 Lucile Chauvet1,2†
Lucile Chauvet1,2† Isabelle Le Huërou-Luron
Isabelle Le Huërou-Luron Amélie Deglaire
Amélie Deglaire- 1STLO, INRAE, L’Institut Agro, Rennes, France
- 2Institut NuMeCan, INRAE, INSERM, Univ Rennes, Saint Gilles, France
- 3SODIAAL International, Centre Recherche AND Innovation, Rennes, France
This review examines the differences in protein composition, digestion, and physiological effects on infants between human milk (HM) and infant formula (IF). The World Health Organization recommends exclusive breastfeeding for the first 6 months of life due to the numerous health benefits associated with it. However, when this is not possible, IF is used as an alternative. Differences between HM and IF remain, particularly in terms of protein composition and structure. Further optimization of IF is needed to better mimic HM and provide similar health benefits. Further improving IF formulation requires implementing a promising strategy, which in turn requires a thorough understanding of the mechanisms of protein digestion and amino acid (AA) absorption, as well as the metabolic and physiological effects of protein composition and structure. These are often altered by heat treatment and processing in IF. The main differences in the protein composition and structure of HM and IF are presented, including a synthesis of knowledge on the non-protein nitrogen (NPN) fraction. This fraction is too often neglected in milks, despite accounting for one fifth of the total nitrogen in HM. The influence of the protein composition and structure of HM and IF on the digestion of dietary protein and dietary AA absorption is compared, highlighting the need for data on the postprandial AA profile in infants from well-designed clinical trials. Finally, this review examines the differences in protein composition and digestion between HM and IF that lead to distinct metabolic, physiological and microbial outcomes. Future research should focus on understanding the role of partially digested proteins and the NPN fraction in shaping the infant gut microbiota and overall health.
1 Introduction
The health benefits of breastfeeding for infants are globally recognized, and the World Health Organization (WHO) recommends an exclusive breastfeeding until 6 months of age (1). However, the rate of exclusive breastfeeding among infants aged 0–6 months remains low worldwide (48%) (1). Non-breastfed infants are therefore fed infant formula (IF), which aims to mimic as much as possible mature human milk (HM). Thanks to increased knowledge and scientific developments, the composition of IF has been improved over the past decades, in the frame of the European regulation (2). The generic formulation and processing route for producing powder IF is described hereafter (3). IFs, for healthy term infants, are commonly formulated from skimmed cow milk (CM), although other protein sources can be used, such as goat milk or soy protein isolates. However, this will not be covered in the present review. In standard IF, the skimmed milk is enriched with whey proteins (WP) to mimic the average casein:whey protein ratio (40:60) of mature HM and thus cover the regulatory aminogram (2). Free amino acids (AAs) may also be added to standard IF to decrease the IF total protein content as some studies have reported that high protein content in IFs may contribute to infant overweight and obesity (4, 5). Purified whey proteins such as lactoferrin (LF), α-lactalbumin, and/or osteopontin can also be used in some specific IFs. Lactose (and sometimes maltodextrin, although not present in HM), fat (usually based on a mixture of vegetable oils, to which milk fat ingredients can be added), minerals and vitamins are used for the IF formulation. The liquid preparation obtained is heat-treated to ensure sanitary safety, the mix is then concentrated usually before or after fat addition and homogenization for emulsion stability (average droplet diameter: 0.1–1 μm). The mix is finally spray dried and packaged (3).
Despite the progress made by industrials in the last decades, discrepancies between HM and IF remain particularly in terms of fine composition and structure, referring here to protein quality, and resulting in different physiological properties in the infant. The present review aims to highlight the differences of composition and structure of proteins between HM and standard IF. The fate of digestion, as well as the metabolic and physiological impacts of HM vs. IF will then be addressed.
2 Proteins and non-protein nitrogen in human milk vs. infant formula
Milk nitrogenous compounds consist of a fraction of protein nitrogen (PN) and, to a lesser extent, a fraction of non-protein nitrogen (NPN). Milk proteins provide bioactive peptides and AAs required for the synthesis of protein contributing to the growth of the infant and the structural and functional development of its organs and tissues, but also for the synthesis of non-protein nitrogen compounds (NPN) (6, 7). Milk NPN fraction, identified as the acid-soluble nitrogen obtained after protein precipitation, consists of more than 10 classes of compounds, the role of which for infant development is still being discussed.
The impact of different IFs and breastfeeding on atopic diseases and growth in pediatric cow’s milk protein allergy, as well as the effects of food processing on allergenicity, have recently been reviewed (8, 9). This topic is not presented here, as the focus is on the impact of the nature of proteins in human milk and cow’s milk-based infant formulas on digestion and gut physiology, which are less documented areas.
2.1 Proteins
The HM protein content (0.8–1.2 g/100 mL) is one of the lowest among other mammal milks, including donkey milk. However, it can rise to as much as 6 g/100 mL in some mammal milks, such as sheep’s milk. This correlated with the growth rate of infants, which is one of the lowest for humans and donkeys, and faster for sheep (10). Milk proteins are classified into three major classes: WPs, caseins and mucins (11). HM is a whey-dominant milk with a casein: WPs ratio of 40:60 in mature milk, while CM, the main source of protein and lactose in IFs, is a casein-dominant milk with a casein: WPs ratio of 80:20 (12, 13). Because of these differences, the manufacture of IFs requires the enrichment of their protein fraction in WPs to mimic the AA profile of HM. This enrichment generally results in an average casein: WPs ratio of 40:60 for IFs, ranging from 30:70 to 80:20 (benchmark on 35 IFs available in France made in 2023). Despite this rebalance, the nature of proteins remains different between HM and IFs, resulting in a higher true protein content in IFs than in HM (14, 15).
2.1.1 Whey proteins
WPs have major relevance for the infant development because of their nutritional input and their bioactive functions, but their nature and concentration vary between HM and IFs.
2.1.1.1 α-Lactalbumin
α-La has a high nutritional value as mainly composed of essential AAs (63.2% of total AA content). α-La is a protein rich in cysteine (6.5% of residues vs. 3.1% in β-LG), and also in tryptophan (3.3% AA residues vs. 2.5% AA residues in β-LG), an essential AA involved in important metabolic pathways allowing brain maturation and the development of sleep–wake rhythm, through serotonin and melatonin synthesis (16). Human and bovine α-La present a similar structure and a high level of homology (~72%) (14), but a few differences in their glycosylation pattern might partly affect their functionality. α-La is the main whey protein in HM, accounting for ~27% of total proteins, while it is only the second most abundant whey protein in IFs, representing ~9.6% of total proteins and being almost three-fold less concentrated than in HM (17, 18). To balance the low-level of α-La in standard IFs, and thus of cysteine and tryptophan, it is necessary to increase the true protein content in IF compared to HM or to formulate an α-La-enriched IF, thus allowing to have a lower protein content in IF. The addition of free AAs is also another way to balance the low-levels of α-La in standard IFs. The latter strategy might be beneficial for the infant (19–21), however this can be costly and has to be done in agreement with the regulation. In addition, sensory and nutritional consequences of this free AA supplementation should be considered, as free AAs could modify the IF organoleptic properties and may not remain stable along the shelf-life of the IF. Finally, free AA bioavailability is expected to differ from that of AA-bound protein. This should be further examined to evaluate the impact of such strategy. Clinical studies have demonstrated that α-LA-enriched IF may promote gastrointestinal tolerance and plasmatic AA concentrations in a similar way to that of HM-fed infants (19, 22–25).
2.1.1.2 β-Lactoglobulin
In HM, β-lactoglobulin (β-LG) is totally absent while it is the most abundant WPs in CM accounting for ~50% of total WPs. β-LG is one of the main allergens found in cow’s milk, although infants with allergies are usually sensitive to several proteins found in cow’s milk, such as caseins or α-La (26). IgE-mediated food allergy reactions to dairy proteins can evolve during the first years of life. An allergic reaction to β-lactoglobulin is commonly reported at birth, before evolving towards caseins and then α-La by the end of the first year (27).
2.1.1.3 Lactoferrin
Lactoferrin (LF) is a protein partially resistant to gastrointestinal digestion. Multiple physiological roles have been reported such as antimicrobial, immunomodulatory, anti-inflammatory, bifidogenic, anticarcinogenic, enzymatic and gene regulation activities (17, 28–32). With an average of 0.15 g/100 mL, LF is the second major WP of HM, accounting for ~16% of total proteins (17). LF level in CM is 10 times lower than in HM. Human and bovine LFs have a rather high homology (69%). LF supplementation in IFs has shown some benefits for infants, with a better weight-gain up to 6 month-old, a reduction of some infant diseases (e.g., lower respiratory tract illnesses), and the modulation of the gut microbiota that better mimics HM-fed infant microbiota (24, 33, 34). In vitro evaluation of the antiviral activity of LFs showed that the commercial sources of available bovine milk LFs, recombinant LFs and native human/bovine milk LFs influenced immune responses by significantly and variously modulating pro-inflammatory cytokine gene expression (35). The variation in LF bioactivity may be due to differences in processing conditions (e.g., thermal and high-pressure treatments), iron saturation, and purity (35, 36).
2.1.1.4 Immunoglobulins
Immunoglobulins (Igs) are the largest milk proteins. They are the main antimicrobial substances in milk and are divided into four classes: IgAs, IgMs, IgEs and IgGs. Taking all classes together, HM contains far more IgAs than CM. In HM, IgAs make up to almost 90% of total Igs (0.6 g/L in mature HM), while the major class in CM is IgGs (0.6 g/L) (37). However, in IFs the reported levels of secretory IgGs and IgAs are very low (38). Bovine milk immunoglobulins could maintain their structure resisting the temperature up to 75°C for 15 s, albeit losing their antigen-binding efficacy (39). In addition, the technical application for Holder pasteurization further influenced the retention rate of Igs and LF in HM (40).
2.1.1.5 Proteose-peptone
A minor fraction, called “proteose-peptone,” is present in WPs. Although little information exists in HM, the “proteose-peptone” fraction has been detailed in CM. It is a complex heterogeneous mixture of heat-resistant WPs divided in two classes according to their origin. The first class comprises non-hydrophobic, highly soluble fragments derived from proteolysis of the N-terminal region of β-casein by bovine plasmin, designated PP5, PP8S and PP8F. The second class consists of a complex heterogeneous group of hydrophobic glycoproteins, the main one being PP3, also known as LP28, and belonging to the fat globule membrane of CM (41). In CM, the “proteose-peptone” fraction accounts for 10% of total WPs. Differences in the “proteose-peptone” fraction composition exist between HM and CM (42). In HM, this fraction contains ~ 45% carbohydrates, whereas in CM it contains only ~ 11%. Regarding IFs, even though its concentration is not well characterized, the “proteose-peptone” fraction is assumed to be present in WP ingredients used for IFs.
2.1.1.6 Glycomacropeptides
WPs used for IF usually derive from cheese making and thus contain a glycomacropeptide (GMP) fraction corresponding to the carboxyl-terminal fragment of casein-κ (43–106) cleaved by the action of chymosin used for cheese making. GMP accounts for 9–15% of total protein in IFs (107). Nutritionally, GMP is of little interest for infants as it lacks aromatic AAs (phenylalanine, tryptophan, tyrosine), cysteine and only has a single methionine residue, whereas it is rich in branched-chain AAs (isoleucine, leucine, valine) and threonine. In HM or IFs derived from ideal whey (obtained after skimmed milk microfiltration), GMP is absent. However, it is released by digestive enzymes during the first step of gastric digestion. It has been reported in adults and animal models that GMP may exert some health-promoting activities (108, 109) and modulate microbiota composition and immune system response (22, 110–114), although these effects are not always observed (115). GMP is considered as a bioactive peptide in the literature (108, 109, 116), but there was no evidence of the concentration at which GMP in IFs may have a physiologic impact (115).
2.1.2 Caseins
Caseins are mainly under micellar forms in HM and IFs (90–95%), but human casein micelles are smaller than the bovine ones (30 to 75 nm in HM vs. 100 to 200 nm in CM) (117). Bovine casein micelles are composed of four casein species - αs1-, αs2-, β- and κ-caseins - with a molar ratio of 4:1:3.5:1.5. However, only three of these are found in HM: αs1-, β- and κ-caseins, with a molar ratio of 1.5:7:1.5 (13, 118). Although HM and CM both contain β-caseins, they only share ~50% sequence homology and have different numbers of phosphorylation sites (0 to 5 in HM vs. 4 to 5 in CM) (118). During gastrointestinal digestion of β-caseins, small peptides rich in phosphorylated AA residues, known casein phosphopeptides (CPPs), are formed. These CPPs facilitate the absorption of calcium, zinc and other divalent cations as they are able to keep these cations soluble (119).
A commercial fraction of purified β-casein can be added to IFs to increase the β-casein content and achieve the proportion of β-casein present in HM. In 2022, using an in vitro digestion model, Huang et al. (120) investigated the effects of supplementing the IF with either β-casein or α-LA on digestion. They highlight that increasing the IF content of one of these proteins resulted in a digestion profile that better mimicked HM digestion. However, further research is needed to confirm these preliminary results.
The interest of non-micellar casein and its impact on IF structure, digestive behavior and physiological consequences will be discussed in a dedicated section in this review.
2.2 Non protein nitrogen
Very few recent reviews have been carried out to synthesize knowledge on this fraction, a too-often neglected fraction in milks, even though it accounts for one fifth of total N in HM (Table 1). The NPN fraction of milk contains more than 10 classes of compounds: urea, peptides, free AAs, creatine, creatinine, uric and orotic acids, ammonia, carnitine, choline, amino alcohols of phospholipids, amino sugars, nucleic acids and nucleotides, polyamines, low molecular weight peptide hormones and other biologically active compounds such as growth factor (121) (Table 2). Their concentration depends on the mammal species, but also on other parameters such as lactation stage, time of the day, diet, prematurity, etc. (122, 123).

Table 1. Concentration of total, protein and non-protein nitrogen, urea nitrogen and free amino acids nitrogen in human milk, cow milk and cow milk-based infant formula (mean ± SD).
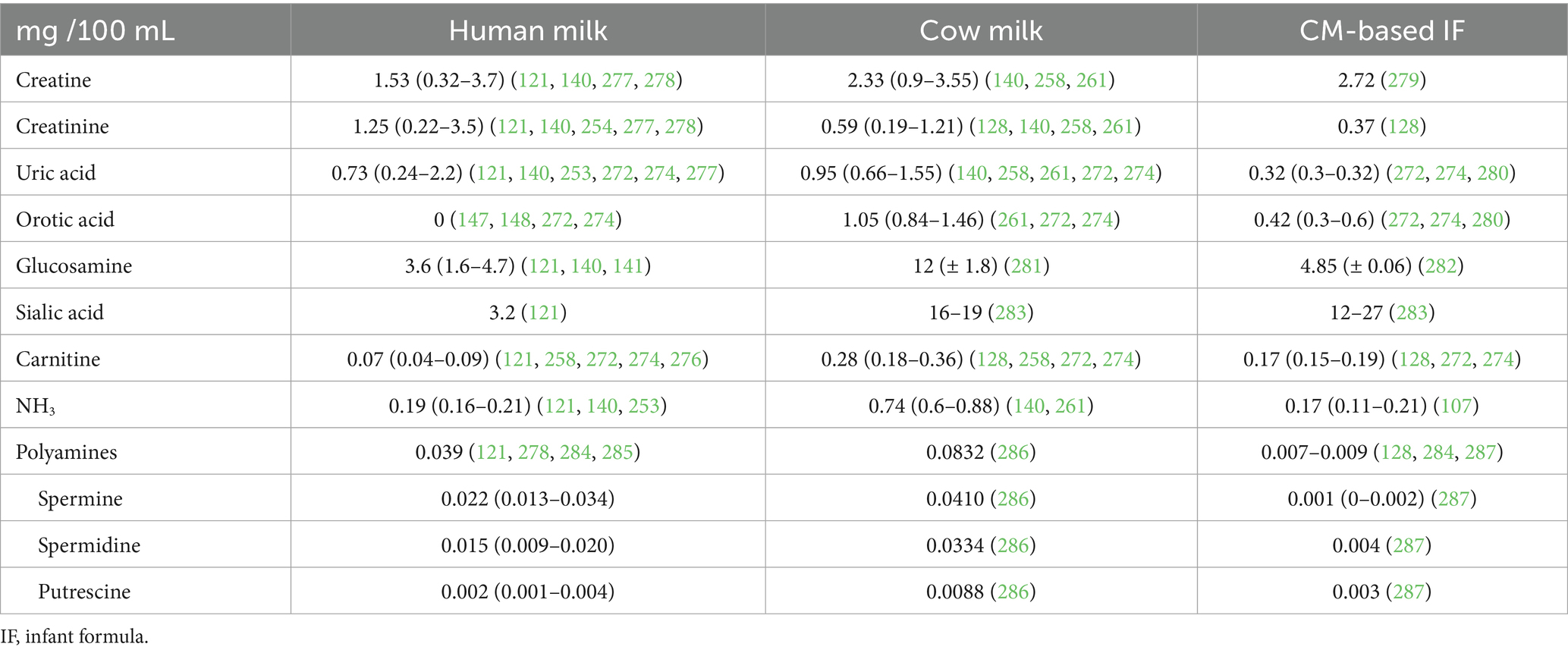
Table 2. Concentration of creatine, creatinine, uric acid, orotic acid, glucosamine, sialic acid, carnitine, ammoniac and polyamines in human milk, cow milk and in cow milk-based infant formula (min - max).
