 Cláudia S. G. P. Pereira
Cláudia S. G. P. Pereira M. Carpena
M. Carpena João C. M. Barreira
João C. M. Barreira M. A. Prieto
M. A. Prieto M. Beatriz P. P. Oliveira
M. Beatriz P. P. Oliveira- 1LAQV/REQUIMTE, Department of Chemical Sciences, Faculty of Pharmacy, University of Porto, Porto, Portugal
- 2Department of Analytical Chemistry and Food Science, Instituto de Agroecoloxía e Alimentación (IAA) - CITEXVI, Universidade de Vigo, Nutrition and Bromatology Group, Vigo, Spain
- 3Mountain Research Centre (CIMO), ESA, Polytechnic Institute of Bragança, Campus de Santa Apolónia, Bragança, Portugal
Viruses remain a major global health challenge due to their strict dependence on host cell machinery and limited therapeutic options. Hydrocolloids (natural and semisynthetic) have gained attention as promising scaffolds for antiviral drugs discovery. Their structural variability, biocompatibility, and low toxicity enable diverse mechanisms of action, including inhibition of viral attachment and entry, disruption of replication, immunomodulation, and in some cases direct virucidal effects. This review examines the antiviral activity of hydrocolloids from three main sources: algal (agar, alginate, carrageenan, fucoidan, laminarin, and ulvan); animal (chitin, chitosan, chondroitin sulphate, dermatan sulphate, keratan sulphate, heparin, heparan sulphate, glycogen, and hyaluronan); and plant (pectin derivatives, starch derivatives, and locust bean gum). Across these groups, antiviral efficacy is strongly modulated by structural determinants such as molecular weight, degree and distribution of sulphation, glycosidic linkages, and branching patterns. Sulphated polysaccharides, in particular, exhibit broad-spectrum activity by blocking early infection steps through electrostatic interactions with viral proteins. Despite their potential, challenges persist, including structural heterogeneity, lack of viral specificity, and anticoagulant side effects in certain sulphated derivatives. Strategies to overcome these limitations include chemical modification, development of semisynthetic derivatives, and nanomaterial engineering to enhance stability, bioavailability, and therapeutic precision. Overall, hydrocolloids represent a versatile and underexplored platform for antiviral therapeutics. Continued efforts toward structural optimization, mechanistic elucidation, and clinical translation are critical to unlock their full potential against current and emerging viral threats.
1 Introduction
Viruses are characterized by a relatively simple architecture, consisting of a protein capsid that encapsulates their genetic (either DNA or RNA) material, together with viral enzymes and, in some cases, an outer lipid envelope. Since they lack the biochemical machinery for autonomous replication, viruses rely entirely on the host cell’s system, making them mandatory intracellular parasites. This hinders the development of virucidal therapies, as effective therapeutic agents must selectively block viral processes without damaging host cells. Accordingly, molecules capable of interfering with one or more stages of the viral replication cycle, thereby halting the onset and/or progression of infection while guaranteeing selective toxicity toward the pathogen without harming host cells, are considered effective antiviral candidates (1).
Moreover, viruses are wide-spread etiological agents, and current antiviral therapies, although valuable, often face challenges such as rapid emergence of resistant strains, limited efficacy across viral families, and potential host toxicity, emphasizing the urgent demand for new antiviral strategies (2). This has created interest in naturally derived biomolecules that may act through broad-spectrum and multitarget mechanisms (3).
Among the potential sources of antiviral compounds, hydrocolloids have emerged as particularly promising candidates due to their high structural variability, wide-ranging biological functions, biocompatibility, and low toxicity. These polysaccharides have increasingly been investigated in their natural state as well as in chemically modified forms designed to enhance their inhibitory properties against clinically significant viruses (4). Their capacity to act through distinct mechanisms of action further highlights their potential as promising candidates as scaffolds to novel antiviral agents (5).
Research has consistently demonstrated that the antiviral properties of hydrocolloids are strongly correlated to the presence of anionic groups, particularly sulphate moieties, which enable electrostatic interactions with viral envelope proteins and host cell receptors (Figure 1), preventing viruses from attaching to host cells and hindering the initial stages of viral infection (6). Moreover, there are additional structural determinants that modulate the antiviral activity of hydrocolloids:
(a) Glycosidic linkages - the type of glycosidic bond can determine the ability of hydrocolloids to inhibit viral enzymes (e.g., proteases), induce viral aggregation, or exert immunomodulatory effects. For instance, β-glucans, which feature β-(1 → 3) and β-(1 → 4) linkages, exhibit potent antiviral effects by enabling binding to immune receptors and stimulating immune responses (e.g., interferon production), leading to enhanced viral defence mechanisms. Conversely, α-glucans with α-(1 → 4) glycosidic linkages, such as starch, lack comparable antiviral activity (7).
(b) Molecular weight - generally, hydrocolloids with higher molecular weight display stronger antiviral effects, as they can form extended networks with viral particles and inhibit infection. High molecular weight hyaluronan, for instance, has demonstrated antiviral activity by blocking viral adhesion to host cells and interfering with the entry of enveloped viruses (7).
(c) Branching patterns - branched hydrocolloids often possess higher structural complexity, increasing their ability to disrupt viral processes. Fucoidans, for example, which are highly branched, exhibit notable antiviral activity by interfering with viral adhesion, entry, and fusion (8).
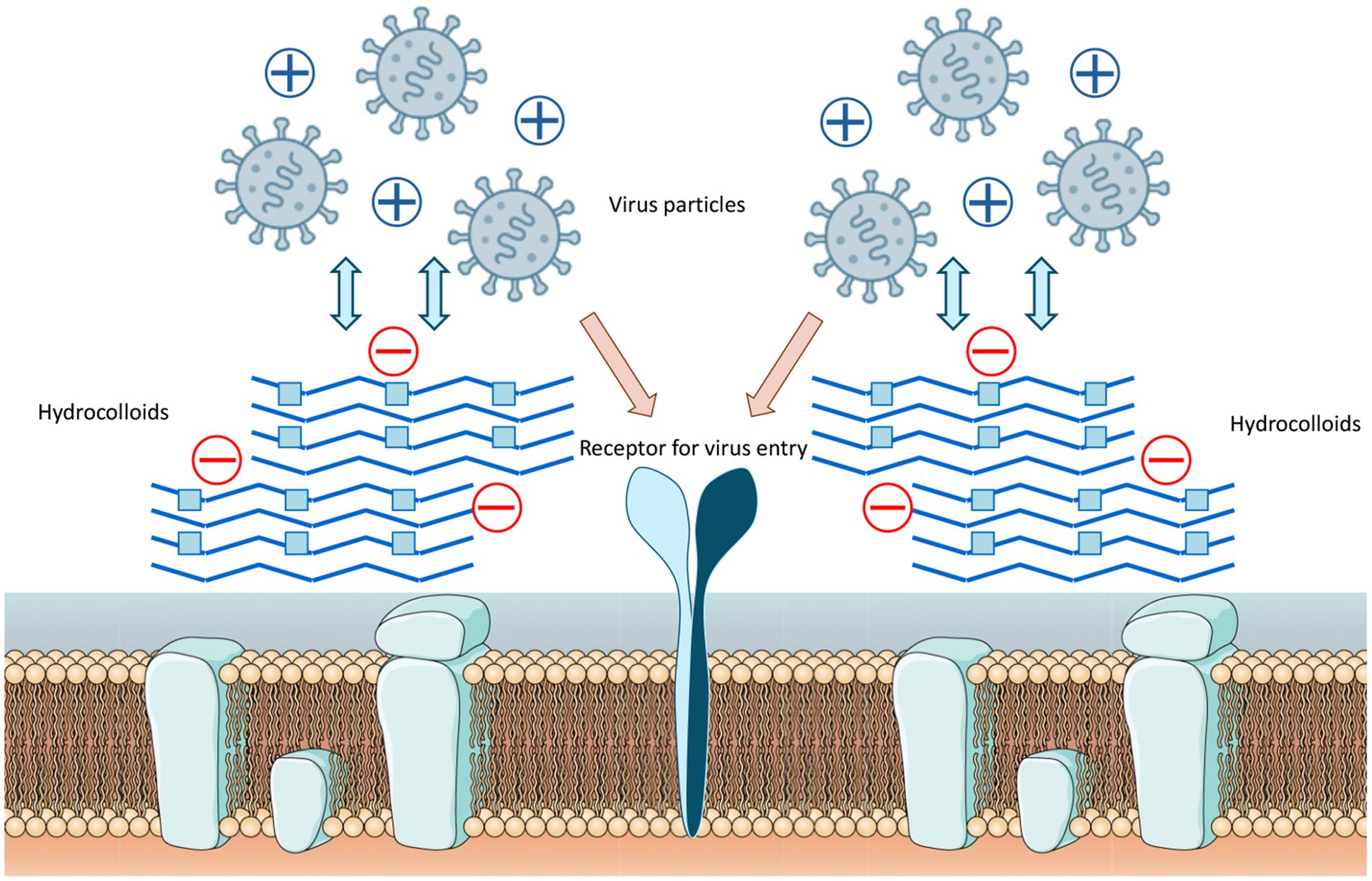
Figure 1. Mechanism of hydrocolloid-mediated viral inhibition by electrostatic interaction. Negatively charged sulphated groups (red circles) interact electrostatically with positively charged amino acids on viral surface proteins, preventing binding to host cell receptors and blocking entry.
Overall, antiviral performance is modulated by parameters such as molecular weight, degree of sulfation, charge density, and conformational flexibility, which enable hydrocolloids to interfere with viral attachment and entry, stimulate immune responses, or directly inhibit replication (9). Likewise, chemical and/or enzymatic modifications (e.g., sulfation, carboxylation, or acetylation) can further strengthen these properties, producing semisynthetic derivatives with enhanced antiviral activity. Depending on these structural features, hydrocolloids may inhibit multiple stages of the virus cycle (Figure 2), including internalization, uncoating, and transcription phases, or even display direct virucidal effects (6). Different antiviral mechanisms (Figure 3) have been described:
(a) Blocking viral attachment and entry - sulphated polysaccharides, such as ulvan, fucoidan, or chondroitin sulphate, can effectively prevent viruses from binding and entering host cells during the initial stages of infection, while showing minimal toxicity at therapeutic doses (10).
(b) Inhibiting viral replication - several hydrocolloids (e.g., chitosan, heparin, ulvan, fucoidan, carrageenan) were shown to be capable of disrupting viral replication, as documented in studies involving herpes simplex virus types 1 and 2 (HSV-1 and HSV-2) and human immunodeficiency virus (HIV) (2).
(c) Immunomodulatory effects - some hydrocolloids (such as heparan sulphate, hyaluronan, alginate, carrageenan, fucoidan, laminarin, ulvan) modulate immune responses affecting lymphocyte proliferation, macrophage activity, inducing increased serum levels of immune-regulating cytokines (IL-2, TNF-α, and IFN-γ), or by altering spleen and thymus parameters (2).
(d) Direct viral inactivation - sulphated hydrocolloids such as alginate demonstrate the capacity to bind virus particles, inhibit early infection steps, and down-regulating host signaling pathways, effectively suppressing viral propagation while maintaining low cytotoxicity (11).
