Abstract
Sustainable agriculture has become a global priority in response to increasing food demand and the challenges confronting agricultural production, such as biotic and abiotic stresses. In this review, we delve into the role of plant diversity in mitigating these stressors within tomato cultivation. Our investigation reveals that the most extensively studied companion species are Vicia villosa Roth, Coriandrum sativum L., and Allium cepa L., while the primary stressors under scrutiny include nutrient deficiencies, aerial pests, and soil-borne pathogenic diseases. Regarding nutrient deficiencies, the cover crop system has demonstrated its capacity to provide essential nutrients directly and indirectly to plants. In addressing aerial pests and pathogens, all cultivation systems exhibit contributions. Finally, we assert that incorporating plant diversity into agroecosystems can effectively counteract various types of stressors. These benefits align with the application of agroecological principles and the development of sustainable agroecosystems. Further assessments of the effects of additional companion plant species are imperative. This should encompass the identification of their distribution, optimal plant quantities, and cultivation systems that enhance their benefits. Ultimately, these evaluations will aid in the formulation of comprehensive guidelines to facilitate the selection and utilization of plant diversity for long-term sustainability.
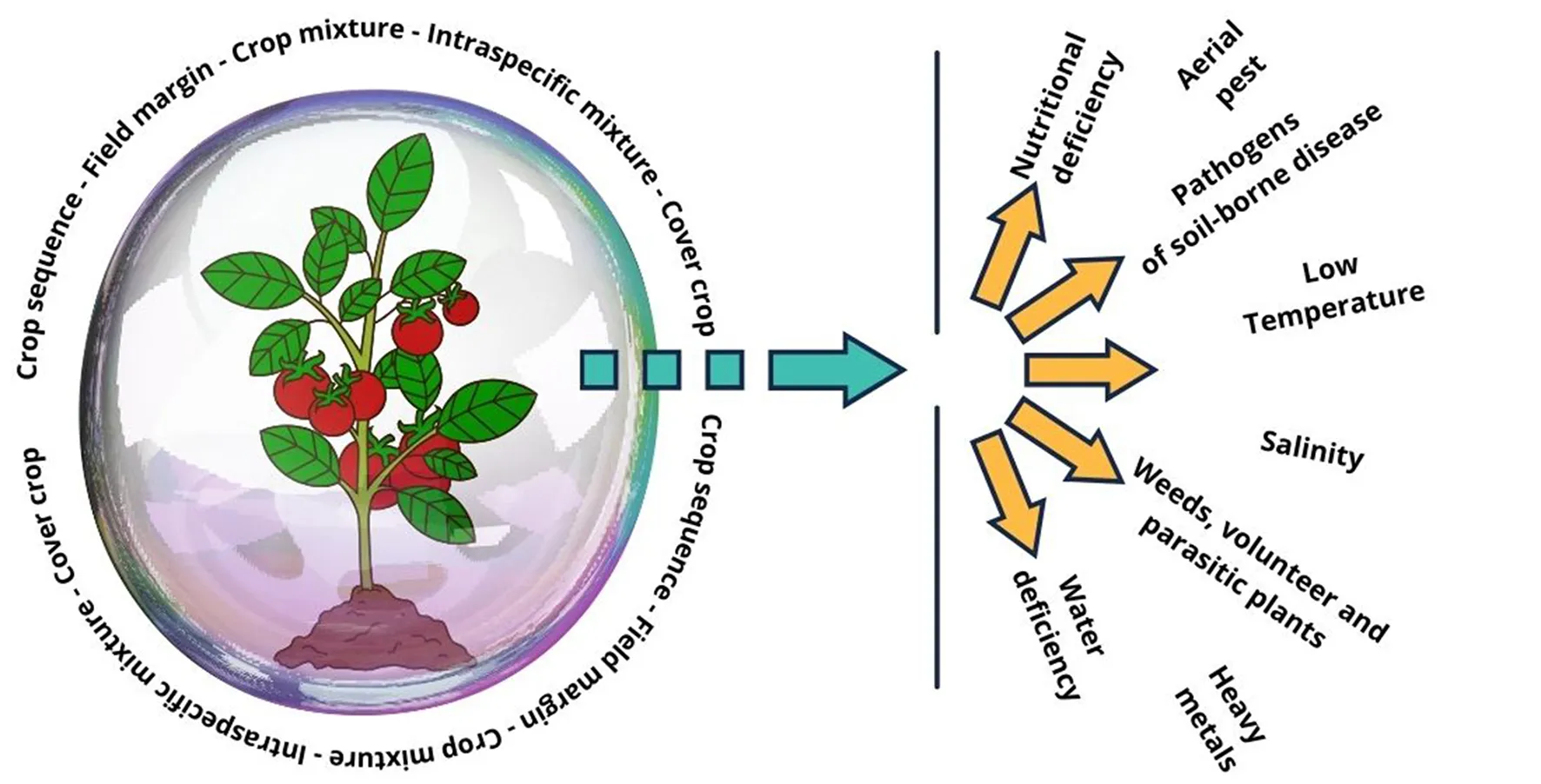
1 Introduction
Agriculture, one of the foundational pillars of human civilization, confronts unparalleled challenges in the twenty first century (Wilson and Lovell, 2016; Sumberg and Giller, 2022). The achievement of successful food production hinges significantly on the growth and development of plants, a process intricately connected to the presence of stress, whether of biotic or abiotic origins (Enebe and Babalola, 2018). Abiotic stress, encompassing salinity, drought, extreme temperatures, and exposure to toxic metals, in conjunction with biotic stress arising from attacks by herbivorous insects and pathogens, presents a determining factor in crop productivity (Benavidez et al., 2002; Song et al., 2015; Zhu, 2016; Inbaraj, 2021). In recent decades, climate change and pollution of water and soil have led to stress conditions appearing more frequently in crops (Volpe et al., 2018; Nawaz et al., 2020; Peck and Mittler, 2020); these factors are also closely related to intensive agricultural practices like continuous monoculture and the excessive application of fertilizers and pesticides (Corrado et al., 2019).
In this critical context, the imperative for an extensive review becomes evident—not only to illuminate how stress impacts crops but also to explore strategies for mitigating its effects. From an agroecological standpoint, conserving biodiversity within agroecosystems has evolved into a foundational principle for alleviating the stresses impacting agricultural production (Wezel et al., 2020). This global diversity encompasses a wide range of plants, animals, and microorganisms chosen for agriculture, as well as the associated wild biodiversity (Wood et al., 2015). As the scientific and agricultural communities increasingly acknowledge the potential of deliberate and associated agrobiodiversity, an invaluable approach known as functional diversity is emerging. Functional diversity, which considers trophic interactions and the functional traits of species, emerges as a promising approach to understanding how agrobiodiversity can effectively counteract the impacts of stress (Calow, 1987; Wood et al., 2015). In this regard, agricultural diversification is the intentional addition of functional biodiversity through different cropping systems at multiple spatial and/or temporal scales (Kremen et al., 2012; Gaba et al., 2015; Tamburini et al., 2020). This review aims to analyze the reported benefits over the last 10 years of agricultural diversification in mitigating different types of biotic and abiotic stresses in tomato (Solanum lycopersicum L), which, due to its status as a high-yielding crop, high economic value, its rich dose of nutrients such as lycopene and carotenoids, as well as its versatility in cooking, is the most produced vegetable in the world and an important food component of the daily diet in most countries (Anwar et al., 2019). A comprehensive review was conducted by consulting two leading academic databases, SCOPUS and Web of Science (WOS), of research articles on tomatoes grown in one of the cropping systems proposed by Gaba et al. (2015): intercropping, crop sequence, field margin, and cover crop.
2 Abiotic stress
Climate change and pollution of water and soil have led to stress conditions such as drought, chilling, water deficits, and heavy metal presence appearing more frequently in crops (Nawaz et al., 2020; Hasan et al., 2023; Raza et al., 2023; Singh et al., 2023). Utilizing plant companions within different crop systems has demonstrated the ability to mitigate various abiotic stresses; notably, soil nutrient deficiency represents the most extensively studied abiotic stress type (Table 1).
