 Jyoti Prakash Tamang1
Jyoti Prakash Tamang1 Fojan Agahi2*
Fojan Agahi2* Birsen Yilmaz3
Birsen Yilmaz3 İbrahim Ender Künili4
İbrahim Ender Künili4 Julie Mardon5*
Julie Mardon5* Tuğçe Bulmus-Tuccar6
Tuğçe Bulmus-Tuccar6 Aleksandra Torbica7
Aleksandra Torbica7 Daniela Nikolovska Nedelkoska8
Daniela Nikolovska Nedelkoska8 Mary-Liis Kütt9
Mary-Liis Kütt9 Jeadran Malagón-Rojas10
Jeadran Malagón-Rojas10 Mayra Alejandra Parada11
Mayra Alejandra Parada11 Baltasar Mayo12,13
Baltasar Mayo12,13 Juana Frias2
Juana Frias2- 1Department of Microbiology, Sikkim University, Gangtok, Sikkim, India
- 2Instituto de Ciencia y Tecnología de Alimentos y Nutrición (ICTAN-CSIC), Madrid, Spain
- 3Department of Biological Sciences, Tata Institute of Fundamental Research, Hyderabad, India
- 4Department of Fishing & Fish Processing Technology, The Faculty of Marine Sciences and Technology, Çanakke Onsekiz Mart University, Çanakkale, Türkiye
- 5Université Clermont Auvergne, INRAE, Lempdes, France
- 6Arla Innovation Center, Arla Foods, Aarhus, Denmark
- 7Institute of Food Technology, University of Novi Sad, Novi Sad, Serbia
- 8Faculty of Technology and Technical Sciences, University "St. Kliment Ohridski"-Bitola, Veles, North Macedonia
- 9äio tech OÜ, Tallinn, Estonia
- 10Observatorio Nacional de Salud, Instituto Nacional de Salud, Bogotá, Colombia
- 11Grupo de Salud Ambiental y Laboral, Instituto Nacional de Salud, Bogotá, Colombia
- 12Instituto de Productos Lácteos de Asturias (IPLA-CSIC), Oviedo, Spain
- 13Investigación Sanitaria del Principado de Asturias (ISPA), Oviedo, Spain
Fermented foods are increasingly recognized for their potential benefits in supporting bone health, attributed to their rich content of bioactive compounds including vitamins K and B, polyphenols, peptides, and fermentation-modified phytates. This review examines how these components, enhanced in bioavailability through fermentation, may modulate bone metabolism via multiple mechanisms: improving mineral absorption, reducing inflammation, regulating oxidative stress, and influencing osteoblast and osteoclast activity. Special attention is given to the gut-bone axis, where fermented foods interact with gut microbiota to produce metabolites such as short-chain fatty acids and immunomodulatory compounds that may further support skeletal health. While preclinical and population-level studies show promising associations, clinical evidence remains limited and heterogeneous. Future research should focus on human trials, strain-specific effects, and long-term outcomes to fully establish the role of fermented foods in osteoporosis prevention and bone health maintenance.
Introduction
Fermentation is an ancient method for food preservation and for producing new food items. Fermented foods worldwide are grouped according to the local ingredients available and the indigenous techniques employed to produce the edible products with desired sensory properties, which are named in accordance with local custom. These groups encompass fermented dairy, fermented grains, fermented meats, fermented fish, fermented legumes, fermented soybeans, fermented vegetables, fermented roots like cassava, and others (Table 1) (1).
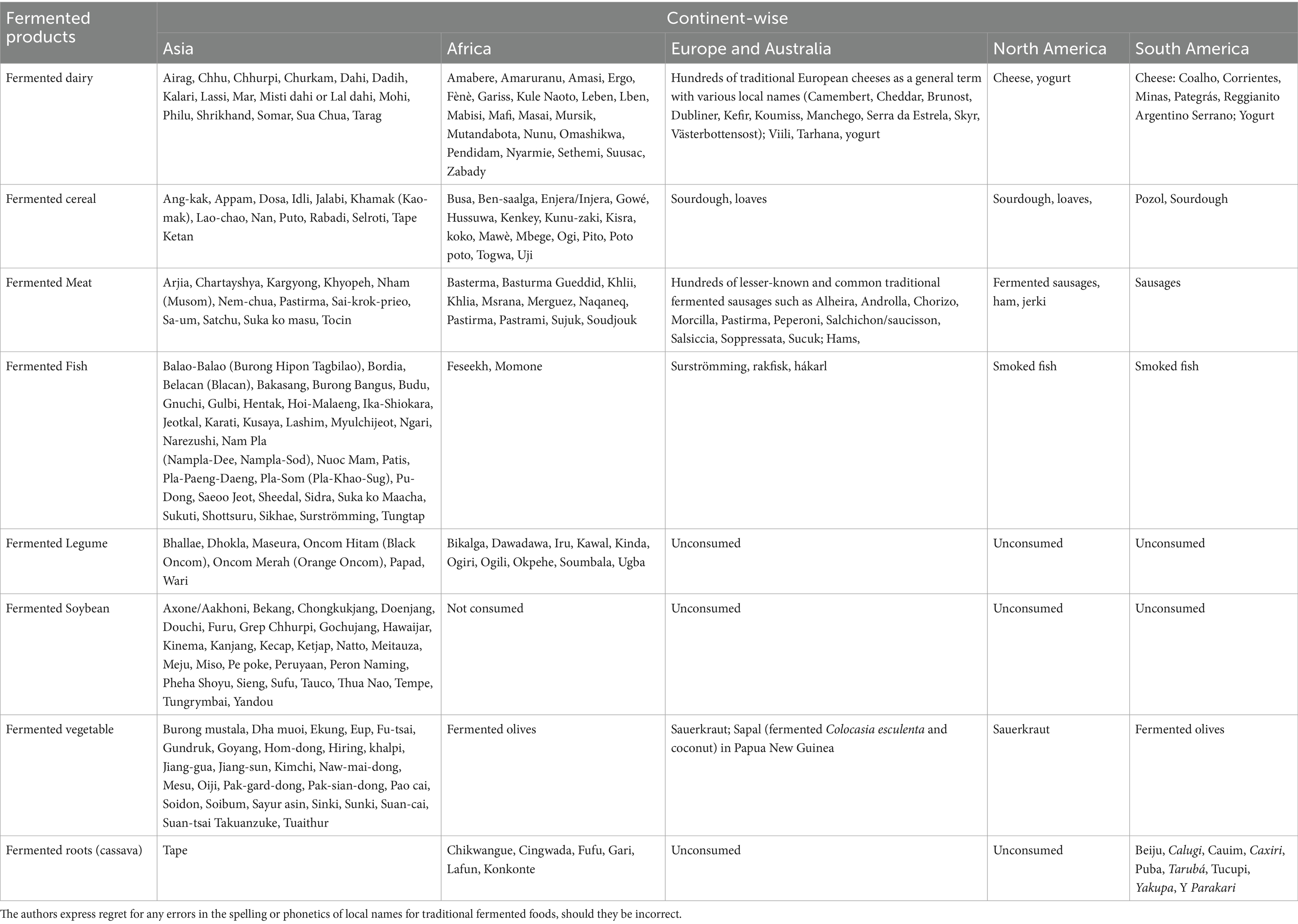
Table 1. Native names of some traditional fermented foods of the world (1).
In regions with widespread pastoral farming, such as the Middle East, Europe, and the Indian subcontinent, abundant milk from cows, sheep, and goats led to the development of fermented dairy products like fermented milk and cheese (2). In contrast, East Asian countries like China, Japan, and Korea, where animal husbandry was less prominent, developed fermented fish and soy-based foods instead (3, 4). In Africa, fermentation traditions rely on local grains (millet, sorghum, maize, wheat) and roots like cassava, alongside naturally fermented dairy from cows, buffalo, and camels (5, 6). Similarly, in Europe, the Americas, and Australia, livestock farming has supported the production of fermented dairy (7) and meat products (8), which are an integral part of local diets.
Traditionally, foods that undergo natural or spontaneous fermentation are produced from either plant or animal origins that do not require starter cultures. Numerous fermented foods are still produced via natural or spontaneous fermentation (9) or by employing back-slopping methods (10, 11). Many traditional fermented foods continue to be made at home, on a small scale, and with traditional practices. Nevertheless, the 20th century witnessed a substantial rise in the availability of starter cultures, which are now commonly used in commercial fermentation processes for products such as wine, dairy, and meat, particularly in Western nations. The application of starter cultures, utilizing the functional strains of food-grade bacteria, yeasts, or molds, represents a major transformation in the fermented foods and beverages market (12).
Historically, the main role of fermented foods was for preservation, but their extra advantages became noticeable as time passed. Prior to the establishment of nutrition science, fermented foods were made to ensure a consistent supply of vitamins, minerals, calories, and other vital nutrients required for survival. Progress in food microbiology and nutrition has indicated that specific beneficial microorganisms are essential in converting raw materials into fermented food products. These microorganisms aid in producing bioactive substances, vitamins, immune system enhancers, short-chain fatty acids (SCFAs), and secondary metabolites, which all contribute to consumer health and well-being (13–15). Genomic techniques supported by multi-omics approaches are increasingly being employed to investigate diverse microbial communities and their bio-functionalities present in fermented foods (16, 17), as well as to detect unidentified biomarkers or genetic signatures for various health and therapeutic purposes (18).
Fermented foods present numerous health benefits, which include improved digestion and regulation of the gut microbiome (19–21), prevention of type 2 diabetes and metabolic syndrome (22, 23), decreased risk of cardiovascular diseases (24), relief from lactose intolerance (25), reduction of inflammation (26), combating obesity (27–29), aiding bone healing (30, 31), exhibiting anti-cancer properties (32, 33), and preventing neurodegenerative diseases (34). Fermented foods are also positioned as key dietary ingredients to promote bone health. Since bone-related diseases represent a wide burden worldwide, it is of great importance to investigate the ability of fermented food to preserve bone health. Indeed, for instance, osteoporosis has a significant impact on individuals’ quality of life and morbidity worldwide, particularly among the elderly and postmenopausal women. Given the increase of the number of bone fractures during the last decades as well as the associated burden to individuals, families, societies, and health-care systems (35, 36), the prevention of bone diseases is a public health priority. Thus, a thorough investigation of the links between fermented foods and bone health is essential. Among these emerging benefits, bone health has gained attention as a novel and promising target for dietary modulation through fermented foods. This interest arises from several factors. First, osteoporosis and low bone mineral density (BMD) are major global public health concerns, particularly affecting aging populations and postmenopausal women (36–38). Second, diet plays a pivotal role in modulating bone metabolism, and the gut–bone axis has emerged as a key mediator linking dietary habits to skeletal outcomes (39). Recent evidence suggests that fermented foods through their unique matrix of nutrients, microbial metabolites, and live microorganisms may influence bone health beyond their basic nutritional content (40–42). This influence is hypothesized to occur through various mechanisms including modulation of gut microbiota, reduction of systemic inflammation, and enhancement of mineral bioavailability (43, 44).
Bone is a dense and structured tissue made up of cells surrounded by a plentiful hard intercellular substance that consists of collagen fibers and calcium phosphate (45). The growth, preservation, and renewal of bone tissues within the human body involve a series of co5rdinated mechanisms that result in tissue development, maintenance, and healing after injury (46). Bone metabolism necessitates essential dietary micronutrients like calcium (47), phosphorus (48), magnesium (49), and vitamin D (50). Minerals are taken up in the upper section of the gastrointestinal tract in an ionized form, which is the typical state under the stomach’s low pH. The uptake of calcium is significantly enhanced by vitamin D, making this vitamin essential for its availability in the body (51). Research has also linked vitamins B complex and K as important factors in maintaining bone health (52). The way B vitamins influence bone physiology seems to be related to their impact on homocysteine metabolism (53), while the action of vitamin K appears to occur through the steroid xenobiotic receptor and/or through the γ-carboxylation of osteocalcin (OC), which is the most prevalent vitamin K-dependent protein specific to bone (54).
In recent years, fermented foods have attracted attention as possible dietary interventions for bone health because of their rich content of bioactive compounds (40–42, 55, 56). Fermentation is a microbial biochemical process that produces numerous metabolites (organic acids, peptides, vitamins, etc.) from the macromolecules of raw materials, some of which may influence bone health (57–59). Certain fermented foods are particularly rich in group B vitamins (riboflavin and folates) and vitamin K (menaquinones) (60, 61), as they are predominantly produced by fermentative microorganisms, especially those belonging to lactic acid bacteria (LAB). In plant-based foods, the absorption of minerals is negatively impacted by the presence of anti-nutritional factors, such as oxalates and phytates (62); these are diminished during fermentation due to the action of microbial enzymes, simultaneously aiding in the release (and availability) of phosphorus (63). Additionally, the low pH of most fermented foods may delay the neutralization of the chyme, thereby prolonging ionization and enhancing absorption (64). Hence, the intake of fermented foods could assist the body in absorbing a greater amount of minerals and improving the intake of vitamins that promote bone health. Preclinical and population studies have suggested potential associations between fermented food consumption and improved bone density or reduced fracture risk (65–67), however, the multiple underlying mechanisms ranging from enhanced nutrient bioavailability and microbiota modulation to anti-inflammatory effects, have not been comprehensively investigated within a single framework.
Phytic acid, vitamins B complex and K, polyphenols, and bioactive peptides are essential biological compounds affecting bone health in humans (68–71). These compounds whether naturally present, enriched, or modified through fermentation, are central to understanding how fermented foods may influence bone metabolism. Recently, it has been reported that fermented foods may enhance bone health via the gut microbiota by stimulating the osteoimmune system, producing SCFAs, and facilitating the absorption of nutrients such as calcium (72).
While most of these compounds are generated or enriched during fermentation, in some cases like phytic acid, fermentation contributes by reducing their levels, thereby indirectly supporting mineral bioavailability and bone health. Despite this reduction, phytic acid itself has been reported to exert context-dependent effects on bone metabolism, and is therefore discussed as a relevant compound within the scope of this review.
However, despite increasing interest, the body of evidence remains fragmented across food types, health claims, and mechanisms. A comprehensive review is thus warranted to systematically explore and consolidate current knowledge on the potential of fermented foods in maintaining or improving bone health, their mechanisms of action, and their relevance in dietary strategies for osteoporosis prevention and bone metabolism modulation. Hence, we attempt to review the various beneficial effects of consuming fermented foods on bone health and the molecular underpinnings of the various mechanisms by which these benefits are conferred. The proposed mechanisms by which fermented foods and their bioactive compounds may support bone health, primarily through modulation of gut microbiota and enhancement of nutrient bioavailability, are illustrated in Figure 1.