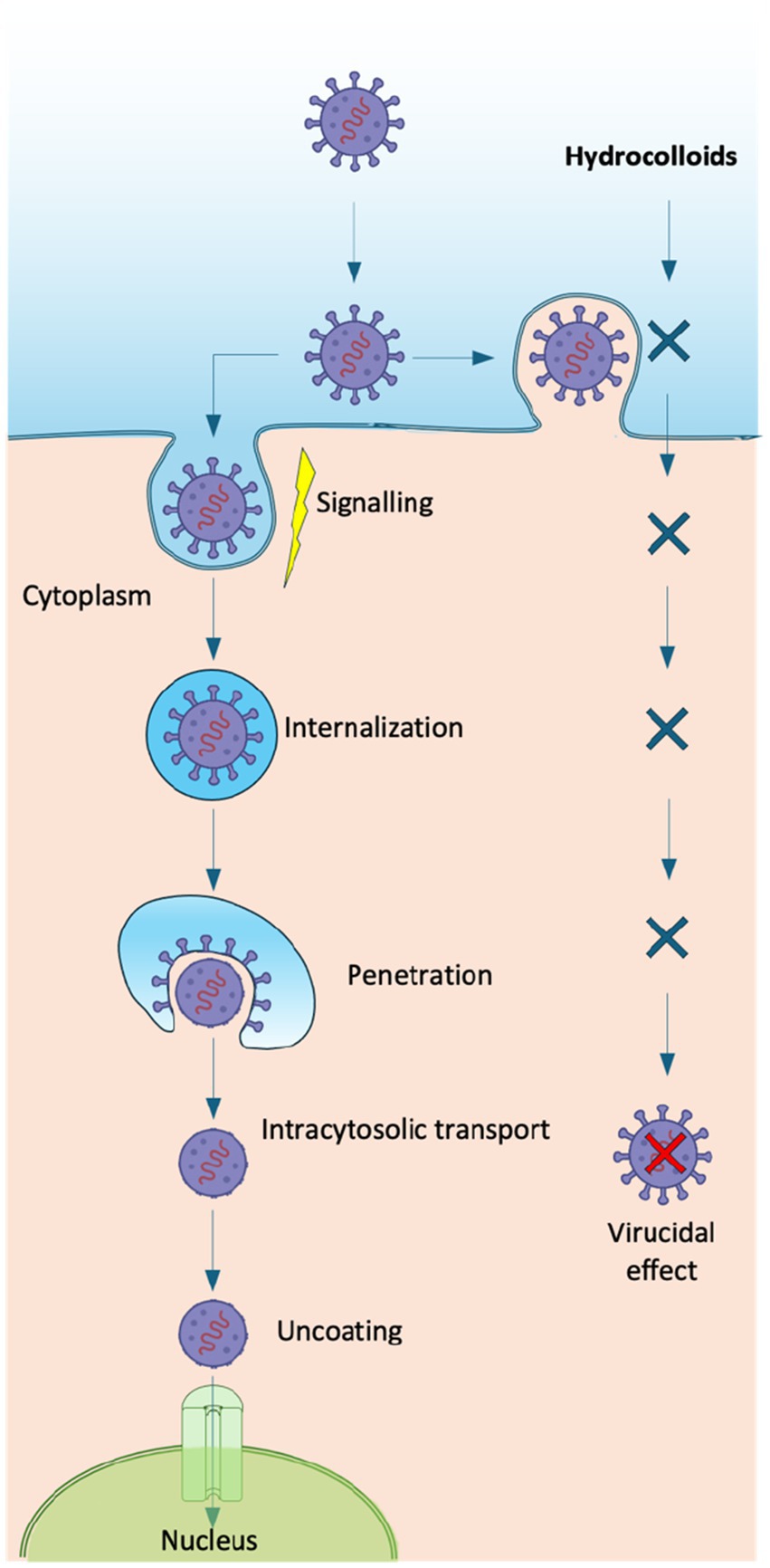
Figure 2. Viral infection pathway and inhibitory role of hydrocolloids. Hydrocolloids block viral attachment, entry, and replication steps, leading to a virucidal effect and preventing the release of the viral genome into the host cell nucleus.
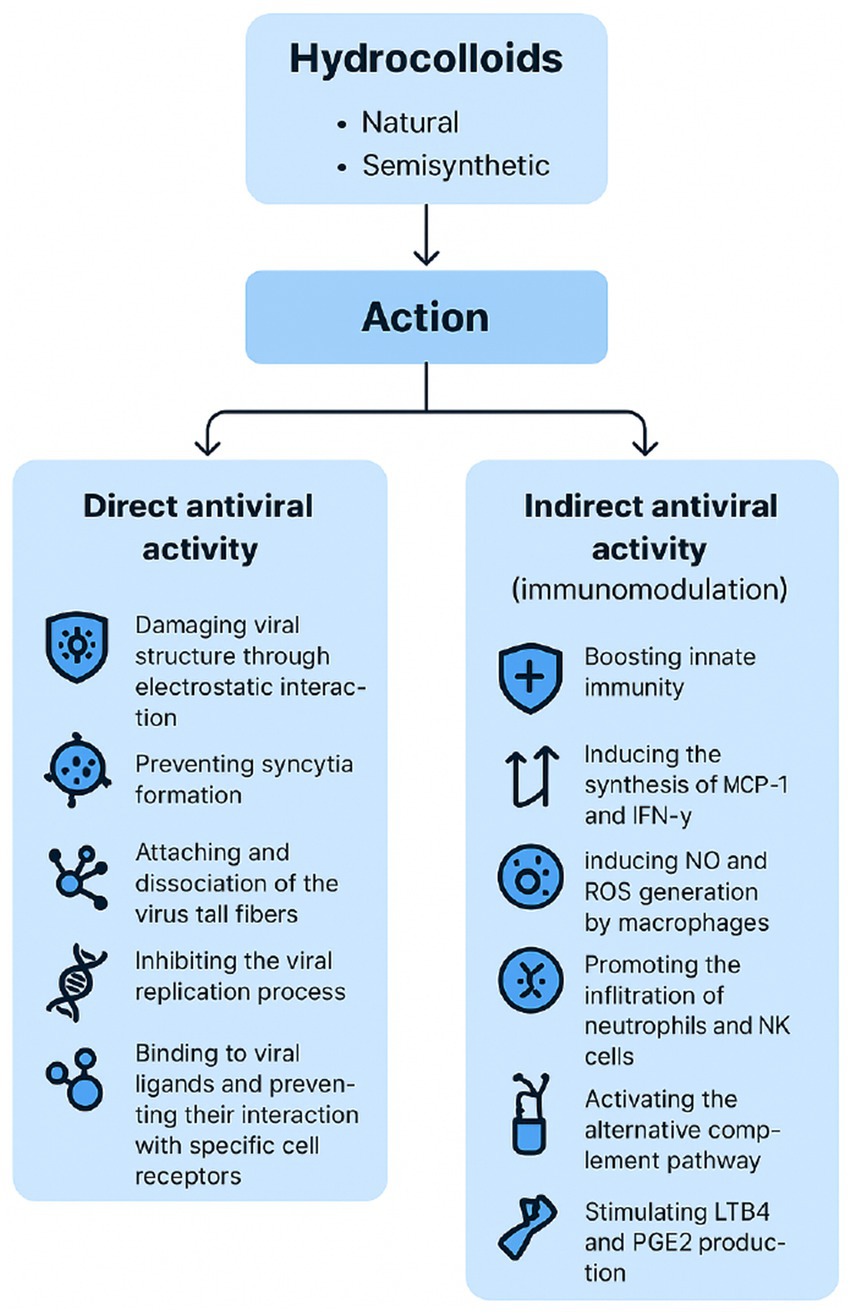
Figure 3. Main direct and indirect antiviral mechanisms of natural and semisynthetic hydrocolloids. Directly, hydrocolloids may block viral attachment and entry, inhibit viral replication, or inactivate viral particles, while is indirect action occurs mainly by modulating host immune responses. These mechanisms are influenced by structural determinants such as sulphation, branching, and molecular weight, which in turn affect clinical translation potential.
The former properties of hydrocolloids are particularly noteworthy, given the limited number of approved antiviral drugs, even in a scenario in which viral diseases are still causing significant morbidity and mortality (12). Consequently, screening hydrocolloids and other molecules with antiviral activity represents a valuable approach to discover new drug candidates and strengthening the available pool of antiviral treatments (13).
Additionally, and unlike small-molecule antivirals, hydrocolloids often exhibit heterogeneous structures that hinder reproducibility, standardization, and generalised use. Moreover, despite compelling in vitro evidence, translation into in vivo systems or clinical models is often inconsistent (1).
This review provides a critical overview of hydrocolloids of algal, animal, and plant origin, either in their native forms or as chemically modified derivatives. For each analysed hydrocolloid, the structural features, reported antiviral spectra, mechanisms of action, strengths, weaknesses and translational potential, are systematically described.
2 Hydrocolloids from algae sources
Algae are a rich source of structurally diverse hydrocolloids, several of which exhibiting notable antiviral activity, which is largely determined by sulphation patterns, molecular weight, and branching frequency. Below, six major algal hydrocolloids are compared regarding the former properties, being also evaluated for their strengths and limitations for therapeutic application.
2.1 Agar
2.1.1 Structure
Agar consists primarily of two fractions: agarose and agaropectin. Agarose, corresponding to about 70% of the polysaccharide, is a linear polymer built from agarobiose (1,3-linked β-D-galactopyranose) and neoagarobiose (1,4-linked 3,6-anhydro-α-L-galactopyranose) units, which often contain alternating sulphate groups. Agaropectin, in contrast, accounts for around 30% of the mixture and is a more heterogeneous fraction, being characterized by a lower frequency of alternating D- and L-galactose residues with acidic side chains such as sulphate and pyruvate, or methoxy groups (9). Agarans, therefore, are commonly found in two fractions: neutral agarose and charged agaropectin, which are differentiated by their ionic composition (4).
2.1.2 Occurrence and antiviral spectrum
The sulphate residues play a crucial role in mediating the antiviral activity, such as it has been observed with the sulphated agarans isolated from Bostrychia montagnei and Acanthophora spicifera, which demonstrated potent inhibition of HSV-1 and HSV-2, with the most active fractions being those enriched in sulphate (Table 1) (4). Beyond sulphated derivatives, agar itself has also shown antiviral potential. When incorporated into food coatings, it was able to reduce murine norovirus (MNV) infectivity, suggesting possible applications in food safety to mitigate the transmission of foodborne viruses (14). Agar was also shown to exert observable effect over plaque development, not only when tested against dengue virus (DENV), but also in assays conducted with encephalomyocarditis virus (ECMV) and Mengo encephalomyelitis virus (MEV), likely by blocking cell-virus interaction through immobilization of virus particles (15).
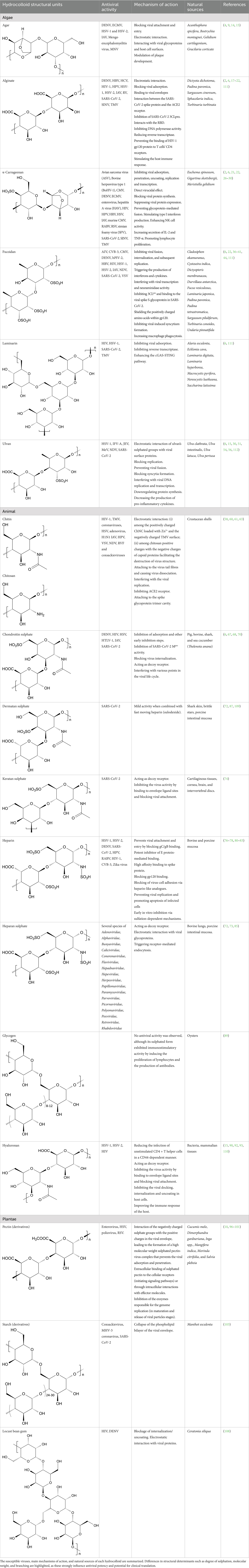
Table 1. Antiviral activity of natural and modified hydrocolloids from animal, algal, and plant sources.
2.1.3 Mechanism of action
Activity is linked to sulphate content, which promotes electrostatic binding to viral envelope proteins and prevents host cell attachment. In the case of MEV, activity was verified to occur by blocking cell-virus interaction through immobilization of virus particles (15).
2.1.4 Critical evaluation
Although agar derivatives demonstrate activity, they typically require higher concentrations than other algal polysaccharides to achieve measurable antiviral effects, raising concerns about their therapeutic relevance. Their greatest potential seems to be more associated to food safety applications such as antiviral coatings, translation into systemic therapy remains unlikely (4).
2.2 Alginate
2.2.1 Structure
Alginates are hydrophilic linear polysaccharides found in the cell walls of brown algae that consist of β-D-mannuronic acid (M), and α-L-glucuronic (G) residues linked through 1,4-glycosidic bonds, organized into homopolymeric (poly-G, poly-M blocks) or heteropolymeric (MG blocks) arrangements (16).
2.2.2 Occurrence and antiviral spectrum
Beyond their traditional industrial use in textiles and paper production, alginates are widely applied in food industry, cosmetics, and biomedical fields (mainly due to its polyanionic structure, which enable many of their biological properties, including antiviral activity). Several species of brown algae, including Padina pavonica, Sargassum cinereum, Turbinaria turbinata, Sphacelaria indica, and Dictyota dichotoma, have yielded alginates with activity (Table 1) against severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) - particularly noticeable in the alginates derived from D. dichotoma, against rubella virus (RV), HSV-1, HIV, and hepatitis B virus (HBV) and also, although in less extent, against influenza A virus (IAV), hepatitis C virus (HCV), and Sindbis virus (SINV) (14, 17–22).
2.2.3 Mechanism of action
Their antiviral potential has been attributed to different mechanisms of action: electrostatic interactions with viral envelopes, inhibition of essential viral enzymes, interference with virus-receptor binding, and modulation of host immune responses (14, 17, 18). Prominently, one of the alginate-derived compounds tested against HIV was shown to block viral adsorption to host cells while also inhibiting reverse transcriptase activity (21).