The NPN fraction in HM accounts for approximately 20% of the total N (11.5–25.9%) in HM while it only accounts for approximately 6% of the total N in CM (4.2–9.9%). In CM-based IFs, the NPN content is variable among brands, accounting on average for 9% of the total N (4.9–13.0%) (Table 1).
The variation of the NPN fraction in CM is often attributed to the variation of the urea level. The urea concentration in CM depends on factors related to the cow’s diet, such as dry matter content, crude protein content, percentage of rumen degradable and rumen non-degradable protein, energy/protein ratio, amount of easily digestible carbohydrates, and water intake. It is also influenced by physiological factors such as breed, body weight, mammary gland health, stage of lactation, age and parity (primiparous or multiparous) of cows, and seasonal factors (124, 125). Milk transport and storage time and processing technologies also play a significant role in modulating NPN concentration. The level of NPN increases with transport and storage time. It is also higher in cheese-derived whey due to its production process (hot or cold ripening) than in ideal whey obtained after skim milk microfiltration (126). The levels of NPN in IFs were reported to be highly dependent on the type of whey used. As a matter of facts, demineralized whey can be obtained from several processes such as ion-exchange and electrodialysis, but also processes such as micro- and ultra-filtration. In a study, Donovan and Lönnerdal (127) demonstrated that the level of NPN was the highest in ion-exchange whey, followed by electrodialyzed and ultrafiltered whey. The ultrafiltered whey contained the lowest amount of peptides in the NPN fraction due to the 10 kDa filtration, whereas the ion exchange demineralized whey contained a low amount of highly charged free AAs (Lysine, Glutamic acid, Arginine) (127). In a more recent study on IFs (126), the authors confirmed that demineralized cheese whey contained a higher content of NPN than the demineralized ideal whey.
2.2.1 Urea
Urea is the main NPN component in HM and CM, accounting for around 45 and 40% of this fraction, respectively. This represents 9% of the total N in HM and less than 3% of total N in CM. There is very little data available on the urea content of CM-based IFs (128), but it is estimated to account for around 43% of the NPN fraction and approximately 4% of the total N in IFs. N-urea becomes available after being released by intestinal bacterial ureases, which are expressed by some Bifidobacterium species (129, 130). Thus, the composition of the microbiota plays an important role in urea production.
2.2.2 Free amino acids
Milk NPN also contains a fraction of free AAs, which is well-documented in the literature for HM, CM and CM-based IFs. The total free AA content (mg N/100 mL) in HM is 4 times higher than in CM. The free AA fraction reaches 13% of the total NPN fraction in HM and 4.7% in CM. In HM and CM, the free AA fraction is mainly composed of proteinogenic AAs (88 and 74% of total free AAs, respectively) and a small fraction of non-proteinogenic AAs, mainly taurine in HM and carnitine in CM. The most abundant free AA is glutamic acid (39 and 27% of total free AAs in HM and CM, respectively), the second most abundant is glutamine for HM (18%), carnitine for CM (19%), and the third most abundant is taurine in HM (10%) and histidine for CM (13%). Taurine is only the fifth most abundant free AA in CM with a concentration 5 times lower than in HM.
In CM-based IFs, free AAs represent an average of only 6% of the NPN, consisting mainly of proteinogenic AAs (55% of the total free AAs). The non-proteinogenic AA fraction mainly consists of taurine (33% of total free AAs) and carnitine (12% of total free AAs), both of which can be added to IFs. As a result, taurine is the first most abundant free AA (33% of total free AAs), followed by carnitine, serine and glutamic acid, in CM-based IFs. Differences between IF brands are observed because different free AAs (other than taurine) can be added and different protein sources can be used for IF formulation.
Free AAs may be beneficial for infants. They can be directly absorbed and metabolized (121, 131). Glutamine and taurine, for example, which are highly represented in the free AA fraction, play essential roles during infant growth. Glutamine, in particular, is the precursor of non-essential AAs, including proline and arginine, which are produced by the intestinal mucosa (132). Taurine indirectly aids fat absorption by conjugating with bile acids to form bile salts (133, 134). However, studies have shown that taurine supplementation in IF does not modulate fat absorption; rather, it contributes to the development of a healthy microbiota, as taurine-conjugated bile salts are less toxic to Bifidobacteria than glycine-conjugated bile salts (135, 136).
2.2.3 Amino sugars
Milk provides N-containing oligosaccharides and amino sugar-containing glycoproteins and glycopeptides. Sialic acids (N-acetyl-neuraminic acid, Neu5Ac in HM and CM; N-gycolylneuraminic acid, Neu-5Gc, in CM), glucosamine and galactosamine are amino sugars present in the NPN fraction (288, 289). Sialic acid is very abundant in HM, with 70 to 83% bound to HM oligosaccharides, 14 to 28% bound to glycoproteins, 2 to 3% in free form, and 0.3% bound to glycolipids (137). Because amino sugar-containing compounds are acid soluble and oligosaccharides are the third most abundant solid component in HM [5 to 15 g/L in mature milk, (138)], nitrogen from amino sugars contributes to a significant portion of the NPN fraction in HM (121). By contrast, oligosaccharides are only present in trace amounts in CM (30 to 60 mg/L) (139). Three studies on HM reported the content of glucosamine (121, 140, 141) and one study reported the content of sialic acid in HM (121). The results showed that glucosamine N accounted for 9.1% of NPN and sialic acid nitrogen for 8.2% of NPN, making them the third and fourth most abundant N compounds in the NPN of HM. Glucosamine and galactosamine contribute to the development of the microbiota, especially Bifidobacteria.
2.2.4 Creatine and creatinine
Creatine and creatinine contribute, respectively, 3.9 and 3.2% to the NPN in HM, and 7.5 and 1.9% in CM. In IF, there was insufficient data to determine the contribution of creatine and creatinine to the NPN fractions.
2.2.5 Uric and orotic acids
Uric acid constitutes 1.9% of NPN in HM, 3% in CM, and 1.5% in CM-based IFs. Orotic acid is absent from HM, despite accounting for 3.4% of NPN in CM. There is no data available for orotic acid in CM-based IFs.
2.2.6 Ammonia, nitrite and nitrate
Ammonia contributes 0.5% of the NPN in HM, and 2.4% of NPN in CM. Interestingly, ammonia contributes an average of only 0.8% of NPN in CM-based IFs.
Nitrite accounts for 0.24% of NPN in HM, whereas it is negligible in CM and CM-based IFs, accounting for just 0.0002 and 0.0001%, respectively. In contrast, nitrate is less abundant in HM than in CM and CM-based IFs, accounting for 0.001, 0.17, and 0.09% of NPN, respectively (142).
2.2.7 Polyamines
The contribution of polyamines (spermine, spermidine, putrescine and cadaverine) to the NPN fraction is low in milk (290). Studies have shown that the polyamine concentration is much higher in HM than in IF (0.093–0.140 mg/100 mL HM vs. 0.014 mg/100 mL IF) and intermediate in CM (0.041 mg/100 mL CM) (128, 143, 144). However, only the contribution of N from polyamines to the NPN fraction in HM has been calculated (0.1% of NPN) (134, 143). There is insufficient data on IFs to determine the contribution of polyamines to the NPN fraction, but it is expected to be negligible in comparison to CM.
Despite their low content, polyamines are thought to be necessary for optimal gastrointestinal growth, as they are assumed to contribute to the regulation of cell growth and proliferation (143), and to modulate the composition of the microbiota. This has been observed in neonatal mice fed a polyamine-supplemented IF (145).
2.2.8 Nucleos(t)ides
The concentration of most nucleos(t)ides tends to decrease gradually as lactation progresses, but the decrease is less abrupt in HM than in CM (146–148). Similar compositions are observed in human and bovine colostrum, whereas differences in composition and concentrations are reported in mature milk (141, 143). Although the concentration of ribonucleotides reported in HM varies widely (10 to 200 μmol/L) (148), it appears that HM contains a higher concentration of nucleos(t)ides than CM. Cytidine and adenosine derivatives represent ~25% of the total nucleotides in HM, a higher amount than in CM. In HM, the concentrations of cytidine monophosphate (CMP), adenosine monophosphate (AMP), guanosine monophosphate (GMP), uridine monophosphate (UMP) were found to be 0.076 ± 0.006 mg N/100 mL, 0.105 ± 0.061 mg N/100 mL, 0.024 ± 0.004 mg N/100 mL, 0.032 ± 0.008 mg N/100 mL, respectively (141, 142, 144). GMP, UMP, and inosine monophosphate (IMP) were not detected in CM (140, 142–144). The usual concentration of ribonucleotides (mono- and diphosphate) is about 1 to 2 orders of magnitude higher than that of ribonucleosides (adenosine, guanosine, thymine, uridine, cytidine) in HM (148). In IF, the concentration of ribonucleos(t)ides appears to be a very negligible fraction of NPN, although insufficient data are currently available. Due to the limited data available and the variability of the values reported in the literature, it is difficult to assess the contribution of nitrogen from ribonucleos(t)ides to the NPN fraction in milk. However, nucleotides can be added to IF, which is a common practice.
2.2.9 Amino alcohols
Amino alcohols such as phosphoethanolamine, ethanolamine, phosphoserine, phosphatidylcholine, sphingosin, which mainly come from MFGM, and unesterified choline contribute to the NPN fraction. However, there is very little data on their concentrations in milk available in the literature. In 1989, Atkinson and Lönnerdal (121) estimated that 0.6 to 2 mg N/100 mL could be derived from phosphoethanolamine and phosphatidylcholine, and 0.3 to 0.9 mg N/100 mL from unesterified choline, in HM. Overall, amino alcohols may account for 2–7% of the NPN fraction in HM. There are insufficient data in the literature to evaluate their contribution to NPN in CM or in CM-based IFs.
Overall, although IFs are designed to closely resemble HM in terms of composition, there are still differences in their fine composition and structure. These differences are mainly due to the different milk source (bovine vs. human). HM is a bioactive fluid whose composition varies during lactation and depends on the mother’s diet. In contrast, IFs which are formulated to meet the infant’s nutritional needs between 0 and 6 months. IFs have a higher protein content to provide sufficient essential AAs to meet the infant’s nutritional needs, whereas HM provides other bioactive components that are partially lacking in IFs (149).
3 Protein digestion and amino acid absorption in infant
At birth, the infant faces many environmental changes, including a change in the way nutrients are delivered. Initially supplied by the placenta during the fetal period, ingested food is destructured throughout the infant’s gastrointestinal tract to allow for nutrient release and absorption, exposing the digestive system to compounds other than those present in the amniotic fluid (150). To meet this challenge, the infant’s organism and metabolism adapt and change during the first 2 years of life. This period is known as the critical period (151) is characterized by the infant’s extreme sensitivity due to the functional immaturity of many tissues and organs. Therefore, it is essential to control the environment to ensure the infant’s optimal growth and metabolic development.
3.1 Digestive specificities in infants
Maturation of digestive functions begins early in utero, with enteral feeding possible as early as 29 weeks of amenorrhea (152), but the digestive system is still immature at birth, which affects the infant’s ability to digest and absorb nutrients, including proteins.
3.1.1 Protein digestion
Protein digestion starts in the gastric phase, where proteolysis is carried out by pepsin, secreted as pepsinogen and activated autocatalytically at a pH below 4 (153). Despite active acid secretion from birth, the infant’s gastric mucosa is still highly susceptible to acidity during the first two to 3 weeks of life, and thus prostaglandins (such as PGE2) stimulate mucus and bicarbonate production, inhibit acid secretion and increase mucosal hydrophobicity (154). As a result, the gastric pH is between 6.0 and 6.5 immediately after feeding and does not reach pH 5.0 until 80 min after feeding due to the high buffering capacity of the milk diet (HM and IF) (155). At this pH, the rate of conversion of pepsinogen to pepsin is low and the activity of pepsin is probably reduced, as its pH optimum has been reported to be 2.0 for the hydrolysis of globular proteins (156) and may be different for the unstructured proteins that are caseins. The level of pepsin activity at 4 weeks postpartum has been reported to be 18% of that in adults (152) and to reach 100% only at 2 years of age (125, 148, 151). Gastric proteolysis in infants is thus incomplete as compared to adults. A previous study (153) showed that the type of diet (HM vs. IF) has no effect on pepsin production during infancy. In vivo, gastric proteolysis in nine full-term neonates averaged 15% of ingested protein (157), while in vitro gastric proteolysis of 5 to 15% of ingested protein is often reported for a static, semi-dynamic, or dynamic model of term neonates (120, 152–155). Overall, this suggests that a large proportion of dietary proteins enter the intestinal compartment partially or non-hydrolyzed. Several studies have reported that the rate of gastric emptying is influenced by the type of diet. HM is assumed to have a faster gastric emptying than IF, with a T1/2 of 36 to 48 min in preterm and term infants for HM (158) and a T1/2 of 65 to 78 min in preterm and term infants for IF (159). Two recent in vivo studies also confirmed faster gastric emptying with HM than with IF (157, 160). Several factors have been reported to influence gastric emptying, including bolus volume, caloric density, lipid quality, and protein type and structure (161).
The pancreas plays an important role for intestinal proteolysis through its secretory activity of major proteases (162). Pancreatic juice contains inactive proteases in the form of zymogens and contribute to pH increase of the acidic gastric chyme thanks to sodium bicarbonate. The activation of these zymogens is the result of cascade mechanisms. Trypsin is the most important pancreatic enzyme, accounting for 20% (w/v) of the total protein in pancreatic juice. Its activity is detected as early as the 16th week of amenorrhea and increases throughout the fetal period, reaching 90% of that of adults at birth (43, 163). In infants, chymotrypsin activity is estimated to be 50 to 60% of that of children over 2 years of age (43). Other pancreatic proteases complete the action of trypsin, such as chymotrypsin, elastases and carboxypeptidases, the latter one being the only exopeptidases among the pancreatic proteases. The intestinal protein hydrolysis is completed by the action of the brush border peptidases. The addition of brush border peptidases to pancreatic proteases in an in vitro digestion model raised the degree of proteolysis from 57 to 74% (44). At the end of the intestinal proteolysis, the luminal nitrogen is composed of small peptides of two to three AA residues (~60 to 70%), free AAs (~25 to 30%), and a small fraction consisting of large peptides and undigested proteins (45). The undigested or unabsorbable nitrogen fraction is then available for fermentation by the colonic microbiota. Fermented nitrogen can either be reused by the bacteria, excreted in the feces or, to a lesser extent, absorbed by the host in the form of ammonia and free AAs (46, 47).
3.1.2 Absorption of proteolysis products
A large proportion of peptides is absorbed in the jejunum, while free AAs are assumed to be mainly absorbed in the ileum (48). Compared with the other two segments, the absorption of free AAs and their derivatives from bacterial fermentation in the colon is low. The complex structure of the intestinal epithelium, whose development begins in the fetus and ends shortly after birth, greatly increases the exchange surface area. Before entering the portal vein, di- and tripeptides may undergo a final stage of proteolysis in the cytoplasm of brush-border enterocytes. Free AAs are absorbed using several transport systems present in the apical and/or basolateral membranes of enterocytes: active (Na+-dependent) transporters and facilitated transport by simple diffusion across the membrane, while di- and tri-peptides are absorbed by a H + -dependent transporter (PepT1) (49). The absorption of AAs and peptides is developmentally regulated and influenced by diet, hormones and growth factors. The transport systems are present at birth, and the transport rates of peptides and most AAs tend to decrease with age (from birth to weaning), although the extent of these changes varies widely among individuals (43). Studies suggested that the high dietary levels of protein or AA lead to the transcriptional activation of the PepT1 gene, resulting in the upregulation of peptide and AA transport (50–52).
3.1.3 Metabolic fate of dietary amino acids
From a quantitative perspective, the appearance of AAs in portal blood depends primarily on the composition and quantity of ingested proteins, as well as their digestibility. The main factors that determine the kinetics of AA appearance are the physicochemical nature of the proteins and how quickly they can transit and/or be digested through the digestive tract (53–55). A large proportion of AAs are used as they are, or can be transaminated for use as other AAs in protein synthesis. Another fraction enters specific metabolic pathways that convert AAs into non-protein nitrogen compounds. A final fraction can be catabolized to produce energy.
Free AAs first circulate through the splanchnic zone before reaching the peripheral circulation. In piglets, it has been reported that approximately 27% of total dietary nitrogen is retained in the splanchnic area following a meal (56). However, splanchnic extraction of AAs varies widely among AAs. For instance, up to 80% of dietary threonine and around 50% of dietary lysine are absorbed by the splanchnic area in piglets (57–59). Studies in adult humans have reported splanchnic extraction rates of up to 96% for glutamic acid, 69% for alanine, and 64% for glutamine. In contrast, the rates were much lower for arginine (38%), phenylalanine (29%) and leucine (21%), expressed as a proportion of enteral intake (60). Several parameters influence the metabolic fate of AAs in the splanchnic zone, such as the tissues with discrepancies between liver and gastrointestinal tissues, the tissue cell characteristics, and the molecular form of AAs (56). After extraction of AAs by the splanchnic zone, the circulating pool of AAs consists of dietary AAs that have escaped splanchnic extraction, AAs from transamination of dietary AAs in the liver, and AAs from proteolysis of endogenous proteins. These circulating AAs are metabolized by peripheral organs. In piglets, 42% of dietary nitrogen retention occurred in the peripheral zone [including 31% in the muscle and 6% in the skin (56)].