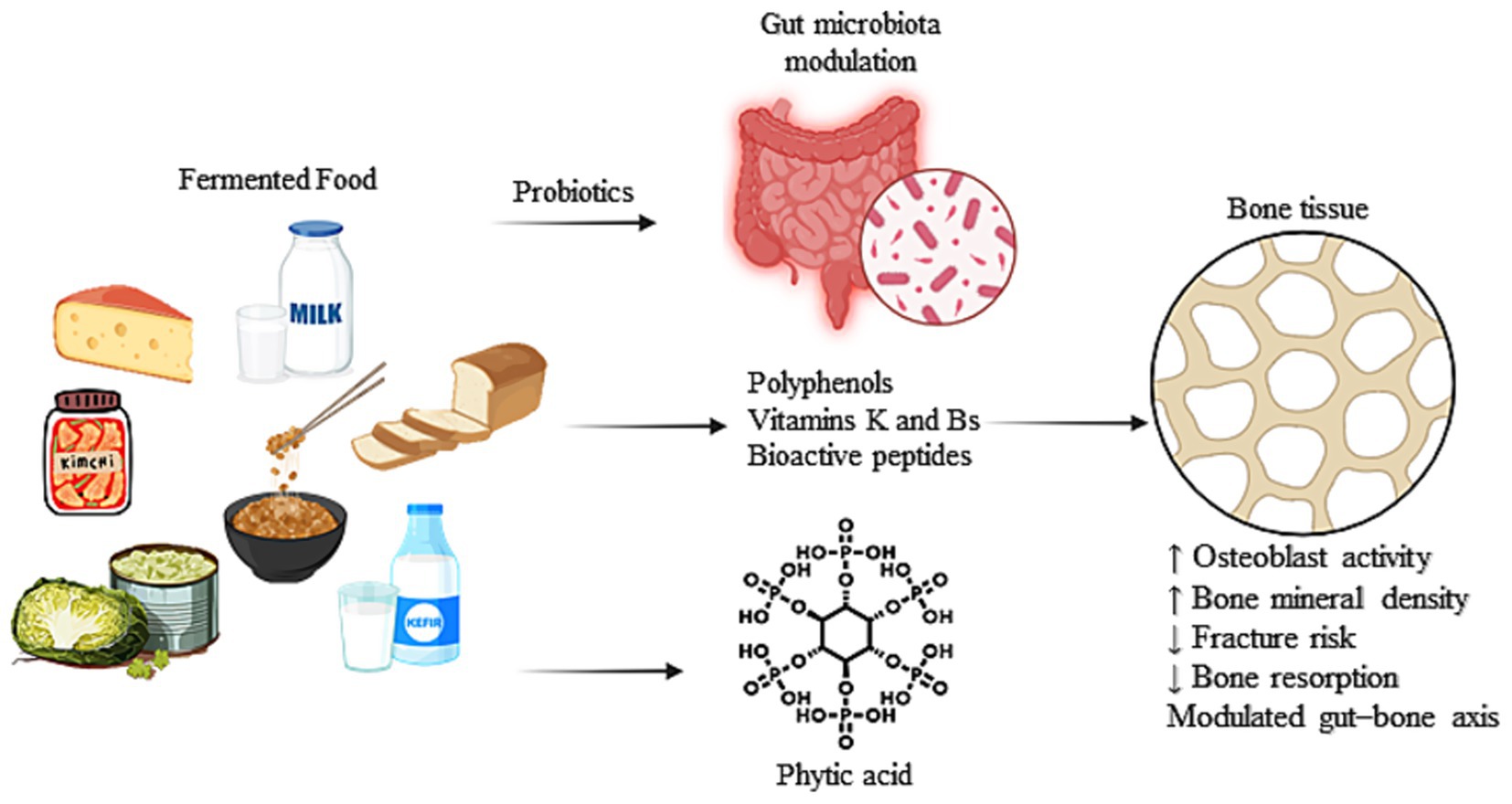
Figure 1. Schematic representation of the effects of fermented food-derived bioactive compounds and microbial modulation on bone metabolism.
Phytic acid and bone health
Phytic acid (C₆H₁₈O₂₄P₆), also known as inositol hexaphosphate, inositol hexakisphosphate, or inositol polyphosphate, is a naturally occurring compound consisting of six dihydrogen phosphate groups linked to inositol (73). At physiological pH levels commonly found in plants, phytic acid exists predominantly as phytate anions, specifically myo-inositol-1,2,3,4,5,6-hexakisphosphates (74). These compounds serve as major phosphorus storage molecules in plant seeds, accounting for approximately 10% of total plant weight and over 60% of total phosphorus content, particularly in cereals, legumes, oilseeds, and nuts (75–77). The structural characteristics of phytates—featuring negatively charged phosphate groups arranged around inositol rings—enable them to form strong chelation complexes with divalent cations, including zinc, iron, calcium, magnesium, manganese, and copper (78, 79). This mineral-binding capacity has traditionally classified phytates as antinutrients, as consumption of raw phytate-rich plant materials can significantly reduce the bioavailability and absorption of essential minerals (80).
However, recent research has fundamentally challenged the traditional view of phytic acid as solely an antinutrient, revealing complex beneficial effects on bone metabolism. Studies have demonstrated that phytic acid contributes to bone development and helps mitigate age-related deterioration of bone marrow mesenchymal stem cells (BMSCs), particularly under hyperglycemic conditions. In diabetic environments, phytic acid enhances osteogenic differentiation and suppresses oxidative stress-induced cellular senescence, primarily through activation of the extracellular signal-regulated kinase signaling pathway (70).
The molecular mechanisms underlying these beneficial effects have been further elucidated through investigations of phytic acid’s role in bone regeneration under diabetic conditions. Phytic acid supplementation restores the osteogenic capacity of BMSCs by modulating the expression of circular RNA (circRNA) circEIF4B (a non-coding RNA that regulates gene expression by acting as a microRNA sponge). This regulatory mechanism promotes osteogenesis by sequestering miR-186-5p and upregulating forkhead box protein O1, while simultaneously stabilizing integrin subunit alpha 5 (ITGA5) mRNA through inhibition of insulin-like growth factor 2 mRNA-binding protein 3 (IGF2BP3) degradation. Notably, the inhibition of circEIF4B impairs the bone-regenerative effects of phytic acid in vivo, confirming the functional importance of this molecular axis in diabetes-related bone disorders (81). Clinical evidence supports these mechanistic findings. In a Mediterranean cohort of postmenopausal women, dietary phytate intake showed positive associations with bone mineral density at multiple skeletal sites, including the femoral neck, total femur, and lumbar spine. This protective effect was most pronounced in women under 66 years of age without type 2 diabetes, with the proposed mechanism involving phytate adsorption onto hydroxyapatite surfaces, thereby inhibiting bone resorption (82).
Additional studies have demonstrated that phytate functions as both a crystallization and dissolution inhibitor by adsorbing onto hydroxyapatite crystal surfaces. In vitro experiments revealed that the anti-resorptive effects of phytate were comparable to alendronate and superior to etidronate. Cross-sectional analyses further showed that phytate intake above 307 mg/day was associated with normal bone mineral density levels in the lumbar spine of postmenopausal women (83).
Clinical interventions have provided additional support for phytate’s bone-protective properties. Oral supplementation with calcium–magnesium phytate (InsP6) significantly reduced urinary calcium excretion and serum β-CrossLaps levels—a marker of bone resorption—in hypercalciuric patients with osteopenia or osteoporosis, suggesting that phytate may help prevent bone demineralization in individuals with excessive bone turnover (84).
Long-term fracture risk assessment has provided compelling evidence for phytate’s protective role. A clinical study evaluating the relationship between urinary phytate concentrations and 10-year fracture risk in recently postmenopausal women demonstrated that those with higher urinary phytate levels (≥1.0 mg/L) had significantly lower FRAX-predicted risks of major osteoporotic and hip fractures compared to women with low phytate levels (≤0.50 mg/L). This protective effect was particularly pronounced in women with established osteoporosis risk factors, such as tobacco or alcohol use, suggesting that higher intake of phytate-rich foods may reduce long-term fracture risk in at-risk populations (85).
While accumulating evidence supports the beneficial effects of phytic acid on bone health, its dual nature requires careful consideration. Recent comprehensive reviews have summarized both the beneficial and potentially adverse effects of phytic acid (IP6) on bone metabolism (86). Although traditionally considered an antinutrient due to its mineral-chelating capacity, IP6 has been shown to bind to hydroxyapatite crystal growth sites, inhibiting pathological calcification. However, this same mechanism raises legitimate concerns about possible interference with physiological bone mineralization processes. Current evidence predominantly supports a protective role of IP6 in bone health through modulation of both extracellular crystallization and intracellular signaling pathways, but the balance between beneficial and potentially harmful effects may depend on dosage, timing, and individual physiological conditions (86).
Understanding this complexity becomes particularly important when considering food processing methods that can modify phytate levels. The breakdown of phytate is mediated by phytase enzymes, which are absent in humans and non-ruminant animals (87). These enzymes occur naturally in plants and microorganisms and exhibit optimal activity at pH levels near 5, though they remain active across pH ranges of 2 to 8 (88). The effectiveness of phytases in degrading phytate can be enhanced through various food processing methods, including cooking, soaking (89), germination (90), and fermentation (91, 92).
Among these methods, fermentation offers particular advantages for promoting bone health by simultaneously reducing pH and enhancing phytase activity (93, 94). The acidic environment created during fermentation not only activates phytases more effectively but also improves calcium absorption—a critical factor for maintaining skeletal integrity (95). For example, sourdough bread made from specific vegetable flours exhibits lower pH compared to original ingredients (96), resulting in up to 45% reduction in phytate content after baking (97). Similarly, vegetable fermentation processes, such as those used to produce kimchi or sauerkraut, reduce pH from approximately 3.9 to 4.4 (98).
The reduced phytate levels in fermented foods compared to raw components (73, 79) can significantly improve mineral bioavailability (Table 2). Studies have demonstrated increased calcium bioavailability in yogurt made from soy milk, with potential benefits for bone health (99). Animal studies have provided additional support, showing that ovariectomized rats fed fermented soybeans with Bacillus subtilis exhibited enhanced bone mineral density after 12 weeks (100). These findings suggest that dietary intake of appropriately fermented foods can help maintain bone health by optimizing the balance between phytate’s beneficial effects and mineral availability.
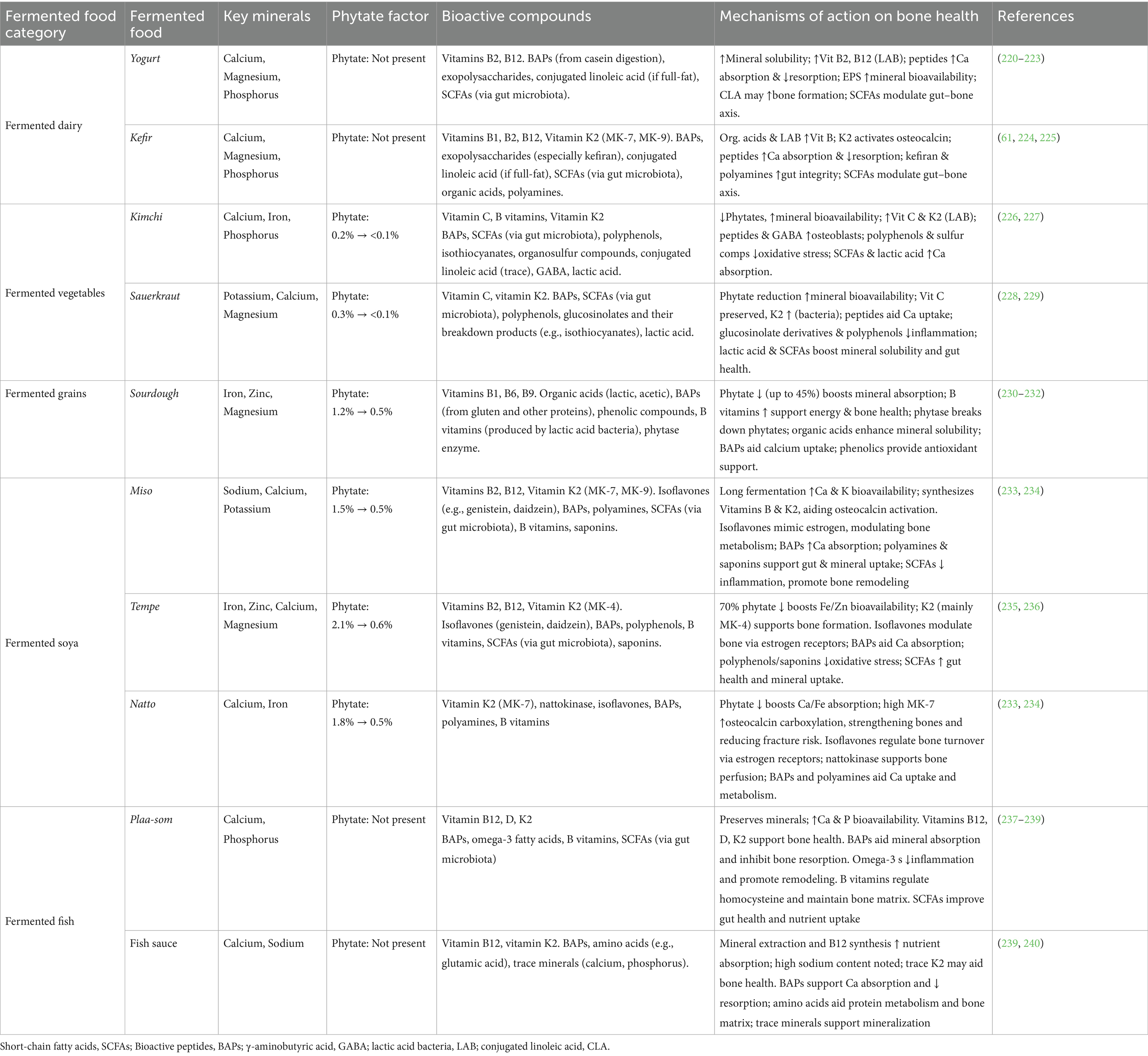
Table 2. Key fermented foods, their bioactive compounds, and mechanisms influencing bone health.
Despite accumulating evidence on the bone-protective effects of phytate, its dual role as both a mineral chelator and functional bioactive compound presents important challenges. While fermentation has emerged as a promising strategy to reduce phytate content and enhance mineral bioavailability, the precise balance between retaining phytate’s benefits and mitigating its antinutritional effects remains unclear. The long-term impact of fermented phytate-rich foods on bone health also requires further investigation. Most existing studies are short-term or observational, limiting conclusions on causality and durability of effects. Additionally, individual differences in phytate metabolism, fermentation efficacy, and dietary patterns further complicate outcome consistency. Interactions between fermented foods and other nutrients, as well as the potential role of gut microbiota in modulating phytate activity post-ingestion, are underexplored areas warranting deeper investigation.
Given the phytic acid dual role as both a mineral chelator and a functional bioactive, it offers a compelling starting point to illustrate how fermentation transforms food components in ways that directly influence bone health. Its modulation during fermentation highlights one of the most fundamental biochemical mechanisms, enhanced mineral bioavailability that underpins the relevance of fermented foods in skeletal support. Building on this foundation, we next explore the role of other key bioactive compounds influenced by fermentation, including vitamins K and B complex, polyphenols, and bioactive peptides. Each contributes to bone health through distinct but complementary pathways, ranging from nutrient signaling and antioxidant activity to hormonal modulation and cellular communication. Finally, these effects converge through the gut-bone axis, where the gut microbiota plays a central role in mediating the systemic impact of fermented foods on bone metabolism.
Vitamins K and B complex for bone health
Vitamin K is essential for maintaining bone health (101), which occurs in two varieties: vitamin K1 (phylloquinone) and vitamin K2 (menaquinones) (102). Vitamin K1 can be found in green leafy vegetables and certain plant oils (103), and vitamin K2 is synthetized by bacteria through fermentation or from animal-derived foods (104), which is critical for bone health (Table 2). For example, the level of vitamin K in natto, the Japanese fermented soybean food, with Bacillus subtilis, rises from 0.5 μg/g in soybeans to 10 μg/g in the finished product fermented with Bacillus subtilis (19). Vitamin K2 plays a crucial role in activating proteins referred to as Gla proteins, which encompass OC, one of the 17 Gla proteins found in humans. The activated variant of OC is significant for the development and preservation of bones (105). There are various forms of vitamin K2; for instance, the short-chain type, MK-4, is synthesized from vitamin K1 in the body and can be found in animal products. Longer-chain variants such as MK-7, MK-8, and MK-9 are synthetized by bacteria (106, 107) and are present in fermented milk foods like cheese (108, 109), as well as in fermented soybean products such as natto from Japan (110) and kinema, native fermented food from the Himalayas (111).