2.2.4 Critical evaluation
Overall, alginates display broad-spectrum activity, often through electrostatic interactions and enzyme inhibition, and are generally biocompatible and low in cytotoxicity, which is advantageous for clinical translation. However, their efficacy is inconsistent across viral families, and in vivo pharmacokinetics remain underexplored. Structural modifications enhance activity, but imply increased structural complexity, which may hinder reproducibility drug development (14).
2.3 Carrageenan
2.3.1 Structure
Carrageenans are linear sulphated galactans consisting of alternating 1,3-linked β-D-galactopyranose and 1,4-linked α-galactopyranose residues. The three main structural types - κ, ι, and λ - differ in the number and position of sulphate ester substituents and in the presence of 3,6-anhydro-D-galactopyranose units: κ-carrageenan carries a single sulphate ester group (25–30%), ι-carrageenan contains two (28–30%), while λ-carrageenan has three (32–39%) (23, 24).
2.3.2 Occurrence and antiviral spectrum
Carrageenans have demonstrated broad-spectrum antiviral properties (Table 1). For instance, carrageenans isolated from Gigartina skottsbergii effectively inhibited the transmission of human papillomavirus (HPV), while also suppressing HSV-1 and HSV-2 and human rhinovirus (HRV) (22, 25). Similarly, carrageenan derived from Meristiella gelidium inhibited HSV activity in vitro murine cytomegalovirus (CMV) in vivo (5).
Likewise, low-molecular-weight κ-carrageenan has been reported to inhibit IAV, HSV-2 and HPV16, respiratory syncytial virus (RSV) (26–30), SARS-CoV-2 (31), and HIV (22).
Moreover, λ-carrageenan from the red alga Gigantina skotsbergii exhibited significant inhibitory effects on rabies virus (RABV), SARS-CoV-2 and IAV (32), while ι-carrageenan from Euchema spinosum has been demonstrated to neutralize SARS-CoV-2 in a concentration-dependent manner, thereby reducing both transmission and progression of infection into the lower respiratory tract (33). In addition, both ι- and λ-carrageenan inhibited DENV replication in Vero and HepG2 cells (21).
2.3.3 Mechanism of action
As with other sulphated hydrocolloids, both the degree of sulphation and the molecular weight are determinant factors for the antiviral activity of carrageenans. However, antiviral efficacy is not strictly proportional to the overall sulphation level; instead, the specific position and spatial arrangement of sulphate groups on the backbone are key factors for antiviral capacity (30). Carrageenans have been shown to act primarily by blocking the initial interactions between viral particles and host cell receptors, inhibiting its adsorption and internalization, and suppressing viral protein expression (25). Carrageenans also display immunomodulatory activity, such as stimulating type I interferon production, enhancing natural killer (NK) cell activity, increasing secretion of IL-2 and TNF-α, and promoting lymphocyte proliferation (21).
2.3.4 Critical evaluation
Interestingly, carrageenan is one of the few hydrocolloids tested in clinical formulations, such as nasal sprays against respiratory viruses, besides displaying broad-spectrum activity. In turn, it holds potential anticoagulant effects and strain variability, which have produced mixed results in clinical translational trials, underscoring the need for controlled head-to-head studies (5).
2.4 Fucoidan
2.4.1 Structure
Fucoidans represent a diverse group of sulphated fucans that are primarily composed of fucose (40–80%), sulphate esters, and uronic acids, along with smaller proportions of other polysaccharides such as alginate and laminarin (34). Among seaweed-derived polysaccharides, fucoidans are regarded as the most structurally heterogeneous, with high variation in both molecular structure and weight (100 to 5,200 kDa) depending on the species (34). Typically, fucoidan constitutes about 25–30% of the dry biomass of brown algae. Its backbone generally consists of α-(1 → 3)-linked L-fucopyranose units or alternating α-(1 → 3) and α-(1 → 4)-L-fucopyranosyl residues. These chains can be further substituted with sulphate or acetate groups and may contain glycosyl side groups such as glucuronic acid, in addition to minor amounts of monosaccharides including D-xylose, D-galactose, D-mannose, and uronic acids (35).
2.4.2 Occurrence and antiviral spectrum
These structurally complex sulphated polysaccharides from brown algae, are well known for their strong anti-inflammatory properties and broad antiviral activity (Table 1), which extends to both RNA and DNA viruses, such as IAV, HSV, HBV, HIV, and vesicular stomatitis virus (VSV) (22, 36). Extracts from Undaria pinnatifida and Kjellmaniella crassifolia were shown to attenuate the effects of IAV by interfering with viral transcription and neuraminidase activity. Fucoidans from Fucus vesiculosus and Laminaria japonica were reported to inhibit parainfluenza virus type 3 and avian influenza virus (AIV) infection, respectively (21), while that derived from Cladosiphon okamuranus suppressed human parainfluenza virus type 2 (hPIV-2) infection in LLC-MK₂ cells (37). Moreover, fucoidans derived from Padina tetrastromatica and Turbinaria conoides were found to significantly reduce SARS-CoV-2 replication in host cells by inhibiting 3CLpro (up to 97%), although without affecting RBD-spike binding (RBD) (38). Likewise, sulphated fucoidans from Dictyota mertensii, Lobophora variegata, Fucus vesiculosus, and Spatoglossum schroederi inhibited HIV replication (39), while those extracted from Padina pavonica, Dictyopteris membranacea, and Fucus evanescens, were active against HSV-1, HSV-2 and coxsackievirus B3 (CVB-3) (40–42).
2.4.3 Mechanism of action
Fucoidans exert their antiviral activity by binding to viral and host proteins, thereby inhibiting viral fusion, internalization, and subsequent replication. Again, the degree of sulphation and branching pattern strongly influence their antiviral efficacy. In addition, fucoidan can also display immunomodulatory effects, such as inducing interferons and cytokines, further supporting host antiviral defence (43). Additional studies demonstrated that fucoidan can also bind to the viral spike S-glycoprotein, thereby preventing SARS-CoV-2 entry into host cells (44), being also active against HIV, either by inhibiting reverse transcriptase (39), or by shielding the positively charged amino acids within gp120 (the viral envelope glycoprotein) through interactions with specific sulphate groups (21).
2.4.4 Critical evaluation
In general, fucoidans exhibit remarkable diversity and strong activity against influenza, HIV, and coronaviruses. However, their structural complexity is a double-edged sword: while varied sulphation enhances antiviral potency, it hinders reproducibility and standardization. Furthermore, in most cases, human pharmacokinetic and safety data are limited, slowing its clinical progress (5).
2.5 Laminarin
2.5.1 Structure
Laminarin, sometimes referred to as laminaran, is a storage glucan naturally present in brown algae. It typically consists of 20–25 glucose residues linked by β-(1 → 3) glycosidic bonds, with occasional β-(1 → 6)-linked side chains. The branching degree can vary depending on environmental factors and the harvesting period of algae (45). There are two types of polymeric chains within laminarin: M-type chains, which terminate with a mannitol residue at the reducing end, and G-type chains, which end with a glucose unit (46).
2.5.2 Occurrence and antiviral spectrum
Laminarin is commonly extracted from species belonging to the Eisenia, Laminaria, Sargassum, and Saccharina genera, and has been associated with diverse antiviral activities (Table 1). For example, laminarin extracted from kelp was active against HIV, HSV-1 tobacco mosaic virus (TMV) (6, 15, 47, 48).
2.5.3 Mechanism of action
Laminarin demonstrated capacity to inhibit both the adsorption and the reverse transcriptase of HIV (6), as well as capacity to enhance the cyclic GMP-AMP synthase stimulator of interferon genes (cGAS-STING), boosting the production of type I interferons in both human and murine cells in its action against viruses such as HSV-1 (48). Interestingly, this effect was markedly reduced in acetylated derivatives of laminarin, as confirmed by viral plaque assays and by analysing changes in viral DNA and protein synthesis (49).
2.5.4 Critical evaluation
In comparison to other hydrocolloids, laminarin stands out for enhancing host immune pathways, offering a unique immunomodulatory advantage. However, antiviral activity is relatively narrow in scope and often attenuated in chemically modified forms, potentially indicating a limited therapeutic relevance (50).
2.6 Ulvan
2.6.1 Structure
Ulvans are the major water-soluble polysaccharides present in the cell walls of green seaweeds. These heteropolysaccharides are characterized by a high degree of sulphation and a complex composition that includes L-rhamnose-3-sulphate, together with several neutral sugars such as xylose, glucose, mannose, galactose, arabinose, and structural polysaccharides like cellulose, xyloglucan, and glucuronan. Ulvans also contain uronic acids like glucuronic acid or iduronic acid (51).
Ulvan’s backbone usually consists of repeating disaccharide units of α-L-rhamnose-3-sulfate linked (1 → 4) to β-D-glucuronic acid (ulvano-biuronic acid A) or α-L-rhamnose-3-sulfate linked (1 → 4) to α-D-iduronic acid (ulvanobiuronic acid B); additionally, it also contains aldobiose fragments known as ulvanobioses (52).
2.6.2 Occurrence and antiviral spectrum
Experimental studies have confirmed its activity against several viruses. For example, ulvan from Ulva pertusa was shown to block replication of VSV (15, 53), while ulvan isolated from Ulva clathrata inhibited Newcastle disease virus (NDV) in vitro by preventing viral fusion (6). Likewise, ulvan from Ulva latuca demonstrated strong inhibitory activity against SARS-CoV-2 (54, 55), while that from Ulva intestinalis was active against measles virus (MeV), HSV-1, and Japanese encephalitis virus (JEV) (6, 21, 30, 56).
2.6.3 Mechanism of action
The antiviral effects (Table 1) of ulvan are generally linked to its negative charge, which enables interactions with viral surface proteins and thereby interferes with the early stages of viral infection, particularly viral adsorption and entry. These inhibitory properties are also influenced by the molecular weight of the polysaccharide (51). The inhibitory activity against SARS-CoV-2 was demonstrated to occur through multiple mechanisms: electrostatic interaction of ulvan’s sulphated groups with viral surface proteins (hindering viral adsorption), suppression of viral replication, and direct virucidal effects (54, 55). Ulvan also showed capacity to block syncytia formation (e.g., against MeV), interfere with viral DNA replication and transcription and downregulating protein synthesis (such as observed with HSV-1), and blocking virus adsorption by effectively decreasing the production of pro-inflammatory cytokines in JEV-infected glial cells (6, 21, 30, 56).
2.6.4 Critical evaluation
Ulvan’s multi-targeted activity (blocking entry, replication, and immune modulation) is promising. However, antiviral effects vary widely across species and extraction methods, hampering reproducibility. Furthermore, in vivo studies are essential before considering it for clinical translational (4).
3 Hydrocolloids from animal sources
Animal-derived hydrocolloids, particularly glycosaminoglycans, are well-characterized regarding their structural features, having also been extensively studied for their interactions with viral proteins. Yet, their potential as antivirals is balanced by constraints such as their anticoagulant activity and variability induced by their natural source.
3.1 Chitin and chitosan
3.1.1 Structure
Chitin is a linear polymer of N-acetyl-D-glucosamine (2-acetylamino-2-deoxy-D-glucose) and D-glucosamine (30), presenting a molecular weight that may vary among 1,030 and 2,500 kDa. Chitosan, in turn, consists of β-(1 → 4)-linked units of 2-acetylamino-2-deoxy-β-D-glucose and 2-amino-2-deoxy-β-D-glucose, reaching lower molecular weights (usually between 100 and 500 kDa) (57). Chitosan is also characterized by a lower degree of acetylation and for lacking inter-sheet hydrogen bonds, making it more hydrosoluble than chitin. Likewise, chitosan is soluble at pH values below 6.5, as the protonation of amino groups imparts polycationic characteristics (58).
3.1.2 Occurrence and antiviral spectrum
Chitin is typically found in the exoskeletons of crustaceans and insects, though it can also be obtained via fungal fermentation processes (30); chitosan is obtained by deacetylation of chitin found in crustaceans exoskeletons (59). They’ve both demonstrated capacity to fight viral infections (Table 1), as demonstrated against TMV, HIV-1, HSV, Friend murine leukaemia helper virus (F-MuLV), adenovirus (in NIH-3 T3 cells), IAV H1N1, HPV, VSV, NDV, SARS-CoV-2, Rift Valley fever virus (RVFV) and coxsackieviruses (assayed in Vero cells) (60, 61).
