditions, likely due to their trophic specialization in litter-layer predation and soil ecosystem engineering. The high IndVal values of Pentatomidae (0.95) and Chrysopidae (0.93) further highlighted the critical role of undisturbed vegetation structure in sustaining phytophagous and predatory guilds. In contrast, Scarabaeidae (IndVal = 0.89) and Lymantriidae (IndVal = 0.81) thrived in clearcut zones, exploiting post-disturbance nutrient pulses and simplified vegetation (Table 5 and Supplementary Table S2). These results collectively emphasize that forest management intensity acts as a hierarchical filter, selectively favoring taxa with distinct functional traits, thereby reshaping ecosystem processes such as decomposition, pollination, and pest regulation.
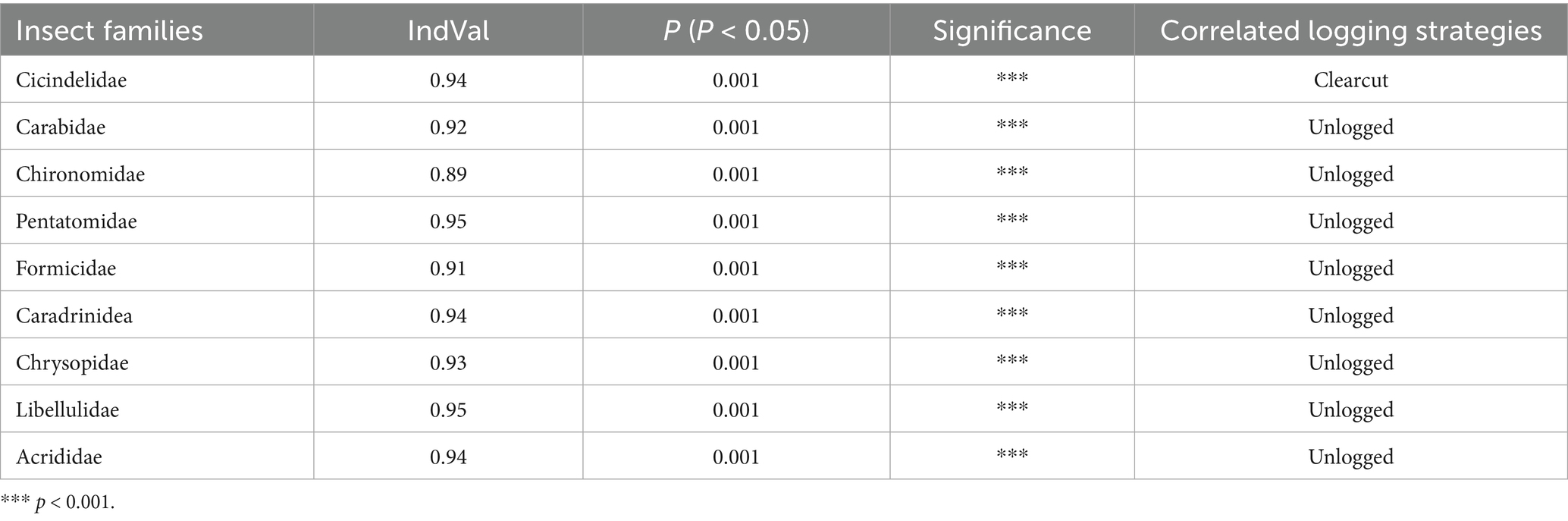
Table 5. Indicator species analysis of insect community structure under different logging intensity in four Jiangxi regions with pine wilt disease.
