 Xiaomei Yang1
Xiaomei Yang1 Qi Feng1*
Qi Feng1* Wei Liu1Honghua Xia1,2
Wei Liu1Honghua Xia1,2 Jutao Zhang1
Jutao Zhang1 Linshan Yang1Chengqi Zhang1Zhiyang Wang3Yonglin Feng3
Linshan Yang1Chengqi Zhang1Zhiyang Wang3Yonglin Feng3- 1Key Laboratory of Ecohydrology of Inland River Basin, Alax Desert Eco-hydrology Experimental Research Station, Qilian Mountains Eco-Environment Research Center in Gansu Province, Northwest Institute of Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou, China
- 2Northwest Institute of Eco-Environment and Resources, University of Chinese Academy of Sciences, Beijing, China
- 3Technology Innovation Center for Mine Geological Environment Restoration in the Alpine and Arid Regions, Lanzhou, China
Background: Mining activities are known to exert significant effects on the structure and function of grassland ecosystems. However, the role of mining grasslands restoration in altering the plant community and soil quality remains poorly understood, especially in alpine regions. Here, we investigated species diversity in grasslands with dynamic changes and different restoration levels in the Tianzhu alpine mining area locating in the Qilian Mountains.
Methods: The plant community structure and species composition of the grasslands with different restoration levels were analyzed by the sample method. We used five different restoration levels: very low recovered degree (VLRD), low recovered degree (LRD), medium recovered degree (MRD), and high recovered degree (HRD), and selected natural grassland (NGL, CK) as the control.
Results: Plant community structure and species composition were significantly higher than those under the VLRD in the Tianzhu alpine mining area (p < 0.05), with HRD > MRD > LRD > VLRD. There were 11 families, 18 genera, and 17 species of plants, mainly in the families of Leguminosae, Asteraceae, Gramineae, Rosaceae, and Salicaceae; among them, Salicaceae and Gramineae played a decisive role in the stability of the community. The ecotype community showed that perennial herbaceous plants were the most dominant, with annual herbaceous plants being the least dominant, and no tree and shrub layers were observed; the dominance index was the highest in VLRD at 0.32, the richness index was the highest in HRD at 2.73, the diversity of HRD was higher at 1.93, soil pH and EC showed a decreasing trend, and SMC, SOC, TN, NO3-N, NH4-N, AN, TP, and AP content showed an increasing trend with the increase of grassland restoration.
Conclusion: In summary, with the increase of restored grassland in the Tianzhu alpine mining area, plant diversity gradually increased and plant community structure gradually diversified, which was close to the plant diversity of NGL. The protection of partially VLRD and LRD grasslands in the mining area should be emphasized, and the mine grassland should be used rationally and scientifically restored.
1 Introduction
Community structure and biodiversity loss have become a global concern; further, there is accumulating evidence suggesting that these losses will negatively impact ecosystem services, which are essential for societies (Wang et al., 2014). Although community structure and species composition determine the presence of biodiversity, the relationship between species diversity and grassland degradation are the core issues in ecological research (Che et al., 2019; Ma et al., 2022). It has been well acknowledged that decreasing soil biodiversity can hinder many ecosystem properties, such as aboveground plant diversity, nutrient retention, and nutrient cycling (Cappelli et al., 2022; Choudoir et al., 2022). When the composition of the plant community changes with the grassland, it can be intuitively predicted through the changes in plants. Therefore, clarifying and quantifying the change law of plant community structure and species diversity with grassland restoration can aid in the accurate prediction of the community structure and biodiversity of varying degrees of restoration on the spatiotemporal scales. However, humans have taken a series of measures to restore the service functions of destroyed ecosystems post unreasonable mining, of which, revegetation is one of the most important approaches (Neldner et al., 2017; Ahirwal et al., 2018; Liu et al., 2019; Gao et al., 2022). In recent years, the study on plant functional species diversity by domestic and foreign scholars have primarily focused on tropical and temperate forest ecosystems (Satake et al., 2021; Thakur et al., 2021); however, there are few reports on the restoration of grassland ecosystems in mining areas, especially in the alpine mining areas of the Qilian mountains.
The utilization of mining is an integral part of socio-economic development. However, over-exploitation in mining areas has aggravated the degradation of the grassland ecological environment (Akala et al., 2015; Wang et al., 2018), resulting in landscape damage to the ecological and geological environment, soil pollution, and vegetation resource pollution; it has largely destroyed the ecological environment of the Qilian mountains national nature reserve (Haddaway et al., 2019; Adator et al., 2021), and its ecological restoration has become a global ecosystem problem. According to the current status of mining area restoration, domestic and foreign experts have actively carried out ecological restoration and governance in alpine mining areas, clarified the current situation and driving forces of mining area restoration, and reported that the impact of mining area restoration on plant community structure, soil nutrient content comparison, underground microbial communities, and diversity were different (Rania et al., 2021). However, the Qilian Mountains alpine mining area has not been used as a study area to measure the degree of grassland restoration and change in biological communities in the mining area (Figueiredo et al., 2023).
Grassland is the largest area and most important land use type in the Qilian mountains in northwest China (Kang et al., 2021); it plays an irreplaceable role in the construction and development of the ecological environment in the western region (Feng, 2019). Due to mining in the Qilian Mountain reserve, the ecological environment has been damaged, such as the destruction of the original grassland and soil erosion. At present, many researchers have managed the ecological environment of the mining area, scientifically and rationally restored the degraded grassland in the mining area, and promoted the restoration of the mining area. Ecological balance, moderate productivity restoration, and economic benefits are crucial (Wang et al., 2021). The Tianzhu alpine grassland belongs to the northeastern margin of the Qilian mountains and plays a pivotal role in the stability of the ecosystem. Mainly dominated by grasses and sedges, it is one of the areas with the alpine ecosystem’s richest and densest species diversity. However, in recent years, grassland plant community diversity has decreased sharply due to unreasonable mining, the number of endangered plants continues to increase, ecosystem stability has been weakened, and grassland productivity has declined (Celestino et al., 2020; Zhu et al., 2022). Many researchers have restored grassland in mining areas in response to this problem. However, the changes in plant community structure, diversity, and soil microorganisms during restoration are yet to be discovered.
In this study, we considered the alpine grassland plant community in a typical Qianmalong mining area on the northeastern margin of the Qilian Mountains as the research sample plots. We specifically addressed the following scientific questions: 1) How the degree is the grassland ecosystem restoration in the alpine mining area of the Qilian Mountains? 2) What is the plant community structure after the grassland ecosystem restoration in the alpine mining area of the Qilian Mountains? 3) What is the dynamic change law of plant community structure, species diversity, and soil properties after the grassland ecosystem restoration in the alpine mining area of the Qilian Mountains with different degrees of restoration? This study aims to help advance the restoration of alpine mining areas and may provide theoretical support for the ecosystem restoration process of alpine mining grassland.
2 Materials and methods
2.1 Study site, experimental design, and soil sampling
The study site (36。91′N, 102。66′E; elevation: 2,821 m) is located in Tianzhu Tibetan Autonomous County, the Qilian mountain Qianmalong Coal mining grassland of the Gansu Province, northwestern China (Figure 1). It is characterized by a temperate continental arid climate with mean annual temperature, precipitation, and sunshine hours of ∼10°C, 416 mm—most of it is orographic rain concentrated from July–September—, and 2926.7 h, respectively. The primary soil type in the study site is a subalpine meadow with organic matter content, total nitrogen and phosphorus, and pH of ∼10.92, 0.64, and 0.65 g kg–1, and 8.00, respectively. The vegetation type in this area is mainly alpine grassland. The vegetation types in the research alpine mining area are rich and diverse, and the main types of grassland plants are Polygonum viviparum, Elymus nutans, Stipa krylovii, and Cyperus rotundus L.