Many studies have explored the relationship between vitamin K intake and bone health. Some research shows inconsistent findings about how vitamin K supplements affect bone health markers (112, 113). Recent research, however, showed a clear link between low vitamin K intake and knee cartilage damage in people with severe knee pain and osteoarthritis, especially among women (114). Additionally, a higher dietary intake of vitamin K was linked to better bone quality and greater BMD (115). Recent research has shown that vitamin K2 supplements can help postmenopausal status by reducing bone loss and enhancing bone quality in Japanese women (116–118).
Numerous studies have investigated the relationship between vitamin K2 sourced from natto and bone health. The effects of vitamin K1 supplements containing MK-7, which is present in natto extract, demonstrated that MK-7 aids in the improved carboxylation of OC (110). Habitual intake of natto is linked to a reduced risk of osteoporotic fractures in Japanese postmenopausal women, independently of bone mineral density (119). Additionally, natto consumption is indirectly associated with lower rates of tooth loss—possibly via improved systemic bone density (120). Collectively, current evidence supports recommending natto to help prevent bone loss in both premenopausal and postmenopausal Japanese women (121, 122). Lundberg et al. (109) examined the influence of Jarlsberg and Camembert cheeses on bone health. They discovered that Jarlsberg cheese, which contains long-chain vitamin K2, enhances total serum OC levels, increases carboxylated OC, and improves the OC ratio, thus fostering bone growth. Within every 100 g of Jarlsberg cheese, there exists 3.0 μg of vitamin K1, 5.2 μg of MK-4, and 1.5 μg of MK (109).
Vitamin K is well-recognized for its role in maintaining bone density as individuals age; however, emerging research also highlights the importance of B vitamins in relation to osteoporosis and fracture risk (123). For instance, one study found that higher intake of vitamin B2 is significantly associated with a reduced risk of femoral osteoporosis and bone loss (124). In a large cohort of US women over the age of 50, elevated levels of homocysteine and methylmalonic acid, biomarkers of poor B-vitamin status, were linked to lower bone mineral density and an increased risk of lumbar spine osteoporosis, despite the widespread fortification of foods with B vitamins (125). These findings emphasize the role of adequate B-vitamin status in supporting bone health and reducing fracture risk. Additionally, a study involving 63,257 adults aged 45 to 74 years reported that higher pyridoxine (vitamin B6) intake was associated with a lower risk of osteoporosis. While several investigations have explored the potential benefits of B-complex vitamins on low bone mineral density, some studies suggest that these vitamins may not directly improve BMD outcomes (125, 126). A positive correlation exists between water-soluble vitamins and BMD, primarily associated with vitamins B12, and C. Conversely, a negative association was observed with fat-soluble vitamins, especially vitamins E and A (126).
Despite the growing body of literature on the role of vitamins K and B in bone health, other researchers have not found an association of unspecified vitamin K intake and bone fracture risk, (e.g., 127) did not find any association between K1 or K2 intake and vertebral fractures in the Hordaland Health Study of 2,994 Chinese community-dwelling men and women aged 65 years and over in Hong Kong. Moreover, results from other studies showed an insignificant effect of vitamin K on lumbar spine and femoral neck BMD (128).
while B-vitamin deficiencies, especially B6, B12, and folate, are linked to elevated homocysteine levels and reduced BMD, causal evidence from intervention trials remains inconclusive. Results from several in vitro and in vivo studies were generally consistent and showed that, although B vitamin deficiency generated significantly higher serum level of homocysteine, there was no significant effect on bone strength and bone area, mineral matrix, callus stiffness, size or tissue composition or bone turnover (129). Similarly, In the Singapore Chinese Health Study, dietary intake of B1 and B3 was not associated with risk of hip fracture in either men or women (130).
The results from observational studies are inconsistent in the associations between K and B vitamins and bone outcomes, however, majority of the randomized clinical trials have not shown protective effects of these vitamins in bone turnover or fracture risk reduction. Hence, while observational and mechanistic studies suggest a potential role for vitamins K and B in bone health, current evidence from randomized clinical trials remains insufficient to support strong clinical recommendations, highlighting the need for well-designed, long-term intervention studies to clarify their therapeutic relevance.
Polyphenols for bone health
In fermented plant-based foods, such as whole-grains, vegetables, and fruits, polyphenols are the main bioactive compounds (131). Generally categorized as flavonoids or non-flavonoids, polyphenols represent a group of secondary metabolites present in plants that exhibit various biological activities (132). According to their structural features, flavonoids can be divided into flavanols, flavones, flavanones, chalcones, and isoflavones (133). Non-flavonoids consist of lignans, astragalus, tannins, and phenolic acids, among others (131).
Numerous studies have reported the beneficial influence of polyphenols on the prevention and treatment of bone health. Overall, the intake of polyphenols may aid in preventing bone loss and decrease fracture risk (134), providing a potential strategy for preventing and managing osteoporosis by impacting bone metabolism, decreasing bone resorption, sustaining bone density, and diminishing osteoclast differentiation (3), while also fostering bone formation and hindering bone resorption (135).
Li et al. provided a theoretical framework for the mechanisms of polyphenol compounds related to bone formation and absorption (136). Polyphenols provide advantageous effects on osteoporotic bone defects by regulating oxidative stress, reducing inflammation, promoting osteogenesis, inhibiting osteoclast differentiation, and inducing osteoclast apoptosis (137).
The fermentation of food is crucial for increasing the beneficial impacts of polyphenols in food (Table 2), as polyphenols that aren’t fermented cannot be absorbed by the intestines, significantly limiting their bioavailability (138). Fermentation enhances the bioaccesibility and bioavailability of polyphenols by transforming intricate polyphenols in food into more straightforward, absorbable forms via beneficial microorganisms and enzymes related to polyphenols (131). These small-molecule free phenols exhibited greater biological activities and bioavailability compared to the macromolecular-bound phenols (131).
In accordance with the aforementioned the results indicate that the total polyphenols amount increases by 30.3% during spontaneous fermentation in the production process of cheonggukjang, a fermented soybean food of Korea (139), and in vinegar made from black wolfberry by 42.9% (140). During the fermentation of cheonggukjang, the total content of flavonols and phenolic acids rises due to the hydrolysis process that converts isoflavone glycosides into aglycones (141). Following a short fermentation period, the levels of aglycone-type isoflavones (daidzein, glycitein, and genistein) increased, while the levels of glycoside-type, malonyl-, and acetyl-type isoflavones decreased (142). This evidence verifies that cheonggukjang serves as a significant source of isoflavone aglycones like daidzein and genistein, even with the brief fermentation duration of soybeans (141). The effectiveness of cheonggukjang in preventing bone mass loss due to osteoporosis was demonstrated by observing reduced bone length and loss in animals fed a diet containing 10% cheonggukjang over a period of 15 weeks (143). According to (144), total isoflavone glycosides decreased and total isoflavone aglycones increased in 70% methanol extracts of three types of standardized chungkookjang inoculated with 1% (v/w) B. subtilis, B. licheniformis, or B. amyloliquefaciens in comparison with unfermented cooked soybeans.
Also, according to the prior research, the bioavailability and bioactivity of isoflavonoids are increased with fermentation, since in vivo studies have shown that isoflavone aglycones (daidzein and genistein) were absorbed more effectively in the human intestine by consuming tempe, Indonesian fermented soybean food, as opposed to a soybean-pieces diet (145, 146). Recently, soy pulp, which is a by-product of black soybean food processing, has been utilized as animal feed or discarded as waste. Among fermented black soybean products, fermented black soybean pulp supports bone metabolism and osteoporosis prevention (147). The research conducted by (147) indicated that genistein levels in black soybean pulp after 12 and 24 h of fermentation with L. acidophilus were 6.8 and 7.2 times higher, respectively, in comparison to controls. One group of ovariectomized rats was treated with black soybean pulp, while a second group of ovariectomized rats was treated with fermented black soybean pulp, indicating that the fermentation of black soy pulp is effective in preventing osteoporosis in ovariectomized rats (147). Finally, numerous studies have shown that (−)-epigallocatechin-3-gallate (EGCG), (−)-epicatechin gallate (ECG), and epigallocatechin (EGC) present in fermented tea are the main compounds that influence bone mass and prevent bone loss (148).
Recent intervention studies investigating the effects of polyphenols, consumed via food or given as isolated compounds, showed inconsistent results regarding bone health (Table 2). While some studies suggest that dietary intervention with polyphenol rich foods may be useful to prevent the incidence and progression of osteoporosis (149), on the other hand, evidence from human intervention studies does not allow a clear conclusion on the effects of dietary polyphenols on bone mineral density and bone turnover markers (150). Consequently, conclusive determinations cannot be reached. The current evidence of polyphenols’ impact on bone health prevention and treatment is based on numerous review articles that address findings from previous decades. The understanding of polyphenols’ effects related to bone health is limited due to the lack of evidence in recent research.
Alongside polyphenols, fermentation also generates bioactive peptides with demonstrated potential to influence bone remodeling. These peptides derived mainly from dairy and soy protein hydrolysis, function through additional pathways such as calcium binding, hormone modulation, and direct stimulation of osteoblast activity, complementing the effects of both polyphenols and vitamins.
Bioactive peptides for bone health
Bioactive peptides (BAPs) are initially encoded within their parent proteins and are released through enzymatic hydrolysis during fermentation and/or digestion (151). BAPs possess various beneficial effects on bone health (71, 152). Fermented foods are recognized for containing BAPs that are derived from the fermentation process by the actions of bacteria such as LAB, along with some Bacillus species (153–155). Nonetheless, there is minimal research regarding how these BAPs influence bone health.
In dairy products, the primary proteins acting as sources for generating BAPs are caseins and whey proteins (156). These peptides typically emerged from the proteolytic activity of LAB (157–160). Employing an untargeted peptidomics approach, researchers discovered BAPs exhibiting antioxidant, antihypertensive, and antidiabetic properties after fermenting whey protein concentrate with strains of Lactobacillus helveticus and Streptococcus thermophilus (161). Specific peptide sequences such as Ile-Pro-Pro and Val-Pro-Pro, which are liberated by the breakdown of milk casein using various strains of Lb. helveticus, are recognized to be effective angiotensin-converting enzyme (ACE) inhibitors and possess antihypertensive effects (162). The impact of lowering blood pressure may also have repercussions for bone health. Elevated blood pressure has been linked to decreased bone density and loss of bone minerals in women at risk for osteoporosis (163, 164).
Casein phosphopeptides (CPPs) are distinctive peptides present in fermented milk and cheese, which are released through the degradation of proteins by enzymes from different types of bacteria (165). Studies on the hydrolysis of β-, αs1-, and αs2-caseins by various strains of S. thermophilus have demonstrated that numerous CPPs are released (166). CPPs may have a significant role in bone health by interacting with metal ions such as iron, calcium, and zinc. This interaction can improve the absorption and dissolution of these minerals (167). Consequently, enhanced calcium absorption from CPPs could contribute to better bone health. In research involving animals, CPPs have also been shown to affect the activity of bone-forming cells (152, 168). CPPs can encourage the growth and maturation of osteoblasts (169). The casein-derived peptide valyl-prolyl-proline, produced during fermentation with Lb. helveticus, had a beneficial impact on bone turnover in rats (170). Lee et al. (171) indicated that milk products fermented with Lactobacillus plantarum A41 and Lactobacillus fermentum SRK414 had anti-osteoporotic benefits on post-menopausal rats by regulating markers associated with bone metabolism. Furthermore, it has been hypothesized that milk-derived peptides with antioxidative or anti-inflammatory activities might affect the signaling pathways engaged in bone remodeling (59).
Kefir is a fermented dairy beverage that includes BAPs generated during its fermentation (172). The impact of kefir peptides (KPs) on bone health has been investigated in multiple studies (Table 2). Chang et al. (173) reported a decrease in pro-inflammatory cytokines present in the serum (IL-1β, IL-6, TNF-α), and markers of bone resorption (CTX-1, RANKL), while enhanced serum indicators of bone formation (P1NP, OPG, OC), effectively averting bone loss in mice with osteoporosis. Additionally, Lai et al. (31) demonstrated that KPs powder aided in fracture recovery by stimulating the development of bone-forming cells in rats. Similarly, Tu et al. (174) reported an increased bone density in mice. A six-month study involving 40 osteoporotic participants evaluated the effects of kefir-fermented milk (1,600 mg) along with calcium bicarbonate (CaCO3, 1,500 mg) on bone metabolism. Those who ingested kefir-fermented milk exhibited significant enhancements in BMD as determined by dual-energy X-ray absorptiometry. Patients whose T-scores were above −1 saw a substantial decline in the serum β C-terminal telopeptide of type I collagen (β-CTX), and serum OC transitioned from negative to positive post-treatment (66). Nonetheless, the specific effects of KPs remain ambiguous in this study.
Some fermented soybean products include a peptide that might have a beneficial impact on bone health by improving calcium absorption (175) (Table 2). Poly-γ-glutamic acid (PGA), a sequence of glutamic acids linked by γ-bonds (176), is present in B. subtilis-fermented items, such as natto (177) and kinema (178). It has been shown that PGA enhances calcium absorption in the intestines of rats. A single administration of PGA resulted in elevated soluble calcium levels in the small intestine (179). These researchers suggested that PGA aids in the creation of a soluble calcium-binding complex, which boosts calcium solubility in the lower small intestine. In humans, Tanimoto et al. (180) examined PGA in a single-blind, randomized, crossover trial, assessing calcium absorption using a double stable isotope technique. They discovered that ingesting two doses of calcium-rich orange juice (200 mg Ca/200 g) and PGA (60 mg/200 g) over a period of 3 to 4 weeks enhanced calcium absorption in healthy post-menopausal women compared to participants who consumed only calcium-fortified orange juice. However, the study had drawbacks, such as a lack of comprehensive details regarding the age, weight, height, and diet of the participants (181). At present, the evidence supporting the function of PGA in enhancing bone health is scarce, and additional research is required to comprehend how it might increase calcium absorption.
Various studies utilizing rodent models have investigated how peptides derived from fermented foods may support bone health. Although peptides were included in these analyses, it is difficult to assert that the impacts on bone metabolism are exclusively attributable to these compounds. For example, Chiang et al. (182) discovered that a 6-week treatment with lactobacilli-fermented soy skim milk aided in diminishing bone loss associated with aging in 13-month-old mice. They proposed that the beneficial effects may arise from both the peptides and other elements such as isoflavones and polysaccharides. BAPs found in fermented dairy products can assist in enhancing bone health by increasing the expression of genes linked to cell growth and the formation of bone cells known as osteoblasts through a specific cellular signaling pathway, which elevates the levels of vital markers for osteoblast formation, including runt-related transcription factor 2 (RUNX2), osteocalcin (OCN), alkaline phosphatase (ALP), and collagen type I alpha 1 chain (COL1A1) (93). Another investigation examined the influence of water-soluble extracts from a long-fermented soybean product from Korea known as doenjang on bone metabolism (183). These extracts comprise small proteins generated from the breakdown of larger proteins during fermentation by microorganisms. The water-soluble extracts from doenjang facilitated the differentiation of osteoblasts by altering gene expression. Moreover, they enhanced the mineralization of osteoblasts in comparison to the control group. The decrease in tartrate-resistant acid phosphatase activity among cells treated with these extracts suggested that the process of bone resorption by osteoclasts was inhibited. Further studies are required to pinpoint the specific water-soluble peptides that contribute to these effects. It is important to recognize that the biological activity of peptides can be influenced by numerous factors, including the kind of protein, the enzymes utilized, the preparation of the proteins, the proportion of substrate to enzyme, the duration, temperature, pH level, the extent of protein breakdown, the peptide structures, the kinds of amino acids, their molecular weights, and among others (161).
Many of these bioactive compounds whether vitamins, peptides, or polyphenols, interact with the host not only directly but also indirectly through the gut microbiota. This emerging axis, linking dietary components to bone metabolism via microbial modulation, serves as a unifying mechanism that integrates the effects of fermented foods across multiple domains of bone health.