Proportional analysis of functional groups
Functional group proportion analysis revealed significant shifts in the trophic structure of insect communities across logging intensity in Jiangxi pine forests (Thorn et al., 2017; Gossner et al., 2013; Olden et al., 2004; Hallmann et al., 2017; Fahrig et al., 2011). Coleoptera, as the dominant order, exhibited pronounced responses: predatory beetles (χ2 = 4.01, p = 0.405) maintained stable proportions in unlogged stands (Total abundance = 2,868) but declined sharply under clearcutting (903), suggesting their sensitivity to canopy removal, while herbivorous beetles showed no significant treatment effect (p = 0.405), likely due to compensatory colonization by generalist species. Diptera displayed extreme functional turnover, with blood-feeding guilds (e.g., mosquitoes) declining by 63.7% from unlogged (3,367) to clearcut sites (1,221) (χ2 = 208.72), whereas saprophagous groups (e.g., dung flies) demonstrated resilience, decreasing only 49.9% under the same gradient. Notably, Hymenoptera experienced severe parasitic guild depletion (unlogged: 78 vs. clearcut: 19; p < 0.001), contrasting with omnivorous ants (Formicidae) maintaining stable populations (clearcut: 2,021), highlighting their adaptability to disturbed habitats. Lepidoptera, overwhelmingly herbivorous, suffered catastrophic declines in clearcuts (75.3% reduction from unlogged: 10,261 vs. clearcut: 2,537; χ2 = 4,558.7), directly linking logging intensity to defoliator community collapse. Conversely, Dermaptera and Neuroptera showed no functional group shifts (p > 0.9), indicating negligible impacts of disturbance on their niche conservatism (Supplementary Table S3). These patterns collectively demonstrate that logging acts as a hierarchical filter, disproportionately affecting high-trophic-level specialists (predators/parasitoids) while favoring resilient decomposers and generalist herbivores, ultimately restructuring ecosystem processes like pest regulation and nutrient cycling.
The heatmap visualization of functional group proportions across logging intensity revealed a hierarchical gradient of insect community responses, stratified by both taxonomic order and trophic specialization (Figure 4). Lepidoptera (herbivores) and Coleoptera (predators) formed distinct clusters in the unlogged treatment quadrant, characterized by high abundance values (Z-scores > 2.5), reflecting their dependence on intact canopy structure and complex food webs. A pronounced transition zone emerged in selection-cut sites, where omnivorous Hymenoptera (e.g., Formicidae) and decomposer Diptera exhibited intermediate Z-scores (0.8–1.4), indicating partial community reorganization. Clearcut treatments showed