Table 1
| Companion species | Crop system | Infrastructure | Benefit | References |
|---|---|---|---|---|
| Low temperature | ||||
| Wheat (Triticum aestivum L.) | Intercropping | - | Mitigates the negative impact of winter climate conditions on tomato flowering, fruit set, and productivity | Sheha et al., 2022 |
| Heavy metals | ||||
| Sweet pepper (Capsicum annuum L.-cv. Cleor); zucchini (Cucurbita pepo L.-cv. Primula); Hairy vetch (Vicia villosa Roth) | Cover crop; intercropping | - | Cover crops reduce the accumulation of As in the roots, stems, leaves, and total biomass of tomato, sweet pepper, and zucchini | Mancinelli et al., 2019 |
| Eclipta prostrata L.; Crassocephalum crepidioides Benth. | - | Inter-cropping with three plant species resulted in maximum root and shoot biomasses for tomato seedlings and reduced Cd content in tomato seedlings to a minimum | Xie et al., 2021 | |
| Nutrient deficit | ||||
| Barley (Hordeum vulgare L.); hairy vetch (V. villosa) | Cover crop | - | Pure vetch provided optimal N status for the following crop but led to increased nitrate leaching. Mixture of 75% vetch and 25% barley ensured an adequate N supply for tomatoes while reducing the concentration of NO3-N in the soil solution | Farneselli et al., 2018 |
| - | Cultivating pure barley or a barley-vetch mixture (with a higher proportion of barley) reduced N leaching but had variable effects on N uptake by tomatoes. This suggests the necessity for adaptable N supplementation to achieve optimal tomato N nutrition and maximize fruit yield | Farneselli et al., 2020 | ||
| Cowpea (Vigna unguiculata L.) | Cover crop | Greenhouse | The combination of cover crops and farmyard manure increases both total N levels and nitrate concentrations in the soil, leading to higher fruit yields in tomato crops | Gatsios et al., 2019 |
| Hairy vetch (V. villosa) | Cover crop | Plastic high tunnel | The use of cover crops increased soil carbon and N stocks and resulted in the highest soil N availability and greater N uptake | Muchanga et al., 2019 |
| Greenhouse | Although the cover crop had a limited short-term nitrogen-supplying effect, it could also function as a long-term nitrogen source, lasting for up to two cycles after the cover crop is used | Sugihara et al., 2016 | ||
| - | The nitrogen obtained from the cover crop was efficiently absorbed with a minimal amount of chemical fertilizers, making it a viable alternative nitrogen fertilizer source | Sugihara et al., 2013 | ||
| - | The cover crop served as a quick-release fertilizer | Sugihara et al., 2014 | ||
| Hairy vetch (V. villosa); rye (Secale cereale L.) | Cover crop | - | The mixture of 67% vetch and 33% rye increased the total N content in the topsoil and resulted in higher tomato yields, with minimal residual N | Muchanga et al., 2020b |
| Plastic high tunnel | A combination of cover crops can release significant amounts of N during both the early and late stages of tomato cultivation | Muchanga et al., 2020a | ||
| Oat (Avena sativa L.); oilseed radish (Raphanus sativus L.); winter cereal rye (S. cereale) | Cover crop | - | Cover crops did not reduce the mineral N content in the soil during the fall season and increased tomato yield. Among all the cover crop species, radish accumulated the highest amount of N in its above-ground parts during the fall | Chahal and Van Eerd, 2021 |
| Oat (A. sativa); oilseed radish (R. sativus); hairy vetch (V. villosa); clover (Trifolium vesiculosum Savi.) | Cover crop | - | The use of radish cover crops increased soil P, Ca, and K levels. This increase in nutrient availability could be one of the factors contributing to the overall increase in total crop production | Galvão et al., 2013 |
| Oilseed radish (R. sativus) | Cover crop | - | The cover crop increases soil mineral N levels during its growth and continues to do so into the tomato growing season | Belfry et al., 2017 |
| Pea (Pisum arvense L.); barley (H. vulgare) | Cover crop | - | The mixture of 25% barley and 75% pea increases N recycling, regardless of the soil management strategy (no-till or conventional tillage) | Tosti et al., 2019 |
| Sunn hemp (Crotalaria junceae L.); millet (Pennisetum americanum L.) | Cover crop | - | No-tillage following cover crops (both millet and sunn hemp) reduces the N requirements while achieving high tomato production | Branco et al., 2017 |
| Faba bean (Vicia faba L.); wheat (T. aestivum); oilseed radish (R. sativus) | Cover crop | - | V. faba enhanced the effects of externally input N | Fracchiolla et al., 2021 |
| Lettuce (Lactuca sativa L. var. Romana, var. Tantan, var. capitata) | Cover crop; crop sequence | High tunnel | Cover crops played a crucial role in preserving plant-available N within organic matter and increasing the availability of Ca, Mg, K, and P. N uptake by the subsequent cash crop was higher in pea monoculture compared to the pea-oat mixture and when used as green manure rather than mulch | Domagała-Swiatkiewicz and Siwek, 2022 |
| Maize (Zea mays L.); alfalfa (Medicago sativa L.) | Crop sequence | - | Incorporating legumes into a tomato-maize rotation reduced the presence of genes associated with denitrification and facilitated the colonization of various bacterial and fungal groups capable of enhancing soil nutrient availability | Samaddar et al., 2021 |
| - | Continuous cultivation with alfalfa as a perennial crop effectively reduced NO3-leaching | Woodward et al., 2022 | ||
| Celery; lettuce; clover | Crop sequence; cover crop | Greenhouse | Incorporating legumes into vegetable crop rotations reduced N input by 25% | Min et al., 2016 |
| Potato onion (Allium cepa L.) | Intercropping | Greenhouse | Intercropping increased the abundance of P-solubilizing microbial communities when exposed to all tested P forms, including Al-P, Fe-P, Ca-P, and O-P | Khashi u Rahman et al., 2021 |
| Greenhouse | Interspecific interactions led to increased growth and P concentration in tomatoes, especially in the absence of P fertilization. However, these interactions harmed the growth of potato-onion plants | Gao et al., 2021b | ||
| Greenhouse | Intercropping enhanced the growth and P uptake of tomato seedlings, but it decreased the growth and P uptake of potato onion plants | Wu et al., 2016 | ||
| Greenhouse | Intercropping with biochar improved soil physicochemical conditions, enhanced plant nutrient uptake, and influenced soil microbial communities | He et al., 2021 | ||
| Greenhouse | Intercropping increased the percentage of root length colonized by hyphae and arbuscules, as well as K content in tomatoes, but decreased these factors in potato-onions | Gao et al., 2021a | ||
| Faba bean (V. faba) | Intercropping; crop sequence | - | Strip cropping increases P uptake. The N inputs from faba beans can potentially provide sufficient N to support tomato production, but these inputs can vary significantly. Therefore, quick estimates are necessary to determine the need for additional fertilization | Warren-Raffa et al., 2022 |
| Veza villosa (V. villosa) | Cover crop | - | Tomato plants grown with vetch showed reduced effects of N toxicity and maintained high photosynthetic rates even with high levels of inorganic nitrogen | Fatima et al., 2012 |
| Salinity | ||||
| Cereal | Crop sequence | - | Introducing fallow cereal immediately after tomatoes or peppers in the rotation leads to a reduction in soil salinity of ~56% (~2 dS/m) | Bani et al., 2021 |
| Salsola soda L. | Intercropping | Greenhouse | Combining S. soda with tomato cultivation in highly saline soils ensure both yield and quality of tomatoes | Karakas et al., 2016 |
| Arthrocaulon macrostachyum L. | Intercropping; crop sequence | Greenhouse | Both intercropping and sequential cropping decreased chloride content, sodium adsorption ratio, and electrical conductivity, leading to reduced soil salinity | Jurado et al., 2024 |
| Water deficiency | ||||
| Velvet bean (Mucuna pruriens L.); Tanzanian sunhemp (Crotalaria ochroleuca G. Don.); purple vetch (Vicia benghalensis L.) | Cover crop | - | The inclusion of cover crops improves soil moisture retention, leading to higher tomato yields and enhanced water utilization efficiency. Moreover, when vetch biomass is integrated into the soil, both above and below ground, it increases the amount and frequency of deep percolation | Karuku et al., 2014 |
| Brown mustard (Brassica juncea L.); wheat (T. aestivum); green beans (Phaseolus vulgaris L.) | Crop sequence | - | The rotation of tomato, bean, wheat, and mustard enhances N-mineralization, resulting in reduced fertilizer and irrigation inputs compared to other rotation schemes | Kayikcioglu et al., 2020 |
| Maize (Zea mays L.) | Intercropping | - | Tomato yields of up to 32 ton ha −1 can be obtained in rainfed conditions when local tomato cultivars adapted to hot and dry climatic conditions are intercropped with corn | Castronuovo et al., 2023 |
The advantages of companion species in various crop systems for mitigating different abiotic stresses.
2.1 Soil nutrient deficiency
Soil nutrient deficiency refers to the lack or insufficiency of essential nutrients for proper plant growth and development in a specific soil area (Roy et al., 2006). Soil nutrients, such as nitrogen, phosphorus, potassium, calcium, magnesium, and various micronutrients, are essential for plant growth (Kathpalia and Bhatla, 2018). Their absence or low availability can limit plant health and have a negative impact on crop quality and yield (Tripathi et al., 2022). Particularly, tomatoes are a crop known for requiring high nutrient inputs because of their extensive vegetative biomass production, heavy fruit load, and long growing season (Neocleous et al., 2021). Used biodiversity can enhance nutrient supply and assimilation, primarily through cover crops; also known as green manures, when a legume is used (Farneselli et al., 2018). A widely used green manure is hairy vetch (V. villosa), which can contribute optimal nitrogen levels to the tomato crop as a fast-release fertilizer, facilitating nitrogen uptake by up to 38% (Sugihara et al., 2013; Muchanga et al., 2019). However, the fast-release of hairy vetch does not prevent N leaching unless used in mixtures with cereals such as barley (Hordeum vulgare L.) or rye (Secale cereale L.); these mixtures increase the carbon/nitrogen ratio, which slow the rate of nitrogen (N) release and favors the retention of up to 47.0% of the nitrogen produced, thus mitigating this problem (Tosti et al., 2014; Farneselli et al., 2020; Muchanga et al., 2020a).
Although cover crops have been shown to supply considerable N, they are typically used in combination with synthetic fertilizers because the N supply of cover crops is unpredictable and variable and not always synchronized with the needs of the tomato plant (Farneselli et al., 2020). However, green manure may act as an alternative N fertilizer, which enhances the efficiency of external nitrogen inputs and can reduce their requirements. For example, the use of hairy vetch, showed a reduction in external nitrogen inputs by at least 50–66%, primarily during the 4 weeks after transplant when N uptake derived from hairy vetch is higher (Sugihara et al., 2013). Branco et al. (2017) also observed that the use of sunn hemp (Crotalaria junceae L.) and millet (Pennisetum americanum L.) as green manures enhanced the efficiency of external nitrogen inputs, which reduced the requirement of external nitrogen input. Acording Yang et al. (2023), the combination of organic and inorganic fertilizers, also have a significant positive effect on the fruit weight, number, size, elemental content, total yield, and plant height of the tomato crop.
While nitrogen represents the main nutrient contribution of green manures, Fatima et al. (2012) found that hairy vetch (V. villosa), in addition to nitrogen, also provides significant potassium inputs to the tomato crop, as in the case of nitrogen, the potassium content was 125 mg kg−1 and nitrogen 23 mg kg−1 in soil without hairy vetch, while 173 mg kg−1 and 56 mg kg−1 was reported in soils with hairy vetch respectively; and Domagała-Swiatkiewicz and Siwek (2022) report that the use of pea (Pisum sativum L.) and a mixture of pea and oat (Avena sativa L.) as green manure or organic mulch significantly improve the availability of other essential elements, including calcium (Ca), magnesium (Mg), potassium (K) and phosphorus (P) compared to the control.
In addition to cover crops, intercrops such as tomatoes with potato-onion (A. cepa) have demonstrated the promotion of a higher abundance of microbial communities capable of solubilizing different forms of phosphorus, which is positively correlated with the availability of this nutrient (Khashi u Rahman et al., 2021).
2.2 Low-temperature
Tomato originating from subtropical regions is notably susceptible to low-temperature stress and suffers injury at temperatures below 13°C (Pandey et al., 2011; Anwar et al., 2019). Cold temperatures lead to issues such as flower abscission, pollen sterility, ovule abortion, and reduced fruit set, ultimately affecting the final yield (Pandey et al., 2011; Albertos et al., 2019).
To safeguard crops against frost, farmers have employed diverse intervention methods, such as wind machines or greenhouse heating, which incur high energy costs and may offer limited protection (Van Ploeg and Heuvelink, 2005; Albertos et al., 2019). In contrast, intercropping emerges as a sustainable alternative to mitigate the effects of cold stress. An illustrative example is the recent study by Sheha et al. (2022), suggesting that intercropping wheat (Triticum aestivum L.) with tomatoes could be an effective measure against cold and frost; their results show that a seeding rate of 50% led to a significant 9.4% increase in fruit yield per plant compared to a 25% seeding rate because increasing the wheat seeding rate enhances protection against cold temperatures by trapping warm air closer to the plants and reducing heat loss through soil convection; they also consider as a crucial factor the wheat sowing date, which must allow for tomato plant growth before facing severe competition with wheat for essential growth resources.
2.3 Salinity
Salinization, primarily caused by irrigation and fertilization practices, affects soil quality and productivity (Tomaz et al., 2020; Martínez-Sias et al., 2022; Khasanov et al., 2023). This elevated concentration of salts dissolved in the soil solution has adverse consequences on tomatoes, including slowed or reduced seed germination, decreased nutrient absorption, and restricted plant growth (Ondrasek et al., 2022; Khasanov et al., 2023).
Various solutions have been proposed to combat salinization, including promoting plant diversity (Ondrasek et al., 2022). For example, in open field conditions, after tomato cultivation irrigated with saline groundwater, the soil reaches a salinity of ~6 dS/m, but if tomato cultivation is followed by sequences of three crops, the salinity decreases to ~1 dS/m; this is possible because, during the rainfed crops period, rain does the leaching process by washing away the salts in the soil; therefore, salinity varies depending on the growing season and rainfall (Bani et al., 2021).