3.1.3 Mechanism of action
Chitin and chitosan can exert their activity by directly interacting with viruses, or by indirectly stimulating antiviral immune responses. Their direct antiviral action often involves electrostatic interactions between their cationic derivatives and the negatively charged viral envelope proteins, damaging viral structures, inactivating viruses after attaching to their tail fibres, or interfering with the viral replication process (60). Their sulphated derivatives blocked the interaction of HIV-1 gp120 with the CD4 receptor in cultured T cells, in addition to hinder viral uptake and cellular penetration (as observed with HSV-1 and F-MuLV) (58). In turn, the N-(2-hydroxypropyl)-3-trimethylammonium chitosan chloride (HTCC) derivative demonstrated activity against several coronaviruses, most likely by inhibiting their interaction with the angiotensin-converting enzyme 2 (ACE2) receptor (58), while quaternized chitosan derivatives were shown to destabilize viral surface proteins by interacting with viral capsid/envelope proteins (62).
Chitin was also used to produce chitin nanocrystals (ChNC), rod-shaped nanostructures with a high aspect ratio (much longer than wide) and surface carboxyl groups that enhance chelation and adsorption. When loaded with Zn2+, ChNCs significantly reduced viral infection, most likely due to electrostatic interactions between the positively charged Zn2+ on ChNCs and the negative charges on virus’ surface (63). In addition, chitosan was reported for preventing viral spread in plant tissues, by specifically activating resistance gene expression and eliciting defence responses (60).
3.1.4 Critical evaluation
Chitosan’s tunability through chemical modification is its greatest strength, enabling tailored derivatives with improved potency. However, their efficacy in vivo is significantly conditioned by the lack of standardized formulations, which hampers systematic in vitro studies and further clinical translation (64).
3.2 Glycosaminoglycans
Research into proteoglycans began in the early 20th century, with work focusing on “chondromucoid” from cartilage and anticoagulant extracts such as heparin from liver tissue. Between 1930 and 1960, major progress was made in identifying and characterizing the polysaccharide components of these substances (then often called mucopolysaccharides). That period led to the elucidation of different structures, such as hyaluronan, dermatan sulphate, keratan sulphate, the isomeric forms of chondroitin sulphate, heparan sulphate, and heparin. Collectively, these polysaccharides were renamed as glycosaminoglycans to emphasize the presence of amino sugars and related carbohydrate units arranged in polymeric chains. Subsequent studies revealed how these chains attach to core proteins within proteoglycans. Glycosaminoglycans are major components of animal connective tissues, and their sulphated derivatives are usually associated with significant antiviral activity (65).
3.2.1 Chondroitin sulphate
3.2.1.1 Structure
The basic unit of this complex glycosaminoglycan consists of a repeating disaccharide of glucuronic acid and N-acetyl-galactosamine linked by β-(1 → 3) bonds. These disaccharides are interconnected by β-(1 → 4) glycosidic bonds, forming a linear chain that reaches a molecular weight ranging between 5 and 50 kDa (66).
3.2.1.2 Occurrence and antiviral spectrum
Together with chitosan, chondroitin sulphate and heparin, two of the glycosaminoglycans described herein, are among the most investigated antiviral animal polysaccharides. Chondroitin sulphate (Table 1) is has shown potent inhibitory activity against HSV-1, DENV (demonstrated in Vero and BHK-21 cell lines) (67), SARS-CoV-2, and bovine coronavirus (BCoV) (68). Moreover, two fucosylated chondroitin sulphates isolated from the sea cucumbers Isostichopus badionotus and Pentacta pygmaea demonstrated strong activity against SARS-CoV-1 and SARS-CoV-2, as, to a lesser extent, against Middle East respiratory syndrome coronavirus (MERS-CoV), highlighting the potential of structurally modified derivatives as broad-spectrum antiviral agents (69).
3.2.1.3 Mechanism of action
The acknowledged effects of chondroitin sulphate include blocking virus internalization, acting as decoy receptors, and interfering with multiple stages of the viral life cycle. In all cases, the effectiveness is highly correlated with the density of sulphate groups and molecular weight (6). In some cases, such as observed with flavivirus, the mechanism of action appears to depend on specific carbohydrate determinants within the polysaccharide backbone, which may function as critical epitopes for the interaction with the virus (67). More recently, the mechanism of action against SARS-CoV-2 was also clarified: chondroitin sulphate can act through competitive inhibition of the viral main protease (Mpro), a key enzyme in the coronavirus replication cycle (70).
3.2.1.4 Critical evaluation
In general, chondroitin sulphate demonstrates promising antiviral activity, but the strong source-dependent variability (e.g., pig, bovine, shark, sea cucumber) complicates the reproducibility. Likewise, additional research is necessary to consistently define the structure-activity relationship.
3.2.2 Dermatan sulphate
3.2.2.1 Structure
Like chondroitin sulphate, dermatan sulphate is composed of approximately equimolar proportions of hexosamine, acetyl groups, uronic acids, and sulphate groups. Both glycosaminoglycans share N-acetyl-D-galactosamine (GalNAc) as building block, but they differ in the type of uronic acid: dermatan sulphate contains L-iduronic acid (IdoA), while chondroitin sulphate contains D-glucuronic acid (GlcA). In dermatan sulphate, the repeating disaccharide sequence is formed through alternating glycosidic linkages: β-(1 → 4) bonds connecting GalNAc to IdoA, and α-(1 → 3) bonds linking IdoA to GalNAc (71).
3.2.2.2 Occurrence and antiviral spectrum
Regarding its viral activity (Table 1), there are no recent papers (published after 2020 till submission of this review) reporting remarkable antiviral activity of dermatan sulphate (therefore the Mode of action subsection was not included herein). Nonetheless, when combined with fast-moving heparin (a mixture commercially known as sulodexide, which is clinically used to prevent and treat vascular diseases), dermatan sulphate improved the clinical outcomes of COVID-19 patients (72).
3.2.2.3 Critical evaluation
Evidence base is sparse, particularly in most recent years. The main translational potential of dermatan sulphate, although unlikely, may lie in combined therapies, such as the exemplified sulodexide, which has been explored in COVID-19 patients for anticoagulant and endothelial-protective effects.
3.2.3 Keratan sulphate
3.2.3.1 Structure
Keratan sulphate consists of repeating disaccharides units made up of D-galactose and N-acetylglucosamine-6-sulphate connected by β-(1 → 4) linkages. Unlike other glycosaminoglycans, keratan sulphate does not attach to the proteoglycan core protein through the typical tetrasaccharide linker. Instead, its three known forms are characterized by distinct attachment modes: in keratan sulphate I, the chains are linked by an asparagine residue through a complex N-glycan; in keratan sulphate II, the connection is through the N-acetylgalactosamine linked to serine or threonine; in keratan sulphate III, the linkage involves a mannose residue connected to serine or threonine (73).
3.2.3.2 Occurrence and antiviral spectrum
Many viruses including IAV, HSV, HIV, and members of the Coronaviridae family, including SARS-CoV-2, use glycosaminoglycans as initial attachment factors to host cells, facilitating their entry. Because of this role, glycosaminoglycans are typically characterized by high bioactivity in result of their interaction with cytokines, cell receptors, and components of the extracellular matrix. However, keratan sulphate, which was identified more recently compared to other glycosaminoglycans and is therefore less extensively studied, has scarce evidence of antiviral activity; even so, in samples obtained from bovine cornea, keratan sulphate has shown moderate activity against SARS-CoV-2 (74).
3.2.3.3 Mechanism of action
As observed in other glycosaminoglycans, keratan sulphate binds to viral surface proteins, interfere with the attachment and potentially blocking host-receptor interactions.
3.2.3.4 Critical evaluation
Keratan sulphate is largely understudied when compared to other glycosaminoglycans. In-depth systematic studies are necessary to determine its therapeutic value.
3.2.4 Heparin
3.2.4.1 Structure
Heparin is a highly sulphated glycosaminoglycan composed by repeating sulphated disaccharide units with prevalence of 2-O-sulphate-α-l-iduronic acid and 6-O-sulfate-N-sulfate-α-D-glucosamine linked by a 1,4-glycosidic bond as the most common unit (75).
3.2.4.2 Occurrence and antiviral spectrum
Heparin is typically derived from porcine intestinal mucosa or bovine lung. Beyond its well-established anticoagulant role, heparin has demonstrated broad-spectrum antiviral activity (Table 1). Early studies showed its activity against HIV, RABV, HSV-1, HSV-2, DENV-2, Zika virus, and human CMV (6, 76, 77).
3.2.4.3 Mechanism of action
Heparin acts by inhibiting the viral attachment (binding specific viral glycoproteins such as HIV gp120 and SARS-CoV-2 RBD spike protein), inhibiting viral enzymes, or supressing virus replication and promoting apoptosis of the infected cells (76–79). Mechanistic insights into heparin’s activity against SARS-CoV-2 suggest multiple antiviral modes of action, such as (i) allosterically hindering spike-ACE2 interactions, (ii) competing with host heparan sulphate proteoglycans that serve as viral coreceptors, and (iii) preventing spike cleavage by furin, thereby reducing viral entry efficiency (80). Heparin also inhibits the main protease (Mpro) of SARS-CoV-2, with low inhibition constant and (Ki < 7 nM) and half maximal inhibitory concentrations (IC50 < 8 nM) (81). It has been further shown to inhibit virus entry and replication in primary bronchial epithelial cells. Given that SARS-CoV-2 infection is initiated predominantly in the nasal epithelium, intranasal administration of heparin has been proposed as a potential strain-agnostic prophylactic strategy (82). Furthermore, its favourable skin permeability, suggests potential utility in topical or inhalation-based formulations, particularly on exposed areas and sites of primary viral entry, e.g., ACE2 rich epithelia of the eye (conjunctiva/lids), nasal cavity, and mouth (83).
3.2.4.4 Critical evaluation
Among glycosaminoglycans, heparin is the most advanced toward clinical application. However, its anticoagulant activity poses major risks for systemic use, limiting the clinical applications to topical or localized formulations (1).
3.2.5 Heparan sulphate
3.2.5.1 Structure
Unlike most glycosaminoglycans, which are modified primarily through O-sulphation, heparan sulphate also undergoes N-sulphation. This structural feature is determinant for its biological activity, as the sulphation of hydroxyl and amino groups on the N-acetylglucosamine units determines how heparan sulphate interacts with proteins, cytokines, and growth factors, ultimately ruling its bioactive roles (73).
From a structural perspective, heparan sulphate is composed of repeating disaccharide units containing N-acetylglucosamine and glucuronic acid. This is closely related to heparin, except for the fact that heparin contains iduronic acid instead of glucuronic acid as the hexuronic component. Heparan sulphate chains are covalently linked to the protein core of the proteoglycan through a serine residue, which is connected by a characteristic tetrasaccharide linker, consisting of one xylose, two galactose units and one glucuronic acid residue (84).
3.2.5.2 Occurrence and antiviral spectrum
Given its verified involvement in viral entry processes, heparan sulphate has been investigated as a potential antiviral target (Table 1). Several viruses, such as HIV-1, SINV, HPV, HSV, human CMV, JEV, DENV, SARS-CoV-2, and Zika virus, as well as members of Alphaviridae, Flaviviridae, Papillomaviridae, Parvoviridae, Picornaviridae, or Retroviridae, were described as using heparan sulphate as an attachment factor to promote the initial contact with host cells (61, 85).
3.2.5.3 Mechanism of action
Heparan sulphate fragments were used to coat viral attachment proteins, thereby preventing them from binding to host cells. However, the development of such therapy is challenging, because the preparation of these fragments is demanding and there is a limited availability of this type of molecules from natural sources (85). When assayed against HIV, heparan sulfate demonstrated high-affinity binding to gp120, leading to an increased concentration of the virus at the cell surface and promoting interactions with CD4 and the co-receptors CCR5 and CXCR4, thereby acting as a competitive inhibitor of viral attachment. Heparan sulphate showed similar interaction with other viral envelope glycoproteins (e.g., HSV, Zika, SARS-CoV-2, and DENV), in all cases reducing infectivity by triggering receptor-mediated endocytosis through additional cellular receptors. A similar effect was observed in non-enveloped viruses such as HPV, where heparan sulphate interacted with the capsid protein L1, triggering conformational changes that promote binding to specific entry receptors and subsequent internalization through endocytosis (72, 86).
In either case, the sulphation pattern, particularly at the 4-O and 6-O of N-acetylglucosamine, have been shown to be crucial for the inhibitory effect of heparan sulphate (87).
3.2.5.4 Critical evaluation
Heparan sulphate act as co-factor for the infection of a diverse array of RNA and DNA viruses, making it a potential target for broad-spectrum antiviral strategies. However, the structural heterogeneity remains a barrier to therapeutic development.
