 Anurag Chaturvedi
Anurag Chaturvedi Vijay Shankar
Vijay Shankar Bibhu Simkhada
Bibhu Simkhada Rachel A. LymanPatrick FreymuthElisabeth HowanskyKatelynne M. Collins
Rachel A. LymanPatrick FreymuthElisabeth HowanskyKatelynne M. Collins Trudy F. C. Mackay*
Trudy F. C. Mackay* Robert R. H. Anholt*
Robert R. H. Anholt*- Center for Human Genetics and Department of Genetics and Biochemistry, Clemson University, Clemson, SC, United States
Arsenic is an ubiquitous environmental toxicant with harmful physiological effects, including neurotoxicity. Modulation of arsenic-induced gene expression in the brain cannot be readily studied in human subjects. However, Drosophila allows quantification of transcriptional responses to neurotoxins at single cell resolution across the entire brain in a single analysis. We exposed Drosophila melanogaster to a chronic dose of NaAsO2 that does not cause rapid lethality and measured survival and negative geotaxis as a proxy of sensorimotor integration. Females survive longer than males but show earlier physiological impairment in climbing ability. Single-nuclei RNA sequencing showed widespread sex-antagonistic transcriptional responses with modulation of gene expression in females biased toward neuronal cell populations and in males toward glial cells. However, differentially expressed genes implicate similar biological pathways. Evolutionary conservation of fundamental processes of the nervous system enabled us to translate arsenic-induced changes in transcript abundances from the Drosophila model to orthologous human neurogenetic networks.
1 Introduction
Arsenic is one of the most hazardous environmental toxicants, affecting over 140 million people globally, with computational estimates suggesting this number may exceed 200 million across more than 70 countries (Podgorski and Berg, 2020). Arsenic toxicity is linked to diverse adverse health outcomes (Kapaj et al., 2006; Jomova et al., 2011; Naujokas et al., 2013), including skin disorders (Kapaj et al., 2006; Naujokas et al., 2013), impaired vision (Freund et al., 2020), neurological damage (Zarazúa et al., 2010; Tyler and Allan, 2014; Anushree et al., 2023; Adebambo et al., 2025), endocrine disruption (Naujokas et al., 2013), respiratory illness (Ortiz et al., 2009; Naujokas et al., 2013; Nie et al., 2020), and gastrointestinal distress (Jomova et al., 2011). Biotransformation of arsenic into methylated derivatives leads to oxidative stress, which can result in genome instability and give rise to a variety of cancers (Muñiz Ortiz et al., 2011; Lee and Yu, 2016; Martinez and Lam, 2021). The accumulation of methylated derivatives also increases risk for cardiovascular disease (Kuo et al., 2021; Karachaliou et al., 2021). Arsenic metabolites can cross the placenta and the blood brain barrier to exert both prenatal and postnatal neurotoxic effects (Tolins et al., 2014; Prakash et al., 2016). These effects are accompanied by epigenetic modifications, which can persist trans-generationally (Rojas et al., 2015; Nohara et al., 2020; Ijomone et al., 2020).
Whereas most studies on model organisms use lethality as an endpoint, in human populations sublethal effects due to chronic exposure are more relevant yet poorly studied (Ganie et al., 2023; Yu et al., 2024). Here, we asked to what extent chronic exposure to arsenic that results in pre-lethal physiological impairments is accompanied by changes in gene expression in the central nervous system; whether such changes are restricted to few distinct neuronal populations or are widespread; whether they include both neuronal and glial cell populations; and whether arsenic-induced changes in transcript abundances are identical for males and females or sexually dimorphic.
Drosophila melanogaster is a powerful model for toxicogenomic studies (Rand, 2010; Rand et al., 2023), since the genetic background and exposure can be controlled precisely and transcriptional responses to neurotoxins can be quantified at single cell resolution across the entire brain in a single analysis. The translational potential of the Drosophila model for arsenic neurotoxicity is underscored by well-established effects of arsenic on mitochondrial function (Prakash et al., 2016; Rahman et al., 2021; Thakur et al., 2021) and oxidative stress-induced DNA damage (Lee and Yu, 2016; Rand et al., 2023), processes that are evolutionarily conserved across phyla. Whereas arsenic and its derivatives, especially meta-arsenite (NaAsO2), are recognized as potent neurotoxins (Tyler and Allan, 2014; Freund et al., 2020; Anushree et al., 2023; Adebambo et al., 2025), their effects on the modulation of gene expression across the human brain cannot be readily investigated. We subjected a standard laboratory strain of D. melanogaster to a chronic dose of NaAsO2 and measured lifespan and climbing ability. We then performed single-nuclei RNA sequencing on male and female brains (Figure 1). We found that the transcriptional response to arsenic neurotoxicity is characterized by widespread sex-specific and sex-antagonistic modulation of gene expression.
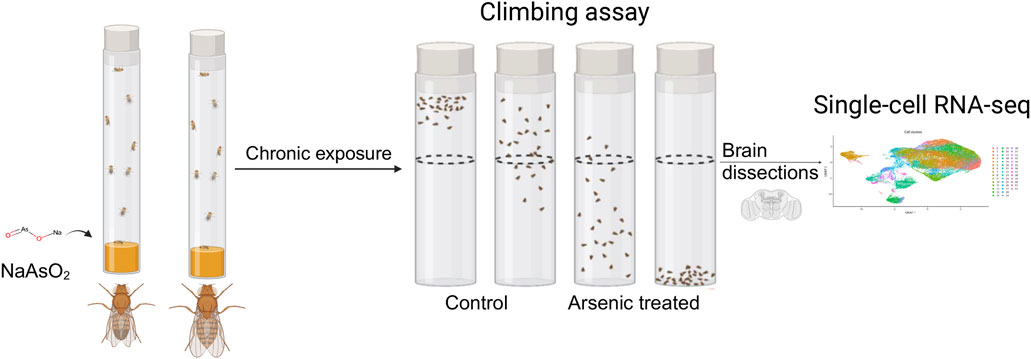
Figure 1. Diagram of the experimental design. Male and female flies aged between 3 and 5 days were exposed to 250 μM NaAsO2, followed by a climbing assay to assess locomotor activity. Single-nuclei RNA-seq analysis was then performed on dissected brains to identify molecular signatures linked to neurotoxicity.
2 Materials and methods
2.1 Drosophila stock
Canton S (B) flies (CSB) were reared on standard cornmeal-yeast-molasses-agar medium at 25°C under a 12:12 h light:dark cycle and 50% humidity. Five males and five females were placed in each vial to mate for 2 days before being cleared to prevent overcrowding. Progeny were collected after eclosion by brief exposure to CO2 anesthesia and aged for 3–5 days prior to experimentation.
2.2 Effective range determination
Sodium (meta) arsenite (NaAsO2) (CAS Number: 7784-46-5) was purchased from Sigma-Aldrich, and a range-finding assessment was conducted using CSB flies to determine the concentration of NaAsO2 that supported survival after 24 and 48 h of exposure for both sexes, following a modified version of a previously established protocol (Holsopple et al., 2023). Two weeks prior to exposure, controlled adult density vials were set up with five female and five male CSB flies per vial, and after 48 h, flies were transferred to fresh vials to promote egg-laying. Adults were removed 48 h later, and after 7 days, progeny were anesthetized using CO2 and sorted into groups of 15–20 same-sex flies. At three to 5 days post-eclosion, following a minimum 3-h recovery period from anesthesia, flies were food-deprived for 20 h in vials containing three pieces of 20 mm Whatman #1 filter paper (Cytiva Cat. #1001-020) wetted with 0.35 mL distilled water. Subsequently, flies were exposed for 24 and 48 h to 0, 100, 150, 200, 250, 300, 350, 400, 600, 800,1200 and 1600 µM NaAsO2 by transferring them to vials containing three pieces of 20 mm filter paper wetted with 0.35 mL of an exposure solution comprising NaAsO2,1.5% yeast, 4% sucrose, and distilled water, with four biological replicates of 15–20 individuals per sex/concentration. Survival was recorded for each replicate and averaged across all replicates, with standard errors calculated as
2.3 Climbing assay
Negative geotaxis was quantified using a climbing assay with a vertically oriented Benzer’s counter-current apparatus on 3–5-day old flies. Control flies were maintained on standard medium at 25°C under a 12:12 h light:dark cycle and 50% humidity without chemical exposure. Treated flies were maintained under the same conditions, but their medium was supplemented with 250 µM NaAsO2. Flies were grouped by sex and exposure day into sets of 20 individuals, and the climbing activity of both control and treatment groups (three replicates each for each sex) was recorded daily in the morning from 9 to 11 a.m. over six consecutive days. A new set of flies was used on each assay day. For each replicate, flies were collected without anesthesia and allowed to acclimate for 1–2 h. During the assay, flies were gently tapped to the bottom of the starting tube and given 15 s to climb into the distal tube. Flies reaching the distal tube were transferred to the next tube, and this process was repeated seven times. At the end of the assay, flies in all eight tubes were frozen, counted, and scored from 1 (indicating no successful crossings) to 8 (indicating successful crossings in all seven trials).
Climbing scores were analyzed using a mixed-effects ANOVA with the PROC MIXED procedure in SAS™ Studio v3.9. The full model used the Type III method to test significance and was defined as follows: Y = µ + treatment (T, fixed) + Sex (S, fixed) + Day (D, fixed) + TxS, TxD, SxD, TxSxD (all fixed) + rep(TxSxD) (random) + ε. In addition, a reduced ANOVA model Y = µ + T (fixed) + rep(T) (random) + ε was used to assess climbing performance by day and sex.
2.4 Lifespan assay
Lifespan assays were conducted on males and females separately by exposing them to either standard medium or medium supplemented with 250 µM NaAsO2. To prevent overcrowding, five males and five females were placed in each vial to mate for 2 days. Progeny were collected after eclosion, and lifespan was assessed by transferring them without anesthesia to new vials with fresh medium daily and recording deaths until all NaAsO2-treated flies had died. We measured 20 replicate vials per treatment, each containing 5 mL of culture medium and three same-sex flies. Survival was recorded as means across all replicates. The full model used the Type III method to test lifespan significance and was defined as follows: Y = µ + treatment (T, fixed) + Sex (S, fixed) + TxS (fixed) + rep(TxS) (random) + ε were analyzed using a mixed-effects ANOVA with the PROC MIXED procedure in SAS™ Studio v3.9.