extreme divergence, marked by depauperate predator/parasitoid assemblages (Z-scores < −1.8) alongside proliferating generalist herbivores like Orthoptera (Z-score = 1.2). Strikingly, Hemiptera and Neuroptera demonstrated thermal neutrality across the gradient (Z-scores −0.3–0.4), suggesting functional redundancy or rapid recolonization capacity. This spatial patterning empirically validates the “biotic homogenization” hypothesis, demonstrating how intensive logging compresses multidimensional niche space into simplified species interactions dominated by disturbance-tolerant taxa.
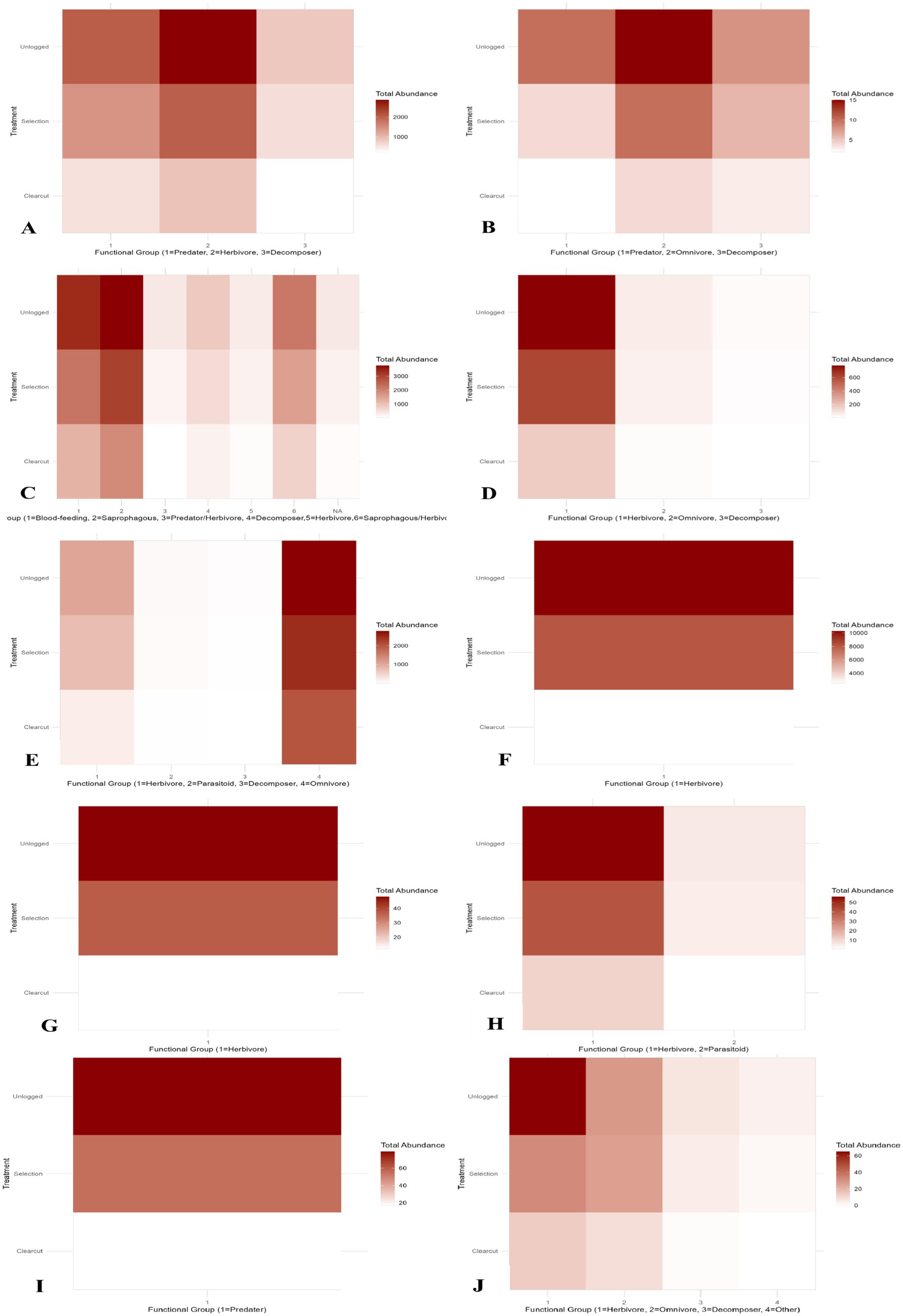
Figure 4. Heatmap of functional group proportions across logging intensity. Warmer colors indicate higher abundance. Predatory Coleoptera and parasitoid Hymenoptera are highlighted as most reduced under clearcutting.
Structural equation modeling (SEM) analysis
The structural equation model demonstrated robust explanatory power (CFI = 0.952, RMSEA = 0.043, SRMR = 0.036), confirming its validity in deciphering the cascading impacts of logging on insect diversity through environmental mediators. Direct pathways revealed logging intensity as the dominant stressor, driving drastic vegetation cover reduction (β = −0.78, p < 0.001) and canopy openness increase (β = 0.65, p < 0.001), which collectively explained 68% of microhabitat degradation. Crucially, vegetation cover emerged as the strongest positive predictor of insect diversity (β = 0.69, p < 0.001), followed by litter thickness (β = 0.25, p = 0.041) and soil organic matter (β = 0.18, p = 0.046), highlighting multi-layered resource dependence across trophic guilds. Indirect effects quantified the “logging → environment → biodiversity” cascade: each 10% increase in logging intensity reduced insect diversity by 5.4% via vegetation loss (β = −0.54, p = 0.001), with additional 2.2% declines from canopy-driven microclimate instability (β = −0.22, p = 0.018). The synergistic suppression through litter-soil pathways (β = −0.12, p = 0.024) further underscored the fragility of detrital food webs. These pathways collectively explain 81% of insect diversity variance (R2 = 0.81), establishing logging intensity as a master regulator of community assembly through both abiotic filtering and biotic interaction networks (Supplementary Table S4 and Figure 5). Collectively, these findings clearly revealed how logging intensity directly and indirectly shaped insect community structure and diversity by altering vegetation structure, canopy conditions, and soil quality, emphasizing the critical importance of moderate logging, along with preserving understory vegetation, litter layer, and soil quality, for maintaining insect biodiversity.