Among glycosaminoglycans, chondroitin sulphate stands out for its potent activity against HSV and SARS-CoV-2 protease; however, its interspecies variability (e.g., pig, shark, sea cucumber) has been hindering an effective clinical translation. In contrast, interest in the antiviral potential of dermatan and keratan sulphates has declined, likely because both compounds seem considerably weaker than heparin. By comparison, heparin and heparan sulphate remain the most promising candidates for clinical translational, with well-validated mechanisms of action; nonetheless, their anticoagulant side effects suggest that localized administration (e.g., intranasal) is preferable to systemic use (1).
3.3 Glycogen
3.3.1 Structure
Glycogen is a highly branched glucose homopolysaccharide linked via α-1,4-glycosidic bonds, with branch points (usually every 8–12 glucose units) formed by α-1,6-glycosidic linkages.
3.3.2 Occurrence and antiviral spectrum
Glycogen is one of the primary carbon and energy reserves in living systems. Despite its importance in metabolism, native glycogen lacks the specific structural features necessary for strong antiviral interactions and therefore exhibits limited activity against viral infections. In turn, chemically modified glycogen derivatives have been shown to exert a variety of biological activities, including immunomodulatory, antitumour, and antiviral activities (88), as demonstrated by its sulphated derivatives against CMV and DENV, as well as stimulation of lymphocyte proliferation (Table 1). In addition to sulphated derivatives, carboxymethylated and phosphorylated forms were also reported to possess potent immunostimulatory properties (89).
3.3.3 Mechanism of action
Sulphated glycogen can attach to surface proteins, interfering with viral binding, in addition to enhancing immune response.
3.3.4 Critical evaluation
In general, native glycogen is largely inactive, requiring extensive chemical modification to display antiviral effects, raising challenges for scaling-up and regulatory approval. In turn, its sulphated derivatives exhibit strong immunomodulatory potential, although it remains poorly characterized.
3.4 Hyaluronan
3.4.1 Structure
Hyaluronan consists of alternating units of β-1,4-D-glucuronic acid and β-1,3-N-acetyl-D-glucosamine (90), presenting varying molecular weight depending on the tissue where it is located (91).
3.4.2 Occurrence and antiviral spectrum
This non-sulphated glycosaminoglycan is an essential component of both extracellular and pericellular matrices of many tissues in the body, including the lungs, where it contributes to development, tissue homeostasis, and injury repair processes (92). It is the only glycosaminoglycan biosynthesized directly at the cell membrane and is not covalently attached to proteoglycans. The antiviral activity of hyaluronan (Table 1) has been demonstrated against a wide range of viruses, including HSV-1, HSV-2, HIV, NDV, VSV, RV, RABV, Ebola virus, Coxsackievirus B5 (COXB5), mumps virus and IAV (H1N1 and H3N2), while only mild activity has been observed against porcine parvovirus (90, 92, 93).
3.4.3 Mechanism of action
Hyaluronan has shown capacity to exert antiviral activity by different mechanisms, such as acting as decoy receptor, limiting the infection of unstimulated CD4 + T helper cells in a CD44-dependent manner, binding to envelope ligand sites, or disrupting viral docking, internalization and uncoating in host cells; it has also been demonstrated to enhance the host immune response by stimulating the release of antiviral mediators (15, 90).
3.4.4 Critical evaluation
Hyaluronan’s lack of sulphation avoids anticoagulants effects, which is a major safety advantage over heparin. However, its antiviral activity is less consistent across viral families, and, like other hydrocolloids, it stills lacking sufficient clinical studies. In turn, its excellent biocompatibility makes it an attractive candidate for topical or adjunctive applications.
4 Hydrocolloids from plant sources
Sulphated polysaccharides with antiviral properties are uncommon in higher plants; however, promising opportunities are emerging from the chemical modification of natural compounds (semisynthetic hydrocolloids). This generally involves a two-step approach, starting by isolating the targeted compound and further subjecting it to sulphation. Such chemically altered molecules present valuable alternatives for developing antiviral therapeutic agents. Sulphated glucans and xylomannans, for instance, have demonstrated strong antiviral efficacy, showing to be active against human CMV, HSV-1, HSV-2, RSV, IAV, and HIV-1 (94).
4.1 Pectin
4.1.1 Structure
Pectin consists of a complex group of polysaccharides primarily composed of a backbone of α-(1 → 4) linked polygalacturonic acid units, intercalated with rhamnose residues, and presenting side chains containing neutral sugars. The carboxyl groups on the galacturonic acid units can be esterified with non-carbohydrate groups such as methyl or acetyl groups.
4.1.2 Occurrence and antiviral spectrum
Pectin is predominantly located in the primary cell walls of plants, especially in soft non-woody tissues (95). Pectin extracted from mango (Mangifera indica) peels showed no activity against RSV, contrasting to its sulphated derivatives, which effectively inhibited RSV-induced cytopathic effects (in Hep-2 cells, a well-established model due to its permissiveness to viral infection) (96). In contrast, other natural pectins, such as those obtained from Cucumis melo, Dimorphandra gardneriana, Inga spp., Morinda citrifolia, and Salvia plebeia, showed antiviral activity in its native form, as reported against RSV, HSV-1 and poliovirus (97–100).
4.1.3 Mechanism of action
Sulphated pectin demonstrated a strong inhibition of the initial stages of the replication cycle, which might be related to the electrostatic interaction of its polyanionic sulphate groups and the positively charged residues in the viral envelope, leading to the formation of high molecular weight sulphated pectin-virus complexes that block viral adsorption and penetration (6). Sulphated pectin can also inhibit viruses through extracellular binding to cellular receptors (triggering signaling pathways) or through intracellular interactions with effector molecules (101); likewise, it can affect the later stages of viral replication, such as maturation and release of viral particles, suggesting a potential interference with enzymes involved in genome replication (10).
4.1.4 Critical evaluation
Sulphated pectins display strong activity at low micromolar concentrations with limited cytotoxicity, whereas native pectin is largely inactive. However, their translation into clinical uses still requires studies to validate their safety and stability in vivo (2).
4.2 Starch
4.2.1 Structure
Starch consists of D-glucose units linked through α-(1 → 4) glycosidic bonds in linear chains (amylose) and α-(1 → 6) at branched points (amylopectin), usually separated by 24–30 glucose units (102).
4.2.2 Occurrence and antiviral spectrum
Native starch is widely recognized as an energy source, and, in its native form, it generally lacks direct antiviral activity. However, starch derivatives obtained by chemical modifications, such as oxidation, carboxymethylation, phosphorylation, esterification, ozonation, and acetylation, have demonstrated antiviral properties (Table 1) (6, 103). More recently, a novel starch derivate prepared through nucleophilic substitution of hydroxyl groups with quaternary ammonium salts has been gaining attention as a promising therapeutic agent due to its antimicrobial properties (104). Actually, quaternized ammonium starch (QAS) achieved 99% viral inactivation rate after just 1 min of contact with the MHV-3 coronavirus (105).
4.2.3 Mechanism of action
The capacity of starch derivatives to inactivate non-envelope viruses (such as coxsackievirus) was shown to occur by directly interacting and disrupting viral particles (6). In the case of QAS, the associated mechanism of action is attributed to electrostatic interactions between the positively charged quaternary ammonium groups and the negatively charged virus surface. As the quaternary amino groups in QAS have limited diffusion into the phospholipid bilayer of the virus, this leads to the disruption of the viral membrane and subsequent viral inactivation (106).
4.2.4 Critical evaluation
Native starch is nearly inactive, whereas its derivatives exhibit relevant potential for topical or surface applications due to their fast-acting antiviral properties. However, their toxicological profile remains insufficiently characterized, limiting regulatory approval and precluding systemic use until safety is adequately established (103).
4.3 Locust bean gum
4.3.1 Structure
Locust bean gum (LBG) is mainly composed by galactomannans consisting of a linear chain of (1 → 4)-linked β-D-mannopyranosyl units with (1 → 6)-linked α-D-galactopyranosyl side chains. The mannose and galactose contents range from 73–86% and 27–14%, respectively, resulting in a typical mannose:galactose ratio of about 4:1 (107).
4.3.2 Occurrence and antiviral spectrum
The antiviral activity of sulphated derivatives of the galactomannan gum derived from the endosperm of Ceratonia siliqua (L.) Taub was evaluated against HIV using an MT-4 cell model. Sulphated LBG demonstrated an anti-HIV activity (Table 1) comparable to that of dextran and curdlan sulphate, two sulphated polysaccharides reported to have the strongest anti-HIV activity (0.1 to 2.4 μg/mL) combined with low cytotoxicity. Furthermore, this anti-HIV effect of sulphated LBG increased proportionally with its degree of sulphation (108). Sulphated LBG was also reported for its activity against DENV in an LLC-MK2 cell model, again in line with the anti-DENV effects observed for dextran and curdlan sulphate. As in the MT-4 cell assays, sulphated LBG showed no acute cytotoxicity in LLC-MK2 cells. The anti-DENV activity was also proportional to the degree of sulphation.
4.3.3 Mechanism of action
The proposed mechanism of action is likely to occur via electrostatic interaction between the negatively charged sulphate groups of sulphated LBG and the positively charged amino groups of poly-L-lysine in viral proteins; independently of the mechanism, the demonstrated antiviral activity is associated with the molecular weight of sulphated LBG (108).
4.3.4 Critical evaluation
Generally, locust bean gum sulphated derivatives stand out for inhibiting HIV and DENV at low concentrations, comparable to those presented by leading algal sulphated polysaccharides. However, its efficacy is highly dependent on the sulphation degree, which may complicate the reproducibility in therapeutic development.
5 Conclusion
Hydrocolloids, especially their sulphated derivatives, have emerged as promising antiviral agents with diverse mechanisms of action, including blocking viral attachment, interfering with entry and fusion, modulating host immune responses, suppressing of replication, and direct virucidal action. Their antiviral potential is strongly influenced by structural features such as degree and position of sulphation, molecular weight, branching patterns, and glycosidic linkages. In fact, a higher sulphation degree often enhances activity by increasing electrostatic interactions with positively charged viral proteins. Importantly, these structural determinants also provide opportunities for therapeutic optimization. For instance, selective O-sulphation has been shown to preserve antiviral potency while reducing anticoagulant effects, thereby widening the therapeutic window. Likewise, tuning molecular weight can balance viral binding affinity with improved tissue penetration and pharmacokinetics, while branching complexity may be harnessed to design semisynthetic mimetics that maximize electrostatic interactions without retaining unwanted biological activities.
Comparatively, algal polysaccharides such as carrageenan and fucoidan demonstrate broad-spectrum activity, with carrageenan already tested in clinical nasal sprays, though efficacy varies. Animal-derived glycosaminoglycans, particularly heparin, exhibit strong mechanistic evidence, but are constrained by anticoagulant risks. Plant-derived hydrocolloids generally require extensive modification to exhibit antiviral properties, entailing regulatory and toxicological challenges.
A critical evaluation shows that their clinical translation is hindered by structural heterogeneity, inconsistent virus-specific activity, and side effects such as anticoagulation. This requires efforts to design derivatives with strong antiviral effects but minimal side effects, or combination approaches with other polysaccharides or synthetic drugs. Importantly, the structural determinants that control antiviral activity also provide levers for therapeutic optimization. For example, selective O-sulphation can preserve antiviral potency while reducing anticoagulant effects, tuning molecular weight can improve tissue penetration, and branching modifications may enhance viral binding while minimizing toxicity. These strategies, combined with formulation approaches such as nanocarrier encapsulation or localized delivery (e.g., intranasal sprays), offer realistic pathways to overcome current limitations and translate hydrocolloids into clinically viable antivirals.
Likewise, and to move forward, future research must prioritize (i) comparative head-to-head evaluations to identify the most promising candidates; (ii) structural modifications that balance potency with safety; (iii) nanocarrier-based strategies to improve bioavailability; (iv) rigorous in vivo and clinical trials addressing dose, pharmacokinetics, and host-virus interactions.
Personalized medicine approaches, integrating insights from virology, immunology, and pharmacology, will also be critical to tailoring treatments to individual patient profiles.
Only by addressing these challenges hydrocolloids will evolve from promising laboratory findings to practical antiviral therapies.
