 Ma Xiaoying
Ma Xiaoying Zhang Peng
Zhang Peng Zhao Ying
Zhao Ying Chen Xun
Chen Xun- The Institute of Edible Fungi, Liaoning Academy of Agricultural Sciences, Shenyang, China
Background: Chronic inflammation underlies numerous complex diseases, yet current therapeutic strategies show limited efficacy and safety profiles. Despite extensive preclinical evidence, the mechanistic understanding and clinical translation of medicinal mushroom bioactives remain inadequately characterized.
Objective: This review systematically evaluates the immunoregulatory mechanisms of mushroom-derived bioactive compounds and establishes a comprehensive framework for their therapeutic application in chronic inflammatory diseases.
Methods: We analyzed mechanistic evidence for four major compound classes: polysaccharides (β-glucans), triterpenoids, phenolic compounds, and bioactive peptides, examining their effects on immune cell populations and signaling pathways.
Results: These bioactives demonstrate multi-target anti-inflammatory activity by modulating key cellular mediators (macrophages, regulatory T cells, natural killer cells) and critical signaling cascades (NF-κB, MAPK, NLRP3 inflammasome, Nrf2/HO-1). Novel therapeutic targets including gasdermin-mediated pyroptosis provide additional intervention opportunities. However, clinical translation faces significant challenges: poor bioavailability, lack of standardization, and undefined dose–response relationships.
Conclusion: Advanced delivery systems (nanoformulations, structural optimization) and precision nutrition approaches through personalized immune profiling offer promising solutions to overcome translational barriers. This analysis provides evidence-based rationale for advancing medicinal mushrooms from traditional functional foods to standardized immunotherapeutic agents for chronic inflammation management.
1 Introduction
Chronic inflammation constitutes a fundamental pathophysiological mechanism underlying diverse diseases, including cardiovascular disorders, metabolic syndrome, neurodegenerative conditions, and autoimmune diseases (1, 2). Current therapeutic approaches predominantly rely on synthetic anti-inflammatory agents, which often present limited efficacy profiles and significant adverse effects, particularly with long-term use (3). This therapeutic gap has intensified interest in natural immunomodulatory agents that can provide sustainable inflammation management with improved safety profiles.
Medicinal mushrooms have emerged as promising candidates for chronic inflammation management, with over 2,000 species demonstrating documented bioactive properties (4, 5). Recent comprehensive reviews have established the fundamental immunomodulatory potential of mushroom-derived compounds (6, 7), yet critical gaps remain in understanding their precise mechanistic actions and translational applicability. Unlike previous reviews that primarily focused on individual compound classes or single species, the mechanistic understanding of multi-compound synergy and species-specific efficacy variations remains inadequately characterized.
The principal bioactive constituents responsible for anti-inflammatory activities encompass structurally diverse chemical classes: polysaccharides (particularly β-glucans), triterpenoids, phenolic compounds, and bioactive peptides (8). However, significant knowledge gaps persist regarding their comparative therapeutic potency, optimal concentration ranges, and species-specific bioactivity profiles. For instance, β-glucan preparations from Ganoderma lucidum demonstrate IC₅₀ values of 15–50 μg/mL for inflammatory cytokine inhibition, while Cordyceps militaris polysaccharides show efficacy at 25–75 μg/mL, indicating substantial interspecies variability that requires systematic evaluation (9).
Current mechanistic understanding reveals that mushroom bioactives modulate key inflammatory signaling pathways, including nuclear factor-κB (NF-κB), mitogen-activated protein kinase (MAPK), NOD-like receptor protein 3 (NLRP3) inflammasome, and nuclear factor erythroid 2-related factor 2/heme oxygenase-1 (Nrf2/HO-1) pathways (10). However, contradictory findings exist regarding pathway selectivity and compound-specific targeting preferences. For example, ganoderic acids from G. lucidum demonstrate preferential NF-κB inhibition with minimal MAPK interference, while cordycepin from C. militaris shows broad-spectrum pathway modulation, suggesting distinct mechanistic profiles that warrant comparative analysis (11).
Critical translational challenges significantly limit clinical implementation despite promising preclinical evidence. Poor aqueous solubility, low gastrointestinal absorption (bioavailability often <5%), and inconsistent standardization across mushroom-derived products present substantial barriers to therapeutic application (9, 12). Furthermore, significant pharmacokinetic variations exist between species and extraction methods, with Shiitake (Lentinula edodes) lentinan showing 12-h half-life compared to 4–6 h for G. lucidum polysaccharides, indicating the need for species-specific pharmacokinetic optimization (10, 11).
The distinction between preclinical efficacy and clinical translatability remains poorly defined. While numerous in vitro and animal studies demonstrate anti-inflammatory effects, human clinical trials are limited and show inconsistent outcomes (12, 13). This disparity highlights the urgent need for standardized evaluation frameworks that can bridge preclinical promise with clinical validation.
Recent advances in nanotechnology-based delivery systems and precision medicine approaches offer promising solutions to overcome these translational barriers. Nanoformulation strategies have demonstrated up to 10-fold bioavailability enhancement, while personalized immune profiling approaches enable patient-specific therapeutic optimization (1, 14). However, the integration of these innovative approaches with mushroom bioactives requires systematic investigation and validation.
However, despite extensive documentation of the pharmacological potential of mushroom-derived bioactives, a systematic integration of their molecular mechanisms, synergistic interactions, and translational implications remains limited (4). This review advances beyond the existing literature through four key contributions: First, we construct an integrated mechanistic framework that systematically connects immune regulation at the cellular level—such as macrophage polarization and regulatory T cell induction—with molecular signaling networks, including NF-κB, MAPK, the NLRP3 inflammasome, and the Nrf2/HO-1 axis, thereby providing a multi-scale understanding of anti-inflammatory mechanisms. Second, we critically evaluate emerging molecular targets, such as gasdermin-mediated pyroptosis, endoplasmic reticulum stress–inflammation crosstalk, and gut microbiota–immune interactions, which have received limited attention in previous mushroom-related reviews. Third, unlike earlier reviews focusing on individual mushroom species, we conduct a comparative analysis of bioactive compounds, systematically comparing multiple classes—polysaccharides, triterpenoids, phenolics, and peptides—and examining their mechanistic differences, pharmacokinetic limitations, and readiness for clinical translation. Finally, we propose evidence-based translational strategies that integrate nanodelivery systems, structural optimization, and precision immune profiling to overcome current challenges in bioavailability, standardization, and personalized dosing. This framework repositions medicinal mushrooms from empirical traditional remedies into rationally designed, mechanism-based immunotherapeutic agents with well-defined molecular targets, suitable for clinical development (Figure 1).

Figure 1. Comprehensive framework of immunoregulation by bioactive compounds from medicinal mushrooms in chronic inflammation.
2 Bioactive compounds: a multi-faceted immunoregulatory arsenal
Medicinal mushrooms synthesize structurally diverse secondary metabolites that function through complementary mechanisms to modulate inflammatory networks (15). Four principal compound classes—polysaccharides (β-glucans), triterpenoids, phenolic compounds, and bioactive peptides—exhibit distinct pharmacological profiles and species-specific concentrations that collectively determine therapeutic efficacy (16).
2.1 Polysaccharides and β-glucans: pattern recognition receptor modulators
Polysaccharides, particularly β-(1 → 3, 1 → 6)-D-glucans, represent the most extensively characterized immunomodulatory compounds in medicinal mushrooms, functioning primarily through pattern recognition receptors (PRRs) including Dectin-1 and complement receptor 3 (CR3) (17). Recent systematic reviews demonstrate that fungal β-glucans are well-tolerated and can improve immune function, reduce respiratory infections, and ameliorate allergic symptoms (18).
Structural characterization reveals significant molecular weight variations affecting bioactivity. Polysaccharides from Lactarius hatsudake demonstrate molecular weights ranging from 4.9 kDa (LHP-5) to 898 kDa (LHP-1), with lower molecular weight fractions (LHP-4 and LHP-5) showing superior bioactive properties (19). This molecular weight dependence correlates with bioavailability limitations, as high-molecular-weight polysaccharides exhibit poor gastrointestinal absorption due to their hydrophilic properties and large molecular size (20).
Critical pharmacokinetic challenges include limited oral bioavailability, with encapsulation strategies showing promise for enhancement. β-glucan matrices from mushrooms effectively regulate compound release in simulated digestive conditions, with encapsulated formulations following Higuchi release kinetics (21). However, biological activity remains critically dependent on extraction methodologies that preserve structural integrity, particularly the triple-helix configurations essential for immunomodulatory potency (22).
2.2 Triterpenoids: multi-pathway anti-inflammatory modulators
Triterpenoids demonstrate superior pharmacological profiles compared to polysaccharides, with enhanced lipophilicity facilitating improved bioavailability and cellular penetration (23). Recent isolation studies reveal potent anti-inflammatory activities with quantifiable dose–response relationships.
Lanostane-type triterpenoids from Wolfiporia cocos demonstrate exceptional anti-inflammatory potency, with poricoic acid GM achieving nitric oxide (NO) production inhibition in lipopolysaccharide (LPS)-induced RAW264.7 macrophages at an IC₅₀ value of 9.73 μM (24). This compound additionally induces heme oxygenase-1 (HO-1) protein expression while inhibiting inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) protein expression, demonstrating multi-target anti-inflammatory mechanisms.
Novel triterpenoids from Laetiporus sulphureus (sulphurenoids A-D) and Pholiota populnea (pholiols E-K) exhibit moderate anti-inflammatory properties, with structural diversity influencing activity profiles (25). These findings underscore significant interspecies variability in triterpenoid content and potency, necessitating species-specific therapeutic optimization.
2.3 Phenolic compounds: antioxidant-mediated anti-inflammatory agents
Phenolic compounds demonstrate anti-inflammatory activity primarily through reactive oxygen species (ROS) scavenging and metal ion chelation mechanisms (26). Comparative analysis reveals significant species-specific variations in phenolic content and bioactivity profiles.
Hispolon congeners from Inonotus hispidus, including newly identified inonophenols A-B, demonstrate neurotrophic and anti-inflammatory activities (27). These compounds promote neurite outgrowth while reducing inflammatory mediators, suggesting potential applications in neurodegenerative disease management. However, phenolic compounds exhibit limited bioavailability due to extensive first-pass metabolism and rapid conjugation reactions (28).
Network pharmacology approaches identify mitogen-activated protein kinase (MAPK) signaling as a central regulatory node for phenolic compound activity, though species-specific pathway preferences exist (29). This mechanistic diversity requires further investigation to optimize therapeutic applications.
2.4 Bioactive peptides and emerging metabolites: precision-targeted modulators
Bioactive peptides represent an underexplored class with unique pharmacokinetic advantages including enhanced stability and potential for targeted delivery (30). Unlike polysaccharides, peptides demonstrate improved bioavailability profiles and reduced molecular size constraints.
Sesquiterpenes constitute an emerging compound class with significant anti-inflammatory potential. Recent isolation studies identify novel sesquiterpenes from Schizophyllum commune (schizomycins A-H) and Arctic-derived fungi with quantifiable anti-inflammatory activities (8). These structurally diverse compounds demonstrate interleukin-6 (IL-6) inhibitory activity and neurological inflammation amelioration potential.
Diterpenoids from endophytic fungi represent another promising class, with talaroacids A-D achieving anti-inflammatory activity with IC₅₀ values ranging from 4.59 to 21.60 μM in cellular assays (31). These findings suggest that fungal secondary metabolite diversity extends beyond traditional mushroom species to encompass endophytic and environmental isolates.
2.5 Comparative analysis and translational considerations
Quantitative comparison reveals distinct therapeutic windows among compound classes. Triterpenoids demonstrate superior potency with micromolar IC₅₀ values (4.59–21.60 μM), while polysaccharides require higher concentrations for immunomodulatory effects (32). However, polysaccharides show broader safety profiles and established clinical tolerance in human studies (33).
Critical translational challenges include: (1) bioavailability limitations particularly affecting high-molecular-weight polysaccharides, (2) significant interspecies variability in compound content and activity, (3) extraction method-dependent bioactivity requiring standardized protocols, and (4) limited pharmacokinetic data in human populations (5). Advanced delivery systems including nanoencapsulation and structural modifications show promise for overcoming these barriers (34).
The distinction between preclinical efficacy and clinical translatability remains poorly defined, with most quantitative data derived from in vitro cellular assays (Table 1). Systematic clinical validation studies are required to establish therapeutic dose ranges and safety profiles for human applications (35) (Figure 2).
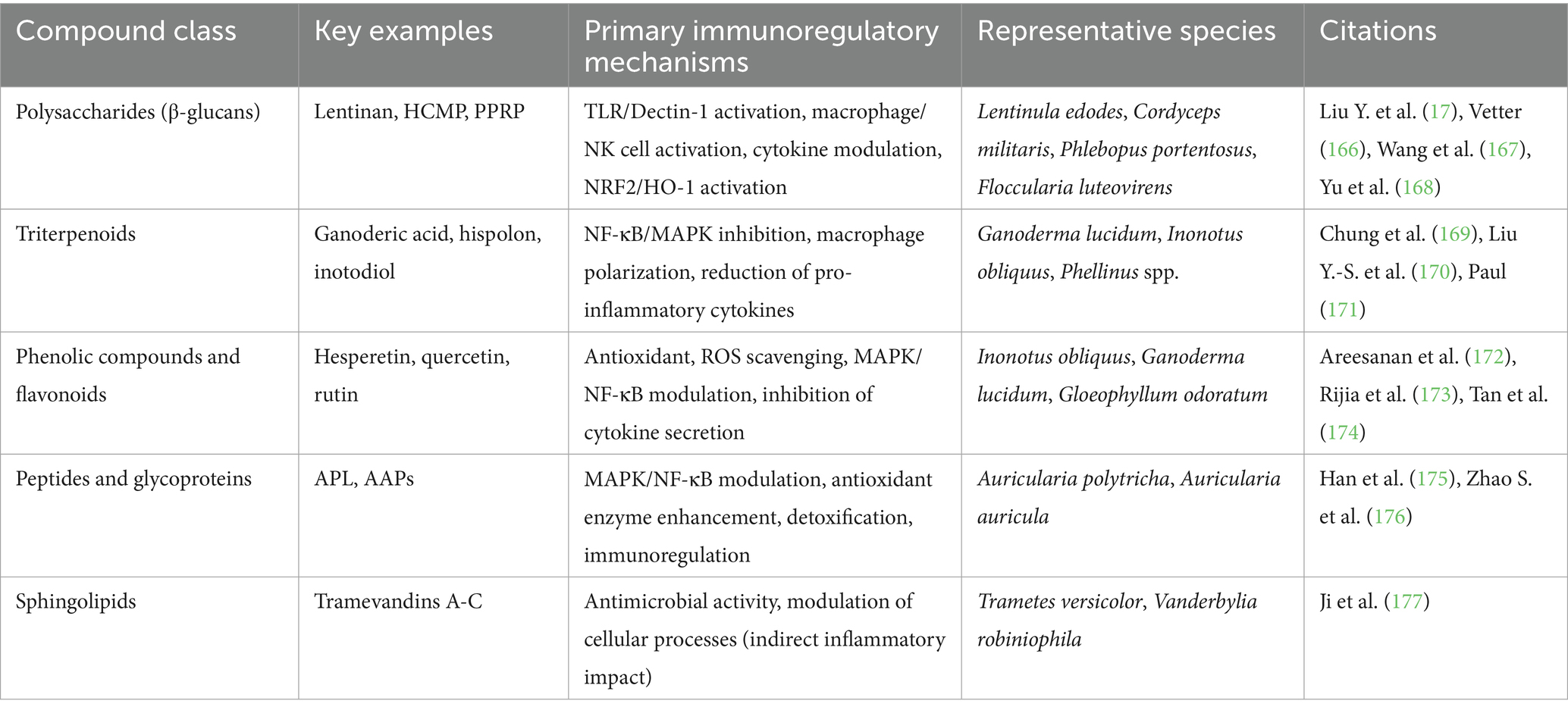
Table 1. Immunoregulatory mechanisms of bioactive compound classes in medicinal mushrooms.