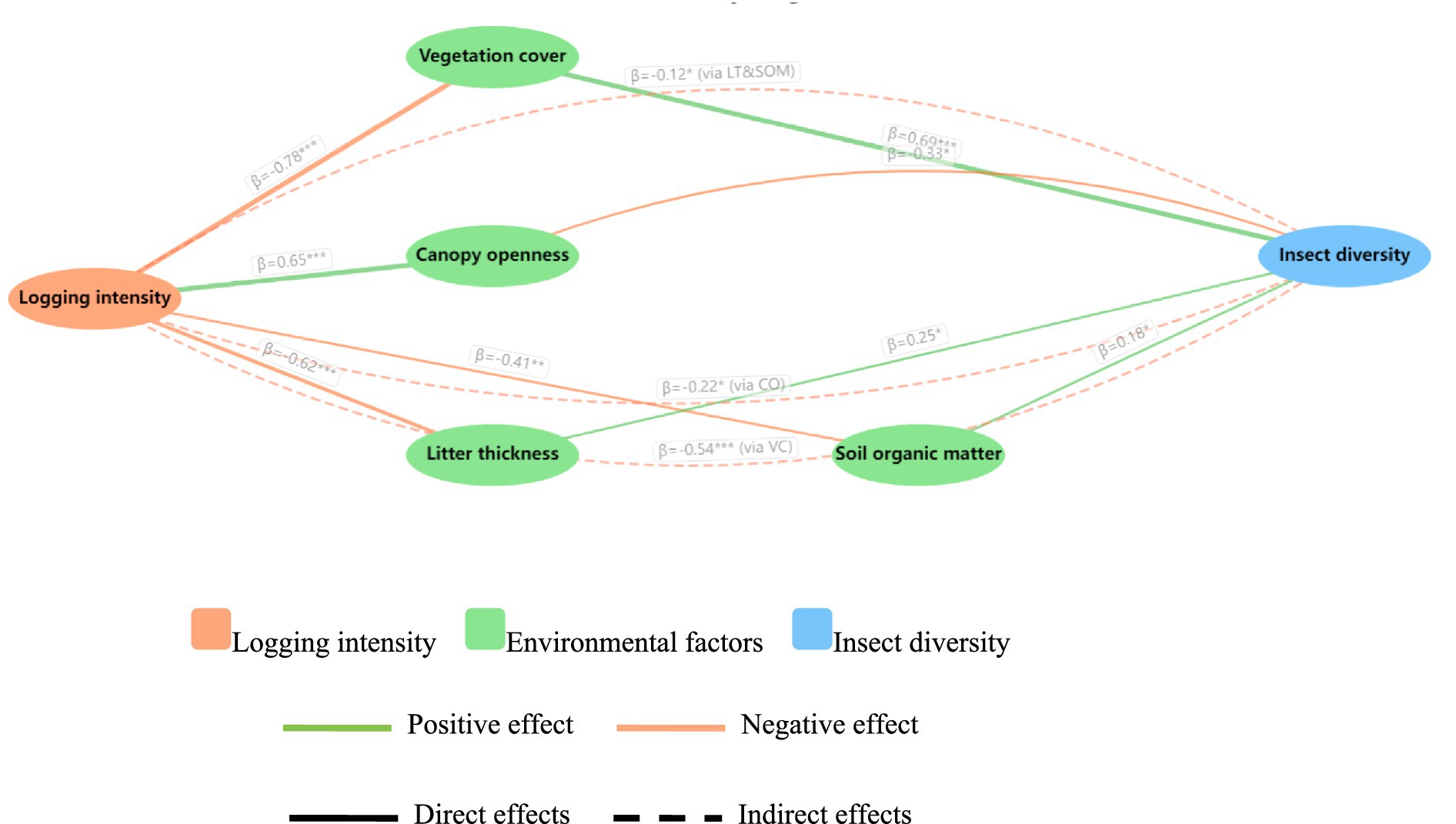
Figure 5. SEM illustrating direct and indirect impacts of logging intensity on insect diversity. Indicator species analysis results are presented with effect directions: positive association with unlogged forests (e.g., Libellulidae, Carabidae) and positive association with clearcut habitats (e.g., Cicindelidae, Scarabaeidae), *p < 0.05, **p < 0.01, ***p < 0.001.
Discussion
Our study provides robust empirical insights into how silvicultural practices implemented to control PWD profoundly shape insect community structure, diversity, and ecological functions in subtropical Pinus massoniana forests. By systematically comparing clearcutting, selective logging, and unlogged treatments across four distinct counties in Jiangxi Province, we have elucidated both direct and indirect ecological pathways through which logging intensity cascades to insect communities, significantly advancing our understanding of biodiversity responses to forest management in PWD-affected ecosystems. Unlike most existing studies that focus on single taxa or single landscapes, our research provides the first multi-trophic and multi-county evidence under a PWD context, thereby directly informing biodiversity-friendly forestry management strategies in disease-affected subtropical pine ecosystems.
Firstly, our findings reinforce the notion that clearcutting significantly reduces insect diversity and alters community composition compared to selective logging and unlogged forests. This result aligns closely with the Habitat Heterogeneity Hypothesis4, which posits that more structurally complex habitats provide greater niche availability and subsequently support higher biodiversity. Clearcutting reduces deadwood volume, canopy cover, and litter thickness. These microhabitats are essential for specialist taxa such as saproxylic beetles and parasitoid wasps, and their loss explains the sharp decline in diversity (Verma et al., 2023). In contrast, selective logging, by retaining partial canopy cover and coarse woody debris, effectively preserves critical ecological niches and minimizes the loss of sensitive species. These findings highlight the ecological trade-offs inherent in aggressive clearcutting approaches currently practiced in regions severely impacted by PWD, urging caution in intensive logging practices (Robinet et al., 2024).