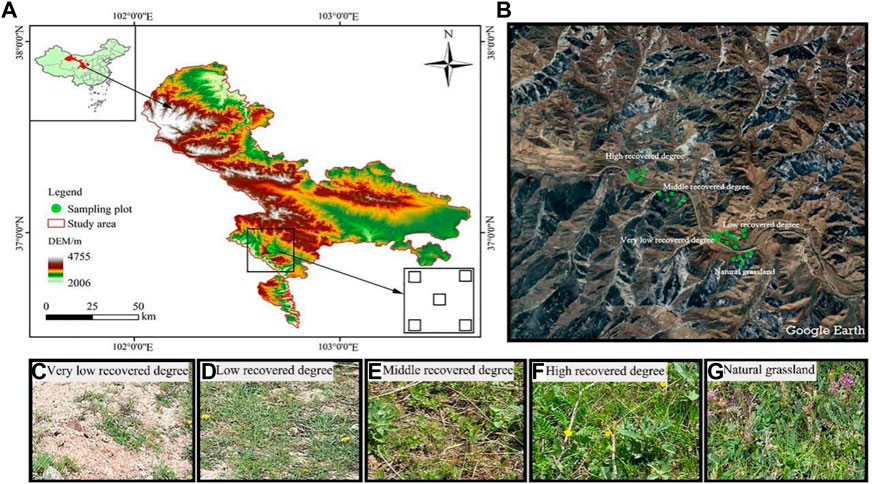
FIGURE 1. Location of the study area in the Chinese Qilian Mountains (A, B) and VLRD, very low recovered degree (C), and LRD, low recovered degree (D) an MRD, middle recovered degree (E) and HRD, high recovered degree (F) and NGL, CK natural grassland (G).
Field investigation was conducted in August 2022; three large quadrats were set up in five different restoration grassland and three small sampling plots (0.5 m × 0.5 m) were set up in each large quadrat (Figures 1A,B). Field investigation and data calculation were based on the method of grassland ecological survey of Prof. Ren (1998), recording the plant species, species number, number of individuals, total community coverage, and species coverage in the quadrats. The circle was measured, the height was measured with a steel tape measure, and the altitude, longitude, and latitude of different locations were recorded using a GPS. The Grassland restoration time started in 2017 in the Qianmalong mine. Based on previous research on the Tianzhu section of Zhuaxi xiulong Township in Tianzhu Tibetan Autonomous County, comprehensive analysis of different grassland vegetation and soil characteristics, and the method of restoring succession sequences based on space restoration instead of time (Ren, 1998) and “Soil erosion classification and grading standard” (SL190-2007) of China, the representative restoration of grassland in Qilian Mountains high cold Tianzhu Qianmalong coal mine was selected for investigation, with five different degrees of grassland recovery: very low recovered degree (VLRD), low recovered degree (LRD), middle recovered degree (MRD), high recovered degree (HRD), and natural grassland (NGL), and natural grassland (NGL) as CK (Table 1).
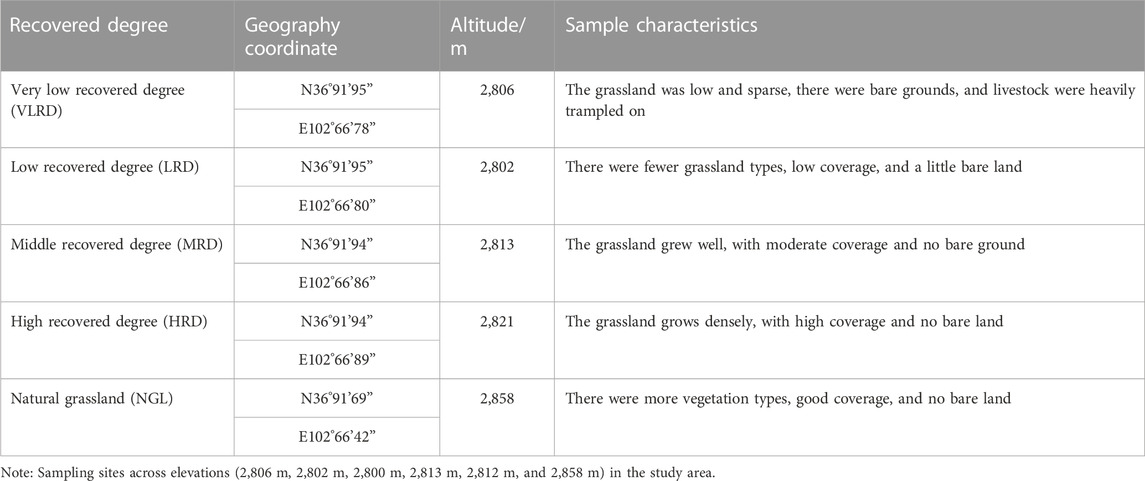
TABLE 1. Sample plots settings.
Soil samples were collected at each sampling plot, from 0 to 20 cm and 20 to 40 cm using a soil auger (Figure 1B). The samples used for soil SMC analysis were passed through a 2-mm sieve determined immediately and then stored at −80°C. Other samples used for analyzing physiochemical soil properties and soil enzyme activity were air-dried and passed through a 2-mm sieve.
2.2 Determination of soil properties
Gravimetric soil moisture content (SMC) was determined by drying to a constant weight at 105°C in an electrothermal blowing oven (DHG - 9,240, Shanghai Jing Macro, Shanghai, China). Soil pH was determined in a soil: water mixture at a ratio of 1:5, and soil organic carbon (SOC) content was determined using the dichromate oxidation method described by Walkley–Black (1934). Total nitrogen (TN) content was measured with an element analyzer (Vario MACRO cube, Elementar INC., Germany), while the total phosphorus (TP) and available phosphorus (AP) were determined by the Sommers-Nelson method. In addition, ammonium nitrogen content (NH4-N), nitrate nitrogen (NO3-N), and alkaline hydrolysis nitrogen (AN) were determined by the Flow-Injection method, Interval-flow analysis method, and Alkali diffusion method (Sommers et al., 1972), respectively.
2.3 Test methods
The community importance value is an important criterion for distinguishing different communities. The main components of each community were determined efficiently.
Important value calculation method: important value (IV) = (relative height + relative cover + relative density)/3 (Zhang, 2004).
Species diversity adopts species richness, diversity index, and uniformity. The indicators such as degree and dominance degree are expressed (Ma, 1994), calculated as follows:
where, H is the Shannon–Wiener species diversity index, Ma is the Margalef richness index, D is the Simpson diversity index, Eve is the Evenness index, Do is the Dominance index, Pi is the important value ratio of species i, N is the sum of the important values of the plants in the transect, Ni is the important value of the ith plant in the plot, and S is the total number of species in the plot.
2.4 Statistical analysis
One-way analysis of variance (ANOVA) was conducted in SPSS 21.0 (IBM Corp., Armonk, NY, United States) to detect vegetation characteristics of different restoration degrees of alpine grassland, the alpine meadow plant diversity variation. The statistical significance of the means was compared by Duncan’s multiple range test using Statistical Product. The important value and diversity index of each plant in the herb layer were calculated by Excel 2010. The bar chart was analyzed and plotted in Sigmaplot 10.0 (Sigmaplot Lab Corporation, Northampton, MA, USA). The correlation was analyzed and plotted in Origin 9.0 (Origin Lab Corporation, Northampton, MA, USA).
3 Results
3.1 Vegetation characteristics of alpine grassland community with different restoration degrees
Differences in vegetation characteristics in different restoration degrees of alpine grassland communities are shown in Figure 2. The grassland communities showed a downward trend and significant differences (p < 0.05) in different recovery levels of alpine grassland, which exhibited the order of HRD > NGL > MRD > LRD > VLRD. The height of the HRD was 2.20, 3.52, and 7.06 times that of the MRD, LRD, and VLRD, while its density (Figure 2A) was 3.88, 5.44, and 8.26 times that of the MRD, LRD, and VLRD, respectively (Figure 2B). The frequency of the HRD was 1.20, 1.73, and 2.49 times that of the MRD, LRD, and VLRD (Figure 2C), while the coverage was 1.44, 2.00, and 3.43 times that of MRD, LRD, and VLRD (Figure 2D), respectively. The aboveground biomass was 1.72, 3.26, and 4.29 times that of MRD, LRD, and VLRD (Figure 2E), while the frequency and coverage of the HRD were close to that of NGL (CK).