As a growing organism, the neonate has a positive nitrogen balance (anabolic predominance), with protein losses inversely related to gestational age (61). However, in 2008, Kalhan and Bier (62) reported that the protein losses expressed as a function of metabolic weight (weight0.75) are not significantly different from those of adults, whereas they are significantly higher when related to body weight. In seven-day-old piglets, the rate of protein accretion was very high and dependent on enteral intake (63). Focusing on specific AAs, the rate of glutamine and phenylalanine appearance in circulating blood, expressed on a body weight basis, was higher in infants than in healthy adults, reflecting higher energy expenditure in the infant (62, 64). It was also demonstrated that a negative relationship between glutamine turnover and the irreversible oxidation of protein (urea synthesis) existed, thus suggesting that glutamine has an important role as a nitrogen source for other synthetic processes and accretion of body proteins in the newborn (64). Finally, tryptophan is an AA of importance for infants because it is a limiting AA in food, being present in low concentration in IFs. Tryptophan released after protein hydrolysis is mainly used for protein biosynthesis. The remaining fraction of free tryptophan is metabolized by the host in the intestine, liver and brain, or by the colonic microbiota (65). The kynurenine pathway in liver is the most significant degradation pathway of the tryptophan unused for protein synthesis (>90% of tryptophan in excess available) (66). It is metabolized to quinolinic acid, which in turn is converted to niacin (vitamin B3). Tryptophan is important for infants because it is also converted in the pineal gland to the neurotransmitters, serotonin and melatonin, which play an important role in sleep regulation. Interestingly, the transport of tryptophan across the blood–brain barrier is in competition with the transport of other large neutral AAs (LNAAs), namely histidine, isoleucine, leucine, methionine, phenylalanine, tyrosine and valine. As a result, a strong correlation between brain tryptophan concentration, brain serotonin concentration and plasma tryptophan/LNAA ratio has been demonstrated (67–69).
3.2 Impact of protein composition and structure on digestion of HM vs. IF
HM is a dynamic fluid that serves as the biological standard for infant nutrition, providing the essential nutrients and thousands of bioactive molecules that play critical roles in protecting against infection and inflammation, contributing to immune maturation, supporting organ development, and promoting healthy microbial colonization. Unlike IF, which targets 0- to 6-month-old infants, HM composition adapts during lactation to meet the specific needs of the developing infant, which vary by stage of lactation, and between term and preterm infants (70).
HM serves as the reference for IF formulation. IFs require numerous ingredients (up to fifty for some IFs) and production steps involving several heat treatments, either on the raw material during ingredient manufacturing or during IF production (3). The formulation and processing route of IFs can differ between industrials, contributing to variability of IFs in their protein composition and potential ingredient interactions. This can affect digestion kinetics and physiological effects (71, 72). With respect to proteins, several factors such as phosphorylation, size, charge, tertiary structure, AA content and glycosylation have been identified as influencing protein degradation (131). Casein and WPs, both in HM and IFs, differ chemically and structurally, influencing their gastric behavior and digestive sensitivity. Interestingly, the ratio of casein to WPs influences gastric emptying and the degree of proteolysis (55, 73). Gastric emptying in infants aged three to 12 months was faster after the ingestion of HM or WP-dominant IF than casein-dominant IF (158), as casein coagulates near its isoelectric point (4.6). In contrast, unaggregated WPs remain soluble at the acidic pH of the infant stomach. In 2020, Halabi et al. (74) demonstrated that the addition of LF within IF induced partial casein micelle disintegration even before heating. Some studies have also explored the effect of casein mineralization and organization on gastric behavior and digestive kinetics (75–77), showing that the lower the mineralization, the faster the proteolysis. These results may explain the variability in gastric half-emptying time reported by different studies for IFs, as the mineralization levels of casein in IFs on the market is highly variable (4 to 12 mmol micellar Ca per 10 g casein (78);) and can also explain why the gastric emptying is slower for IF than for HM, due to the lower mineralisation level of HM caseins [~3.2 mmol micellar Ca per 10 g casein; (79)]. This can induce a different pattern of protein coagulation in the stomach, in addition to the different structure of human and bovine casein micelles (80–82) and the different size of fat droplets in HM and IF. Furthermore, there are different variants of β-casein (A1 and A2) in cow’s milk and IFs. Milk based on A2 β-casein is available on some markets. It has been reported that A2 β-casein cow’s milk has a different casein micelle size and different curd formation properties. This could potentially result in reduced gut discomfort related to milk (83). However, whether this remains true within the IF context requires further investigation.
Thermal treatments of proteins, such as those employed in ingredient and IF processing, are known to alter both protein structure and gastric behavior. Heat treatments increase the resistance of casein to gastric hydrolysis by forming casein/WP aggregates (84). A recent work also showed that the heat-induced denaturation of LF within IF significantly increased its susceptibility to hydrolysis (85). Changes in the surface properties of casein micelles, after WP binding, enhanced casein hydrolysis within IF (85) and increased the pH at which casein coagulates from ~4.9 to ~5.4 (86). The casein micelle organization was proved to be strongly dependent on the β-LG and LF contents in IF and on the heating temperature (74). The dairy protein ingredients used in IF formulations may be dry or liquid, depending on the manufacturer. A main factor influencing protein denaturation and aggregation during the initial processing of the liquid raw materials is the heat intensity (87–89). For dairy protein ingredients, the production process (serum, concentrate or isolate) appears to affect the rate of protein denaturation and aggregation, resulting in differences in digestibility (90).
Focusing on WPs, several studies have shown that β-LG in its native form remains intact in the stomach. Heat treatment can either accelerate its proteolysis in the intestine due to protein chain unfolding phenomenon (91–95), or reduce its digestibility by forming compact, aggregated WPs that hinder enzyme access to cleavage sites (96, 97). A study investigating the impact of heat treatment on proteins within IF revealed that IF containing α-La and LF preserved a higher proportion of native WPs than IF containing β-LG for high heat treatments (90°C/15 s, 75°C/15 min) (98). In its native form, LF has been shown to be resistant to pepsin, enabling it to exert its full bioactivity in the gut. However, the pasteurization of HM results in the partial gastric hydrolysis of LF, as demonstrated in vitro under infant digestive conditions (99, 100).
The Maillard reaction (glycation), favored by the heat treatment that occurs during IF production and storage due to the presence of primary amines and of lactose (a reducing sugar), can also affect protein digestibility directly or indirectly by sterically hindering the cleavage sites of digestive enzymes (90). In 2020, Zenker et al. (101) showed that high levels of glycation in IFs increase the size of peptides during digestion, indicating reduced proteolytic efficiency. Reduced glycation levels in IFs may enhance the gastrointestinal comfort of infants, as indicated by the clinical trial of Sheng et al. (102). However, the latter result is confounded by the different nature of the prebiotics administered in the various IFs. In addition, Maillard reaction products, such as carboxymethyl lysine, may also affect intestinal physiology (103, 104).
A recent in vitro and in vivo study on HM and IF (164) showed that the microstructure of the digesta differs between HM and IF, and that gastric proteolysis of α-La and casein is significantly lower for HM than for IF. In addition, it was demonstrated that the quality (structure and composition) of dairy protein ingredients within IF formulation significantly influenced the microstructure of the IFs. These differences were found to modulate proteolysis kinetics as well as the breakdown of the emulsion during the early gastric phase (126), highlighting the importance of considering the quality of protein ingredients in IF manufacturing. Whenever possible, IFs should be designed to closely resemble HM, including its digestive behavior (3, 105).
The true ileal digestibility (TID) of HM vs. IF was recently measured in Yucatan mini-piglets as a model for human infants (106). It was shown that the TID of total nitrogen was lower for HM than for IF due to the greater proportion of NPN in HM. NPN remains largely unabsorbed and is transferred to microbiota. As previously discussed in this review, this may have physiological relevance. Additionally, the TID of seven AAs differed significantly, particularly for lysine, phenylalanine, threonine, valine, alanine, proline and serine. The digestible indispensable AA score (DIAAS) of IF was lower than that of HM (83 vs. 101). Moughan et al. highlighted that the current FAO (2013) recommendations for AA requirements for infants may require revision, as the recommended AA concentrations were not corrected for hydrolysis time and digestibility. This resulted in lower values for leucine, lysine and threonine (more than 16% difference) and histidine and tryptophan (more than 30% difference).
3.3 Impact of protein composition and structure on absorption of dietary amino acids of HM vs. IF
The kinetics of AA appearance in plasma can be significantly impacted by several parameters, with the gastric emptying rate being the most important factor (55, 165). Previous studies on the casein fraction in IFs have shown that the degree of casein mineralization affects their supramolecular organization, which in turn influences their behavior in the stomach (76, 78). In other dairy matrices, it has been confirmed that the supramolecular organization of casein influences the gastric emptying rate and, subsequently, postprandial AA kinetics (77, 166). Another factor influencing the appearance of AA in the plasma is the rate of protein digestion, which in turn is modulated by protein structure. Specifically, the structure of WPs is easily modified in IFs, due to the numerous heat treatments. In vivo studies comparing postprandial AA concentrations in neonatal piglets fed native or denatured WP solutions have shown that higher plasma levels of essential AAs are observed within the first 60 min with native WP than with denatured WP (167). Similarly, in rodents fed diets containing 40% IF with either native or heat-denatured WPs (168), the consumption of native WPs resulted in higher plasma levels of total AAs than heat-denatured WPs.
A study in neonatal mini-piglets investigated how the composition and structure of protein ingredients within IFs affects plasma AA kinetics and concentrations (107). Although no difference in plasma AA kinetics was observed, both preprandial and postprandial AA concentrations in the plasma were modulated by the quality of the protein within the IFs. Interestingly, the whey origin within the IFs (either cheese or ideal whey) was found to modify AA homeostasis, resulting in increased plasma total and essential AA concentrations in piglets fed cheese whey IFs compared to ideal whey IFs. In line with previous studies (169), the importance of using cheese whey to increase plasma threonine concentration was emphasized, given that the cheese whey contains a significant amount of GMP, a threonine-rich peptide (107). Several other studies focusing on the casein/WP ratio in IFs found that an increased WP content in IFs was associated with higher plasma threonine concentrations, which were directly related to the AA profile of the predominant proteins (23, 170, 171). Several authors have suggested that increased threonine intake may be related to the limited ability of infants to eliminate threonine, given that it leads to higher plasma threonine levels (107, 169, 170, 172, 173). Interestingly, formula-fed term infants have been reported to have higher baseline and postprandial plasma concentrations of threonine, urea and valine, than breastfed term infants (174).
Several studies have attempted to compare the levels of AAs in the plasma of infants fed HM vs. formula. Most emphasized that the different protein profile in IFs compared to HM impacts the plasma concentration of AAs, as well as urea and other compounds, in preterm (169, 170, 172) and term (23, 170, 175–177) infants. However, only one study was conducted under presumed isonitrogenous conditions. This study showed that the postprandial plasma concentration of essential AAs was ~18% lower in HM-fed infants than in formula-fed infants. This suggests that the nature and structure of proteins may play a modulating role in AA absorption (178), in addition to the molecular form of nitrogen. It is likely that nitrogen was present in HM in lower proportions as true proteins than in IF.
Comparable data on HM and IF is lacking in this area due to differences in the design of the studies carried out, the nitrogen content of the matrices under study, the absence of a dietary adaptation period prior to sampling, the lack of sampling kinetics and imprecision regarding the timing of blood sampling in relation to the last meal. Furthermore, plasma AA concentrations, as measured in the systemic circulation, are influenced by first-pass extraction from splanchnic tissues, as well as by protein turnover and AA metabolism in the whole organism. These factors could differ between matrices (i.e., within IFs and HM).
4 Infant diet impact on microbiota, intestinal and brain development
Early nutrition plays an essential role in programming adult health and disease, including metabolic, cardiovascular, and immune diseases, as well as programming food preferences and eating behaviors (179, 180). The health benefits provided by HM are not yet fully realized with IF feeding. Therefore, improving the functional effects of IFs is an important goal in order to reduce the gap in terms of physiological and metabolic health between breastfed and formula-fed infants.
In the short term, HM has been associated with a lower risk of neonatal mortality. HM appears to be protective against sudden infant death syndrome as compared to IF (181). The reduced risk prevalence was a function of the breastfeeding rate, with a greater reduction for exclusive or predominant breastfeeding. Furthermore, there is convincing evidence that HM reduces the risk of developing diarrhea (0–5 years) (181–183) and acute otitis media (0–2 years) (183–185). In the long term, HM-fed infants may be at a lower risk (− 22%) of being overweight or obese in adulthood compared to formula-fed infant (4, 186–190). Furthermore, HM may prevent type II diabetes in adulthood compared to IF (181, 191–193).
The infant development initiated in utero continues from birth up to 2–3 years of age. Many changes occur during the early postnatal period, especially regarding intestinal development through morphological changes, functional maturation such as microbiota establishment (194, 195) and brain development (synaptogenesis and myelination) (196). Environmental factors such as infant nutrition (HM vs. IF) are essential determinants of the postnatal development, reported to modulate the gut microbiota (197–199), and thus the global microbiota-gut-brain axis (199, 200).
4.1 Microbiota, a key determinant of the infant health through its role on the gut brain-axis
Gut microbiota represents a complex bacterial ecosystem that colonizes the digestive tract. Itferments indigestible nutrients (such as HMOs or urea) and produces various metabolites including SCFAs (201). The early development of microbiota is under the influence of many parameters like the mode of delivery, the maternal genetic, the use of antibiotics and the environment as well as the maternal and infant nutrition (15, 180, 202, 203).
A lower fecal α-diversity was observed in HM-fed infants compared to formula-fed infants at 40 days of age, the difference being further reduced at 6 months of age (204–207). Indeed, gut microbiota abundance and evenness increased in HM-fed infants after 3 months of age unlike that in formula-fed infants for whom these indexes were already high at 40 days of age gut microbiota (207). A lower α-diversity was also measured in different intestinal segments (ileum, colon and rectum) in 3 weeks-old HM-fed piglets than in formula-fed piglets (198, 206), suggesting a specific role of HM components. Actinomycetota (formerly Actinobacteria) is the major phyla in gut microbiota of 3- and 6-month-old HM-fed and formula-fed infants, representing over 42 to 74% of the total phyla. The microbiota of HM-fed infants is characterized by a lower relative abundance of Bacillota (formerly Firmicutes) than that of formula-fed infants (205, 207), but Bacillota remains the second major phylum in 6-month-old HM-fed infants. Dissimilarities also exist at the genus level. Bifidobacterium (including Bifidobacterium spp., Bifidobacterium breve, Bifidobacterium infantis, Bifidobacterium longum) is frequently described as the dominant genus in the fecal content of 0- to 9-month-old HM-fed infants unlike that in formula-fed infants (203, 207–209), although some studies reported no difference (210, 211).
Besides HMOs, lipids and lactose (193, 212–214), it has been demonstrated that proteins and NPN are able to modulate the microbiota composition (15). Firstly, the impact of the nitrogenous fraction on microbiota shaping is frequently associated to proteins having immunomodulatory properties such as LF, lysozyme or Igs, but can also be due to other proteins and to NPN. Regarding immunological proteins, LF decreases the iron availability for bacteria due to its bacteriostatic property and is able to decrease E. coli, Pseudomonas aeruginosa and Candida albicans (215). Moreover, lysozyme can hydrolyze the peptidoglycan polymers found in the cell walls of bacteria, thereby lysing Gram-positive bacteria. However, it can also act synergistically with LF, contributing to the degradation of Gram-negative bacteria (216). Other studies have suggested that the undigested proteins from IF could also contribute, albeit to a lesser extent, to modulating the gut microbiota. Supplementating IF with α-La and GMP has been shown to increase the relative abundance of Clostridiaceae, Enterobacteriaceae and Streptococcus in preterm piglets after 19 days of feeding (217), whereas no difference was observed in six-month-old term infants (218).
Other nitrogenous compounds that could influence the microbiota include NPN, particularly urea, for which a role has been reported. Indeed, some bacteria possess urease genes, including certain Bifidobacterium species (e.g., B. longum subsp. Infantis); meaning that urea could act as a growth factor for these species (130, 219). Supplementation the IF with nucleotides was shown to decrease the ratio of Bacteroides-, Porphyromonas-, Prevotella- to Bifidobacterium-species in the fecal microbiota of 20-week-old infants (220).
4.2 Impact of infant diet on gut immune and barrier functions and brain development and their relationships with gut microbiota
Food in direct contact with the intestinal epithelium plays a recognized role in the maturation of the intestinal barrier, immune and endocrine functions. The structural and functional development of the gut depends on the type of the diet consumed (HM vs. IF). Intestinal growth is enhanced by bioactive components of HM, such as growth factors, absent or present in low amount in IF (194, 221).