Author contributions
CP: Conceptualization, Investigation, Methodology, Writing – original draft, Writing – review & editing. MC: Conceptualization, Investigation, Writing – original draft. JB: Conceptualization, Validation, Writing – review & editing. MP: Conceptualization, Supervision, Writing – review & editing. MO: Conceptualization, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work received financial support from the PT national funds (FCT/MECI, Fundação para a Ciência e Tecnologia and Ministério da Educação, Ciência e Inovação) through the project UID/50006 - Laboratório Associado para a Química Verde - Tecnologias e Processos Limpos. Authors also thank the EU-FORA Fellowship Program (EUBA-EFSA-2023-ENREL-01-ALGAESAFE-GA14). JB acknowledges FCT for his contract (FCT-Tenure: 2023.11031.TENURE.023); CP (2021.09490.BD) and MC (2024.05795.BD) thank FCT for their PhD grants.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. To refine Figure 2.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. De Jesús-González, LA, León-Juárez, M, Lira-Hernández, FI, Rivas-Santiago, B, Velázquez-Cervantes, MA, Méndez-Delgado, IM, et al. Advances and challenges in antiviral development for respiratory viruses. Pathogens. (2025) 14:20. doi: 10.3390/pathogens14010020
2. Sharma, A, Shahid, A, Banerjee, R, and Kumar, KJ. Emerging insights into the structure-activity relationship of water-soluble polysaccharides in antiviral therapy. Euro J Med Chem Rep. (2024) 10:100122. doi: 10.1016/J.EJMCR.2023.100122
3. Andrei, G. Vaccines and antivirals: grand challenges and great opportunities. Front Virol. (2021) 1:666548. doi: 10.3389/fviro.2021.666548
4. Panggabean, JA, Adiguna, SP, Rahmawati, SI, Ahmadi, P, Zainuddin, EN, Bayu, A, et al. Antiviral activities of algal-based sulfated polysaccharides. Molecules. (2022) 27:1178. doi: 10.3390/molecules27041178
5. Ray, B, Ali, I, Jana, S, Mukherjee, S, Pal, S, Ray, S, et al. Antiviral strategies using natural source-derived sulfated polysaccharides in the light of the COVID-19 pandemic and major human pathogenic viruses. Viruses. (2022) 14:35. doi: 10.3390/v14010035
6. Claus-Desbonnet, H, Nikly, E, Nalbantova, V, Karcheva-Bahchevanska, D, Ivanova, S, Pierre, G, et al. Polysaccharides and their derivatives as potential antiviral molecules. Viruses. (2022) 14:426. doi: 10.3390/v14020426
7. Kwon, PS, Oh, H, Kwon, SJ, Jin, W, Zhang, F, Fraser, K, et al. Sulfated polysaccharides effectively inhibit SARS-CoV-2 in vitro. Cell Discov. (2020) 6:50. doi: 10.1038/S41421-020-00192-8
8. Kim, M, Kim, SR, Park, J, Mun, SH, Kwak, M, Ko, HJ, et al. Structure and antiviral activity of a pectic polysaccharide from the root of Sanguisorba officinalis against enterovirus 71 in vitro/vivo. Carbohydr Polym. (2022) 281:119057. doi: 10.1016/J.CARBPOL.2021.119057
9. Alfinaikh, RS, Alamry, KA, and Hussein, MA. Sustainable and biocompatible hybrid materials-based sulfated polysaccharides for biomedical applications: a review. RSC Adv. (2025) 15:4708–67. doi: 10.1039/D4RA07277D
10. Lu, W, Yang, Z, Chen, J, Wang, D, and Zhang, Y. Recent advances in antiviral activities and potential mechanisms of sulfated polysaccharides. Carbohydr Polym. (2021) 272:118526. doi: 10.1016/j.carbpol.2021.118526
11. Wang, S, Wang, W, Hou, L, Qin, L, He, M, Li, W, et al. A sulfated glucuronorhamnan from the green seaweed Monostroma nitidum: characteristics of its structure and antiviral activity. Carbohydr Polym. (2020) 227:115280. doi: 10.1016/J.CARBPOL.2019.115280
12. Aw, DZH, Zhang, DX, and Vignuzzi, M. Strategies and efforts in circumventing the emergence of antiviral resistance against conventional antivirals. NPJ Antimicrobials Resistance. (2025) 3:54. doi: 10.1038/s44259-025-00125-z
13. Tahamid Tusar, M T, Hossen, Z, Gazi, H R, Haq, N, Jubayer, AAl, Islam, M M, et al., High-throughput screening of natural antiviral drug candidates against white spot syndrome virus targeting VP28 in Penaeus monodon: computational drug design approaches J Genet Eng Biotechnol (2025) 23:100455 doi: 10.1016/J.JGEB.2024.100455
14. Cerqueira, MA, Leite, ACCO, Tomás, AL, Reichel, A, Silva, PM, Santos, NC, et al. Edible alginate-based films with anti-SARS-CoV-2 activity. Food Microbiol. (2023) 113:104251. doi: 10.1016/J.FM.2023.104251
15. Bai, RG, and Tuvikene, R. Potential antiviral properties of industrially important marine algal polysaccharides and their significance in fighting a future viral pandemic. Viruses. (2021) 13:1817. doi: 10.3390/V13091817
16. Jarrah, RM, Potes, MDA, Vitija, X, Durrani, S, Ghaith, AK, Mualem, W, et al. Alginate hydrogels: a potential tissue engineering intervention for intervertebral disc degeneration. J Clin Neurosci. (2023) 113:32–7. doi: 10.1016/j.jocn.2023.05.001
17. Hashimoto, A, Takayama, R, Lin, D-J, Cano-Vicent, A, Hashimoto, R, Takayama, K, et al. Biocompatible films of calcium alginate inactivate enveloped viruses such as SARS-CoV-2. Polymers. (2022) 14:1483. doi: 10.3390/POLYM14071483
18. Rohilla, D, Srivastava, AK, Singh, RP, Yadav, P, Singh, SK, Kumar, D, et al. Algae polysaccharides (carrageenan and alginate)—a treasure-trove of antiviral compounds: an in silico approach to identify potential candidates for inhibition of S1-RBD spike protein of SARS-CoV2 Stress 3 555–569 (2023) doi: 10.3390/STRESSES3030039 3
19. El-Sheekh, M, Kassem, WMA, Alwaleed, EA, and Saber, H. Optimization and characterization of brown seaweed alginate for antioxidant, anticancer, antimicrobial, and antiviral properties. Int J Biol Macromol. (2024) 278:134715. doi: 10.1016/J.IJBIOMAC.2024.134715
20. Yang, C, Li, D, Wang, S, Xu, M, Wang, D, Li, X, et al. Inhibitory activities of alginate phosphate and sulfate derivatives against SARS-CoV-2 in vitro. Int J Biol Macromol. (2023) 227:316–28. doi: 10.1016/J.IJBIOMAC.2022.11.311
21. Dong, X, Qiu, Y, Jia, N, Wu, Y, Nie, Q, Wen, J, et al. Recent advances of edible marine algae-derived sulfated polysaccharides in antiviral treatments: challenges vs. opportunities. Front Nutr. (2025) 12:1561119. doi: 10.3389/FNUT.2025.1561119/XML
22. Sansone, C, Brunet, C, Noonan, DM, and Albini, A. Marine algal antioxidants as potential vectors for controlling viral diseases. Antioxidants. (2020) 9:392. doi: 10.3390/ANTIOX9050392
23. Jabeen, F, Zil-E-Aimen, N, Ahmad, R, Mir, S, Awwad, NS, and Ibrahium, HA. Carrageenan: structure, properties and applications with special emphasis on food science. RSC Adv. (2025) 15:22035–62. doi: 10.1039/d5ra03296b
24. Krylova, NV, Kravchenko, AO, Iunikhina, OV, Pott, AB, Likhatskaya, GN, Volod’ko, AV, et al. Influence of the structural features of carrageenans from red algae of the far eastern seas on their antiviral properties. Mar Drugs. (2022) 20:60. doi: 10.3390/md20010060
25. Hans, N, Malik, A, and Naik, S. Antiviral activity of sulfated polysaccharides from marine algae and its application in combating COVID-19: mini review. Bioresour Technol Rep. (2020) 13:100623. doi: 10.1016/J.BITEB.2020.100623
26. Salih, AEM, Thissera, B, Yaseen, M, Hassane, ASI, El-seedi, HR, Sayed, AM, et al. Marine sulfated polysaccharides as promising antiviral agents: a comprehensive report and modeling study focusing on SARS cov-2. Mar Drugs. (2021) 19:406. doi: 10.3390/md19080406
27. Wang, S, Luo, Y, Huang, L, Wang, S, Hao, C, Sun, L, et al. The inhibition effects and mechanisms of sulfated chitooligosaccharides on influenza a virus in vitro and in vivo. Carbohydr Polym. (2022) 286:119316. doi: 10.1016/j.carbpol.2022.119316
28. Fröba, M, Große, M, Setz, C, Rauch, P, Auth, J, Spanaus, L, et al. Iota-carrageenan inhibits replication of sars-cov-2 and the respective variants of concern alpha, beta, gamma and delta. Int J Mol Sci. (2021) 22:13202. doi: 10.3390/ijms222413202
29. Morokutti-Kurz, M, Fröba, M, Graf, P, Große, M, Grassauer, A, Auth, J, et al. Iota-carrageenan neutralizes SARS-CoV-2 and inhibits viral replication in vitro. PLoS One. (2021) 16:e0237480. doi: 10.1371/journal.pone.0237480
30. Wei, Q, Fu, G, Wang, K, Yang, Q, Zhao, J, Wang, Y, et al. Advances in research on antiviral activities of Sulfated polysaccharides from seaweeds. Pharmaceuticals. (2022) 15:581. doi: 10.3390/PH15050581
31. Schutz, D, Conzelmann, C, Fois, G, Groß, R, Weil, T, Wettstein, L, et al. Carrageenan-containing over-the-counter nasal and oral sprays inhibit SARS-CoV-2 infection of airway epithelial cultures. Am J Physiol Lung Cell Mol Physiol. (2021) 320:L750–6. doi: 10.1152/AJPLUNG.00552.2020
32. Jang, Y, Shin, H, Lee, MK, Kwon, OS, Shin, JS, Kim, Y i, et al. Antiviral activity of lambda-carrageenan against influenza viruses and severe acute respiratory syndrome coronavirus 2. Sci Rep. (2021) 11:821. doi: 10.1038/S41598-020-80896-9
33. Morokutti-Kurz, M, Unger-Manhart, N, Graf, P, Rauch, P, Kodnar, J, Große, M, et al. The saliva of probands sucking an iota-carrageenan containing lozenge inhibits viral binding and replication of the Most predominant common cold viruses and SARS-CoV-2. Int J Gen Med. (2021) 14:5241. doi: 10.2147/IJGM.S325861
34. Chadwick, M, Carvalho, LG, Vanegas, C, and Dimartino, S. A comparative review of alternative fucoidan extraction techniques from seaweed. Mar Drugs. (2025) 23:23. doi: 10.3390/md23010027
35. Zhao, Y, Ning, L, Zhu, P, Jiang, J, Yao, Z, and Zhu, B. The origin, properties, structure, catalytic mechanism, and applications of fucoidan-degrading enzymes. Mar Drugs. (2025) 23:23. doi: 10.3390/md23030097
36. Li, Z, Li, L, Cai, S, Xu, X, Zhang, X, Du, K, et al. The STING-mediated antiviral effect of fucoidan from Durvillaea Antarctica. Carbohydr Polym. (2024) 331:121899. doi: 10.1016/J.CARBPOL.2024.121899
37. Richards, C, Williams, NA, Fitton, JH, Stringer, DN, Karpiniec, SS, and Park, AY. Oral fucoidan attenuates lung pathology and clinical signs in a severe influenza a mouse model. Mar Drugs. (2020) 18:246. doi: 10.3390/MD18050246
38. Hans, N, Gupta, S, Patel, AK, Naik, S, and Malik, A. Deciphering the role of fucoidan from brown macroalgae in inhibiting SARS-CoV-2 by targeting its main protease and receptor binding domain: invitro and insilico approach. Int J Biol Macromol. (2023) 248:125950. doi: 10.1016/J.IJBIOMAC.2023.125950