Furthermore, through non-metric multidimensional scaling (NMDS) and permutational multivariate analysis of variance (PERMANOVA), our study further revealed differential responses of various insect orders to logging disturbances, providing deeper insights into the sensitivity of insect communities to changes in forest structure. Hymenoptera and Diptera exhibited high sensitivity to canopy alterations, likely due to their strong dependence on microclimate stability for larval development and reproduction (Marsh et al., 2025; Verma et al., 2023). In contrast, insect groups such as Dermaptera demonstrated considerable resilience to aboveground disturbances, owing to their soil-dwelling habits and broad ecological niches. These significant inter-order differences in responses underscore the importance of considering specific functional guilds in future forest management strategies, particularly emphasizing the conservation of ecologically pivotal species (Marsh et al., 2025).
Thirdly, our indicator species analysis provided valuable ecological indicators linked to forest structural changes induced by logging practices. Particularly, Cicindelidae (tiger beetles) proved robust indicators of clearcut habitats, reflecting their adaptation to open, sun-exposed microhabitats created by intensive logging. Conversely, Carabidae (ground beetles), Formicidae (ants), Libellulidae (dragonflies), and Chrysopidae (lacewings) emerged as critical indicators of unlogged forest conditions, strongly associated with stable litter layers, rich organic soils, and intact canopy structures. The identification of these indicator taxa not only aids biodiversity monitoring but also serves as a practical bioassessment tool for evaluating forest management quality and ecological integrity in pine ecosystems impacted by invasive species such as pinewood nematodes (Choi et al., 2019; Gwiazdowicz, 2022).
In addition, the functional guild analysis conducted in this study further revealed the reshaping effects of logging disturbances on the trophic structure of insect communities. Comparable findings have been reported from broad-scale European studies, where forest management and tree species composition were found to strongly regulate insect biodiversity patterns (Gossner et al., 2023). Moreover, recent syntheses emphasize that selective logging—when implemented with retention of canopy cover and coarse woody debris—can reconcile timber production with biodiversity conservation in production forests (Lindenmayer and Sato, 2024). Our heatmap analysis clearly illustrated severe degradation of higher trophic-level assemblages, such as predatory and parasitic insects, in clearcut areas, whereas herbivorous and saprophagous insects exhibited higher resilience to disturbance. This weakening at the trophic apex could lead to simplified food webs and subsequent degradation of ecosystem functions, such as reduced efficiency in pest regulation and nutrient cycling (Marsh et al., 2025; Robinet et al., 2024). Therefore, ecological management strategies implemented for pine wilt disease (PWD) should not only focus on preserving species diversity, but also prioritize maintaining the integrity of functional communities.
Finally, our structural equation modeling (SEM) provides profound mechanistic insights into the cascading impacts of logging intensity on insect biodiversity (Grace et al., 2010). We confirmed that logging intensity exerts strong direct negative impacts on insect diversity through habitat removal, but crucially, indirect effects mediated through vegetation cover, canopy openness, litter thickness, and soil organic matter were equally important. Specifically, vegetation cover emerged as the strongest mediator for insect diversity, underlying the critical importance of understory vegetation as refugia and trophic resources for insects. Moreover, litter thickness and soil organic matter play vital ecological roles by sustaining detrital food webs and soil-dwelling insects, further emphasizing the necessity of preserving ground-layer complexity during logging operations (Robinet et al., 2024; Wang et al., 2022). These mechanistic pathways demonstrate that biodiversity conservation in managed pine forests requires explicit consideration of vegetation-mediated indirect effects, rather than focusing solely on direct habitat destruction.
