 Niraj Chaudhary1
Niraj Chaudhary1 Luis G. Villa-Diaz1,2*
Luis G. Villa-Diaz1,2*- 1Department of Biological Sciences, Oakland University, Rochester, MI, United States
- 2Department of Bioengineering, Oakland University, Rochester, MI, United States
The advancement of human pluripotent stem cell (hPSC) culture systems has revolutionized the landscape of preclinical drug discovery and toxicological evaluation. Progressing innovations from feeder-dependent and xenogeneic matrices to chemically defined, xeno-free, and fully synthetic platforms have addressed long-standing challenges in reproducibility, safety, and clinical translation. Developments in recombinant extracellular matrix proteins, synthetic peptide substrates, and polymer-based coatings have enabled the generation of Good Manufacturing Practice (GMP)-compliant, scalable hPSC cultures while minimizing biological variability and immunogenic risks. Integration of automation, artificial intelligence (AI), and three-dimensional (3D) bioprocessing technologies aims at further enhancement of standardization, quality control, and throughput. In the context of pharmaceutical research, hPSC-derived cellular models now underpin high-throughput drug screening and mechanistic toxicological assays, offering superior human relevance compared to traditional animal models. Despite these advances, barriers such as cellular immaturity, inter-batch variability, and limited regulatory acceptance persist, underscoring the need for further protocol standardization and technological refinement. This review provides a comprehensive analysis of current animal-free hPSC culture platforms, critically examines their strengths and limitations, and discusses future directions for advancing their application in drug discovery and predictive toxicology. The ongoing evolution of hPSC technologies promises to accelerate the development of safer, more effective therapeutic agents and to reshape the future of human disease modeling and pharmacological research.
1 Introduction: from the challenges to the potential of hPSC culture
1.1 hPSCs in the era of translational biomedicine
Human pluripotent stem cells (hPSCs) —including human embryonic stem cells (hESCs) derived from pre-implantation embryos (Thomson et al., 1998) and human induced pluripotent stem cells (hiPSCs) reprogrammed from somatic cells (Takahashi et al., 2007) are characterized by two fundamental capabilities: the ability to self-renew indefinitely in vitro while preserving pluripotency and the capacity to differentiate into virtually all specialized cell types of the human body, encompassing approximately 400 distinct lineages (Hatton et al., 2025). These attributes render hPSCs as essential tools for applications in regenerative medicine, drug development, disease modeling, and studies of human embryogenesis. To fully harness their therapeutic and research potential, it is critical to maintain hPSCs under defined culture conditions that support self-renewal and minimize spontaneous differentiation with minimal or no exposure to animal-derived products (Villa-Diaz et al., 2013).
Early hPSC culture methods utilized mouse embryonic fibroblast (MEF) feeder layers and fetal bovine serum-based media (Thomson et al., 1998). Subsequent advancements enabled feeder-free systems, incorporating Matrigel (Xu et al., 2001) and defined media such as mTeSR1 (Ludwig et al., 2006a). New developments brought in recombinant proteins such as vitronectin (Braam et al., 2008) and laminin-511/521 (Miyazaki et al., 2008), synthetic peptides such as Synthemax (Melkoumian et al., 2010) and StemAdhere (Nagaoka et al., 2010), and polymers like PMEDSAH (Villa-Diaz et al., 2010), advancing the development of fully synthetic substrates and three-dimensional culture systems optimized for large-scale expansion. Despite improvements, challenges remain, notably due to the risk of xenogeneic contamination and batch-to-batch variability associated with Matrigel and the substantial cost of defined culture components (Zhou et al., 2022).
Recent advances have focused on developing chemically defined, xeno-free culture systems to enable safe and efficient expansion of hPSCs. These defined conditions aim to eliminate variability and ensure scalability for large-scale production, crucial for clinical applications and research advancements. GMP for hPSCs ideally requires xeno-free, fully defined, and scalable culture systems and automated processes (Liu et al., 2020). Machine learning—often grouped under the broader umbrella of artificial intelligence (AI)—combined with automation can optimize hPSC culture, improving reproducibility and scalability for therapies and research. In addition, combining synthetic materials, including low-cost polymers and 3D configurations, with high-throughput processing compatible with pluripotency might create automated pluripotent stem cell factories (Park et al., 2023).
This review succinctly outlines the advancements in hPSC culture, highlighting the application of synthetic coatings as substrates to facilitate the indefinite proliferation of hPSCs in vitro.