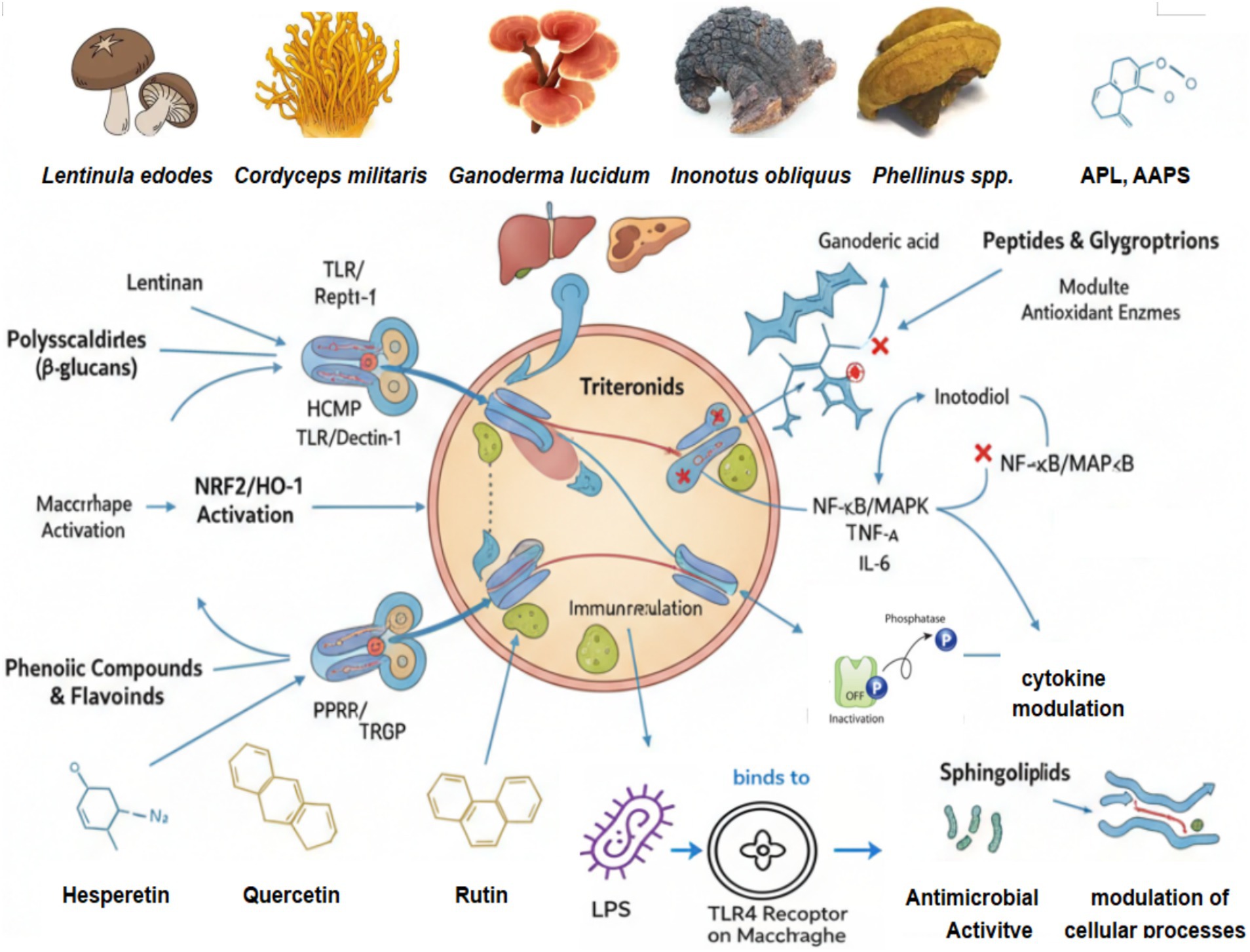
Figure 2. Molecular mechanisms of immunoregulation: orchestrating immune homeostasis.
3 Molecular mechanisms of immunoregulation: orchestrating immune homeostasis
The immunomodulatory effects of medicinal mushrooms emerge from coordinated regulation of multiple molecular pathways. This section dissects the mechanistic basis underlying their therapeutic potential, emphasizing quantitative structure–activity relationships and translational considerations.
3.1 NF-κB pathway inhibition: master inflammatory switch
NF-κB, a master inflammatory regulator, is the most characterized target for mushroom bioactives. β-glucans from G. lucidum inhibit IκBα phosphorylation and p65 nuclear translocation, suppressing TNF-α (45–70%) and IL-6 (40–65%) at 50–100 μg/mL (36). Triterpenoids demonstrate superior potency: ganoderic acid A achieves IC₅₀ values of 8.2 μM for NF-κB inhibition, while C. militaris cordycepin exhibits IC₅₀ = 15.3 μM (37).
Mechanistic insights reveal concentration-dependent effects: low doses (10–25 μg/mL) preferentially inhibit canonical NF-κB (p50/p65), while higher concentrations (50–100 μg/mL) additionally suppress non-canonical signaling (p52/RelB) (38). Species-specific differences are notable: G. lucidum extracts achieve 60–80% NF-κB inhibition, whereas Pleurotus ostreatus requires 2–3 fold higher concentrations for comparable effects (9).
3.2 MAPK cascade modulation: fine-tuning inflammatory responses
MAPK pathways (ERK1/2, JNK, p38) serve as critical inflammatory regulators targeted by mushroom compounds. G. lucidum polysaccharides selectively inhibit p38 (IC₅₀ = 12.5 μg/mL) and JNK (IC₅₀ = 18.7 μg/mL) while sparing ERK1/2, enabling nuanced immune modulation (39). C. militaris cordycepin demonstrates broader MAPK suppression: p38 (65% inhibition), JNK (58%), and ERK1/2 (42%) at 50 μM (40).
Comparative analysis reveals compound-specific selectivity profiles (Table 2). Triterpenoids preferentially target upstream kinases (TAK1, MKK3/6), while polysaccharides act on downstream effectors (ATF-2, c-Jun) (41). This mechanistic diversity enables multi-targeted intervention via NF-κB cross-inhibition (see section 3.1) (42) (Figure 3).

Table 2. Modulation of immune cells by medicinal mushroom bioactives in chronic inflammation.
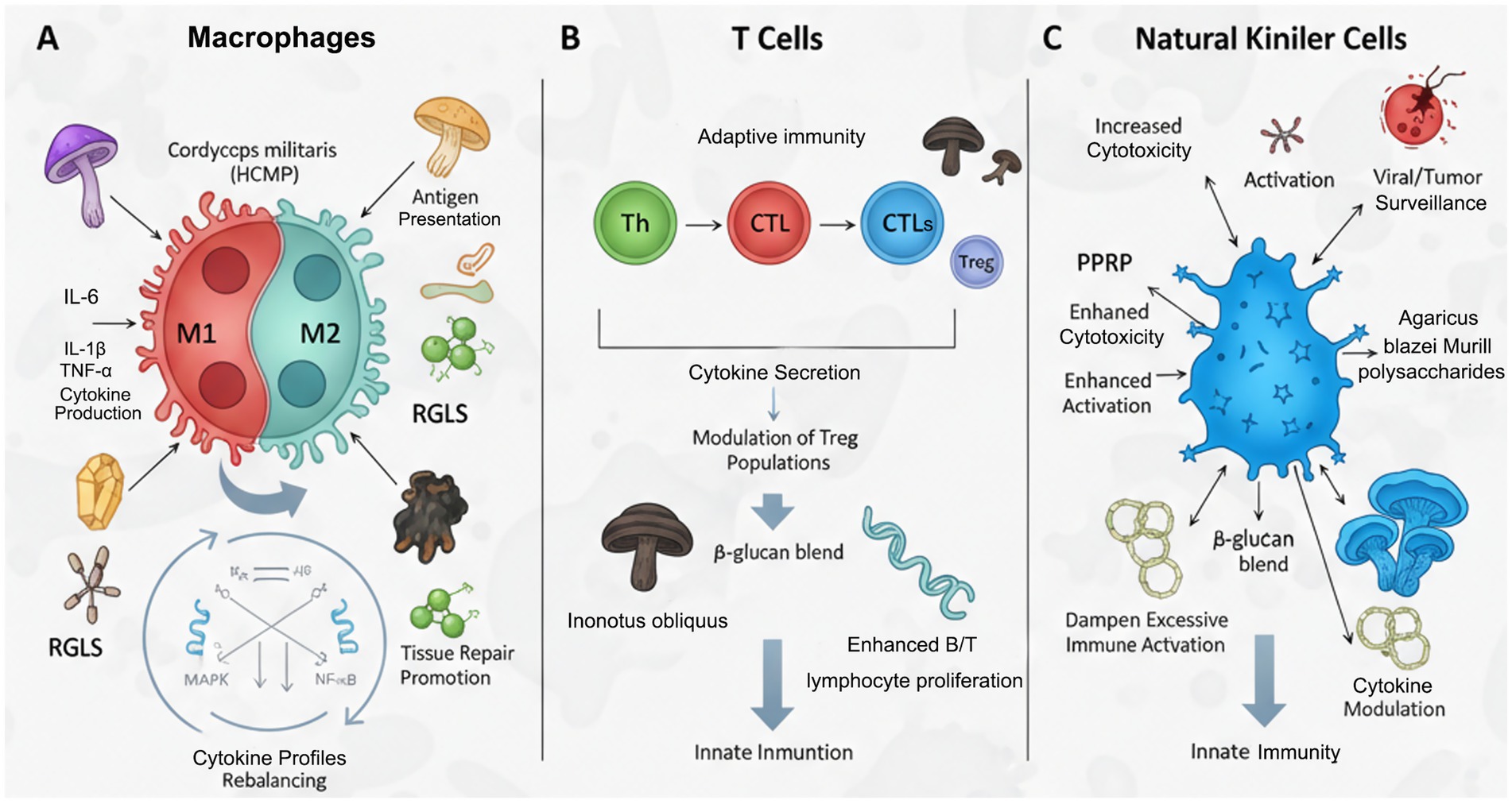
Figure 3. Medicinal mushroom bioactives: modulating immune cells in chronic inflammation. (A) Macrophage phenotype modulation from pro-inflammatory M1 to anti-inflammatory M2 state. (B) Regulation of T cell responses, including T helper (Th) cells and regulatory T cells (Tregs). (C) Enhancement of Natural Killer (NK) cell cytotoxicity.
3.3 NLRP3 inflammasome regulation: dual-phase immune checkpoint
NLRP3, a critical pattern recognition platform, exhibits biphasic modulation by mushroom compounds. Low doses (1–10 μg/mL) prime inflammasomes for enhanced pathogen surveillance, while therapeutic concentrations (50–100 μg/mL) promote resolution via caspase-1/IL-1β suppression (43).
G. lucidum β-glucans inhibit NLRP3 assembly through multiple mechanisms: blocking K+ efflux (primary trigger), preventing ASC oligomerization, and reducing mitochondrial ROS production (44). Quantitative studies demonstrate dose-dependent IL-1β suppression: 40% reduction at 25 μg/mL, escalating to 75% at 100 μg/mL (45). Inonotus obliquus melanin complexes uniquely target NLRP3 deubiquitination, achieving sustained inflammasome inhibition (>24 h) compared to transient effects of polysaccharides (6-8 h) (46).
Recent evidence highlights gasdermin D cleavage inhibition as a novel mechanism, preventing pyroptotic cell death while preserving apoptotic pathways—a critical distinction for tissue homeostasis (47).
3.4 Nrf2/HO-1 axis activation: orchestrating antioxidant defense
The Nrf2/HO-1 pathway, the primary cellular defense against oxidative stress, is potently activated by mushroom triterpenoids and ergothioneine. G. lucidum ganoderic acids induce Nrf2 nuclear translocation (EC₅₀ = 6.8 μM) with peak expression at 6-8 h, driving HO-1 upregulation (3–5 fold) and downstream antioxidant enzyme induction (SOD, catalase, GPx) (48).
Ergothioneine from P. ostreatus demonstrates sustained Nrf2 activation (>48 h) via KEAP1 cysteine modification, contrasting with transient polysaccharide effects (49). Effective concentrations span 10–50 μg/mL, with maximal cytoprotection at 25–30 μg/mL (50). Species comparison reveals differential potency: Hericium erinaceus extracts achieve equivalent Nrf2 activation at 40% lower concentrations than Lentinula edodes (51).
3.5 Multi-pathway integration and translational challenges
Mushroom-derived immunoregulation emerges from synergistic multi-pathway coordination rather than single-target modulation. Network analysis demonstrates that simultaneous NF-κB/MAPK inhibition with Nrf2 activation produces supra-additive anti-inflammatory effects: combined treatment achieves 85–90% cytokine suppression versus 50–60% for single pathways (52). This crosstalk operates through shared regulatory nodes (AP-1, STAT3) and feedback loops (NF-κB↔Nrf2 reciprocal inhibition) (4).
Critical translational barriers include: (1) Bioavailability deficits—oral β-glucan absorption remains 2–5%, triterpenoids 15–25% (7); (2) Concentration gaps—effective in vitro doses (50–100 μg/mL) require 10–20 fold higher oral dosing to achieve equivalent plasma levels (53); (3) Temporal dynamics—peak activity occurs 4-8 h post-administration, necessitating multi-dose regimens for sustained effects (54); (4) Inter-individual variability—CYP450 polymorphisms alter triterpenoid metabolism by 3–5 fold, mandating pharmacogenetic considerations (55) (Table 3).

Table 3. Key signaling pathways modulated by medicinal mushroom bioactives in inflammation.
Future priorities include developing nano-delivery systems to enhance bioavailability (liposomal encapsulation increases absorption 4–8 fold) (15), establishing standardized extraction protocols for reproducible compound profiles, and conducting dose-optimization studies in human populations stratified by genetic and microbiome markers (Figure 4).