In the greenhouse context, introducing salt-resistant companion plants has proven to be an effective strategy for reducing soil salinity and safeguarding tomato production. For example, the yield of a tomato grown with companion plants of Salsola soda L. in soils with high salt content increased from 20.3 g plant−1 when the tomato was grown alone to 229.7 g plant−1 because S. soda absorbs and stores soil salts in its tissues (Karakas et al., 2016). The halophyte Arthrocaulon macrostachyum L., in intercropping and sequential cropping with tomato, also reduced soil salinity under moderately saline conditions and enhanced tomato yield (Jurado et al., 2024).
2.4 Water deficiency
Tomato crops have high water requirements, making scarcity a limiting factor that can cause delays in plant development and reduce the number of fruits in clusters (Alomari-Mheidat et al., 2023). There are few studies on how cover crops can contribute to avoiding the impacts of water deficiencies; however, Schomberg et al. (2023) mention that they offer a promising solution because they can reduce runoff, enhance infiltration and minimize evaporation which favors water storage and soil moisture conservation. In this regard (Karuku et al., 2014), studied how purple vetch (Vicia benghalensis L.) as a cover crop improves tomato yield and water use efficiency by 80% and 57% above control, respectively. In addition to cover crops, intercropping local tomato cultivars adapted to hot and dry conditions with corn allows yields of up to 32 ton ha −1 under rainfed conditions (Castronuovo et al., 2023).
2.5 Heavy metals toxicity
Heavy metal toxicity is widespread in agricultural soils across the globe (Ur Rahman et al., 2023). The origin and impact of these pollutants on agriculture vary depending on every heavy metal and crop (Selvi et al., 2019; Joshi and Gururani, 2023; Rashid et al., 2023). Agronomic interventions such as phytoremediation using hyper-accumulator plants to remove contaminants from soil and water is one of the effective methods to remove heavy metals (Elango et al., 2022). For instance, low concentrations of arsenic (As) can stimulate tomato growth, but high concentrations inhibit germination, reduce root and shoot development, lower yield, disrupt photosynthesis and mineral nutrition, and induce necrosis (Sandil et al., 2021). To manage this issue, Mancinelli et al. (2019) explored biodiversity; their study revealed that when preceded by V. villosa as green manure, tomato crops accumulated lower levels of As in their total biomass and yielded higher crop yields.
Cadmium (Cd) is found naturally in the environment and is also generated through human activities, such as phosphate fertilizers (Rahim et al., 2022). Cadmium toxicity can disturb the uptake and translocation of essential mineral nutrients in plants, affecting plant metabolism and inhibiting growth and development (Qin et al., 2020). However, Xie et al. (2021) investigated the effects of intercropping tomatoes with the accumulator plant Eclipta prostrata L. and hyperaccumulator plant Crassocephalum crepidioides Benth; their study demonstrated a significant increase in the biomass of tomato seedlings; additionally, Cd contents in the roots and shoots of tomato seedlings decreased by 17.35% and 22.35%, respectively.
3 Biotic stress
The widespread application of pesticides has caused imbalances inside and outside the plots where they are applied (Maurya et al., 2019). For example, they have caused the resistance of many pests to insecticides, affected beneficial organisms such as pollinators, predators, and parasitoids, and damaged non-target organisms several kilometers from the point of their original release (Aktar et al., 2009; Abad et al., 2020). Agricultural diversification offers a viable alternative to reduce the impacts of crop pests and diseases and dependence on pesticides, as well as to control weeds, although in the latter case, there are still few studies, as shown in Table 2.
Table 2
| Companion species | Crop system | Infrastructure | Benefit | References |
|---|---|---|---|---|
| Aerial pest | ||||
| Asteraceae | Crop sequence | Greenhouse | Tomato plants cultivated in soil previously occupied by Asteraceae family plants displayed significant resistance to caterpillar feeding by Manduca sexta L. | Ingerslew and Kaplan, 2018 |
| - | Field margin | - | Field margins increase the diversity of natural enemies and decrease damage caused by Lepidoptera pests like Tuta absoluta Meyrick. However, total crop damage increases with the expansion of arable land | Balzan et al., 2015 |
| Buckwheat (Fagopyrum esculentum Moench.) | Field margin | - | The parasitism rates of Nezara viridula L. eggs by Trissolcus basalis Wollaston decrease as the distance from the field margin with buckwheat increases | Foti et al., 2019 |
| Sown flower strips; semi-natural margins | Field margin | - | Sown flower strips serve as protective covers against sap-sucking pests, increasing the parasitism rate of aphids, reducing the rate of foliar damage caused by lepidopterans, and enhancing associations with natural enemies. Their effectiveness is further improved when combined with the conservation of diverse seminatural vegetation in the existing field margin | Balzan and Moonen, 2014 |
| Sown mixtures; natural vegetation | Field margin | - | The abundance and diversity of beneficial arthropods were greater in the sown mixture than in natural vegetation | Kati et al., 2021 |
| Basil (Ocimum basilicum L.) | Intercropping | - | Companion cropping with a row of basil planted between adjacent tomato plants and covered with Agronet significantly reduced Bemisia tabaci Genn. infestation in tomatoes by 68.7% | Mutisya et al., 2016 |
| Coriander (Coriandrum sativum L.) | Intercropping | Greenhouse | Coriander volatiles attract Cycloneda sanguinea L. adults, and females also use coriander plants as oviposition sites. Larvae hatching on coriander find nearby aphid-infested tomato plants within about 3 days after hatching | Togni et al., 2016 |
| - | Intercropping reduces the damage caused by T. absoluta and Spodoptera eridania Cramer in organic production systems | Marouelli et al., 2013 | ||
| Persian clover (Trifolium resupinatum L.) | Intercropping | - | Intercropping reduces the density and increases parasitism rates of Helicoverpa armigera Hubner eggs and larvae, especially with row ratios of tomato-clover 1:2 and 2:2. Additionally, the average number of predatory bugs increases with a higher proportion of clover/tomato rows | Abad et al., 2020 |
| Sesame (Sesamum indicum L.) | Intercropping | - | Nesidiocoris tenuis Reuter are zoophytophagous mirids used successfully as an alternative for controlling B. tabaci. However, this control agent can become a pest of tomatoes when prey is scarce. Intercropping helps maintain the populations of N. tenuis with minimal damage to tomatoes | Castillo et al., 2022 |
| Solanum lycopersicum var. cerasiforme | Intraspecific mixture | - | Monocropping wild tomatoes and intercropping wild and cultivated tomato plants both result in reduced levels of T. absoluta infestation | Miano et al., 2022 |
| Wild oregano (Plectranthus amboinicus Lour.) | Intercropping | Greenhouse | In a greenhouse insect-proof cage, tomato plants intercropped with wild oregano from southern Martinique had 1.5 fewer adult whiteflies than tomato plants grown alone after 96 h of exposure. The behavior of B. tabaci appears to be influenced by the origin of the plant | Pouët et al., 2022 |
| Coriander (C. sativum); Greek basil (Ocimum minimum Labiatae) | Intercropping | Open field; greenhouse | Planting tomatoes with coriander and Greek basil resulted in an 84% and 79% reduction in adult B. tabaci populations compared to monocropping | Carvalho et al., 2017 |
| English marigold (Calendula officinalis L.) | Intercropping | Greenhouse | Intercropping significantly inhibits the population development of Trialeurodes vaporariorum Westwood | Conboy et al., 2019 |
| Marigold (Tagetes erecta L.) | Intercropping | - | Intercropping, combined with one release of Trichogramma chilonis Ishii, the application of Nimbecidine, and a spray of Bacillus thuringiensis Berliner kurstaki, significantly reduces the number of eggs and larval population of H. armigera, with minimal negative impact on non-target organisms | Khokhar and Rolania, 2022 |
| Sainfoin (Onobrychis viciifolia Scop.) | Intercropping | - | Intercropping reduces the number of T. absoluta eggs, larvae, and galleries. It also increases the diversity index for the species composition of T. absoluta predators, such as Orius niger Wolff and N. tenuis, and raises the larval parasitism rate | Zarei et al., 2019 |
| Coriander (C. sativum) | Intercropping | - | The presence of coriander plants created visual and olfactory barriers that made tomato plants less attractive to B. tabaci. As a result, intercropping with overhead sprinkler irrigation reduced the colonization levels of B. tabaci and decreased the incidence of Begomovirus-infected plants | Togni et al., 2018 |
| Coriander (C. sativum); marigold (Tagetes minuta L.); sorghum (Sorghum bicolor L.) | Crop mixture (surrounding crop) | - | Intercropping reduced the incidence of thrips, resulting in a decrease in the percentage of plants and fruits with symptoms of viral disease. Additionally, it led to a reduced incidence of drilling by Neoleucinodes elegantalis Guenee | Gomes et al., 2012 |
| Coriander (C. sativum) and dill (Anethum graveolens L.) | Intercropping | - | The reduction in whitefly population between the intercropping treatments ranged from 56.00% to 72.00% with coriander and 33.00% to 58.00% with dill, when compared to tomato as a sole crop | Padala et al., 2023 |
| - | Field margin | - | Field margins increased the densities of natural enemies, reduced pest damage to the crop, decreased weed cover, and consequently, resulted in higher yields in tomato crops. Pest control was more effective during the late crop stage compared to early-season sampling | Segre et al., 2020 |
| Soil-borne pathogens | ||||
| Maize (Z. mays) | Cover crop | - | Crop rotation enhances the effectiveness of the antagonist Pasteuria penetrans in suppressing root-knot nematode (RKN) populations in subsequent crop cycles | Shahid et al., 2020 |
| Crop sequence | Glasshouse; open field | The application of crop rotation consistently led to a reduction in the numbers of second-stage juvenile RKN on roots, combined with the benefits of the Pochonia chlamydosporia biological control agent | Luambano et al., 2015 | |
| Sunn hemp (Crotalaria juncea L.) | Cover crop | - | Sunn hemp is effective in the suppression of Meloidogyne incognita Kofoid and White | Marquez and Hajihassani, 2023 |
| Nemat (Eruca sativa Mill.); canola (Brassica napus L.) | Cover crop; crop sequence | Greenhouse | Using nemat as a cover or rotation crop suppressed Meloidogyne javanica. In the case of Criconemoides xenoplax, the population declined over time when canola was implemented as a cover crop | Kruger et al., 2015 |
| Wheat-resistant cultivar Lassik (Triticum aestivum L.) | Crop sequence | Greenhouse | Tomato exhibited reduced root galling by RKN when grown in soil from micro plots that had previously included RKN-resistant wheat | Williamson et al., 2013 |
| Castor (Ricinus communis L.) | Intercropping | Greenhouse | The root exudates from intercropped plants reduced the nematode soil population and inhibited M. incognita infection by regulating nematode chemotaxis | Dong et al., 2018 |
| Crown daisy (Chrysanthemum coronarium L.) | Intercropping | Greenhouse | The potent bioactivity of lauric acid, which is a specific crown daisy root exudate, serves as both a lethal trap and a repellent for M. incognita. The reaction depends on the concentration of the exudate | Dong et al., 2014 |