2.5 Brain dissection and single nucleus RNA sequencing (snRNA-seq)
We anesthetized CSB flies on ice and dissected 10 brains per sex after 6 days of exposure to control medium or medium supplemented with 250 µM NaAsO2 in cold Dulbecco’s phosphate-buffered saline (PBS). Brains were flash frozen in a dry ice-ethanol bath, and stored at −80°C. We prepared the samples for snRNA-seq as described in the 10X Genomics Gene Expression (GEX) protocol (CGOOO338). 200 μL of chilled Lysis Buffer was added to each tube containing the brain samples and incubated on ice for 5 min. The brains were mechanically dissociated through stepwise trituration with a P200 micropipette (5 times), a 23G needle (5 times), and a 27G needle (5 times). The dissociated samples were passed through a 30 μm MACS SmartStrainer (Miltenyi Biotec B.V. & Co. KG), followed by rinsing with 800 μL of lysis buffer, and collected into 5 mL tubes. Each sample was then passed through a 10 μm strainer (Celltrics, Görlitz, Germany), followed by rinsing with 1 mL PBS + 1% BSA, and collected into a new 5 mL tube. A 20 μL aliquot was removed for quantitative analysis, and the remaining sample was centrifuged at 500 rcf for 5 min at 4°C. After centrifugation, the supernatant was removed, and the pellet was resuspended in a chilled diluted nuclei buffer (Baker et al., 2021). Single nuclei were counted using a hemocytometer with trypan blue exclusion and proceeded to GEM generation using the microfluidics Chromium Controller (10X Genomics, Pleasanton, CA). Libraries were prepared following the 10X Genomics v3.1 protocols. Fragment sizes were determined using Agilent Tapestation High Sensitivity D5000 for amplified cDNA (Agilent Technologies Inc., Santa Clara, CA) and High Sensitivity D1000 ScreenTape assay for libraries. Concentrations of amplified cDNA and final libraries were measured using a Qubit 1X dsDNA HS kit (Invitrogen, Waltham, MA) and cDNA amplification and indexing PCR were performed with 12 cycles each. Final libraries were sequenced on an Illumina NovaSeq6000 (Illumina Inc., San Diego, CA).
2.6 Sn-RNA-seq analysis
The mkfastq pipeline in Cell Ranger v3.1 (10X Genomics) was used to convert BCL files into demultiplexed FASTQ files, and the mkref pipeline indexed the D. melanogaster reference genome (release 6, GCA_000001215.4 from NCBI Genbank). Alignment was performed using the count pipeline in Cell Ranger v3.1 with the expected cell count set to 5,000. Raw expression counts were imported into Seurat v3.10 in R (Baker et al., 2021; Mokashi et al., 2021; Butler et al., 2018), normalized using the scTransform (Hafemeister and Satija, 2019) pipeline with regularized negative binomial regression, and integrated using the SCT method.
Cell clustering was performed using the Louvain algorithm (Blondel et al., 2008) with a resolution parameter 1 to optimize community formation. Resolution parameter was selected by iterating between 0.1 and 2.0 to identify stable plateaus that correspond to numbers of cell-type clusters. Dimensionality reduction was achieved using the RunUMAP and FindNeighbors functions with 10 dimensions. Thirty-five cell-type clusters were identified through unsupervised clustering (FindClusters), and annotation of the top 20 markers for each cell type was based on the most frequently occurring identity for cell types using expression scores from BGEE (Bastian et al., 2025). Only cell types with an expression score >90 were considered. Annotation of cell type was based on the most frequently occurring identity based on the top 20 marker genes. Clusters 0 and 13 were further subclustered at a resolution of 0.2 to identify the heterogeneous cell identities, resulting in 9 and 4 subclusters, respectively. The subclusters are denoted by appending the subcluster number to the cluster ID, separated by an underscore (“_”).
2.7 Analysis of NaAsO2-induced differential expression of genes and genetic networks in the fly brain
Pearson residuals from the scTransform (Hafemeister and Satija, 2019) pipeline were used for differential expression analysis. The MAST algorithm in the FindMarkers function was employed to calculate differential expression within clusters after merging clusters with the same identity, incorporating cellular detection rates as a covariate (Finak et al., 2015). P-values were adjusted for multiple-hypothesis testing using the Benjamini–Hochberg method (Benjamini and Hochberg, 1995) and adjusted P-values ≤ 0.05 were considered statistically significant.
Gene enrichment analyses were conducted using PAthway, Network, and Gene-set Enrichment Analysis (PANGEA) (Hu et al., 2023). The analysis incorporated GO hierarchy categories (GO Biological Processes, GO Molecular Function, GO Cellular Component) and pathway resources (KEGG) (Kanehisa et al., 2025) Pathway D.mel, PANTHER (Mi and Thomas, 2009) pathway D.mel, REACTOME (Grenter et al., 2024) pathway) to identify overrepresented functional categories.
These analyses were performed separately for upregulated and downregulated genes in each cluster and sex, with results further filtered using a Benjamini–Hochberg (Benjamini and Hochberg, 1995) adjusted P-value threshold of ≤ 0.05.
The scaled data generated from the sctransform pipeline (Hafemeister and Satija, 2019) for differentially expressed genes across all clusters in male and female samples were extracted to construct co-expression networks. Pairwise Spearman correlations were computed using these scaled datasets, and the resulting correlation networks were visualized with Cytoscape version 3.7.2 (Shannon et al., 2003). To create protein-protein interaction networks, gene IDs were converted to gene symbols using the FlyBase Consortium’s “Query-by-symbols/ID” tool. Interactions between gene products were then calculated by filtering for differentially expressed genes from the known physical interaction database from FlyBase (release v2024_v2) within Cytoscape (Shannon et al., 2003). In addition, genes that were connected to at least six differentially expressed genes were computationally recruited and included within the network. The network was subclustered using MCODE plugin and default parameters (Bader and Hogue, 2003). Smaller MCODE clusters with high degree of interconnectivity were merged. Human orthologs for Drosophila genes were identified using the Drosophila RNAi Screening Center Integrative Ortholog Prediction Tool (DIOPT) v9.0 (Hu et al., 2011). Multiple human orthologs for the same Drosophila gene were resolved by selecting the human gene with the highest DIOPT score. Functional characterization of the subclusters were accomplished using Gene Ontology enrichment analysis and over-representation tests. Only categories with Benjamini–Hochberg adjusted P-value ≤ 0.05 for the over-representation tests were considered.
To compare protein-protein interaction networks between Drosophila and humans, the Drosophila network was filtered for nodes with predicted human orthologs (DIOPT ≥ 3). String-DB v11.5 interaction database was filtered for these human orthologs as well as a combined interaction score threshold ≥500. The threshold for the combined interaction score was determined by constructing a histogram for distribution of the scores in the filtered network. The two networks were computationally compared using the DyNet plugin within Cytoscape (Goenwan et al., 2016). We used a Monte Carlo Permutation Procedure to assess the statistical significance of the observed connectivity within the network composed of the shared edges. Briefly, we generated a null distribution of network connectivity indices (average number of neighbors) from randomly sampled subnetworks from the score-filtered protein-protein interaction database, with the same number of genes as the observed network. P-value was calculated as the ratio between the rank of the observed connectivity index and the total number of permutations (N = 10,000). Construction of the null distribution using randomly sampled networks and calculation of the connectivity indices were performed using the igraph R package v1.4.2 in R.
3 Results
3.1 Chronic arsenic exposure reduces lifespan and climbing ability
We generated survival curves using a range of NaAsO2 concentrations and identified 250 µM as the highest concentration that showed no mortality after 24 h of exposure in the w1118 Canton S (B) (CSB) laboratory strain (Figure 2). Prolonged exposure to this concentration resulted in 50% mortality by day 15 (Figure 3A; Supplementary Table 1) with significant effects of treatment (P < 0.0001) and sex × treatment (P < 0.0004) (Supplementary Table 2). To further assess the phenotypic effects of NaAsO2 exposure, we performed a climbing assay to quantify locomotor activity driven by negative geotaxis. We observed a sexually dimorphic physiological response to chronic NaAsO2 exposure, with females exhibiting a significant reduction in climbing ability as early as day 1, while males showed a decline in performance only after 6 days (Figure 3B; Supplementary Tables 2, 3). Thus, chronic exposure to concentrations of NaAsO2 below acute lethal levels can induce neurotoxicity that leads to adverse and sex-specific effects on survival and locomotor function.
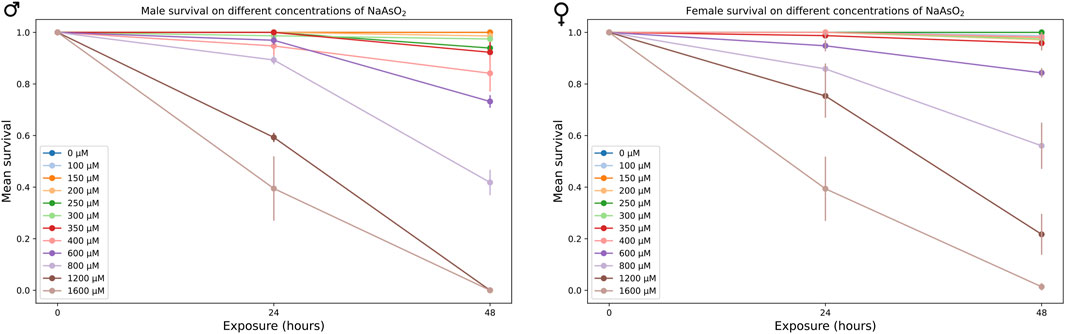
Figure 2. Mean survival of male and female CSB flies exposed to varying concentrations of NaAsO2, after 24 and 48 h.