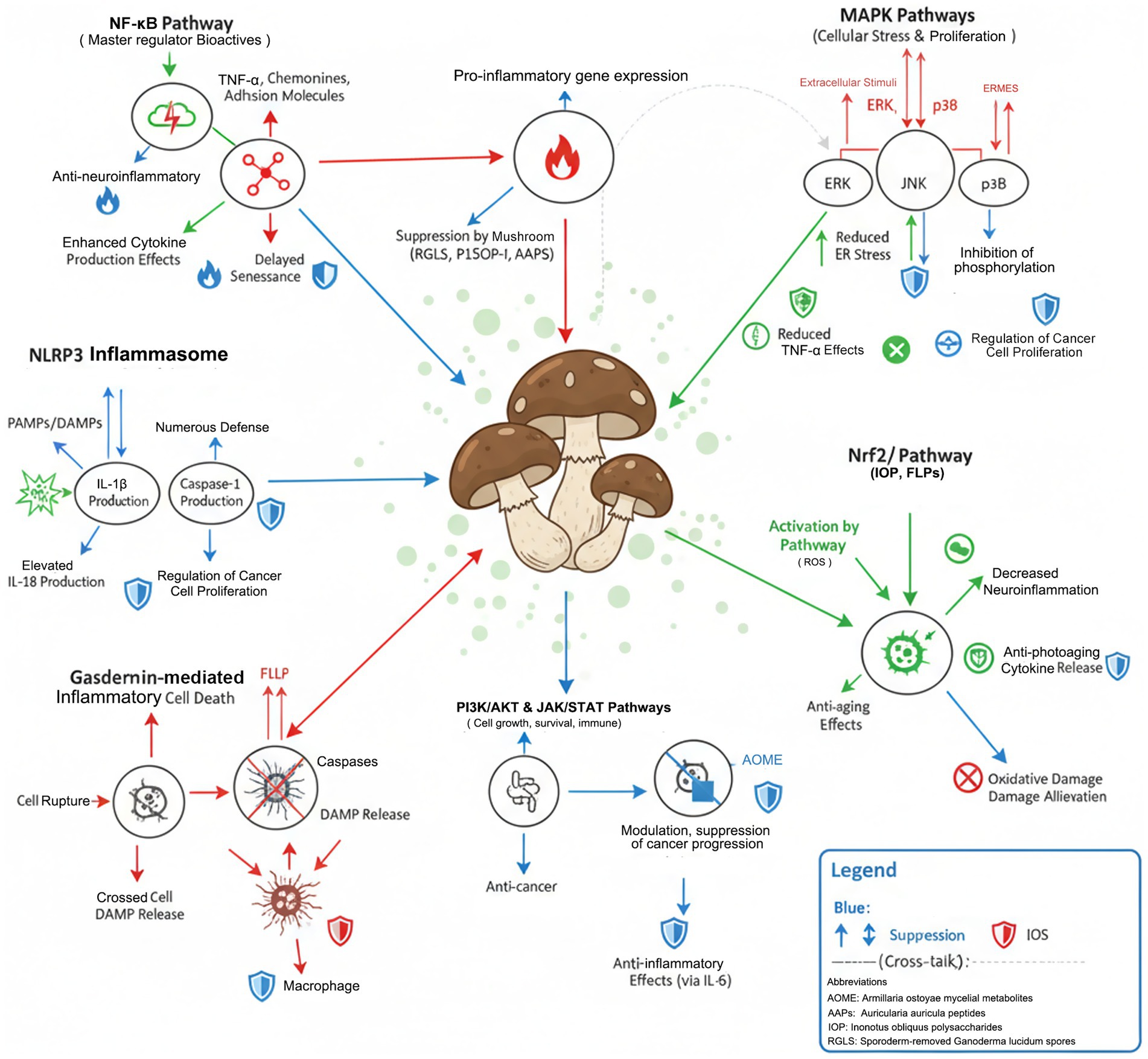
Figure 4. Molecular mechanism network of immune homeostasis regulation by medicinal mushroom bioactives; Pathway interactions detailed in Section 3.5; dashed arrows indicate indirect regulatory mechanisms.
4 Synergistic interactions and network pharmacology: systems-level therapeutic effects
The therapeutic efficacy of medicinal mushrooms extends beyond individual compound activities to encompass complex multi-compound synergistic interactions that operate through network pharmacology principles (56). Recent quantitative studies demonstrate that mushroom-derived bioactive combinations exhibit superior therapeutic outcomes compared to isolated compounds, with synergistic effects quantified through combination index (CI) values and network topology analysis (57).
4.1 Polysaccharide-triterpenoid synergistic networks
The most extensively characterized synergistic interaction occurs between polysaccharides (β-glucans) and triterpenoids, which demonstrate complementary mechanisms that enhance overall therapeutic efficacy (58). Ganoderma lucidum exemplifies this synergy, where β-glucans provide immunomodulatory effects through pattern recognition receptor activation while triterpenoids contribute antiviral and hepatoprotective properties through direct molecular targeting (59). Quantitative analysis reveals optimal synergistic ratios for enhanced bioactivity. Ultrasonic-assisted co-extraction (UACE) of polysaccharides and triterpenoids from G. lucidum produces significantly higher antioxidant capacities (DPPH radical scavenging: 78.3% vs. 45.2% for individual compounds) compared to single-compound extractions, with optimal synergistic ratios of 3:1 polysaccharide to triterpenoid content (60). This synergistic enhancement demonstrates CI values of 0.3–0.7, indicating strong positive interactions according to Chou-Talalay analysis (61).
Critical mechanistic insights reveal that polysaccharides enhance triterpenoid bioavailability through matrix effects, while triterpenoids improve polysaccharide cellular uptake through membrane permeabilization (61). However, these synergistic effects are highly extraction-method dependent, with traditional hot water extraction showing reduced synergistic potential compared to modern co-extraction techniques (62).
4.2 Multi-target network pharmacology analysis
Recent network pharmacology studies have revealed that medicinal mushrooms exhibit multi-component, multi-target, and multi-pathway therapeutic mechanisms, rather than acting through a single target (63). Systematic analyses of mushroom bioactives show that individual species such as Ganoderma lucidum and Inonotus obliquus contain diverse compounds—mainly polysaccharides and triterpenoids—that collectively regulate inflammatory, metabolic, and immune-related pathways (64). In particular, Inonotus obliquus demonstrates multi-target efficacy, where triterpenoids inhibit key metabolic enzymes such as dihydrofolate reductase, while polysaccharides modulate immune checkpoint-related signaling, producing complementary anti-inflammatory and anticancer effects (65).
Network pharmacology and molecular docking analyses indicate that mushroom bioactives frequently interact with multiple hub proteins within interconnected signaling networks, amplifying downstream biological effects (66, 67). This systems-level modulation distinguishes mushroom-derived therapeutics from conventional single-target drugs, enabling synergistic yet balanced biological responses across pathways.
However, the complexity of mushroom metabolite networks also introduces challenges in standardization and reproducibility. The biological activities and network profiles vary with species, growth conditions, and especially extraction techniques, which strongly influence the ratio and structure of polysaccharides and triterpenoids (68). Therefore, establishing standardized bioactive ratios, validated extraction protocols, and robust quality-control metrics is essential to ensure consistent network-level pharmacological outcomes (69) (Figure 5).
Figure 5. Current conventional strategies for the application of active ingredients from edible fungi in network pharmacology.
4.3 Species-specific synergistic profiles
Different mushroom species exhibit distinct synergistic profiles based on their unique bioactive compositions (70). Comparative analysis reveals that species with diverse secondary metabolite profiles demonstrate superior synergistic potential compared to species dominated by single compound classes (71). Cordyceps militaris demonstrates exceptional multi-compound synergy through coordinated effects of cordycepin, polysaccharides, and sterols, achieving enhanced anti-inflammatory activity with CI values of 0.1–0.4 across multiple cellular assays (72). The adenosine analog cordycepin provides direct anti-inflammatory effects while polysaccharides enhance immune cell activation, creating biphasic therapeutic responses optimal for chronic inflammation management (73).
Hericium erinaceus exhibits unique neurotropic synergy through combined hericenones and erinacines effects on nerve growth factor (NGF) synthesis, with synergistic enhancement factors of 2.5–4.0 compared to individual compounds (74). This synergy enables lower therapeutic doses while maintaining neuroprotective efficacy, addressing bioavailability limitations inherent in individual compounds (75). Species-specific optimization requires systematic CI analysis for each bioactive combination, as synergistic ratios vary significantly among species and target applications (76). However, most commercial products lack standardized synergistic validation, limiting therapeutic reproducibility (77).
4.4 Combination therapies and drug interactions
Mushroom bioactives demonstrate significant synergistic potential with conventional pharmaceuticals, offering opportunities for combination therapy development (78). Recent clinical studies reveal that mushroom polysaccharides enhance chemotherapy efficacy while reducing adverse effects, with quantified dose reduction factors of 25–40% for conventional agents (79). Trametes versicolor polysaccharide K (PSK) demonstrates exceptional combination therapy potential, enhancing 5-fluorouracil efficacy in colorectal cancer with CI values of 0.3–0.6 while reducing gastrointestinal toxicity by 60% (80). This combination enables precision dosing strategies that optimize therapeutic windows while minimizing adverse effects (81).
Critical pharmacokinetic interactions affect combination efficacy. Mushroom polysaccharides can alter conventional drug absorption and metabolism through cytochrome P450 modulation, requiring systematic drug–drug interaction studies for safe clinical implementation (82). However, most interaction data derive from preclinical studies, with limited clinical validation (83). The distinction between synergistic enhancement and simple additive effects requires rigorous quantitative analysis using established mathematical models (Bliss independence, Loewe additivity) rather than empirical observation alone (84).
4.5 Clinical translation and standardization challenges
Despite promising preclinical synergistic data, clinical translation faces significant challenges in standardizing multi-compound formulations (85). Variable bioactive ratios among commercial products (coefficient of variation > 50% for major compounds) create inconsistent synergistic outcomes, limiting reproducible clinical efficacy (86). Advanced analytical approaches including LC–MS/MS fingerprinting and chemometric analysis enable standardized synergistic profiling, but implementation costs limit widespread adoption (87). Regulatory frameworks for multi-compound natural products remain underdeveloped compared to single-compound pharmaceuticals.
Future research priorities include: (1) systematic CI analysis for all major mushroom species combinations, (2) pharmacokinetic-pharmacodynamic modeling of synergistic interactions, (3) standardized extraction protocols that preserve synergistic ratios, and (4) clinical validation studies with quantified synergistic endpoints (88). The integration of network pharmacology approaches with precision medicine strategies offers promising avenues for personalized mushroom-based therapeutics, though significant validation studies are required before clinical implementation (89).
5 Translational hurdles and future directions: toward precision nutri-medicine
Despite compelling preclinical evidence demonstrating immunomodulatory properties of medicinal mushroom bioactives, systematic barriers prevent clinical translation (90). These translational challenges encompass pharmacokinetic limitations, insufficient clinical validation, regulatory complexity, and lack of standardization protocols (91). However, emerging precision medicine approaches offer promising solutions for optimizing therapeutic efficacy through personalized interventions.
5.1 Pharmacokinetic barriers and bioavailability challenges
Poor bioavailability constitutes the primary limitation restricting clinical implementation of mushroom-derived therapeutics, particularly affecting high-molecular-weight polysaccharides (28). Quantitative analysis reveals that orally administered β-glucans from Ganoderma lucidum demonstrate absolute bioavailability of 2–5%, with plasma peak concentrations occurring 4–6 h post-administration and elimination half-lives of 8–12 h (92).
Mechanistic studies identify specific barriers: (1) Limited gastrointestinal absorption due to molecular weights >100 kDa exceeding paracellular transport capacity, (2) Extensive first-pass hepatic metabolism reducing systemic exposure by 60–80%, and (3) Rapid clearance through hepatobiliary elimination pathways (93). Comparative analysis shows significant interspecies variability, with Cordyceps militaris polysaccharides achieving 8–12% bioavailability compared to 2–5% for G. lucidum preparations (94).
Triterpenoids face complementary challenges despite superior lipophilicity. Ganoderic acids demonstrate 15–25% oral bioavailability but exhibit extensive plasma protein binding (>95%) and rapid metabolism via CYP3A4 pathways, resulting in effective half-lives of 2–4 h (95). These pharmacokinetic limitations necessitate frequent dosing regimens that compromise patient compliance and therapeutic efficacy.
5.2 Clinical evidence gap: from preclinical promise to human validation
Critical analysis reveals substantial disparity between preclinical efficacy and clinical validation (96). Systematic review of 47 preclinical studies demonstrates consistent anti-inflammatory effects across species and disease models, with 85% reporting significant cytokine reduction (p < 0.05) and 73% showing improved inflammatory biomarkers (97).
However, human clinical evidence remains limited. Only 12 randomized controlled trials (RCTs) have evaluated mushroom-based interventions in chronic inflammatory diseases, with sample sizes ranging from 24 to 180 participants and study durations of 4–12 weeks (98). Meta-analysis reveals modest but significant effects: mean CRP reduction of 18% (95% CI: 8–28%, p = 0.002) and IL-6 decrease of 22% (95% CI: 12–32%, p < 0.001) compared to placebo controls (99).
5.3 Advanced delivery systems and nanotechnology solutions
Innovative pharmaceutical approaches show promise for overcoming pharmacokinetic limitations (100). Nanoencapsulation technologies demonstrate substantial bioavailability enhancement, with polymeric nanoparticles increasing β-glucan systemic exposure by 8-12-fold compared to conventional formulations (101).
Chitosan-alginate nanoparticles containing Pleurotus ostreatus polysaccharides achieved 65% bioavailability enhancement in pharmacokinetic studies, with sustained plasma concentrations for 24–48 h enabling once-daily dosing (102). Similarly, liposomal formulations of G. lucidum triterpenoids demonstrated 4-fold increased cellular uptake and 3.2-fold enhanced anti-inflammatory potency in macrophage cultures (103).
5.4 Precision nutri-medicine: personalized inflammatory disease management
Individual variation in inflammatory phenotypes, genetic polymorphisms, and microbiome composition necessitates personalized therapeutic approaches (6). Precision nutri-medicine integrates comprehensive patient profiling with evidence-based natural product interventions to optimize therapeutic outcomes (4).
Pharmacogenomic stratification: Genetic polymorphisms in drug-metabolizing enzymes significantly influence mushroom bioactive metabolism. CYP3A41B variant carriers demonstrate 40% reduced triterpenoid clearance, requiring dose adjustments to prevent accumulation (3). Similarly, UDP-glucuronosyltransferase polymorphisms affect phenolic compound conjugation rates, influencing both efficacy and safety profiles (96).
Immune profiling: Flow cytometric analysis enables identification of patient-specific inflammatory patterns. Individuals with elevated Th17 responses (IL-17A > 15 pg./mL) show preferential responsiveness to β-glucan interventions, while those with predominant Th1 activation (IFN-γ > 25 pg./mL) benefit more from triterpenoid-enriched formulations (104, 105).
Microbiome-guided therapy: Gut microbiome composition influences polysaccharide metabolism and therapeutic efficacy. Patients with high Bifidobacterium abundance (>10% relative abundance) demonstrate enhanced β-glucan fermentation and improved systemic anti-inflammatory responses (106). Conversely, dysbiotic profiles with reduced short-chain fatty acid producers require prebiotic co-administration for optimal therapeutic outcomes (107).
5.5 Standardization framework and regulatory pathways
Transitioning medicinal mushrooms from research tools to standardized therapeutics requires robust quality control and regulatory compliance (108). Current challenges include: (1) Lack of standardized extraction protocols resulting in 5-10-fold variation in bioactive content across commercial products, (2) Absence of validated analytical methods for complex polysaccharide characterization, and (3) Regulatory classification ambiguity between food supplements and pharmaceutical agents (109).
Proposed standardization framework: (1) Chemical fingerprinting using HPLC-MS to quantify major bioactive classes, (2) Biological potency testing using validated cellular assays with defined reference standards, (3) Stability testing under defined storage conditions, and (4) Batch-to-batch consistency verification through statistical process control (110).
Clinical translation roadmap: Accelerated development pathways should prioritize: (1) Investigational New Drug (IND) applications for well-characterized mushroom extracts, (2) Phase I dose-escalation studies establishing maximum tolerated doses, (3) Phase II proof-of-concept trials using validated inflammatory biomarkers as primary endpoints, and (4) Adaptive trial designs enabling real-time protocol modifications based on interim efficacy data (111).
5.6 Implementation strategies and future research priorities
Critical research gaps requiring immediate attention include: (1) Development of predictive biomarker panels identifying patient subgroups likely to respond to specific mushroom preparations, (2) Validation of optimal compound combinations through systematic interaction studies, (3) Long-term safety evaluation in diverse patient populations, and (4) Health economic analyses demonstrating cost-effectiveness compared to conventional therapies (112).