| Black-jack (Bidens pilosa L.) | Intercropping | - | Intercropping reduced the number of galls and egg masses on M. incognita susceptible host plants by 3–9 times compared to the control group | Kihika-Opanda et al., 2022 |
| Chinese leek (Allium tuberosum Rottler ex Spreng) | Intercropping | Greenhouse | The gall indexes of tomato plants intercropped with Chinese leek decreased by 41.1%. Chinese leek exhibits high resistance to RKNs, demonstrates strong nematocidal activity against M. incognita, and significantly reduces the incidence of diseases caused by nematodes | Huang et al., 2016 |
| Moringa (Moringa oleifera Lam.) | Intercropping | Greenhouse | The lowest values of M. incognita and Rotylenchulus reniformis Linford and Oliveira, as well as their rate of increase and root gall index, were observed when the highest number of moringa seedlings per pot (four) were grown | Ismail, 2013 |
| Marigold (T. erecta); garlic (Allium sativum L.) | Intercropping; crop sequence | Greenhouse | Intercropping tomato with antagonistic marigold, followed by tomato intercropped with garlic, resulting in a reduction in root galls, egg masses, and J2 nematodes of RKNs per plant compared to tomato monoculture | Miheret et al., 2019 |
| Arugula (E. sativa.) | - | The use of fresh arugula as a soil amendment reduced tomato nematode infection. Therefore, it can be considered a promising companion plant for tomato growers to control M. incognita | Aissani et al., 2015 | |
| Marigold (Tagetes patula L.); basil (O. basilicum); lettuce (L. sativa); white mustard (Sinapis alba L.) | Intercropping | Greenhouse | All the tested companion plants suppressed the development of Meloidogyne spp., with white mustard and marigold showing the highest effectiveness at 53.45% and 46.38%, respectively | Tringovska et al., 2015 |
| Garlic (Allium sativum L.); Madagascar periwinkle (Catharanthus roseus L.); Yarrow (Achillea millefolium L.) | Intercropping | Laboratory; greenhouse | The root exudates of garlic and yarrow did not attract the J2s of M. javanica. The root exudates of marigolds were highly and Madagascar periwinkle slightly attractive; but despite their attractiveness, J2s did not complete their cycle inside the roots in marigolds and Madagascar periwinkle | Cavalcanti et al., 2023 |
| Rye (S. cereale); Indian mustard (B. juncea); white mustard (S. alba); arugula (E. sativa) | Cover crop (Anaerobic soil disinfestation) | - | The mean populations of Rhizoctonia solani Kuhn throughout the year were lower than those in the untreated control and similar to those in the biofumigant control when using cover crops such as mustard/arugula and rye for anaerobic soil disinfestation treatment | McCarty et al., 2014 |
| Celery (Apium graveolens L.) | Crop sequence | Greenhouse | Rotating with celery may increase the abundance and diversity of fungi in continuous tomato cropping substrates while reducing the relative abundance of harmful fungi such as Pseudogymnoascus, Gibberella, and Pyrenochaeta | Lyu et al., 2020 |
| Rice (Oryza sativa L.) | Crop sequence | - | Crop sequencing reduced the presence of the pathogenic fungal genus Fusarium and increased the presence of potentially beneficial bacterial phyla, such as Acidobacteria and Chloroflexi. The total phosphorus content was a determining factor for both bacterial and fungal communities | Ma et al., 2021 |
| Wheat (T. aestivum) | Crop sequence | - | After 4 years of consecutive rotation, there was an increase in the soil's suppressive response against F. oxysporum, a decrease in their pathogenicity and abundance in the tomato rhizosphere, and an increase in the growth of tomato shoots | De Corato et al., 2020 |
| Basil (O. basilicum) | Intercropping | Greenhouse | Intercropping increased plant biomass and significantly reduced the severity of F. oxysporum in tomatoes. Furthermore, the addition of arbuscular mycorrhizal fungi to the intercrop enhanced the host plant's tolerance by influencing tomato root morphology and exudation dynamics | Raza et al., 2022 |
| Potato-onion (A. cepa) | Intercropping | - | Intercropping increased the relative abundances of certain taxa and various operational taxonomic units that have the potential to enhance plant growth. Simultaneously, it decreased the relative abundances of some potential plant pathogens, such as Cladosporium | Li et al., 2020 |
| - | Intercropping can enhance tomato resistance against F. oxysporum by improving soil enzymes and increasing antifungal enzyme activity in the soil | Sweellum and Naguib, 2023 | ||
| Greenhouse | Intercropping altered the composition of the tomato rhizosphere microbiome by increasing the colonization of specific Bacillus sp., which has been shown to reduce the growth of V. dahliae and induce systemic resistance in tomato plants | Zhou et al., 2023 | ||
| Greenhouse | Root exudates from tomatoes accompanied by potato-onion reduced the mycelial growth and spore germination of V. dahliae. However, these effects were not observed with root exudates from potato-onion alone | Fu et al., 2015 | ||
| Crop sequence | - | The root exudates from Chinese onion accessions increased tomato growth and reduced the incidence of F. oxysporum | Liu et al., 2013 | |
| - | Laboratory | Root exudate inhibited the growth of V. dahlia | Li et al., 2018 | |
| Marigold (T. erecta) | Intercropping | - | Intercropping with plastic mulching reduces disease intensity compared to tomato monocropping. Marigold serves as a barrier to the movement of conidia, and plastic mulching creates a microclimate that reduces the germination of Alternaria solani spores | Jambhulkar et al., 2015 |
| - | Intercropping with the use of plastic mulch reduces the intensity of A. solani. Marigold serves as a barrier to the movement of conidia, and plastic mulch prevents evapotranspiration, reducing canopy relative humidity | Jambhulkar et al., 2016 | ||
| Tall fescue (F. arundinacea) | Intercropping | Greenhouse | Intercropping tomatoes with tall fescue resulted in a reduced incidence and severity of R. solani, and the antifungal activities of root exudates from both species increased in the intercropping system | Zhou et al., 2019 |
| Sunn hemp (C. juncea); Japanese millet (Echinochloa crusgalli L.) | Cover crop | - | In monoculture, the disease incidence was 33%, whereas in the rotation with sunn hemp and Japanese millet, it decreased to 9%. These benefits continued into a second season, with a 17% reduction in Ralstonia solanacearum Smith in the rotation plots. Furthermore, tomato plants in the rotation exhibited lower levels of root galls caused by M. incognita infection | Chellemi et al., 2013 |
| Weed and parasitic plants | ||||
| Hairy vetch (V. villosa) | Cover crop | - | Hairy vetch, when used as a cover crop, helps reduce weed growth | Campiglia et al., 2015 |
| Soybean; maize; sweet corn; bean | Crop sequence | - | Crop rotation reduces volunteer plants in tomato crops and can also decrease the sources of inoculum for diseases transmitted by seeds, such as bacterial spots caused by Xanthomonas perforans Jones | Moura et al., 2020 |
| V. villosa; Vicia sativa L.; Secale cereale L.; x Triticosecale Wittmack | Cover crop | - | Cover crops reduced weed dry weight in all tillage systems and increased the nitrogen level in tomato leaves. Soil nitrogen and organic carbon contents remained unaffected throughout the experiments | Samedani and Meighani, 2022 |
| Vigna sinensis L.; Hibiscus sabdariffa L.; H. vulgare; Sorghum vulgare Pers. | Crop sequence | Glasshouse | Tomato cultivation following service species resulted in increased tomato growth and improved control of Orobanche | Qasem, 2019 |
Advantages of partner plant species in various crop systems for mitigating diverse biotic stresses.
3.1 Aerial pest
Pests cause 20% of global annual crop losses (Mateos-Fernández et al., 2022). This vulnerability of crops is observed, particularly in monoculture systems (Ratnadass et al., 2021). Incorporating high functional biodiversity through different cropping systems contributes to maintaining ecological functions that help pest management (Altieri et al., 2015). For aerial pests case, intercropping has proven to be particularly effective because companion plants play multiple roles, serving as repellents, reservoirs for natural enemies, and creating visual and olfactory barriers (Togni et al., 2016; Carvalho et al., 2017; Pouët et al., 2022). For example, Mutisya et al. (2016) observed that a row of aromatic basil (Ocimum basilicum L.) as companion cropping between adjacent tomato rows significantly lowered whitefly (Bemisia tabaci Genn) infestation in tomatoes by 68.7% and resulted in 13.75 t/ha−1 as tomato yield compared to 5.9 t/ha−1 in the control because attractive nature of the basil plant makes it a better host for insects such as B. tabaci. Moreover, the essential oils and volatiles of wild oregano (Plectranthus amboinicus Lour.), coriander (Coriandrum sativum L.), and Greek basil (Ocimum minimum Labiatae) have been shown to repel B. tabaci when grown with tomatoes (Carvalho et al., 2017; Pouët et al., 2022); additionally, C. sativum shown to reduces the tomato damage caused by Tuta absoluta Meyrick and Spodoptera eridania Cramer, and attract females of Cycloneda sanguinea L. who use coriander plants as oviposition sites and help control aphids of infested tomato plants (Marouelli et al., 2013; Togni et al., 2016). Another companion plant that attracts predators is sesame (Sesamum indicum L.), which helps to maintain the zoophytophagus mirid Nesidiocoris tenuis Reuter populations during the entire tomato cropping season and reduces up to 75% of the whitefly numbers compared to untreated tomatoes. Management of B. tabaci also contributes to reduced disease transmission, such as Begomovirus (Togni et al., 2018).
The secretion of phenolic compounds, essential oils, and volatile organic compounds helps the companion plants to act as repellents or olfactory barriers. This mechanism can also attract beneficial organisms, to which the companion plants provide additional resources such as food and shelter (Abad et al., 2020; Castillo et al., 2022). Therefore, aromatic plants are commonly selected as companion plants for pest management (Carvalho et al., 2017; Conboy et al., 2019).
The depletion of natural landscape habitats, linked to monocultures and agricultural intensification, has increased pest pressure (Balzan et al., 2015). One approach to enhance habitat complexity is the implementation of field margins, which provide flowering resources and alternate prey necessary to enhance natural enemies' abundance and richness (Segre et al., 2020). Plants with strong scents and abundant nectar are generally used for the margins to complement the existing natural diversity (Balzan and Moonen, 2014; Foti et al., 2019). Certain plant families, such as Asteraceae, Fabaceae, and Apiaceae, have demonstrated the ability to foster a reservoir of parasitoids and predators (Kati et al., 2021). It's worth noting that the studies we reviewed primarily focus on observations made in open fields and within ~200 meters from the margins.
Other cropping systems, such as intraspecific mixtures and crop sequences, have also shown promise in mitigating damage caused by aerial pests (Ingerslew and Kaplan, 2018; Miano et al., 2022). However, a more comprehensive understanding of their benefits is needed.
3.2 Soil-borne pathogens
Monoculture reduces microbial diversity and soil organic matter, consequently giving rise to soil-borne pathogens such as fungi, bacteria, actinomycetes, and nematodes (Hooper et al., 2000; Lyu et al., 2020). According to this review, in the context of tomato crops, nematodes and fungi have been the primary focus of research.