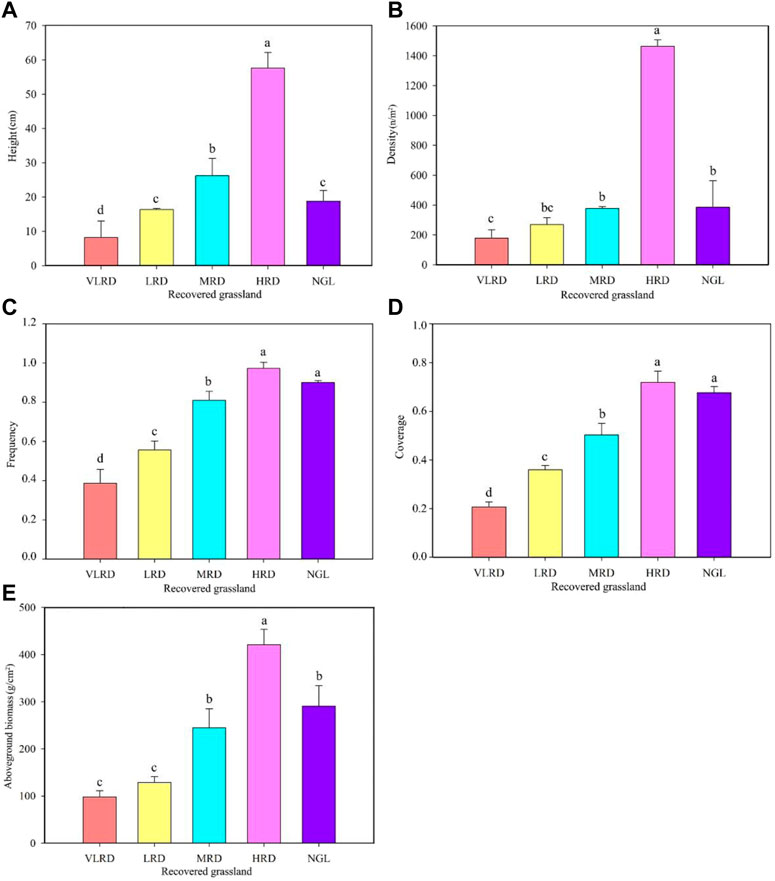
FIGURE 2. Vegetation Characteristics of alpine grassland under different restoration degrees. (A), Height; (B), Density; (C), Frequency; (D), Coverage; (E), Aboveground biomass. VLRD, very low recovered degree; LRD, low recovered degree; MRD, middle recovered degree; HRD, high recovered degree; NGL, CK natural grassland. Different letters above error bars between treatments indicate significant differences at the 0.05 level.
3.2 Plant family, genus, and species structure of alpine grassland community with different restoration degrees
Sequences in plant family, genus, and species structure in different restoration degrees of alpine grassland are shown in Table 2. It showed that the order of species composition with different degradation degrees of alpine grassland communities was HRD > MRD > LRD > VLRD. The sum of the individual numbers of different communities was consistent with the composition of species. According to the statistical sample survey data, there were 17 species of herbaceous plant in our study plot, which belonged to 11 families and 18 genera; of them, Asteraceae had the most species (5 genera and 5 species), followed by 2 genera and 2 species of Leguminae and Sedges, and 1 genus and 1 species of Poaceae, Yak seedlings, Ranunculaceae, Lessons, Rosaceae, Plantainae, Apiaceae, and Chenopodaceae. The community structure was simple, and the family composition was relatively scattered. In all samples, there were five species of Carex spp, Chenopodium glaucum, Cyperus rotundus L., Oxytropis ochrocephala, and Potentilla chinensis, and four plant species with different degrees of recovery belonged to the Legume, Asteraceae, Grasses, Rosaceae, and Sedges families were high; of them, five major families accounted for 90% of the total plant species of this community in the HRD, 80% of the MRD, 73% of the LRD, and 64% of the VLRD, mainly because Legumes, Poaceae, and other plants were adaptable to low temperature and arid environment; further, compared with other plants, these five major families of plants were adaptable to growth in alpine grassland mining areas. Among them, the number of species composition of grassland communities with high coverage was 3.50 times that of natural grassland in mining areas.
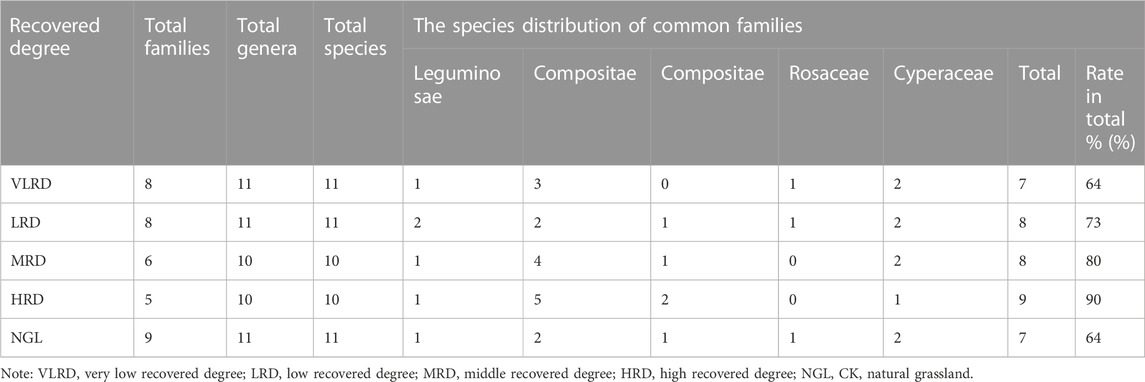
TABLE 2. Composition of main plant families, genera and species at different degrees of recovery.
3.3 Life-form structure characteristics of plant communities of alpine grassland communities with different restoration degrees
Characteristics in the life-form structure of plant communities in different restoration degrees of alpine grassland are shown in Table 3. According to the Whittaker growth type system, the life form is distinguished by the degree of lignification of plant stems in the community (Yang et al., 2016). The herbaceous plants in the study community were divided into perennial and annual herbs. In terms of life forms, perennial herbaceous species were the most dominant, with 15 species belonging to 10 families and 15 genera, and annual herbaceous species were the least dominant, with 2 species belonging to 2 families and 2 genera, accounting for 88.20% and 11.80% of the total species, respectively (Table 3). The number of perennial and annual herbaceous species was large in the grassland communities of alpine mining areas with different degrees of recovery. The number of perennial herbaceous species were relatively abundant and numerous.
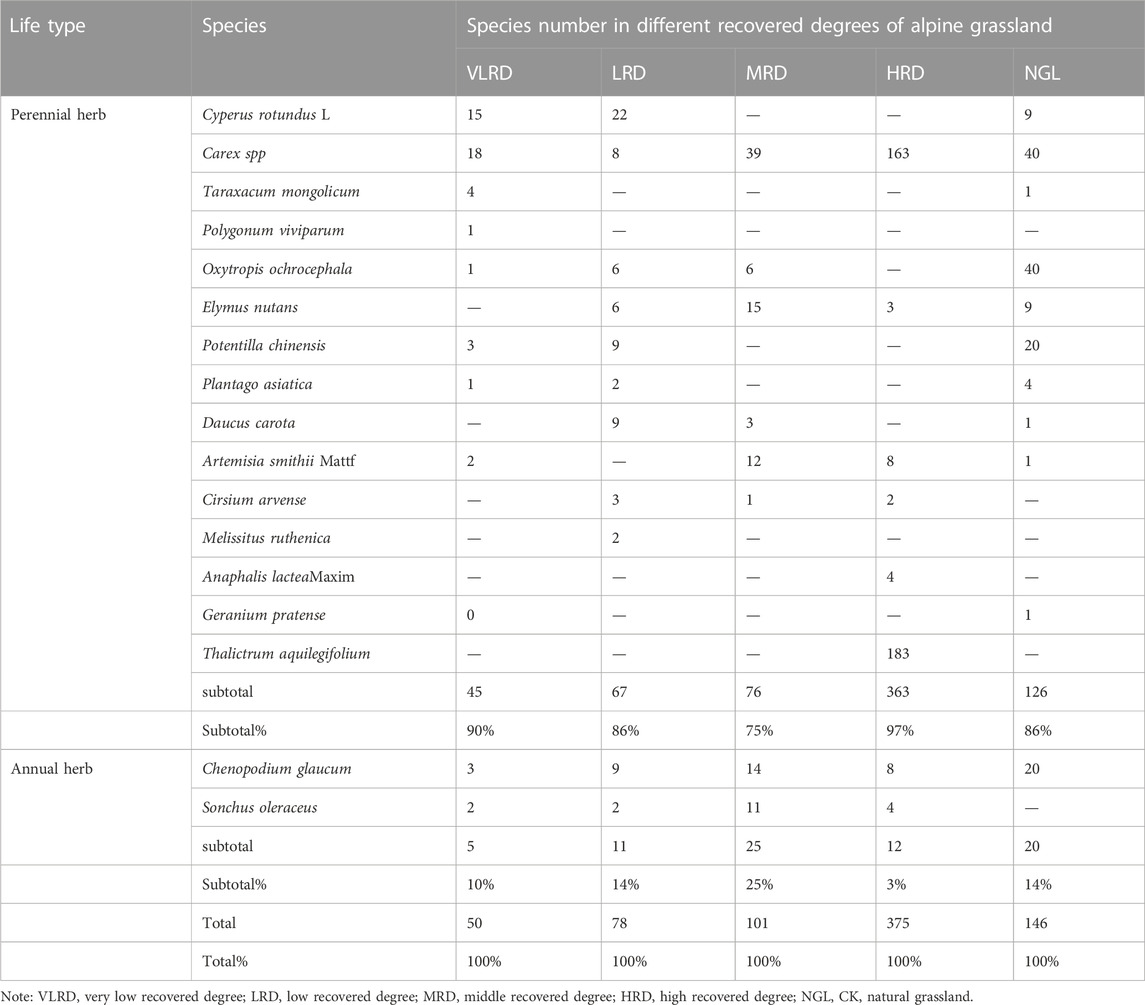
TABLE 3. Composition and quantitative characteristics of community in different recovered degrees.