Gut microbiota for bone health
The human gut microbiota is a diverse community of a high number of microorganisms including bacteria, viruses, fungi, and archaea, residing primarily in the gastrointestinal tract. These microbes play essential roles in metabolism (aiding digestion, extracting nutrients, synthesizing vitamins) such as K and B group, and producing SCFAs; barrier function (maintaining the structural integrity of the gut lining and protecting against pathogen colonization); immune regulation (modulating immune responses, promoting immune tolerance, and influencing systemic inflammation); and neurological effects (affecting the gut-brain axis, impacting cognition, behavior, and neurological health) (72, 184, 185) (Table 2). A balanced gut microbiota is fundamental for overall health, while disturbances, termed dysbiosis, are linked to metabolic, immune, neurodegenerative, and gastrointestinal diseases (72, 184). Emerging research has shown a strong connection between gut microbiota and bone health. This connection has been explained through several mechanisms which have been highlighted in this section. Additionally, interventions altering the gut microbiome such as through diet, prebiotics, or probiotics may positively influence bone density and strength, although findings are still preliminary and sometimes inconsistent (186).
The gut microbiota can enhance bone health through various mechanisms, such as immune responses, hormone synthesis, producing beneficial compounds like SCFAs, aiding in the absorption of nutrients like calcium, and communicating with the brain through neurotransmitter production (187). The microorganisms present in fermented foods can function as probiotics, including Lactobacillus and Bifidobacterium, generate beneficial peptides and compounds, and convert phenolic compounds into more active forms that may support bone health (188). Fermented foods contain specific microbial groups, especially LAB, that engage with the gut microbiome and can positively influence bone metabolism following consumption (40, 66, 174, 189, 190). Changes in the types or numbers of microorganisms present in the gut can result in dysbiosis, which leads to inflammation in the intestines and modifies gut health (191). When inflammation occurs in the intestines, Th17 cells become activated, resulting in the release of pro-inflammatory substances such as TNF-α, IL-1β, IL-6, and IL-17. Marahleh et al. (192) indicated that these pro-inflammatory substances elevate the levels of receptor activator of NF-κB (RANK) ligand (RANKL). RANKL is crucial for the activation of osteoclasts and contributes to bone loss (193, 194). Out of these substances, interleukin 1β is among the most potent pro-inflammatory agents, greatly enhancing bone loss in both laboratory and living models by elevating RANKL levels (195). The activation of TLR5 increases the ratio of RANKL to osteoprotegerin (OPG), which is a natural inhibitor of RANKL, in cells responsible for bone formation, resulting in the production of more osteoclasts and accelerated bone loss (196). RANKL promotes osteoclast maturation by binding to RANK on the surface of osteoclast precursors and stimulates their conversion into osteoclasts that destroy bone (192, 196). It has been demonstrated that diets rich in fermented foods have been demonstrated to enhance bone health by influencing bone formation, owing to the gut bacteria capable of surviving in the digestive tract and interacting with other gut bacteria, which may contribute to improved bone health (197). Despite substantial evidence linking bone health to gut bacteria, many facets of osteoimmunology remain to be thoroughly investigated.
Additionally, gut bacteria might be related to bone health via the endocrine system (198). The initial substance identified that connects the gut and bones was the insulin-like growth factor 1 (IGF-1), primarily produced in the liver in response to food intake and influenced by gut microbiota and specific microbes, such as Lb. plantarum, found in fermented foods (197, 199). The IGF-1 assists in bone growth and remodeling (200). There is a possibility that bacteria influence bone health through SCFAs, which are generated when bacteria metabolize fiber, can also result in increased IGF-1 (201). In addition to IGF-1, gut bacteria have been indicated to influence other hormones associated with bone health, including serotonin, parathyroid hormone, glucagon-like peptide 1, and leptin (202). Hydrogen sulfide emerges as another significant factor in microbial endocrinology that regulates bone metabolism (203). Specific intestinal bacteria, such as Escherichia, Fusobacterium, Desulfovibrio, Streptococcus, Klebsiella, Clostridium, Salmonella, and Enterobacter, convert cysteine into hydrogen sulfide, pyruvate, and ammonia (196, 204, 205). Hydrogen sulfide, a new metabolite generated by gut bacteria, originates from cysteine, homocysteine, and sulfate-reducing bacteria (206), may involve in bone marrow mesenchymal stem cells (207). When there is too little hydrogen sulfide, it disrupts stem cell differentiation, causing irregular calcium flow within the cells and disrupting signaling pathways that are crucial for bone formation (207). Sodium hydrosulfide, a standard source of hydrogen sulfide, was found to reduce RANKL/OPG mRNA levels in human mesenchymal stem cells, which play a role in supporting osteoclasts (208).
Despite the abovementioned findings, it is crucial to note that a recent systematic review has shown that targeting the gut microbiota did not show consistent improvements in bone health. Some studies report positive changes in bone density or strength, while others find no significant effect (186). You et al. have studied relationship between age-dependent microbial change and age-related bone loss in mice and reported that the bone loss associated with age was not dependent on the gut microbiome (209). This directly challenges prior models suggesting microbiota alterations underlie age-associated skeletal deterioration. These findings indicate that there is a need to conduct large longitudinal human cohorts with multi omics, standardized methods, identification of causal microbial strains/metabolites, and assessment of long-term safety and bidirectional signaling.
Conclusion and future research prospects
Bone health is a multifactorial condition influenced by a combination of dietary components, gut microbiota, and host metabolic processes. This review examined how various bioactive compounds found in fermented foods such as vitamins (K and B groups), polyphenols, bioactive peptides, and fermentation-modified phytates may contribute to bone metabolism through multiple mechanisms (Table 2). These include enhancement of calcium and mineral bioavailability, modulation of oxidative stress and inflammation, and regulation of bone cell signalling pathways involved in osteoblast and osteoclast activity. Additionally, the interplay between fermented foods and gut microbiota particularly the production of SCFAs and immunomodulatory metabolites suggests a potentially important role of the gut-bone axis in skeletal homeostasis.
Despite these findings, the evidence base for fermented foods as modulators of bone health remains less developed than that for traditional approaches such as calcium and vitamin D supplementation or pharmacological interventions. While some preclinical and population studies report associations between fermented food intake and improved bone mineral density or reduced fracture risk, the bioavailability and efficacy of such food-derived compounds are not yet well quantified in humans (40, 210, 211). Moreover, the dose response relationship and long-term impact of fermented food consumption on clinically relevant bone outcomes remain unclear.
A critical gap in the current literature is the limited number of well-designed human clinical trials that evaluate fermented foods in comparison to, or in combination with, conventional bone health interventions. The variability in microbial strains, food matrices, and fermentation processes further complicates the interpretation of findings and the development of standardized dietary recommendations (212–214). Furthermore, while the mechanistic roles of isolated nutrients such as vitamin K2 or polyphenols have been described, few studies investigate the synergistic or antagonistic effects that may occur within complex fermented food systems.
Recent developments in microbiome research and nutritional metabolomics highlight the need for a more personalized approach. Emerging data suggest that individual responses to fermented foods may vary depending on host genetics, baseline nutrient status, and gut microbial composition (215). Additionally, microbial-derived postbiotics such as SCFAs, indoles, or peptidoglycans have been shown to influence bone remodelling pathways, offering a novel direction for future investigation (216). The potential application of synbiotic formulations (fermented foods combined with prebiotics or probiotics) also warrants further study, particularly in populations at high risk for osteoporosis, such as postmenopausal women (217, 218).
To advance this field, future research should focus on establishing the bioavailability and efficacy of fermented food components compared to conventional treatments, identifying strain or product-specific effects on bone health, and evaluating the long-term impact of fermented food consumption on bone quality and fracture outcomes through large scale, controlled human studies. Moreover, interdisciplinary efforts are needed to integrate insights from nutrition science, microbiology, endocrinology, and clinical research to fully elucidate the mechanisms by which fermented foods contribute to skeletal health. Future research should prioritize several critical areas to advance the application of fermented foods in bone health management. First, large-scale randomized controlled trials (RCTs) are essential to confirm the effects of specific fermented foods or bioactives, such as vitamin K2 (MK-7), casein-derived peptides, or fermented soy isoflavones on BMD, turnover markers, and fracture outcomes in humans. Second, stratified analyses based on age, sex, menopausal status, gut microbiota composition, and baseline nutrient levels are needed to identify subpopulations that may benefit most from targeted interventions. Third, comparative studies are warranted to evaluate the synergistic effects of multiple bioactives within complex food matrices versus isolated compounds. Additionally, future investigations should integrate multi-omics approaches (e.g., metagenomics, metabolomics, transcriptomics) to decipher the role of the gut microbiota in mediating host responses to fermented food intake (18).
Technological advances in fermentation also open opportunities for developing functional foods with optimized microbial strains to deliver higher concentrations of bone-beneficial metabolites. Regulatory challenges around standardization and health claims, especially in functional or fortified fermented products, should be addressed to facilitate clinical translation and policy guidance. Furthermore, traditional fermented food systems from regions like India, Japan, Korea, and Africa represent a rich but underexplored source of bioactive compounds, deserving deeper study and possible integration into global dietary strategies (43, 219). Ultimately, bridging preclinical and clinical research through collaborative, interdisciplinary, and longitudinal studies will be essential to unlock the full therapeutic potential of fermented foods in bone health maintenance and osteoporosis prevention.
In conclusion, while fermented foods represent a promising and culturally diverse dietary strategy for supporting bone health, their clinical utility remains to be clearly established. This review underscores the need for targeted research to validate their effectiveness, explore their interaction with the gut microbiome, and position them within the broader context of dietary and therapeutic approaches to osteoporosis prevention and management.
The evidence gathered in this review suggests that incorporating fermented foods into the diet may be a beneficial strategy to complement bone health through natural food-based approaches. However, these effects should be viewed as supportive and not as substitutes for clinically established bone health interventions, such as calcium and vitamin D supplementation or pharmacological treatments when indicated.
Author contributions
JT: Writing – original draft, Validation, Conceptualization. FA: Writing – review & editing. BY: Writing – review & editing. İK: Writing – review & editing. JM: Writing – review & editing. TB-T: Writing – review & editing. AT: Writing – review & editing. DNN: Writing – review & editing. M-LK: Conceptualization, Writing – review & editing. JM-R: Writing – review & editing, Supervision, Conceptualization. MP: Writing – review & editing. BM: Validation, Writing – review & editing, Supervision, Conceptualization. JF: Validation, Conceptualization, Supervision, Writing – review & editing, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This review was supported by COST ACTION-PIMENTO CA20128 by the European Cooperation in Science and Technology (www.cost.eu). This research revision has been carried out under the Cost Action CA20128 “Promoting the Innovation in Fermented Foods, PIMENTO funded by the European Union.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Tamang, JP, and Thapa, N. Beneficial microbiota in ethnic fermented foods and beverages In: FJ Bruijn, H Smidt, LS Cocolin, M Sauer, D Dowling, and L Thomashow, editors. Good microbes in medicine, food production, biotechnology, bioremediation, and agriculture. Hoboken, New Jersey, USA: Wiley (2022). 130–48.