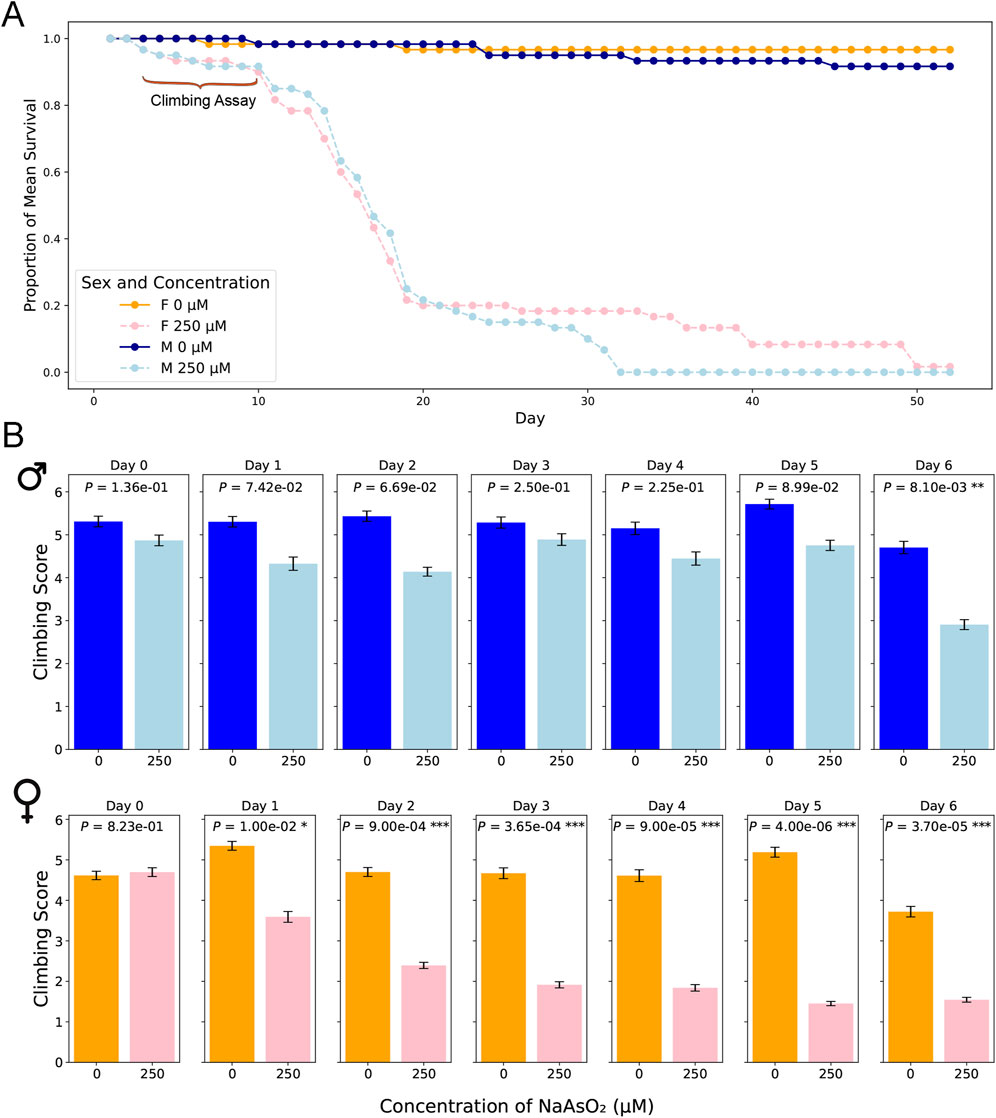
Figure 3. Lifespan and locomotion upon chronic NaAsO2, exposure. (A) The impact of NaAsO2 exposure on survival of male and female D. melanogaster exposed to control medium or medium supplemented with 250 μM NaAsO2. (B) The impact of NaAsO2 exposure on climbing ability.
3.2 Sexually antagonistic transcriptional responses to chronic NaAsO2 exposure in the Drosophila brain at single cell resolution
We performed snRNA-seq analysis for pools of brains from CSB males and females collected after 6 days of exposure to 250 µM NaAsO2 and from control flies of the same age not exposed to NaAsO2. We profiled whole brain transcriptomes of 68,709 single nuclei (Supplementary Table 4) and identified 35 cell clusters representing 20 major neural and glial cell types, including those from the optic lobe, mushroom body, and blood-brain barrier (Figure 4; Supplementary Figure 1; Supplementary Table 5). Our differential expression analysis revealed changes in 767 genes, with 53.7% (412 genes) with altered transcript abundances in both males and females, 18.6% (143 genes) unique to females, and 27.6% (212 genes) unique to males (Supplementary Figures 2, 3; Supplementary Table 6). The differentially expressed genes were biased toward neurons in females and glia in males (Figure 5).
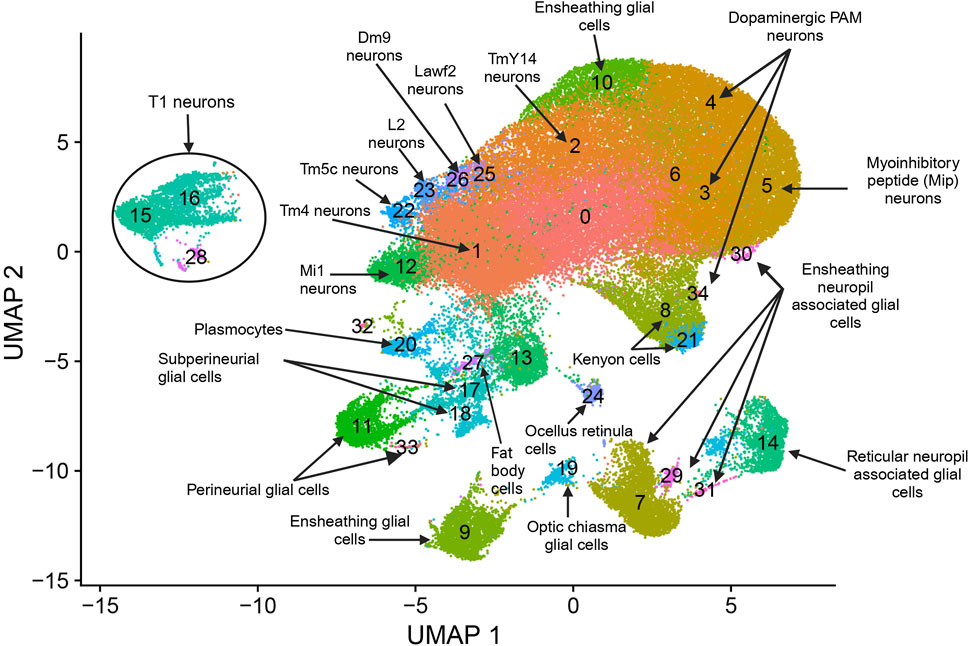
Figure 4. snRNA-seq analysis of brains from flies exposed to chronic NaAsO2, exposure. Cells were clustered according to their gene expression patterns using the unsupervised Shared Nearest Neighbor (SNN) clustering algorithm. Each dot represents an individual cell, with colors indicating the cluster to which the cell belongs. Cell type identification for each cluster was performed by annotating the top 20 marker genes. These markers were determined based on the most frequently occurring cell type identities using expression scores obtained from the BGEE database (Bastian et al., 2025).
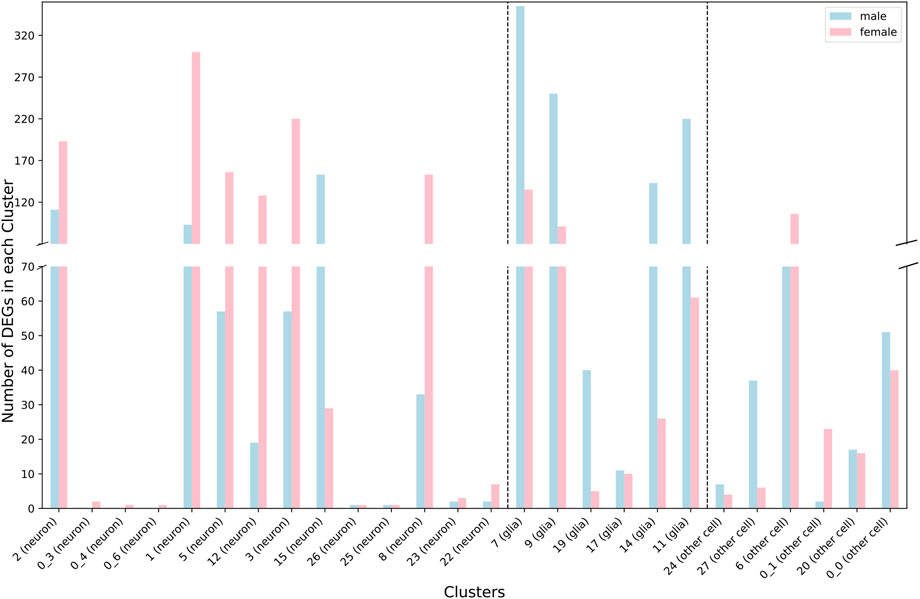
Figure 5. Distribution of differentially expressed genes between neurons and glia across cell clusters for males (blue) and females (pink) separately.
We observed that most gene expression differences between control and NaAsO2-treated flies were sexually antagonistic, with 94.7% (390 of the 412 shared genes) of differentially expressed genes in females showing downregulation, while 82.7% (341 of the 412 shared genes) in males exhibited upregulation across cell types (Figure 6; Supplementary Figure 4). Cell type-specific sex-specific expression was also evident, with 10 genes in females and 26 genes in males exhibiting expression profiles in multiple distinct cell types (Supplementary Table 7).