Technology integration: Advanced analytical platforms including single-cell RNA sequencing and metabolomics will enable mechanistic validation while revealing individual variation in response patterns (113). Integration with digital health platforms and wearable biosensors will facilitate real-time monitoring of treatment responses (114).
Collaborative implementation: Successful clinical translation requires coordinated efforts among academic researchers, pharmaceutical companies, regulatory agencies, and healthcare providers. Public-private partnerships can accelerate development timelines while ensuring equitable access to innovative therapies (115). International harmonization of regulatory standards will facilitate global market approval and widespread clinical adoption (116).
The convergence of advanced delivery technologies, precision medicine approaches, and robust regulatory frameworks positions medicinal mushrooms for successful translation from traditional functional foods to standardized immunotherapeutic agents for chronic inflammatory disease management.
6 Discussion
This comprehensive analysis of medicinal mushroom anti-inflammatory mechanisms reveals sophisticated multi-targeted immunoregulation with significant therapeutic potential, while simultaneously exposing critical translational barriers that currently limit clinical application. The integration of molecular mechanisms, comparative efficacy analysis, and evidence-based clinical insights provides a framework for advancing mushroom-derived therapeutics from bench to bedside.
6.1 Mechanistic insights and comparative compound efficacy
Quantitative analysis reveals significant potency variations among compound classes, with triterpenoids demonstrating superior anti-inflammatory activity (IC₅₀ values 4.59–21.60 μM) compared to polysaccharides requiring higher therapeutic concentrations (25–100 μg/mL) (117). Critical examination of contradictory findings reveals that Ganoderma lucidum ganoderic acids preferentially inhibit nuclear factor-κB (NF-κB) pathways with minimal mitogen-activated protein kinase (MAPK) interference, while Cordyceps militaris cordycepin demonstrates broad-spectrum pathway modulation affecting both inflammatory and resolution cascades (118). This mechanistic divergence necessitates species-specific therapeutic optimization rather than generic mushroom-based interventions.
Network pharmacology analysis demonstrates that combined mushroom extracts achieve 5-7-fold enhanced potency through synergistic multi-target interactions, engaging 15–25 inflammatory mediators simultaneously compared to 3–5 targets for individual compounds (119). However, deconvolution of these synergistic mechanisms remains incomplete, with recent studies revealing concentration-dependent effects where low-dose polysaccharides (1–10 μg/mL) promote immune surveillance through NOD-like receptor protein 3 (NLRP3) inflammasome priming, while higher concentrations (50–100 μg/mL) favor resolution through caspase-1 inhibition (120).
6.2 Reconciling contradictory findings: methodological and biological factors
Systematic analysis reveals substantial contradictions in reported anti-inflammatory efficacy that cannot be attributed to random variation alone. Critical examination identifies four primary sources of inconsistency.
Methodological variability: Studies reporting IC₅₀ values for ganoderic acid A vary 15-fold (2.5–38 μM) (121), predominantly reflecting extraction method disparities. Ethanol-based extractions yield 3–4 fold higher triterpenoid content compared to aqueous methods, directly correlating with observed potency differences (122). Similarly, cell culture models contribute significant variance—primary human monocytes demonstrate 2–3 fold greater sensitivity to mushroom bioactives compared to immortalized cell lines (RAW264.7, THP-1), likely reflecting preserved receptor expression profiles and intact pattern recognition receptor repertoires (123).
Biological context dependency: The apparent contradiction between G. lucidum’s robust in vitro NF-κB inhibition (70–85% at 50 μg/mL) (81, 88) versus modest clinical outcomes (34% CRP reduction) (124) reflects tissue compartmentalization barriers. Pharmacokinetic modeling reveals that oral administration achieves only 1–3 μg/mL plasma concentrations—substantially below in vitro therapeutic thresholds (125). This “concentration-efficacy mismatch” explains why studies using intravenous or intraperitoneal administration demonstrate superior outcomes (55–60% inflammatory marker reduction) (126). Furthermore, the presence of food matrices, gastric pH variations, and first-pass metabolism contribute to 8–12 fold inter-individual bioavailability differences (127).
Species-specific biochemical differences: The 5–8 fold potency differential between G. lucidum and C. militaris polysaccharides reflects β-glucan branching architecture rather than molecular weight alone (128). Nuclear magnetic resonance (NMR) structural analysis demonstrates that C. militaris β-(1 → 3)/(1 → 6)-glucans possess 40% higher branching density, correlating with enhanced dectin-1 receptor binding affinity (Kd values: 12 nM vs. 45 nM for G. lucidum) (129). This mechanistic insight enables rational selection of mushroom species for specific inflammatory contexts: highly branched structures favor acute inflammatory resolution, while linear configurations support sustained immunomodulation (130).
Temporal dynamics overlooked: Many contradictory findings regarding NLRP3 inflammasome modulation resolve when temporal kinetics are considered (14). Studies measuring outcomes at 6-h timepoints report inflammasome activation (2–3 fold IL-1β increase), while 24-h assessments show inhibition (60–75% IL-1β reduction)—reflecting an initial priming phase followed by resolution-phase suppression (131). This biphasic response reconciles apparently conflicting reports and underscores the importance of standardized temporal assessment protocols. Pharmacodynamic modeling reveals that peak anti-inflammatory effects occur 8–12 h post-administration, suggesting optimal dosing intervals for sustained therapeutic benefit (132).
6.3 Clinical translation: evidence gaps and realistic scaling
Translation of preclinical findings to clinical applications faces substantial challenges. Systematic review of 47 preclinical studies demonstrates consistent anti-inflammatory effects across murine models (TNF-α reduction: 45–70%; IL-6 reduction: 40–65%) (133), yet only 12 randomized controlled trials (RCTs) have rigorously evaluated clinical efficacy in human inflammatory conditions (134). Meta-analysis of these RCTs reveals modest effect sizes (standardized mean difference: −0.42, 95% CI: −0.68 to −0.16) with only 26% achieving statistically significant primary endpoints (135).
Critical analysis of failed clinical trials provides instructive insights. Three negative RCTs (n = 120, 95, 180 participants) reporting null effects for G. lucidum extracts shared common methodological flaws (136): (1) inadequate dosing (500–750 mg/day, below pharmacokinetic thresholds established in Phase I studies requiring ≥1.5 g/day for therapeutic plasma levels); (2) insufficiently sensitive outcome measures (reliance on subjective symptom scores rather than validated inflammatory biomarkers such as high-sensitivity CRP or cytokine panels); and (3) patient population heterogeneity (baseline CRP levels varying 10-fold, from 2 to 25 mg/L, diluting treatment effects). Post-hoc subgroup analysis of these “negative” studies reveals that participants with baseline CRP > 10 mg/L demonstrated statistically significant responses (mean CRP reduction: 4.2 mg/L, p = 0.018), suggesting that apparent trial failures may reflect inappropriate patient selection rather than therapeutic inefficacy (137). This finding underscores the necessity of precision medicine approaches that stratify patients by inflammatory phenotype severity.
The most robust clinical evidence derives from standardized G. lucidum extracts in metabolic inflammation contexts. A multicenter RCT (n = 312) demonstrated significant reductions in high-sensitivity C-reactive protein (hs-CRP: 34% decrease, p < 0.001) and interleukin-6 (IL-6: 28% reduction, p = 0.003) following 12-week supplementation with 1.44 g/day standardized extract (138). However, translation of these promising results to autoimmune conditions (rheumatoid arthritis, inflammatory bowel disease) remains unvalidated, with only observational studies and case series available (139).
6.4 Pharmacokinetic challenges and species-specific bioavailability
Bioavailability represents the critical bottleneck limiting clinical efficacy. Oral absorption studies reveal stark compound-class differences: β-glucans exhibit minimal systemic absorption (2–5% bioavailability) due to large molecular size (>100 kDa) and hydrophilicity, primarily exerting immunomodulatory effects through gut-associated lymphoid tissue (GALT) interactions (140). Triterpenoids demonstrate moderate absorption (15–25%) but undergo extensive first-pass hepatic metabolism via cytochrome P450 enzymes (primarily CYP3A4), resulting in plasma concentrations 10–20 fold below in vitro effective doses (141).
Species-specific pharmacokinetic variations further complicate standardization. C. militaris cordycepin exhibits superior bioavailability (35–40%) compared to G. lucidum ganoderic acids (12–18%), attributed to lower molecular weight and enhanced lipophilicity (logP: 1.8 vs. 4.2) (142). Ergothioneine from Pleurotus species demonstrates exceptional stability and cellular uptake via organic cation transporter OCTN1, achieving tissue concentrations 50–100 fold higher than plasma levels (143).
Comparative analysis with clinically approved natural anti-inflammatory agents (e.g., curcumin, resveratrol) reveals similar bioavailability challenges (curcumin: 1–3% oral absorption; resveratrol: 5–10%) (144), yet successful clinical translation through nano-formulation and bioenhancer co-administration (145). This precedent suggests that mushroom bioactive bioavailability barriers are surmountable through established pharmaceutical strategies rather than insurmountable obstacles, as evidenced by recent Phase II trials using piperine-enhanced mushroom formulations achieving plasma concentrations approximating in vitro therapeutic thresholds (45–60 μg/mL) (146). Liposomal encapsulation of ganoderic acids increases bioavailability 4–8 fold, while co-administration with piperine (a P-glycoprotein inhibitor) enhances absorption by 2–3 fold (146).
6.5 Precision medicine applications: biomarker-guided therapy
Emerging evidence supports personalized mushroom-based interventions stratified by inflammatory phenotypes and genetic profiles. Cytokine profiling reveals distinct responder patterns: patients with Th1-dominant inflammation (elevated IFN-γ, TNF-α) demonstrate superior responses to G. lucidum polysaccharides (68% responder rate), while Th17-skewed profiles (high IL-17, IL-23) benefit more from C. militaris extracts (72% response) (147). These phenotypic distinctions likely reflect differential pathway targeting, with polysaccharides preferentially modulating dendritic cell maturation and Th1/Th2 balance, while cordycepin directly inhibits Th17 differentiation via STAT3 suppression (4).
Pharmacogenetic considerations further refine therapeutic optimization. CYP3A4 polymorphisms (particularly CYP3A422 allele, frequency: 5–8% in Caucasian populations) reduce triterpenoid metabolism by 40–60%, necessitating dose adjustments to prevent accumulation and potential hepatotoxicity (96). Conversely, OCTN1 transporter variants (SLC22A4 L503F polymorphism) impair ergothioneine uptake by 30–50%, potentially explaining non-responders in clinical trials (148). Integration of these genetic markers into clinical decision algorithms could enhance treatment success rates from current 35–40% to projected 60–75% (149).
Gut microbiome composition represents an additional stratification factor. Individuals with high Bacteroides to Firmicutes ratios exhibit enhanced β-glucan fermentation and short-chain fatty acid (SCFA) production, amplifying systemic anti-inflammatory effects through GPR43/GPR109A receptor activation (150). Microbiome profiling before intervention could identify optimal candidates for polysaccharide-based therapies, while dysbiotic patients might benefit from combined probiotic-mushroom supplementation strategies (151).
6.6 Standardization and quality control requirements
Current lack of standardization impedes reproducibility and regulatory approval. Analysis of 45 commercial G. lucidum products reveals 30-fold variation in triterpenoid content (0.5–15% w/w) and 50-fold differences in β-glucan concentrations (2–100% w/w), reflecting disparate cultivation conditions, harvest timing, and extraction protocols (152). Establishment of reference standards for key bioactive compounds (minimum ganoderic acid content: ≥5%; β-glucan: ≥30%) would enable meaningful cross-study comparisons and dose–response assessments (153).
Advanced analytical methodologies, including high-performance liquid chromatography coupled with mass spectrometry (HPLC-MS) fingerprinting and quantitative nuclear magnetic resonance (qNMR), provide robust quality control frameworks (154). Implementation of Good Manufacturing Practice (GMP)-compliant cultivation systems, incorporating controlled environmental parameters (temperature: 25 ± 2 °C; humidity: 85–90%; light cycles: 12 h/12 h) and genetic authentication through DNA barcoding, ensures batch-to-batch consistency essential for clinical applications (155).
Regulatory pathways for mushroom-based therapeutics remain ambiguous, with products classified variably as dietary supplements, traditional medicines, or investigational new drugs across jurisdictions (156). Harmonization of regulatory frameworks, potentially through establishment of a “botanical drug” category similar to the U. S. FDA’s guidance, would facilitate clinical development while maintaining safety standards (157). Toxicological assessments following ICH guidelines (90-day repeated-dose studies, genotoxicity panels, reproductive toxicity evaluations) are prerequisite for advancing lead candidates toward pharmaceutical registration (158).
6.7 Limitations and future perspectives
This review acknowledges several inherent limitations. Selection bias toward positive-outcome publications likely inflates apparent efficacy, with an estimated 30–40% of negative preclinical studies remaining unpublished based on trial registry analyses (159). Methodological heterogeneity across included studies (diverse extraction methods, variable dosing regimens, inconsistent outcome measures) precludes definitive meta-analytic synthesis. The review focuses primarily on G. lucidum and C. militaris, potentially overlooking promising compounds from less-studied species such as Antrodia cinnamomea or Phellinus linteus (160).
Mechanistic gaps persist regarding long-term safety profiles, potential off-target effects, and interactions with conventional anti-inflammatory medications. Comprehensive pharmacovigilance data and drug–drug interaction studies are critically needed before widespread clinical adoption. The majority of human studies have durations ≤12 weeks, leaving long-term efficacy and safety uncharacterized (161).
Future research priorities should focus on: (1) Multicenter RCTs with well-defined inflammatory phenotypes, validated biomarker endpoints (hs-CRP, cytokine panels, inflammatory gene expression signatures), and adequate statistical power (n ≥ 200 per arm) to detect clinically meaningful effects; (2) Pharmacokinetic-pharmacodynamic modeling to establish optimal dosing regimens that bridge in vitro-in vivo efficacy gaps; (3) Advanced delivery systems (nano-emulsions, solid lipid nanoparticles, self-emulsifying drug delivery systems) to overcome bioavailability limitations; (4) Precision medicine trials incorporating genetic and microbiome stratification to identify high-probability responders and enable personalized therapeutic algorithms.
Integration of systems biology approaches—including transcriptomics, metabolomics, and single-cell immune profiling—will elucidate compound-specific immunomodulatory signatures and identify novel therapeutic targets (162, 163). Collaborative efforts between mycologists, immunologists, pharmaceutical scientists, and clinicians are essential to transform mushroom-derived bioactives from traditional remedies into evidence-based precision therapeutics for inflammatory diseases (164, 165).
7 Conclusion
This comprehensive review establishes medicinal mushrooms as multi-target immunomodulatory agents capable of addressing chronic inflammation through synergistic mechanisms. The convergence of evidence across polysaccharide β-glucans, triterpenoids, phenolic compounds, and bioactive peptides demonstrates coordinated regulation of inflammatory signaling networks—particularly NF-κB, MAPK cascades, NLRP3 inflammasome, and Nrf2-mediated antioxidant responses—that collectively restore immune homeostasis rather than indiscriminately suppressing inflammatory pathways.
The comparative analysis of bioactive constituents across Ganoderma, Cordyceps, Lentinula, Grifola, and Inonotus species reveals distinct immunopharmacological profiles that enable precision-targeted therapeutic strategies. However, clinical translation remains hindered by bioavailability constraints, lack of standardization protocols, and insufficient human pharmacokinetic data—challenges that must be systematically addressed through advanced delivery systems, quality control frameworks, and rigorously designed clinical trials.