2. Bittante, G, Amalfitano, N, Bergamaschi, M, Patel, N, Haddi, M-L, Benabid, H, et al. Composition and aptitude for cheese-making of milk from cows, buffaloes, goats, sheep, dromedary camels, and donkeys. J Dairy Sci. (2022) 105:2132–52. doi: 10.3168/jds.2021-20961
3. Tamang, JP, Cotter, PD, Endo, A, Han, NS, Kort, R, Liu, SQ, et al. Fermented foods in a global age: east meets west. Comp Rev Food Sci Food Safe. (2020) 19:184–217. doi: 10.1111/1541-4337.12520
4. Narzary, Y, Das, S, Goyal, AK, Lam, SS, Sarma, H, and Sharma, D. Fermented fish products in south and southeast Asian cuisine: indigenous technology processes, nutrient composition, and cultural significance. J Ethn Food. (2021) 8:33. doi: 10.1186/s42779-021-00109-0
5. Obafemi, YD, Oranusi, SU, Ajanaku, KO, Akinduti, PA, Leech, J, and Cotter, PD. African fermented foods: overview, emerging benefits, and novel approaches to microbiome profiling. npj Sci Food. (2022) 6:15. doi: 10.1038/s41538-022-00130-w
6. Agyei, D, Owusu-Kwarteng, J, Akabanda, F, and Akomea-Frempong, S. Indigenous African fermented dairy products: processing technology, microbiology and health benefits. Crit Rev Food Sci Nutr. (2020) 60:991–1006. doi: 10.1080/10408398.2018.1555133
7. Narvhus, JA, and Abrahamsen, RK. Traditional and modern Nordic fermented milk products: a review. Int Dairy J. (2023) 142:105641. doi: 10.1016/j.idairyj.2023.105641
8. Van Reckem, E, Geeraerts, W, Charmpi, C, Van Der Veken, D, De Vuyst, L, and Leroy, F. Exploring the link between the geographical origin of European fermented foods and the diversity of their bacterial communities: the case of fermented meats. Front Microbiol. (2019) 10:2302. doi: 10.3389/fmicb.2019.02302
9. Capozzi, V, Fragasso, M, Romaniello, R, Berbegal, C, Russo, P, and Spano, G. Spontaneous food fermentations and potential risks for human health. Fermentation. (2017) 3:49. doi: 10.3390/fermentation3040049
10. Tamang, JP, Watanabe, K, and Holzapfel, WH. Review: diversity of microorganisms in global fermented foods and beverages. Front Microbiol. (2016) 7:377. doi: 10.3389/fmicb.2016.00377
11. Kim, D-H, Jeong, D, Song, K-Y, and Seo, K-H. Comparison of traditional and backslopping methods for kefir fermentation based on physicochemical and microbiological characteristics. LWT. (2018) 97:503–7. doi: 10.1016/j.lwt.2018.07.023
12. Comunian, R, and Chessa, L. Development and application of starter cultures. Fermentation. (2024) 10:512. doi: 10.3390/fermentation10100512
13. Deveci, G, Çelik, E, Ağagündüz, D, Bartkiene, E, Rocha, JMF, and Özogul, F. Certain fermented foods and their possible health effects with a focus on bioactive compounds and microorganisms. Fermentation. (2023) 9:923. doi: 10.3390/fermentation9110923
14. Abbaspour, N. Fermentation’s pivotal role in shaping the future of plant-based foods: an integrative review of fermentation processes and their impact on sensory and health benefits. Appl Food Res. (2024) 4:100468. doi: 10.1016/j.afres.2024.100468
15. Khayatan, D, Nouri, K, Momtaz, S, Roufogalis, BD, Alidadi, M, Jamialahmadi, T, et al. Plant-derived fermented products: An interesting concept for human health. Current Develop Nutrition. (2024) 8:102162. doi: 10.1016/j.cdnut.2024.102162
16. Shi, H, An, F, Lin, H, Li, M, Wu, J, and Wu, R. Advances in fermented foods revealed by multi-omics: a new direction toward precisely clarifying the roles of microorganisms. Front Microbiol. (2022) 13:1044820. doi: 10.3389/fmicb.2022.1044820
17. Wen, L, Yang, L, Chen, C, Li, J, Fu, J, Liu, G, et al. Applications of multi-omics techniques to unravel the fermentation process and the flavor formation mechanism in fermented foods. Crit Rev Food Sci Nutr. (2024) 64:8367–83. doi: 10.1080/10408398.2023.2199425
18. Walsh, AM, Leech, J, Huttenhower, C, Delhomme-Nguyen, H, Crispie, F, Chervaux, C, et al. Integrated molecular approaches for fermented food microbiome research. FEMS Microbiol Rev. (2023) 47:fuad001. doi: 10.1093/femsre/fuad001
19. Li, Q, Li, N, Cai, W, Xiao, M, Liu, B, and Zeng, F. Fermented natural product targeting gut microbiota regulate immunity and anti-inflammatory activity: a possible way to prevent COVID-19 in daily diet. J Funct Foods. (2022) 97:105229. doi: 10.1016/j.jff.2022.105229
20. Leeuwendaal, NK, Stanton, C, O’Toole, PW, and Beresford, TP. Fermented foods, health and the gut microbiome. Nutrients. (2022) 14:1527. doi: 10.3390/nu14071527
21. Balasubramanian, R, Schneider, E, Gunnigle, E, Cotter, PD, and Cryan, JF. Fermented foods: harnessing their potential to modulate the microbiota-gut-brain axis for mental health. Neurosci Biobehav Rev. (2024) 158:105562. doi: 10.1016/j.neubiorev.2024.105562
22. Chan, M, Baxter, H, Larsen, N, Jespersen, L, Ekinci, EI, and Howell, K. Impact of botanical fermented foods on metabolic biomarkers and gut microbiota in adults with metabolic syndrome and type 2 diabetes: a systematic review protocol. BMJ Open. (2019) 9:e029242. doi: 10.1136/bmjopen-2019-029242
23. Yilmaz, B, Elibol, E, Shangpliang, HNJ, Ozogul, F, and Tamang, JP. Microbial communities in home-made and commercial kefir and their hypoglycemic properties. Fermentation. (2022) 8:590. doi: 10.3390/fermentation8110590
24. Li, KJ, Burton-Pimentel, KJ, Vergères, G, Feskens, EJM, and Brouwer-Brolsma, EM. Fermented foods and cardiometabolic health: definitions, current evidence, and future perspectives. Front Nutr. (2022) 9:976020. doi: 10.3389/fnut.2022.976020
25. Ibrahim, SA, Gyawali, R, Awaisheh, SS, Ayivi, RD, Silva, RC, Subedi, K, et al. Fermented foods and probiotics: An approach to lactose intolerance. J Dairy Res. (2021) 88:357–65. doi: 10.1017/S0022029921000625
26. Paul, AK, Lim, CL, Apu MdAI,, Dolma, KG, Gupta, M, De Lourdes Pereira, M, et al. Are fermented foods effective against inflammatory diseases? IJERPH. (2023) 20:2481. doi: 10.3390/ijerph20032481
27. Jalili, M, Nazari, M, and Magkos, F. Fermented foods in the Management of Obesity: mechanisms of action and future challenges. IJMS. (2023) 24:2665. doi: 10.3390/ijms24032665
28. Jung, H, Yun, Y-R, Hong, SW, and Shin, S. Association between kimchi consumption and obesity based on BMI and abdominal obesity in Korean adults: a cross-sectional analysis of the health examinees study. BMJ Open. (2024) 14:e076650. doi: 10.1136/bmjopen-2023-076650
29. Tian, Y, Li, F, Du, L, Peng, D, Yang, Z, Li, J, et al. Fermented fruits ameliorate obesity by controlling food intake and regulating lipid metabolism in high-fat dietary mice. J Funct Foods. (2024) 114:106072. doi: 10.1016/j.jff.2024.106072
30. Guo, X, Zhong, K, Zou, L, Xue, H, Zheng, S, Guo, J, et al. Effect of Lactobacillus casei fermented milk on fracture healing in osteoporotic mice. Front Endocrinol. (2022) 13:1041647. doi: 10.3389/fendo.2022.1041647
31. Lai, J-C, Li, H-P, Chang, GR-L, Lan, Y-W, Chen, Y-H, Tseng, Y-S, et al. Kefir peptides promote osteogenic differentiation to enhance bone fracture healing in rats. Life Sci. (2022) 310:121090. doi: 10.1016/j.lfs.2022.121090
32. Gad, AI, Orabi, MM, Abou-Taleb, KA, Abdelghani, DY, and Amin, SA. In vitro digestive system simulation and anticancer activity of soymilk fermented by probiotics and synbiotics immobilised on agro-industrial residues. Sci Rep. (2024) 14:18518. doi: 10.1038/s41598-024-68086-3
33. Nami, Y, Tavallaei, O, Kiani, A, Moazami, N, Samari, M, Derakhshankhah, H, et al. Anti-oral cancer properties of potential probiotic lactobacilli isolated from traditional milk, cheese, and yogurt. Sci Rep. (2024) 14:6398. doi: 10.1038/s41598-024-57024-y
34. Kumar, MR, Azizi, NF, Yeap, SK, Abdullah, JO, Khalid, M, Omar, AR, et al. Clinical and preclinical studies of fermented foods and their effects on Alzheimer’s disease. Antioxidants. (2022) 11:883. doi: 10.3390/antiox11050883
35. Wu, A-M, Bisignano, C, James, SL, Abady, GG, Abedi, A, Abu-Gharbieh, E, et al. Global, regional, and national burden of bone fractures in 204 countries and territories, 1990–2019: a systematic analysis from the global burden of disease study 2019. Lancet Healthy Longevity. (2021) 2:e580–92. doi: 10.1016/S2666-7568(21)00172-0
36. Shen, Y, Huang, X, Wu, J, Lin, X, Zhou, X, Zhu, Z, et al. The global burden of osteoporosis, low bone mass, and its related fracture in 204 countries and territories, 1990-2019. Front Endocrinol. (2022) 13:882241. doi: 10.3389/fendo.2022.882241
37. Smit, AE, Meijer, OC, and Winter, EM. The multi-faceted nature of age-associated osteoporosis. Bone Reports. (2024) 20:101750. doi: 10.1016/j.bonr.2024.101750
38. Salari, N, Ghasemi, H, Mohammadi, L, Behzadi, MH, Rabieenia, E, Shohaimi, S, et al. The global prevalence of osteoporosis in the world: a comprehensive systematic review and meta-analysis. J Orthop Surg Res. (2021) 16:609. doi: 10.1186/s13018-021-02772-0
39. Zhang, Y-W, Song, P-R, Wang, S-C, Liu, H, Shi, Z-M, and Su, J-C. Diets intervene osteoporosis via gut-bone axis. Gut Microbes. (2024) 16:2295432. doi: 10.1080/19490976.2023.2295432
40. Ong, AM, Kang, K, Weiler, HA, and Morin, SN. Fermented Milk products and bone health in postmenopausal women: a systematic review of randomized controlled trials, prospective cohorts, and case-control studies. Adv Nutr. (2020) 11:251–65. doi: 10.1093/advances/nmz108
41. Rizzoli, R, Biver, E, and Brennan-Speranza, TC. Nutritional intake and bone health. Lancet Diabetes Endocrinol. (2021) 9:606–21. doi: 10.1016/S2213-8587(21)00119-4
42. Savaiano, DA, and Hutkins, RW. Yogurt, cultured fermented milk, and health: a systematic review. Nutr Rev. (2021) 79:599–614. doi: 10.1093/nutrit/nuaa013
43. Valentino, V, Magliulo, R, Farsi, D, Cotter, PD, O’Sullivan, O, Ercolini, D, et al. Fermented foods, their microbiome and its potential in boosting human health. Microb Biotechnol. (2024) 17:e14428. doi: 10.1111/1751-7915.14428
44. Chu, X, Xing, H, Chao, M, Xie, P, and Jiang, L. Gut microbiota modulation in osteoporosis: probiotics, prebiotics, and natural compounds. Meta. (2025) 15:301. doi: 10.3390/metabo15050301
45. Šromová, V, Sobola, D, and Kaspar, P. A brief review of bone cell function and importance. Cells. (2023) 12:2576. doi: 10.3390/cells12212576
46. Lopes, D, Martins-Cruz, C, Oliveira, MB, and Mano, JF. Bone physiology as inspiration for tissue regenerative therapies. Biomaterials. (2018) 185:240–75. doi: 10.1016/j.biomaterials.2018.09.028
47. Price, CT, Langford, JR, and Liporace, FA. Essential nutrients for bone health and a review of their availability in the average north American diet. TOORTHJ. (2012) 6:143–9. doi: 10.2174/1874325001206010143
48. Serna, J, and Bergwitz, C. Importance of dietary phosphorus for bone metabolism and healthy aging. Nutrients. (2020) 12:3001. doi: 10.3390/nu12103001
49. Rondanelli, M, Faliva, MA, Tartara, A, Gasparri, C, Perna, S, Infantino, V, et al. An update on magnesium and bone health. Biometals. (2021) 34:715–36. doi: 10.1007/s10534-021-00305-0
50. Fleet, JC. Vitamin D-mediated regulation of intestinal calcium absorption. Nutrients. (2022) 14:3351. doi: 10.3390/nu14163351
51. Shkembi, B, and Huppertz, T. Calcium absorption from food products: food matrix effects. Nutrients. (2021) 14:180. doi: 10.3390/nu14010180
52. Kuroda, T, Uenishi, K, Ohta, H, and Shiraki, M. Multiple vitamin deficiencies additively increase the risk of incident fractures in Japanese postmenopausal women. Osteoporos Int. (2019) 30:593–9. doi: 10.1007/s00198-018-4784-2
53. Fratoni, V, and Brandi, M. B vitamins, homocysteine and bone health. Nutrients. (2015) 7:2176–92. doi: 10.3390/nu7042176
54. Tsugawa, N, and Shiraki, M. Vitamin K nutrition and bone health. Nutrients. (2020) 12:1909. doi: 10.3390/nu12071909
55. Matía-Martín, P, Torrego-Ellacuría, M, Larrad-Sainz, A, Fernández-Pérez, C, Cuesta-Triana, F, and Rubio-Herrera, MÁ. Effects of Milk and dairy products on the prevention of osteoporosis and osteoporotic fractures in Europeans and non-Hispanic whites from North America: a systematic review and updated Meta-analysis. Adv Nutr. (2019) 10:S120–43. doi: 10.1093/advances/nmy097
56. Guo, J, Givens, DI, and Heitmann, BL. Association between dairy consumption and cardiovascular disease events, bone fracture and all-cause mortality. PLoS One. (2022) 17:e0271168. doi: 10.1371/journal.pone.0271168
57. Duscha, A, Hegelmaier, T, Dürholz, K, Desel, C, Gold, R, Zaiss, MM, et al. Propionic acid beneficially modifies osteoporosis biomarkers in patients with multiple sclerosis. Ther Adv Neurol Disord. (2022) 15:17562864221103935. doi: 10.1177/17562864221103935
58. Jensen, MB, Daugintis, A, and Jakobsen, J. Content and bioaccessibility of vitamin K (Phylloquinone and Menaquinones) in cheese. Foods. (2021) 10:2938. doi: 10.3390/foods10122938
59. Bu, T, Zheng, J, Liu, L, Li, S, and Wu, J. Milk proteins and their derived peptides on bone health: biological functions, mechanisms, and prospects. Comp Rev Food Sci Food Safe. (2021) 20:2234–62. doi: 10.1111/1541-4337.12707
60. Levit, R, Savoy De Giori, G, Moreno De LeBlanc, A, and LeBlanc, JG. Recent update on lactic acid bacteria producing riboflavin and folates: application for food fortification and treatment of intestinal inflammation. J Appl Microbiol. (2021) 130:1412–24. doi: 10.1111/jam.14854
61. Tarvainen, M, Fabritius, M, and Yang, B. Determination of vitamin K composition of fermented food. Food Chem. (2019) 275:515–22. doi: 10.1016/j.foodchem.2018.09.136
62. Popova, A, and Mihaylova, D. Antinutrients in plant-based foods: a review. TOBIOTJ. (2019) 13:68–76. doi: 10.2174/1874070701913010068
63. Faridi, A, Gitoee, A, and France, J. A meta-analysis of the effects of nonphytate phosphorus on broiler performance and tibia ash concentration. Poult Sci. (2015) 94:2753–62. doi: 10.3382/ps/pev280
64. Dai, J, Tao, L, Shi, C, Yang, S, Li, D, Sheng, J, et al. Fermentation improves calcium bioavailability in Moringa oleifera leaves and prevents bone loss in calcium-deficient rats. Food Sci Nutr. (2020) 8:3692–703. doi: 10.1002/fsn3.1653
65. Bashir, HH, Hasnain, MA, Abbas, A, Lee, J-H, and Moon, G-S. The impact of fermented dairy products and probiotics on bone health improvement. Food Sci Anim Resour. (2025) 45:449–67. doi: 10.5851/kosfa.2025.e7
66. Tu, M-Y, Chen, H-L, Tung, Y-T, Kao, C-C, Hu, F-C, and Chen, C-M. Short-term effects of kefir-fermented Milk consumption on bone mineral density and bone metabolism in a randomized clinical trial of osteoporotic patients. PLoS One. (2015) 10:e0144231. doi: 10.1371/journal.pone.0144231
67. Bose, S, and Sharan, K. Effect of probiotics on postmenopausal bone health: a preclinical meta-analysis. Br J Nutr. (2024) 131:567–80. doi: 10.1017/S0007114523002362
68. Martiniakova, M, Babikova, M, Mondockova, V, Blahova, J, Kovacova, V, and Omelka, R. The role of macronutrients, micronutrients and flavonoid polyphenols in the prevention and treatment of osteoporosis. Nutrients. (2022) 14:523. doi: 10.3390/nu14030523
69. Aaseth, JO, Finnes, TE, Askim, M, and Alexander, J. The importance of vitamin K and the combination of vitamins K and D for calcium metabolism and bone health: a review. Nutrients. (2024) 16:2420. doi: 10.3390/nu16152420
70. Liu, D-Y, Wu, J, Zhou, H-Y, Lv, J-X, Cai, K-Z, and Tang, C-B. Phytic acid improves osteogenesis and inhibits the senescence of human bone marrow mesenchymal stem cells under high-glucose conditions via the ERK pathway. Chem Biol Interact. (2024) 387:110818. doi: 10.1016/j.cbi.2023.110818
71. Xu, Z, Zhang, R, Chen, H, Zhang, L, Yan, X, Qin, Z, et al. Characterization and preparation of food-derived peptides on improving osteoporosis: a review. Food Chemistry: X. (2024) 23:101530. doi: 10.1016/j.fochx.2024.101530
72. Zhang, Y, Wang, H, Sang, Y, Liu, M, Wang, Q, Yang, H, et al. Gut microbiota in health and disease: advances and future prospects. MedComm. (2024) 5:e70012. doi: 10.1002/mco2.70012
73. Sharma, N, Angural, S, Rana, M, Puri, N, Kondepudi, KK, and Gupta, N. Phytase producing lactic acid bacteria: cell factories for enhancing micronutrient bioavailability of phytate rich foods. Trends Food Sci Technol. (2020) 96:1–12. doi: 10.1016/j.tifs.2019.12.001
74. Pires, SMG, Reis, RS, Cardoso, SM, Pezzani, R, Paredes-Osses, E, Seilkhan, A, et al. Phytates as a natural source for health promotion: a critical evaluation of clinical trials. Front Chem. (2023) 11:1174109. doi: 10.3389/fchem.2023.1174109
75. Feizollahi, E, Mirmahdi, RS, Zoghi, A, Zijlstra, RT, Roopesh, MS, and Vasanthan, T. Review of the beneficial and anti-nutritional qualities of phytic acid, and procedures for removing it from food products. Food Res Int. (2021) 143:110284. doi: 10.1016/j.foodres.2021.110284