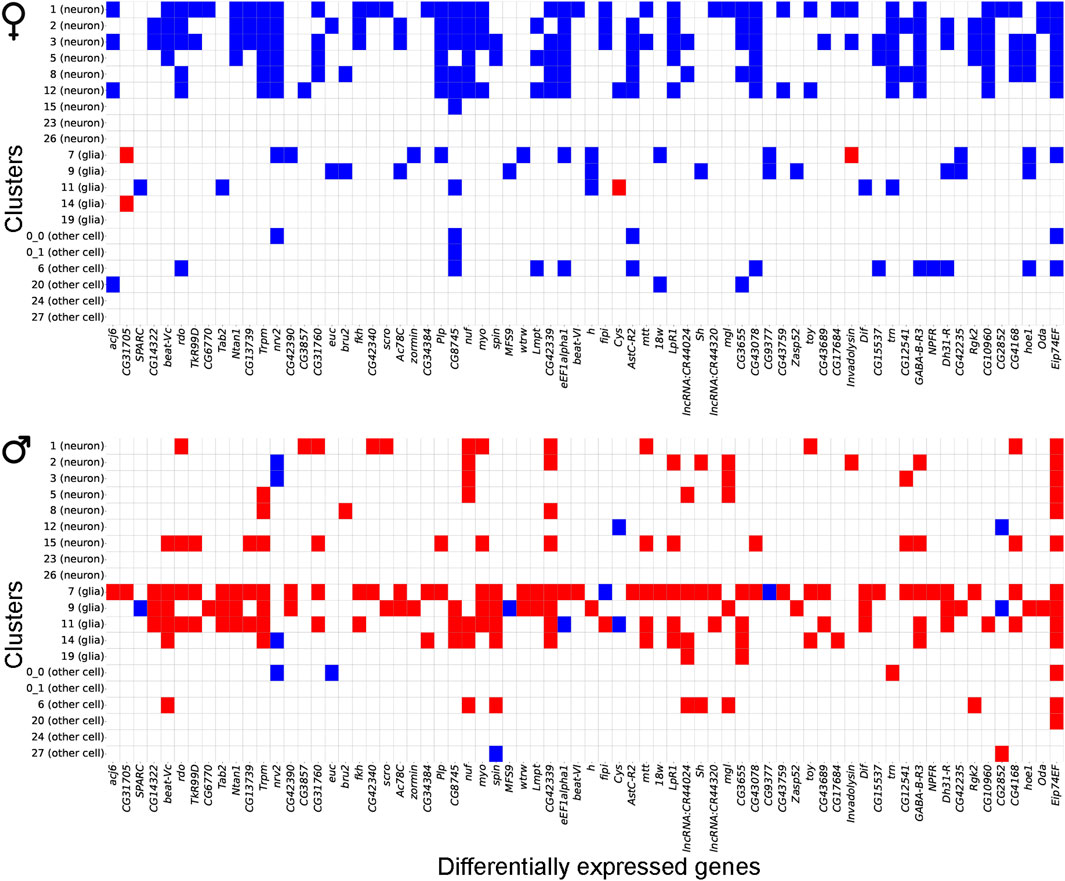
Figure 6. Gene expression patterns between males and females following chronic NaAsO2, exposure. Heatmap showing antagonistic gene expression patterns between males and females for 70 representative genes out of 412 shared DEGs across neuronal and glial cell types.
Further analysis of differentially expressed gene profiles revealed dysregulation of genes associated with all major neurotransmitter receptor and transport systems, including cholinergic, dopaminergic, GABAergic, and glutamatergic systems (Figure 7). These NaAsO2-induced changes in gene expression involved receptor subunits and transporters critical for neurotransmitter synthesis, release, reuptake, and signaling (Figure 7). We observed downregulation of both inhibitory and excitatory neurotransmitter-associated genes in both sexes. For example, Vglut (vesicular glutamate transporter), Vmat (vesicular monoamine transporter), and Gat (GABA transporter) were downregulated in both sexes, albeit in different cell types. Thus, chronic exposure to NaAsO2 leads to widespread disruption in synaptic communication.
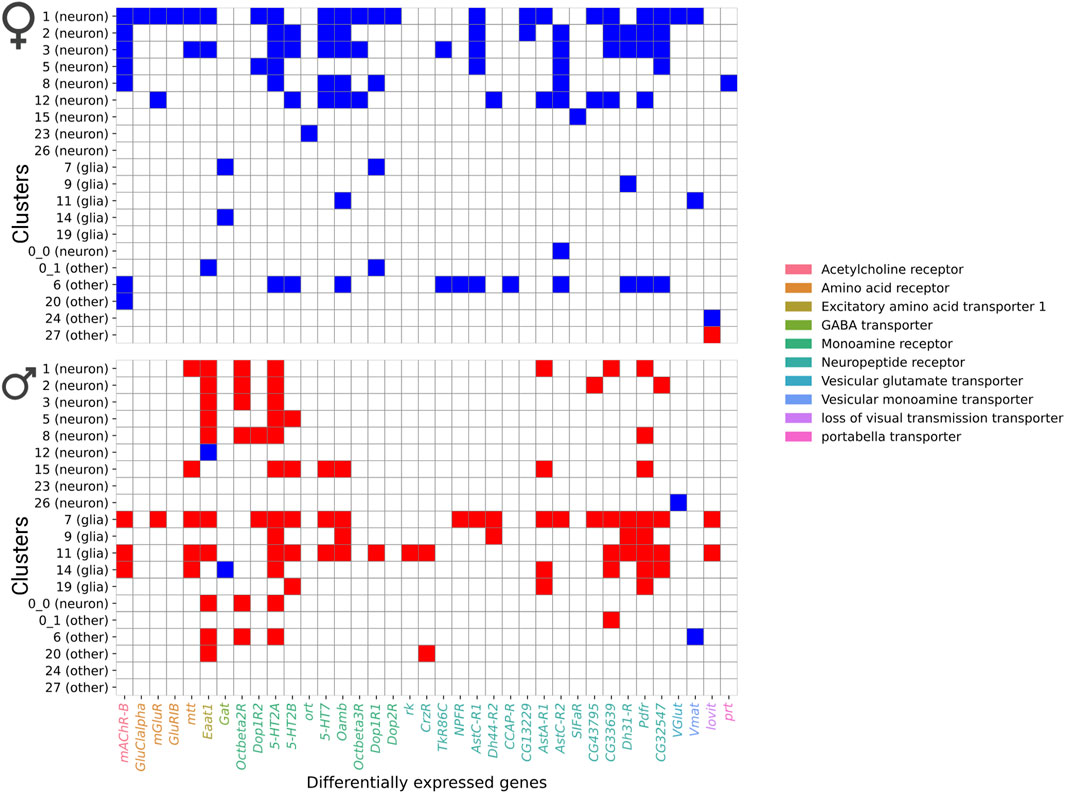
Figure 7. Heatmap showing differentially expressed genes in neurotransmitter receptor and transporter categories. Blue shows downregulated genes and red shows upregulated genes.
3.3 Differential expression of genes associated with detoxification
The Drosophila fat body, functionally analogous to mammalian adipose tissue and liver, plays a central role in heavy metal detoxification (Musselman et al., 2013). We recovered head fat body tissue in our dissected brain samples. Metallothionein (Mtna) and Turandot (Tot) genes are critical for responding to heavy metal toxicity (Mahapatra and Rand, 2012). We observed the highest upregulation of gene expression for Mtna in the fat body (cluster 27), with a 42-fold increase in males and a 43-fold increase in females. Conversely, we found significant downregulation for TotA (70-fold) and TotC (48-fold) in the fat body of males (Supplementary Table 6).
Functional enrichment analysis of upregulated genes in males revealed pathways related to the metabolism of xenobiotics by cytochrome P450 and members of the Glutathione S-transferase (GST) gene family (Supplementary Table 8). Cyp28d1 features prominently in both males and females, where it is upregulated in Cluster 7, associated with ensheathing neuropil-associated glial cells. In males, Cyp28d1 is also upregulated in optic chiasma glial cells (Cluster 19) and downregulated in fat body cells (Cluster 27). Expression of Cyp28a5 is altered in Clusters 7 and 9 in males. Additional members of the cytochrome P450 family (Cyp4g15, Cyp6w1 and Cyp311a1) also show upregulation in male glial cells (Cluster 17). In females, expression of GstD1 is downregulated in Kenyon cells (Cluster 8), while it is upregulated in fat body cells in males (Cluster 27). Similarly, GstE6 and GstE9 are both upregulated in fat body cells in males, whereas GstD9 is downregulated in ensheathing glial cells (Cluster 9).
Members of the UDP-glucuronosyl transferase (UGT) gene family also show differential changes in brain transcript abundances in flies exposed to NaAsO2 (Supplementary Table 6). In males, expression of Ugt49C1 is increased in fat body cells (Cluster 27) and ensheathing glial cells (Cluster 9) and perineurial glial cells (Cluster 11). Conversely, expression of Ugt35B1 is reduced in males in ensheathing glial cells (Cluster 9) and ensheathing neuropil-associated glial cells (Cluster 7). In females, Ugt35E2 is downregulated in ensheathing glial cells (Cluster 9), and Ugt35B1 is reduced in ensheathing neuropil-associated glial cells (Cluster 7). These patterns indicate largely cell-and sex-specific roles of NaAsO2-induced Cytochrome P450s, GSTs, and UGTs in detoxification.
Complementing these detoxification enzymes, transcripts encoding MsrA (Methionine sulfoxide reductase A) and Sod3 (Superoxide dismutase 3), which neutralize superoxide radicals generated during oxidative stress (Orr and Sohal, 1994; Ruan et al., 2002; Kirby et al., 2002), are upregulated in males, particularly in fat body cells (Cluster 27), with upregulation in ensheathing glial cells (Cluster 9), perineurial glial cells (Cluster 11), and ensheathing neuropil-associated glial cells (Cluster 7). Expression of Sestrin (Sesn), which plays a vital role in oxidative stress response (Chen et al., 2021), is downregulated in females in Myoinhibitory peptide (MIP) neurons, ensheathing neuropil-associated glial cells, and dopaminergic PAM neurons; and upregulated in males in ensheathing glial cells, reticular neuropil-associated glial cells, ensheathing neuropil-associated glial cells, and perineurial glial cells.
3.4 Differential expression of genes associated with the blood brain barrier and ensheathing glia
The Drosophila blood-brain barrier (BBB) consists of perineurial and sub-perineurial cells, which play critical roles in maintaining neural homeostasis (Hindle and Bainton, 2014; Calderon et al., 2022; Contreras et al., 2024). In perineurial cells (Cluster 11), downregulated genes were enriched in the Notch signaling pathway, previously shown to be essential for maintaining the BBB integrity in subperineurial glia by regulating cell proliferation, differentiation, and the expression of tight junction proteins (Currier et al., 2023). In males, upregulated genes in these cells were predominantly enriched in the heterotrimeric G-protein signaling pathways, specifically Gαi - and Gαs-mediated signaling pathways (Supplementary Table 8).