It is acknowledged that the postnatal development of intestinal permeability in humans and porcine models follows a bell curve (222–224). During the first weeks of life, higher permeability is most commonly reported in HM-fed infants as compared to formula-fed infants (225), and in HM-fed and sow’s milk-fed piglets as compared to formula-fed piglets (199, 226). This has been associated with a reduced expression of genes involved in tight junction proteins in HM-fed piglets compared to formula-fed ones (199, 227). However, conflicting data have been observed in animal models, such as no change in jejunal or ileal permeability and/or increased expression of tight junction proteins in sow’s milk- vs. formula-fed piglets (206, 226, 228), and no change in the lactulose/mannitol ratio in breastfed infants (229–231). Such discrepancies may be related to the specificity of epithelial permeability in the intestinal segments studied in animal models, which cannot easily be recapitulated by an overall measurement such as the lactulose/mannitol ratio in infants. High intestinal permeability can lead to an increased passage of molecules across the epithelium, thereby promoting immune system development and tolerance to commensal bacteria and dietary antigens (226). Accordingly, HM-induced enhancement of the mucosal immune system has been reported in breastfed infants (232–234) and in piglets fed only HM (199). Fecal calprotectin content, a valuable marker of the state of intestinal mucosal inflammatory infiltration, was higher in HM-fed infants compared to formula-fed infants during the first weeks of life (225, 231, 235–237).
Several immunological factors, such as LF and IGs (IgG, IgA, IgM), which are more abundant in HM than in IF (38), anti-inflammatory cytokines (such as TGF-β and IL-10), pro-inflammatory cytokines [IL-1b, IL-6, IL-8, IL-12, TNF-α, IFN-γ; (15)] and other minor proteins present in HM but not in IF (17), are likely to contribute to the immune system boost in addition to dietary-induced changes in microbiota composition. The importance of the Bacillota phylum in inducing a pre-weaning peak in intestinal inflammatory markers has been demonstrated in rodents and piglets. Accordingly, our study comparing HM-fed and IF-fed piglets supports the relationship between microbiota and the mucosal immune system maturation. Several significant positive correlations were observed between Anaerovibrio, Mitsuokella and Veillonella genera belonging to Veillonellaceae family (Bacillota phylum) and genes encoding anti- and pro-inflammatory cytokines (IL-10, IL-10Ra, SOCS3, CCL2, IL-1bR, IL-8, TNF-α) and cellular signaling (ICAM1, MYD88) (199). This boost of the mucosal immune system has been shown to be essential for both immune ontogeny and regulation of susceptibility to immunopathologies later in life (199, 238). Moreover, in infants, positive correlations between fecal calprotectin excretion and colonization by some taxa of the Bacillota phylum support the role of bacteria in maturation of the intestinal immune system (239). The quality of the protein ingredients in IFs has also been reported to moderately affect the gut physiology and microbiota of three-week-old mini-piglets used as a model of human infants (115). This suggests that the quality of WP and casein (structure and composition) may mediate some of the physiological properties of IFs.
Similar to the gut, brain development begins in utero and continues throughout the first years of life. The postnatal period is crucial for the development of the central nervous system (240). While some steps in the development of neurons that begin at birth, such as visual and auditory processing, are rapidly developed, others, such as synaptogenesis and synaptic refinement, take longer to be fully developed (240, 241). These processes occur from birth to 3 years of age. It has been established that HM and IF have different effects on brain development and function. A large observational study (>17,000 healthy infants) has provided strong evidence that HM is more beneficial for optimal infant neurodevelopment than IF (242). It has also been demonstrated that HM is associated with higher myelination than IF at 2 years of age. Better myelination resulted in superior language and motor function in HM-fed infants, which is consistent with the superior cognitive performance observed in HM-fed infants compared to IF-fed infants during the first 6 months of life (243–245). Later in life, better cognitive function was observed in 10-19-year-olds who were breastfed as infants, compared to those who were formula-fed during infancy (246). However, several other environmental factors may affect child development. Supporting these clinical observations in infants, HM and IF diets were found to induce different genes expression profiles related to blood–brain barrier, endocrine and immune functions, neurosynaptogenesis, and metabolite levels in various brain regions, particularly the hypothalamus and hippocampus, in piglets (199).
The difference in brain development profiles between infants fed HM and IF can mainly be explained by differences in food composition related to components of the milk fat globule membrane (MFGM), such as polar lipids (244, 247) and to polyunsaturated fatty acids (248–251). Furthermore, a recent study in piglets demonstrated that the composition and structure of WPs and caseins did not affect the expression of several genes associated with hypothalamic development (115).
5 Conclusion
HM is a biofluid that provides the necessary nutrients to support infant growth. It contains many components that have been shown to affect metabolism, gut physiology, and the development of infant gut microbiota. Proteins are among the most important of these components for optimal growth. When formulating IF, the quality of the protein ingredient must be considered, although difficulties remain due to the different protein profiles of bovine and human milk. The nutritional value of milk from other mammals requires further investigation. There is a lack of data on the causal relationship between the protein composition and quality of IF and the plasma AA profile, highlighting the need for convincing studies. Furthermore, the quantity of HM compounds that reach the colon, and the impact of partially digested proteins (e.g., HM-derived peptides) on the composition of the microbiota, must be investigated to fully comprehend the role of the HM nitrogenous fraction in shaping the infant gut microbiota. In particular, the NPN fraction may be important, despite not yet being considered in current IF formulations.
Author contributions
LC: Writing – review & editing, Writing – original draft, Investigation. EC: Investigation, Writing – review & editing, Writing – original draft. ML: Writing – review & editing, Writing – original draft. IH-L: Funding acquisition, Writing – review & editing, Supervision, Writing – original draft, Validation, Conceptualization, Project administration. AD: Supervision, Conceptualization, Validation, Writing – review & editing, Funding acquisition, Writing – original draft, Project administration.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. WHO, UNICEF . Global breastfeeding scorecard 2023. Rates of breastfeeding increase around the world through improved protection and support. Geneva: WHO (2023).
2. European Union . Commission delegated regulation (EU) 2016/127. (2016). Available online at: http://data.europa.eu/eli/reg_del/2016/127/oj/eng (Accessed March 24, 2021).
3. Deglaire, A, Lee, J, Lanotte, L, Croguennec, T, Le Floch-Fouéré, C, Jeantet, R, et al. Towards more biomimetic and sustainable infant formula: challenges and future opportunities. Trends Food Sci Technol. (2023) 137:109–23. doi: 10.1016/j.tifs.2023.05.010
4. Koletzko, B, von Kries, R, Closa, R, Escribano, J, Scaglioni, S, Giovannini, M, et al. Lower protein in infant formula is associated with lower weight up to age 2 y: a randomized clinical trial. Am J Clin Nutr. (2009) 89:1836–45. doi: 10.3945/ajcn.2008.27091
5. Totzauer, M, Luque, V, Escribano, J, Closa-Monasterolo, R, Verduci, E, ReDionigi, A, et al. Effect of lower versus higher protein content in infant formula through the first year on body composition from 1 to 6 years: follow-up of a randomized clinical trial. Obesity (Silver Spring). (2018) 26:1203–10. doi: 10.1002/oby.22203
6. Haschke, F, Haiden, N, and Thakkar, SK. Nutritive and bioactive proteins in breastmilk. Ann Nutr Metab. (2016) 69:16–26. doi: 10.1159/000452820
7. Lönnerdal, B . Human milk proteins In: LK Pickering, AL Morrow, GM Ruiz-Palacios, and RJ Schanler, editors. Protecting infants through human Milk: Advances in Experimental Medicine and Biology. Boston, MA: Springer US (2004). 11–25.
8. Li, T, Zheng, Q, Xu, J, Li, Y, Zhang, M, Zhang, B, et al. Comparison of 11 formulas and breastfeeding for atopic dermatitis and growth in pediatric cow’s Milk protein allergy: a systematic review and network Meta-analysis of 23 randomized controlled trials. J Evid Based Med. (2025) 18:e70026. doi: 10.1111/jebm.70026
9. Gonzalez, PM, Cassin, AM, Durban, R, and Upton, JEM. Effects of food processing on Allergenicity. Curr Allergy Asthma Rep. (2025) 25:191. doi: 10.1007/s11882-024-01191-5
10. Jenness, R . Lactational performance of various mammalian species. J Dairy Sci. (1986) 69:869–85. doi: 10.3168/jds.S0022-0302(86)80478-7
11. Lönnerdal, B . Nutritional and physiologic significance of human milk proteins. Am J Clin Nutr. (2003) 77:1537S–43S. doi: 10.1093/ajcn/77.6.1537S
12. Cheftel, JC, and Lorient, D. Les propriétés fonctionnelles des protéines laitières et leur amélioration. Lait. (1982) 62:435–83. doi: 10.1051/lait:1982617-62029
13. Léonil, J, Michalski, MC, and Martin, P. Les structures supramoléculaires du lait: structure et impact nutritionnel de la micelle de caséine et du globule gras. INRAE Prod Anim. (2013) 26:129–44. doi: 10.20870/productions-animales.2013.26.2.3142
14. Heine, WE, Klein, PD, and Reeds, PJ. The importance of α-Lactalbumin in infant nutrition. J Nutr. (1991) 121:277–83. doi: 10.1093/jn/121.3.277
15. Boudry, G, Charton, E, Huerou-Luron, I, Ferret-Bernard, S, Le Gall, S, Even, S, et al. The relationship between breast Milk components and the infant gut microbiota. Front Nutr. (2021) 8:629740. doi: 10.3389/fnut.2021.629740
16. Heine, WE . The significance of tryptophan in infant nutrition In: G Huether, W Kochen, TJ Simat, and H Steinhart, editors. Tryptophan, serotonin, and melatonin: Basic aspects and applications. Boston, MA: Springer US (1999). 705–10.
17. Chatterton, DEW, Rasmussen, JT, Heegaard, CW, Sørensen, ES, and Petersen, TE. In vitro digestion of novel milk protein ingredients for use in infant formulas: research on biological functions. Trends Food Sci Technol. (2004) 15:373–83. doi: 10.1016/j.tifs.2003.12.004
18. Nasirpour, A, Scher, J, and Desobry, S. Baby foods: formulations and interactions (a review). Crit Rev Food Sci Nutr. (2006) 46:665–81. doi: 10.1080/10408390500511896
19. Davis, AM, Harris, BJ, Lien, EL, Pramuk, K, and Trabulsi, J. Alpha-lactalbumin-rich infant formula fed to healthy term infants in a multicenter study: plasma essential amino acids and gastrointestinal tolerance. Eur J Clin Nutr. (2008) 62:1294–301. doi: 10.1038/sj.ejcn.1602848
20. Heine, W, Radke, M, Wutzke, KD, Peters, E, and Kundt, G. Alpha-Lactalbumin-enriched low-protein infant formulas: a comparison to breast milk feeding. Acta Paediatr. (1996) 85:1024–8. doi: 10.1111/j.1651-2227.1996.tb14210.x
21. Tinghäll Nilsson, U, Hernell, O, Lönnerdal, B, Hartvigsen, ML, Jacobsen, LN, Kvistgaard, AS, et al. Low-protein formulas with alpha-Lactalbumin-enriched or Glycomacropeptide-reduced whey: effects on growth, nutrient intake and protein metabolism during early infancy: a randomized, double-blinded controlled trial. Nutrients. (2023) 15:1010. doi: 10.3390/nu15041010
22. Kelleher, SL, Chatterton, D, Nielsen, K, and Lönnerdal, B. Glycomacropeptide and alpha-lactalbumin supplementation of infant formula affects growth and nutritional status in infant rhesus monkeys. Am J Clin Nutr. (2003) 77:1261–8. doi: 10.1093/ajcn/77.5.1261
23. Sandström, O, Lönnerdal, B, Graverholt, G, and Hernell, O. Effects of alpha-lactalbumin-enriched formula containing different concentrations of glycomacropeptide on infant nutrition. Am J Clin Nutr. (2008) 87:921–8. doi: 10.1093/ajcn/87.4.921
24. O’Callaghan, DM, O’Mahony, J, Ramanujam, KS, and Burgher, AM. Dehydrated dairy products infant formulae In: DM O’Callaghan , editor. Encyclopedia of dairy sciences. San Diego: Academic Press (2011). 135–45.
25. Almeida, C, Mendonça Pereira, B, Leandro, K, Costa, M, Spisso, B, and Conte-Junior, C. Bioactive compounds in infant formula and their effects on infant nutrition and health: a systematic literature review. Int J Food Sci. (2021) 2021:1–31. doi: 10.1155/2021/8850080
26. D’Auria, E, Mameli, C, Piras, C, Cococcioni, L, Urbani, A, Zuccotti, GV, et al. Precision medicine in cow’s milk allergy: proteomics perspectives from allergens to patients. J Proteome. (2018) 188:173–80. doi: 10.1016/j.jprot.2018.01.018
27. Boutin, A, Liabeuf, V, Agabriel, C, Cleach, I, and Vitte, J. Profil de sensibilisation moléculaire aux protéines du lait de vache: évolution de 0 à 16 ans. Rev Fr Allergol. (2015) 55:219. doi: 10.1016/j.reval.2015.02.022
28. Brock, J . Lactoferrin: a multifunctional immunoregulatory protein? Immunol Today. (1995) 16:417–9. doi: 10.1016/0167-5699(95)80016-6
29. Håversen, LA, Engberg, I, Baltzer, L, Dolphin, G, Hanson, LÅ, and Mattsby-Baltzer, I. Human Lactoferrin and peptides derived from a surface-exposed helical region reduce experimental Escherichia coli urinary tract infection in mice. Infect Immun. (2000) 68:5816–23. doi: 10.1128/IAI.68.10.5816-5823.2000
30. Elass, E, Masson, M, Mazurier, J, and Legrand, D. Lactoferrin inhibits the lipopolysaccharide-induced expression and proteoglycan-binding ability of Interleukin-8 in human endothelial cells. Infect Immun. (2002) 70:1860–6. doi: 10.1128/IAI.70.4.1860-1866.2002
31. Lönnerdal, B . Nutritional roles of lactoferrin. Curr Opin Clin Nutr Metab Care. (2009) 12:293–7. doi: 10.1097/MCO.0b013e328328d13e
32. Karav, S, Le Parc, A, Nobrega, L, de Moura Bell, JM, Frese, SA, Kirmiz, N, et al. Oligosaccharides released from Milk glycoproteins are selective growth substrates for infant-associated Bifidobacteria. Appl Environ Microbiol. (2016) 82:3622–30. doi: 10.1128/AEM.00547-16
33. Roberts, AK, Chierici, R, Sawatzki, G, Hill, MJ, Volpato, S, and Vigi, V. Supplementation of an adapted formula with bovine lactoferrin: 1. Effect on the infant faecal flora. Acta Paediatr. (1992) 81:119–24. doi: 10.1111/j.1651-2227.1992.tb12186.x
34. King, JC, Cummings, GE, Guo, N, Trivedi, L, Readmond, BX, Keane, V, et al. A double-blind, placebo-controlled, pilot study of bovine lactoferrin supplementation in bottle-fed infants. J Pediatr Gastroenterol Nutr. (2007) 44:245–51. doi: 10.1097/01.mpg.0000243435.54958.68
35. Jiang, R, Du, X, and Lönnerdal, B. Effects of different sources of lactoferrin on cytokine response to SARS-COV-2, respiratory syncytial virus, and rotavirus infection in vitro. Biochem Cell Biol. (2025) 103:1–12. doi: 10.1139/bcb-2024-0146
36. Parrón, JA, Ripollés, D, Ramos, SJ, Pérez, MD, Semen, Z, Rubio, P, et al. Antirotaviral potential of lactoferrin from different origin: effect of thermal and high pressure treatments. Biometals. (2018) 31:343–55. doi: 10.1007/s10534-018-0088-4
37. Goedhart, AC, and Bindels, JG. The composition of human milk as a model for the design of infant formulas: recent findings and possible applications. Nutr Res Rev. (1994) 7:1–23. doi: 10.1079/NRR19940004
38. Su, M-Y, Broadhurst, M, Liu, C-P, Gathercole, J, Cheng, W-L, Qi, X-Y, et al. Comparative analysis of human milk and infant formula derived peptides following in vitro digestion. Food Chem. (2017) 221:1895–903. doi: 10.1016/j.foodchem.2016.10.041
39. Mainer, G, Sánchez, L, Ena, JM, and Calvo, M. Kinetic and thermodynamic parameters for heat denaturation of bovine Milk IgG, IgA and IgM. J Food Sci. (1997) 62:1034–8. doi: 10.1111/j.1365-2621.1997.tb15032.x
40. Müller, K, Toll, LJ, Manzardo, OA, Baumgartner, J, Nickel, E, Wenzel, F, et al. Holder pasteurization: comparison of water-Bath and dry-tempering devices. Front Pediatr. (2022) 10:853. doi: 10.3389/fped.2022.879853
41. Karamoko, G, Anihouvi, P, and Blecker, C. Évolution des connaissances sur les fonctionnalités de la fraction protéose-peptone du lait: propriétés techno-fonctionnelles et biologiques (synthèse bibliographique). Biotechnology, Agronomy, Society and Environment. (2013). Available online at: https://popups.uliege.be:443/1780-4507/index.php?id=9976 (Accessed June 8, 2020).