In contrast, the annual herbs were relatively single and small in number; among them, perennial herbs accounted for the largest proportion of HRD, reaching 97%. On the contrary, the proportion of perennial herbs in MRD, LRD, VLRD, and NGL was large, reaching 75%, 86%, 90%, and 86%. respectively. The proportion of annual herbs with medium coverage was the largest, reaching 25%. The community structure of HRD grassland was slightly higher than that of NGL. The perennial herbaceous layer determined the community structure, system function, and stability. Hence, the community structure of HRD grassland was the most stable, followed by MRD, LRD, and VLRD grasslands.
3.4 Community value of alpine grassland community with different restoration degrees
The values of plant communities in different restoration degrees of alpine grassland are shown in Table 4. Perennial herbaceous plants have dominant positions in different communities and play an important role in community structure, ecosystem function, and stability. The dominant species of HRD and MRD of Carex spp were 29.43 and 29.11, respectively, and the sub-dominant species were Cyperus rotundus L. and Elymus nutans; the important values of HRD grassland were 25.86 and 10.67, while the important values of MRD were 19.70 and 13.38, respectively. The main associated species of HRD grassland were Chenopodium glaucum and Sonchus oleraceus, with important values of 7.46 and 6.14; the dominant species of Carex spp and Cyperus rotundus L. were the dominant species in the VLRD grassland, with important values of 39.96 and 30.86. The important values of other associated species were not much different, below 10. Comparative analysis of the herbaceous communities of alpine grassland in the four recovery degree mining areas showed that the critical value of perennial herbs in the community was ∼70. Hence, it occupied an important position in the recovery community of the alpine grassland mining area. However, the dominant position of species in each community was unclear, and the recovery grassland with an HRD was the best, close to NGL.
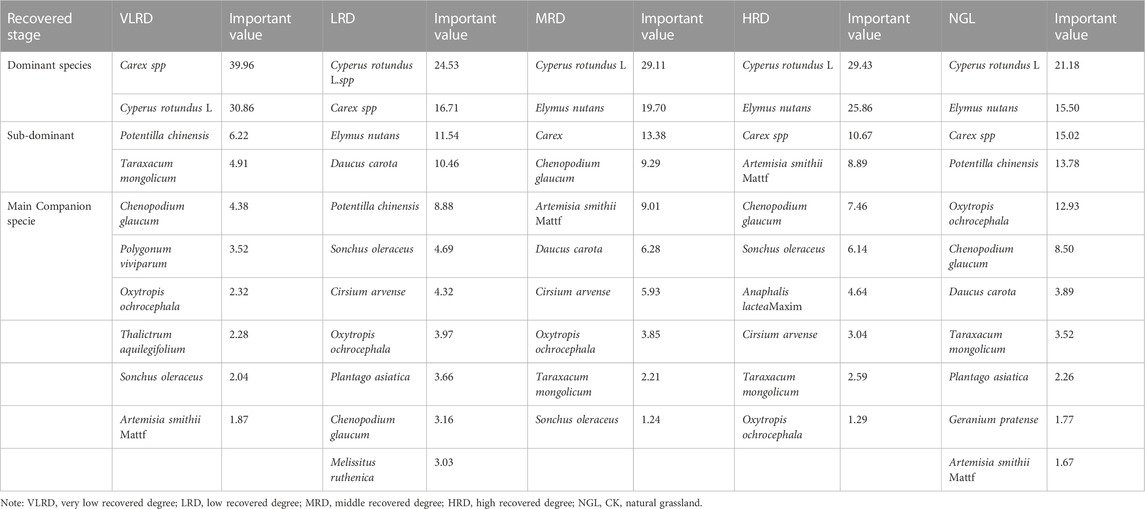
TABLE 4. The important value of dominant and sub-dominant species under different recovered stages.
3.5 Characteristics of community life structure with different restoration degrees
Compared with the two life forms, the trend of all types of restored grassland was the same: perennial herbaceous plants were markedly dominant, and the sum of important value was more significant than 86.40%, followed by annual herbs, accounting for a smaller proportion (Table 5). The important values of different life forms of the same restoration grassland were generally manifested as perennial herbs > annual herbs, among which, the important values of perennial herbs in alpine grasslands with HRD and MRD were relatively large. In contrast, the important values of annual herbs with MRD and HRD and alpine grassland with MRD were similar. The important values of annual herbs in alpine grassland with VLRD were more significant than those of the other four restoration grasslands. Among the alpine grassland type communities with different degrees of recovery, most of the species were perennial herbs, and only a small number of species were annual herbs; perennial herbaceous plants mainly include Poaceae, Asteraceae, Legumes, Rosaceae, and Sedges, and annual herbs were primarily limited to the two species of Sonchus oleraceus and Chenopodium glaucum. Compared with alpine grasslands with different recovery levels, alpine grassland with HRD had the most species, mostly perennial herbaceous plants, and the number of annual herbaceous plant species in MRD and LRD were roughly the same. Among the degraded grassland communities, perennial herbaceous plants mainly included Melissitus ruthenica, Oxytropis ochrocephala, Carex spp, Geranium pratense, and Cyperus rotundus L.; annual herbaceous plants mainly included Sonchus oleraceus and Chenopodium glaucum.

TABLE 5. Important values of the life forms of communities in different recovered degree grassland/%.
3.6 Changes in grassland species diversity of alpine grassland community with different restoration degrees
Variations of species diversity in different restoration degrees of alpine grassland are shown in Figure 3. The analysis of species diversity with different restoration degrees revealed that the species richness of HRD grassland was significantly higher than that of MRD, LRD, and VLRD grasslands (p < 0.05); among them, HRD grassland was the most abundant with 13 species, followed by MRD with 12 species, LRD with 11 species, and at least 10 species in VLRD grassland. The Simpson index showed that the differences between HRD grassland and MRD grassland were higher than that of VLRD grassland. However, the differences in the Simpson index were not significant, with values of 0.81 and 0.83, respectively. The Shannon–Wiener index of HRD and MRD grasslands were higher than that of VLRD grassland; however, the differences in the Shannon-Wiener index were not significant, with values of 1.93 and 1.99, respectively. The VLRD grassland diversity index was the lowest, indicating that the community’s species composition was simple. The dominance index of alpine grassland types with different restoration degrees differed significantly (p < 0.05). The dominance index of VLRD grassland was significantly higher than that of other grasslands with a degree of restoration, at 0.32. The uniformity indices of HRD and MRD grasslands were higher than those of VLRD grassland. The differences were not significant, and the values were 0.84 and 0.86, respectively.