BBB cells establish contacts with ensheathing neuropil-associated glial cells (Jang et al., 2017). In ensheathing glial cells, NaAsO2 exposure elicited distinct sex-specific responses. In females, downregulated genes were enriched in pathways associated with the transport of small molecules, SLC-mediated transmembrane transport, and GABA-B receptor activation. In contrast, upregulated genes in males were enriched in heterotrimeric G-protein signaling pathways and the endothelin signaling pathway. Notably, components of GPCR signaling, Notch signaling, MAPK signaling and adenylate cyclase inhibitory pathways were enriched in ensheathing glial cells of both sexes (Supplementary Table 8). However, these pathways exhibited antagonistic responses in differential gene expression patterns between males and females (Supplementary Table 6).
3.5 Differential expression of genes associated with vision and locomotion
Since there were profound effects of chronic NaAsO2 exposure on negative geotaxis (Figure 3), we examined the extent to which genes associated with sensorimotor integration and locomotion showed NaAsO2-induced changes in gene expression in the brain. We identified 102 genes in 10 male cell clusters (0_0, 1, 2, 6, 7, 8, 9, 11, 14, 15) and 96 genes in nine female clusters (1, 2, 3, 5, 7, 8, 9, 11, 22), enriched for locomotion-related functions in both neuronal and glial cell types (Supplementary Table 8). We observed the most significant responses to NaAsO2 exposure in genes involved in memory, vision, and motion. Notably, the Rhodopsin 2 (Rh2) gene exhibited the largest downregulation, particularly in the ocellus retinula cells, with an average 25-fold and 13-fold decrease in males and females, respectively (Supplementary Table 6). Genes involved in these processes were organized in co-expression networks, highlighting multiple neuronal cell types in the optic lobes implicated in vision and locomotion, including Tm4, TmY14, T1, Tm5c, L2, Lawf2, Dm9, LC17, T3, Tm9 and Mi1 neurons (Liu C. et al., 2012) (Figure 8).
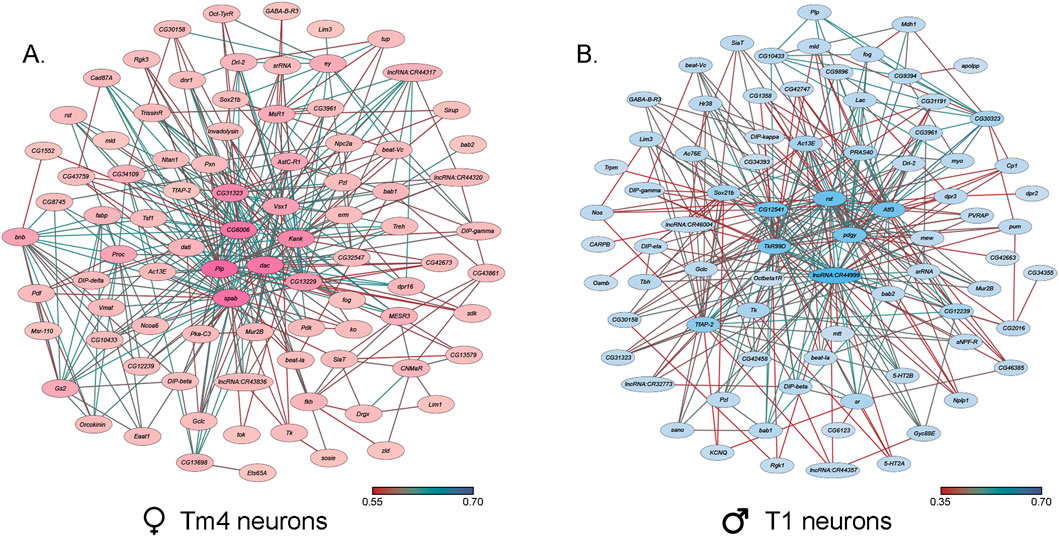
Figure 8. Co-expression networks of neurons in the optic lobes that exhibit the largest responses to NaAsO2, exposure. (A) Co-expression network of Tm4 neurons in female cluster 1. (B) Co-expression network of T1 neurons in male cluster 15. The darker colors in both co-expression networks represent a higher number of connections.
Expression of Arrestin 2 (Arr2), associated with phototransduction, was downregulated in photoreceptor neurons in females and multiple male neuron types, including myoinhibitory peptide (MIP) neurons, which regulate reproductive behaviors (Burke et al., 2012), and dopaminergic PAM neurons, critical for reward processing and learning (Yamagata et al., 2016; Mohr et al., 2018; Silva et al., 2021) (Supplementary Table 6). In males, expression of the Ndae1 (Na+/H+ antiporter) gene, which can mediate clearance of toxic arsenic anions (Schoofs et al., 2017), is upregulated in optic chiasma glial cells, reticular neuropil-associated glial cells, and Tm4 neurons. In females, Ndae1 expression is downregulated in Kenyon cells, MIP neurons, and dopaminergic PAM neurons. Additionally, differentially expressed genes in Kenyon cells, which mediate experience-dependent modification of behavior, showed sex-specific responses in G protein-coupled receptor signaling pathways. These results emphasize the wide-ranging effects of NaAsO2 exposure on neuronal function, with distinct impacts on cells associated with sensory processing and motor coordination.
3.6 Global protein-protein interaction networks
Although the arsenic-induced changes in transcript abundances differ in a sex-specific manner, the overarching biological processes disrupted by NaAsO2 exposure are shared between males and females (Supplementary Table 9). For example, in pathways related to neurotransmitter signaling and synaptic transmission, genes involved in receptor activation and signal propagation were upregulated in males (Figure 7). In contrast, genes associated with neurotransmitter synthesis and vesicle transport were predominantly downregulated in females. Similarly, G protein-coupled receptor (GPCR) signaling and calcium signaling in oxidative stress response pathways were enriched in both sexes but were modulated through different sets of genes.
We analyzed global protein-protein interaction networks of differentially expressed gene products across all cell clusters for both males and females to identify biological functions impacted by NaAsO2 exposure. Among differentially expressed genes in the Drosophila brain 72.35% had human orthologs. Despite pronounced sexual dimorphism and antagonism in gene expression responses to arsenic, arsenic-induced neurotoxicity affected the same pathways in both males and females but involved distinct components within these pathways. We constructed global protein-protein interaction networks in which we computationally recruited known physically interacting partners with the products of the differentially expressed genes (Figure 9). The global network consists of 258 nodes and reveals distinct subnetworks, including a large subnetwork associated with neuronal development and differentiation, a subnetwork of cell adhesion proteins recruited through cDIP, and a cluster of ribosomal proteins (Figure 9). Based on evolutionarily conserved orthology, we constructed a human network corresponding to the Drosophila global protein-protein interaction network (Figures 10A,B). Comparisons between these networks reveal common edges that show a conserved core structure of protein-protein interactions associated with the transcriptional response to arsenic neurotoxicity (Figure 10C). These include ribosomal protein interactions, signal transduction pathways, and developmental processes (Figure 10C). Statistical analysis shows that the likelihood of obtaining this orthologous network architecture due to randomness is P < 0.0001 (Supplementary Figure 5).
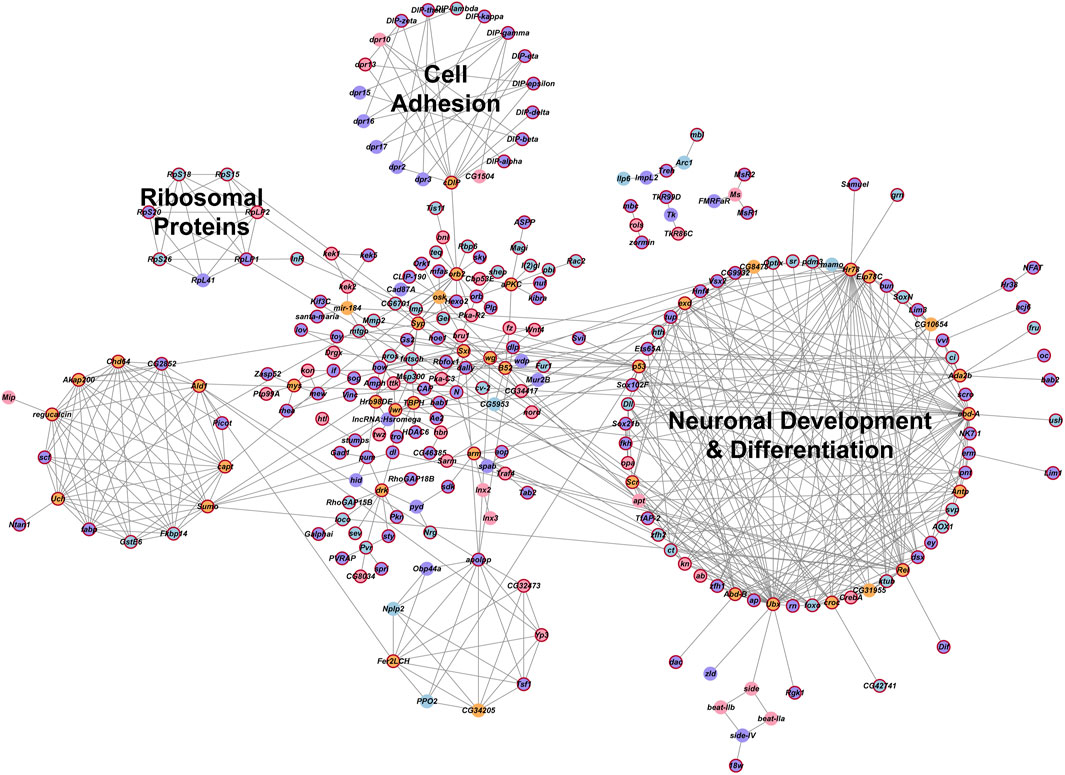
Figure 9. Global gene interaction network. The protein-protein interaction network was constructed within Cytoscape, based on interactions among genes that were differentially expressed (Benjamini–Hochberg adjusted p-value ≤0.05) across all clusters in the male and female datasets. Crimson red borders indicate genes with human orthologs. Circles representing genes are color-coded: purple indicates genes differentially expressed in both males and females, blue indicates genes differentially expressed only in males, pink indicates genes differentially expressed only in females and orange indicates inferred genes that were computationally recruited from the FlyBase Interaction Database. Annotations of these groups represent the processes that are enriched for the genes within these groups. A Benjamini and Hochberg (1995) adjusted p-value ≤0.05 was considered a significant enrichment in the statistical overrepresentation tests.