39. Salehi, B, Sharifi-Rad, J, Seca, AML, Pinto, DCGA, Michalak, I, Trincone, A, et al. Current trends on seaweeds: looking at chemical composition, phytopharmacology, and cosmetic applications. Molecules. (2019) 24:4182. doi: 10.3390/MOLECULES24224182
40. Amri, S, Bouslama, L, Mzoughi, Z, Nouira, F, Majdoub, H, and Bouraoui, A. Chemical characterization and evaluation of antiviral activity of two fucoidans extracted from Mediterranean brown seaweeds, Padina pavonica and Dictyopteris membranacea. Lett Appl Microbiol. (2025) 78:ovaf002. doi: 10.1093/LAMBIO/OVAF002
41. Akl, FMA, El-Sheekh, MM, Makhlof, MEM, and Ahmed, SI. Antimicrobial, antidiabetic, antiviral, and antioxidant activities of fucoidan extracted from the brown seaweed Padina pavonica. BMC Biotechnol. (2025) 25:70. doi: 10.1186/S12896-025-01004-1
42. Krylova, NV, Ermakova, SP, Lavrov, VF, Leneva, IA, Kompanets, GG, Iunikhina, OV, et al. The comparative analysis of antiviral activity of native and modified fucoidans from brown algae Fucus evanescens in vitro and in vivo. Mar Drugs. (2020) 18:224. doi: 10.3390/MD18040224
43. Liyanage, NM, Nagahawatta, DP, Jayawardena, TU, Sanjeewa, KKA, Jayawrdhana, HHACK, Kim, JI, et al. Sulfated polysaccharides from seaweeds: a promising strategy for combatting viral diseases—a review. Mar Drugs. (2023) 21:461. doi: 10.3390/md21090461
44. Lakshmana Senthil, S. A comprehensive review to assess the potential, health benefits and complications of fucoidan for developing as functional ingredient and nutraceutical. Int J Biol Macromol. (2024) 277:134226. doi: 10.1016/J.IJBIOMAC.2024.134226
45. Christensen, MD, Allahgholi, L, Dobruchowska, JM, Moenaert, A, Guðmundsson, H, Friðjónsson, Ó, et al. Laminarins and their derivatives affect dendritic cell activation and their crosstalk with T cells. Int J Biol Macromol. (2025) 306:141287. doi: 10.1016/J.IJBIOMAC.2025.141287
46. Li, Y, Zheng, Y, Zhang, Y, Yang, Y, Wang, P, Imre, B, et al. Brown Algae Carbohydrates: Structures, Pharmaceutical Properties, and Research Challenges. Mar Drugs. (2021) 19:620. doi: 10.3390/md19110620
47. Pramanik, S, Singh, A, Abualsoud, BM, Deepak, A, Nainwal, P, Sargsyan, AS, et al. From algae to advancements: laminarin in biomedicine. RSC Adv. (2024) 14:3209–31. doi: 10.1039/D3RA08161C
48. Xu, L, Lyu, J, Qiu, Z, Liu, Q, Hu, H, Zhao, L, et al. Laminaran potentiates cGAS-STING signaling to enhance antiviral responses. Int Immunopharmacol. (2025) 147:114014. doi: 10.1016/J.INTIMP.2025.114014
49. Long, H, Xiao, J, Wang, X, Liang, M, Fan, Y, Xu, Y, et al. Laminarin acetyl esters: synthesis, conformational analysis and anti-viral effects. Int J Biol Macromol. (2022) 216:528–36. doi: 10.1016/J.IJBIOMAC.2022.06.208
50. Deng, C, Chen, D, Yang, L, Zhang, Y, Jin, C, Li, Y, et al. The role of cGAS-STING pathway ubiquitination in innate immunity and multiple diseases. Front Immunol. (2025) 16:1522200. doi: 10.3389/fimmu.2025.1522200
51. Li, C, Tang, T, Du, Y, Jiang, L, Yao, Z, Ning, L, et al. Ulvan and Ulva oligosaccharides: a systematic review of structure, preparation, biological activities and applications. Bioresour Bioprocess. (2023) 10:66. doi: 10.1186/s40643-023-00690-z
52. Glasson, CRK, Luiten, CA, Carnachan, SM, Daines, AM, Kidgell, JT, Hinkley, SFR, et al. Structural characterization of ulvans extracted from blade (Ulva ohnoi) and filamentous (Ulva tepida and Ulva prolifera) species of cultivated Ulva. Int J Biol Macromol. (2022) 194:571–9. doi: 10.1016/J.IJBIOMAC.2021.11.100
53. Chi, Y, Zhang, M, Wang, X, Fu, X, Guan, H, and Wang, P. Ulvan lyase assisted structural characterization of ulvan from Ulva pertusa and its antiviral activity against vesicular stomatitis virus. Int J Biol Macromol. (2020) 157:75–82. doi: 10.1016/J.IJBIOMAC.2020.04.187
54. Shefer, S, Robin, A, Chemodanov, A, Lebendiker, M, Bostwick, R, Rasmussen, L, et al. Fighting SARS-CoV-2 with green seaweed Ulva sp. extract: extraction protocol predetermines crude ulvan extract anti-SARS-CoV-2 inhibition properties in in vitro Vero-E6 cells assay. PeerJ. (2021) 9:e12398. doi: 10.7717/PEERJ.12398/SUPP-5
55. Binsuwaidan, R, El-Masry, TA, El-Sheekh, M, Seadawy, MG, Makhlof, MEM, Aboukhatwa, SM, et al. Prospective antiviral effect of Ulva lactuca aqueous extract against COVID-19 infection. Mar Drugs. (2024) 22:22. doi: 10.3390/md22010030
56. Syndel Caroline Ribeiro, M, Araujo da Silva, F, Pereira, B, dos Santos, L, Danyelle Santos Araujo, T, Monteiro, R, et al. Characterization and evaluation of the biological activities of Ulvan extracted from Ulva lactuca with a focus on the antiviral activity against herpes simplex virus type 1. J Adv Biol Biotechnol. (2025) 28:587–601. doi: 10.9734/JABB/2025/V28I22018
57. Thambiliyagodage, C, Jayanetti, M, Mendis, A, Ekanayake, G, Liyanaarachchi, H, and Vigneswaran, S. Recent advances in chitosan-based applications—a review. Materials. (2023) 16:2073. doi: 10.3390/ma16052073
58. Tan, RSL, Hassandarvish, P, Chee, CF, Chan, LW, and Wong, TW. Chitosan and its derivatives as polymeric anti-viral therapeutics and potential anti-SARS-CoV-2 nanomedicine. Carbohydr Polym. (2022) 290:119500. doi: 10.1016/J.CARBPOL.2022.119500
59. Harugade, A, Sherje, AP, and Pethe, A. Chitosan: a review on properties, biological activities and recent progress in biomedical applications. React Funct Polym. (2023) 191:105634. doi: 10.1016/j.reactfunctpolym.2023.105634
60. Safarzadeh, M, Sadeghi, S, Azizi, M, Rastegari-Pouyani, M, Pouriran, R, and Haji Molla Hoseini, M. Chitin and chitosan as tools to combat COVID-19: a triple approach. Int J Biol Macromol. (2021) 183:235–44. doi: 10.1016/J.IJBIOMAC.2021.04.157
61. Gopal, J, Muthu, M, Pushparaj, SSC, and Sivanesan, I. Anti-COVID-19 credentials of chitosan composites and derivatives: future scope? Antibiotics. (2023) 12:665. doi: 10.3390/ANTIBIOTICS12040665
62. Teotia, A, Laurén, I, Borandeh, S, and Seppälä, J. Quaternized chitosan derivatives as viable antiviral agents: structure-activity correlations and mechanisms of action. ACS Appl Mater Interfaces. (2023) 15:18707–19. doi: 10.1021/ACSAMI.3C01421/ASSET/IMAGES/LARGE/AM3C01421_0009.JPEG
63. Cao, Z, Ma, X, Lv, D, Wang, J, Shen, Y, Peng, S, et al. Synthesis of chitin nanocrystals supported Zn2+ with high activity against tobacco mosaic virus. Int J Biol Macromol. (2023) 250:126168. doi: 10.1016/J.IJBIOMAC.2023.126168
64. Mawazi, SM, Kumar, M, Ahmad, N, Ge, Y, and Mahmood, S. Recent applications of chitosan and its derivatives in antibacterial, anticancer, wound healing, and tissue engineering fields. Polymers. (2024) 16:1351. doi: 10.3390/POLYM16101351
65. Merry, CLR, Lindahl, U, Couchman, J, and Esko, JD. Proteoglycans and Sulfated glycosaminoglycans. Essentials of glycobiology. Cold Spring Harbor (NY): Cold Spring Harbor Laboratory Press. (2022)
66. Wang, K, Qi, L, Zhao, L, Liu, J, Guo, Y, and Zhang, C. Degradation of chondroitin sulfate: mechanism of degradation, influence factors, structure-bioactivity relationship and application. Carbohydr Polym. (2023) 301:120361. doi: 10.1016/j.carbpol.2022.120361
67. Lee, MF, Wu, YS, and Poh, CL. Molecular mechanisms of antiviral agents against dengue virus. Viruses. (2023) 15:705. doi: 10.3390/V15030705
68. Möller, S, Theiß, J, Deinert, TIL, Golat, K, Heinze, J, Niemeyer, D, et al. High-sulfated glycosaminoglycans prevent coronavirus replication. Viruses. (2022) 14:413. doi: 10.3390/V14020413/S1
69. Zoepfl, M, Dwivedi, R, Kim, SB, McVoy, MA, and Pomin, VH. Antiviral activity of marine sulfated glycans against pathogenic human coronaviruses. Sci Rep. (2023) 13:1–12. doi: 10.1038/s41598-023-31722-5
70. Li, J, and Li, SJ. Chondroitin sulfate binds to main protease of SARS-CoV-2 and efficaciously inhibits its activity. Int J Biol Macromol. (2025) 306:141547. doi: 10.1016/J.IJBIOMAC.2025.141547
71. Chen, C, Zhang, X, Zhang, W, Ding, D, Loka, RS, Zhao, K, et al. Dermatan sulfate: structure, biosynthesis, and biological roles. Biomolecules. (2025) 15:1158. doi: 10.3390/BIOM15081158
72. Zhang, Q, Pavlinov, I, Ye, Y, and Zheng, W. Therapeutic development targeting host heparan sulfate proteoglycan in SARS-CoV-2 infection. Front Med (Lausanne). (2024) 11:1364657. doi: 10.3389/FMED.2024.1364657/XML
73. Morris, JS, and Dawson, PA. Pathogens that infect mammalian cells via sulfonated glycosaminoglycans. Front Cell Infect Microbiol. (2025) 15:1613923. doi: 10.3389/FCIMB.2025.1613923/XML
74. Carvajal-Barriga, EJ, and Fields, RD. Sulfated polysaccharides as multi target molecules to fight COVID 19 and comorbidities. Heliyon. (2023) 9:e13797. doi: 10.1016/J.HELIYON.2023.E13797
75. Kjellén, L, and Lindahl, U. Heparin and heparan sulfate—the essence of sequence. Proteoglycan Res. (2024) 2:2. doi: 10.1002/pgr2.70006
76. Wang, Y, Zhang, Y, Wang, P, Jing, T, Hu, Y, and Chen, X. Research Progress on antiviral activity of heparin. Curr Med Chem. (2024) 31:7–24. doi: 10.2174/0929867330666230203124032
77. Kim, SY, Koetzner, CA, Payne, AF, Nierode, GJ, Yu, Y, Wang, R, et al. Glycosaminoglycan compositional analysis of relevant tissues in zika virus pathogenesis and in vitro evaluation of heparin as an antiviral against zika virus infection. Biochemistry. (2019) 58:1155–66. doi: 10.1021/ACS.BIOCHEM.8B01267/SUPPL_FILE/BI8B01267_SI_001.PDF
78. Tree, JA, Turnbull, JE, Buttigieg, KR, Elmore, MJ, Coombes, N, Hogwood, J, et al. Unfractionated heparin inhibits live wild type SARS-CoV-2 cell infectivity at therapeutically relevant concentrations. Br J Pharmacol. (2021) 178:626–35. doi: 10.1111/BPH.15304
79. Tandon, R, Sharp, JS, Zhang, F, Pomin, VH, Ashpole, NM, Mitra, D, et al. Effective inhibition of SARS-CoV-2 entry by heparin and enoxaparin derivatives. J Virol. (2021) 95:e01987-20. doi: 10.1128/JVI.01987-20
80. Paiardi, G, Richter, S, Oreste, P, Urbinati, C, Rusnati, M, and Wade, RC. The binding of heparin to spike glycoprotein inhibits SARS-CoV-2 infection by three mechanisms. J Biol Chem. (2022) 298:101507. doi: 10.1016/J.JBC.2021.101507
81. Li, J, Zhang, Y, Pang, H, and Li, SJ. Heparin interacts with the main protease of SARS-CoV-2 and inhibits its activity. Spectrochim Acta A Mol Biomol Spectrosc. (2021) 267:120595. doi: 10.1016/J.SAA.2021.120595