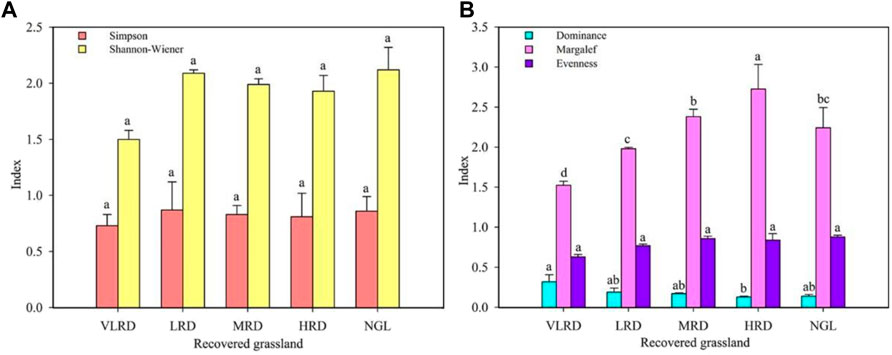
FIGURE 3. Plant diversity variation of the alpine meadow under different restoration degrees. (A), Simpson and Shannon–Wiener; (B), Dominance, Margalef and Evenness. VLRD, very low recovered degree; LRD, low recovered degree; MRD, middle recovered degree; HRD, high recovered degree; NGL, CK natural grassland. Different letters above error bars between treatments indicate significant differences at the 0.05 level.
3.7 Soil physical properties
The soil physical properties varied significantly among different recovered grassland (Figure 4). Specifically, soil pH, EC, and SMC in the 20–40 cm soil depths were considerably higher than in 10–20 cm soil depths (p < 0.05); soil pH and EC changed at different restoration levels, gradually decreasing with increasing restoration levels (Figures 4A,B), while soil SMC increased with increasing restoration levels (Figure 4C). Compared with the different restoration levels, the VLRD and LRD sample sites exhibited 40% and 18.85% higher soil pH at 10–20 cm soil depths than that of the NGL, while the VLRD, LRD, MRD, and HRD sample site exhibited 35.45%, 23.58%, 0.07%, and 0.06% higher soil pH at 20–40 cm soil depths than that of the NGL and the soil EC and SMC results were similar. In addition, the HRD recovery effect was close to that of the NGL, with the best recovery effect.
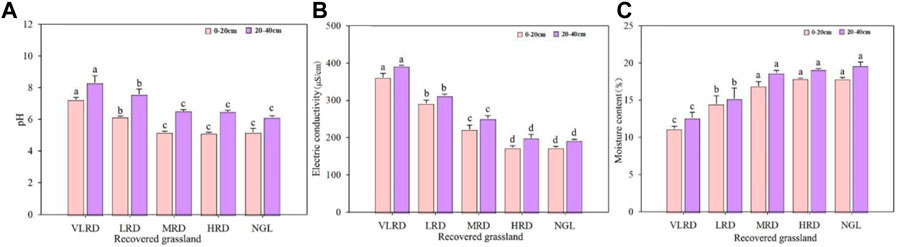
FIGURE 4. Soil physical properties of alpine meadow, pH (A), EC (B), and SMC (C), under different restoration degrees. VLRD, very low recovered degree; LRD, low recovered degree; MRD, middle recovered degree; HRD, high recovered degree; NGL, CK natural grassland. Different letters above error bars between treatments indicate significant differences at the 0.05 level.
3.8 Soil chemical properties
Differences in soil chemical properties at 0–40 cm depths under different restoration degrees grassland are shown in Figure 5. Soil SOC, TN, NO3-N, NH4-N, AN, TP, and AP at 0–20 cm soil depths were significantly higher than those at 20–40 cm soil depths (p < 0.05); the contents of SOC, TN, NO3-N, NH4-N, AN, TP, and AP were significantly higher in HRD grassland than those in VLRD (p < 0.05). The changes at different restoration levels gradually increased with increasing restoration levels at 10–20 cm and 20–40 cm soil depths. In different recovered grassland, the HRD recovery effect was close to that of the NGL, with the best recovery effect.
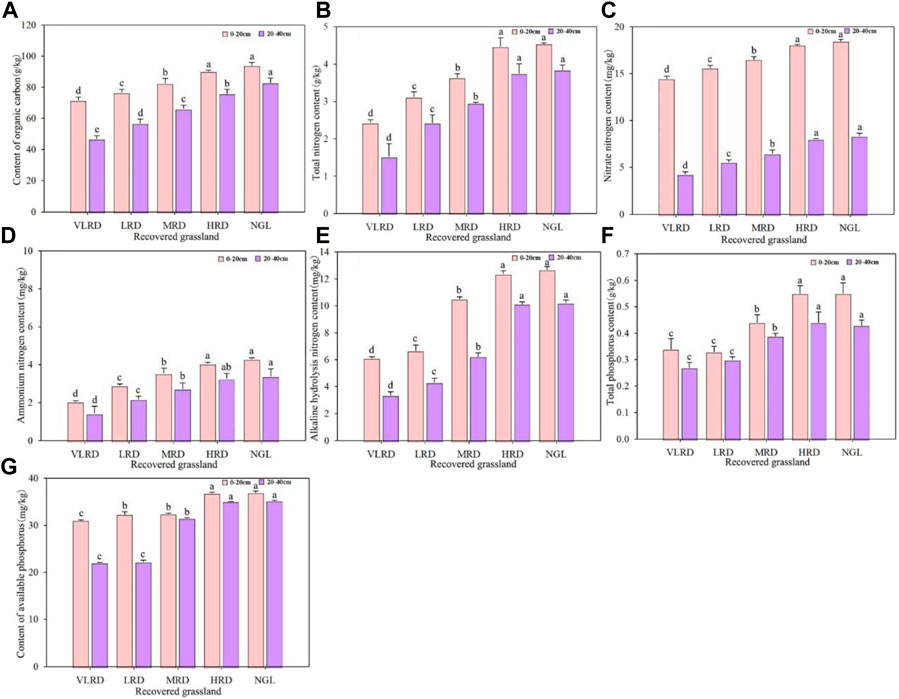
FIGURE 5. Soil chemical properties of alpine meadow, SOC (A), TN (B), NO3-N (C), NH4-N (D), AN (E), TP (F), and AP (G), under different restoration degrees. VLRD, very low recovered degree; LRD, low recovered degree; MRD, middle recovered degree; HRD, high recovered degree; NGL, CK natural grassland. Different letters above error bars between treatments indicate significant differences at the 0.05 level.
3.9 Potential driving factors of plant community diversity and soil physiochemical properties
Pearson correlation analysis of plant community biomass and soil physicochemical properties (Figure 6) revealed that soil SMC, AN, TP, NH4-N, TN, AP, SOC, and NO3-N contents at 0–20 cm soil depths were significantly and positively correlated with plant frequency, coverage, and biomass (p < 0.01), and soil pH and EC were significantly negatively correlated with plant frequency, coverage, and biomass (p > 0.05); further, soil TN was significantly and positively correlated with plant height (R = 0.67, p < 0.01) (Figure 6A). At 20–40 cm soil depths, SMC, AN, TP, NH4-N, TN, AP, SOC, and NO3-N contents were significantly and positively correlated with plant frequency, coverage, and biomass (p < 0.01), while soil pH and EC were significantly negatively correlated with plant frequency, coverage, and biomass (p > 0.05). Soil TP was significantly and positively correlated with plant height (R = 0.71, p < 0.01) (Figure 6B).
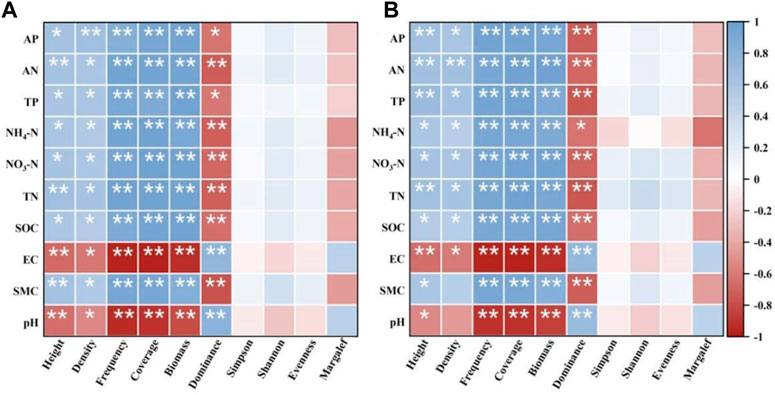
FIGURE 6. Correlations between plant community diversity and soil physiochemical properties in alpine recovered grassland at 0–20 cm soil depths (A) and 20–40 cm soil depths (B). pH, soil pH; SMC, soil moisture content; EC, soil electric conductivity; SOC, soil organic carbon; AP, soil available phosphorus; TP, soil total phosphorus; TN, soil total nitrogen; NO3-N, soil nitrate nitrogen; NH4-N, soil ammonium nitrogen; AN, soil alkaline nitrogen.