1.2 Challenges in mechanistic investigations concerning feeder cells and xenogeneic elements
The incorporation of feeder cells and xenogeneic elements, including MEFs and matrices derived from animals, such as Matrigel, played a crucial role in the initial cultivation of hPSCs. Although these systems demonstrate the capacity of hPSCs for self-renewal and pluripotency, they also present considerable risks that obstruct mechanistic inquiries and translational applications (Table 1).

Table 1. Risks associated with feeder cells and xenogeneic components in hPSC culture.
Table 1 summarizes the potential hazards of feeder cells and xenogeneic components in hPSC culture, detailing impacts on quality, safety, and translation. Feeder systems can introduce zoonotic contaminants, including viruses, mycoplasma, and prions (Cobo et al., 2005). This risk is compounded by batch-to-batch variability in feeders or serum that drives inconsistent proliferation kinetics and unscheduled differentiation, undermining assay reproducibility (Villa-Diaz et al., 2009; Ieyasu et al., 2017). Prolonged culture on animal-derived matrices further exacerbates instability by increasing chromosomal abnormalities and other genomic lesions (Mikkola et al., 2006), while incorporation of non-human sialic acids such as Neu5Gc heightens immunogenicity and the likelihood of rejection in human recipients (Martin et al., 2005). Persistent paracrine cues from feeder cells bias lineage specification, and undefined feeder-derived factors reshape transcriptomes and signaling networks, eroding phenotype fidelity (Shoji et al., 2018; Eiselleova et al., 2008; Wan et al., 2022). Because such xenogeneic systems are difficult to qualify under GMP, they remain unsuitable for therapeutic-grade manufacturing and regulated testing (Unger et al., 2008). Collectively, these considerations justify a transition to defined, xeno-free culture systems that support consistent expansion and translational testing of hPSCs.
1.2.1 Feeder-free culture systems and their benefits in understanding the biology of hPSCs
The development of feeder-free culture systems marks a pivotal step toward standardizing hPSC cultivation and enhancing mechanistic studies of stem cell biology (Table 2). Unlike traditional feeder-dependent approaches, which rely on MEFs and animal-derived components, advanced feeder-free systems offer defined, xeno-free environments that reduce variability, eliminate cross-species contamination, and align with clinical manufacturing standards (Kawase and Nakatsuji, 2023). Although early feeder-free methods often used Matrigel (Xu et al., 2001), which is a complicated and inconsistent extract from mouse tumors (Hughes et al., 2010), advancements have resulted in the use of more reliable and clinically suitable materials, like recombinant laminin isoforms and synthetic polymer coatings (Villa-Diaz et al., 2013; Celiz et al., 2014a). These systems maintain hPSC self-renewal and pluripotency while providing a tractable platform for dissecting the molecular pathways that govern cell fate decisions. Consequently, feeder-free technologies not only support safe and scalable stem cell expansion for regenerative therapies but also enable reproducible, reductionist investigations into hPSC biology (Villa-Diaz et al., 2013; Kawase and Nakatsuji, 2023; Celiz et al., 2014a).

Table 2. Comparative evaluation of feeder-free vs. feeder-based culture systems in hPSC research.
Feeder-free culture is superior for most hPSC expansion and differentiation, in addition to provide benefits in safety, reproducibility, scalability, automation, and mechanistic control (Yuan et al., 2015; Villa-Diaz et al., 2009; Dakhore et al., 2018; Ozawa et al., 2023; Soares et al., 2014; Englund et al., 2010; Mei et al., 2010; James et al., 2005; Wang et al., 2005; Hanna et al., 2010; Schwedhelm et al., 2019). Defined media and extracellular matrices lower xenogeneic inputs, align with translational workflows, and decrease experimental noise, whereas feeder layers introduce undefined factors that degrade reproducibility and drive lot variability (Yuan et al., 2015; Villa-Diaz et al., 2009; Dakhore et al., 2018; Ozawa et al., 2023; Soares et al., 2014; Englund et al., 2010; Mei et al., 2010; James et al., 2005; Wang et al., 2005). Targeted supplementation of core pathways (FGF2/TGF-β with BMP antagonism) stabilizes pluripotency and improves protocol transferability (Mei et al., 2010; James et al., 2005; Wang et al., 2005; Hanna et al., 2010). Operationally, feeder-free platforms reduce QC burden and support scale-up and automation, including real-time monitored aggregate workflows in stirred systems (Ozawa et al., 2023; Soares et al., 2014; Schwedhelm et al., 2019). Engineered adhesion substrates reproducibly replace feeder-derived ECM and support clonal growth (Yuan et al., 2015; Mei et al., 2010). Although feeders can yield faster proliferation, defined systems show more predictable kinetics and scheduling (Yuan et al., 2015; Villa-Diaz et al., 2009; Englund et al., 2010; Mei et al., 2010; James et al., 2005; Wang et al., 2005). In feeder-free conditions, costs shift toward reagents but are offset by reduced labor and scalable, weekend-light operations (Dakhore et al., 2018; Soares et al., 2014; Englund et al., 2010; Schwedhelm et al., 2019). Overall, feeder-free culture increases standardization, safety, and causal interpretability while sustaining pluripotency (Yuan et al., 2015; Ozawa et al., 2023; Soares et al., 2014; Mei et al., 2010; James et al., 2005; Wang et al., 2005; Hanna et al., 2010).