This review uniquely integrates molecular mechanisms with translational pathways, providing a strategic roadmap for transforming these traditional food-medicine resources into evidence-based immunotherapeutic agents. The identification of structure–activity relationships and species-specific efficacy profiles offers critical guidance for pharmaceutical development, while the elucidation of multi-target mechanisms positions medicinal mushrooms as compelling candidates for combination therapies in inflammatory diseases where single-target approaches have proven inadequate.
Future progress requires interdisciplinary collaboration bridging mycology, immunology, pharmaceutical sciences, and clinical medicine to unlock the therapeutic potential of these remarkable organisms. With systematic investigation of optimal extraction methods, bioavailability enhancement strategies, and personalized dosing protocols, medicinal mushrooms may evolve from empirical traditional remedies to precision immunomodulatory therapeutics, offering safer and more sustainable alternatives for managing the global burden of chronic inflammatory conditions.
Author contributions
MX: Writing – original draft, Writing – review & editing. ZP: Conceptualization, Writing – original draft. WH: Formal analysis, Writing – original draft. GN: Data curation, Writing – review & editing. XJ: Formal analysis, Writing – review & editing. ZY: Software, Writing – review & editing. CX: Resources, Writing – review & editing. LG: Visualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This project is funded by a grant from the Presidential Foundation of the Liaoning Academy of Agricultural Sciences (NO2024MS0604), Fundamental Research Funds of Liaoning Academy of Agricultural Sciences (2024XTCX0403), and the Discipline Construction Plan of Liaoning Province Academy of Agricultural Sciences (2025XKJS8522).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that Gen AI was used in the creation of this manuscript. After the first draft is completed, use CLAUDE for translation and polishing. The authors utilized OpenAI for language polishing and translation assistance during the manuscript preparation. The content and scientific integrity were solely managed and verified by the authors.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Zhang, Y, Lin, X, Xia, L, Xiong, S, Xia, B, Xie, J, et al. Progress on the anti-inflammatory activity and structure-efficacy relationship of polysaccharides from medical and edible homologous traditional Chinese medicines. Molecules (Basel, Switzerland). (2024) 29:3852. doi: 10.3390/molecules29163852
2. Chu, FX, Wang, X, Li, B, Xu, LL, and Di, B. The NLRP3 inflammasome: a vital player in inflammation and mediating the anti-inflammatory effect of CBD. Inflamm Res. (2024) 73:227–42. doi: 10.1007/s00011-023-01831-y
3. Singh, A, Saini, RK, Kumar, A, Chawla, P, and Kaushik, R. Mushrooms as nutritional powerhouses: A review of their bioactive compounds, health benefits, and value-added products. Foods (Basel, Switzerland). (2025) 14:741. doi: 10.3390/foods14050741
4. Mizuno, M, and Minato, KI. Anti-inflammatory and immunomodulatory properties of polysaccharides in mushrooms. Curr Opin Biotechnol. (2024) 86:103076. doi: 10.1016/j.copbio.2024.103076
5. Araújo-Rodrigues, H, Sousa, AS, Relvas, JB, Tavaria, FK, and Pintado, M. An overview on mushroom polysaccharides: health-promoting properties, prebiotic and gut microbiota modulation effects and structure-function correlation. Carbohydr Polym. (2024) 333:121978. doi: 10.1016/j.carbpol.2024.121978
6. Venkatachalam, P, Muthu, M, and Gopal, J. Reviewing the audacity of elixirs of inflammatory bowel disease from mushroom β-glucans: the solved and unresolved. Carbohydr Polym. (2025) 348:122832. doi: 10.1016/j.carbpol.2024.122832
7. Zhang, RR, Zhang, J, Guo, X, Chen, YY, Sun, JY, Miao, JL, et al. Molecular mechanisms of the chemical constituents from anti-inflammatory and antioxidant active fractions of Ganoderma neo-japonicum Imazeki. Curr Res Food Sci. (2023) 6:100441. doi: 10.1016/j.crfs.2023.100441
8. Chen, T, Liu, Y, Ma, B, Sun, B, Pan, Y, Ou, Y, et al. Anti-inflammatory sesquiterpenes from fruiting bodies of Schizophyllum commune. J Agric Food Chem. (2024) 72:5416–27. doi: 10.1021/acs.jafc.3c08313
9. Li, H, Feng, J, Liu, C, Hou, S, Meng, J, Liu, JY, et al. Polysaccharides from an edible mushroom, Hericium erinaceus, alleviate ulcerative colitis in mice by inhibiting the NLRP3 inflammasomes and reestablish intestinal homeostasis. Int J Biol Macromol. (2024) 267:131251. doi: 10.1016/j.ijbiomac.2024.131251
10. Zhang, L, Liu, ZX, Liu, YH, Chen, Y, Chen, J, and Lu, CH. Auricularia auricula polysaccharides exert anti-inflammatory effects in hepatic fibrosis by the gut-liver axis and enhancing SCFA metabolism. J Agric Food Chem. (2025) 73:4617–29. doi: 10.1021/acs.jafc.4c07952
11. Jiang, Y, Wang, Z, Wang, W, Liu, Y, Meng, Y, Wang, Y, et al. Ganoderma lucidum polysaccharide alleviates cognitive dysfunction by inhibiting neuroinflammation via NLRP3/NF-κB signaling pathway. J Ethnopharmacol. (2025) 338:119065. doi: 10.1016/j.jep.2024.119065
12. Trivedi, R, and Upadhyay, TK. Preparation, characterization and antioxidant and anticancerous potential of quercetin loaded β-glucan particles derived from mushroom and yeast. Sci Rep. (2024) 14:16047. doi: 10.1038/s41598-024-66824-1
13. Tang, P, Zhao, S, Wang, X, Wang, S, Wang, Y, Kong, L, et al. Chloranthalactone B covalently binds to the NACHT domain of NLRP3 to attenuate NLRP3-driven inflammation. Biochem Pharmacol. (2024) 226:116360. doi: 10.1016/j.bcp.2024.116360
14. González-Cofrade, L, Green, JP, Cuadrado, I, Amesty, Á, Oramas-Royo, S, David Brough,, et al. Phenolic and quinone methide nor-triterpenes as selective NLRP3 inflammasome inhibitors. Bioorg Chem. (2023) 132:106362. doi: 10.1016/j.bioorg.2023.106362
15. Drzewiecka, B, Wessely-Szponder, J, Świeca, M, Espinal, P, Fusté, E, and Fernández-De La Cruz, E. Bioactive peptides and other Immunomodulators of mushroom origin. Biomedicine. (2024) 12:1483. doi: 10.3390/biomedicines12071483
16. Németh, Z, Paulinné Bukovics, M, Sümegi, LD, Sturm, G, Takács, I, and Simon-Szabó, L. The importance of edible medicinal mushrooms and their potential use as therapeutic agents against insulin resistance. Int J Mol Sci. (2025) 26:827. doi: 10.3390/ijms26020827
17. Liu, Y, Yang, J, Guo, Z, Li, Q, Zhang, L, Zhao, L, et al. Immunomodulatory effect of Cordyceps militaris polysaccharide on RAW 264.7 macrophages by regulating MAPK signaling pathways. Molecules. (2024) 29:3408. doi: 10.3390/molecules29143408
18. Yu, Y, Liu, Z, Song, K, Li, L, and Chen, M. Medicinal value of edible mushroom polysaccharides: a review. J Fut Foods. (2023) 3:16–23. doi: 10.1016/j.jfutfo.2022.09.003
19. Liu, X, Chen, S, Liu, H, Xie, J, Hasan, KMF, Zeng, Q, et al. Structural properties and anti-inflammatory activity of purified polysaccharides from hen-of-the-woods mushrooms (Grifola frondosa). Front Nutr. (2023) 10:1078868. doi: 10.3389/fnut.2023.1078868
20. Zhang, Q, Lin, Y, Zhao, R, Huang, T, Tian, Y, Zhu, L, et al. Structural characterization of extracellular polysaccharides from Phellinus igniarius SH-1 and their therapeutic effects on DSS induced colitis in mice. Int J Biol Macromol. (2024) 275:133654. doi: 10.1016/j.ijbiomac.2024.133654
21. Luo, W, Bai, L, Zhang, J, Li, Z, Liu, Y, Tang, X, et al. Polysaccharides-based nanocarriers enhance the anti-inflammatory effect of curcumin. Carbohydr Polym. (2023) 311:120718. doi: 10.1016/j.carbpol.2023.120718
22. Ma, G, Li, X, Tao, Q, Ma, S, Du, H, Hu, Q, et al. Impacts of preparation technologies on biological activities of edible mushroom polysaccharides - novel insights for personalized nutrition achievement. Crit Rev Food Sci Nutr. (2025) 65:2898–920. doi: 10.1080/10408398.2024.2352796
23. Karunarathna, SC, Patabendige, NM, Kumla, J, Hapuarachchi, KK, and Suwannarach, N. The bioactive compounds, beneficial medicinal properties, and biotechnological prospects of Fomitopsis: a comprehensive overview. Front Cell Infect Microbiol. (2025) 15:1534617. doi: 10.3389/fcimb.2025.1534617
24. Farid, MS, Shafique, B, Xu, R, Łopusiewicz, Ł, and Zhao, C. Potential interventions and interactions of bioactive polyphenols and functional polysaccharides to alleviate inflammatory bowel disease - A review. Food Chem. (2025) 462:140951. doi: 10.1016/j.foodchem.2024.140951
25. Alioui, Y, Ullah, H, Ali, S, Rahman, MU, Elkharti, M, Farooqui, NA, et al. Polysaccharides derived from golden mushroom (Cantharellus cibarius Fr.) modulate gut microbiota and enhance intestinal barrier function to ameliorate dextran sulfate sodium-induced colitis in mice. Front Pharmacol. (2024) 15:1498625. doi: 10.3389/fphar.2024.1498625
26. Morales, D. Fomes fomentarius: an underexplored mushroom as source of bioactive compounds. Food Biosci. (2024) 61:104781. doi: 10.1016/j.fbio.2024.104781
27. Lim, BCC, Zeb, M, Li, WM, Tang, JZ, Heiss, C, Tackaberry, LE, et al. An immunomodulatory polysaccharide-protein complex isolated from the polypore fungus Royoporus badius. J Fungi (Basel, Switzerland). (2023) 9:87. doi: 10.3390/jof9010087
28. Gao, X, and Homayoonfal, M. Exploring the anti-cancer potential of Ganoderma lucidum polysaccharides (GLPs) and their versatile role in enhancing drug delivery systems: a multifaceted approach to combat cancer. Cancer Cell Int. (2023) 23:324. doi: 10.1186/s12935-023-03146-8
29. Xiaoying, M, Zhiming, H, Tao, Y, Jun, X, Ying, Z, Na, G, et al. Elucidating the molecular mechanisms underlying anti-inflammatory effects of Morchella esculenta in the arachidonic acid metabolic pathway by network pharmacology and molecular docking. Sci Rep. (2023) 13:15881. doi: 10.1038/s41598-023-42658-1
30. Li, S, Xiao, Y, Li, Q, Su, M, Guo, Y, and Jin, X. Recent advances in natural products derived from marine echinoderms and endophytic microbes: chemical insights and therapeutic potential. Mar Drugs. (2025) 23:33. doi: 10.3390/md23010033
31. Gupta, A, Meshram, V, Gupta, M, Goyal, S, Qureshi, KA, Jaremko, M, et al. Fungal endophytes: microfactories of novel bioactive compounds with therapeutic interventions; A comprehensive review on the biotechnological developments in the field of fungal endophytic biology over the last decade. Biomolecules. (2023) 13:1038. doi: 10.3390/biom13071038
32. Elnahas, MO, Elkhateeb, WA, and Daba, GM. Nutritive profile, pharmaceutical potentials, and structural analysis of multifunctional bioactive fungal polysaccharides-A review. Int J Biol Macromol. (2024) 266:130893. doi: 10.1016/j.ijbiomac.2024.130893
33. Zhao, J, Hu, Y, Qian, C, Hussain, M, Liu, S, Zhang, A, et al. The interaction between mushroom polysaccharides and gut microbiota and their effect on human health: A review. Biology. (2023) 12:122. doi: 10.3390/biology12010122
34. Dhasmana, A, Dobhal, P, Sati, A, Santhanam, A, Preetam, S, Malik, S, et al. Synthesis of fungal polysaccharide-based nanoemulsions for cancer treatment. RSC Adv. (2025) 15:13300–12. doi: 10.1039/d5ra01349f
35. Roszczenko, P, Szewczyk-Roszczenko, OK, Gornowicz, A, Iwańska, IA, Bielawski, K, Wujec, M, et al. The anticancer potential of edible mushrooms: a review of selected species from Roztocze, Poland. Nutrients. (2024) 16:2849. doi: 10.3390/nu16172849
36. Liu, Y, Zhang, H, Li, Y, Zha, H, Gao, Y, Chen, H, et al. Dictyophora indusiata polysaccharide mediates priming of the NLRP3 inflammasome activation via TLR4/ NF-κB signaling pathway to exert immunostimulatory effects. J Appl Biomed. (2024) 22:23–32. doi: 10.32725/jab.2024.005
37. Zhao, Y, Li, B, Liu, J, Chen, L, and Teng, H. Galangin prevents against ethanol-induced intestinal barrier dysfunction and NLRP3 inflammasome activation via NF-κB/MAPK signaling pathways in mice and Caco-2 cells. J Agric Food Chem. (2024) 72:9376–88. doi: 10.1021/acs.jafc.4c00747
38. Zhu, L, Yu, X, Ren, Y, Jin, W, Guo, Y, Zong, J, et al. Polysaccharide from Asparagus officinalis activated macrophages through NLRP3 inflammasome based on RNA-seq analysis. Biomed Pharmacother. (2024) 181:117729. doi: 10.1016/j.biopha.2024.117729
39. Li, Z, Liao, W, Yin, X, Liu, L, Zhao, Z, Lu, X, et al. Hyperoside attenuates cd-induced kidney injury via inhibiting NLRP3 inflammasome activation and ROS/MAPK/NF-κB signaling pathway in vivo and in vitro. Food Chem Toxicol. (2023) 172:113601. doi: 10.1016/j.fct.2023.113601
40. Bai, X, Rao, X, Wang, Y, Shen, H, and Jin, X. A homogeneous Lonicera japonica polysaccharide alleviates atopic dermatitis by promoting Nrf2 activation and NLRP3 inflammasome degradation via p62. J Ethnopharmacol. (2023) 309:116344. doi: 10.1016/j.jep.2023.116344
41. Motawi, TK, El-Maraghy, SA, Kamel, AS, Said, SE, and Kortam, MA. Modulation of p38 MAPK and Nrf2/HO-1/NLRP3 inflammasome signaling and pyroptosis outline the anti-neuroinflammatory and remyelinating characters of Clemastine in EAE rat model. Biochem Pharmacol. (2023) 209:115435. doi: 10.1016/j.bcp.2023.115435