Principal component analysis was used to determine the correlation between soil physicochemical properties and the degree of soil recovery. The principal component analysis of soil biological factors at 0–20 cm soil depths of the recovered area of the Qianmalong coal mine revealed the first axis explained 92.67%, with the corresponding second axis explaining 2.54%, and the cumulative of the first two axes explaining 95.20% (Figure 7A). Soil physicochemical properties at 0–20 cm soil depths from HRD and NGL were positively correlated with SMC, AN, TP, NH4-N, TN, AP, SOC, and NO3-N, while the HRD was negatively correlated with pH at 0–20 cm soil depths. The first axis explained 93.63% of the 20–40 cm soil depths, with the corresponding second axis explaining 2.17% and the cumulative of the first two axes explaining 95.80% (Figure 7B). Soil physicochemical properties at 20–40 cm soil depths from HRD and NGL were positively correlated with SMC, AN, TP, NH4-N, TN, AP, SOC, and NO3-N, while the HRD was negatively correlated with pH at 20–40 cm soil depths.
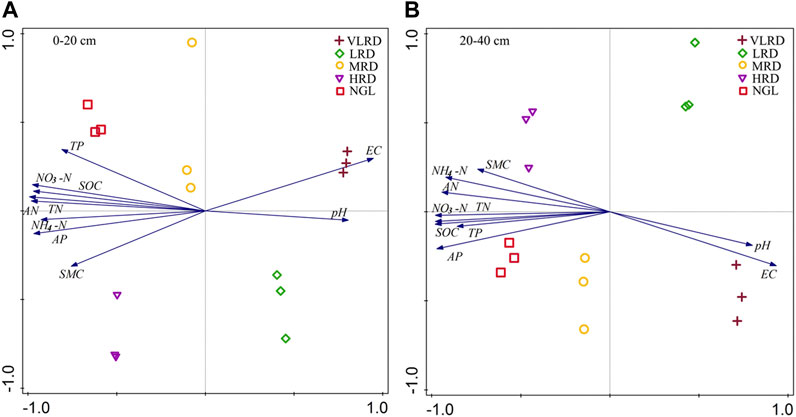
FIGURE 7. Principal component analysis (PCA) of soil physiochemical properties in alpine recovered grassland at 0–20 cm soil depths (A) and 20–40 cm soil depths (B). Arrows indicate the direction and magnitude of the soil physiochemical index associated. The length of the arrows in the PCA plot corresponds to the strength of the correlation variables. Each graph represents the recovered grassland for each sample (three replicates per sample, 1–3). Data are based on three replicates (1–3) per sample. pH, soil pH; SMC, soil moisture content; EC, soil electric conductivity; SOC, soil organic carbon; AP, soil available phosphorus; TP, soil total phosphorus; TN, soil total nitrogen; NO3-N, soil nitrate nitrogen; NH4-N, soil ammonium nitrogen; AN, soil alkaline nitrogen.
4 Discussion
4.1 Changes in plant characteristics, plant family, genus, species structure, community life structure, and community value
The structure of plant community families, genera, and species not only reflects the community characteristics of plants but also reflects the habitat conditions of plant communities (Zhao et al., 2022). The community species with different degrees of recovery in this study were mainly concentrated in five major families: Legumes, Goldenrods, Grasses, Roses, and Sedges; other families had fewer and scattered species. These five major families had strong ecological adaptability and could better adapt to the semi-arid climate and poor soil ecological restoration environment in the Qilian mountain Tianzhu mining area of China, which was the main part of the degradation degree community at each stage (Table 2). The composition of species was manifested in the fact that most species belong to a few families, and minority species belong to most families. As confirmed by our study, the plant community composition and species diversity in Northwest China are consistent with this community structure and party appearance (Huang et al., 2007). With the increase of grassland recovery coverage in the mining area, the biomass and total cover of alpine grassland increased significantly (Figure 2); the changing trend is similar to the result of Tian et al. (2022). In addition, our results confirmed that VLRD and LRD grasslands might be associated with roadside overgrazing due to their short recovery time and lack of stable growth patterns, thus requiring enhanced management of restored areas and prohibition of grazing.
The community life type structure is the main component of the community structure, which is the external expression of the comprehensive reflection of the habitat factors (Yang et al., 2017). The importance value usually indicates ecological adaptability, and its size is the only indicator to determine the dominant and subdominant species (Kaiser et al., 2016). The turnover of dominant species is a sign of community succession with different degrees of recovery (Ahirwal et al., 2018). This study found that the grassland in the alpine mining area at each restoration level mainly consisted of perennial herbs, followed by annual herbs (Table 3), which accounted for a small proportion. The former dominant species of Salicaceae gradually lost their dominance. Annual herbs appeared, eventually forming VLRD and LRD grassland types, with the dominance of Carex spp (Table 4); in this study, the plants exhibited strong adaptability to cold and arid climatic environments (Yang et al., 2016; Li et al., 2021). The HRD grassland was close to the community life type structure of NGL, which indicated that the restoration method was reasonable, with a noticeable restoration effect (Table 5). Due to the restoration of mine land, it was also possible that its species composition was simple and perennial herbs had absolute dominance due to the direct limitation of environmental conditions, resulting in a single population community development (Xue et al., 2006). It was also possible that the influence of precipitation and restoration time made the species composition and structure of perennial herbs more stable (Lehmann et al., 2020), while annual herbs were less stable, which is consistent with observations of Zhang et al. (2016), who conducted a study on vegetation community characteristics of alpine grassland mining areas on the Qinghai-Tibet Plateau, China. Therefore, annual herbaceous plants have short life cycles, most of which are axial-rooted. In contrast, typical alpine grasslands have high-quality forage grasses of the genus Gramineae and Sonchus, which are the most important resources for ecological conservation and belong to dense clumps of fibrous-rooted perennial herbaceous plants; these species will slowly increase with the restoration of the mine grassland environment and recovery time (Kang et al., 2021), effectively playing the role of soil and water conservation and change the ecological damage of the mining environment (Konig et al., 2023).
4.2 Changes in community species diversity
The community’s diversity needs to be comprehensively measured by three aspects of community richness, evenness, and ecological dominance, which can reflect the structural level of community species composition (De et al., 2016). The results of this study showed that with the increasing degree of mine grassland restoration, the diversity of species in mine-restored grassland showed an increasing trend; the HRD grassland community had the highest species richness and diversity index the lowest dominance. The VLRD grassland community had the lowest diversity index and community evenness but the highest dominance; the number of species was significantly lower than in other sample sites. The community stability and community evenness of HRD grassland were higher, indicating that the restoration method by mulching and seeding were suitable for local ecological restoration (Figure 3). The community stability of HRD grassland was similar to that of the undamaged NGL, accompanied by the emergence of ecologically protected plants of the genus Gramineae and Artemisia, which improved the process of mine restoration. This is consistent with the findings of Xu et al. (2005) and Chen et al. (2020) on the succession pattern of restored grasslands and with the study of Gornish et al. (2017) scholars on the negative relationship between the diversity index and ecological dominance. Proper grazing can reduce the competitive ability of dominant community species, increase the diversity level of the community, provide invasion and growth of weaker or new species, and enrich the diversity of community species to some extent (Huang et al., 2007), leading to high evenness, high richness, and low dominance of MRD grassland community.