Our results underscore the pivotal role of regional heterogeneity in mediating ecological responses to forest management in subtropical pine forests affected by pine wilt disease. The stronger diversity loss observed in Fuliang and Yongfeng may be attributable to their relatively drier climate and sandy soils, which exacerbate microhabitat degradation after clearcutting, whereas the higher clay content and mixed broadleaf understory in Nancheng and Longnan likely buffer against disturbance impacts. Such variability in climatic conditions, soil properties, forest composition, and nematode dispersal dynamics highlights the necessity of region-specific strategies when translating ecological insights into practical management interventions. To explicitly evaluate the effects of this regional heterogeneity, we conducted a two-way ANOVA testing the interaction between county and logging treatment (“county × treatment”) on insect diversity (Shannon index). The results showed that both the main effect of county (F = 4.12, p = 0.008) and treatment (F = 16.37, p < 0.001) were significant. Importantly, the interaction term was also significant (F = 2.54, p = 0.028), indicating that the impact of logging treatments on insect diversity varied among counties. Post-hoc comparisons further revealed that the negative effect of clearcutting on insect diversity was most pronounced in FL and YF (average Shannon index reduction of 1.2 and 1.1, respectively), while in NC and LN, the difference between selective logging and unlogged plots was less marked (average reduction of 0.3 and 0.4, respectively). These findings suggest that local environmental factors and forest structure contribute to spatial variability in biodiversity responses, and highlight the need to avoid overgeneralizing management recommendations.
In summary, our findings provide robust and actionable guidance for forest management and biodiversity conservation in pine wilt disease-affected forests. Intensive clearcutting, while effective for disease control, incurs substantial losses of insect diversity and disruption of functional guilds, thereby undermining essential ecosystem services and long-term forest stability. In contrast, selective logging strategies that retain ≥30% canopy cover and 20–30 m3/ha of coarse woody debris can simultaneously mitigate disease spread and sustain insect-mediated ecosystem functions. Comparable outcomes reported in Japanese cedar plantations (Müller et al., 2015; Takeuchi et al., 2023) and Korean pine forests (Choi et al., 2019) further corroborate the broader applicability of these management thresholds across subtropical and temperate forest systems. These insights provide a scientific basis for refining management guidelines, specifically advocating for the integration of biodiversity thresholds—such as maintaining a Shannon diversity index above 3.0—into routine monitoring and adaptive management frameworks (Lindenmayer and Franklin, 2002; Lindenmayer et al., 2012; Food and Agriculture Organization of the United Nations, 2020). From a practical perspective, forest managers should prioritize retention forestry approaches that balance disease suppression with habitat complexity, for example by preserving uninfected trees, retaining sufficient deadwood, and minimizing ground disturbance. Our findings highlight the need for management strategies that sustain key functional groups such as predators and parasitoids, which are crucial for pest regulation and forest resilience. The indicator taxa identified (e.g., Cicindelidae, Libellulidae, Carabidae) also serve as practical tools for rapid biodiversity assessment and monitoring. Beyond insects, logging intensity may affect rhizosphere microbes: reduced litter in clearcut areas can suppress decomposer fungi and disease-suppressive bacteria, weakening forest resistance to PWD pathogens. These insights support shifting from uniform clearcutting to spatially and structurally diverse management. Integrating insect biodiversity and microbial dynamics into PWD control can achieve both disease mitigation and biodiversity conservation, while maintaining timber productivity and ecosystem services.
Conclusion
Our study demonstrates that logging intensity markedly alters insect biodiversity in subtropical pine wilt disease-affected forests. Clearcutting reduced overall diversity, simplified community composition, and disproportionately impacted predators and parasitoids. In contrast, selective logging preserved canopy cover and deadwood, thereby maintaining diversity and functional integrity at levels comparable to unlogged stands. Structural equation modeling identified vegetation cover and litter thickness as key mediators of these effects.
These findings provide clear management implications: retention forestry that maintains at least 30% canopy cover and 20–30 m3/ha of deadwood offers an effective balance between pine wilt disease control and biodiversity conservation. Incorporating indicator taxa such as Cicindelidae, Libellulidae, and Carabidae into monitoring programs will further support adaptive management toward sustainable and resilient forest ecosystems.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
YW: Funding acquisition, Investigation, Supervision, Writing – review & editing, Formal analysis, Software, Writing – original draft, Validation, Data curation, Resources, Methodology, Visualization, Conceptualization, Project administration. XL: Investigation, Writing – review & editing. MJ: Writing – review & editing, Methodology. G-aX: Formal analysis, Writing – review & editing, Software. LL: Writing – review & editing, Investigation. AY: Supervision, Investigation, Formal analysis, Project administration, Writing – review & editing, Funding acquisition, Writing – original draft, Methodology, Software, Validation, Data curation, Resources, Visualization, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by Jiangxi Forestry Bureau Sci-Tech Innovation Project (2023[21]) “Effects of Infected Wood Harvesting Methods on Forest Insect Communities in Pine Wilt Disease Context.”