1.3 Redefining hPSC expansion: a journey from non-defined media to automated, AI-guided bioprocessing
hPSC have revolutionized regenerative medicine through their unique abilities to self-renew indefinitely and differentiate into all cell types. The evolution of hPSC culture methods represents a remarkable journey from traditional feeder-dependent systems to sophisticated automated platforms. As clinical applications demand scalable, standardized production, the field has progressed toward chemically defined conditions, synthetic substrates, and AI-guided bioprocessing, marking a transformative shift in stem cell manufacturing capabilities.
Building on this foundation, automation has emerged as a critical innovation to address the demands of consistent, large-scale hPSC production. Automation in stem cell culture has emerged as a pivotal advancement. Initial efforts, initiated over a decade ago, employed microcarrier-based and bioreactor platforms to support the large-scale expansion of neural progenitor cells (Sen et al., 2002). These systems offered important benefits, like high-throughput screening capabilities, consistent and reproducible culture conditions, and a reduced risk of microbial contamination (Liu et al., 2020). Nevertheless, early technologies encountered limitations such as elevated operational costs, uncontrolled cell aggregation leading to necrosis, and poor mechanical stability of culture materials like hydrogels (Kawase and Nakatsuji, 2023; Torizal et al., 2022). Recent advances have introduced chemically defined, xeno-free media and synthetic substrates to improve reproducibility, scalability, and GMP compliance (Rivera-Ordaz et al., 2021). Automated platforms now integrate high-throughput robotics, real-time monitoring, and standardized passaging (Schwedhelm et al., 2019; Tristan et al., 2021; Thomas et al., 2009). Closed-loop systems combining microfluidics and AI enable dynamic quality control, reducing manual variability and supporting scalable, clinically aligned biomanufacturing (Park et al., 2019).
Complementing automation, the incorporation of AI has further enhanced control and efficiency in stem cell bioprocessing. AI-guided bioprocessing in stem cell research emerged around 2014, introducing data-driven optimization to enhance the efficiency, reproducibility, and standardization of cell culture workflows (Joutsijoki et al., 2014). AI facilitates automated high-throughput screening, real-time monitoring, and decision-making algorithms that reduce human error and improve process consistency. Despite these advantages, challenges remain, including variability in experimental reproducibility and the need for standardized, high-quality training datasets. Current applications of AI in stem cell biology include predictive modeling in disease research, automated phenotyping, and the development of complex three-dimensional organoids (Park et al., 2019; Herland et al., 2020).
In parallel, advancements in three-dimensional (3D) culture have introduced more physiologically relevant platforms for hPSC expansion and differentiation. 3D culture systems for hPSC emerged in the mid-2000s as a strategy to more closely replicate the in vivo microenvironment (Mohr et al., 2006). These systems offer significant advantages, including enhanced scalability for large-scale expansion and improved nutrient and gas exchange, both critical for clinical translation. Nonetheless, several technical challenges, such as uncontrolled cell aggregation, which can lead to necrotic core formation, and difficulties in achieving uniform cell distribution are associated with 3D culture systems (Wu et al., 2014). Recent advancements have focused on engineering defined biomaterials, including peptide-based hydrogels and nanofibrous matrices, to provide structural support and improve culture homogeneity (Zhou et al., 2022).