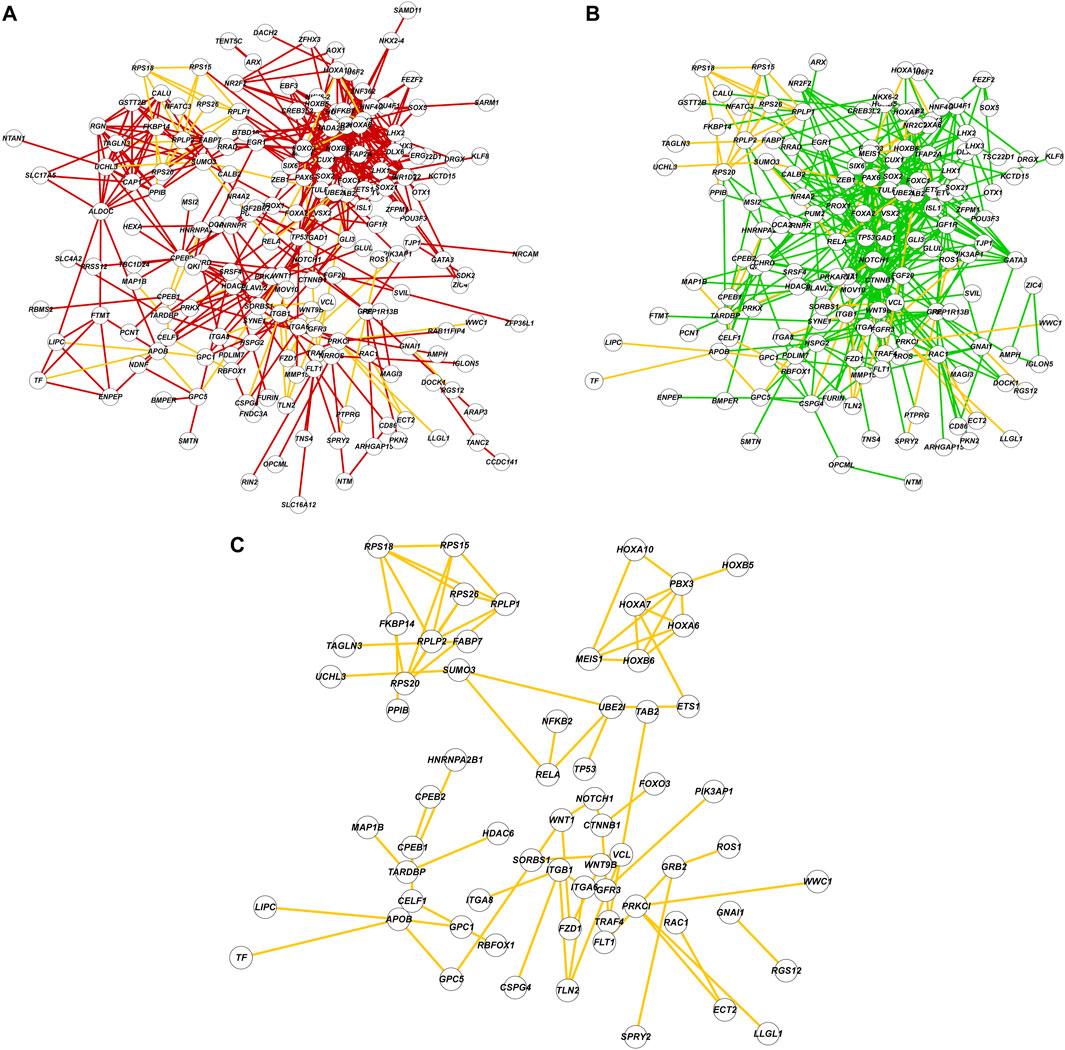
Figure 10. Comparisons of protein-protein interaction networks. (A) The protein-protein interaction network constructed with Drosophila genes and FlyBase physical interaction database. Nodes are labeled with human orthologs of Drosophila genes. (B) The protein-protein interaction network constructed with human orthologs and interactions from String-DB database. (C) A subnetwork composed of shared edges (yellow) between (A) and (B).
4 Discussion
Exposure to NaAsO2 at a concentration that does not result in immediate lethality affects Drosophila locomotor ability sooner in females than in males. The transcriptional response in the brain after 6 days of chronic exposure to NaAsO2 is widespread and different between males and females, yet gene ontology analysis suggests that these sexually dimorphic responses converge through different mechanisms on the same biological processes. The transcriptional responses to arsenic exposure are markedly sex antagonistic with responses in females biased toward neurons and in males toward glia. This may imply a neuroprotective function of glia in males since males are more resistant to chronic arsenic exposure than females (Figure 3B). The mechanisms that underlie sex antagonistic modulation of arsenic-induced gene expression are not known, but may arise from differences in chromatin accessibility, differences in chromosomal conformations due to the presence of different sex chromosomes affecting intrachromosomal interactions, as well as antagonistic regulation of the doublesex (dsx) gene, which shows NaAsO2-induced altered transcript abundance and is part of the sex determination pathway (Steinmann-Zwicky, 1994).
Chronic exposure to NaAsO2 leads to differential expression of more than 5% of the Drosophila genome, indicating a highly polygenic response that is likely to affect multiple organismal phenotypes. We used innate negative geotaxis as a proxy phenotype for NaAsO2 neurotoxicity as it can readily be quantified using a climbing assay. It is, however, likely that phototaxis is also affected by NaAsO2 exposure, based on the prominent changes in transcript abundances in cell clusters associated with vision. In addition, gene ontology enrichment analysis implicates possible additional physiological, developmental and behavioral phenotypes that may be affected by NaAsO2, including locomotion, mating and courtship, feeding, sleep and circadian rhythm as well as neurodevelopment and nervous system function, including sensory perception. Locomotion and circadian rhythm have previously been implicated in studies on arsenic toxicity in Drosophila (Ortiz et al., 2009).
Differential regulation of genes associated with detoxification processes and response to oxidative stress features prominently in the transcriptional response to chronic NaAsO2 exposure, most notably in males in glial cell populations, in concordance with their greater resilience to NaAsO2 than females as reflected in their climbing ability. In addition, many genes associated with locomotion were differentially expressed upon chronic exposure to NaAsO2, including the X-chromosome-linked genes spin, sr, tim, zfh1, dac, ey, rhea, robo2, and Sh, also in line with the adverse effect of NaAsO2 on climbing ability.
Over 60% of differentially expressed genes occur in multiple cell clusters in both males and females. In females, the top 10 genes (Orcokinin, Pdf, Ms, fabp, CG12239, Nplp1, lncRNA:CR40469, CG46385, CG8745, and ATP8B) were differentially expressed across multiple clusters. In males, srRNA, TotC, CG12239, Eip74 EF, Pzl, lncRNA:CR46004, apolpp, 5-HT2A, TotA, sr, Obp44a, Eaat1, puc, MtnA, and CG34393 showed widespread differential expression (Supplementary Table 10).
Orcokinin and Nplp1 are differentially expressed in females in 20 and 18 clusters, respectively, and have been implicated in sleep, locomotion, and feeding (Silva et al., 2021). In males, Eip74 EF, a target of the ecdysone signaling pathway, associated with metamorphosis, apoptosis, immune response, and stress (Thummel, 1996; Liu N. et al., 2012; Xiong et al., 2016; Chalmers et al., 2023) is differentially regulated in 13 clusters. The gene puc (puckered), a negative regulator of JNK signaling, that contributes to apoptosis, stress responses, and immune function (McEwen et al., 2005) is differentially expressed in 10 cell clusters in response to chronic NaAsO2 exposure. Additionally, transcript abundance of the 5-HT2A serotonin receptor, which modulates neural signaling related to feeding and locomotion (Munneke et al., 2022), is altered in 12 cell clusters. These examples underscore the widespread pleiotropic effects on diverse physiological and behavioral phenotypes that are likely to result from arsenic exposure.
Previous studies have shown extensive genetic variation in susceptibility to cadmium and lead (Zhou et al., 2017) in an advanced intercross population derived from the inbred lines of the Drosophila Genetic Reference Panel (DGRP) (Mackay et al., 2012; Huang et al., 2014). Variation in sexual dimorphism in susceptibility to 4-methylimidazole has also been documented in the DGRP (Collins et al., 2024). It is, however, likely that the widespread changes in gene regulation across multiple cell populations in the brain and the sexual dimorphism and sexual antagonism that are characteristic of the neurotranscriptional response to NaAsO2 will also be representative for transcriptional responses to other neurotoxins. Furthermore, we analyzed concordance between Drosophila and human protein-protein interaction networks and found statistically significant conservation of functional relationships indicating our ability to infer neurotoxic mechanisms in the human brain from the Drosophila model.
Data availability statement
Raw data for climbing and lifespan measurements are included in the supplementary tables. All single-nuclei RNA sequences data generated in this study have been submitted to the NCBI Gene Expression Omnibus (GEO; https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE290993) under accession numbers GSE290993, GSM8827136, GSM8827137, GSM8827138 and GSM8827139. R code that was used to perform Seurat-based and Network-based analyses is available at GitHub repositories https://github.com/Anuragbio/As-snRNAseq and https://github.com/vshanka23/mcpp_network_significance respectively.