42. Bezkorovainy, A, Nichols, JH, and Sly, DA. Proteose-peptone fractions of human and bovine milk. Int J Biochem. (1976) 7:639–42. doi: 10.1016/0020-711X(76)90105-1
43. Lebenthal, E, and Lee, PC. Development of functional responses in human exocrine pancreas. Pediatrics. (1980) 66:556–60.
44. Picariello, G, Miralles, B, Mamone, G, Sánchez-Rivera, L, Recio, I, Addeo, F, et al. Role of intestinal brush border peptidases in the simulated digestion of milk proteins. Mol Nutr Food Res. (2015) 59:948–56. doi: 10.1002/mnfr.201400856
45. Gray, GM, and Cooper, HL. Protein digestion and absorption. Gastroenterology. (1971) 61:535–44. doi: 10.1016/S0016-5085(19)33506-1
46. Huizenga, JR, Gips, CH, and Tangerman, A. The contribution of various organs to ammonia formation: a review of factors determining the arterial ammonia concentration. Ann Clin Biochem. (1996) 33:23–30. doi: 10.1177/000456329603300103
47. Huneau, JF, Mahe, S, and Tomé, D. La digestion et l’absorption des protéines dans l’intestin In: JF Huneau , editor. Les protéines: le métabolisme et les besoins chez l’homme. Paris: IFN Institut Francais pour la Nutrition (1997). 7–22.
49. Boudry, G, David, ES, Douard, V, Monteiro, IM, Le Huerou-Luron, I, and Ferraris, RP. Role of intestinal transporters in neonatal nutrition: carbohydrates, proteins, lipids, minerals, and vitamins. J Pediatr Gastroenterol Nutr. (2010) 51:380–401. doi: 10.1097/MPG.0b013e3181eb5ad6
50. Shiraga, T, Miyamoto, K-I, Tanaka, H, Yamamoto, H, Taketani, Y, Morita, K, et al. Cellular and molecular mechanisms of dietary regulation on rat intestinal H+/peptide transporter PepT1. Gastroenterology. (1999) 116:354–62. doi: 10.1016/S0016-5085(99)70132-0
51. Chen, H, Pan, Y, Wong, EA, and Webb, KE. Dietary protein level and stage of development affect expression of an intestinal peptide transporter (cPepT1) in chickens1. J Nutr. (2005) 135:193–8. doi: 10.1093/jn/135.2.193
52. Sitrin, M . Digestion and absorption of carbohydrates and proteins In: PS Leung , editor. The Gastrointestinal System: Gastrointestinal, Nutritional and Hepatobiliary Physiology. Berlin: Springer Science and Business (2014). 137–58.
53. Galibois, I, Simoes Nunes, C, Rérat, A, and Savoie, L. Net appearance of amino acids in portal blood during the digestion of casein or rapeseed proteins in the pig. Can J Physiol Pharmacol. (1989) 67:1409–17. doi: 10.1139/y89-226
54. Rérat, A . Nutritional supply of proteins and absorption of their hydrolysis products: consequences on metabolism. Proc Nutr Soc. (1993) 52:335–44. doi: 10.1079/PNS19930069
55. Boirie, Y, Dangin, M, Gachon, P, Vasson, MP, Maubois, JL, and Beaufrère, B. Slow and fast dietary proteins differently modulate postprandial protein accretion. Proc Natl Acad Sci U S A. (1997) 94:14930–5. doi: 10.1073/pnas.94.26.14930
56. Bos, C, Stoll, B, Fouillet, H, Gaudichon, C, Guan, X, Grusak, MA, et al. Postprandial intestinal and whole body nitrogen kinetics and distribution in piglets fed a single meal. Am J Physiol Endocrinol Metab. (2005) 288:E436–46. doi: 10.1152/ajpendo.00263.2004
57. van der Schoor, SR, Reeds, PJ, Stellaard, F, Wattimena, JD, Sauer, PJ, Büller, HA, et al. Lysine kinetics in preterm infants: the importance of enteral feeding. Gut. (2004) 53:38–43. doi: 10.1136/gut.53.1.38
58. Schaart, MW, Schierbeek, H, van der Schoor, SRD, Stoll, B, Burrin, DG, Reeds, PJ, et al. Threonine utilization is high in the intestine of piglets. J Nutr. (2005) 135:765–70. doi: 10.1093/jn/135.4.765
59. Hamard, A, Sève, B, and Le Floc’h, N. A moderate threonine deficiency differently affects protein metabolism in tissues of early-weaned piglets. Comp Biochem Physiol A Mol Integr Physiol. (2009) 152:491–7. doi: 10.1016/j.cbpa.2008.12.002
60. Stoll, B, and Burrin, DG. Measuring splanchnic amino acid metabolism in vivo using stable isotopic tracers. J Anim Sci. (2006) 84:E60–72. doi: 10.2527/2006.8413_supple60x
61. Denne, SC, and Poindexter, BB. Differences between metabolism and feeding of preterm and term infants In: PJ Thureen and WW Hay, editors. Neonatal nutrition and metabolism. Cambridge: Cambridge University Press (2006). 437–44.
62. Kalhan, SC, and Bier, DM. Protein and amino acid metabolism in the human newborn. Annu Rev Nutr. (2008) 28:389–410. doi: 10.1146/annurev.nutr.28.061807.155333
63. Stoll, B, Chang, X, Fan, MZ, Reeds, PJ, and Burrin, DG. Enteral nutrient intake level determines intestinal protein synthesis and accretion rates in neonatal pigs. Am J Physiol Gastrointest Liver Physiol. (2000) 279:G288–94. doi: 10.1152/ajpgi.2000.279.2.G288
64. Parimi, PS, Devapatla, S, Gruca, L, O’Brien, AM, Hanson, RW, and Kalhan, SC. Glutamine and leucine nitrogen kinetics and their relation to urea nitrogen in newborn infants. Am J Physiol Endocrinol Metab. (2002) 282:E618–25. doi: 10.1152/ajpendo.00403.2001
65. Gostner, JM, Geisler, S, Stonig, M, Mair, L, Sperner-Unterweger, B, and Fuchs, D. Tryptophan metabolism and related pathways in psychoneuroimmunology: the impact of nutrition and lifestyle. Neuropsychobiology. (2020) 79:89–99. doi: 10.1159/000496293
66. O’Mahony, SM, Clarke, G, Borre, YE, Dinan, TG, and Cryan, JF. Serotonin, tryptophan metabolism and the brain-gut-microbiome axis. Behav Brain Res. (2015) 277:32–48. doi: 10.1016/j.bbr.2014.07.027
67. Ruddick, JP, Evans, AK, Nutt, DJ, Lightman, SL, Rook, GAW, and Lowry, CA. Tryptophan metabolism in the central nervous system: medical implications. Expert Rev Mol Med. (2006) 8:1–27. doi: 10.1017/S1462399406000068
68. Le Floc’h, N, Otten, W, and Merlot, E. Tryptophan metabolism, from nutrition to potential therapeutic applications. Amino Acids. (2011) 41:1195–205. doi: 10.1007/s00726-010-0752-7
69. Fernstrom, JD . Large neutral amino acids: dietary effects on brain neurochemistry and function. Amino Acids. (2013) 45:419–30. doi: 10.1007/s00726-012-1330-y
70. Ballard, O, and Morrow, AL. Human Milk composition: nutrients and bioactive factors. Pediatr Clin N Am. (2013) 60:49–74. doi: 10.1016/j.pcl.2012.10.002
71. Macierzanka, A, Sancho, AI, Mills, ENC, Rigby, NM, and Mackie, AR. Emulsification alters simulated gastrointestinal proteolysis of β-casein and β-lactoglobulin. Soft Matter. (2009) 5:538–50. doi: 10.1039/B811233A
72. Zhang, Q, Cundiff, JK, Maria, SD, McMahon, RJ, Wickham, MSJ, Faulks, RM, et al. Differential digestion of human milk proteins in a simulated stomach model. J Proteome Res. (2014) 13:1055–64. doi: 10.1021/pr401051u
73. Meyer, R, Foong, R-X, Thapar, N, Kritas, S, and Shah, N. Systematic review of the impact of feed protein type and degree of hydrolysis on gastric emptying in children. BMC Gastroenterol. (2015) 15:137. doi: 10.1186/s12876-015-0369-0
74. Halabi, A, Deglaire, A, Hennetier, M, Violleau, F, Burel, A, Bouhallab, S, et al. Structural characterization of heat-induced protein aggregates in model infant milk formulas. Food Hydrocoll. (2020) 107:105928. doi: 10.1016/j.foodhyd.2020.105928
75. Wang, X, Ye, A, Lin, Q, Han, J, and Singh, H. Gastric digestion of milk protein ingredients: Study using an in vitro dynamic model. J Dairy Sci. (2018) 101:6842–52. doi: 10.3168/jds.2017-14284
76. Huppertz, T, and Lambers, TT. Influence of micellar calcium phosphate on in vitro gastric coagulation and digestion of milk proteins in infant formula model systems. Int Dairy J. (2020) 107:104717. doi: 10.1016/j.idairyj.2020.104717
77. Boulier, A, Denis, S, Henry, G, Guérin, S, Alric, M, Meunier, N, et al. Casein structures differently affect postprandial amino acid delivery through their intra-gastric clotting properties. Food Chem. (2023) 415:135779. doi: 10.1016/j.foodchem.2023.135779
78. Huppertz, T, and Timmer, C. Salt equilibria in nutritional formulae for infants and young children. Int Dairy J. (2020) 110:104805. doi: 10.1016/j.idairyj.2020.104805
79. Holt, C, and Jenness, R. Interrelationships of constituents and partition of salts in milk samples from eight species. Comp Biochem Physiol A Physiol. (1984) 77:275–82. doi: 10.1016/0300-9629(84)90060-4
80. Yang, T, Liu, D, Tang, J, Tao, X, Zhang, J, Liu, X, et al. Formation of casein micelles simulating human milk casein composition from bovine caseins: micellar structure and in vitro infant gastrointestinal digestion. Food Hydrocoll. (2024) 149:109610. doi: 10.1016/j.foodhyd.2023.109610
81. He, T, Rombouts, W, Einerhand, AWC, Hotrum, N, and Van De Velde, F. Gastric protein digestion of goat and cow milk infant formula and human milk under simulated infant conditions. Int J Food Sci Nutr. (2022) 73:28–38. doi: 10.1080/09637486.2021.1921705
82. Sood, SM, Herbert, PJ, and Slatter, CW. Structural studies on casein micelles of human Milk: dissociation of β-casein of different phosphorylation levels induced by cooling and Ethylenediaminetetraacetate. J Dairy Sci. (1997) 80:628–33. doi: 10.3168/jds.s0022-0302(97)75980-0
83. Giribaldi, M, Lamberti, C, Cirrincione, S, Giuffrida, MG, and Cavallarin, L. A2 Milk and BCM-7 peptide as emerging parameters of Milk quality. Front Nutr. (2022) 9:375. doi: 10.3389/fnut.2022.842375
84. Dupont, D, Boutrou, R, Menard, O, Jardin, J, Tanguy, G, Schuck, P, et al. Heat treatment of Milk during powder manufacture increases casein resistance to simulated infant digestion. Food Dig. (2010) 1:28–39. doi: 10.1007/s13228-010-0003-0
85. Halabi, A, Croguennec, T, Bouhallab, S, Dupont, D, and Deglaire, A. Modification of protein structures by altering the whey protein profile and heat treatment affects in vitro static digestion of model infant milk formulas. Food Funct. (2020) 11:6933–45. doi: 10.1039/D0FO01362E
86. Donato, L, and Guyomarc’H, F. Formation and properties of the whey protein/kappa-casein complexes in heated skim milk – a review. Dairy Sci Technol. (2009) 89:3–29. doi: 10.1051/dst:2008033
87. Singh, H, and Creamer, LK. Denaturation, aggregation and heat stability of milk protein during the manufacture of skim milk powder. J Dairy Res. (1991) 58:269–83. doi: 10.1017/S002202990002985X
88. Morgan, F, Léonil, J, Mollé, D, and Bouhallab, S. Modification of bovine β-Lactoglobulin by glycation in a powdered state or in an aqueous solution: effect on association behavior and protein conformation. J Agric Food Chem. (1999) 47:83–91. doi: 10.1021/jf9804387
89. Mehta, BM, and Deeth, HC. Blocked lysine in dairy products: formation, occurrence, analysis, and nutritional implications. Compr Rev Food Sci Food Saf. (2016) 15:206–18. doi: 10.1111/1541-4337.12178
90. van Lieshout, GA, Lambers, TT, Bragt, MC, and Hettinga, KA. How processing may affect milk protein digestion and overall physiological outcomes: a systematic review. Crit Rev Food Sci Nutr. (2019) 1:1–24. doi: 10.1080/10408398.2019.1646703
91. Guo, MR, Fox, PF, Flynn, A, and Kindstedt, PS. Susceptibility of β-Lactoglobulin and sodium Caseinate to proteolysis by pepsin and trypsin. J Dairy Sci. (1995) 78:2336–44. doi: 10.3168/jds.S0022-0302(95)76860-6
92. Kitabatake, N, and Kinekawa, Y-I. Digestibility of bovine Milk whey protein and β-Lactoglobulin in vitro and in vivo. J Agric Food Chem. (1998) 46:4917–23. doi: 10.1021/jf9710903
93. Lorieau, L, Halabi, A, Ligneul, A, Hazart, E, Dupont, D, and Floury, J. Impact of the dairy product structure and protein nature on the proteolysis and amino acid bioaccessiblity during in vitro digestion. Food Hydrocoll. (2018) 82:399–411. doi: 10.1016/j.foodhyd.2018.04.019
94. Rahaman, T, Vasiljevic, T, and Ramchandran, L. Shear, heat and pH induced conformational changes of wheat gluten – impact on antigenicity. Food Chem. (2016) 196:180–8. doi: 10.1016/j.foodchem.2015.09.041
95. Singh, TK, Øiseth, SK, Lundin, L, and Day, L. Influence of heat and shear induced protein aggregation on the in vitro digestion rate of whey proteins. Food Funct. (2014) 5:2686–98. doi: 10.1039/c4fo00454j
96. Lindberg, T, Engberg, S, Sjöberg, LB, and Lönnerdal, B. In vitro digestion of proteins in human milk fortifiers and in preterm formula. J Pediatr Gastroenterol Nutr. (1998) 27:30–6. doi: 10.1097/00005176-199807000-00006
97. Wada, Y, and Lönnerdal, B. Effects of different industrial heating processes of milk on site-specific protein modifications and their relationship to in vitro and in vivo digestibility. J Agric Food Chem. (2014) 62:4175–85. doi: 10.1021/jf501617s
98. Halabi, A, Deglaire, A, Hamon, P, Bouhallab, S, Dupont, D, and Croguennec, T. Kinetics of heat-induced denaturation of proteins in model infant milk formulas as a function of whey protein composition. Food Chem. (2020) 302:125296. doi: 10.1016/j.foodchem.2019.125296
99. de Oliveira, S, Deglaire, A, Ménard, O, Bellanger, A, Rousseau, F, Henry, G, et al. Holder pasteurization impacts the proteolysis, lipolysis and disintegration of human milk under in vitro dynamic term newborn digestion. Food Res Int. (2016) 88:263–75. doi: 10.1016/j.foodres.2015.11.022
100. Nebbia, S, Giribaldi, M, Cavallarin, L, Bertino, E, Coscia, A, Briard-Bion, V, et al. Differential impact of holder and high temperature short time pasteurization on the dynamic in vitro digestion of human milk in a preterm newborn model. Food Chem. (2020) 328:127126. doi: 10.1016/j.foodchem.2020.127126
101. Zenker, HE, van Lieshout, GAA, van Gool, MP, Bragt, MCE, and Hettinga, KA. Lysine blockage of milk proteins in infant formula impairs overall protein digestibility and peptide release. Food Funct. (2020) 11:358–69. doi: 10.1039/C9FO02097G
102. Sheng, XY, Buthmanaban, V, Van Lieshout, GAA, and Parikh, P. Reduced occurrence of gastrointestinal symptoms in Chinese infants fed minimally processed commercially available formula: a Cross-sectional observational Study. J Nutr Metab. (2020) 2020:1–10. doi: 10.1155/2020/1807397
103. Elmhiri, G, Mahmood, D, Niquet-Leridon, C, Jacolot, P, Firmin, S, Guigand, L, et al. Formula-derived advanced glycation end products are involved in the development of long-term inflammation and oxidative stress in kidney of IUGR piglets. Mol Nutr Food Res. (2015) 59:939–47. doi: 10.1002/mnfr.201400722
104. Baskara, I, Niquet-Leridon, C, Anton, P, and Delayre-Orthez, C. Neoformed compounds from the Maillard reaction in infant formulas: a new risk factor for allergy? EMJ Allergy Immunol. (2017) 2:87–93.