Together, these emerging technologies converge to define the future landscape of stem cell biomanufacturing. In summary, the integration of 3D culture systems, automated bioprocessing, and AI is poised to transform large-scale hPSC manufacturing. Future directions will emphasize the combination of 3D platforms with bioreactor-based automation to enable robust, reproducible, and GMP-compliant production for regenerative and personalized therapies. AI, particularly when integrated with nanotechnology and 3D bioprinting, will enhance predictive modeling, adaptive control, and tissue engineering precision. These improvements are crucial for consistent hPSC production and improved reliability in applications such as disease modeling, drug screening, and clinical translation, marking a pivotal shift toward next-generation stem cell biomanufacturing.
2 Comparative analysis of animal-free culture systems for human pluripotent stem cells: approaches, challenges, and clinical implications
hPSC expansion requires defined culture conditions to preserve self-renewal and pluripotency. Traditional systems use xenogeneic components such as feeder cells and Matrigel, introducing variability and risks of contamination. In response, xeno-free platforms using chemically defined media, recombinant human proteins, synthetic matrices, and plant-derived substrates (Liu et al., 2020) have been developed to enhance consistency, safety, and regulatory compliance. This section examines animal-free hPSC culture systems across the field’s evolution, spanning the past Traditional and Transitional periods, the current robotic, AI and machine-learning era and the future industrial-automated period aimed at scalable manufacturing (Figure 1).

Figure 1. Comparative evolution of animal-free hPSC culture systems. Illustration showing the shift from feeder/xeno substrates and serum media to defined xeno-free matrices and media, then to robotic/AI-assisted automation, and finally industrial closed-bioreactor manufacturing.
2.1 Chemically defined and xeno-free media for animal-free hPSC culture
Chemically defined and xeno-free media represent a major advancement in hPSC culture, enabling consistent maintenance of pluripotency while eliminating the variability and risks associated with animal-derived components. Traditional systems such as mTeSR™1 (47), although feeder-free, contained bovine serum albumin and other xenogeneic additives. In response, next-generation formulations like Essential 8 (E8) (Chen et al., 2011) were developed to include only defined components—such as insulin, transferrin, FGF2, and TGF-β—while excluding serum albumin and β-mercaptoethanol to achieve full chemical definition. Further innovations have led to xeno-free commercial media, including TeSR-E8 (Chen et al., 2011), NutriStem hPSC XF (Biological Industries. Press Release, 2016), and StemFit® AK02N(50), which support GMP-compliant hPSC culture using recombinant or human-derived proteins on defined substrates (Skorik et al., 2020; Timilsina et al., 2023). Table 3 summarizes key features of these and other chemically defined, xeno-free media designed to support safe and reproducible hPSC culture in feeder-free conditions.

Table 3. Comparison of chemically defined and xeno-free media.
Table 3 summarizes key features of these and other chemically defined, xeno-free media designed to support safe and reproducible hPSC culture in feeder-free conditions. The development of chemically defined, xeno-free media has significantly advanced hPSC culture by improving reproducibility, reducing contamination risks, and facilitating compliance with clinical manufacturing standards. Media formulations now commonly utilize recombinant human proteins or plant-derived analogs, such as plant-produced FGF2, EGF, and albumin, to achieve fully animal-origin-free systems (Lee et al., 2022). These innovations enable robust, scalable, and standardized platforms suitable for both research and therapeutic applications. Continued refinement of these systems will be essential for supporting the safe and efficient clinical translation of stem cell–based therapies.
2.2 Substrates for feeder-free culture
In addition to defined media, an animal-free hPSC culture system requires a suitable substrate or matrix for cell attachment. hPSCs survival and self-renewal are compromised by untreated tissue culture plastic and traditionally depend on either feeder cell layers or complex extracellular matrix (ECM) mixtures for adhesion and growth factor support.