Author contributions
AC: Writing – review and editing, Methodology, Investigation, Writing – original draft, Formal Analysis, Conceptualization, Visualization, Data curation. VS: Software, Investigation, Visualization, Writing – review and editing, Methodology, Formal Analysis. BS: Investigation, Methodology, Writing – review and editing. RL: Writing – review and editing, Methodology. PF: Methodology, Writing – review and editing. EH: Investigation, Writing – review and editing. KC: Investigation, Writing – review and editing. TM: Conceptualization, Project administration, Supervision, Writing – review and editing, Resources, Visualization, Funding acquisition. RA: Funding acquisition, Writing – original draft, Resources, Project administration, Supervision, Writing – review and editing, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This project has received funding from the European Union’s Horizon 2020 Research and Innovation program under Grant Agreement No 965406. The work presented in this publication was performed as part of ASPIS. The results and conclusions reflect only the authors’ views, and the European Commission cannot be held responsible for any use that may be made of the information contained therein. AC is supported by a fellowship from the Self Family Foundation. The Genomics and Bioinformatics Cores of the Center for Human Genetics are supported in part by grant 1P20GM139769 from the National Institute of General Medical Sciences of the National Institutes of Health to TFCM and RRHA.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ftox.2025.1636431/full#supplementary-material
References
Adebambo, T. H., Medina-Flores, F. M., Zhang, S. L., and Lerit, D. A. (2025). Arsenic impairs Drosophila neural stem cell mitotic progression and sleep behavior in a tauopathy model. G3 15, jkaf049. doi:10.1093/g3journal/jkaf049
Anushree, N., Ali, M. Z., Bilgrami, A. L., and Ahsan, J. (2023). Acute exposure to arsenic affects pupal development and neurological functions in Drosophila melanogaster. Toxics 11, 327. doi:10.3390/toxics11040327
Bader, G. D., and Hogue, C. W. (2003). An automated method for finding molecular complexes in large protein interaction networks. BMC Bioinforma. 4, 2–27. doi:10.1186/1471-2105-4-2
Baker, B. M., Mokashi, S. S., Shankar, V., Hatfield, J. S., Hannah, R. C., Mackay, T. F. C., et al. (2021). The Drosophila brain on cocaine at single-cell resolution. Genome Res. 31, 1927–1937. doi:10.1101/gr.268037.120
Bastian, F. B., Cammarata, A. B., Carsanaro, S., Detering, H., Huang, W. T., Joye, S., et al. (2025). Bgee in 2024: focus on curated single-cell RNA-seq datasets, and query tools. Nucleic Acids Res. 53, D878–D885. doi:10.1093/nar/gkae1118
Benjamini, Y., and Hochberg, Y. (1995). Controlling the false discovery rate - a practical and powerful approach to multiple testing. J. R. Stat. Soc. B 57, 289–300. doi:10.1111/j.2517-6161.1995.tb02031.x
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., and Lefebvre, E. (2008). Fast unfolding of communities in large networks. J. Stat. Mech. P10008 2008, P10008. doi:10.1088/1742-5468/2008/10/P10008
Burke, C. J., Huetteroth, W., Owald, D., Perisse, E., Krashes, M. J., Das, G., et al. (2012). Layered reward signalling through octopamine and dopamine in Drosophila. Nature 492, 433–437. doi:10.1038/nature11614
Butler, A., Hoffman, P., Smibert, P., Papalexi, E., and Satija, R. (2018). Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat. Biotechnol. 36, 411–420. doi:10.1038/nbt.4096
Calderon, M. R., Mori, M., Kauwe, G., Farnsworth, J., Ulian-Benitez, S., Maksoud, E., et al. (2022). Delta/Notch signaling in glia maintains motor nerve barrier function and synaptic transmission by controlling matrix metalloproteinase expression. Proc. Natl. Acad. Sci. U. S. A. 119, e2110097119. doi:10.1073/pnas.2110097119
Chalmers, M. R., Kim, J., and Kim, N. C. (2023). Eip74EF is a dominant modifier for ALS-FTD-linked VCP(R152H) phenotypes in the Drosophila eye model. BMC Res. Notes 16, 30. doi:10.1186/s13104-023-06297-z
Chen, Z., Zhang, W., Wang, F., Mu, R., and Wen, D. (2021). Sestrin protects Drosophila midgut from mercury chloride-induced damage by inhibiting oxidative stress and stimulating intestinal regeneration. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 248, 109083. doi:10.1016/j.cbpc.2021.109083
Collins, K. M., Howansky, E., Macon-Foley, S. C., Adonay, M. E., Shankar, V., Lyman, R. F., et al. (2024). Drosophila toxicogenomics: genetic variation and sexual dimorphism in susceptibility to 4-methylimidazole. Hum. Genomics 18, 119. doi:10.1186/s40246-024-00689-3
Contreras, E. G., Kautzmann, S., and Klämbt, C. (2024). The Drosophila blood-brain barrier invades the nervous system in a GPCR-dependent manner. Front. Cell. Neurosci. 18, 1397627. doi:10.3389/fncel.2024.1397627
Currier, T. A., Pang, M. M., and Clandinin, T. R. (2023). Visual processing in the fly, from photoreceptors to behavior. Genetics 224, iyad064. doi:10.1093/genetics/iyad064
Finak, G., McDavid, A., Yajima, M., Deng, J., Gersuk, V., Shalek, A. K., et al. (2015). MAST: a flexible statistical framework for assessing transcriptional changes and characterizing heterogeneity in single-cell RNA sequencing data. Genome Biol. 16, 278. doi:10.1186/s13059-015-0844-5
Freund, P., Al-Shafai, L., Mankovskii, G., Howarth, D., and Margolin, E. (2020). Clinicopathological correlates: chronic arsenic toxicity causing bilateral symmetric progressive optic neuropathy. J. Neuroophthalmol. 40, 423–427. doi:10.1097/WNO.0000000000000961
Ganie, S. Y., Javaid, D., Hajam, Y. A., and Reshi, M. S. (2023). Arsenic toxicity: sources, pathophysiology and mechanism. Toxicol. Res. 13, tfad111. doi:10.1093/toxres/tfad111
Goenawan, I. H., Bryan, K., and Lynn, D. J. (2016). DyNet: visualization and analysis of dynamic molecular interaction networks. Bioinformatics 32, 2713–2715. doi:10.1093/bioinformatics/btw187
Grentner, A., Ragueneau, E., Gong, C., Prinz, A., Gansberger, S., Oyarzun, I., et al. (2024). ReactomeGSA: new features to simplify public data reuse. Bioinformatics 40, btae338. doi:10.1093/bioinformatics/btae338
Hafemeister, C., and Satija, R. (2019). Normalization and variance stabilization of single-cell RNA-seq data using regularized negative binomial regression. Genome Biol. 20, 296. doi:10.1186/s13059-019-1874-1
Hindle, S. J., and Bainton, R. J. (2014). Barrier mechanisms in the Drosophila blood-brain barrier. Front. Neurosci. 8, 414. doi:10.3389/fnins.2014.00414
Holsopple, J. M., Smoot, S. R., Popodi, E. M., Colbourne, J. K., Shaw, J. R., Oliver, B., et al. (2023). Assessment of chemical toxicity in adult Drosophila melanogaster. J. Vis. Exp. 10, 3791–65029. doi:10.3791/65029
Hu, Y., Comjean, A., Attrill, H., Antonazzo, G., Thurmond, J., Chen, W., et al. (2023). PANGEA: a new gene set enrichment tool for Drosophila and common research organisms. Nucleic Acids Res. 51, W419–W426. doi:10.1093/nar/gkad331
Hu, Y., Flockhart, I., Vinayagam, A., Bergwitz, C., Berger, B., Perrimon, N., et al. (2011). An integrative approach to ortholog prediction for disease-focused and other functional studies. BMC Bioinforma. 12, 357–16. doi:10.1186/1471-2105-12-357
Huang, W., Massouras, A., Inoue, Y., Peiffer, J., Ràmia, M., Tarone, A. M., et al. (2014). Natural variation in genome architecture among 205 Drosophila melanogaster Genetic Reference Panel lines. Genome Res. 24, 1193–1208. doi:10.1101/gr.171546.113
Ijomone, O. M., Ijomone, O. K., Iroegbu, J. D., Ifenatuoha, C. W., Olung, N. F., and Aschner, M. (2020). Epigenetic influence of environmentally neurotoxic metals. Neurotoxicology 81, 51–65. doi:10.1016/j.neuro.2020.08.005
Jang, Y. H., Chae, H. S., and Kim, Y. J. (2017). Female-specific myoinhibitory peptide neurons regulate mating receptivity in Drosophila melanogaster. Nat. Commun. 8, 1630. doi:10.1038/s41467-017-01794-9
Jomova, K., Jenisova, Z., Feszterova, M., Baros, S., Liska, J., Hudecova, D., et al. (2011). Arsenic: toxicity, oxidative stress and human disease. J. Appl. Toxicol. 31, 95–107. doi:10.1002/jat.1649
Kanehisa, M., Furumichi, M., Sato, Y., Matsuura, Y., and Ishiguro-Watanabe, M. (2025). KEGG: biological systems database as a model of the real world. Nucleic Acids Res. 53, D672–D677. doi:10.1093/nar/gkae909
Kapaj, S., Peterson, H., Liber, K., and Bhattacharya, P. (2006). Human health effects from chronic arsenic poisoning--a review. J. Environ. Sci. Health A Tox. Hazard Subst. Environ. Eng. 41, 2399–2428. doi:10.1080/10934520600873571
Karachaliou, C., Sgourou, A., Kakkos, S., and Kalavrouziotis, I. (2021). Arsenic exposure promotes the emergence of cardiovascular diseases. Rev. Environ. Health 37, 467–486. doi:10.1515/reveh-2021-0004
Kirby, K., Hu, J., Hilliker, A. J., and Phillips, J. P. (2002). RNA interference-mediated silencing of Sod2 in Drosophila leads to early adult-onset mortality and elevated endogenous oxidative stress. Proc. Natl. Acad. Sci. U. S. A. 99, 16162–16167. doi:10.1073/pnas.252342899