105. Bourlieu, C, Deglaire, A, Cassia de Oliveira, S, Ménard, O, Le Gouar, Y, Carrière, F, et al. Towards infant formula biomimetic of human milk structure and digestive behaviour. OCL. (2017) 24:D206. doi: 10.1051/ocl/2017010
106. Charton, E, Henry, G, Cahu, A, Le-Gouar, Y, Dahirel, P, Moughan, PJ, et al. Ileal digestibility of nitrogen and amino acids in human Milk and an infant formula as determined in neonatal Minipiglets. J Nutr. (2023) 153:1063–74. doi: 10.1016/j.tjnut.2023.02.025
107. Chauvet, L, Brunel, A, Gouar, Y, Guerin, S, Janvier, R, Henry, G, et al. Protein ingredient quality within infant formulas impacts plasma amino acid concentrations in neonatal minipiglets. J Nutr. (2024) 154:2029–41. doi: 10.1016/j.tjnut.2024.05.009
108. Thomä-Worringer, C, Sørensen, J, and López-Fandiño, R. Health effects and technological features of caseinomacropeptide. Int Dairy J. (2006) 16:1324–33. doi: 10.1016/j.idairyj.2006.06.012
109. Córdova-Dávalos, LE, Jiménez, M, and Salinas, E. Glycomacropeptide bioactivity and health: a review highlighting action mechanisms and signaling pathways. Nutrients. (2019) 11:598. doi: 10.3390/nu11030598
110. Royle, PJ, McIntosh, GH, and Clifton, PM. Whey protein isolate and glycomacropeptide decrease weight gain and alter body composition in male Wistar rats. Br J Nutr. (2008) 100:88–93. doi: 10.1017/S0007114507883000
111. Requena, P, González, R, López-Posadas, R, Abadía-Molina, A, Suárez, MD, Zarzuelo, A, et al. The intestinal antiinflammatory agent glycomacropeptide has immunomodulatory actions on rat splenocytes. Biochem Pharmacol. (2010) 79:1797–804. doi: 10.1016/j.bcp.2010.02.008
112. Chen, Q, Cao, J, Jia, Y, Liu, X, Yan, Y, and Pang, G. Modulation of mice fecal microbiota by administration of casein glycomacropeptide. Microbiol Res. (2012) 3:3–e3. doi: 10.4081/mr.2012.e3
113. Sawin, EA, De Wolfe, TJ, Aktas, B, Stroup, BM, Murali, SG, Steele, JL, et al. Glycomacropeptide is a prebiotic that reduces Desulfovibrio bacteria, increases cecal short-chain fatty acids, and is anti-inflammatory in mice. Am J Physiol Gastrointest Liver Physiol. (2015) 309:G590–601. doi: 10.1152/ajpgi.00211.2015
114. Ntemiri, A, Chonchúir, F, O’Callaghan, TF, Stanton, C, Ross, RP, and O’Toole, PW. Glycomacropeptide sustains microbiota diversity and promotes specific taxa in an artificial colon model of elderly gut microbiota. J Agric Food Chem. (2017) 65:1836–46. doi: 10.1021/acs.jafc.6b05434
115. Chauvet, L, Randuineau, G, Guérin, S, Cahu, A, Janvier, R, Dahirel, P, et al. The protein ingredient quality of infant formula influences colonic physiology and microbiota and the serum metabolomic profile in neonatal mini-piglets. Food Res Int. (2025) 200:115511. doi: 10.1016/j.foodres.2024.115511
116. Neelima, SR, Rajput, YS, and Mann, B. Chemical and functional properties of glycomacropeptide (GMP) and its role in the detection of cheese whey adulteration in milk: A review. Dairy Sci Technol. (2013) 93:21–43. doi: 10.1007/s13594-012-0095-0
117. Calapaj, GG . An electron microscope study of the ultrastructure of bovine and human casein micelles in fresh and acidified milk. J Dairy Res. (1968) 35:1–6. doi: 10.1017/S0022029900018719
118. Kunz, C, and Lönnerdal, B. Casein Micelles and Casein Subunits in Human Milk. Boca Raton, FL: CRC Press, pp. 9–27. (1989).
119. Hansen, M, Sandström, B, and Lönnerdal, B. The effect of casein phosphopeptides on zinc and calcium absorption from high phytate infant diets assessed in rat pups and Caco-2 cells. Pediatr Res. (1996) 40:547–52. doi: 10.1203/00006450-199610000-00006
120. Huang, J, Zhang, L, Lan, H, and Zhou, P. How to adjust α-lactalbumin and β-casein ratio in milk protein formula to give a similar digestion pattern to human milk? J Food Compos Anal. (2022) 110:104536. doi: 10.1016/j.jfca.2022.104536
121. Atkinson, S, and Lonnerdal, B. Proteins and non-protein nitrogen in human Milk. 1st ed. Boca Raton, FL: CRC Press (1989). 260 p.
122. Thurl, S, Munzert, M, Henker, J, Boehm, G, Müller-Werner, B, Jelinek, J, et al. Variation of human milk oligosaccharides in relation to milk groups and lactational periods. Br J Nutr. (2010) 104:1261–71. doi: 10.1017/S0007114510002072
123. Italianer, MF, Naninck, EFG, Roelants, JA, van der Horst, GTJ, Reiss, IKM, van Goudoever, JB, et al. Circadian variation in human milk composition, a systematic review. Nutrients. (2020) 12:2328. doi: 10.3390/nu12082328
124. Block, E, Depatie, C, Lefebvre, D, and Petitclerc, D. L’urée du lait: les sources de variation et les implications. Conseil des productions animales du Québec (1998).
125. Bendelja Ljoljić, D, Dolenčić Špehar, I, Prpić, Z, Vnučec, I, and Samaržija, D. Urea concentration in goat milk: importance of determination and factors of variability. J Cent Eur Agric. (2020) 21:707–21. doi: 10.5513/JCEA01/21.4.3058
126. Chauvet, L, Ménard, O, Le Gouar, Y, Henry, G, Jardin, J, Hennetier, M, et al. Protein ingredient quality of infant formulas impacts their structure and kinetics of proteolysis under in vitro dynamic digestion. Food Res Int. (2023) 169:112883. doi: 10.1016/j.foodres.2023.112883
127. Donovan, SM, and Lonnerdal, B. Non-Protein Nitrogen and True Protein in Infant Formulas. Acta Paediatr. (1989) 78:497–504. doi: 10.1111/j.1651-2227.1989.tb17927.x
128. Prosser, CG, Mclaren, RD, Frost, D, Agnew, M, and Lowry, DJ. Composition of the non-protein nitrogen fraction of goat whole milk powder and goat milk-based infant and follow-on formulae. Int J Food Sci Nutr. (2008) 59:123–33. doi: 10.1080/09637480701425585
129. Fomon, SJ, Matthews, DE, Bier, DM, Rogers, RR, Rebouche, CJ, Edwards, BB, et al. Bioavailability of dietary urea nitrogen in the infant. J Pediatr. (1987) 111:221–4. doi: 10.1016/s0022-3476(87)80071-9
130. Schimmel, P, Kleinjans, L, Bongers, RS, Knol, J, and Belzer, C. Breast milk urea as a nitrogen source for urease positive Bifidobacterium infantis. FEMS Microbiol Ecol. (2021) 97:fiab019. doi: 10.1093/femsec/fiab019
131. Dallas, DC, Underwood, MA, Zivkovic, AM, and German, JB. Digestion of protein in premature and term infants. J Nutr Disord Ther. (2012) 2:112. doi: 10.4172/2161-0509.1000112
132. Reeds, PJ, Burrin, DG, Stoll, B, and Jahoor, F. Intestinal glutamate metabolism. J Nutr. (2000) 130:978S–82S. doi: 10.1093/jn/130.4.978S
133. Militante, JD, and Lombardini, JB. Taurine: evidence of physiological function in the retina. Nutr Neurosci. (2002) 5:75–90. doi: 10.1080/10284150290018991
134. Dallas, DC, Sah, BN, Beverly, RL, You, X, Hilliard, MA, Sela, DA, et al. Nonprotein nitrogen and protein-derived peptides in human milk In: MK Mcguire and D O’connor, editors. Human Milk. lst ed. Cambridge, MA: Academic Press (2021). 299–336.
135. Grill, JP, Perrin, S, and Schneider, F. Bile salt toxicity to some bifidobacteria strains: role of conjugated bile salt hydrolase and pH. Can J Microbiol. (2000) 46:878–84. doi: 10.1139/w00-066
136. Noriega, L, Cuevas, I s, Margolles, A, and de los Reyes-Gavilán, CG. Deconjugation and bile salts hydrolase activity by Bifidobacterium strains with acquired resistance to bile. Int Dairy J. (2006) 16:850–5. doi: 10.1016/j.idairyj.2005.09.008
137. Wang, B, Yu, B, Karim, M, Hu, H, Sun, Y, McGreevy, P, et al. Dietary sialic acid supplementation improves learning and memory in piglets2. Am J Clin Nutr. (2007) 85:561–9. doi: 10.1093/ajcn/85.2.561
138. Wang, M, Monaco, MH, Hauser, J, Yan, J, Dilger, RN, and Donovan, SM. Bovine Milk oligosaccharides and human Milk oligosaccharides modulate the gut microbiota composition and volatile fatty acid concentrations in a preclinical neonatal model. Microorganisms. (2021) 9:884. doi: 10.3390/microorganisms9050884
139. Leeuwen, SS, Poele, EM, Chatziioannou, A, Benjamins, E, Haandrikman, A, and Dijkhuizen, L. Goat Milk oligosaccharides: their diversity, quantity, and functional properties in comparison to human Milk oligosaccharides. J Agric Food Chem. (2020) 68:13469–85. doi: 10.1021/acs.jafc.0c03766
140. Hambraeus, L . Proprietary milk versus human breast milk in infant feeding. A critical appraisal from the nutritional point of view. Pediatr Clin N Am. (1977) 24:17–36. doi: 10.1016/s0031-3955(16)33384-3
141. Svanberg, U, Gebre-Medhin, M, Ljungqvist, B, and Olsson, M. Breast milk composition in Ethiopian and Swedish mothers. III. Amino acids and other nitrogenous substances. Am J Clin Nutr. (1977) 30:499–507. doi: 10.1093/ajcn/30.4.499
142. Hord, N, Ghannam, J, Garg, H, Berens, P, and Bryan, N. Nitrate and nitrite content of human, formula, bovine, and soy milks: implications for dietary nitrite and nitrate recommendations. Breastfeed Med. (2011) 6:393–9. doi: 10.1089/bfm.2010.0070
143. Buts, JP, De Keyser, N, De Raedemaeker, L, Collette, E, and Sokal, EM. Polyamine profiles in human milk, infant artificial formulas, and semi-elemental diets. J Pediatr Gastroenterol Nutr. (1995) 21:44–9. doi: 10.1097/00005176-199507000-00007
144. Atiya Ali, M, Strandvik, B, Sabel, K-G, Palme Kilander, C, Strömberg, R, and Yngve, A. Polyamine levels in breast milk are associated with mothers’ dietary intake and are higher in preterm than full-term human milk and formulas. J Hum Nutr Diet. (2014) 27:459–67. doi: 10.1111/jhn.12156
145. Gómez-Gallego, C, Collado, MC, Ilo, T, Jaakkola, U-M, Bernal, MJ, Periago, MJ, et al. Infant formula supplemented with polyamines alters the intestinal microbiota in neonatal BALB/cOlaHsd mice. J Nutr Biochem. (2012) 23:1508–13. doi: 10.1016/j.jnutbio.2011.10.003
146. Gil, A, and Sanchez-Medina, F. Acid-soluble nucleotides of cow’s, goat’s and sheep’s milks, at different stages of lactation. J Dairy Res. (1981) 48:35–44. doi: 10.1017/s0022029900021427
147. Gil, A, and Sanchez-Medina, F. Acid-soluble nucleotides of human milk at different stages of lactation. J Dairy Res. (1982) 49:301–7. doi: 10.1017/s0022029900022391
148. Schlimme, E, Martin, D, and Meisel, H. Nucleosides and nucleotides: natural bioactive substances in milk and colostrum. Br J Nutr. (2000) 84:59–68. doi: 10.1017/s0007114500002269
149. Moughan, PJ, Deglaire, A, Yan, Y, Wescombe, P, Lim, WXJ, Stroebinger, N, et al. Amino acid requirements of the infant: the amino acid composition of human breast milk. Front Nutr. (2024) 11:565. doi: 10.3389/fnut.2024.1446565
150. Abrahamse, E, Minekus, M, van Aken, GA, van de Heijning, B, Knol, J, Bartke, N, et al. Development of the digestive system—experimental challenges and approaches of infant lipid digestion. Food Dig. (2012) 3:63–77. doi: 10.1007/s13228-012-0025-x
151. Paparo, L, di Costanzo, M, di Scala, C, Cosenza, L, Leone, L, Nocerino, R, et al. The influence of early life nutrition on epigenetic regulatory mechanisms of the immune system. Nutrients. (2014) 6:4706–19. doi: 10.3390/nu6114706
152. Bourlieu, C, Ménard, O, Bouzerzour, K, Mandalari, G, Macierzanka, A, Mackie, AR, et al. Specificity of infant digestive conditions: some clues for developing relevant in vitro models. Crit Rev Food Sci Nutr. (2014) 54:1427–57. doi: 10.1080/10408398.2011.640757
153. Hamosh, M . Digestion in the newborn. Clin Perinatol. (1996) 23:191–209. doi: 10.1016/S0095-5108(18)30238-0
154. Kelly, EJ, and Newell, SJ. Gastric ontogeny: clinical implications. Arch Dis Child Fetal Neonatal Ed. (1994) 71:F136–41. doi: 10.1136/fn.71.2.f136
155. De Oliveira, S . Impact de la pasteurisation et de l’homogénéisation sur la digestion du lait maternel chez le nouveau-né: Etudes in vitro et in vivo. [These de doctorat]. Rennes, Agrocampus Ouest. (2016). Available online at: https://www.theses.fr/2016NSARB284.
156. Piper, DW, and Fenton, BH. pH stability and activity curves of pepsin with special reference to their clinical importance. Gut. (1965) 6:506–8. doi: 10.1136/gut.6.5.506
157. Henderson, TR, Hamosh, M, Armand, M, Mehta, NR, and Hamosh, P. Gastric proteolysis in the preterm infant: protein digestion is limited and is not affected by diet, human Milk or formula † 580. Pediatr Res. (1998) 43:101–1. doi: 10.1203/00006450-199804001-00601
158. Billeaud, C, Guillet, J, and Sandler, B. Gastric emptying in infants with or without gastro-oesophageal reflux according to the type of milk. Eur J Clin Nutr. (1990) 44:577–83.