76. Kumar, A, Singh, B, Raigond, P, Sahu, C, Mishra, UN, Sharma, S, et al. Phytic acid: blessing in disguise, a prime compound required for both plant and human nutrition. Food Res Int. (2021) 142:110193. doi: 10.1016/j.foodres.2021.110193
77. Zhang, YY, Stockmann, R, Ng, K, and Ajlouni, S. Revisiting phytate-element interactions: implications for iron, zinc and calcium bioavailability, with emphasis on legumes. Crit Rev Food Sci Nutr. (2022) 62:1696–712. doi: 10.1080/10408398.2020.1846014
78. Kumar, A, Kumar, V, Krishnan, V, Hada, A, Marathe, A, C, P, et al. Seed targeted RNAi-mediated silencing of GmMIPS1 limits phytate accumulation and improves mineral bioavailability in soybean. Sci Rep. (2019) 9:7744. doi: 10.1038/s41598-019-44255-7
79. Wang, R, and Guo, S. Phytic acid and its interactions: contributions to protein functionality, food processing, and safety. Comp Rev Food Sci Food Safe. (2021) 20:2081–105. doi: 10.1111/1541-4337.12714
80. Gupta, RK, Gangoliya, SS, and Singh, NK. Reduction of phytic acid and enhancement of bioavailable micronutrients in food grains. J Food Sci Technol. (2015) 52:676–84. doi: 10.1007/s13197-013-0978-y
81. Wu, J, Li, X, Nie, H, Shen, Y, Guo, Z, Huihan Chu, C, et al. Phytic acid promotes high glucose-mediated bone marrow mesenchymal stem cells osteogenesis via modulating circEIF4B that sponges miR-186-5p and complexes with IGF2BP3. Biochem Pharmacol. (2024) 222:116118. doi: 10.1016/j.bcp.2024.116118
82. Sanchis, P, Prieto, RM, Konieczna, J, Grases, F, Abete, I, Salas-Salvadó, J, et al. Estimated Phytate intake is associated with bone mineral density in Mediterranean postmenopausal women. Nutrients. (2023) 15:1791. doi: 10.3390/nu15071791
83. Sanchis, P, López-González, Á-A, Costa-Bauzá, A, Busquets-Cortés, C, Riutord, P, Calvo, P, et al. Understanding the protective effect of Phytate in bone decalcification related-diseases. Nutrients. (2021) 13:2859. doi: 10.3390/nu13082859
84. Guimerà, J, Martínez, A, Bauza, JL, Sanchís, P, Pieras, E, and Grases, F. Effect of phytate on hypercalciuria secondary to bone resorption in patients with urinary stones: pilot study. Urolithiasis. (2022) 50:685–90. doi: 10.1007/s00240-022-01357-8
85. Lopez-Gonzalez, AA. Urinary phytate concentration and risk of fracture determined by the FRAX index in a group of postmenopausal women. Turk J Med Sci. (2019) 49:458–63. doi: 10.3906/sag-1806-117
86. Yoshiko, Y, and Vucenik, I. Inositol Hexaphosphate in bone health and disease. Biomolecules. (2024) 14:1072. doi: 10.3390/biom14091072
87. Rebello, S, Jose, L, Sindhu, R, and Aneesh, EM. Molecular advancements in the development of thermostable phytases. Appl Microbiol Biotechnol. (2017) 101:2677–89. doi: 10.1007/s00253-017-8195-7
88. Sandez Penidez, SH, Velasco Manini, MA, Gerez, CL, and Rollán, GC. Partial characterization and purification of phytase from Lactobacillus plantarum CRL1964 isolated from pseudocereals. J Basic Microbiol. (2020) 60:787–98. doi: 10.1002/jobm.202000236
89. Shi, L, Arntfield, SD, and Nickerson, M. Changes in levels of phytic acid, lectins and oxalates during soaking and cooking of Canadian pulses. Food Res Int. (2018) 107:660–8. doi: 10.1016/j.foodres.2018.02.056
90. Maldonado-Alvarado, P, Pavón-Vargas, DJ, Abarca-Robles, J, Valencia-Chamorro, S, and Haros, CM. Effect of germination on the nutritional properties, Phytic acid content, and Phytase activity of quinoa (Chenopodium quinoa Willd). Foods. (2023) 12:389. doi: 10.3390/foods12020389
91. Joudaki, H, Aria, N, Moravej, R, Rezaei Yazdi, M, Emami-Karvani, Z, and Hamblin, MR. Microbial Phytases: properties and applications in the food industry. Curr Microbiol. (2023) 80:374. doi: 10.1007/s00284-023-03471-1
92. Vilanculos, SL, Svanberg, U, and Andlid, T. Phytate degradation in composite wheat/cassava/sorghum bread: effects of phytase-secreting yeasts and addition of yeast extracts. Food Sci Nutr. (2024) 12:216–26. doi: 10.1002/fsn3.3754
93. Samtiya, M, Aluko, RE, Puniya, AK, and Dhewa, T. Enhancing micronutrients bioavailability through fermentation of plant-based foods: a concise review. Fermentation. (2021) 7:63. doi: 10.3390/fermentation7020063
94. Mukherjee, A, Breselge, S, Dimidi, E, Marco, ML, and Cotter, PD. Fermented foods and gastrointestinal health: underlying mechanisms. Nat Rev Gastroenterol Hepatol. (2024) 21:248–66. doi: 10.1038/s41575-023-00869-x
95. Ahmed, MI, Xu, X, Sulieman, AA, Na, Y, and Mahdi, AA. The effect of fermentation time on in vitro bioavailability of iron, zinc, and calcium of kisra bread produced from koreeb (Dactyloctenium aegyptium) seeds flour. Microchem J. (2020) 154:104644. doi: 10.1016/j.microc.2020.104644
96. Karaman, K, Sagdic, O, and Durak, MZ. Use of phytase active yeasts and lactic acid bacteria isolated from sourdough in the production of whole wheat bread. LWT. (2018) 91:557–67. doi: 10.1016/j.lwt.2018.01.055
97. Li, Z, Zhou, M, Cui, M, Wang, Y, and Li, H. Improvement of whole wheat dough fermentation for steamed bread making using selected phytate-degrading Wickerhamomyces anomalus P4. J Cereal Sci. (2021) 100:103261. doi: 10.1016/j.jcs.2021.103261
98. Song, HS, Lee, SH, Ahn, SW, Kim, JY, Rhee, J-K, and Roh, SW. Effects of the main ingredients of the fermented food, kimchi, on bacterial composition and metabolite profile. Food Res Int. (2021) 149:110668. doi: 10.1016/j.foodres.2021.110668
99. Gan, J, Kong, X, Wang, K, Chen, Y, Du, M, Xu, B, et al. Effect of fermentation using different lactic acid bacteria strains on the nutrient components and mineral bioavailability of soybean yogurt alternative. Front Nutr. (2023) 10:1198456. doi: 10.3389/fnut.2023.1198456
100. Yang, H-J, Zhang, T, Yue, Y, Jeong, S-J, Ryu, M-S, Wu, X, et al. Protective effect of long-term fermented soybeans with abundant Bacillus subtilis on glucose and bone metabolism and memory function in Ovariectomized rats: modulation of the gut microbiota. Foods. (2023) 12:2958. doi: 10.3390/foods12152958
101. Rodríguez-Olleros Rodríguez, C, and Díaz Curiel, M. Vitamin K and bone health: a review on the effects of vitamin K deficiency and supplementation and the effect of non-vitamin K antagonist oral anticoagulants on different bone parameters. J Osteoporos. (2019) 2019:1–8. doi: 10.1155/2019/2069176
102. Halder, M, Petsophonsakul, P, Akbulut, AC, Pavlic, A, Bohan, F, Anderson, E, et al. Vitamin K: double bonds beyond coagulation insights into differences between vitamin K1 and K2 in health and disease. IJMS. (2019) 20:896. doi: 10.3390/ijms20040896
103. Sim, M, Lewis, JR, Prince, RL, Levinger, I, Brennan-Speranza, TC, Palmer, C, et al. The effects of vitamin K-rich green leafy vegetables on bone metabolism: a 4-week randomised controlled trial in middle-aged and older individuals. Bone Reports. (2020) 12:100274. doi: 10.1016/j.bonr.2020.100274
104. Mladěnka, P, Macáková, K, Kujovská Krčmová, L, Javorská, L, Mrštná, K, Carazo, A, et al. Vitamin K – sources, physiological role, kinetics, deficiency, detection, therapeutic use, and toxicity. Nutr Rev. (2022) 80:677–98. doi: 10.1093/nutrit/nuab061
105. Stock, M, and Schett, G. Vitamin K-Dependent Proteins in Skeletal Development and Disease. IJMS. (2021) 22:9328. doi: 10.3390/ijms22179328
106. Liu, Y, Van Bennekom, EO, Zhang, Y, Abee, T, and Smid, EJ. Long-chain vitamin K2 production in Lactococcus lactis is influenced by temperature, carbon source, aeration and mode of energy metabolism. Microb Cell Factories. (2019) 18:129. doi: 10.1186/s12934-019-1179-9
107. Lee, SY, Hu, X, and Stuckey, DC. Optimised “green solvent” extraction of long-chain menaquinones (vitamin K2) from wet Lactococcus lactis biomass. Sep Purif Technol. (2022) 287:120560. doi: 10.1016/j.seppur.2022.120560
108. Walther, B, Guggisberg, D, Schmidt, RS, Portmann, R, Risse, M-C, Badertscher, R, et al. Quantitative analysis of menaquinones (vitamin K2) in various types of cheese from Switzerland. Int Dairy J. (2021) 112:104853. doi: 10.1016/j.idairyj.2020.104853
109. Lundberg, HE, Glasø, M, Chhura, R, Shukla, AA, Austlid, T, Sarwar, Z, et al. Effect on bone anabolic markers of daily cheese intake with and without vitamin K2: a randomised clinical trial. BMJNPH. (2022) 5:182–90. doi: 10.1136/bmjnph-2022-000424
110. Sato, T, Inaba, N, and Yamashita, T. MK-7 and its effects on bone quality and strength. Nutrients. (2020) 12:965. doi: 10.3390/nu12040965
111. Kharnaior, P, and Tamang, JP. Metagenomic-Metabolomic Mining of Kinema, a naturally fermented soybean food of the eastern Himalayas. Front Microbiol. (2022) 13:868383. doi: 10.3389/fmicb.2022.868383
112. Torbergsen, AC, Watne, LO, Frihagen, F, Wyller, TB, and Mowè, M. Effects of nutritional intervention upon bone turnover in elderly hip fracture patients. Randomized controlled trial. Clin Nutr ESPEN. (2019) 29:52–8. doi: 10.1016/j.clnesp.2017.11.012
113. Alonso, N, Meinitzer, A, Fritz-Petrin, E, Enko, D, and Herrmann, M. Role of vitamin K in bone and muscle metabolism. Calcif Tissue Int. (2022) 112:178–96. doi: 10.1007/s00223-022-00955-3
114. Liao, Z, Chang, J, Zhu, Z, Han, W, Meng, T, Zheng, S, et al. Associations between dietary intake of vitamin K and changes in symptomatic and structural changes in patients with knee osteoarthritis. Arthritis Care Res. (2023) 75:1503–10. doi: 10.1002/acr.24964
115. Elshaikh, AO, Shah, L, Joy Mathew, C, Lee, R, Jose, MT, and Cancarevic, I. Influence of vitamin K on bone mineral density and osteoporosis. Cureus. (2020) 12:e10816. doi: 10.7759/cureus.10816
116. Zhang, Y, Liu, Z, Duan, L, Ji, Y, Yang, S, Zhang, Y, et al. Effect of low-dose vitamin K2 supplementation on bone mineral density in middle-aged and elderly Chinese: a randomized controlled study. Calcif Tissue Int. (2020) 106:476–85. doi: 10.1007/s00223-020-00669-4
117. Ma, M, Ma, Z, He, Y, Sun, H, Yang, B, Ruan, B, et al. Efficacy of vitamin K2 in the prevention and treatment of postmenopausal osteoporosis: a systematic review and meta-analysis of randomized controlled trials. Front Public Health. (2022) 10:979649. doi: 10.3389/fpubh.2022.979649
118. Rusu, ME, Bigman, G, Ryan, AS, and Popa, D-S. Investigating the effects and mechanisms of combined vitamin D and K supplementation in postmenopausal women: An up-to-date comprehensive review of clinical studies. Nutrients. (2024) 16:2356. doi: 10.3390/nu16142356
119. Kojima, A, Ikehara, S, Kamiya, K, Kajita, E, Sato, Y, Kouda, K, et al. Natto intake is inversely associated with osteoporotic fracture risk in postmenopausal Japanese women. J Nutr. (2020) 150:599–605. doi: 10.1093/jn/nxz292
120. Iwasaki, M, Sato, M, Yoshihara, A, Saito, T, Kitamura, K, Ansai, T, et al. A 5-year longitudinal association between dietary fermented soya bean (natto) intake and tooth loss through bone mineral density in postmenopausal women: the Yokogoshi cohort study. Gerodontology. (2021) 38:267–75. doi: 10.1111/ger.12523
121. Sim, M, Strydom, A, Blekkenhorst, LC, Bondonno, NP, McCormick, R, Lim, WH, et al. Dietary vitamin K1 intake is associated with lower long-term fracture-related hospitalization risk: the Perth longitudinal study of ageing women. Food Funct. (2022) 13:10642–50. doi: 10.1039/D2FO02494B
122. Hirata, H, Kitamura, K, Saito, T, Kobayashi, R, Iwasaki, M, Yoshihara, A, et al. Association between dietary intake and bone mineral density in Japanese postmenopausal women: the Yokogoshi cohort study. Tohoku J Exp Med. (2016) 239:95–101. doi: 10.1620/tjem.239.95
123. Clements, M, Heffernan, M, Ward, M, Hoey, L, Doherty, LC, Hack Mendes, R, et al. A 2-year randomized controlled trial with low-dose B-vitamin supplementation shows benefits on bone mineral density in adults with lower B12 status. J Bone Miner Res. (2020) 37:2443–55. doi: 10.1002/jbmr.4709
124. Yang, Q, Zhang, L, Sun, D, Jie, S, Tao, X, Meng, Q, et al. Dietary riboflavin (vitamin B2) intake and osteoporosis in U.S. female adults: unveiling of association and exploration of potential molecular mechanisms. Nutr J. (2025) 24:53. doi: 10.1186/s12937-025-01103-x
125. Bailey, RL, Looker, AC, Lu, Z, Fan, R, Eicher-Miller, HA, Fakhouri, TH, et al. B-vitamin status and bone mineral density and risk of lumbar osteoporosis in older females in the United States. Am J Clin Nutr. (2015) 102:687–94. doi: 10.3945/ajcn.115.108787
126. Zhang, R, Huang, Q, Su, G, Wei, M, Cui, Y, Zhou, H, et al. Association between multiple vitamins and bone mineral density: a cross-sectional and population-based study in the NHANES from 2005 to 2006. BMC Musculoskelet Disord. (2023) 24:113. doi: 10.1186/s12891-023-06202-6
127. Chan, R, Leung, J, and Woo, J. No association between dietary vitamin K intake and fracture risk in Chinese community-dwelling older men and women: a prospective study. Calcif Tissue Int. (2012) 90:396–403. doi: 10.1007/s00223-012-9586-5
128. Xie, C, Gong, J, Zheng, C, Zhang, J, Gao, J, Tian, C, et al. Effects of vitamin K supplementation on bone mineral density at different sites and bone metabolism in the middle-aged and elderly population: a meta-analysis and systematic review of randomized controlled trials. Bone Joint Res. (2024) 13:750–63. doi: 10.1302/2046-3758.1312.BJR-2024-0053.R1
129. Dai, Z, and Koh, W-P. B-vitamins and bone health–a review of the current evidence. Nutrients. (2015) 7:3322–46. doi: 10.3390/nu7053322
130. Dai, Z, Wang, R, Ang, LW, Yuan, J-M, and Koh, W-P. Dietary B vitamin intake and risk of hip fracture: the Singapore Chinese health study. Osteoporos Int. (2013) 24:2049–59. doi: 10.1007/s00198-012-2233-1
131. Yang, F, Chen, C, Ni, D, Yang, Y, Tian, J, Li, Y, et al. Effects of fermentation on bioactivity and the composition of polyphenols contained in polyphenol-rich foods: a review. Foods. (2023) 12:3315. doi: 10.3390/foods12173315
132. Roy, A, Khan, A, Ahmad, I, Alghamdi, S, Rajab, BS, Babalghith, AO, et al. Flavonoids a bioactive compound from medicinal plants and its therapeutic applications. Biomed Res Int. (2022) 2022:5445291. doi: 10.1155/2022/5445291
133. Chen, S, Wang, X, Cheng, Y, Gao, H, and Chen, X. A review of classification, biosynthesis, biological activities and potential applications of flavonoids. Molecules. (2023) 28:4982. doi: 10.3390/molecules28134982
134. Dini, I, and Grumetto, L. Recent advances in natural polyphenol research. Molecules. (2022) 27:8777. doi: 10.3390/molecules27248777
135. Ramesh, P, Jagadeesan, R, Sekaran, S, Dhanasekaran, A, and Vimalraj, S. Flavonoids: classification, function, and molecular mechanisms involved in bone Remodelling. Front Endocrinol. (2021) 12:779638. doi: 10.3389/fendo.2021.779638
136. Li, Y, Li, L, Li, X, Luo, B, Ye, Q, Wang, H, et al. A mechanistic review of chinese medicine polyphenols on bone formation and resorption. Front Pharmacol. (2022) 13:1017538. doi: 10.3389/fphar.2022.1017538
137. Xv, D, Cao, Y, Hou, Y, Hu, Y, Li, M, Xie, C, et al. Polyphenols and functionalized hydrogels for osteoporotic bone regeneration. Macromol Rapid Commun. (2025) 46:e2400653. doi: 10.1002/marc.202400653
138. Luca, SV, Macovei, I, Bujor, A, Miron, A, Skalicka-Woźniak, K, Aprotosoaie, AC, et al. Bioactivity of dietary polyphenols: the role of metabolites. Crit Rev Food Sci Nutr. (2020) 60:626–59. doi: 10.1080/10408398.2018.1546669
139. Piao, Y-Z, and Eun, J-B. Physicochemical characteristics and isoflavones content during manufacture of short-time fermented soybean product (cheonggukjang). J Food Sci Technol. (2020) 57:2190–7. doi: 10.1007/s13197-020-04255-2
140. Li, S, Tao, Y, Li, D, Wen, G, Zhou, J, Manickam, S, et al. Fermentation of blueberry juices using autochthonous lactic acid bacteria isolated from fruit environment: fermentation characteristics and evolution of phenolic profiles. Chemosphere. (2021) 276:130090. doi: 10.1016/j.chemosphere.2021.130090
141. Lee, D, Kim, MJ, Ahn, J, Lee, SH, Lee, H, Kim, JH, et al. Nutrikinetics of isoflavone metabolites after fermented soybean product (Cheonggukjang) ingestion in ovariectomized mice. Mol Nutr Food Res. (2017) 61:1700322. doi: 10.1002/mnfr.201700322
142. Kim, I-S. Current perspectives on the beneficial effects of soybean Isoflavones and their metabolites for humans. Antioxidants. (2021) 10:1064. doi: 10.3390/antiox10071064
143. Kim, J-S, Lee, H, Nirmala, FS, Jung, CH, Jang, Y-J, Ha, T-Y, et al. Dry-fermented soybean food (Cheonggukjang) ameliorates senile osteoporosis in the senescence-accelerated mouse prone 6 model. J Med Food. (2019) 22:1047–57. doi: 10.1089/jmf.2018.4335
144. Yang, HJ, Kwon, DY, Moon, NR, Kim, MJ, Kang, HJ, Jung, DY, et al. Soybean fermentation with Bacillus licheniformis increases insulin sensitizing and insulinotropic activity. Food Funct. (2013) 4:1675–84. doi: 10.1039/c3fo60198f
145. Slavin, JL, Karr, SC, Hutchins, AM, and Lampe, JW. Influence of soybean processing, habitual diet, and soy dose on urinary isoflavonoid excretion. Am J Clin Nutr. (1998) 68:1492S–5S.