Among the most common pathogenic nematodes affecting tomato crops are the root-knot nematodes (RKN), belonging to the genus Meloidogyne (Seid et al., 2015). Several cultivation systems have been employed to manage them, including cover cropping, crop sequencing, and intercropping. For crop sequencing, companion plants from the Poaceae family, such as wheat and maize, have been utilized; in the maize inclusion case, the presence and effectiveness of antagonists or biological control agents like Pasteuria penetrans and Pochonia chlamydosporia are enhanced and resulted in a 72% decrease in numbers of egg masses, 38% in root galling and 46% regarding female nematode populations over the control after the final harvest (Luambano et al., 2015; Shahid et al., 2020). In the case of cover crops, plants from the Brassicaceae family offer a biocidal effect due to the release of specific biologically active compounds during maceration and incorporation processes (Kruger et al., 2015). Also, Fabaceae plants, such as C. juncea, are employed as cover crops (Marquez and Hajihassani, 2023), which favor the abundance and richness of soil communities, which results effectively in suppressing M. incognita (Scaglione et al., 2023). In the intercropping system, species like Allium tuberosum Rottl, Ricinus communis L., Chrysanthemum coronarium L., and Bidens pilosa L. release exudates that inhibit egg hatching or act as nematicides (Dong et al., 2014, 2018; Huang et al., 2016; Kihika-Opanda et al., 2022).
While companion species in intercropping systems offer benefits for controlling soil pests, they sometimes can lead to reduced yields of the main crop due to competition among plants (Tringovska et al., 2015). In such cases, it is essential to conduct studies to identify the minimum population of companion plants that can continue to provide benefits without significantly affecting tomato yield (Castillo et al., 2022).
To control pathogenic fungi, crop rotation and intercropping stand out as widely studied cropping systems; particularly, crop rotation enhances the suppression of pathogens and improves tomato plant growth by inducing changes in microbial composition and soil chemical parameters (De Corato et al., 2020). For instance, Apium graveolens L., due to its potent allelochemicals that alter soil pH, enhances the abundance and diversity of fungi, reducing the abundance of harmful organisms (Lyu et al., 2020).
Both intercropping and crop rotation induce changes in microbial communities and improve the soil environment (Li et al., 2020; Zhou et al., 2023). Additionally, they boost tomato resistance against pathogens. For example, when the tomato grew with A. cepa, it exhibited heightened activity in antifungal enzymes and increased content of phenols bound to the cell wall, effectively curbing the growth and spread of Fusarium oxysporum Schl. (Sweellum and Naguib, 2023). Moreover, root exudates from tomatoes accompanied by A. cepa significantly inhibit the mycelia growth and spore germination of Verticillium dahlia Kleb (Fu et al., 2015). Another way in which mixed cultures reduce pathogen damage is by acting as barriers to the movement of conidia, as demonstrated by the use of Tagetes erecta L. against Alternaria solani (Jambhulkar et al., 2015).
3.3 Weeds, volunteer plants, and parasitic plants
Weeds compete with crops for essential resources such as light, water, and nutrients; additionally, they can serve as alternative hosts for crop pests and pathogens (Moura et al., 2020; Christina et al., 2021). The few existing studies provide a glimpse of the benefits of cover crops in weed management, mainly in no-tillage tomato crops, but they are not sufficient to understand all the variables that may influence the success of this practice against this type of stress, so there are still challenges regarding the use of cover crops in weed control (Campiglia et al., 2015; Roberts and Mattoo, 2018; Samedani and Meighani, 2022).
In the case of parasitic plants, Orobanche spp. is a problematic parasitic species for agriculture that is difficult to control; trap species such as Vigna sinensis L., Hibiscus sabdariffa L., H. vulgare, Sorghum vulgare Pers. stimulate the germination of parasite seeds and then destroy them before the parasite flowers, reducing the parasite in the subsequent tomato crop by 73% (Qasem, 2019).
Volunteer plants, which can sprout from fruit that remains in the soil after mechanical harvesting of tomato, represent a source of inoculum for many crop diseases, such as Xanthomonas perforans Jones; in this regard, when tomatoes grow with soybean, corn, sweet corn, and bean, the number of volunteer plants and sources of X. perforans inoculum is reduced (Moura et al., 2020).
4 Conclusions and future directions
Biodiversity offers numerous advantages for tomato cultivation. These benefits directly mitigate the impacts of various types of biotic and abiotic stress and indirectly align with agroecological principles such as reduced inputs, improved soil health, synergy, and recycling. The most extensively studied stressors encompass: (1) Soil nutrient deficiency: Often addressed with companion species like V. villosa; (2) Aerial Pests: Effectively managed with aromatic plants such as C. sativum; and (3) Soil-Borne Pathogens: A. cepa is a common choice for combatting pathogenic fungi. Different cropping systems come into play, with cover crops being prominent for nutritional deficiencies and intercropping for aerial pests and soil-borne pathogens.
Future research should focus on assessing companion species that can mitigate less explored stress types such as cold temperatures, heavy metal contamination, salinity, water scarcity, and weed infestations. These evaluations should consider factors like species distribution, plant quantities, and the cultivation system that maximizes companion species benefits.
To select companion species for evaluation it's recommended to incorporate local knowledge through participatory methods while also developing general guidelines or principles for species selection. A couple of proposals arising from this review are: (1) Aromatic plants show significant potential in managing aerial pests and soil-borne disease pathogens, and (2) Cover crops contribute directly and indirectly to provide nutrients. Using legume-cereal mixtures to balance the carbon-nitrogen ratio prevents nutrient leaching.
Statements
Author contributions
VC-L: Data curation, Formal analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review & editing, Conceptualization. CG-E: Formal analysis, Methodology, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. RP-P: Conceptualization, Formal analysis, Resources, Supervision, Validation, Writing – review & editing. CR: Conceptualization, Formal analysis, Resources, Supervision, Validation, Visualization, Writing – review & editing. JÁ-L: Data curation, Formal analysis, Methodology, Software, Supervision, Validation, Visualization, Writing – review & editing. IM: Data curation, Formal analysis, Resources, Software, Supervision, Validation, Visualization, Writing – review & editing. LB-O: Data curation, Methodology, Project administration, Software, Supervision, Validation, Visualization, Writing – review & editing. FG-P: Data curation, Formal analysis, Resources, Software, Supervision, Validation, Visualization, Writing – review & editing. JD-J: Data curation, Formal analysis, Resources, Software, Supervision, Validation, Visualization, Writing – review & editing. NL-V: Data curation, Formal analysis, Resources, Software, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by the Instituto Politécnico Nacional through grant agreement SIP-20231866 and Consejo Nacional de Humanidades, Ciencia y Tecnología (CONAHCYT-Mexico) for the Scholarship (660555).
Acknowledgments
We thank the reviewers for their suggestions in a previous version of the manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1
AbadM. K. R.FathiS. A. A.Nouri-GanbalaniG.Amiri-BesheliB. (2020). Influence of tomato/clover intercropping on the control of Helicoverpa armigera (Hübner). Int. J. Trop. Insect. Sci.40, 39–48. 10.1007/s42690-019-00048-z
2
AissaniN.UrgegheP. P.OplosC.SabaM.ToccoG.PetrettoG. L.et al. (2015). Nematicidal activity of the volatilome of eruca sativa on meloidogyne incognita. J. Agric. Food Chem.63, 6120–6125. 10.1021/acs.jafc.5b02425
3
AktarW.SenguptaD.ChowdhuryA. (2009). Impact of pesticides use in agriculture: their benefits and hazards. Interdiscip. Toxicol.2, 1–12. 10.2478/v10102-009-0001-7
4
AlbertosP.WagnerK.PoppenbergerB. (2019). Cold stress signalling in female reproductive tissues. Plant Cell. Environ.42, 846–853. 10.1111/pce.13408
5
Alomari-MheidatM.CorellM.Castro-ValdecantosP.AndreuL.MorianaA.Martín-PalomoM. J. (2023). Effect of water stress and rehydration on the cluster and fruit quality of greenhouse tomatoes. Agronomy13, 563. 10.3390/agronomy13020563
6
AltieriM. A.NichollsC. I.HenaoA.LanaM. A. (2015). Agroecology and the design of climate change-resilient farming systems. Agron Sustain Dev35, 869–890. 10.1007/s13593-015-0285-2
7
AnwarR.FatimaT.MattooA. K. (2019). Tomatoes: A Model Crop of Solanaceous Plants in Oxford Research Encyclopedia of Environmental Science.Oxford: Oxford University Press.
8
BalzanM. V.BocciG.MoonenA.-C. (2015). Landscape complexity and field margin vegetation diversity enhance natural enemies and reduce herbivory by Lepidoptera pests on tomato crop. BioControl61, 141–154. 10.1007/s10526-015-9711-2
9
BalzanM. V.MoonenA. C. (2014). Field margin vegetation enhances biological control and crop damage suppression from multiple pests in organic tomato fields. Entomol Exp Appl150, 45–65. 10.1111/eea.12142
10
BaniA.DaghariI.HatiraA.ChaabaneA.DaghariH. (2021). Sustainable management of a cropping system under salt stress conditions (Korba, Cap-Bon, Tunisia). Environ. Sci. Pollut. Res.28, 46469–46476. 10.1007/s11356-020-09767-0
11
BelfryK. D.TruemanC.VynR. J.LoewenS. A.Van EerdL. L. (2017). Winter cover crops on processing tomato yield, quality, pest pressure, nitrogen availability, and profit margins. PLoS ONE12, e0180500. 10.1371/journal.pone.0180500
12
BenavidezA.RamírezH.RobledoV.MaitiR.CornejoE.HernándezJ.et al. (2002). Ecofisiología y Bioquímica del estrés en Plantas. Saltillo: Univseridad Autonóma Antonio Narro.
13
BrancoR. B.BlatS. F.GimenesT. G.NowakiR. H.AraújoH. S.SallesF. A. (2017). Nitrogen fertilization of vegetables cultivated under no-tillage after cover crops. Hortic. Bras.35, 103–110. 10.1590/s0102-053620170116
14
CalowP. (1987). Towards a definition of functional ecology. Funct. Ecol.1, 57. 10.2307/2389358
15
CampigliaE.RadicettiE.MancinelliR. (2015). Cover crops and mulches influence weed management and weed flora composition in strip-tilled tomato (Solanum lycopersicum). Weed Res.55, 416–425. 10.1111/wre.12156
16
CarvalhoM. G.BortolottoO. C.VenturaM. U. (2017). Aromatic plants affect the selection of host tomato plants by Bemisia tabaci biotype B. Entomol. Exp. Appl.162, 86–92. 10.1111/eea.12534
17
CastilloJ.RodaA.QureshiJ.Pérez-HedoM.UrbanejaA.StanslyP. (2022). Sesame as an alternative host plant to establish and retain predatory mirids in open-field tomatoes. Plants11, 2779. 10.3390/plants11202779
18
CastronuovoD.CardoneL.CruoglioM.LelaL.BenedettoN.CarlucciV.et al. (2023). Productivity and quality of different tomato cultivars under intercropping system with maize and dry farming conditions in Southern Italy. Italus Hortus30, 3. 10.26353/j.itahort/2023.1.0316
19
CavalcantiV. P.TerraW. C.da SilvaJ. C. P.OliveiraA. J. M.FonsecaK. M. F.SilvaB. M.et al. (2023). Attractive response of Meloidogyne javanica varies among non-host plants, while all of them reduce the nematode population when intercropped with host plants. Plant Soil. 12, 1–17. 10.1007/s11104-023-06194-1
20
ChahalI.Van EerdL. L. (2021). Cover crops increase tomato productivity and reduce nitrogen losses in a temperate humid climate. Nutr. Cycl. Agroecosyst.119, 195–211. 10.1007/s10705-020-10105-6
21
ChellemiD. O.RosskopfE. N.Kokalis-BurelleN. (2013). The effect of transitional organic production practices on soilborne pests of tomato in a simulated microplot study. Phytopathology103, 792–801. 10.1094/PHYTO-09-12-0243-R
22
ChristinaM.NegrierA.MarnotteP.ViaudP.MansuyA.AuzouxS.et al. (2021). A trait-based analysis to assess the ability of cover crops to control weeds in a tropical island. Eur. J. Agron.128, 126316. 10.1016/j.eja.2021.126316
23
ConboyN. J. A.McDanielT.OrmerodA.GeorgeD.GatehouseA. M. R.WhartonE.et al. (2019). Companion planting with French marigolds protects tomato plants from glasshouse whiteflies through the emission of airborne limonene. PLoS ONE14, e0213071. 10.1371/journal.pone.0213071
24
CorradoC.ElenaT.GiancarloR.StefanoC. (2019). “The Role of Agrobiodiversity in Sustainable Food Systems Design and Management,” in Genetic Diversity in Horticultural Plants, ed. D. Nandwani (Cham: Springer), 245–271.