4.3 Changes in potential driving factors of plant community diversity soil physiochemical properties
The community structure of the soil is sensitive to changes in soil physicochemical properties (Kathryn et al., 2012). Several studies have reported that the soil pH increases with increasing soil depth and decreases with increasing grass restoration (Bates et al., 2011; Lin et al., 2019). This study found a similar result: soil pH was higher at 20–40 cm soil depths. The reason is that the alpine grassland mainly contains herbaceous vegetation, wherein the root system is primarily gathered at 0–20 cm soil depths; therefore, microorganisms are enriched in the inter-root soil. The root system and the inter-root microorganism respiration produce CO2, and the inter-root water combination renders the pH of the surface soil negligible. As confirmed by our study, the trend of SMC, SOC, TN, NO3-N, NH4-N, TP, AN, and AP decreased with increasing soil depth and increased with increasing degradation, mainly because soil nutrient source is primarily concentrated in the upper layer; in addition to the low precipitation of alpine grassland, the root-rich surface soil is rich in bound water, root secretions, and microorganisms, resulting in relatively high nutrient content in the surface layer of soil (Xu et al., 2020). The results of PCA showed that the indices of recovered mine grassland with HRD were positively correlated with that of NGL, mainly because of the high vegetation biomass and cover in HRD, increased uptake of mineralized and animal excretion of fast-acting nutrients by plants, and increased soil nutrient content of HRD at the surface—with the degree of recovery (Liu et al., 2019).
In summary, our results confirmed that the communities of VLRD and LRD grasslands in the mine area of the Tianzhu Qianmalong coal mine were unstable and had less plant forage value. This was accompanied by the appearance of harmful plants, poor recovery, and is prone to degradation into bald spots, which should be improved with later fence protection on both sides of the road, fence closure, and a grazing ban for recreation (Li et al., 2021). Furthermore, secondary seeding maintenance can accelerate the recovery of poorly recovered grassland, and nurturing measures should be carried out (Fieldsjohnson et al., 2012), to improve the overall ecological restoration of the mine site and the ecological restoration of the mining ecosystem.
5 Conclusion
Vegetation characteristics, community structure, and species diversity of four different degrees of restoration and natural grassland in the Tianzhu Qianmalong coal mining area grassland were analyzed. We arrived at the following conclusions: the overall plant community showed a decreasing trend with the increase of restoration level of mine grassland, and the difference was significant (p < 0.05), in the order of HRD > NGL > MRD > LRD > VLRD. There were 11 families, 18 genera, and 17 species of plants in the study area; of them, 12% were Legumes with high ecological adaptability of plant communities, 29% were Asteraceae, 5% were Gramineae, 5% were Rosaceae, and 12% were Salicaceae. The four different restoration degrees and natural grassland vegetation types were Carex spp + Cyperus rotundus L., Cyperus rotundus L. + Carex spp, Cyperus rotundus L. + Elymus nutans, Cyperus rotundus L. + Elymus nutans, and Cyperus rotundus L. + Elymus nutans. The ecotype community of restored grassland in the alpine mining area showed the predominance of perennial herbaceous plants, with annual herbaceous plants being the least dominant and no tree or shrub layer. The very low recovered degree species dominance index of grassland communities was the highest in the stage of different degrees of restoration grassland in the alpine mining area. The community species richness and diversity indices were the highest in HRD grasslands. Soil pH and EC showed a decreasing trend, and SMC, SOC, TN, NO3-N, NH4-N, AN, TP, and AP content exhibited an increasing trend with the increase of grassland restoration. The index of mine restored grassland with HRD was closely related to the index of NGL with the best restoration effect.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
XY, QF, and WL: conception and design of the research. XY and HX: acquisition of data and drafting the manuscript. XY and JZ: analysis and interpretation of data. XY and CZ: statistical analysis. YF, ZW, and LY: revision of manuscript for important intellectual content. All authors contributed to the article and approved the submitted version.
Funding
This work was supported by the National Key R&D Program of China (No. 2022YFF1303301, 2022YFF1302603), the National Natural Science Fund of China (No. 52179026, 42001035, 42101115), the Science and Technology Program of Gansu Province (No. 22JR5RA072, 22JR5RA068), the Postdoctoral Funding Program of Gansu Province (No. E339880139), and the Open Fund of Technology Innovation Center for Mine Geological Environment Restoration in the Alpine and Arid Regions (No. HHGCKK2204).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adator, S. W., and Li, J. F. (2021). Evaluating the environmental and economic impact of mining for post-mined land restoration and land-use: A review. J. Environ. Manage. 279, 111623. doi:10.1016/j.jenvman.2020.111623
Ahirwal, J., and Maiti, S. K. (2018). Development of technosol properties and recovery of carbon stock after 16 years of revegetation on coal mine degraded lands, India. Catena 166, 114–123. doi:10.1016/j.catena.2018.03.026
Akala, V. A., and Lal, R. (2015). Potential of mine land reclamation for soil organic carbon sequestration in Ohio. Land Degrad. Dev. 11, 289–297. doi:10.1002/1099-145x
Bates, S. T., Berg-Lyons, D., Caporaso, J. G., Walters, W. A., Knight, R., and Fierer, N. (2011). Examining the global distribution of dominant archaeal populations in soil. ISME J. 5, 908–917. doi:10.1038/ismej.2010.171
Cappelli, S. L., Domeignoz-Horta, L. A., and Loaiza, V. (2022). Plant biodiversity promotes sustainable agriculture directly and via belowground effects. Trends Plant Sci. 9, 1878–1880. doi:10.1016/j.tplants.2022.02.003
Celestino, Q. S., Michel-Pierre, F., and Rimi, R. (2020). Plant functional traits on tropical ultramafic habitats affected by fire and mining: Insights for reclamation. Diversity 12, 248. doi:10.3390/d12060248
Che, R., Wang, Y., and Li, K. (2019). Degraded patch formation significantly changed microbial community composition in alpine meadow soils. Soil Tillage Res. 195, 104426. doi:10.1016/j.still.2019.104426
Chen, J., Mo, L., Zhang, Z., Nan, J., Xu, D., Chao, L., et al. (2020). Evaluation of the ecological restoration of a coal mine dump by exploring the characteristics of microbial communities. Appl. Soil Ecol. 147, 103430. doi:10.1016/j.apsoil.2019.103430
Choudoir, M. J., and Deangelis, K. M. (2022). A framework for integrating microbial dispersal modes into soil ecosystem ecology. iScience 25, 103887. doi:10.1016/j.isci.2022.103887
De Quadros, P. D., Zhalnina, K., Davis-Richardson, A. G., Drew, J. C., Menezes, F. B., Flávio, A. D. O., et al. (2016). Coal mining practices reduce the microbial biomass, richness and diversity of soil. Appl. Soil Ecol. 98, 195–203. doi:10.1016/j.apsoil.2015.10.016
Feng, Q. (2019). Theory and Technology of ecosystem protection and restoration in qilian mountains [M]. Beijing, China: Science Press. (In Chinese).