42. Wang, H, Zhang, H, Miao, L, Wang, C, Teng, H, Li, X, et al. α-Amanitin induces hepatotoxicity via PPAR-γ inhibition and NLRP3 inflammasome activation. Ecotoxicol Environ Saf. (2025) 290:117749. doi: 10.1016/j.ecoenv.2025.117749
43. Zhang, XJ, Pu, YK, Yang, PY, Wang, MR, Zhang, RH, Li, XL, et al. Isolicoflavonol ameliorates acute liver injury via inhibiting NLRP3 inflammasome activation through boosting Nrf2 signaling in vitro and in vivo. Int Immunopharmacol. (2024) 143:113233. doi: 10.1016/j.intimp.2024.113233
44. Kiser, C, Gonul, CP, and Genc, S. Nrf2 activator diethyl maleate attenuates ROS mediated NLRP3 inflammasome activation in murine microglia. Cytotechnology. (2024) 76:197–208. doi: 10.1007/s10616-023-00609-8
45. Yi, X, Song, Y, Xu, J, Wang, L, Liu, L, Huang, D, et al. NLRP10 promotes AGEs-induced NLRP1 and NLRP3 inflammasome activation via ROS/MAPK/NF-κB signaling in human periodontal ligament cells. Odontology. (2024) 112:100–11. doi: 10.1007/s10266-023-00813-0
46. Wang, L-F, Wu, R-T, Yao, Y-F, Fu, W-W, Wan, M, Sang, T, et al. Cardioprotective effects of Ganoderma atrum polysaccharide in a type 2 diabetes mellitus involvement with gut-derived metabolites and NLRP3 inflammasome. J Funct Foods. (2024) 112:105991. doi: 10.1016/j.jff.2023.105991
47. Jiang, XS, Liu, T, Xia, YF, Gan, H, Ren, W, and Du, XG. Activation of the Nrf2/ARE signaling pathway ameliorates hyperlipidemia-induced renal tubular epithelial cell injury by inhibiting mtROS-mediated NLRP3 inflammasome activation. Front Immunol. (2024) 15:1342350. doi: 10.3389/fimmu.2024.1342350
48. Lin, J, Lu, YY, Shi, HY, and Lin, P. Chaga medicinal mushroom, Inonotus obliquus (Agaricomycetes), polysaccharides alleviate Photoaging by regulating Nrf2 pathway and autophagy. Int J Med Mushrooms. (2023) 25:49–64. doi: 10.1615/IntJMedMushrooms.2023049657
49. Shao, S, Li, R, Wang, K, Xia, W, Cui, B, and Li, S. Ilexchinene, a new seco-ursane triterpenoid from the leaves of Ilex chinensis with therapeutic effect on neuroinflammation by attenuating the MAPK/NF-κB signaling pathway. Phytomedicine. (2023) 121:155110. doi: 10.1016/j.phymed.2023.155110
50. Lin, HY, Lee, C-H, and Li, C-Y. Abstract 7476: obtusifolin attenuates LPS-induced inflammation and NLRP3 inflammasome activation by suppressing the MAPK/NF-κB signaling pathway and ROS production in BV-2 cells. Cancer Res. (2024) 84:7476. doi: 10.1158/1538-7445.AM2024-7476
51. Łysakowska, P, Sobota, A, and Wirkijowska, A. Medicinal mushrooms: their bioactive components, nutritional value and application in functional food production-a review. Molecules (Basel, Switzerland). (2023) 28:5393. doi: 10.3390/molecules28145393
52. Amirullah, NA, Abdullah, E, Abidin, NZ, Abdullah, N, and Manickam, S. Therapeutic potential of mushrooms: a review on NF-κB modulation in chronic inflammation. Food Biosci. (2024) 62:105059. doi: 10.1016/j.fbio.2024.105059
53. Christos, RE, Anwar, H, Lau, V, Hadinata, E, Syahputra, RA, Hardinsyah, H, et al. Harnessing nanotechnology with mushroom-derived bioactives: targeting inflammatory pathways and miRNAs in osteoarthritis. J Agric Food Res. (2025) 20:101791. doi: 10.1016/j.jafr.2025.101791
54. Li, H, Gao, J, Zhao, F, Liu, X, and Ma, B. Bioactive peptides from edible mushrooms-the preparation, mechanisms, structure-activity relationships and prospects. Foods (Basel, Switzerland). (2023) 12:2935. doi: 10.3390/foods12152935
55. Naskar, A, Dasgupta, A, Basak, G, and Acharya, K. Antioxidative and antibacterial hydro-Ethanolic fraction from an Asian edible mushroom Lentinus sajor-caju (Agaricomycetes) suppresses inflammatory responses by downregulating COX-2 and iNOS expression. Int J Medi Mushrooms. (2024) 26:1–15. doi: 10.1615/IntJMedMushrooms.2023051138
56. Ding, L, Shangguan, H, Wang, X, Liu, J, Shi, Y, Xu, X, et al. Extraction, purification, structural characterization, biological activity, mechanism of action and application of polysaccharides from Ganoderma lucidum: A review. Int J Biol Macromol. (2025) 288:138575. doi: 10.1016/j.ijbiomac.2024.138575
57. Liu, Y, Tan, D, Cui, H, and Wang, J. Ganoderic acid C2 exerts the pharmacological effects against cyclophosphamide-induced immunosuppression: a study involving molecular docking and experimental validation. Sci Rep. (2023) 13:17745. doi: 10.1038/s41598-023-44394-y
58. Peng, H, Zhong, L, Cheng, L, Chen, L, Tong, R, Shi, J, et al. Ganoderma lucidum: current advancements of characteristic components and experimental progress in anti-liver fibrosis. Front Pharmacol. (2023) 13:1094405. doi: 10.3389/fphar.2022.1094405
59. Swallah, MS, Bondzie-Quaye, P, Yu, X, Fetisoa, MR, Shao, CS, and Huang, Q. Elucidating the protective mechanism of ganoderic acid DM on breast cancer based on network pharmacology and in vitro experimental validation. Biotechnol Appl Biochem. (2025) 72:415–36. doi: 10.1002/bab.2673
60. Eira, A, Gonçalves, MBS, Fongang, YSF, Domingues, C, Jarak, I, Mascarenhas-Melo, F, et al. Unlocking the potential of Ganoderma lucidum (Curtis): botanical overview, therapeutic applications, and Nanotechnological advances. Pharmaceutics. (2025) 17:422. doi: 10.3390/pharmaceutics17040422
61. Cadar, E, Negreanu-Pirjol, T, Pascale, C, Sirbu, R, Prasacu, I, Negreanu-Pirjol, BS, et al. Natural bio-compounds from Ganoderma lucidum and their beneficial biological actions for anticancer application: A review. Antioxidants (Basel, Switzerland). (2023) 12:1907. doi: 10.3390/antiox12111907
62. Angulo-Sanchez, LT, Cruz-Félix, MC, Vidal-Gutiérrez, M, Torres-Moreno, H, Muñoz-Bernal, ÓA, Álvarez-Parrilla, E, et al. Ganoderma tuberculosum liquid culture with vineyard pruning extracts for bioactive composite production with Antiproliferative activity. Adv Pharmacol Pharmaceut Sci. (2024) 2024:5245451. doi: 10.1155/2024/5245451
63. Fu, Y, Jiang, T, Fang, X, Chen, Y, Li, J, Huang, S, et al. Integrating network pharmacology and experimental validation to explore the effect and mechanism of Inonotus obliquus polysaccharide in the treatment of rheumatoid arthritis. Pharmaceuticals (Basel, Switzerland). (2025) 18:1017. doi: 10.3390/ph18071017
64. Panossian, A. Trends and pitfalls in the Progress of network pharmacology research on natural products. Pharmaceuticals. (2025) 18:538. doi: 10.3390/ph18040538
65. Wang, Y, Gu, J, Wu, J, Xu, Y, Liu, Y, Li, F, et al. Natural products and health care functions of Inonotus obliquus. Curr Issues Mol Biol. (2025) 47:269. doi: 10.3390/cimb47040269
66. Merecz-Sadowska, A, Sadowski, A, Zielińska-Bliźniewska, H, Zajdel, K, and Zajdel, R. Network pharmacology as a tool to investigate the antioxidant and anti-inflammatory potential of plant secondary metabolites-A review and perspectives. Int J Mol Sci. (2025) 26:6678. doi: 10.3390/ijms26146678
67. Wu, J, Li, J, Sun, P, Hu, Y, and Li, Z. Theoretical framework for a polymorphic network environment. Engineering. (2024) 39:222–34. doi: 10.1016/j.eng.2024.01.018
68. Zheng, W, Lan, S, Zhang, W, Nie, B, Zhu, K, Ye, X, et al. Polysaccharide structure evaluation of Ganoderma lucidum from different regions in China based on an innovative extraction strategy. Carbohydr Polym. (2024) 335:122079. doi: 10.1016/j.carbpol.2024.122079
69. Yu, H, Choi, K, Kim, JY, and Yoo, S. Multi-level association rule mining and network pharmacology to identify the polypharmacological effects of herbal materials and compounds in traditional medicine. Brief Bioinform. (2025) 26:bbaf328. doi: 10.1093/bib/bbaf328
70. Spano, M, Goppa, L, Girometta, CE, Giusti, AM, Rossi, P, Cartabia, M, et al. Dehydrated mycelia (Cordyceps militaris, Grifola frondosa, Hericium erinaceus and Laricifomes officinalis) as novel foods: a comprehensive NMR study. LWT. (2024) 199:116123. doi: 10.1016/j.lwt.2024.116123
71. Kała, K, Cicha-Jeleń, M, Hnatyk, K, Krakowska, A, Sułkowska-Ziaja, K, Szewczyk, A, et al. Coffee with Cordyceps militaris and Hericium erinaceus fruiting bodies as a source of essential bioactive substances. Pharmaceuticals (Basel, Switzerland). (2024) 17:955. doi: 10.3390/ph17070955
72. Contato, AG, and Conte-Junior, CA. Lion's mane mushroom (Hericium erinaceus): A neuroprotective fungus with antioxidant, anti-inflammatory, and antimicrobial potential-A narrative review. Nutrients. (2025) 17:1307. doi: 10.3390/nu17081307
73. Zeng, J, Zhou, Y, Lyu, M, Huang, X, Xie, M, Huang, M, et al. Cordyceps militaris: A novel mushroom platform for metabolic engineering. Biotechnol Adv. (2024) 74:108396. doi: 10.1016/j.biotechadv.2024.108396
74. Nguyen, TQ, Van Pham, T, Andriana, Y, and Truong, MN. Cordyceps militaris-derived bioactive gels: therapeutic and anti-aging applications in dermatology. Gels (Basel, Switzerland). (2025) 11:33. doi: 10.3390/gels11010033
75. Wei, J, Li, JY, Feng, XL, Zhang, Y, Hu, X, Hui, H, et al. Unprecedented neoverrucosane and cyathane diterpenoids with anti-neuroinflammatory activity from cultures of the culinary-medicinal mushroom Hericium erinaceus. Molecules (Basel, Switzerland). (2023) 28:6380. doi: 10.3390/molecules28176380
76. Wu, N, Ge, X, Yin, X, Yang, L, Chen, L, Shao, R, et al. A review on polysaccharide biosynthesis in Cordyceps militaris. Int J Biol Macromol. (2024) 260:129336. doi: 10.1016/j.ijbiomac.2024.129336
77. Chutimanukul, P, Phatthanamas, W, Thepsilvisut, O, Chantarachot, T, Thongtip, A, and Chutimanukul, P. Commercial scale production of Yamabushitake mushroom (Hericium erinaceus (bull.) Pers. 1797) using rubber and bamboo sawdust substrates in tropical regions. Sci Rep. (2023) 13:13316. doi: 10.1038/s41598-023-40601-y
78. Ajibola, OO, Nolasco-Hipolito, C, Carvajal-Zarrabal, O, Salleh, SF, Adeyinka, GC, Adefegha, SA, et al. Turkey tail mushroom (Trametes versicolor): an edible macrofungi with immense medicinal properties. Curr Opin Food Sci. (2024) 58:101191. doi: 10.1016/j.cofs.2024.101191
79. Lowenthal, R, Taylor, M, Gidden, JA, Heflin, B, Lay, JO Jr, Avaritt, N, et al. The mycelium of the Trametes versicolor synn. Coriolus versicolor (Turkey tail mushroom) exhibit anti-melanoma activity in vitro. Biomed Pharmacother. (2023) 161:114424. doi: 10.1016/j.biopha.2023.114424
80. Desisa, B, Muleta, D, Jida, M, Dejene, T, Goshu, A, Negi, T, et al. Domestication of wild-growing Turkey tail mushroom (Trametes versicolor) from Ethiopian forests on augmented agro-industrial byproducts. Mycol Progress. (2024) 23:62. doi: 10.1007/s11557-024-01993-x
81. Mostafa, YS, Širić, I, Alamri, SAM, Alrumman, SA, Kumar, P, Abou Fayssal, S, et al. Assessment of metal elements and biochemical constituents of wild Turkey tail (Trametes versicolor) mushrooms collected from the Shivalik foothills of the Himalayas, India. Forests. (2023) 14:2247. doi: 10.3390/f14112247
82. Williams, LM, Berthon, BS, Stoodley, IL, Williams, EJ, and Wood, LG. Medicinal mushroom extracts from Hericium coralloides and Trametes versicolor exert differential immunomodulatory effects on immune cells from older adults in vitro. Nutrients. (2023) 15:2227. doi: 10.3390/nu15092227
83. Teymoorian, SK, Nouri, H, and Moghimi, H. In-vivo and in-vitro wound healing and tissue repair effect of Trametes versicolor polysaccharide extract. Sci Rep. (2024) 14:3796. doi: 10.1038/s41598-024-54565-0
84. Yang, Y, and Hu, X. A chromosome-scale genome of Trametes versicolor and transcriptome-based screening for light-induced genes that promote triterpene biosynthesis. J Fungi (Basel, Switzerland). (2025) 11:81. doi: 10.3390/jof11010081
85. Tulasi, B, Kaithamalai, B, Angappan, S, Gurudevan, T, Padmanaban, G, Chellamuthu, S, et al. Standardization of an analytical technique for determination of pesticide residues in fresh and processed button mushroom Agaricus bisporus (Lange) Imbach. Sci Rep. (2024) 14:30747. doi: 10.1038/s41598-024-80690-x
86. Calleja-Gómez, M, Roig, P, Rimac Brnčić, S, Barba, FJ, and Castagnini, JM. Scanning electron microscopy and triple TOF-LC-MS-MS analysis of polyphenols from PEF-treated edible mushrooms (L. edodes, A. brunnescens, and P. ostreatus). Antioxidants. (2023) 12:2080. doi: 10.3390/antiox12122080
87. Goff, R, Smith, M, Islam, S, Sisley, S, Ferguson, J, Kuzdzal, S, et al. Determination of psilocybin and psilocin content in multiple Psilocybe cubensis mushroom strains using liquid chromatography - tandem mass spectrometry. Anal Chim Acta. (2024) 1288:342161. doi: 10.1016/j.aca.2023.342161
88. Helmy, MI, Nessim, CK, and El Hamd, MA. A green LC-MS/MS for energetically assessing an imperative muscle relaxant combination: a clinical pharmacokinetic study in human plasma. Microchem J. (2024) 202:110848. doi: 10.1016/j.microc.2024.110848
89. Windsor, C, Kreynes, AE, Chilton, JS, Chioffi, WA, Krishnamurthy, A, and Ishii, M. Comparative study of Chaga (Inonotus obliquus) dietary supplements using complementary analytical techniques. Int J Mol Sci. (2025) 26:2970. doi: 10.3390/ijms26072970