25
De CoratoU.PatrunoL.AvellaN.SalimbeniR.LacollaG.CucciG.et al. (2020). Soil management under tomato-wheat rotation increases the suppressive response against Fusarium wilt and tomato shoot growth by changing the microbial composition and chemical parameters. Appl. Soil Ecol.154, 103601. 10.1016/j.apsoil.2020.103601
26
Domagała-SwiatkiewiczI.SiwekP. (2022). Effect of field pea (Pisum sativum subsp. arvense (L.) Asch.) and pea-oat (Avena sativa L.) biculture cover crops on high tunnel vegetable under organic production system. Organic Agric.12, 91–106. 10.1007/s13165-021-00383-x
27
DongL.LiX.HuangC.LuQ.LiB.YaoY.et al. (2018). Reduced Meloidogyne incognita infection of tomato in the presence of castor and the involvement of fatty acids. Sci. Hortic.237, 169–175. 10.1016/j.scienta.2018.03.066
28
DongL.LiX.HuangL.GaoY.ZhongL.ZhengY.et al. (2014). Lauric acid in crown daisy root exudate potently regulates root-knot nematode chemotaxis and disrupts Mi-flp-18 expression to block infection. J. Exp. Bot.65, 131–141. 10.1093/jxb/ert356
29
ElangoD.DeviK. D.JeyabalakrishnanH. K.RajendranK.Thoomatti HaridassV. K.DharmarajD.et al. (2022). Agronomic, breeding, and biotechnological interventions to mitigate heavy metal toxicity problems in agriculture. J. Agric. Food Res.10, 100374. 10.1016/j.jafr.2022.100374
30
EnebeM. C.BabalolaO. O. (2018). The influence of plant growth-promoting rhizobacteria in plant tolerance to abiotic stress: a survival strategy. Appl. Microbiol. Biotechnol.102, 7821–7835. 10.1007/s00253-018-9214-z
31
FarneselliM.BenincasaP.TostiG.GuiducciM.TeiF. (2020). Combining green manuring and fertigation maximizes tomato crop yield and minimizes nitrogen losses. Agronomy10, 977. 10.3390/agronomy10070977
32
FarneselliM.TostiG.OnofriA.BenincasaP.GuiducciM.PannacciE.et al. (2018). Effects of N sources and management strategies on crop growth, yield and potential N leaching in processing tomato. Eur. J. Agron.98, 46–54. 10.1016/j.eja.2018.04.006
33
FatimaT.TeasdaleJ. R.BunceJ.MattooA. K. (2012). Tomato response to legume cover crop and nitrogen: differing enhancement patterns of fruit yield, photosynthesis and gene expression. Funct. Plant Biol.39, 246. 10.1071/FP11240
34
FotiM. C.PeriE.WajnbergE.ColazzaS.RostásM. (2019). Contrasting olfactory responses of two egg parasitoids to buckwheat floral scent are reflected in field parasitism rates. J. Pest Sci.92, 747–756. 10.1007/s10340-018-1045-2
35
FracchiollaM.RennaM.DuranteM.MitaG.SerioF.CazzatoE. (2021). Cover crops and manure combined with commercial fertilizers differently affect yield and quality of processing tomato (Solanum lycopersicum L.) organically grown in puglia. Agriculture11, 757. 10.3390/agriculture11080757
36
FuX.WuX.ZhouX.LiuS.ShenY.WuF. (2015). Companion cropping with potato onion enhances the disease resistance of tomato against Verticillium dahliae. Front. Plant Sci.6, 726. 10.3389/fpls.2015.00726
37
GabaS.LescourretF.BoudsocqS.EnjalbertJ.HinsingerP.JournetE. P.et al. (2015). Multiple cropping systems as drivers for providing multiple ecosystem services: from concepts to design. Agron. Sustain. Dev.35, 607–623. 10.1007/s13593-014-0272-z
38
GalvãoA. G.ResendeJ. T. V.deMorales, R. G. F.LustosaS. B.DiasD. M.MarodinJ. C. (2013). Tomato yield and soil chemical attributes depending on previous cover crops. Hortic Bras31, 68–73. 10.1590/S0102-05362013000100011
39
GaoD.PanX.Khashiu RahmanM.ZhouX.WuF. (2021a). Common mycorrhizal networks benefit to the asymmetric interspecific facilitation via K exchange in an agricultural intercropping system. Biol. Fertil. Soils57, 959–971. 10.1007/s00374-021-01561-5
40
GaoD.PanX.ZhouX.WeiZ.LiN.WuF. (2021b). Phosphorus fertilization and intercropping interactively affect tomato and potato onion growth and rhizosphere arbuscular mycorrhizal fungal community. Arch. Agron. Soil Sci.67, 919–933. 10.1080/03650340.2020.1768530
41
GatsiosA.NtatsiG.CeliL.Said-PullicinoD.TampakakiA.GiannakouI.et al. (2019). Nitrogen nutrition optimization in organic greenhouse tomato through the use of legume plants as green manure or intercrops. Agronomy9, 766. 10.3390/agronomy9110766
42
GomesF. B.FortunatoL.PachecoA. L. V.AzevedoL. H.de FreitasN.HommaS. K. (2012). Incidência de pragas e desempenho produtivo de tomateiro orgânico em monocultivo e policultivo. Hortic. Bras.30, 756–761. 10.1590/S0102-05362012000400032
43
HasanN.PushpalathaR.ManivasagamV. S.ArlikattiS.CibinR. (2023). Global sustainable water management: a systematic qualitative review. Water Res. Manage.37, 5255–5272. 10.1007/s11269-023-03604-y
44
HeX.XieH.GaoD.KhashiU.RahmanM.ZhouX.WuF. (2021). Biochar and intercropping with potato–onion enhanced the growth and yield advantages of tomato by regulating the soil properties, nutrient uptake, and soil microbial community. Front. Microbiol.12, 695447. 10.3389/fmicb.2021.695447
45
HooperD.BignellD.BrownV.BrussaardL.DangerfieldM.WallD.et al. (2000). Interactions between aboveground and belowground biodiversity in terrestrial ecosystems: patterns, mechanisms, and feedbacks. Bioscience50, 1049–106110.1641/0006-3568(2000)050(1049:IBAABB)2.0.CO;2
46
HuangY.MaoZ.XieB. (2016). Chinese leek (Allium tuberosum Rottler ex Sprengel) reduced disease symptom caused by root-knot nematode. J. Integr. Agric.15, 364–372. 10.1016/S2095-3119(15)61032-2
47
InbarajM. P. (2021). Plant-microbe interactions in alleviating abiotic stress—a mini review. Front. Agron.3, 667903. 10.3389/fagro.2021.667903
48
IngerslewK. S.KaplanI. (2018). Distantly related crops are not better rotation partners for tomato. J. Appl. Ecol.55, 2506–2516. 10.1111/1365-2664.13156
49
IsmailA. E. (2013). Feasibility of growing Moringa oleifera as a mix-crop along with tomato for control of Meloidogyne incognita and Rotylenchulus reniformis in Egypt. Arch. Phytopathol. Plant Prot.46, 1403–1407. 10.1080/03235408.2013.768062
50
JambhulkarP. P.JambhulkarN.MeghwalM.AmetaG. S. (2016). Altering conidial dispersal of alternaria solani by modifying microclimate in tomato crop canopy. Plant Pathol. J.32, 508–518. 10.5423/PPJ.OA.06.2015.0101
51
JambhulkarP. P.MeghwalM. L.AmetaG. S. (2015). Microclimatic modification of tomato crop canopy to alter conidial dispersal of alternaria solani. Vegetos An Int. J. Plant Res.28, 155. 10.5958/2229-4473.2015.00098.1
52
JoshiN. C.GururaniP. (2023). A mini review on heavy metal contamination in vegetable crops. Int. J. Environ. Anal. Chem.12, 1–12. 10.1080/03067319.2023.2210058
53
JuradoC.Díaz-VivancosP.GregorioB. E.Acosta-MotosJ. R.HernándezJ. A. (2024). Effect of halophyte-based management in physiological and biochemical responses of tomato plants under moderately saline greenhouse conditions. Plant Physiol. Biochem.206, 108228. 10.1016/j.plaphy.2023.108228
54
KarakasS.CulluM.KayaC.DikilitasM. (2016). Halophytic companion plants improve growth and physiological parameters of tomato plants grown under salinity. Pak. J. Bot.48, 21–28.
55
KarukuG. N.GacheneC. K. K.KaranjaN.CornelisW. N.VerplackeH. (2014). Effect of different cover crop residue management practices in soil moisture content under a tomato crop (Licopersicon esculentum). Trop. Subtrop. Agroecosyst.17, 509–523.
56
KathpaliaR.BhatlaS. C. (2018). “Plant mineral nutrition,” in Plant Physiology, Development and Metabolism (Singapore: Springer Singapore), 37–81. 10.1007/978-981-13-2023-1_2
57
KatiV.KaramaounaF.EconomouL.MylonaP. V.SamaraM.MitroiuM. D.et al. (2021). Sown wildflowers enhance habitats of pollinators and beneficial arthropods in a tomato field margin. Plants10, 1003. 10.3390/plants10051003
58
KayikciogluH. H.DumanI.AsciogulT. K.BozokalfaM. K.ElmaciÖ. L. (2020). Effects of tomato-based rotations with diversified pre-planting on soil health in the Mediterranean soils of Western Turkey. Agric. Ecosyst. Environ.299, 1–10. 10.1016/j.agee.2020.106986
59
KhasanovS.KulmatovR.LiF.van AmstelA.BartholomeusH.AslanovI.et al. (2023). Impact assessment of soil salinity on crop production in Uzbekistan and its global significance. Agric. Ecosyst. Environ.342, 108262. 10.1016/j.agee.2022.108262
60
Khashi u RahmanM.WangX.GaoD.ZhouX.WuF. (2021). Root exudates increase phosphorus availability in the tomato/potato onion intercropping system. Plant Soil464, 45–62. 10.1007/s11104-021-04935-8
61
KhokharS.RolaniaK. (2022). Efficacy of different management modules against tomato fruit borer, Helicoverpa armigera (Hübner). Int. J. Trop. Insect Sci.42, 2731–2738. 10.1007/s42690-022-00806-6
62
Kihika-OpandaR.TchouassiD. P.Ng'ang'aM. M.BeckJ. J.TortoB. (2022). Chemo-ecological insights into the use of the non-host plant vegetable black-jack to protect two susceptible solanaceous crops from root-knot nematode parasitism. J. Agric. Food Chem.70, 6658–6669. 10.1021/acs.jafc.2c01748
63
KremenC.IlesA.BaconC. (2012). Diversified farming systems: an agroecological, systems-based alternative to modern industrial agriculture. Ecol. Soc. 17, art44. 10.5751/ES-05103-170444
64
KrugerD. H. M.FourieJ. C.MalanA. P. (2015). Control potential of brassicaceae cover crops as green manure and their host status for Meloidogyne javanica and Criconemoides xenoplax. South Afr. J. Enol. Viticulture36, 165–174. 10.21548/36-1-949
65
LiH. Y.ZhouX. G.WuF. Z. (2018). Effects of root exudates from potato onion on Verticillium dahliae. Allelopathy J.43, 217–222. 10.26651/allelo.j./2018-43-2-1142
66
LiN.GaoD.ZhouX.ChenS.LiC.WuF. (2020). Intercropping with potato-onion enhanced the soil microbial diversity of tomato. Microorganisms8, 834. 10.3390/microorganisms8060834
67
LiuS. Q.WuF. Z.WenX. Y. (2013). Allelopathic effects of root exudates of Chinese onion on tomato growth and the pathogen Fusarium oxysporum(Sch1) f.sp lycopersici. Allelopathy J.31, 387–403.