Fieldsjohnson, C. W., Zipper, C. E., Burger, J. A., and Evans, D. M. (2012). Forest restoration on steep slopes after coal surface mining in appalachian USA: Soil grading and seeding effects. For. Ecol. Manag. 270, 126–134. doi:10.1016/j.foreco.2012.01.018
Figueiredo, M. A., da Silva, T. H., Pinto, O. H. B., Leite, M. G. P., de Oliveira, F. S., Messias, M. C. T. B., et al. (2023). Metabarcoding of soil fungal communities in rupestrian grassland areas preserved and degraded by mining: Implications for restoration. Microb. Ecol. 2, 153. doi:10.1007/s00248-023-02177-y
Gao, Z. W., Liu, Z. Y., Wu, Z. H., Huang, J., Zhang, L., Zhang, G. G., et al. (2022). Soil nematode communities and functions: Ecological package restoration responses in damaged desert steppe open-pit mining ecosystems. Land Degrad. Dev. 11, 1–14. doi:10.1002/ldr.4208
Gornish, E. S., Lennox, M. S., Lewis, D., Tate, K. W., and Jackson, R. D. (2017). Comparing herbaceous plant communities in active and passive riparian restoration. Plos One 12, e0176338. doi:10.1371/journal.pone.0176338
Haddaway, N. R., Cooke, S. J., and Lesser, P. (2019). Evience of the impacts of metal mining and the effectiveness of mining mitigation measures on social-ecological systems in arctic and boreal regions: A systematic map protocol. Environ. Evid. 8, 9. doi:10.1186/s13750-019-0152-8
Huang, D., Wang, K., and Wu, W. L. (2007). Dynamics of soil physical and chemical properties and vegetation succession characteristics during grassland desertification under sheep grazing in an agro-pastoral transition zone in Northern China. J. Arid. Environ. 70, 120–136. doi:10.1016/j.jaridenv.2006.12.009
Kaiser, K., Wemheuer, B., Korolkow, V., Wemheuer, F., Nacke, H., Schoning, I., et al. (2016). Driving forces of soil bacterial community structure, diversity, and function in temperate grasslands and forests. Sci. Rep. 6, 33696. doi:10.1038/srep33696
Kang, B. T., Bowatte, S., and Hou, F. J. (2021). Soil microbial communities and their relationships to soil properties at different depths in an alpine meadow and desert grassland in the Qilian mountain range of China. J. Arid. Environ. 184, 104316. doi:10.1016/j.jaridenv.2020.104316
Kathryn, G. E., Debenport, S., Anderson, S., and Fierer, N. (2012). Digging deeper to find unique microbial communities: The strong effect of depth on the structure of bacterial and archaeal communities in soil. Soil Biol. biochem. 50, 58–65. doi:10.1016/j.soilbio.2012.03.011
Konig, L. A., Medina-Vega, J. A., Longo, R. M., Zuidema, P. A., and Jakovac, C. C. (2023). Restoration success in former Amazonian mines is driven by soil amendment and forest proximity. Philo. Trans. Ro. B-Bio. Sci. 378, 1867. doi:10.1098/rstb.2021.0086
Lehmann, J., Bossio, D. A., Kögel-Knabner, I., and Rillig, M. C. (2020). The concept and future prospects of soil health. Nat. Rev. Earth Environ. 1, 544–553. doi:10.1038/s43017-020-0080-8
Li, W. L., Liu, C. L., Wang, W. Y., Zhou, H. K., Xue, Y. T., Xu, J., et al. (2021). Effects of different grazing disturbances on the plant diversity and ecological functions of alpine grassland ecosystem on the Qinghai-Tibetan plateau. Front. Plant Sci. 12, 765070. doi:10.3389/fpls.2021.765070
Lin, Y., Ye, G., Kuzyakov, Y., Liu, D., Fan, J., and Ding, W. (2019). Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil boil. biochem. 134, 187–196. doi:10.1016/j.soilbio.2019.03.030
Liu, Y., Lei, S. G., and Gong, C. G. (2019). Comparison of plant and microbial communities between an artificial restoration and a natural restoration topsoil in coal mining subsidence area. Environ. Earth Sci. 78, 204. doi:10.1007/s12665-019-8195-2
Ma, K. P. (1994). A measure of biological community diversity A measure of alpha diversity (Top). Biodiversity 2, 162–168. (In Chinese).
Ma, R., Hu, F., Xu, C., Liu, J., and Zhao, S. (2022). Response of soil aggregate stability and splash erosion to different breakdown mechanisms along natural vegetation restoration. Catena 208, 105775. doi:10.1016/j.catena.2021.105775
Neldner, V. J., and Ngugi, M. R. (2017). Establishment of woody species across 26 years of revegetation on a Queensland coal mine. Ecol. Manag. Restor. 18, 75–78. doi:10.1111/emr.12243
Rania, R., Joana, D., and Omar, D. (2021). A tunnel under an in-pit mine waste dump to improve environmental and landscape recovery of the site. Minerals 11, 2–11. doi:10.3390/min11060566
Ren, J. Z. (1988). Grass industry scientific research methods [M]. Beijing: China Agricultural Publishing House, 122–136. (In Chinese).
Satake, A., Nagahama, A., and Sasaki, E. (2021). A cross-scale approach to unravel the molecular basis of plant phenology in temperate and tropical climates. New phytol. 233, 2340–2353. doi:10.1111/nph.17897
Sommers, L. E., and Nelson, D. W. (1972). Determination of total phosphorus in soils: A rapid perchloric acid digestion procedure. Soil Sci. Soc. Am. J. 36, 902–904. doi:10.2136/sssaj1972.03615995003600060020x
Thakur, M. P., Wim, H. P., and Wilschut, R. A. (2021). Plant-Soil feedback and temporal dynamics of plant diversity-productivity relationships. Trends ecol. evol. 36, 651–661. doi:10.1016/j.tree.2021.03.011
Tian, X. M., Yin, R. P., Wang, J., Dong, L., Cheng, B., Liu, H., et al. (2022). Microtopographic reconstruction improves soil erosion resistance and vegetation characteristics on the slopes of large dump sites in semi-arid areas. Front. Environ. Sci. 10, 930519. doi:10.3389/fenvs.2022.930519
Walkley, A., and Black, I. A. (1934). An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 37, 29–38. doi:10.1097/00010694-193401000-00003
Wang, C., Bender, S. F., Widmer, F., and Heijden, M. G. (2014). Soil biodiversity and soil community composition determine ecosystem multifunctionality. Proc. Natl. Acad. Sci. 111, 5266–5270. doi:10.1073/pnas.1320054111
Wang, D., Zhang, B., Zhu, L. L., Yang, Y. S., and Li, M. M. (2018). Soil and vegetation development along a 10-year restoration chronosequence in tailing dams in the Xiaoqinling gold region of Central China. Catena 167, 250–256. doi:10.1016/j.catena.2018.05.007
Wang, Y. C., Liu, G. X., and Yu, H. (2021). Meadow degradation increases spatial turnover rates of the fungal community through both niche selection and dispersal limitation. Sci. total Environ. 798, 149362. doi:10.1016/j.scitotenv.2021.149362
Xu, A., Li, L., Xie, J., Wang, X., Coulter, J. A., Liu, C., et al. (2020). Effect of long-term nitrogen addition on wheat yield, nitrogen use efficiency, and residual soil nitrate in a semiarid area of the loess plateau of China. Sustainability 12, 1735. doi:10.3390/su12051735
Xu, L. L., Zhang, X. Z., Shi, P. L., and Yu, G. R. (2005). Establishment of apparent quantum yield and maximum ecosystem assimilation on Tibetan Plateau alpine meadow ecosystem. Sci. China Earth Sci. 48, 141–147. doi:10.1360/05zd0014
Xue, D., Yao, H., and Huang, C. (2006). Microbial biomass, N mineralization and nitrification, enzyme activities, and microbial community diversity in tea orchard soils. Plant Soil 288, 319–331. doi:10.1007/s11104-006-9123-2
Yang, T., Adams, J. M., Shi, Y., He, J. S., Jing, X., Chen, L., et al. (2017). Soil fungal diversity in natural grasslands of the Tibetan plateau: Associations with plant diversity and productivity. New Phytol. 215, 756–765. doi:10.1111/nph.14606
Yang, X. M., Ma, J. P., Tang, J. N., Yang, T. B., and Ghebrezgabher, M. G. (2016). Dynamics of desert vegetation and its response to climate changes during 1982–2013 in the Hexi Corridor of Northwest China. IGARSS 2016. IEEE Int. Geosci. Remote Sens. Symp. 11, 4495–4497. doi:10.1109/IGARSS.2016.7730172
Zhang, Y., Dong, S. K., Gao, Q. Z., Liu, S. L., Zhou, H. K., Ganjurjav, H., et al. (2016). Climate change and human activities altered the diversity and composition of soil microbial community in alpine grasslands of the Qinghai-Tibetan Plateau. Sci. Total Environ. 562, 353–363. doi:10.1016/j.scitotenv.2016.03.221
Zhao, M. T., Wang, C. Q., He, Y. S., and He, T. H. (2022). Soil characteristics and response thresholds of salt meadow on lake beaches of the Ordos platform. Front. Environ. Sci. 10, 1050757. doi:10.3389/fenvs.2022.1050757
Keywords: alpine mining areas, grasslands, restoration, plant diversity, community, Qilian Mountains
Citation: Yang X, Feng Q, Liu W, Xia H, Zhang J, Yang L, Zhang C, Wang Z and Feng Y (2023) Community structure and plant diversity under different degrees of restored grassland in mining areas of the Qilian Mountains, Northwestern China. Front. Environ. Sci. 11:1191599. doi: 10.3389/fenvs.2023.1191599
Received: 05 April 2023; Accepted: 18 May 2023;
Published: 05 June 2023.
Edited by:
Adriano Sfriso, Ca’ Foscari University of Venice, ItalyCopyright © 2023 Yang, Feng, Liu, Xia, Zhang, Yang, Zhang, Wang and Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qi Feng, cWlmZW5nQGx6Yi5hYy5jbg==