146. Khosravi, A, and Razavi, SH. Therapeutic effects of polyphenols in fermented soybean and black soybean products. J Funct Foods. (2021) 81:104467. doi: 10.1016/j.jff.2021.104467
147. Hong, G-E, Mandal, PK, Lim, K-W, and Lee, C-H. Fermentation increases isoflavone aglycone contents in black soybean pulp. Asian J Anim Vet Adv. (2012) 7:502–11. doi: 10.3923/ajava.2012.502.511
148. Xu, Q, Yu, Y, and Chen, K. The potential application of fermented tea as a drink for regulating bone mass. Front Pharmacol. (2024) 15:1353811. doi: 10.3389/fphar.2024.1353811
149. Chisari, E, Shivappa, N, and Vyas, S. Polyphenol-rich foods and osteoporosis. CPD. (2019) 25:2459–66. doi: 10.2174/1381612825666190722093959
150. Austermann, K, Baecker, N, Stehle, P, and Heer, M. Putative effects of nutritive polyphenols on bone metabolism in vivo—evidence from human studies. Nutrients. (2019) 11:871. doi: 10.3390/nu11040871
151. Cruz-Casas, DE, Aguilar, CN, Ascacio-Valdés, JA, Rodríguez-Herrera, R, Chávez-González, ML, and Flores-Gallegos, AC. Enzymatic hydrolysis and microbial fermentation: the most favorable biotechnological methods for the release of bioactive peptides. Food Chem: Molecular Sci. (2021) 3:100047. doi: 10.1016/j.fochms.2021.100047
152. Ahn, C-B, and Je, J-Y. Bone health-promoting bioactive peptides. J Food Biochem. (2019) 43:e12529. doi: 10.1111/jfbc.12529
153. Chaudhary, A, Bhalla, S, Patiyal, S, Raghava, GPS, and Sahni, G. FermFooDb: a database of bioactive peptides derived from fermented foods. Heliyon. (2021) 7:e06668. doi: 10.1016/j.heliyon.2021.e06668
154. Guo, Q, Chen, P, and Chen, X. Bioactive peptides derived from fermented foods: preparation and biological activities. J Funct Foods. (2023) 101:105422. doi: 10.1016/j.jff.2023.105422
155. Hiago Bellaver, E, Eliza Redin, E, Militão Da Costa, I, Schittler Moroni, L, and Pinto, KA. Food peptidomic analysis of bovine milk fermented by Lacticaseibacillus casei LBC 237: in silico prediction of bioactive peptides and anticancer potential. Food Res Int. (2024) 180:114060. doi: 10.1016/j.foodres.2024.114060
156. Saubenova, M, Oleinikova, Y, Rapoport, A, Maksimovich, S, Yermekbay, Z, and Khamedova, E. Bioactive peptides derived from whey proteins for health and functional beverages. Fermentation. (2024) 10:359. doi: 10.3390/fermentation10070359
157. Raveschot, C, Cudennec, B, Coutte, F, Flahaut, C, Fremont, M, Drider, D, et al. Production of bioactive peptides by Lactobacillus species: from gene to application. Front Microbiol. (2018) 9:2354. doi: 10.3389/fmicb.2018.02354
158. Peighambardoust, SH, Karami, Z, Pateiro, M, and Lorenzo, JM. A review on health-promoting, biological, and functional aspects of bioactive peptides in food applications. Biomolecules. (2021) 11:631. doi: 10.3390/biom11050631
159. Nielsen, SD, Jakobsen, LMA, Geiker, NRW, and Bertram, HC. Chemically acidified, live and heat-inactivated fermented dairy yoghurt show distinct bioactive peptides, free amino acids and small compounds profiles. Food Chem. (2022) 376:131919. doi: 10.1016/j.foodchem.2021.131919
160. Samtiya, M, Samtiya, S, Badgujar, PC, Puniya, AK, Dhewa, T, and Aluko, RE. Health-promoting and therapeutic attributes of Milk-derived bioactive peptides. Nutrients. (2022) 14:3001. doi: 10.3390/nu14153001
161. Solieri, L, Valentini, M, Cattivelli, A, Sola, L, Helal, A, Martini, S, et al. Fermentation of whey protein concentrate by Streptococcus thermophilus strains releases peptides with biological activities. Process Biochem. (2022) 121:590–600. doi: 10.1016/j.procbio.2022.08.003
162. Sultan, S, Huma, N, Butt, MS, Aleem, M, and Abbas, M. Therapeutic potential of dairy bioactive peptides: a contemporary perspective. Crit Rev Food Sci Nutr. (2018) 58:105–15. doi: 10.1080/10408398.2015.1136590
163. Canoy, D, Harvey, NC, Prieto-Alhambra, D, Cooper, C, Meyer, HE, Åsvold, BO, et al. Elevated blood pressure, antihypertensive medications and bone health in the population: revisiting old hypotheses and exploring future research directions. Osteoporos Int. (2022) 33:315–26. doi: 10.1007/s00198-021-06190-0
164. Du, X-P, Zheng, M-L, Yang, X-C, and Zheng, M-L. High blood pressure is associated with increased risk of future fracture, but not vice versa. Sci Rep. (2024) 14:8005. doi: 10.1038/s41598-024-58691-7
165. Zhang, Y, Guo, Y, Jiang, C, Xie, A, Yue, X, and Li, M. A review of casein phosphopeptides: from enrichment identification to biological properties. Food Biosci. (2024) 59:104217. doi: 10.1016/j.fbio.2024.104217
166. Miclo, L, Roux, É, Genay, M, Brusseaux, É, Poirson, C, Jameh, N, et al. Variability of hydrolysis of β-, αs1 -, and αs2 -caseins by 10 strains of Streptococcus thermophilus and resulting bioactive peptides. J Agric Food Chem. (2012) 60:554–65. doi: 10.1021/jf202176d
167. Tenenbaum, M, Deracinois, B, Dugardi, C, Matéos, A, Romelard, A, Auger, J, et al. Identification, production and bioactivity of casein phosphopeptides – a review. Food Res Int. (2022) 157:111360. doi: 10.1016/j.foodres.2022.111360
168. Rizzoli, R, and Biver, E. Effects of fermented Milk products on bone. Calcif Tissue Int. (2018) 102:489–500. doi: 10.1007/s00223-017-0317-9
169. Huang, W, Lao, L, Deng, Y, Li, Z, Liao, W, Duan, S, et al. Preparation, characterization, and osteogenic activity mechanism of casein phosphopeptide-calcium chelate. Front Nutr. (2022) 9:960228. doi: 10.3389/fnut.2022.960228
170. Narva, M, Rissanen, J, Halleen, J, Vapaatalo, H, Väänänen, K, and Korpela, R. Effects of bioactive peptide, Valyl-prolyl-proline (VPP), and Lactobacillus helveticus fermented Milk containing VPP on bone loss in Ovariectomized rats. Ann Nutr Metab. (2007) 51:65–74. doi: 10.1159/000100823
171. Lee, CS, Kim, J-Y, Kim, BK, Lee, IO, Park, NH, and Kim, SH. Lactobacillus-fermented milk products attenuate bone loss in an experimental rat model of ovariectomy-induced post-menopausal primary osteoporosis. J Appl Microbiol. (2021) 130:2041–62. doi: 10.1111/jam.14852
172. Dalabasmaz, S, De La Torre, EP, Gensberger-Reigl, S, Pischetsrieder, M, and Rodríguez-Ortega, MJ. Identification of potential bioactive peptides in sheep Milk kefir through Peptidomic analysis at different fermentation times. Foods. (2023) 12:2974. doi: 10.3390/foods12152974
173. Chang, GR-L, Lin, W-Y, Fan, H-C, Tu, M-Y, Liu, Y-H, Yen, C-C, et al. Kefir peptides ameliorate osteoporosis in AKR1A1 knockout mice with vitamin C deficiency by promoting osteoblastogenesis and inhibiting osteoclastogenesis. Biomed Pharmacother. (2022) 156:113859. doi: 10.1016/j.biopha.2022.113859
174. Tu, M-Y, Han, K-Y, Chang, GR-L, Lai, G-D, Chang, K-Y, Chen, C-F, et al. Kefir peptides prevent estrogen deficiency-induced bone loss and modulate the structure of the gut microbiota in Ovariectomized mice. Nutrients. (2020) 12:3432. doi: 10.3390/nu12113432
175. Harahap, IA, Kuligowski, M, Schmidt, M, and Suliburska, J. The impact of soybean products and probiotics on calcium bioaccessibility from organic and inorganic calcium salts in an in vitro digestion model. Food Chem Advan. (2023) 2:100269. doi: 10.1016/j.focha.2023.100269
176. Elbanna, K, Alsulami, FS, Neyaz, LA, and Abulreesh, HH. Poly (γ) glutamic acid: a unique microbial biopolymer with diverse commercial applicability. Front Microbiol. (2024) 15:1348411. doi: 10.3389/fmicb.2024.1348411
177. Johnson, LC, Akinmola, AT, and Scholz, C. Poly(glutamic acid): from natto to drug delivery systems. Biocatal Agric Biotechnol. (2022) 40:102292. doi: 10.1016/j.bcab.2022.102292
178. Prakash Tamang, J, Kharnaior, P, and Pariyar, P. Whole genome sequencing of the poly-γ-glutamic acid-producing novel Bacillus subtilis Tamang strain, isolated from spontaneously fermented kinema. Food Res Int. (2024) 190:114655. doi: 10.1016/j.foodres.2024.114655
179. Yang, L-C, Wu, J-B, Ho, G-H, Yang, S-C, Huang, Y-P, and Lin, W-C. Effects of poly-γ-glutamic acid on calcium absorption in rats. Biosci Biotechnol Biochem. (2008) 72:3084–90. doi: 10.1271/bbb.80024
180. Tanimoto, H, Fox, T, Eagles, J, Satoh, H, Nozawa, H, Okiyama, A, et al. Acute effect of poly-γ-glutamic acid on calcium absorption in post-menopausal women. J Am Coll Nutr. (2007) 26:645–9. doi: 10.1080/07315724.2007.10719642
181. Cao, Z-H, Green-Johnson, JM, Buckley, ND, and Lin, Q-Y. Bioactivity of soy-based fermented foods: a review. Biotechnol Adv. (2019) 37:223–38. doi: 10.1016/j.biotechadv.2018.12.001
182. Chiang, S, Liao, J, and Pan, T. Effect of bioactive compounds in lactobacilli-fermented soy skim milk on femoral bone microstructure of aging mice. J Sci Food Agric. (2012) 92:328–35. doi: 10.1002/jsfa.4579
183. Seol, JY, Youn, YN, Koo, M, Kim, HJ, and Choi, SY. Influence of water-soluble extracts of long-term fermented Doenjang on bone metabolism bioactivity and breast cancer suppression. Food Sci Biotechnol. (2016) 25:517–24. doi: 10.1007/s10068-016-0072-0
184. Paul, JK, Azmal, M, Haque, ASNB, Meem, M, and Ghosh, A. Unlocking the secrets of the human gut microbiota: comprehensive review on its role in different diseases. World J Gastroenterol. (2025) 31:328–335. doi: 10.3748/wjg.v31.i5.99913
185. Khalil, M, Di Ciaula, A, Mahdi, L, Jaber, N, Di Palo, DM, Graziani, A, et al. Unraveling the role of the human gut microbiome in health and diseases. Microorganisms. (2024) 12:2333. doi: 10.3390/microorganisms12112333
186. Barry, DJ, Wu, SSX, and Cooke, MB. The relationship between gut microbiota, muscle mass and physical function in older individuals: a systematic review. Nutrients. (2024) 17:81. doi: 10.3390/nu17010081
187. Zemanova, N, Omelka, R, Mondockova, V, Kovacova, V, and Martiniakova, M. Roles of gut microbiome in bone homeostasis and its relationship with bone-related diseases. Biology. (2022) 11:1402. doi: 10.3390/biology11101402
188. Latif, A, Shehzad, A, Niazi, S, Zahid, A, Ashraf, W, Iqbal, MW, et al. Probiotics: mechanism of action, health benefits and their application in food industries. Front Microbiol. (2023) 14:1216674. doi: 10.3389/fmicb.2023.1216674
189. Chiang, S-S, and Pan, T-M. Antiosteoporotic effects of Lactobacillus -fermented soy skim milk on bone mineral density and the microstructure of femoral bone in ovariectomized mice. J Agric Food Chem. (2011) 59:7734–42. doi: 10.1021/jf2013716
190. Eor, JY, Tan, PL, Son, YJ, Lee, CS, and Kim, SH. Milk products fermented by Lactobacillus strains modulate the gut–bone axis in an ovariectomised murine model. Int J Dairy Technol. (2020) 73:743–56. doi: 10.1111/1471-0307.12708
191. Zeng, MY, Inohara, N, and Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. (2017) 10:18–26. doi: 10.1038/mi.2016.75
192. Marahleh, A, Kitaura, H, Ohori, F, Kishikawa, A, Ogawa, S, Shen, W-R, et al. TNF-α directly enhances osteocyte RANKL expression and promotes osteoclast formation. Front Immunol. (2019) 10:2925. doi: 10.3389/fimmu.2019.02925
193. Li, L, Rao, S, Cheng, Y, Zhuo, X, Deng, C, Xu, N, et al. Microbial osteoporosis: the interplay between the gut microbiota and bones via host metabolism and immunity. MicrobiologyOpen. (2019) 8:e00810. doi: 10.1002/mbo3.810
194. Duffuler, P, Bhullar, KS, and Wu, J. Targeting gut microbiota in osteoporosis: impact of the microbial based functional food ingredients. Food Sci Human Wellness. (2024) 13:1–15. doi: 10.26599/FSHW.2022.9250001
195. Ruscitti, P, Cipriani, P, Carubbi, F, Liakouli, V, Zazzeroni, F, Di Benedetto, P, et al. The role of IL-1 β in the bone loss during rheumatic diseases. Mediat Inflamm. (2015) 2015:782382. doi: 10.1155/2015/782382
196. Ono, T, Hayashi, M, Sasaki, F, and Nakashima, T. RANKL biology: bone metabolism, the immune system, and beyond. Inflamm Regener. (2020) 40:2. doi: 10.1186/s41232-019-0111-3
197. Lyu, Z, Hu, Y, Guo, Y, and Liu, D. Modulation of bone remodeling by the gut microbiota: a new therapy for osteoporosis. Bone Res. (2023) 11:31. doi: 10.1038/s41413-023-00264-x
198. Jensen, EA, Young, JA, Mathes, SC, List, EO, Carroll, RK, Kuhn, J, et al. Crosstalk between the growth hormone/insulin-like growth factor-1 axis and the gut microbiome: a new frontier for microbial endocrinology. Growth Hormon IGF Res. (2020) 53-54:101333. doi: 10.1016/j.ghir.2020.101333
199. Zaiss, MM, Jones, RM, Schett, G, and Pacifici, R. The gut-bone axis: how bacterial metabolites bridge the distance. J Clin Invest. (2019) 129:3018–28. doi: 10.1172/JCI128521
200. Fang, J, Zhang, X, Chen, X, Wang, Z, Zheng, S, Cheng, Y, et al. The role of insulin-like growth factor-1 in bone remodeling: a review. Int J Biol Macromol. (2023) 238:124125. doi: 10.1016/j.ijbiomac.2023.124125
201. Yan, J, and Charles, JF. Gut microbiota and IGF-1. Calcif Tissue Int. (2018) 102:406–14. doi: 10.1007/s00223-018-0395-3
202. Tu, Y, Kuang, X, Zhang, L, and Xu, X. The associations of gut microbiota, endocrine system and bone metabolism. Front Microbiol. (2023) 14:1124945. doi: 10.3389/fmicb.2023.1124945
203. Hao, Y, Wang, H, Fang, L, Bian, J, Gao, Y, and Li, C. H2S donor and bone metabolism. Front Pharmacol. (2021) 12:661601. doi: 10.3389/fphar.2021.661601
204. Gambari, L, Lisignoli, G, Cattini, L, Manferdini, C, Facchini, A, and Grassi, F. Sodium hydrosulfide inhibits the differentiation of osteoclast progenitor cells via NRF2-dependent mechanism. Pharmacol Res. (2014) 87:99–112.