Kuo, C.-C., Balakrishnan, P., Gribble, M. O., Best, L. G., Goessler, W. J. G., Umans, J. G., et al. (2021). The association of arsenic exposure and arsenic metabolism with all-cause, cardiovascular and cancer mortality in the Strong Heart Study. Environ. Int. 159, 107029. doi:10.1016/j.envint.2021.107029
Lee, C. H., and Yu, H. S. (2016). Role of mitochondria, ROS, and DNA damage in arsenic induced carcinogenesis. Front. Biosci. 8, 312–320. doi:10.2741/s465
Liu, C., Plaçais, P. Y., Yamagata, N., Pfeiffer, B. D., Aso, Y., Friedrich, A. B., et al. (2012). A subset of dopamine neurons signals reward for odour memory in Drosophila. Nature 488, 512–516. doi:10.1038/nature11304
Liu, N., Landreh, M., Cao, K., Abe, M., Hendriks, G. J., Kennerdell, J. R., et al. (2012). The microRNA miR-34 modulates ageing and neurodegeneration in Drosophila. Nature 482, 519–523. doi:10.1038/nature10810
Mackay, T. F. C., Richards, S., Stone, E. A., Barbadilla, A., Ayroles, J. F., Zhu, D., et al. (2012). The Drosophila melanogaster genetic reference Panel. Nature 482, 173–178. doi:10.1038/nature10811
Mahapatra, C. T., and Rand, M. D. (2012). Methylmercury tolerance is associated with the humoral stress factor gene Turandot A. Neurotoxicol. Teratol. 34, 387–394. doi:10.1016/j.ntt.2012.04.007
Martinez, V. D., and Lam, W. L. (2021). Health effects associated with pre- and perinatal exposure to arsenic. Front. Genet. 12, 664717. doi:10.3389/fgene.2021.664717
McEwen, D. G., Peifer, M., and Puckered, M. (2005). A Drosophila MAPK phosphatase, ensures cell viability by antagonizing JNK-induced apoptosis. Development 132, 3935–3946. doi:10.1242/dev.01949
Mi, H., and Thomas, P. (2009). PANTHER pathway: An ontology-based pathway database coupled with data analysis tools. Methods Mol. Biol. 563, 123–140. doi:10.1007/978-1-60761-175-2_7
Mohr, S. E., Rudd, K., Hu, Y., Song, W. R., Gilly, Q., Buckner, M., et al. (2018). Zinc detoxification: A functional genomics and transcriptomics analysis in Drosophila melanogaster cultured cells. G3 (Bethesda) 8, 631–641. doi:10.1534/g3.117.300447
Mokashi, S. S., Shankar, V., MacPherson, R. A., Hannah, R. C., Mackay, T. F. C., and Anholt, R. R. H. (2021). Developmental alcohol exposure in Drosophila: Effects on adult phenotypes and gene expression in the brain. Front. Psychiatry 12, 699033. doi:10.3389/fpsyt.2021.699033
Muñiz Ortiz, J. G., Shang, J., Catron, B., Landero, J., Caruso, J. A., and Cartwright, I. L. (2011). A transgenic Drosophila model for arsenic methylation suggests a metabolic rationale for differential dose-dependent toxicity endpoints. Toxicol. Sci. 121, 303–311. doi:10.1093/toxsci/kfr074
Munneke, A. S., Chakraborty, T. S., Porter, S. S., Gendron, C. M., and Pletcher, S. D. (2022). The serotonin receptor 5-HT2A modulates lifespan and protein feeding in Drosophila melanogaster. Front. Aging 3, 1068455. doi:10.3389/fragi.2022.1068455
Musselman, L. P., Fink, J. L., Ramachandran, P. V., Patterson, B. W., Okunade, A. L., Maier, E., et al. (2013). Role of fat body lipogenesis in protection against the effects of caloric overload in Drosophila. J. Biol. Chem. 288, 8028–8042. doi:10.1074/jbc.M112.371047
Naujokas, M. F., Anderson, B., Ahsan, H., Aposhian, H. V., Graziano, J. H., Thompson, C., et al. (2013). The broad scope of health effects from chronic arsenic exposure: update on a worldwide public health problem. Environ. Health Perspect. 121, 295–302. doi:10.1289/ehp.1205875
Nie, X., Wang, Y., Zhao, H., Guo, M., Liu, Y., and Xing, M. (2020). As(3+) or/and Cu(2+) exposure triggers oxidative stress imbalance, induces inflammatory response and apoptosis in chicken brain. Ecotoxicol. Environ. Saf. 203, 110993. doi:10.1016/j.ecoenv.2020.110993
Nohara, K., Suzuki, T., and Okamura, K. (2020). Gestational arsenic exposure and paternal intergenerational epigenetic inheritance. Toxicol. Appl. Pharmacol. 409, 115319. doi:10.1016/j.taap.2020.115319
Orr, W. C., and Sohal, R. S. (1994). Extension of life-span by overexpression of superoxide dismutase and catalase in Drosophila melanogaster. Science 263, 1128–1130. doi:10.1126/science.8108730
Ortiz, J. G. M., Opoka, R., Kane, D., and Cartwright, I. L. (2009). Investigating arsenic susceptibility from a genetic perspective in Drosophila reveals a key role for glutathione synthetase. Toxicol. Sci. 107, 416–426. doi:10.1093/toxsci/kfn192
Podgorski, J., and Berg, M. (2020). Global threat of arsenic in groundwater. Science 368, 845–850. doi:10.1126/science.aba1510
Prakash, C., Soni, M., and Kumar, V. (2016). Mitochondrial oxidative stress and dysfunction in arsenic neurotoxicity: A review. J. Appl. Toxicol. 36, 179–188. doi:10.1002/jat.3256
Rahman, M. A., Hannan, M. A., Uddin, M. J., Rahman, M. S., Rashid, M. M., and Kim, B. (2021). Exposure to environmental arsenic and emerging risk of Alzheimer's disease: perspective mechanisms, management strategy, and future directions. Toxics 9, 188. doi:10.3390/toxics9080188
Rand, M. D. (2010). Drosophotoxicology: the growing potential for Drosophila in neurotoxicology. Neurotoxicol. Teratol. 32, 74–83. doi:10.1016/j.ntt.2009.06.004
Rand, M. D., Tennessen, J. M., Mackay, T. F. C., and Anholt, R. R. H. (2023). Perspectives on the Drosophila melanogaster model for advances in toxicological science. Curr. Protoc. 3, e870. doi:10.1002/cpz1.870
Rojas, D., Rager, J. E., Smeester, L., Bailey, K. A., Drobná, Z., Rubio-Andrade, M., et al. (2015). Prenatal arsenic exposure and the epigenome: identifying sites of 5-methylcytosine alterations that predict functional changes in gene expression in newborn cord blood and subsequent birth outcomes. Toxicol. Sci. 143, 97–106. doi:10.1093/toxsci/kfu210
Ruan, H., Tang, X. D., Chen, M. L., Joiner, M. L., Sun, G., Brot, N., et al. (2002). High-quality life extension by the enzyme peptide methionine sulfoxide reductase. Proc. Natl. Acad. Sci. U. S. A. 99, 2748–2753. doi:10.1073/pnas.032671199
Schoofs, L., De Loof, A., and Van Hiel, M. B. (2017). Neuropeptides as regulators of behavior in insects. Annu. Rev. Entomol. 62, 35–52. doi:10.1146/annurev-ento-031616-035500
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi:10.1101/gr.1239303
Silva, V., Palacios-Muñoz, A., Volonté, M., Frenkel, L., Ewer, J., and Ons, S. (2021). Orcokinin neuropeptides regulate reproduction in the fruit fly, Drosophila melanogaster. Insect Biochem. Mol. Biol. 139, 103676. doi:10.1016/j.ibmb.2021.103676
Steinmann-Zwicky, M. (1994). Sex determination of the Drosophila germ line: tra and dsx control somatic inductive signals. Development 120, 707–716. doi:10.1242/dev.120.3.707
Thakur, M., Rachamalla, M., Niyogi, S., Datusalia, A. K., and Flora, S. J. S. (2021). Molecular mechanism of arsenic-induced neurotoxicity including neuronal dysfunctions. Int. J. Mol. Sci. 22, 10077. doi:10.3390/ijms221810077
Thummel, C. S. (1996). Flies on steroids-Drosophila metamorphosis and the mechanisms of steroid hormone action. Trends Genet. 12, 306–310. doi:10.1016/0168-9525(96)10032-9
Tolins, M., Ruchirawat, M., and Landrigan, P. (2014). The developmental neurotoxicity of arsenic: cognitive and behavioral consequences of early life exposure. Ann. Glob. Health 80, 303–314. doi:10.1016/j.aogh.2014.09.005
Tyler, C. R., and Allan, A. M. (2014). The effects of arsenic exposure on neurological and cognitive dysfunction in human and rodent studies: A Review. Curr. Environ. Health Rep. 1, 132–147. doi:10.1007/s40572-014-0012-1
Xiong, X. P., Kurthkoti, K., Chang, K. Y., Li, J. L., Ren, X., Ni, J. Q., et al. (2016). miR-34 Modulates innate immunity and ecdysone signaling in Drosophila. PLoS Pathog. 12, e1006034. doi:10.1371/journal.ppat.1006034
Yamagata, N., Hiroi, M., Kondo, S., Abe, A., and Tanimoto, H. (2016). Suppression of dopamine neurons mediates reward. PLoS Biol. 14, e1002586. doi:10.1371/journal.pbio.1002586
Yu, G., Wu, L., Su, Q., Ji, X., Zhou, J., Wu, S., et al. (2024). Neurotoxic effects of heavy metal pollutants in the environment: Focusing on epigenetic mechanisms. Environ. Pollut.(Barking, Essex 1987) 345, 123563. doi:10.1016/j.envpol.2024.123563
Zarazúa, S., Ríos, R., Delgado, J. M., Santoyo, M. E., Ortiz-Pérez, D., and Jiménez-Capdeville, M. E. (2010). Decreased arginine methylation and myelin alterations in arsenic exposed rats. Neurotoxicology 31, 94–100. doi:10.1016/j.neuro.2009.10.014
Keywords: neurotoxicity, single cell RNA sequencing, behavioral genetics, sexual dimorphism, genetic networks
Citation: Chaturvedi A, Shankar V, Simkhada B, Lyman RA, Freymuth P, Howansky E, Collins KM, Mackay TFC and Anholt RRH (2025) Arsenic toxicity in the Drosophila brain at single cell resolution. Front. Toxicol. 7:1636431. doi: 10.3389/ftox.2025.1636431
Received: 27 May 2025; Accepted: 25 June 2025;
Published: 10 July 2025.
Edited by:
Konrad Andrzej Szychowski, University of Information Technology and Management in Rzeszow, PolandReviewed by:
Sudipta Dutta, Texas A and M University, United StatesChayan Munshi, Ethophilia Research Foundation, India
Copyright © 2025 Chaturvedi, Shankar, Simkhada, Lyman, Freymuth, Howansky, Collins, Mackay and Anholt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Trudy F. C. Mackay, dG1hY2theUBjbGVtc29uLmVkdQ==; Robert R. H. Anholt, cmFuaG9sdEBjbGVtc29uLmVkdQ==