68
LuambanoN. D.NarlaR. D.WanjohiW. J.KimenjuJ. W.KerryB. R. (2015). Integrated management of root-knot nematodes in a tomato-maize crop system using the biocontrol fungus Pochonia clamydosporia. Crop Prot.71, 45–50. 10.1016/j.cropro.2015.01.013
69
LyuJ.JinL.JinN.XieJ.XiaoX.HuL.et al. (2020). Effects of different vegetable rotations on fungal community structure in continuous tomato cropping matrix in greenhouse. Front. Microbiol.11, 829. 10.3389/fmicb.2020.00829
70
MaX.DuM.LiuP.TangY.LiH.YuanQ.et al. (2021). Alternation of soil bacterial and fungal communities by tomato–rice rotation in hainan island in southeast of China. Arch. Microbiol.203, 913–925. 10.1007/s00203-020-02086-5
71
MancinelliR.RadicettiE.MuleoR.MarinariS.BravoI.PapettiP. (2019). Can hairy vetch cover crop affects arsenic accumulation in vegetable crops?Agriculture9, 89. 10.3390/agriculture9050089
72
MarouelliW. A.LageD. A.daC.GravinaC. S.Michereff FilhoM.de SouzaR. B. (2013). Sprinkler and drip irrigation in the organic tomato for single crops and when intercropped with coriander. Revista Ciência Agron.44, 825–833. 10.1590/S1806-66902013000400020
73
MarquezJ.HajihassaniA. (2023). Successional effects of cover cropping and deep tillage on suppression of plant-parasitic nematodes and soilborne fungal pathogens. Pest Manag. Sci.79, 2737–2747. 10.1002/ps.7450
74
Martínez-SiasV. A.Martínez-HernándezJ.deJ.Zúñiga-EstradaL.Martínez-MontoyaJ. F. (2022). Mejoradores de suelo salino-sódico y su efecto en el desarrollo de jitomate (Solanum lycopersicum). Ecosistemas y Recursos Agropecuarios 9, 3086. 10.19136/era.a9n1.3086
75
Mateos-FernándezR.PetekM.GerasymenkoI.JuteršekM.BaeblerŠ.KallamK.et al. (2022). Insect pest management in the age of synthetic biology. Plant Biotechnol. J.20, 25–36. 10.1111/pbi.13685
76
MauryaP. K.MalikD. S.SharmaA. (2019). “Impacts of pesticide application on aquatic environments and fish diversity,” in Contaminants in Agriculture and Environment: Health Risks and Remediation.Haridwar: Agro Environ Media - Agriculture and Ennvironmental Science Academy, 111–128.
77
McCartyD. G.Eichler InwoodS. E.OwnleyB. H.SamsC. E.WszelakiA. L.ButlerD. M. (2014). Field evaluation of carbon sources for anaerobic soil disinfestation in tomato and bell pepper production in tennessee. HortScience49, 272–280. 10.21273/HORTSCI.49.3.272
78
MianoR. N.AyeloP. M.MusauR.HassanaliA.MohamedS. A. (2022). Electroantennogram and machine learning reveal a volatile blend mediating avoidance behavior by Tuta absoluta females to a wild tomato plant. Sci. Rep.12, 8965. 10.1038/s41598-022-13125-0
79
MiheretS.SeidA.HailuN. (2019). Distribution and management of root-knot nematodes (Meloidogyne spp.) in tomato (Lycopersicum esculentum) in North Shoa Zone, Ethiopia. Pakistan J. Nematol.37, 123–134. 10.18681/pjn.v37.i02.p123-134
80
MinJ.LuK.SunH.XiaL.ZhangH.ShiW. (2016). Global warming potential in an intensive vegetable cropping system as affected by crop rotation and nitrogen rate. Clean44, 766–774. 10.1002/clen.201400785
81
MouraD. R.YamadaJ. K.de AlbuquerqueL. C.de Carvalho PontesN. (2020). Crop rotation reduces the density of volunteer plants in processing tomato fields and the inoculum of bacterial spot. Eur. J. Plant Pathol.156, 299–304. 10.1007/s10658-019-01868-y
82
MuchangaR. A.HirataT.ArakiH. (2019). Hairy vetch and livestock compost improve soil carbon and nitrogen, and fresh-market tomato yield. HortScience54, 1023–1030. 10.21273/HORTSCI13828-18
83
MuchangaR. A.HirataT.UchidaY.HatanoR.ArakiH. (2020a). Soil carbon and nitrogen and tomato yield response to cover crop management. Agron. J.112, 1636–1648. 10.1002/agj2.20098
84
MuchangaR. A.UchidaY.HirataT.HatanoR.ArakiH. (2020b). Dynamics of N derived from 15N-labeled rye in soil–tomato system as influenced by cover crop residue management. Hort J.89, 394–402. 10.2503/hortj.UTD-132
85
MutisyaS.SaidiM.OpiyoA.NgouajioM.MartinT. (2016). Synergistic effects of agronet covers and companion cropping on reducing whitefly infestation and improving yield of open field-grown tomatoes. Agronomy6, 1–14. 10.3390/agronomy6030042
86
NawazR.AbbasiN. A.HafizI. A.KhalidA. (2020). Increasing level of abiotic and biotic stress on Kinnow fruit quality at different ecological zones in climate change scenario. Environ. Exp. Bot.171, 103936. 10.1016/j.envexpbot.2019.103936
87
NeocleousD.NikolaouG.NtatsiG.SavvasD. (2021). Nitrate supply limitations in tomato crops grown in a chloride-amended recirculating nutrient solution. Agric. Water Manag.258, 107163. 10.1016/j.agwat.2021.107163
88
OndrasekG.RathodS.ManoharaK. K.GireeshC.AnanthaM. S.SakhareA. S.et al. (2022). Salt stress in plants and mitigation approaches. Plants11, 717. 10.3390/plants11060717
89
PadalaV. K.KumarP. S.RamyaN.JayanthiP. D. K. (2023). Aromatic plant odours of Anethum graveolens and Coriandrum sativum repel whitefly, Bemisia tabaci in tomato. Curr. Sci.124, 231–238. 10.18520/cs/v124/i2/231-238
90
PandeyS. K.NookarajuA.UpadhyayaC. P.GururaniM. A.VenkateshJ.KimD. H.et al. (2011). An update on biotechnological approaches for improving abiotic stress tolerance in tomato. Crop Sci.51, 2303–2324. 10.2135/cropsci2010.10.0579
91
PeckS.MittlerR. (2020). Plant signaling in biotic and abiotic stress. J. Exp. Bot.71, 1649–1651. 10.1093/jxb/eraa051
92
PouëtC.DeletreE.RhinoB. (2022). Repellency of wild oregano plant volatiles, plectranthus amboinicus, and their essential oils to the silverleaf whitefly, bemisia tabaci, on tomato. Neotrop. Entomol.51, 133–142. 10.1007/s13744-021-00921-y
93
QasemJ. R. (2019). Branched broomrape (Orobanche ramosa L.) control in tomato (Lycopersicon esculentum Mill.) by trap crops and other plant species in rotation. Crop Prot.120, 75–83. 10.1016/j.cropro.2019.02.021
94
QinS.LiuH.NieZ.RengelZ.GaoW.LiC.et al. (2020). Toxicity of cadmium and its competition with mineral nutrients for uptake by plants: a review. Pedosphere30, 168–180. 10.1016/S1002-0160(20)60002-9
95
RahimH. U.AkbarW. A.AlataloJ. M. (2022). A comprehensive literature review on cadmium (Cd) status in the soil environment and its immobilization by biochar-based materials. Agronomy12, 877. 10.3390/agronomy12040877
96
RashidA.SchutteB. J.UleryA.DeyholosM. K.SanogoS.LehnhoffE. A.et al. (2023). Heavy metal contamination in agricultural soil: environmental pollutants affecting crop health. Agronomy13, 1521. 10.3390/agronomy13061521
97
RatnadassA.AvelinoJ.FernandesP.LetourmyP.BabinR.DeberdtP.et al. (2021). Synergies and tradeoffs in natural regulation of crop pests and diseases under plant species diversification. Crop Prot.146, 105658. 10.1016/j.cropro.2021.105658
98
RazaA.CharaghS.Najafi-KakavandS.AbbasS.ShoaibY.AnwarS.et al. (2023). Role of phytohormones in regulating cold stress tolerance: physiological and molecular approaches for developing cold-smart crop plants. Plant Stress8, 100152. 10.1016/j.stress.2023.100152
99
RazaS. M. J.AkhterA.WahidF.HashemA.Abd-AllahE. F. (2022). Rhizophagus intraradices and tomato-basil companionship affect root morphology and root exudate dynamics in tomato under Fusarium wilt disease stress. Appl. Ecol. Environ. Res.20, 235–249. 10.15666/aeer/2001_235249
100
RobertsD.MattooA. (2018). Sustainable agriculture—enhancing environmental benefits, food nutritional quality and building crop resilience to abiotic and biotic stresses. Agriculture8, 8. 10.3390/agriculture8010008
101
RoyR. N.FinckA.BlairG. J.TandonH. L. S. (2006). Plant Nutrition for Food Security. A Guide for Integrated Nutrient Management. Rome: FAO.