159. Cavell, B . Gastric emptying in infants fed human milk or infant formula. Acta Paediatr Scand. (1981) 70:639–41.
160. Le Roux, L, Ménard, O, Chacon, R, Dupont, D, Jeantet, R, Deglaire, A, et al. Are faba bean and pea proteins potential whey protein substitutes in infant formulas? An in vitro dynamic digestion approach. Foods. (2020) 9:362. doi: 10.3390/foods9030362
161. Siegel, M, Krantz, B, and Lebenthal, E. Effect of fat and carbohydrate composition on the gastric emptying of isocaloric feedings in premature infants. Gastroenterology. (1985) 89:785–90. doi: 10.1016/0016-5085(85)90573-6
162. Keller, PJ, and Allan, BJ. The protein composition of human pancreatic juice. J Biol Chem. (1967) 242:281–7. doi: 10.1016/S0021-9258(19)81461-8
163. McClean, P, and Weaver, LT. Ontogeny of human pancreatic exocrine function. Arch Dis Child. (1993) 68:62–5. doi: 10.1136/adc.68.1_Spec_No.62
164. Charton, E, Menard, O, Cochet, M-F, Le Gouar, Y, Jardin, J, Henry, G, et al. Human milk vs. infant formula digestive fate: in vitro dynamic digestion and in vivo mini-piglet models lead to similar conclusions. Food Res Int. (2024) 196:115070. doi: 10.1016/j.foodres.2024.115070
165. Gaudichon, C, Roos, N, Mahé, S, Sick, H, Bouley, C, and Tomé, D. Gastric emptying regulates the kinetics of nitrogen absorption from 15N-labeled milk and 15N-labeled yogurt in miniature pigs. J Nutr. (1994) 124:1970–7. doi: 10.1093/jn/124.10.1970
166. Deglaire, A, Fromentin, C, Fouillet, H, Airinei, G, Gaudichon, C, Boutry, C, et al. Hydrolyzed dietary casein as compared with the intact protein reduces postprandial peripheral, but not whole-body, uptake of nitrogen in humans. Am J Clin Nutr. (2009) 90:1011–22. doi: 10.3945/ajcn.2009.27548
167. Welch-Jernigan, RJ, Abrahamse, E, Stoll, B, Smith, O, Wierenga, PA, van de Heijning, BJM, et al. Postprandial amino acid kinetics of Milk protein mixtures are affected by composition, but not denaturation, in neonatal piglets. Curr Dev Nutr. (2019) 3:nzy102. doi: 10.1093/cdn/nzy102
168. Calvez, J, Blais, A, Deglaire, A, Gaudichon, C, Blachier, F, and Davila, A-M. Minimal processed infant formula vs conventional shows comparable protein quality and increased postprandial plasma amino acid kinetics in rats. Br J Nutr. (2023) 131:1115–24. doi: 10.1017/S0007114523002696
169. Rigo, J, Boehm, G, Georgi, G, Jelinek, J, Nyambugabo, K, Sawatzki, G, et al. An infant formula free of glycomacropeptide prevents hyperthreoninemia in formula-fed preterm infants. J Pediatr Gastroenterol Nutr. (2001) 32:127–30. doi: 10.1097/00005176-200102000-00006
170. Darling, PB, Dunn, M, Sarwar, G, Brookes, S, Ball, RO, and Pencharz, PB. Threonine kinetics in preterm infants fed their mothers’ milk or formula with various ratios of whey to casein. Am J Clin Nutr. (1999) 69:105–14. doi: 10.1093/ajcn/69.1.105
171. Janas, LM, Picciano, MF, and Hatch, TF. Indices of protein metabolism in term infants fed human milk, whey-predominant formula, or cow’s milk formula. Pediatrics. (1985) 75:775–83. doi: 10.1542/peds.75.4.775
172. Boehm, G, Cervantes, H, Georgi, G, Jelinek, J, Sawatzki, G, Wermuth, B, et al. Effect of increasing dietary threonine intakes on amino acid metabolism of the central nervous system and peripheral tissues in growing rats. Pediatr Res. (1998) 44:900–6. doi: 10.1203/00006450-199812000-00013
173. Edmonds, MS, Gonyou, HW, and Baker, DH. Effect of excess levels of methionine, tryptophan, arginine, lysine or threonine on growth and dietary choice in the pig. J Anim Sci. (1987) 65:179–85. doi: 10.2527/jas1987.651179x
174. Slupsky, CM, He, X, Hernell, O, Andersson, Y, Rudolph, C, Lönnerdal, B, et al. Postprandial metabolic response of breast-fed infants and infants fed lactose-free vs regular infant formula: a randomized controlled trial. Sci Rep. (2017) 7:3640. doi: 10.1038/s41598-017-03975-4
175. Karlsland Akeson, PM, Axelsson, IE, and Räihä, NC. Protein and amino acid metabolism in three- to twelve-month-old infants fed human milk or formulas with varying protein concentrations. J Pediatr Gastroenterol Nutr. (1998) 26:297–304. doi: 10.1097/00005176-199803000-00011
176. Lien, EL, Davis, AM, and Euler, ARMulticenter Study Group. Growth and safety in term infants fed reduced-protein formula with added bovine alpha-lactalbumin. J Pediatr Gastroenterol Nutr. (2004) 38:170–6. doi: 10.1097/00005176-200402000-00013
177. Räihä, NCR, Fazzolari-Nesci, A, Cajozzo, C, Puccio, G, Monestier, A, Moro, G, et al. Whey predominant, whey modified infant formula with protein/energy ratio of 1.8 g/100 kcal: adequate and safe for term infants from birth to four months. J Pediatr Gastroenterol Nutr. (2002) 35:275–81. doi: 10.1097/00005176-200209000-00008
178. Moro, G, Minoli, I, Boehm, G, Georgi, G, Jelinek, J, and Sawatzki, G. Postprandial plasma amino acids in preterm infants: influence of the protein source. Acta Paediatr. (1999) 88:885–9. doi: 10.1080/08035259950168838
179. Nicklaus, S, and Remy, E. Early origins of overeating: tracking between early food habits and later eating patterns. Curr Obes Rep. (2013) 2:179–84. doi: 10.1007/s13679-013-0055-x
180. Wang, M, Monaco, MH, and Donovan, SM. Impact of early gut microbiota on immune and metabolic development and function. Semin Fetal Neonatal Med. (2016) 21:380–7. doi: 10.1016/j.siny.2016.04.004
181. Mosca, F, and Giannì, ML. Human milk: composition and health benefits. Pediatr Med Chir. (2017) 39:155. doi: 10.4081/pmc.2017.155
182. Horta, BL . Loret De Mola C, orld health organization. Short-term effects of breastfeeding: a systematic review on the benefits of breastfeeding on diarrhoea and pneumonia mortality (2013). Available online at: https://www.who.int/publications/i/item/9789241506120 (Accessed May 5, 2025).
183. Boone, K, Geraghty, S, and Keim, S. Feeding at the breast and expressed Milk feeding: associations with otitis media and diarrhea in infants. J Pediatr. (2016) 174:118–25. doi: 10.1016/j.jpeds.2016.04.006
184. Ip, S, Chung, M, Raman, G, Chew, P, Magula, N, DeVine, D, et al. Breastfeeding and maternal and infant health outcomes in developed countries. Evid Rep Technol Assess (Full Rep). (2007) 153:1–186.
185. Bowatte, G, Tham, R, Allen, KJ, Tan, DJ, Lau, M, Dai, X, et al. Breastfeeding and childhood acute otitis media: a systematic review and meta-analysis. Acta Paediatr. (2015) 104:85–95. doi: 10.1111/apa.13151
186. Hanning, R, Paes, B, and Atkinson, S. Protein metabolism and growth of term infants in response to a reduced-protein, 40:60 whey: casein formula with added tryptophan. Am J Clin Nutr. (1992) 56:1004–11. doi: 10.1093/ajcn/56.6.1004
187. Monteiro, POA, and Victora, CG. Rapid growth in infancy and childhood and obesity in later life--a systematic review. Obes Rev. (2005) 6:143–54. doi: 10.1111/j.1467-789X.2005.00183.x
188. Stettler, N, Stallings, VA, Troxel, AB, Zhao, J, Schinnar, R, Nelson, SE, et al. Weight gain in the first week of life and overweight in adulthood: a cohort study of European American subjects fed infant formula. Circulation. (2005) 111:1897–903. doi: 10.1161/01.CIR.0000161797.67671.A7
189. Koletzko, B, von Kries, R, Monasterolo, R, Subías, J, Scaglioni, S, Giovannini, M, et al. Infant feeding and later obesity risk. Adv Exp Med Biol. (2009) 646:15–29. doi: 10.1007/978-1-4020-9173-5_2
190. Michaelsen, KF, and Greer, FR. Protein needs early in life and long-term health. Am J Clin Nutr. (2014) 99:718S–22S. doi: 10.3945/ajcn.113.072603
191. Savino, F, Benetti, S, Liguori, SA, Sorrenti, M, and Cordero Di Montezemolo, L. Advances on human milk hormones and protection against obesity. Cell Mol Biol (Noisy-le-grand). (2013) 59:89–98.
192. Horta, BL, Loret de Mola, C, and Victora, CG. Long-term consequences of breastfeeding on cholesterol, obesity, systolic blood pressure and type 2 diabetes: a systematic review and meta-analysis. Acta Paediatr. (2015) 104:30–7. doi: 10.1111/apa.13133
193. Lemaire, M, Dou, S, Cahu, A, Formal, M, Le Normand, L, Romé, V, et al. Addition of dairy lipids and probiotic Lactobacillus fermentum in infant formula programs gut microbiota and entero-insular axis in adult minipigs. Sci Rep. (2018) 8:11656. doi: 10.1038/s41598-018-29971-w
194. Xu, RJ . Development of the newborn GI tract and its relation to colostrum/milk intake: a review. Reprod Fertil Dev. (1996) 8:35–48. doi: 10.1071/rd9960035
195. Koenig, JE, Spor, A, Scalfone, N, Fricker, AD, Stombaugh, J, Knight, R, et al. Succession of microbial consortia in the developing infant gut microbiome. Proc Natl Acad Sci USA. (2011) 108:4578–85. doi: 10.1073/pnas.1000081107
196. Knickmeyer, RC, Gouttard, S, Kang, C, Evans, D, Wilber, K, Smith, JK, et al. A structural MRI study of human brain development from birth to 2 years. J Neurosci. (2008) 28:12176–82. doi: 10.1523/JNEUROSCI.3479-08.2008
197. Marques, T, Wall, R, Ross, RP, Fitzgerald, GF, Ryan, CA, and Stanton, C. Programming infant gut microbiota: influence of dietary and environmental factors. Curr Opin Biotechnol. (2010) 21:149–56. doi: 10.1016/j.copbio.2010.03.020
198. Brink, LR, Matazel, K, Piccolo, BD, Bowlin, AK, Chintapalli, SV, Shankar, K, et al. Neonatal diet impacts bioregional microbiota composition in piglets fed human breast Milk or infant formula. J Nutr. (2019) 149:2236–46. doi: 10.1093/jn/nxz170
199. Charton, E, Bourgeois, A, Bellanger, A, Gouar, Y, Dahirel, P, Cahu, A, et al. Infant nutrition affects the microbiota-gut-brain axis: comparison of human milk vs. infant formula feeding in the piglet model. Front Nutr. (2022) 9:42. doi: 10.3389/fnut.2022.976042
200. Ratsika, A, Codagnone, MC, O’Mahony, S, Stanton, C, and Cryan, JF. Priming for life: early life nutrition and the microbiota-gut-brain Axis. Nutrients. (2021) 13:423. doi: 10.3390/nu13020423
201. Valdes, AM, Walter, J, Segal, E, and Spector, TD. Role of the gut microbiota in nutrition and health. BMJ. (2018) 361:k2179. doi: 10.1136/bmj.k2179
202. Gómez-Gallego, C, Recio, I, Gómez-Gómez, V, Ortuño, I, Bernal, MJ, Ros, G, et al. Effect of processing on polyamine content and bioactive peptides released after in vitro gastrointestinal digestion of infant formulas. J Dairy Sci. (2016) 99:924–32. doi: 10.3168/jds.2015-10030
203. Levin, AM, Sitarik, AR, Havstad, SL, Fujimura, KE, Wegienka, G, Cassidy-Bushrow, AE, et al. Joint effects of pregnancy, sociocultural, and environmental factors on early life gut microbiome structure and diversity. Sci Rep. (2016) 6:31775. doi: 10.1038/srep31775
204. Pannaraj, PS, Li, F, Cerini, C, Bender, JM, Yang, S, Rollie, A, et al. Association between breast Milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. (2017) 171:647–54. doi: 10.1001/jamapediatrics.2017.0378
205. Ho, NT, Li, F, Lee-Sarwar, KA, Tun, HM, Brown, BP, Pannaraj, PS, et al. Meta-analysis of effects of exclusive breastfeeding on infant gut microbiota across populations. Nat Commun. (2018) 9:4169. doi: 10.1038/s41467-018-06473-x
206. Brink, LR, Mercer, KE, Piccolo, BD, Chintapalli, SV, Elolimy, A, Bowlin, AK, et al. Neonatal diet alters fecal microbiota and metabolome profiles at different ages in infants fed breast milk or formula. Am J Clin Nutr. (2020) 111:1190–202. doi: 10.1093/ajcn/nqaa076
207. Ma, J, Li, Z, Zhang, W, Zhang, C, Zhang, Y, Mei, H, et al. Comparison of gut microbiota in exclusively breast-fed and formula-fed babies: a study of 91 term infants. Sci Rep. (2020) 10:15792. doi: 10.1038/s41598-020-72635-x
208. Jost, T, Lacroix, C, Braegger, CP, and Chassard, C. New insights in gut microbiota establishment in healthy breast fed neonates. PLoS One. (2012) 7:e44595. doi: 10.1371/journal.pone.0044595
209. Stewart, CJ, Ajami, NJ, O’Brien, JL, Hutchinson, DS, Smith, DP, Wong, MC, et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature. (2018) 562:583–8. doi: 10.1038/s41586-018-0617-x
210. Penders, J, Thijs, C, Vink, C, Stelma, FF, Snijders, B, Kummeling, I, et al. Factors influencing the composition of the intestinal microbiota in early infancy. Pediatrics. (2006) 118:511–21. doi: 10.1542/peds.2005-2824
211. Adlerberth, I, and Wold, AE. Establishment of the gut microbiota in Western infants. Acta Paediatr. (2009) 98:229–38. doi: 10.1111/j.1651-2227.2008.01060.x
212. Boehm, G, and Moro, G. Structural and functional aspects of prebiotics used in infant Nutrition1. J Nutr. (2008) 138:1818S–28S. doi: 10.1093/jn/138.9.1818s
213. Stein-Thoeringer, CK, Nichols, KB, Lazrak, A, Docampo, MD, Slingerland, AE, Slingerland, JB, et al. Lactose drives Enterococcus expansion to promote graft-versus-host disease. Science. (2019) 366:1143–9. doi: 10.1126/science.aax3760
214. Lemaire, M, Ménard, O, Cahu, A, Nogret, I, Briard-Bion, V, Cudennec, B, et al. Addition of dairy lipids and probiotic Lactobacillus fermentum in infant formulas modulates proteolysis and lipolysis with moderate consequences on gut physiology and metabolism in Yucatan piglets. Front Nutr. (2021) 8:615248. doi: 10.3389/fnut.2021.615248
215. Arnold, RR, Brewer, M, and Gauthier, JJ. Bactericidal activity of human lactoferrin: sensitivity of a variety of microorganisms. Infect Immun. (1980) 28:893–8. doi: 10.1128/iai.28.3.893-898.1980
216. Ellison, RT, and Giehl, TJ. Killing of gram-negative bacteria by lactoferrin and lysozyme. J Clin Invest. (1991) 88:1080–91. doi: 10.1172/JCI115407
217. Nielsen, C, Hui, Y, Nguyen, D, Ahnfeldt, A, Burrin, DG, Hartmann, B, et al. Alpha-lactalbumin enriched whey protein concentrate to improve gut, immunity and brain development in preterm pigs. Nutrients. (2020) 12:245. doi: 10.3390/nu12010245
218. Brück, WM, Redgrave, M, Tuohy, KM, Lönnerdal, B, Graverholt, G, Hernell, O, et al. Effects of bovine alpha-lactalbumin and casein glycomacropeptide-enriched infant formulae on faecal microbiota in healthy term infants. J Pediatr Gastroenterol Nutr. (2006) 43:673–9. doi: 10.1097/01.mpg.0000232019.79025.8f
219. You, X . Human milk non-protein nitrogen metabolism by infant associated bifidobacteria. (2019). Available online at: https://hdl.handle.net/20.500.14394/18040 (Accessed May 5, 2025).
220. Singhal, A, Macfarlane, G, Macfarlane, S, Lanigan, J, Kennedy, K, Elias-Jones, A, et al. Dietary nucleotides and fecal microbiota in formula-fed infants: a randomized controlled trial1. Am J Clin Nutr. (2008) 87:1785–92. doi: 10.1093/ajcn/87.6.1785
221. Yamashiro, Y, Sato, M, Shimizu, T, Oguchi, S, Maruyama, K, and Kitamura, S. Possible biological growth factors in breast milk and postnatal development of the gastrointestinal tract. Acta Paediatr Jpn. (1989) 31:417–23. doi: 10.1111/j.1442-200x.1989.tb01327.x
222. Shulman, RJ, Schanler, RJ, Lau, C, Heitkemper, M, Ou, CN, and Smith, EO. Early feeding, antenatal glucocorticoids, and human milk decrease intestinal permeability in preterm infants. Pediatr Res. (1998) 44:519–23. doi: 10.1203/00006450-199810000-00009
223. De Quelen, F, Chevalier, J, Rolli-Derkinderen, M, Mourot, J, Neunlist, M, and Boudry, G. N–3 polyunsaturated fatty acids in the maternal diet modify the postnatal development of nervous regulation of intestinal permeability in piglets. J Physiol. (2011) 589:4341–52. doi: 10.1113/jphysiol.2011.214056
224. Desaldeleer, C, Ferret-Bernard, S, de Quelen, F, Le Normand, L, Perrier, C, Savary, G, et al. Maternal 18:3n-3 favors piglet intestinal passage of LPS and promotes intestinal anti-inflammatory response to this bacterial ligand. J Nutr Biochem. (2014) 25:1090–8. doi: 10.1016/j.jnutbio.2014.05.014
225. Lee, GO, McCormick, BJ, Seidman, JC, Kosek, MN, Haque, R, Olortegui, MP, et al. Infant nutritional status, feeding practices, enteropathogen exposure, socioeconomic status, and illness are associated with gut barrier function as assessed by the lactulose mannitol test in the MAL-ED birth cohort. Am J Trop Med Hyg. (2017) 97:281–90. doi: 10.4269/ajtmh.16-0830
226. Le Huërou-Luron, I, Blat, S, and Boudry, G. Breast- v. formula-feeding: impacts on the digestive tract and immediate and long-term health effects. Nutr Res Rev. (2010) 23:23–36. doi: 10.1017/S0954422410000065
227. Elolimy, AA, Washam, C, Byrum, S, Chen, C, Dawson, H, Bowlin, AK, et al. Formula diet alters the ileal metagenome and transcriptome at weaning and during the postweaning period in a porcine model. mSystems. (2020) 5:20. doi: 10.1128/msystems.00457-20
228. Boudry, G, Morise, A, Seve, B, and Le Huërou-Luron, I. Effect of Milk formula protein content on intestinal barrier function in a porcine model of LBW neonates. Pediatr Res. (2011) 69:4–9. doi: 10.1203/PDR.0b013e3181fc9d13
229. Weaver, LT . The impact of milk and weaning diet on gastrointestinal permeability in English and Gambian infants. Trans R Soc Trop Med Hyg. (1988) 82:784–9. doi: 10.1016/0035-9203(88)90236-2
230. Catassi, C, Bonucci, A, Coppa, GV, Carlucci, A, and Giorgi, PL. Intestinal permeability changes during the first month: effect of natural versus artificial feeding. J Pediatr Gastroenterol Nutr. (1995) 21:383–6. doi: 10.1097/00005176-199511000-00003
231. Castanet, M, Costalos, C, Haiden, N, Hascoet, J-M, Berger, B, Sprenger, N, et al. Early effect of supplemented infant formulae on intestinal biomarkers and microbiota: a randomized clinical trial. Nutrients. (2020) 12:1481. doi: 10.3390/nu12051481
232. Laubereau, B, Brockow, I, Zirngibl, A, Koletzko, S, Gruebl, A, von Berg, A, et al. Effect of breast-feeding on the development of atopic dermatitis during the first 3 years of life—results from the GINI-birth cohort study. J Pediatr. (2004) 144:602–7. doi: 10.1016/j.jpeds.2003.12.029
233. Calder, PC, Krauss-Etschmann, S, de Jong, EC, Dupont, C, Frick, J-S, Frokiaer, H, et al. Early nutrition and immunity - progress and perspectives. Br J Nutr. (2006) 96:774–90.