205. Grassi, F, Tyagi, AM, Calvert, JW, Gambari, L, Walker, LD, Yu, M, et al. Hydrogen sulfide is a novel regulator of bone formation implicated in the bone loss induced by estrogen deficiency. J Bone Miner Res. (2016) 31:949–63. doi: 10.1002/jbmr.2757
206. Gambari, L, Lisignoli, G, Cattini, L, Manferdini, C, Facchini, A, Grassi, F, et al. Sodium hydrosulfide inhibits the differentiation of osteoclast progenitor cells via NRF2-dependent mechanism. Pharmacological Research. (2014) 87:99–112. doi: 10.1016/j.phrs.2014.06.014
207. Liu, Y, Yang, R, Liu, X, Zhou, Y, Qu, C, Kikuiri, T, et al. Hydrogen sulfide maintains mesenchymal stem cell function and bone homeostasis via regulation of Ca2+ channel Sulfhydration. Cell Stem Cell. (2014) 15:66–78. doi: 10.1016/j.stem.2014.03.005
208. Gambari, L, Grigolo, B, and Grassi, F. Hydrogen sulfide in bone tissue regeneration and repair: state of the art and new perspectives. IJMS. (2019) 20:5231. doi: 10.3390/ijms20205231
209. You, X, Yan, J, Herzog, J, Nobakhti, S, Campbell, R, Hoke, A, et al. Bone loss with aging is independent of gut microbiome in mice. Bone Res. (2024) 12:65. doi: 10.1038/s41413-024-00366-0
210. Rizzoli, R, and Chevalley, T. Nutrition and osteoporosis prevention. Curr Osteoporos Rep. (2024) 22:515–22. doi: 10.1007/s11914-024-00892-0
211. Diez-Ozaeta, I, and Astiazaran, OJ. Fermented foods: An update on evidence-based health benefits and future perspectives. Food Res Int. (2022) 156:111133. doi: 10.1016/j.foodres.2022.111133
212. Sawant, SS, Park, H-Y, Sim, E-Y, Kim, H-S, and Choi, H-S. Microbial fermentation in food: impact on functional properties and nutritional enhancement—a review of recent developments. Fermentation. (2025) 11:15. doi: 10.3390/fermentation11010015
213. Li, KJ, Brouwer-Brolsma, EM, Burton-Pimentel, KJ, Vergères, G, and Feskens, EJM. A systematic review to identify biomarkers of intake for fermented food products. Genes Nutr. (2021) 16:5. doi: 10.1186/s12263-021-00686-4
214. Caffrey, EB, Perelman, D, Ward, CP, Sonnenburg, ED, Gardner, CD, and Sonnenburg, JL. Unpacking food fermentation: clinically relevant tools for fermented food identification and consumption. Adv Nutr. (2025) 16:100412. doi: 10.1016/j.advnut.2025.100412
215. Park, I, and Mannaa, M. Fermented foods as functional systems: microbial communities and metabolites influencing gut health and systemic outcomes. Foods. (2025) 14:2292. doi: 10.3390/foods14132292
216. Feng, B, Lu, J, Han, Y, Han, Y, Qiu, X, and Zeng, Z. The role of short-chain fatty acids in the regulation of osteoporosis: new perspectives from gut microbiota to bone health: a review. Medicine. (2024) 103:e39471. doi: 10.1097/MD.0000000000039471
217. Turbić, A, Vandenput, L, Gandham, A, and Lorentzon, M. Effects of Synbiotic supplementation on bone and metabolic health in Caucasian postmenopausal women: rationale and design of the OsteoPreP trial. Nutrients. (2024) 16:4219. doi: 10.3390/nu16234219
218. Wei, J, Ding, W, Song, K, Zhang, Y, Luo, Q, and Qi, C. Next-generation probiotics and engineered BEVs for precision therapeutics in osteoporosis. Front Nutr. (2025) 12:1581971. doi: 10.3389/fnut.2025.1581971
219. Cuamatzin-García, L, Rodríguez-Rugarcía, P, El-Kassis, EG, Galicia, G, Meza-Jiménez, MDL, Baños-Lara, MDR, et al. Traditional fermented foods and beverages from around the world and their health benefits. Microorganisms. (2022) 10:1151. doi: 10.3390/microorganisms10061151
220. Hoppner, K, and Lampi, B. Total Folate, Pantothenic Acid and Biotin Content of Yogurt Products. Canadian Institute of Food Science and Technology Journal. (1990) 23:223–5. doi: 10.1016/S0315-5463(90)70247-7
221. Fabian, E, Majchrzak, D, Dieminger, B, Meyer, E, and Elmadfa, I. Influence of Probiotic and Conventional Yoghurt on the Status of Vitamins B1, B2 and B6 in Young Healthy Women. Ann Nutr Metab. (2008) 52:29–36. doi: 10.1159/000114408
222. Laiño, JE, Juarez Del Valle, M, Savoy De Giori, G, and LeBlanc, JGJ. Development of a high folate concentration yogurt naturally bio-enriched using selected lactic acid bacteria. LWT - Food Science and Technology (2013) 54:1–5. doi: 10.1016/j.lwt.2013.05.035
223. Deshwal, GK, Tiwari, S, Kumar, A, Raman, RK, and Kadyan, S. Review on factors affecting and control of post-acidification in yoghurt and related products. Trends in Food Science & Technology. (2021) 109:499–512. doi: 10.1016/j.tifs.2021.01.057
224. Kök-Taş, T, Seydim, AC, Özer, B, and Guzel-Seydim, ZB. Effects of different fermentation parameters on quality characteristics of kefir. Journal of Dairy Science. (2013) 96:780–9. doi: 10.3168/jds.2012-5753
225. Kneifel, W, and Mayer, HK. Vitamin profiles of kefirs made from milks of different species. Int J of Food Sci Tech. (1991) 26:423–8. doi: 10.1111/j.1365-2621.1991.tb01985.x
226. Tarvainen, M, Fabritius, M, and Yang, B. Determination of vitamin K composition of fermented food. Food Chemistry. (2019) 275:515–22.
227. Jeong, SH, Jung, JY, Lee, SH, Jin, HM, and Jeon, CO. Microbial succession and metabolite changes during fermentation of dongchimi, traditional Korean watery kimchi. International Journal of Food Microbiology. (2013) 164:46–53. doi: 10.1016/j.ijfoodmicro.2013.03.016
228. Jung, SJ, Kim, MJ, and Chae, SW. Quality and functional characteristics of kimchi made with organically cultivated young Chinese cabbage (olgari-baechu). Journal of Ethnic Foods. (2016) 3:150–8. doi: 10.1016/j.jef.2016.05.003
229. Du, R, Song, G, Zhao, D, Sun, J, Ping, W, and Ge, J. Lactobacillus casei starter culture improves vitamin content, increases acidity and decreases nitrite concentration during sauerkraut fermentation. Int J of Food Sci Tech. (2018) 53:1925–31. doi: 10.1111/ijfs.13779
230. Yang, X, Hu, W, Xiu, Z, Jiang, A, Yang, X, Saren, G, et al. Microbial Community Dynamics and Metabolome Changes During Spontaneous Fermentation of Northeast Sauerkraut From Different Households. Front Microbiol. (2020) 11:1878. doi: 10.3389/fmicb.2020.01878
231. Gänzle, MG, Loponen, J, and Gobbetti, M. Proteolysis in sourdough fermentations: mechanisms and potential for improved bread quality. Trends in Food Science & Technology. (2008) 19:513–21. doi: 10.1016/j.tifs.2008.04.002
232. Van Kerrebroeck, S, Bastos, FCC, Harth, H, and De Vuyst, L. A low pH does not determine the community dynamics of spontaneously developed backslopped liquid wheat sourdoughs but does influence their metabolite kinetics. International Journal of Food Microbiology. (2016) 239:54–64. doi: 10.1016/j.ijfoodmicro.2016.07.019
233. De Vuyst, L, Comasio, A, and Kerrebroeck, SV. Sourdough production: fermentation strategies, microbial ecology, and use of non-flour ingredients. Critical Reviews in Food Science and Nutrition. (2023) 63:2447–79. doi: 10.1080/10408398.2021.1976100
234. Xu, L, Cai, WX, and Xu, BJ. A Systematic Assesment on Vitamins (B2, B12) and GABA Profiles in Fermented Soy Products Marketed in China: VITAMINS B2, B12 AND GABA IN FERMENTED SOY PRODUCTS. Journal of Food Processing and Preservation. (2017) 41:e13126. doi: 10.1111/jfpp.13126
235. Tanaka, R, and Tanaka, R. Development of vitamin K analysis method using column switching high-performance liquid chromatography method and analysis results of various food items for vitamin K content. Food Anal Methods. (2024) 17:1218–28. doi: 10.1007/s12161-024-02643-4
236. Astuti, M, Meliala, A, Dalais, FS, and Wahlqvist, ML. Tempe, a nutritious and healthy food from Indonesia. Asia Pac J Clin Nutr. (2000) 9:322–5. doi: 10.1046/j.1440-6047.2000.00176.x
237. Azeke, MA, Greiner, R, and Jany, K-D. PURIFICATION AND CHARACTERIZATION OF TWO INTRACELLULAR PHYTASES FROM THE TEMPEH FUNGUS RHIZOPUS OLIGOSPORUS: TWO INTRACELLULAR PHYTASES FROM RHIZOPUS OLIGOSPORUS. Journal of Food Biochemistry. (2011) 35:213–27. doi: 10.1111/j.1745-4514.2010.00377.x
238. Marui, J, Boulom, S, Panthavee, W, Momma, M, Kusumoto, K-I, Nakahara, K, et al. Culture-independent bacterial community analysis of the salty-fermented fish paste products of Thailand and Laos. Bioscience of Microbiota, Food and Health. (2015) 34:45–52. doi: 10.12938/bmfh.2014-018
239. Cai, H, Tao, L, Zhou, X, Liu, Y, Sun, D, Ma, Q, et al. Lactic acid bacteria in fermented fish: Enhancing flavor and ensuring safety. Journal of Agriculture and Food Research. (2024) 16:101206. doi: 10.1016/j.jafr.2024.101206
240. Yamanaka, T, Katsuura, K, Koseki, K, Bito, T, Prangthip, P, Umebayashi, Y, et al. Characterization of Vitamin B12 Compounds from Commercially Available Fish Sauce Products. ACS Food Sci Technol. (2023) 3:1196–202. doi: 10.1021/acsfoodscitech.3c00128
Keywords: fermented foods, bone health, phytase, bio-peptide, vitamin K, gut-bone
Citation: Tamang JP, Agahi F, Yilmaz B, Künili İE, Mardon J, Bulmus-Tuccar T, Torbica A, Nikolovska Nedelkoska D, Kütt M-L, Malagón-Rojas J, Parada MA, Mayo B and Frias J (2025) Characterization of fermented foods: bone health. Front. Nutr. 12:1648775. doi: 10.3389/fnut.2025.1648775
Edited by:
Lee Sin Chang, USCI University, MalaysiaReviewed by:
Tanu Tomar, Lovely Professional University, IndiaAlice Njolke Mafe, Taraba State University, Nigeria
Copyright © 2025 Tamang, Agahi, Yilmaz, Künili, Mardon, Bulmus-Tuccar, Torbica, Nikolovska Nedelkoska, Kütt, Malagón-Rojas, Parada, Mayo and Frias. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fojan Agahi, Zm9qYW4uYWdhaGlAdXYuZXM=; Julie Mardon, anVsaWUubWFyZG9uQHZldGFncm8tc3VwLmZy