82. Lee, LYY, Suryadinata, R, McCafferty, C, Ignjatovic, V, Purcell, DFJ, Robinson, P, et al. Heparin inhibits SARS-CoV-2 replication in human nasal epithelial cells. Viruses. (2022) 14:2620. doi: 10.3390/V14122620/S1
83. Gupta, Y, Maciorowski, D, Zak, SE, Kulkarni, CV, Herbert, AS, Durvasula, R, et al. Heparin: a simplistic repurposing to prevent SARS-CoV-2 transmission in light of its in-vitro nanomolar efficacy. Int J Biol Macromol. (2021) 183:203–12. doi: 10.1016/J.IJBIOMAC.2021.04.148
84. Casale, J, and Crane, JS. Biochemistry, glycosaminoglycans. StatPearls (2023) Available online at: https://www.ncbi.nlm.nih.gov/books/NBK544295/ [Accessed August 27, 2025]
85. Revuelta, J, Rusu, L, Frances-Gomez, C, Trapero, E, Iglesias, S, Pinilla, EC, et al. Synthetic heparan sulfate mimics based on chitosan derivatives show broad-spectrum antiviral activity. Commun Biol. (2025) 8:360. doi: 10.1038/S42003-025-07763-Z
86. Fuochi, V, Furnari, S, Floresta, G, Patamia, V, Zagni, C, Drago, F, et al. Antiviral efficacy of heparan sulfate and enoxaparin sodium against SARS-CoV-2. Arch Pharm (Weinheim). (2025) 358:e2400545. doi: 10.1002/ARDP.202400545
87. Watanabe, T, Takeda, K, Hiemori, K, Minamisawa, T, and Tateno, H. A glycosaminoglycan microarray identifies the binding of SARS-CoV-2 spike protein to chondroitin sulfate E. FEBS Lett. (2021) 595:2341–9. doi: 10.1002/1873-3468.14173
88. Fu, Z, Zhang, X, Liu, J, Li, J, Zeng, Y, Yang, J, et al. Enzymatic synthesis and immunomodulatory activity of highly branched α-D-glucans with glycogen-like structure. Int J Biol Macromol. (2023) 237:123882. doi: 10.1016/J.IJBIOMAC.2023.123882
89. Reddy Shetty, P, Batchu, UR, Buddana, SK, Sambasiva Rao, KRS, and Penna, S. A comprehensive review on α-D-glucans: structural and functional diversity, derivatization and bioapplications. Carbohydr Res. (2021) 503:108297. doi: 10.1016/J.CARRES.2021.108297
90. Zamboni, F, Wong, CK, and Collins, MN. Hyaluronic acid association with bacterial, fungal and viral infections: can hyaluronic acid be used as an antimicrobial polymer for biomedical and pharmaceutical applications? Bioact Mater. (2022) 19:458. doi: 10.1016/J.BIOACTMAT.2022.04.023
91. Snetkov, P, Zakharova, K, Morozkina, S, Olekhnovich, R, and Uspenskaya, M. Hyaluronic acid: the influence of molecular weight on structural, physical, physico-chemical, and degradable properties of biopolymer. Polymers (Basel). (2020) 12:1800. doi: 10.3390/POLYM12081800
92. Tang, F, Reeves, SR, Brune, JE, Chang, MY, Chan, CK, Waldron, P, et al. Inter-alpha-trypsin inhibitor (IαI) and hyaluronan modifications enhance the innate immune response to influenza virus in the lung. Matrix Biol. (2024) 126:25–42. doi: 10.1016/J.MATBIO.2024.01.004
93. Shebl, RI, Amer, ME, Abuamara, TMM, Matar, ER, Ahmed, HF, Gomah, TA, et al. Staphylococcus aureus derived hyaluronic acid and bacillus Calmette-Guérin purified proteins as immune enhancers to rabies vaccine and related immuno-histopathological alterations. Clin Exp Vaccine Res. (2021) 10:229–39. doi: 10.7774/CEVR.2021.10.3.229
94. Luan, F, Ji, Y, Peng, L, Liu, Q, Cao, H, Yang, Y, et al. Extraction, purification, structural characteristics and biological properties of the polysaccharides from Codonopsis pilosula: a review. Carbohydr Polym. (2021) 261:117863. doi: 10.1016/J.CARBPOL.2021.117863
95. Xue, Y, Li, H, and Kang, X. Arabinan-rich architectures in pectin rhamnogalacturonan I domain unveiled by termite digestion: in situ structural insights from solid-state NMR. Carbohydr Polym. (2025) 361:123662. doi: 10.1016/J.CARBPOL.2025.123662
96. Barboza, MGL, Dyna, AL, Lima, TF, Tavares, ER, Yamada-Ogatta, SF, Deduch, F, et al. In vitro antiviral effect of sulfated pectin from Mangifera indica against the infection of the viral agent of childhood bronchiolitis (respiratory syncytial virus - RSV). Int J Biol Macromol. (2024) 280:135387. doi: 10.1016/J.IJBIOMAC.2024.135387
97. Agostinho, KF, Rechenchoski, DZ, Faccin-Galhardi, LC, De Sousa, ALN, Cunha, AP, Ricardo, NMPS, et al. Cucumis melo pectin as potential candidate to control herpes simplex virus infection. FEMS Microbiol Lett. (2021) 368:fnab013. doi: 10.1093/femsle/fnab013
98. de Godoi, AM, Faccin-Galhardi, LC, Rechenchoski, DZ, Arruda, TBMG, Cunha, AP, de Almeida, RR, et al. Structural characterization and antiviral activity of pectin isolated from Inga spp. Int J Biol Macromol. (2019) 139:925–31. doi: 10.1016/j.ijbiomac.2019.07.212
99. Santos, DG, Cunha, AP, Ribeiro, ACB, Brito, DHA, Alenca, LMR, Farias, DF, et al. Characterization and biological activity of native and sulfated noni (Morinda citrifolia Linn.) pectin. J Braz Chem Soc. (2023) 35. doi: 10.21577/0103-5053.20230156
100. Liang Y yu,, Li K wei,, Niu F jv,, Li Y,, Wei H cheng,, Dai Y lei,, et al. Salvia plebeia R. Br. Polysaccharides (SPP) against RSV (respiratory syncytial virus) infection: antiviral effect and mechanisms of action. Biomed Pharma. (2021) 141:111843. doi: 10.1016/j.biopha.2021.111843
101. Jabeen, M, Dutot, M, Fagon, R, Verrier, B, and Monge, C. Seaweed sulfated polysaccharides against respiratory viral infections. Pharmaceutics. (2021) 13:13. doi: 10.3390/pharmaceutics13050733
102. Zhao, D, Li, Z, Xia, J, Kang, Y, Sun, P, Xiao, Z, et al. Research progress of starch as microencapsulated wall material. Carbohydr Polym. (2023) 318:121118. doi: 10.1016/j.carbpol.2023.121118
103. Ribeiro de Carvalho, G, Kudaka, AM, Fares Sampar, J, Alvares, LE, Delarmelina, C, Duarte, MCT, et al. Quaternization of cassava starch and determination of antimicrobial activity against bacteria and coronavirus. Carbohydr Res. (2024) 538:109098. doi: 10.1016/J.CARRES.2024.109098
104. Hou, X, Wang, H, Shi, Y, and Yue, Z. Recent advances of antibacterial starch-based materials. Carbohydr Polym. (2023) 302:120392. doi: 10.1016/j.carbpol.2022.120392
105. de Moura Junior, CF, Ochi, D, Freitas, ED, Kerwald, J, d’Ávila, MA, and Beppu, MM. Synthesis and characterization of n-phosphonium chitosan and its virucidal activity evaluation against coronavirus. Int J Biol Macromol. (2023) 246:125665. doi: 10.1016/j.ijbiomac.2023.125665
106. Guerrero-Bermea, C, Rodríguez Fuentes, N, Cervantes-Uc, JM, Alcántara Quintana, LE, Díaz-Barriga, F, Pérez-Vázquez, F, et al. Antiviral capacity of polypropylene/(1-hexadecyl) trimethyl-ammonium bromide composites against COVID-19. Polym Eng Sci. (2022) 62:4129–35. doi: 10.1002/PEN.26172
107. Flores García, Y, del Martín Campo Solís, MF, Gómez-Angulo, JH, Martínez Preciado, AH, Silva-Jara, JM, de la Padilla Rosa, JD, et al. Industrial applications, principal sources, and extraction of galactomannans: a review. Foods. (2025) 14:1587. doi: 10.3390/FOODS14091587
108. Geronço, MS, da Silveira Ramos, IFda, Silva Filho, EC, dos Santos Rizzo, M, Ribeiro, AB, and da Costa, MP Are structurally modified galactomannan derivatives biologically active? Polysaccharides (2021) 2 1–15 doi: 10.3390/POLYSACCHARIDES2010001
109. Yao, Y, Tang, H, Ma, H, Liu, Z, Huang, J, Yang, X, et al. Chondroitin sulfate/dermatan sulfate hybrid chains from swim bladder: isolation, structural analysis, and anticoagulant activity. Mar Drugs. (2024) 22:9. doi: 10.3390/MD22010009/S1
110. Kwiatkowska, A, and Granicka, LH. Anti-viral surfaces in the fight against the spread of coronaviruses. Membranes (Basel). (2023) 13:464. doi: 10.3390/MEMBRANES13050464
111. Gheda, S, Karkour, AM, Shafay, SEl, GabAllah, M, Cotas, J, and Pereira, L. Polysaccharides from brown seaweeds (Padina boergesenii and Sargassum euryphyllum) as promising inhibitors of SARS-CoV-2: characterization, mechanisms, and therapeutic potential. Macromol (2025), Vol 5:18 Doi:doi: 10.3390/MACROMOL5020018
112. Dong, M, Jiang, Y, Wang, C, Yang, Q, Jiang, X, and Zhu, C. Determination of the extraction, physicochemical characterization, and digestibility of Sulfated polysaccharides in seaweed-Porphyra haitanensis. Mar Drugs. (2020) 18:539. doi: 10.3390/md18110539
Glossary
GalNAc - N-acetyl-D-galactosamine
ACE2 - angiotensin-converting enzyme 2
AIV - avian influenza virus
ASV - avian sarcoma virus
BCoV - bovine coronavirus
BoHV-1 - bovine herpesvirus type 1
ChNC - chitin nanocrystals
CVB-3 - coxsackievirus B3
COXB5 - Coxsackievirus B5
cGAS-STING - cyclic GMP-AMP synthase stimulator of interferon genes
CMV - cytomegalovirus
DENV - dengue virus
ECMV - encephalomyocarditis virus
F-MuLV - friend murine leukaemia helper virus
GlcA - D-glucuronic acid
HAV - hepatitis A virus
HBV - hepatitis B virus
HCV - hepatitis C virus
HSV-1 - herpes simplex virus type-1
HSV-2 - herpes simplex virus type 2
HIV - human immunodeficiency virus
hPIV-2 - human parainfluenza virus type 2
HPV - human papillomavirus
HRV - human rhinovirus
HTCC - N-(2-hydroxypropyl)-3-trimethylammonium chitosan chloride
MNV - murine norovirus
IdoA - L-iduronic acid
IAV - influenza A virus
JEV - Japanese encephalitis virus
LBG - locust bean gum
MeV - measles virus
MEV - Mengo encephalomyelitis virus
MERS-CoV - Middle East respiratory syndrome coronavirus
NDV - Newcastle disease virus
QAS - quaternized ammonium starch
RABV - rabies virus
RBD - receptor-binding domain
RSV - respiratory syncytial virus
RVFV - Rift Valley fever virus
RV - rubella virus
SARS-CoV-2 - severe acute respiratory syndrome coronavirus 2
SFV - simian foamy virus
SINV; - Sindbis virus
TMV - tobacco mosaic virus
VSV - vesicular stomatitis virus
Keywords: hydrocolloid sources and derivatization, antiviral mechanisms of action, susceptible viruses, structure-activity relationship, complementary therapeutic strategies
Citation: Pereira CSGP, Carpena M, Barreira JCM, Prieto MA and Oliveira MBPP (2025) Antiviral activity of natural and modified hydrocolloids: main sources, susceptible viruses, structure-activity relationship and mechanisms of action. Front. Nutr. 12:1696022. doi: 10.3389/fnut.2025.1696022
Edited by:
Marcus O. Muench, Vitalant Research Institute, United StatesReviewed by:
Saikat De, The Scripps Research Institute, United StatesAhmad Mostafa, Al-Azhar University, Egypt
Dinanath Gaikwad, Bharati Vidyapeeth College of Pharmacy, Kolhapur, India
Arun Teotia, University of Helsinki, Finland
Copyright © 2025 Pereira, Carpena, Barreira, Prieto and Oliveira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: João C. M. Barreira, amJhcnJlaXJhQGlwYi5wdA==