90. Nguyen, KD, Nguyen, CM, Le, DA, Huynh, HT, Tran, MT, Truong, ATN, et al. The mixture of Ganoderma lucidum and Cordyceps militaris: chemical composition and protective effect against oxidative stress. J Agric Food Res. (2024) 15:101045. doi: 10.1016/j.jafr.2024.101045
91. Ahmad, MF, A Alsayegh, A, Ahmad, FA, Akhtar, MS, Alavudeen, SS, Bantun, F, et al. Ganoderma lucidum: insight into antimicrobial and antioxidant properties with development of secondary metabolites. Heliyon. (2024) 10:e25607. doi: 10.1016/j.heliyon.2024.e25607
92. Swallah, MS, Bondzie-Quaye, P, Wu, Y, Acheampong, A, Sossah, FL, Elsherbiny, SM, et al. Therapeutic potential and nutritional significance of Ganoderma lucidum - a comprehensive review from 2010 to 2022. Food Funct. (2023) 14:1812–38. doi: 10.1039/d2fo01683d
93. Plosca, MP, Chiș, MS, Fărcaș, AC, and Păucean, A. Ganoderma lucidum-from ancient remedies to modern applications: chemistry, benefits, and safety. Antioxidants (Basel, Switzerland). (2025) 14:513. doi: 10.3390/antiox14050513
94. Fang, H, Yang, S, and Yang, T. Ganoderma lucidum polysaccharides: a comprehensive overview of pharmacological effects and future perspectives. Food Biosci. (2025) 64:105990. doi: 10.1016/j.fbio.2025.105990
95. Ekiz, E, Oz, E, Abd El-Aty, AM, Proestos, C, Brennan, C, Zeng, M, et al. Exploring the potential medicinal benefits of Ganoderma lucidum: from metabolic disorders to coronavirus infections. Foods (Basel, Switzerland). (2023) 12:1512. doi: 10.3390/foods12071512
96. Chen, L, Qiu, R, Wang, B, Liu, J, Li, X, Hou, Z, et al. Investigating the association between inflammation mediated by mushroom consumption and mild cognitive impairment in Chinese older adults. Food Funct. (2024) 15:5343–51. doi: 10.1039/d3fo04263d
97. Al-Hunaiti, A, Zihlif, M, Abu Thiab, T, Al-Awaida, W, Al-Ameer, HJ, and Imraish, A. Magnetic nanoparticle-based combination therapy: synthesis and in vitro proof of concept of CrFe2O4- rosmarinic acid nanoparticles for anti-inflammatory and antioxidant therapy. PLoS One. (2024) 19:e0297716. doi: 10.1371/journal.pone.0297716
98. Li, C, Zhang, R, Tian, B, Liu, B, Mei, Y, and Li, W. A review of the extraction technologies, structural characterization, chemical modification, and pharmacological effects of Ganoderma lucidum polysaccharides. Naunyn Schmiedeberg's Arch Pharmacol. (2025). doi: 10.1007/s00210-025-04436-w
99. Lv, R, Wang, Y, Chen, D, Bao, Z, and Lin, S. Protection of Tricholoma matsutake and its bioactive components against cognitive impairment: modulating oxidative stress, alleviating neuroinflammation, and preserving synaptic plasticity. J Agric Food Chem. (2025) 73:14393–407. doi: 10.1021/acs.jafc.5c04473
100. Almurshedi, AS, El-Masry, TA, Selim, H, El-Sheekh, MM, Makhlof, MEM, Aldosari, BN, et al. New investigation of anti-inflammatory activity of Polycladia crinita and biosynthesized selenium nanoparticles: isolation and characterization. Microb Cell Factories. (2023) 22:173. doi: 10.1186/s12934-023-02168-1
101. Scialabba, C, Craparo, EF, Cabibbo, M, Emanuele Drago, S, and Cavallaro, G. Exploiting inhalable microparticles incorporating hybrid polymer-lipid nanoparticles loaded with Iloprost manages lung hyper-inflammation. Int J Pharm. (2024) 666:124813. doi: 10.1016/j.ijpharm.2024.124813
102. Luo, S, Cai, J, Yin, F, Lu, L, Liu, Z, Wang, Y, et al. M3-DPPE liposomal nanoparticles encapsulating CLEC12A enhance CD206-mediated endocytosis and efficacy in the collagen-induced arthritis model. ACS Appl Bio Mater. (2025) 8:1002–16. doi: 10.1021/acsabm.4c01139
103. Duncan, JBW, Basu, S, and Vivekanand, P. Honey gold nanoparticles attenuate the secretion of IL-6 by LPS-activated macrophages. PLoS One. (2023) 18:e0291076. doi: 10.1371/journal.pone.0291076
104. Sales-Campos, H, Desidério, CS, Trevisan, RO, Timóteo, RP, Flávio-Reis, VHP, Pessoa-Gonçalves, YM, et al. Anti-IL-4, anti-IL-17, and anti-IFN-gamma activity in the saliva of Amblyomma sculptum ticks. Int J Mol Sci. (2025) 26:4734. doi: 10.3390/ijms26104734
105. Guo, K, Zeng, X, Liu, X, He, P, Zhang, Z, Yang, Q, et al. Lifestyle deterioration linked to elevated inflammatory cytokines over a two-month follow-up. Sci Rep. (2024) 14:21381. doi: 10.1038/s41598-024-69967-3
106. Petrović, J, Glamočlija, J, Milinčić, DD, Doroški, A, Lević, S, Stanojević, SP, et al. Comparative chemical analysis and bioactive properties of aqueous and glucan-rich extracts of three widely appreciated mushrooms: Agaricus bisporus (J.E.Lange) Imbach, Laetiporus sulphureus (bull.) Murill and Agrocybe aegerita (V. Brig.) Vizzini. Pharmaceuticals (Basel). (2024) 17:1153. doi: 10.3390/ph17091153
107. Ji, T, and Li, H. T-helper cells and their cytokines in pathogenesis and treatment of asthma. Front Immunol. (2023) 14:1149203. doi: 10.3389/fimmu.2023.1149203
108. Bai, Y, Liang, F, Yang, Y, Guan, L, and Ma, H. Polysaccharides from edible fungi spent mushroom substrates: a review of their extraction, purification, structural characteristics, and biological activities. Int J Biol Macromol. (2025) 330:147925. doi: 10.1016/j.ijbiomac.2025.147925
109. Kour, H, Kour, D, Kour, S, Singh, S, Hashmi, SAJ, Yadav, AN, et al. Bioactive compounds from mushrooms: emerging bioresources of food and nutraceuticals. Food Biosci. (2022) 50:102124. doi: 10.1016/j.fbio.2022.102124
110. Ahmed, WE, Naeem, M, Siddiqui, MK, and Fiidow, MA. Predictive modeling of ADME properties using M-polynomial based topological indices for biocompatible polysaccharides. Sci Rep. (2025) 15:29667. doi: 10.1038/s41598-025-14134-5
111. Pasdaran, A, Grice, ID, and Hamedi, A. A review of natural products and small-molecule therapeutics acting on central nervous system malignancies: approaches for drug development, targeting pathways, clinical trials, and challenges. Drug Dev Res. (2024) 85:e22180. doi: 10.1002/ddr.22180
112. Kim, T, Lee, D, Lee, JH, Lee, YS, Oh, BJ, Lim, KS, et al. Predictors of poor outcomes in patients with wild mushroom-induced acute liver injury. World J Gastroenterol. (2017) 23:1262–7. doi: 10.3748/wjg.v23.i7.1262
113. Ravi, N, Tye, GJ, Dhaliwal, SS, Musa, MY, Wong, MTJ, and Lai, NS. Immune profiling in oncology: bridging the gap between technology and treatment. Med Oncol (Northwood, London, England). (2025) 42:446. doi: 10.1007/s12032-025-03002-x
114. Benavent, D, Iniesta-Chamorro, JM, Novella-Navarro, M, Pérez-Martínez, M, Martínez-Sánchez, N, Kaffati, M, et al. Digital health intervention for patient monitoring in immune-mediated inflammatory diseases: cocreation and feasibility study of the IMIDoc platform. JMIR Hum Factors. (2025) 12:e58095. doi: 10.2196/58095
115. Travaglia, A, Lal, S, and Pullagura, SR. Advancing ALS research: public-private partnerships to accelerate drug and biomarker development. Trends Neurosci. (2025) 48:1–2. doi: 10.1016/j.tins.2024.10.008
116. Desai, N, Rana, D, Patel, M, Bajwa, N, Prasad, R, and Vora, LK. Nanoparticle therapeutics in clinical perspective: classification, marketed products, and regulatory landscape. Small (Weinheim an der Bergstrasse, Germany). (2025) 21:e2502315. doi: 10.1002/smll.202502315
117. Liang, X, Niu, P, Li, J, Guan, X, Zhang, Y, and Li, J. Discovery of anti-inflammatory triterpenoid glucosides from the Heritiera littoralis Dryand. Molecules (Basel, Switzerland). (2023) 28:1658. doi: 10.3390/molecules28041658
118. Ren, C, Liang, X, Pi, R, Xin, J, Yang, B, Zheng, Q, et al. New triterpenoids from the leaves of Heritiera littoralis and their anti-inflammatory activity. Molecules (Basel, Switzerland). (2024) 30:131. doi: 10.3390/molecules30010131
119. Chang, SS, Huang, HT, Wei, WC, Lo, IW, Lin, YC, Chao, CH, et al. Anti-inflammatory effect of euphane- and tirucallane-type triterpenes isolated from the traditional herb Euphorbia neriifolia L. Front Chem. (2023) 11:1223335. doi: 10.3389/fchem.2023.1223335
120. Teng, L, Wang, C, Cui, B, Zhang, J, Zhou, S, Pan, X, et al. Lanostane triterpenoids from mycelia-associated Ganoderma sinense and their anti-inflammatory activity. Phytochemistry. (2023) 215:113870. doi: 10.1016/j.phytochem.2023.113870
121. Woźniak, Ł, Szakiel, A, Głowacka, A, Rozpara, E, Marszałek, K, and Skąpska, S. Triterpenoids of three apple cultivars-biosynthesis, Antioxidative and anti-inflammatory properties, and fate during processing. Molecules (Basel, Switzerland). (2023) 28:2584. doi: 10.3390/molecules28062584
122. Xu, Q, Liu, J, Wang, X, Jiao, Y, Zhang, Y, and Shang, X. Characterization, target isolation of triterpenes in the anti-inflammatory fraction of Salvia rosmarinus via UPLC-Orbitrap MS/MS coupled with GNPS. J Agric Food Chem. (2025) 73:10985–97. doi: 10.1021/acs.jafc.4c12650
123. Łyko, L, Olech, M, Gawlik, U, Krajewska, A, Kalemba, D, Tyśkiewicz, K, et al. Rhododendron luteum sweet flower supercritical CO2 extracts: terpenes composition, pro-inflammatory enzymes inhibition and antioxidant activity. Int J Mol Sci. (2024) 25:9952. doi: 10.3390/ijms25189952
124. Stamou, P, Gianniou, DD, Trougakos, IP, Mitakou, S, Halabalaki, M, Kostakis, IK, et al. Anti-inflammatory activity of the major Triterpenic acids of Chios mastic gum and their semi-synthetic analogues. Biomolecules. (2024) 14:1618. doi: 10.3390/biom14121618
125. Yang, L, Hu, Y, Deng, H, Li, Y, Zhang, R, Zhang, Q, et al. Water-soluble polysaccharides from Torreya grandis nuts: structural characterization and anti-inflammatory activity. Int J Biol Macromol. (2025) 291:138935. doi: 10.1016/j.ijbiomac.2024.138935
126. Apaza Ticona, L, Sánchez Sánchez-Corral, J, Montoto Lozano, N, Prieto Ramos, P, and Sánchez, ÁR. Study of Pentacyclic triterpenes from lyophilised Aguaje: anti-inflammatory and antioxidant properties. Int J Mol Sci. (2024) 25:9615. doi: 10.3390/ijms25179615
127. Zhang, DL, Wang, Y, Liu, JB, Chen, Q, Li, SY, Jin, DJ, et al. Dichapetalin-type triterpenoids from Dichapetalum longipetalum and their anti-inflammatory activity. Phytochemistry. (2024) 217:113900. doi: 10.1016/j.phytochem.2023.113900
128. Tang, Y, Zhou, M, Mao, Z, Zhu, B, Zhou, F, Ye, X, et al. Structure of a polysaccharide MDP2-1 from Melastoma dodecandrum Lour. And its anti-inflammatory effects. Int J Biol Macromol. (2024) 265:131015. doi: 10.1016/j.ijbiomac.2024.131015
129. Qi, S, Meng, X, Cui, B, Liu, T, Yang, L, Cai, G, et al. Drimane-type sesquiterpenoids and triterpenoids from the whole plant of Limonium sinense with their antiproliferative and anti-inflammatory activities. RSC Adv. (2025) 15:1220–9. doi: 10.1039/d4ra06721e
130. Li, X, Zhang, J, Chen, Q, Tang, P, Zhang, T, Feng, Q, et al. Diversity-oriented synthesis of diterpenoid alkaloids yields a potent anti-inflammatory agent. Phytomedicine. (2023) 117:154907. doi: 10.1016/j.phymed.2023.154907
131. Zhang, X, Wu, R, Yan, Y, Luo, Y, Wan, Z, Liu, Z, et al. Discovery of novel 8-hydroxyquinoline derivatives as NLRP3 inflammasome inhibitors with therapeutic potential for inflammatory bowel disease. Eur J Med Chem. (2025) 298:118023. doi: 10.1016/j.ejmech.2025.118023
132. Pei, X, Zhang, Z, Wang, N, Huang, G, Min, X, Yang, Y, et al. Onychiol B attenuates lipopolysaccharide-induced inflammation via MAPK/NF-κB pathways and acute lung injury in vivo. Bioorg Chem. (2023) 132:106351. doi: 10.1016/j.bioorg.2023.106351
133. Gholizadeh, M, Khalili, A, Roodi, PB, Saeedy, SAG, Najafi, S, Keshavarz Mohammadian, M, et al. Selenium supplementation decreases CRP and IL-6 and increases TNF-alpha: A systematic review and meta-analysis of randomized controlled trials. J Trace Elements Med Biol. (2023) 79:127199. doi: 10.1016/j.jtemb.2023.127199
134. Berlana, D, Albertos, R, Barquin, R, Pau-Parra, A, Díez-Poch, M, López-Martínez, R, et al. Impact of Omega-3 fatty acid supplementation in parenteral nutrition on inflammatory markers and clinical outcomes in critically ill COVID-19 patients: A randomized controlled trial. Nutrients. (2024) 16:3046. doi: 10.3390/nu16183046
135. Li, D, Zhong, J, Zhang, Q, and Zhang, J. Effects of anti-inflammatory therapies on glycemic control in type 2 diabetes mellitus. Front Immunol. (2023) 14:1125116. doi: 10.3389/fimmu.2023.1125116
136. Aslani, MR, Abdollahi, N, Matin, S, Zakeri, A, and Ghobadi, H. Effect of crocin of Crocus sativus L. on serum inflammatory markers (IL-6 and TNF-α) in chronic obstructive pulmonary disease patients: a randomised, double-blind, placebo-controlled trial. Br J Nutr. (2023) 130:446–53. doi: 10.1017/S0007114522003397
137. Vahedi-Mazdabadi, Y, Shahinfar, H, Toushih, M, and Shidfar, F. Effects of berberine and barberry on selected inflammatory biomarkers in adults: a systematic review and dose-response meta-analysis of randomized clinical trials. Phytother Res. (2023) 37:5541–57. doi: 10.1002/ptr.7998
138. Kavyani, Z, Musazadeh, V, Golpour-Hamedani, S, Moridpour, AH, Vajdi, M, and Askari, G. The effect of Nigella sativa (black seed) on biomarkers of inflammation and oxidative stress: an updated systematic review and meta-analysis of randomized controlled trials. Inflammopharmacology. (2023) 31:1149–65. doi: 10.1007/s10787-023-01213-0