2.2.1 Matrigel as the gold standard substrate for iPSC culture: benefits and limitations
Matrigel has been the most used substrate for feeder-free culture of hPSC, including hiPSCs, due to its robust support for cell adhesion, proliferation, and maintenance of pluripotency (Xu et al., 2001). Derived from Engelbreth-Holm-Swarm (EHS) mouse sarcoma (Rodin et al., 2010; Kleinman et al., 1986), Matrigel is composed primarily of laminin, collagen IV, entactin, and perlecan, along with a complex mix of growth factors and over 1,800 unique proteins (Hughes et al., 2010). Despite its widespread utility in basic research, Matrigel’s undefined composition, xenogeneic origin, and batch-to-batch variability pose significant challenges for standardization and clinical translation (Dakhore et al., 2018; Ozawa et al., 2023). Although Matrigel remains a reliable matrix for experimental hPSC culture, its limitations have prompted a shift toward xeno-free, chemically defined, and GMP-compliant substrates for therapeutic applications (Melkoumian et al., 2010; Villa-Diaz et al., 2010). Advances in recombinant extracellular matrix proteins (e.g., laminin-511, vitronectin), synthetic peptides, and defined hydrogels offer improved reproducibility, safety, and scalability (Zhou et al., 2022; Ozawa et al., 2023; Miyazaki et al., 2012). These emerging platforms align with regulatory requirements and are more compatible with automation and bioreactor systems, facilitating the transition toward robust, clinical-grade iPSC expansion and differentiation (Celiz et al., 2014a).
2.2.2 Synthetic and recombinant substrates for feeder-free culture
To eliminate the variability and xenogeneic risks associated with Matrigel and feeder cell systems, recombinant ECM proteins and fully synthetic substrates have been developed as defined alternatives for feeder-free hPSC culture.
Recombinant human ECM proteins have proven effective as defined substrates (Table 4). Laminins and vitronectin are two key adhesion ligands for hPSCs. Among these, vitronectin and fibronectin were among the first applied, with defined fragments like vitronectin-N supporting hPSC adhesion via αvβ5 integrins and maintaining self-renewal in xeno-free conditions (Braam et al., 2008). However, both vitronectin and fibronectin may show lower attachment efficiency or require higher coating concentrations compared to Matrigel (Badenes et al., 2016). In contrast, laminin-511 and laminin-521—basement membrane proteins expressed in the embryonic inner cell mass and placenta—more closely replicate the native hPSC niche (Xu et al., 2001). Laminin-511 supports long-term self-renewal (Rodin et al., 2010), while laminin-521 promotes single-cell survival (Rodin et al., 2014), clonal expansion, and sustained pluripotency. These substrates act primarily through α6β1 integrin signaling in hPSCs (Villa-Diaz et al., 2016) and are commercially available in defined, clinical-grade formats (e.g., iMatrix-511), making them widely used in GMP-compliant protocols for hiPSC expansion and directed differentiation.
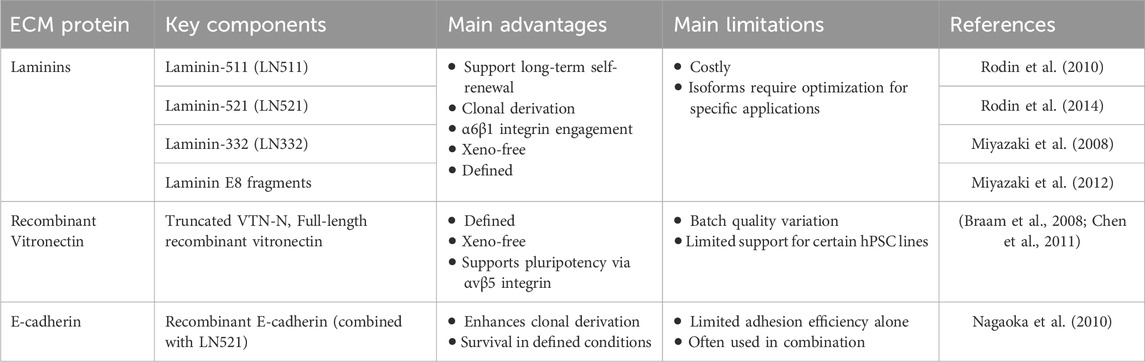
Table 4. Recombinant human ECM protein substrates for hPSC maintenance.
On the other hand, synthetic peptide-based substrates have emerged as defined and xeno-free alternatives to traditional extracellular matrix coatings for hPSC culture (Table 5). Vitronectin-derived peptides such as VN1:KGGPQVTRGDVFTMP (Deng et al., 2013), VN2: CGGKKQRFRHRNRKG (Park et al., 2015), and commercially available Synthemax (a vitronectin peptide–acrylate conjugate) (Melkoumian et al., 2010) support robust cell adhesion, self-renewal, and long-term maintenance of pluripotency under feeder-free conditions. These substrates are scalable and chemically defined, making them well-suited for research and preclinical applications (Badenes et al., 2014). However, their performance may vary across different hPSC lines and often requires optimization. RGD-containing peptides, including heparin-binding variants GKKQRFRHRNRKG (Klim et al., 2010) and optimized vitronectin mimetics (e.g., Ac-KGGPQVTRGDTYRAY) (Zhou et al., 2018) function by mimicking natural integrin-binding domains to promote cell attachment and survival. While promising for translational use, these systems are sensitive to peptide density and conformation, which may affect reproducibility. Overall, synthetic peptide substrates represent a critical step toward standardized, xeno-free platforms for hPSC expansion and differentiation.