234. Jeon, M-T, Kim, K-S, Kim, ES, Lee, S, Kim, J, Hoe, H-S, et al. Emerging pathogenic role of peripheral blood factors following BBB disruption in neurodegenerative disease. Ageing Res Rev. (2021) 68:101333. doi: 10.1016/j.arr.2021.101333
235. Savino, F, Castagno, E, Calabrese, R, Viola, S, Oggero, R, and Miniero, R. High faecal calprotectin levels in healthy, exclusively breast-fed infants. Neonatology. (2010) 97:299–304. doi: 10.1159/000255161
236. Li, F, Ma, J, Geng, S, Wang, J, Ren, F, and Sheng, X. Comparison of the different kinds of feeding on the level of fecal calprotectin. Early Hum Dev. (2014) 90:471–5. doi: 10.1016/j.earlhumdev.2014.06.005
237. Asgarshirazi, M, Shariat, M, Nayeri, F, Dalili, H, and Abdollahi, A. Comparison of fecal calprotectin in exclusively breastfed and formula or mixed fed infants in the first six months of life. Acta Med Iran. (2017) 55:53–8.
238. Al Nabhani, Z, Dulauroy, S, Marques, R, Cousu, C, Al Bounny, S, Déjardin, F, et al. A weaning reaction to microbiota is required for resistance to Immunopathologies in the adult. Immunity. (2019) 50:1276–1288.e5. doi: 10.1016/j.immuni.2019.02.014
239. Rougé, C, Butel, M-J, Piloquet, H, Ferraris, L, Legrand, A, Vodovar, M, et al. Fecal calprotectin excretion in preterm infants during the neonatal period. PLoS One. (2010) 5:e11083. doi: 10.1371/journal.pone.0011083
240. Borre, YE, O’Keeffe, GW, Clarke, G, Stanton, C, Dinan, TG, and Cryan, JF. Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med. (2014) 20:509–18. doi: 10.1016/j.molmed.2014.05.002
241. Jena, A, Montoya, CA, Mullaney, JA, Dilger, RN, Young, W, McNabb, WC, et al. Gut-brain Axis in the early postnatal years of life: a developmental perspective. Front Integr Neurosci. (2020) 14:44. doi: 10.3389/fnint.2020.00044
242. Kramer, MS, Aboud, F, Mironova, E, Vanilovich, I, Platt, RW, Matush, L, et al. Breastfeeding and child cognitive development: new evidence from a large randomized trial. Arch Gen Psychiatry. (2008) 65:578–84. doi: 10.1001/archpsyc.65.5.578
243. Deoni, SCL, Dean, DC, Piryatinsky, I, O’Muircheartaigh, J, Waskiewicz, N, Lehman, K, et al. Breastfeeding and early white matter development: a cross-sectional study. NeuroImage. (2013) 82:77–86. doi: 10.1016/j.neuroimage.2013.05.090
244. Timby, N, Lönnerdal, B, Hernell, O, and Domellöf, M. Cardiovascular risk markers until 12 mo of age in infants fed a formula supplemented with bovine milk fat globule membranes. Pediatr Res. (2014) 76:394–400. doi: 10.1038/pr.2014.110
245. Deoni, S, Dean, D, Joelson, S, O’Regan, J, and Schneider, N. Early nutrition influences developmental myelination and cognition in infants and young children. NeuroImage. (2018) 178:649–59. doi: 10.1016/j.neuroimage.2017.12.056
246. Horta, BL, Loret de Mola, C, and Victora, CG. Breastfeeding and intelligence: a systematic review and meta-analysis. Acta Paediatr. (2015) 104:14–9. doi: 10.1111/apa.13139
247. Timby, N, Domellöf, M, Lönnerdal, B, and Hernell, O. Supplementation of infant formula with bovine Milk fat globule membranes. Adv Nutr. (2017) 8:351–5. doi: 10.3945/an.116.014142
248. Koletzko, B, Rodriguez-Palmero, M, Demmelmair, H, Fidler, N, Jensen, R, and Sauerwald, T. Physiological aspects of human milk lipids. Early Hum Dev. (2001) 65:S3–S18. doi: 10.1016/s0378-3782(01)00204-3
249. Carlson, SE, Ford, AJ, Werkman, SH, Peeples, JM, and Koo, WW. Visual acuity and fatty acid status of term infants fed human milk and formulas with and without docosahexaenoate and arachidonate from egg yolk lecithin. Pediatr Res. (1996) 39:882–8. doi: 10.1203/00006450-199605000-00024
250. Birch, EE, Hoffman, DR, Uauy, R, Birch, DG, and Prestidge, C. Visual acuity and the essentiality of docosahexaenoic acid and arachidonic acid in the diet of term infants. Pediatr Res. (1998) 44:201–9. doi: 10.1203/00006450-199808000-00011
251. Birch, EE, Carlson, SE, Hoffman, DR, Fitzgerald-Gustafson, KMM, Fu, VL, Drover, JR, et al. The DIAMOND (DHA intake and measurement of neural development) Study: a double-masked, randomized controlled clinical trial of the maturation of infant visual acuity as a function of the dietary level of docosahexaenoic acid. Am J Clin Nutr. (2010) 91:848–59. doi: 10.3945/ajcn.2009.28557
252. Janas, LM, and Picciano, MF. The nucleotide profile of human milk. Pediatr Res. (1982) 16:659–62. doi: 10.1203/00006450-198208000-00014
253. Atkinson, S, Anderson, G, and Bryan, M. Human milk: comparison of the nitrogen composition in milk from mothers of premature and full-term infants. Am J Clin Nutr. (1980) 33:811–5. doi: 10.1093/ajcn/33.4.811
254. Neville, MC, Keller, RP, Seacat, J, Casey, CE, Allen, JC, and Archer, P. Studies on human lactation. I. Within-feed and between-breast variation in selected components of human milk. Am J Clin Nutr. (1984) 40:635–46. doi: 10.1093/ajcn/40.3.635
255. Ross, SA, and Clark, RM. Nitrogen distribution in human milk from 2 to 16 weeks postpartum. J Dairy Sci. (1985) 68:3199–201. doi: 10.3168/jds.S0022-0302(85)81227-3
256. Agostoni, C, Carratù, B, Boniglia, C, Riva, E, and Sanzini, E. Free amino acid content in standard infant formulas: comparison with human milk. J Am Coll Nutr. (2000) 19:434–8. doi: 10.1080/07315724.2000.10718943
257. Carratù, B, Boniglia, C, Scalise, F, Ambruzzi, AM, and Sanzini, E. Nitrogenous components of human milk: non-protein nitrogen, true protein and free amino acids. Food Chem. (2003) 81:357–62. doi: 10.1016/S0308-8146(02)00430-2
258. Park, Y, Juárez, M, Ramos, M, and Haenlein, G. Physico-chemical characteristics of goat and sheep milk. Small Rumin Res. (2007) 68:88–113. doi: 10.1016/j.smallrumres.2006.09.013
259. Grote, V, Verduci, E, Scaglioni, S, Vecchi, F, Contarini, G, Giovannini, M, et al. Breast milk composition and infant nutrient intakes during the first 12 months of life. Eur J Clin Nutr. (2016) 70:250–6. doi: 10.1038/ejcn.2015.162
260. Ng-Kwai-Hang, KF, Hayes, JF, Moxley, JE, and Monardes, HG. Percentages of protein and nonprotein nitrogen with varying fat and somatic cells in bovine Milk. J Dairy Sci. (1985) 68:1257–62. doi: 10.3168/jds.S0022-0302(85)80954-1
261. DePeters, EJ, and Ferguson, JD. Nonprotein nitrogen and protein distribution in the milk of cows. J Dairy Sci. (1992) 75:3192–209. doi: 10.3168/jds.S0022-0302(92)78085-0
262. Baker, LD, Ferguson, JD, and Chalupa, W. Responses in urea and true protein of milk to different protein feeding schemes for dairy cows. J Dairy Sci. (1995) 78:2424–34. doi: 10.3168/jds.S0022-0302(95)76871-0
263. Hadjipanayiotou, M . Composition of ewe, goat and cow milk and of colostrum of ewes and goats. Small Rumin Res. (1995) 18:255–62. doi: 10.1016/0921-4488(95)00697-3
264. Mustafa, A, Chouinard, P, and Christensen, D. Effects of feeding micronised flaxseed on yield and composition of milk from Holstein cows. J Sci Food Agric. (2003) 83:920–6. doi: 10.1002/jsfa.1430
265. Couvreur, S, Hurtaud, C, Marnet, PG, Faverdin, P, and Peyraud, JL. Composition of milk fat from cows selected for milk fat globule size and offered either fresh pasture or a corn silage-based diet. J Dairy Sci. (2007) 90:392–403. doi: 10.3168/jds.S0022-0302(07)72640-1
266. Olalla, M, Ruiz-López, D, Navarro, M, Artacho, R, Cabrera, C, Giménez, R, et al. Nitrogen fractions of Andalusian goat milk compared to similar types of commercial milk. Food Chem. (2009) 113:835–8. doi: 10.1016/j.foodchem.2008.10.022
267. Ruska, D, and Jonkus, D. Crude protein and non-protein nitrogen content in dairy cow Milk. Proc Latvia Univ Agric. (2014) 32:36–40. doi: 10.2478/plua-2014-0011
268. Kapadiya, D, Prajapati, D, Jain, A, Mehta, B, Darji, V, and Aparnathi, K. Comparison of Surti goat milk with cow and buffalo milk for gross composition, nitrogen distribution, and selected minerals content. Vet World. (2016) 9:710–6. doi: 10.14202/vetworld.2016.710-716
269. Rafiq, S, Huma, N, Pasha, I, Sameen, A, Mukhtar, O, and Khan, MI. Chemical composition, nitrogen fractions and amino acids profile of Milk from different animal species. Asian Australas J Anim Sci. (2016) 29:1022–8. doi: 10.5713/ajas.15.0452
270. Caroprese, M, Mancino, R, Ciliberti, MG, Di Luccia, A, La Gatta, B, and Albenzio, M. Fatty acid profile and coagulating ability of milk from Jersey and Friesian cows fed whole flaxseed. J Dairy Res. (2017) 84:14–22. doi: 10.1017/S002202991600073X
271. Biadała, A . Goat’s milk-derived bioactive components - a review. Mljekarstvo. (2018) 68:239–53. doi: 10.15567/mljekarstvo.2018.0401
272. Ferreira, I . Quantification of non-protein nitrogen components of infant formulae and follow-up milks: comparison with cows’ and human milk. Br J Nutr. (2003) 90:127–33. doi: 10.1079/bjn2003882
273. Harzer, G, Franzke, V, and Bindels, JG. Human milk nonprotein nitrogen components: changing patterns of free amino acids and urea in the course of early lactation. Am J Clin Nutr. (1984) 40:303–9. doi: 10.1093/ajcn/40.2.303
274. Ferreira, I, Gomes, A, and Ferreira, M. Determination of sugars, and some other compounds in infant formulae, follow-up milks and human milk by HPLC-UV/RI. Carbohydr Polym. (1998) 37:225–9. doi: 10.1016/S0144-8617(98)00064-2
275. Sarwar, G, Botting, HG, Davis, TA, Darling, P, and Pencharz, PB. Free amino acids in milks of human subjects, other primates and non-primates. Br J Nutr. (1998) 79:129–31. doi: 10.1079/bjn19980023
276. Smilowitz, J, O’Sullivan, A, Barile, D, German, J, Lönnerdal, B, and Slupsky, C. The human milk metabolome reveals diverse oligosaccharide profiles. J Nutr. (2013) 143:1709–18. doi: 10.3945/jn.113.178772
277. Macy, IG . Composition of human colostrum and milk. Am J Dis Child (1911). (1949) 78:589–603. doi: 10.1001/archpedi.1949.02030050604009
278. Schimmel, P, Stahl, B, Knol, J, and Belzer, C. The infant gut microbiota: in pursuit of non-protein nitrogen. Gut Microbes. (2023) 15:2211917. doi: 10.1080/19490976.2023.2211917
279. Edison, EE, Brosnan, ME, Aziz, K, and Brosnan, JT. Creatine and guanidinoacetate content of human milk and infant formulas: implications for creatine deficiency syndromes and amino acid metabolism. Br J Nutr. (2013) 110:1075–8. doi: 10.1017/s000711451300010x
280. Ferreira, I, and Ferreira, M. Simultaneous determination of sugars, uric and orotic acids in infant formulae by HPLC-UV/RI. J Liq Chromatogr Relat Technol. (1997) 20:3419–29. doi: 10.1080/10826079708005841
281. Takatsu, Z, Matsumoto, H, Hanaki, K, Zhang, Z, Hou, R, Wang, H, et al. Glucosamine contents of milk hydrolysates from various mammals. Thai J Vet Med. (2020) 50:583–7. doi: 10.56808/2985-1130.3065
282. Xinmin, W, Ruili, Z, Zhihua, L, Yuanhong, W, and Tingfu, J. Determination of glucosamine and lactose in milk-based formulae by high-performance liquid chromatography. J Food Compos Anal. (2008) 21:255–8. doi: 10.1016/j.jfca.2007.10.006
283. Spichtig, V, Michaud, J, and Austin, S. Determination of sialic acids in milks and milk-based products. Anal Biochem. (2010) 405:28–40. doi: 10.1016/j.ab.2010.06.010
284. Romain, N, Dandrifosse, G, Jeusette, F, and Forget, P. Polyamine concentration in rat milk and food, human milk, and infant formulas. Pediatr Res. (1992) 32:58–63. doi: 10.1203/00006450-199207000-00011
285. Gómez-Gallego, C, Kumar, H, García-Mantrana, I, du Toit, E, Suomela, J-P, Linderborg, KM, et al. Breast Milk polyamines and microbiota interactions: impact of mode of delivery and geographical location. Ann Nutr Metab. (2017) 70:184–90. doi: 10.1159/000457134
286. Löser, C . Polyamines in human and animal milk. Br J Nutr. (2000) 84:55–8. doi: 10.1017/s0007114500002257
287. Muñoz-Esparza, NC, Comas-Basté, O, Latorre-Moratalla, ML, Veciana-Nogués, MT, and Vidal-Carou, MC. Differences in polyamine content between human Milk and infant formulas. Foods. (2021) 10:2866. doi: 10.3390/foods10112866
288. Abrahamse, E, Thomassen, GGM, Renes, IB, Wierenga, PA, and Hettinga, KA. Gastrointestinal protein hydrolysis kinetics: opportunities for further infant formula improvement. Nutrients. (2022) 14:1512. doi: 10.3390/nu14071512
289. Halabi, A, Croguennec, T, Ménard, O, Briard-Bion, V, Jardin, J, Le Gouar, Y, et al. Protein structure in model infant milk formulas impacts their kinetics of hydrolysis under in vitro dynamic digestion. Food Hydrocoll. (2022) 126:107368. doi: 10.1016/j.foodhyd.2021.107368
Keywords: milk protein, protein structure, non-protein nitrogen, digestion, plasma amino acid, gut microbiota, gut physiology, metabolism
Citation: Chauvet L, Charton E, Lemaire M, Le Huërou-Luron I and Deglaire A (2025) Human milk vs. cow-milk based infant formula proteins: structure, digestion and physiological impacts. Front. Nutr. 12:1635919. doi: 10.3389/fnut.2025.1635919
Edited by:
Claude Billeaud, Independent Paediatrics Nutrition Expert, Bordeaux, FranceReviewed by:
Sophie Gallier, Dairy Goat Co-Operative, New ZealandAndre Leke, Centre Hospitalier Universitaire (CHU) d’Amiens, France
Copyright © 2025 Chauvet, Charton, Lemaire, Le Huërou-Luron and Deglaire. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Amélie Deglaire, QW1lbGllLmRlZ2xhaXJlQGluc3RpdHV0LWFncm8uZnI=; Isabelle Le Huërou-Luron, SXNhYmVsbGUubHVyb25AaW5yYWUuZnI=
†These authors have contributed equally to this work
‡These authors have contributed equally to this work and share senior authorship