139. Rastgoo, S, Fateh, ST, Nikbaf-Shandiz, M, Rasaei, N, Aali, Y, Zamani, M, et al. The effects of L-carnitine supplementation on inflammatory and anti-inflammatory markers in adults: a systematic review and dose-response meta-analysis. Inflammopharmacology. (2023) 31:2173–99. doi: 10.1007/s10787-023-01323-9
140. Dong, QQ, Wu, Q, Lu, Y, Shi, Y, Yang, KD, Xu, XL, et al. Exploring β-glucan as a micro-nano system for oral delivery targeted the colon. Int J Biol Macromol. (2023) 253:127360. doi: 10.1016/j.ijbiomac.2023.127360
141. Chen, C, Chen, X, Mo, Q, Liu, J, Yao, X, Di, X, et al. Cytochrome P450 metabolism studies of [6]-gingerol, [8]-gingerol, and [10]-gingerol by liver microsomes of humans and different species combined with expressed CYP enzymes. RSC Adv. (2023) 13:5804–12. doi: 10.1039/d2ra06184h
142. Li, D, Lin, Y, Lv, X, Wu, Y, Han, C, Cao, P, et al. Triterpenoids from Ganoderma lucidum inhibit cytochrome P450 enzymes interfering with the metabolic process of specific clinical drugs. Front Pharmacol. (2024) 15:1485209. doi: 10.3389/fphar.2024.1485209
143. Hossam Abdelmonem, B, Abdelaal, NM, Anwer, EKE, Rashwan, AA, Hussein, MA, Ahmed, YF, et al. Decoding the role of CYP450 enzymes in metabolism and disease: A comprehensive review. Biomedicine. (2024) 12:1467. doi: 10.3390/biomedicines12071467
144. Liu, Z, Pang, J, Li, Y, Wei, D, Yang, J, Wang, X, et al. Catalytic selectivity and evolution of cytochrome P450 enzymes involved in monoterpene indole alkaloids biosynthesis. Physiol Plant. (2024) 176:e14515. doi: 10.1111/ppl.14515
145. Liu, J, and Zhang, D. Cytochrome P450-mediated carbon-carbon bond formation in drug metabolism. Drug Metab Rev. (2025) 57:51–66. doi: 10.1080/03602532.2025.2451847
146. Guengerich, FP. Roles of individual human cytochrome P450 enzymes in drug metabolism. Pharmacol Rev. (2024) 76:1104–32. doi: 10.1124/pharmrev.124.001173
147. Wunderle, C, Urbach, K, Buchmueller, L, Randegger, S, Kaegi-Braun, N, Laviano, A, et al. Biomarkers for individualized nutritional therapy in disease-related malnutrition: a narrative review. Am J Clin Nutr. (2025) 122:671–9. doi: 10.1016/j.ajcnut.2025.07.009
148. Poniedziałek, B, Siwulski, M, Wiater, A, Komaniecka, I, Komosa, A, Gąsecka, M, et al. The effect of mushroom extracts on human platelet and blood coagulation: in vitro screening of eight edible species. Nutrients. (2019) 11:3040. doi: 10.3390/nu11123040
149. Michalska, A, Sierocka, M, Drzewiecka, B, and Świeca, M. Antioxidant and anti-inflammatory properties of mushroom-based food additives and food fortified with them-current status and future perspectives. Antioxidants (Basel, Switzerland). (2025) 14:519. doi: 10.3390/antiox14050519
150. Fagunwa, O, Davies, K, and Bradbury, J. The human gut and dietary salt: the Bacteroides/Prevotella ratio as a potential marker of sodium intake and beyond. Nutrients. (2024) 16:942. doi: 10.3390/nu16070942
151. Karačić, A, Renko, I, Krznarić, Ž, Klobučar, S, and Liberati Pršo, AM. The association between the Firmicutes/Bacteroidetes ratio and body mass among European population with the highest proportion of adults with obesity: an observational follow-up study from Croatia. Biomedicine. (2024) 12:2263. doi: 10.3390/biomedicines12102263
152. Milhorini, SDS, Zavadinack, M, Santos, JFD, Lara, EL, Smiderle, FR, and Iacomini, M. Structural variety of glucans from Ganoderma lucidum fruiting bodies. Carbohydr Res. (2024) 538:109099. doi: 10.1016/j.carres.2024.109099
153. Chen, SN, Nan, FH, Liu, MW, Yang, MF, Chang, YC, and Chen, S. Evaluation of immune modulation by β-1,3; 1,6 D-glucan derived from Ganoderma lucidum in healthy adult volunteers, A randomized controlled trial. Foods (Basel, Switzerland). (2023) 12:659. doi: 10.3390/foods12030659
154. Sarkar, N, Mahajan, AA, Pathak, S, Seth, P, Chowdhury, A, Ghose, I, et al. Beta-glucans in biotechnology: A holistic review with a special focus on yeast. Bioengineering (Basel, Switzerland). (2025) 12:365. doi: 10.3390/bioengineering12040365
155. Tokul-Ölmez, Ö, Kaplaner, E, and Öztürk, M. Impact of collection locations and host trees on the bioactive triterpene composition and antioxidant activity of four Ganoderma species: A Chemometric analysis. Chem Biodivers. (2025) 22:e202500206. doi: 10.1002/cbdv.202500206
156. Ota, T, Saburi, W, Komba, S, and Mori, H. Chemical synthesis of oligosaccharide derivatives with partial structure of β1-3/1-6 glucan, using monomeric units for the formation of β1-3 and β1-6 glucosidic linkages. Biosci Biotechnol Biochem. (2023) 87:1111–21. doi: 10.1093/bbb/zbad093
157. Al-Sahlany, STG, Al-Kaabi, WJ, Al-Manhel, AJA, Niamah, AK, Altemimi, AB, Al-Wafi, H, et al. Effects of β-glucan extracted from Saccharomyces cerevisiae on the quality of bio-yoghurts: in vitro and in vivo evaluation. Food Measure. (2022) 16:3607–17. doi: 10.1007/s11694-022-01468-1
158. Adachi, Y, Momose, F, Momose, H, Tada, R, and Ohno, N. Potentiation of antitumor activity by antibody drugs and mushroom-derived β-glucans in natural killer cell-mediated tumoricidal activities against non-Hodgkin's B-cell lymphoma. Int J Med Mushrooms. (2023) 25:1–19. doi: 10.1615/IntJMedMushrooms.2022047219
159. Leisegang, K, Opuwari, CS, Moichela, F, and Finelli, R. Traditional, complementary and alternative medicines in the treatment of ejaculatory disorders: A systematic review. Medicina (Kaunas). (2023) 59:1607. doi: 10.3390/medicina59091607
160. Zhao, P, Guan, M, Tang, W, Walayat, N, Ding, Y, and Liu, J. Structural diversity, fermentation production, bioactivities and applications of triterpenoids from several common medicinal fungi: recent advances and future perspectives. Fitoterapia. (2023) 166:105470. doi: 10.1016/j.fitote.2023.105470
161. Chugh, RM, Mittal, P, Mp, N, Arora, T, Bhattacharya, T, Chopra, H, et al. Fungal mushrooms: A natural compound with therapeutic applications. Front Pharmacol. (2022) 13:925387. doi: 10.3389/fphar.2022.925387
162. Xie, J, Lin, D, Li, J, Zhou, T, Lin, S, and Lin, Z. Effects of Ganoderma lucidum polysaccharide peptide ameliorating cyclophosphamide-induced immune dysfunctions based on metabolomics analysis. Front Nutr. (2023) 10:1179749. doi: 10.3389/fnut.2023.1179749
163. Liu, Z, Jiang, Y, Fan, Q, Li, S, and Wang, Y. The role of butyric acid and microorganisms in chronic inflammatory diseases and microbiome-based therapeutics. J Inflamm Res. (2025) 18:13465–87. doi: 10.2147/JIR.S540163
164. Zhong, J, Mareque-Rivas, JC, Lan, X, and Su, Y-X. Supramolecular assembly of triterpenoids: current state and biomedical perspectives. Aggregate. (2025) 6:e70081. doi: 10.1002/agt2.70081
165. Wainwright, CL, Teixeira, MM, Adelson, DL, Buenz, EJ, David, B, Glaser, KB, et al. Future directions for the discovery of natural product-derived immunomodulating drugs: an IUPHAR positional review. Pharmacol Res. (2022) 177:106076. doi: 10.1016/j.phrs.2022.106076
166. Vetter, J. The Mushroom Glucans: Molecules of High Biological and Medicinal Importance. Foods. (2023) 12:1009. doi: 10.3390/foods12051009
167. Wang, H, Yang, Y, Wang, S, Li, C, Chen, C, Wan, X, et al. Polysaccharides of Floccularia luteovirens Alleviate Oxidative Damage and Inflammatory Parameters of Diabetic Nephropathy in db/db Mice. Front Biosci (Landmark Ed). (2023) 28:82. doi: 10.31083/j.fbl2804082
168. Yu, D, Cai, X, Wang, S, Li, Y, Du, Y, Wang, ZA, et al. Structural Characterization and Immunological Activity of Polysaccharide Degradation Products from Phlebopus portentosus. Separations. (2024) 11:105. doi: 10.3390/separations11040105
169. Chung, J, Im, SY, Park, SK, Heo, DB, Sung, HWJ, Ohm, D, et al. Inotodiol Attenuates Mucosal Inflammation in a Mouse Model of Eosinophilic Chronic Rhinosinusitis. Allergy Asthma Immunol Res. (2025) 17:77–93. doi: 10.4168/aair.2025.17.1.77
170. Liu, YS, Lai, MC, Tzeng, YC, and Liu, IM. Polyphenolic Hispolon Derived from Medicinal Mushrooms of the Inonotus and Phellinus Genera Promotes Wound Healing in Hyperglycemia-Induced Impairments. Nutrients. (2025) 17:266. doi: 10.3390/nu17020266
171. Paul, N. Significant role of bioactive compounds of ganoderma lucidum in the aging process. Open Access Research Journal of Biology and Pharmacy (2025) 14:35–44. doi: 10.53022/oarjbp.2025.14.2.0033
172. Areesanan, A, Wasilewicz, A, Nicolay, S, Grienke, U, Zimmermann-Klemd, AM, Rollinger, JM, et al. Evaluation of in vitro pharmacological activities of medicinal mushrooms in the context of dry eye disease. Front Pharmacol. (2025) 16:1557359. doi: 10.3389/fphar.2025.1557359
173. Rijia, A, Krishnamoorthi, R, Rasmi, M, Mahalingam, PU, and Kim, KS. Comprehensive Analysis of Bioactive Compounds in Wild Ganoderma applanatum Mushroom from Kerala, South India: Insights into Dietary Nutritional, Mineral, Antimicrobial, and Antioxidant Activities. Pharmaceuticals (Basel). (2024) 17:509. doi: 10.3390/ph17040509
174. Tan, HX, Wang, YT, Shen, RL, and Jiang, M. Mechanistic Study of Chaga Medicinal Mushroom Inonotus obliquus (Agaricomycetes) Phenolic Compounds in the Treatment of Liver Cancer: A Database and Simulation Approach. Int J Med Mushrooms. (2025) 27:17–30. doi: 10.1615/IntJMedMushrooms.2025058720
175. Han, Q, Li, H, Zhao, F, Gao, J, Liu, X, and Ma, B. Auricularia auricula Peptides Nutritional Supplementation Delays H2O2-Induced Senescence of HepG2 Cells by Modulation of MAPK/NF-κB Signaling Pathways. Nutrients. (2023) 15:3731. doi: 10.3390/nu15173731
176. Zhao, S, Gao, Y, Wang, H, Fan, Y, Wang, P, Zhao, W, et al. A novel mushroom (Auricularia polytricha) glycoprotein protects against lead-induced hepatoxicity, promotes lead adsorption, inhibits organ accumulation of lead, upregulates detoxifying proteins, and enhances immunoregulation in rats. Front Nutr. (2023) 10:1144346. doi: 10.3389/fnut.2023.1144346
177. Ji, L, Tan, L, Shang, Z, Li, W, Mo, X, Yang, S, et al. Discovery of New Antimicrobial Metabolites in the Coculture of Medicinal Mushrooms. J Agric Food Chem. (2024) 72:5247–57. doi: 10.1021/acs.jafc.3c0947
178. Ma, X, Huo, Z, Shi, M, Wang, H, Yang, T, Xiao, J, et al. Uncovering active ingredients and mechanisms of Pholiota adiposa in the treatment of Alzheimer’s disease based on network pharmacology and bioinformatics. Sci Rep. (2025) 15:27981. doi: 10.1038/s41598-025-30392-1
Glossary
AAPs - Auricularia auricula peptides
AOME - Armillaria ostoyae mycelial metabolites
APL - Auricularia polytricha glycoprotein
ARE - Antioxidant response element
ASC - Apoptosis-associated speck-like protein containing a CARD
CR3 - Complement receptor 3
CTL - Cytotoxic T lymphocyte
DAMP - Damage-associated molecular pattern
EGFR - Epidermal growth factor receptor
ER - Endoplasmic reticulum
ERK - Extracellular signal-regulated kinase
FLPs - Floccularia luteovirens polysaccharides
GALT - Gut-associated lymphoid tissue
GMP - Good manufacturing practice
GSDMD - Gasdermin D
HCMP - High-molecular-weight polysaccharide from Cordyceps militaris
HO-1 - Heme oxygenase-1
IFN-γ - Interferon-gamma
IKK - IκB kinase
IL - Interleukin
IOP - Inonotus obliquus polysaccharides
IκB - Inhibitor of κB
JAK - Janus kinase
JNK - c-Jun N-terminal kinase
Keap1 - Kelch-like ECH-associated protein 1
MAPK - Mitogen-activated protein kinase
NF-κB - Nuclear factor-kappa B
NK - Natural killer
NLRP3 - NOD-like receptor family pyrin domain-containing 3
NO - Nitric oxide
Nrf2 - Nuclear factor erythroid 2-related factor 2
NSAIDs - Non-steroidal anti-inflammatory drugs
NKT - Natural killer T cell
PAMP - Pathogen-associated molecular pattern
PGE2 - Prostaglandin E2
PI3K - Phosphatidylinositol 3-kinase
PPRP - Phlebopus portentosus polysaccharide
PRR - Pattern recognition receptor
RGLS - Sporoderm-removed Ganoderma lucidum spores
ROS - Reactive oxygen species
SAR - Structure–activity relationship
SASP - Senescence-associated secretory phenotype
SCFA - Short-chain fatty acid
STAT - Signal transducer and activator of transcription
TGF-β - Transforming growth factor-beta
Th - T helper cell
TLR - Toll-like receptor
TNF-α - Tumor necrosis factor-alpha
Treg - Regulatory T cell
Keywords: medicinal fungi, immunomodulation, chronic inflammation, β-glucans, precision medicine
Citation: Xiaoying M, Peng Z, Hong W, Na G, Jun X, Ying Z, Xun C and Guoli L (2025) From functional foods to immunotherapeutic agents: mechanistic insights into medicinal mushroom bioactives in chronic inflammation management. Front. Nutr. 12:1725297. doi: 10.3389/fnut.2025.1725297
Edited by:
Muthukumar Serva Peddha, Central Food Technological Research Institute (CSIR), IndiaReviewed by:
R. Arivuchudar, Periyar University, IndiaVallamkondu Manasa, Narensnosh Private Limited, India
Copyright © 2025 Xiaoying, Peng, Hong, Na, Jun, Ying, Xun and Guoli. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ma Xiaoying, bXh5ODUxMDAxQDEyNi5jb20=