102
SamaddarS.SchmidtR.TautgesN. E.ScowK. (2021). Adding alfalfa to an annual crop rotation shifts the composition and functional responses of tomato rhizosphere microbial communities. Appl. Soil Ecol.167, 104102. 10.1016/j.apsoil.2021.104102
103
SamedaniB.MeighaniF. (2022). Effect of cover crops residue on weed control and yield in conservation tillage tomato (Lycopersicon esculentum Mill.) production. Weed Biol. Manag.22, 59–67. 10.1111/wbm.12254
104
SandilS.ÓváriM.DobosyP.VetésiV.EndrédiA.TakácsA.et al. (2021). Effect of arsenic-contaminated irrigation water on growth and elemental composition of tomato and cabbage cultivated in three different soils, and related health risk assessment. Environ. Res.197, 111098. 10.1016/j.envres.2021.111098
105
ScaglioneJ.MonticoS.MonteroG. (2023). Efectos a corto plazo de los cultivos de cobertura sobre propiedades y macrofauna del suelo. Ecosistemas y Recursos Agropecuarios10, 3645. 10.19136/era.a10n2.3645
106
SchombergH. H.WhiteK. E.ThompsonA. I.BagleyG. A.BurkeA.GarstG.et al. (2023). Interseeded cover crop mixtures influence soil water storage during the corn phase of corn-soybean-wheat no-till cropping systems. Agric. Water Manag.278, 108167. 10.1016/j.agwat.2023.108167
107
SegreH.SegoliM.CarmelY.ShwartzA. (2020). Experimental evidence of multiple ecosystem services and disservices provided by ecological intensification in Mediterranean agro-ecosystems. J. Appl. Ecol.57, 2041–2053. 10.1111/1365-2664.13713
108
SeidA.FininsaC.MeketeT.DecraemerW.WesemaelW. M. L. (2015). Tomato (Solanum lycopersicum) and root-knot nematodes (Meloidogyne spp.) – a century-old battle. Nematology17, 995–1009. 10.1163/15685411-00002935
109
SelviA.RajasekarA.TheerthagiriJ.AnanthaselvamA.SathishkumarK.MadhavanJ.et al. (2019). Integrated remediation processes toward heavy metal removal/recovery from various environments-a review. Front. Environ. Sci.7, 66. 10.3389/fenvs.2019.00066
110
ShahidM.GowenS. R.PembrokeB. (2020). Evaluation of efficacy of Pasteuria penetrans alone and in combination with Verticillium chlamidosporium against Meloidogyne javanica. Plant Protection4, 117–124. 10.33804/pp.004.03.3411
111
ShehaA. M.El-MehyA. A.MohamedA. S.SalehS. A. (2022). Different wheat intercropping systems with tomato to alleviate chilling stress, increase yield and profitability. Annal. Agric. Sci.67, 136–145. 10.1016/j.aoas.2022.06.005
112
SinghA.PandeyH.PandeyS.LalD.ChauhanD.AparnaH.et al. (2023). Drought stress in maize: stress perception to molecular response and strategies for its improvement. Funct. Integr. Genomics23, 296. 10.1007/s10142-023-01226-6
113
SongY.ChenD.LuK.SunZ.ZengR. (2015). Enhanced tomato disease resistance primed by arbuscular mycorrhizal fungus. Front. Plant Sci.6, 1–13. 10.3389/fpls.2015.00786
114
SugiharaY.UenoH.HirataT.ArakiH. (2013). Uptake and distribution of nitrogen derived from hairy vetch used as a cover crop by tomato plant. J. Jap. Soc. Hortic. Sci.82, 30–38. 10.2503/jjshs1.82.30
115
SugiharaY.UenoH.HirataT.ArakiH. (2014). Hairy vetch derived-n uptake by tomato grown in a pot containing fast-and slow-release N fertilizer. J. Jap. Soc. Hortic. Sci.83, 222–228. 10.2503/jjshs1.CH-061
116
SugiharaY.UenoH.HirataT.KomatsuzakiM.ArakiH. (2016). Contribution of N derived from a hairy vetch incorporated in the previous year to tomato N uptake under hairy vetch-tomato rotational cropping system. Hort J.85, 217–223. 10.2503/hortj.MI-073
117
SumbergJ.GillerK. E. (2022). What is ‘conventional' agriculture?Glob Food Sec.32, 100617. 10.1016/j.gfs.2022.100617
118
SweellumT. A.NaguibD. M. (2023). Tomato potato onion intercropping induces tomato resistance against soil borne pathogen, Fusarium oxysporum through improvement soil enzymatic status, and the metabolic status of tomato root and shoot. J. Plant Dis. Prot.130, 245–261. 10.1007/s41348-022-00699-0
119
TamburiniG.BommarcoR.WangerT. C.KremenC.van der HeijdenM. G. A.LiebmanM.et al. (2020). Agricultural diversification promotes multiple ecosystem services without compromising yield. Sci. Adv.6, 1715. 10.1126/sciadv.aba1715
120
TogniP.MarouelliW. A.Inoue-NagataA. K.PiresC. S. S.SujiiE. R. (2018). Integrated cultural practices for whitefly management in organic tomato. J. Appl. Entomol.142, 998–1007. 10.1111/jen.12558
121
TogniP.VenzonM.MunizC.MartinsE.PalliniA.SujiiE. (2016). Mechanisms underlying the innate attraction of an aphidophagous coccinellid to coriander plants: Implications for conservation biological control. Biol. Control92, 77–84. 10.1016/j.biocontrol.2015.10.002
122
TomazA.PalmaP.AlvarengaP.GonçalvesM. C. (2020). “Soil salinity risk in a climate change scenario and its effect on crop yield,” in Climate Change and Soil Interactions, eds. P. M. N. Vara and M. Pietrzykowski (Amsterdam: Elsevier), 351–396.
123
TostiG.BenincasaP.FarneselliM.GuiducciM.OnofriA.TeiF. (2019). Processing tomato–durum wheat rotation under integrated, organic and mulch-based no-tillage organic systems: yield, N balance and N loss. Agronomy9, 718. 10.3390/agronomy9110718
124
TostiG.BenincasaP.FarneselliM.TeiF.GuiducciM. (2014). Barley–hairy vetch mixture as cover crop for green manuring and the mitigation of N leaching risk. Eur. J. Agron.54, 34–39. 10.1016/j.eja.2013.11.012
125
TringovskaI.YankovaV.MarkovaD.MihovM. (2015). Effect of companion plants on tomato greenhouse production. Sci. Hortic.186, 31–37. 10.1016/j.scienta.2015.02.016
126
TripathiR.TewariR.SinghK. P.KeswaniC.MinkinaT.SrivastavaA. K.et al. (2022). Plant mineral nutrition and disease resistance: a significant linkage for sustainable crop protection. Front. Plant Sci.13, 883970. 10.3389/fpls.2022.883970
127
Ur RahmanS.LiY.HussainS.HussainB.KhanW.-D.RiazL.et al. (2023). Role of phytohormones in heavy metal tolerance in plants: a review. Ecol. Indic.146, 109844. 10.1016/j.ecolind.2022.109844
128
Van PloegD.HeuvelinkE. (2005). Influence of sub-optimal temperature on tomato growth and yield: a review. J. Hortic. Sci. Biotechnol.80, 652–659. 10.1080/14620316.2005.11511994
129
VolpeV.ChitarraW.CasconeP.VolpeM. G.BartoliniP.MonetiG.et al. (2018). The association with two different arbuscular mycorrhizal fungi differently affects water stress tolerance in tomato. Front. Plant Sci.9, 1480. 10.3389/fpls.2018.01480
130
Warren-RaffaD.MiglioreM.CampanelliG.LeteoF.TrincheraA. (2022). Effects of faba bean strip cropping in an outdoor organic tomato system on soil nutrient availability, production, and N budget under different fertilizations. Agronomy12, 1372. 10.3390/agronomy12061372
131
WezelA.HerrenB. G.KerrR. B.BarriosE.GonçalvesA. L. R.SinclairF. (2020). Agroecological principles and elements and their implications for transitioning to sustainable food systems. A review. Agron. Sustain. Dev.40, 40. 10.1007/s13593-020-00646-z
132
WilliamsonV. M.ThomasV.FerrisH.DubcovskyJ. (2013). An aegilops ventricosa translocation confers resistance against root-knot nematodes to common wheat. Crop Sci.53, 1412–1418. 10.2135/cropsci2012.12.0681
133
WilsonM.LovellS. (2016). Agroforestry—the next step in sustainable and resilient agriculture. Sustainability8, 574. 10.3390/su8060574
134
WoodS. A.KarpD. S.DeClerckF.KremenC.NaeemS.PalmC. A. (2015). Functional traits in agriculture: agrobiodiversity and ecosystem services. Trends Ecol. Evol.30, 531–539. 10.1016/j.tree.2015.06.013
135
WoodwardE.Raij-HoffmanI.ScowK.TautgesN. (2022). Alfalfa reduces winter nitrate leaching relative to organic and conventional annual vegetable systems: resin bag field measurements and modeling with HYDRUS-1D. J. Soil Water Conserv.77, 450–465. 10.2489/jswc.2022.00155
136
WuX.WuF.ZhouX.FuX.TaoY.XuW.et al. (2016). Effects of intercropping with potato onion on the growth of tomato and rhizosphere alkaline phosphatase genes diversity. Front. Plant Sci.7, 846. 10.3389/fpls.2016.00846
137
XieY.WangL.YangL.YanW.HeZ.TangY.et al. (2021). Intercropping with Eclipta prostrata and Crassocephalum crepidioides decrease cadmium uptake of tomato seedlings. Int. J. Environ. Anal. Chem.101, 1231–1239. 10.1080/03067319.2019.1678606
138
YangJ.MattooA. K.LiuY.ZvomuyaF.HeH. (2023). Trade-offs of organic and organic-inorganic fertilizer combinations in tomato quality and yield: a global meta-analysis (1992–2021). Eur. J. Agron. 151, 126985. 10.1016/j.eja.2023.126985
139
ZareiE.FathiS. A. A.HassanpourM.GolizadehA. (2019). Assessment of intercropping tomato and sainfoin for the control of Tuta absoluta (Meyrick). Crop Prot.120, 125–133. 10.1016/j.cropro.2019.02.024
140
ZhouX.ZhangJ.Khashi u RahmanM.GaoD.WeiZ.WuF.et al. (2023). Interspecific plant interaction via root exudates structures the disease suppressiveness of rhizosphere microbiomes. Mol. Plant16, 849–864. 10.1016/j.molp.2023.03.009
141
ZhouY.CenH.TianD.WangC.ZhangY. (2019). A tomato and tall fescue intercropping system controls tomato stem rot. J. Plant Interact.14, 637–647. 10.1080/17429145.2019.1689582
142
ZhuJ. K. (2016). Abiotic stress signaling and responses in plants. Cell167, 313–324. 10.1016/j.cell.2016.08.029
Summary
Keywords
agrobiodiversity, agricultural diversification, agroecology, green manures, plant diversity, integrated pest management
Citation
Cruz-López V, Granados-Echegoyen CA, Pérez-Pacheco R, Robles C, Álvarez-Lopeztello J, Morales I, Bastidas-Orrego LM, García-Pérez F, Dorantes-Jiménez J and Landero-Valenzuela N (2024) Plant diversity as a sustainable strategy for mitigating biotic and abiotic stresses in tomato cultivation. Front. Sustain. Food Syst. 8:1336810. doi: 10.3389/fsufs.2024.1336810
Received
11 November 2023
Accepted
22 January 2024
Published
06 February 2024
Volume
8 - 2024
Edited by
Liming Ye, Ghent University, Belgium
Reviewed by
Autar Krishen Mattoo, Agricultural Research Service (USDA), United States
Updates
Copyright
© 2024 Cruz-López, Granados-Echegoyen, Pérez-Pacheco, Robles, Álvarez-Lopeztello, Morales, Bastidas-Orrego, García-Pérez, Dorantes-Jiménez and Landero-Valenzuela.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rafael Pérez-Pacheco cagranad@uacam.mx
†These authors share first authorship
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.