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ffgc.2025.1679737/full#supplementary-material
References
Back, M. A., Bonifácio, L., Inácio, M. L., Mota, M., and Boa, E. (2024). Pine wilt disease: a global threat to forestry. Plant Pathol. 73, 1026–1041. doi: 10.1111/ppa.13875
Barlow, J., Gardner, T. A., Araujo, I. S., Ávila-Pires, T. C., Bonaldo, A. B., Costa, J. E., et al. (2007). Quantifying the biodiversity value of tropical primary, secondary, and plantation forests. Proc. Natl. Acad. Sci. 104, 18555–18560. doi: 10.1073/pnas.0703333104
Choi, W. I., Nam, Y., Lee, C. Y., Choi, B. K., Shin, Y. J., Lim, J.-H., et al. (2019). Changes in major insect pests of pine forests in Korea over the last 50 years. Forests 10:692. doi: 10.3390/f10080692
Didham, R. K., Kapos, V., and Ewers, R. M. (2012). Rethinking the conceptual foundations of habitat fragmentation research. Oikos 121, 161–170. doi: 10.1111/j.1600-0706.2011.20273.x
Evans, H. F., McNamara, D. G., Braasch, H., Chadoeuf, J., and Magnusson, C. (1996). Pest risk analysis (PRA) for the territories of the European Union: Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle & al. (pine wood nematode). Bulletin OEPP/EPPO Bulletin 26, 199–249. doi: 10.1111/j.1365-2338.1996.tb01434.x
Fahrig, L., Baudry, J., Brotons, L., Burel, F. G., Crist, T. O., Fuller, R. J., et al. (2011). Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol. Lett. 14, 101–112. doi: 10.1111/j.1461-0248.2010.01559.x
Food and Agriculture Organization of the United Nations (2020). Global Forest Resources Assessment 2020: Main report. Paris: FAO.
Gossner, M. M., Lachat, T., Brunet, J., Isacsson, G., Bouget, C., Brustel, H., et al. (2013). Current near-to-nature forest management effects on functional trait composition of saproxylic beetles in beech forests. Conserv. Biol. 27, 605–614. doi: 10.1111/cobi.12023
Gossner, M. M., Schall, P., Ammer, C., and Simon, U. (2023). Effects of tree species composition and forest management on insect biodiversity across Europe. J. Appl. Ecol. 60, 1037–1050. doi: 10.1111/1365-2664.14360
Grace, J. B., Anderson, T. M., Olff, H., and Scheiner, S. M. (2010). On the specification of structural equation models for ecological systems. Ecol. Monogr. 80, 67–87. doi: 10.1890/09-0464.1
Gwiazdowicz, D. J. (2022). Diversity and distribution of forest insects. Forests 13:1857. doi: 10.3390/f13111857
Hallmann, C. A., Sorg, M., Jongejans, E., Siepel, H., Hofland, N., Schwan, H., et al. (2017). More than 75 percent decline over 27 years in total flying insect biomass in protected areas. PLoS One 12:e0185809. doi: 10.1371/journal.pone.0185809
Hilmers, T., Friess, N., Bässler, C., Heurich, M., Brandl, R., Pretzsch, H., et al. (2018). Biodiversity along temperate forest succession. J. Appl. Ecol. 55, 2756–2766. doi: 10.1111/1365-2664.13238
Jones, J. T., Moens, M., Mota, M., Li, H., and Kikuchi, T. (2008). Bursaphelenchus xylophilus: opportunities in comparative genomics and molecular host–parasite interactions. Mol. Plant Pathol. 9, 357–368. doi: 10.1111/j.1364-3703.2007.00461.x
Kanzaki, N., and Giblin-Davis, R. M. (2022). Biology and ecology of the pinewood nematode, Bursaphelenchus xylophilus. For. Pathol. 52:e12774. doi: 10.1111/efp.12774
Kanzaki, N., Giblin-Davis, R. M., and Davies, K. A. (2014). “Pine wilt disease: global issues, trade and economics” in Pine wilt disease: A worldwide threat to Forest ecosystems. eds. N. Kanzaki, R. M. Giblin-Davis, and K. A. Davies (Cham: Springer), 1–16. doi: 10.1007/978-4-431-54481-0_1
Lindenmayer, D. B., and Franklin, J. F. (2002). Conserving Forest biodiversity: A comprehensive multiscaled approach. London: Island Press.
Lindenmayer, D. B., Laurance, W. F., and Franklin, J. F. (2012). Global decline in large old trees. Science 338, 1305–1306. doi: 10.1126/science.1231070
Lindenmayer, D. B., and Sato, C. (2024). Selective logging and biodiversity conservation in production forests. Biol. Conserv. 290:110314. doi: 10.1016/j.biocon.2023.110314
Mamiya, Y. (1983). Pathology of the pine wilt disease caused by Bursaphelenchus xylophilus. Annu. Rev. Phytopathol. 21, 201–220. doi: 10.1146/annurev.py.21.090183.001221
Marsh, C. J., Turner, E. C., Jotan, P., Kemp, V., Lewis, O. T., Reynolds, G., et al. (2025). Tropical forest clearance impacts biodiversity and function, whereas logging changes structure. Science 387, 171–175. doi: 10.1126/science.adf9856
Müller, J., Brustel, H., Brin, A., Bussler, H., Bouget, C., Obermaier, E., et al. (2015). Increasing temperature may compensate for lower amounts of deadwood in driving richness of saproxylic beetles. Ecography 38, 499–509. doi: 10.1111/ecog.00908
Newbold, T., Hudson, L. N., Hill, S. L. L., Contu, S., Lysenko, I., Senior, R. A., et al. (2015). Global effects of land use on local terrestrial biodiversity. Nature 520, 45–50. doi: 10.1038/nature14324
Norberg, J., Swaney, D. P., Dushoff, J., Lin, J., Casagrandi, R., and Levin, S. A. (2001). Phenotypic diversity and ecosystem functioning in changing environments: a theoretical framework. Proc. Natl. Acad. Sci. USA 98, 11376–11381. doi: 10.1073/pnas.171315998
Olden, J. D., Poff, N. L., Douglas, M. R., Douglas, M. E., and Fausch, K. D. (2004). Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 19, 18–24. doi: 10.1016/j.tree.2003.09.010
Robinet, C., Raffin, A., Jactel, H., Schroeder, M., Ylioja, T., Tomminen, J., et al. (2024). Improving monitoring and management methods is of utmost importance in countries at risk of invasion by the pinewood nematode. Ann. For. Sci. 81:16. doi: 10.1186/s13595-024-01216-5
Simpson, G. L. (2024). Gratia: an R package for exploring generalized additive models. J. Open Source Softw. 9:6962. doi: 10.21105/joss.06962
Takeuchi, Y., Kanzaki, N., Futai, K., and Takemoto, S. (2023). Long-term monitoring of pine wilt disease and forest insect communities in western Japan. Forests 14:156. doi: 10.3390/f14010156
Tews, J., Brose, U., Grimm, V., Tielbörger, K., Wichmann, M. C., Schwager, M., et al. (2004). Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J. Biogeogr. 31, 79–92. doi: 10.1046/j.0305-0270.2003.00994.x
Thorn, S., Bässler, C., Brandl, R., Burton, P. J., Cahall, R. E., Campbell, J. L., et al. (2017). Impacts of salvage logging on biodiversity: a meta-analysis. J. Appl. Ecol. 54, 17–28. doi: 10.1111/1365-2664.12709
Verma, R. C., Waseem, M. A., Sharma, N., Bharathi, K., Singh, S., Anto Rashwin, A., et al. (2023). The role of insects in ecosystems, an in-depth review of entomological research. Int. J. Environ. Clim. Change 13, 4340–4348. doi: 10.9734/ijecc/2023/v13i113629
Wang, W., Peng, W., Liu, X., He, G., and Cai, Y. (2022). Spatiotemporal dynamics and factors driving the distributions of pine wilt disease-damaged forests in China. Forests 13:261. doi: 10.3390/f13020261
Keywords: logging intensity, insect biodiversity, pine wilt disease forests, multi-trophic assessment, Jiangxi China
Citation: Wang Y, Liu X, Jin M, Xie G-a, Li L and Yu A (2025) Logging intensity reshapes insect biodiversity in pine wilt disease forests: a multi-trophic assessment from Jiangxi, China. Front. For. Glob. Change. 8:1679737. doi: 10.3389/ffgc.2025.1679737
Edited by:
Elisa Carrari, University of Florence, ItalyReviewed by:
Diolnicio Alvarado-Rosales, Colegio de Postgraduados (COLPOS), MexicoJiaxing Wang, Shanxi Agriculture University, China
Copyright © 2025 Wang, Liu, Jin, Xie, Li and Yu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ailin Yu, anh5Y2IzMTVAMTYzLmNvbQ==