Table 5. Synthetic peptide-based culture systems for hPSC maintenance.
Synthetic polymer-based substrates have been developed as defined, xeno-free platforms for the maintenance and expansion of hPSCs (Table 6). Polyzwitterion, acrylate, and methacrylate polymers such as PMEDSAH (Villa-Diaz et al., 2010), APMAAm (Higuchi et al., 2015), and Poly (HPhMA-co-HEMA) (Deng et al., 2013) support long-term self-renewal and pluripotency in feeder-free conditions, although their synthesis and surface coating protocols can be complex. Other synthetic polymers, including Poly (TCDMDA-blend-BA) (Park et al., 2015), PMVE-alt-MA (Chen et al., 2017), and, PVB (Otsuji et al., 2014) offer biocompatibility and GMP potential; however, they often require cell-type-specific optimization. Temperature-responsive polymers like PEGMA brush (Celiz et al., 2014b), PVA (Forghani et al., 2017), and PNIPAM (Otsuji et al., 2014) enable enzyme-free passaging and are promising for automation and large-scale production; however, they are still in the experimental stage due to concerns regarding stability and reproducibility. Collectively, these materials contribute to the development of scalable, standardized hPSC culture systems compatible with clinical applications.

Table 6. Synthetic polymer-based culture systems for hPSC maintenance.
In summary, the development of synthetic and recombinant substrates has markedly improved feeder-free culture systems for hPSCs by introducing defined, xeno-free, and scalable alternatives to biologically derived matrices. Although further work is needed to address issues related to cost-efficiency, cell-line-specific optimization, and regulatory validation, these substrates represent a pivotal advancement toward the establishment of standardized, GMP-compliant platforms for clinical-grade stem cell manufacturing.
2.3 Three-dimensional (3D) culture systems
Three-dimensional (3D) culture systems have emerged as essential platforms for hPSC maintenance and expansion, offering improved physiological relevance compared to traditional two-dimensional systems. By facilitating enhanced cell–cell and cell–matrix interactions, 3D cultures better replicate in vivo conditions and enable scalable production required for downstream clinical and research applications. Common 3D strategies include aggregate cultures (scaffold-free and scaffold-supported), microwell/ULA plate–based aggregation, microcarrier-based suspension systems, microencapsulation in hydrogels, bio-printed constructs, acoustic/levitation assembly, and organ-on-chip (microfluidic) platforms, each with distinct biochemical and mechanical properties (Zhou et al., 2022). These approaches have been adapted for feeder-free and xeno-free culture, aligning with regulatory and GMP standards. Table 7 outlines the key components, advantages, and limitations of the major 3D hPSC culture systems currently used in the field.

Table 7. Three-dimensional (3D) culture systems for hPSC maintenance.
As summarized in Table 7, each 3D culture system has specific strengths and limitations. Aggregate methods are cost-effective and easy to implement but may suffer from poor size control and shear stress-related damage (Otsuji et al., 2014; Lancaster et al., 2013). Microwell/ULA plate systems provide tight size control and high uniformity but are constrained by plate scale and throughput (Mohr et al., 2010). On the other hand, scaffold-supported aggregates embed spheroids in defined hydrogels (e.g., PEG-DA, GelMA, fibrin), adding tissue-like architecture but inheriting matrix variability and diffusion limits. Microcarrier-based systems also offer high surface area for large-scale expansion, with common coatings such as vitronectin, laminin-521, or synthetic peptides; however, challenges associated include coating cost, cell–bead separation, and shear sensitivity (Oh et al., 2009). Microencapsulation protects cells and standardizes aggregate size using alginate, hyaluronic acid, or PNIPAAm–PEG hydrogels, yet is limited by nutrient diffusion, difficult cell retrieval, and material costs (Kim et al., 2013). Bio-printed constructs enable spatial control and vasculature templating in printable bioinks (GelMA, PEGDA, fibrin, alginate), however, face print-induced shear and rheology constraints (Faulkner-Jones et al., 2015). Acoustic/levitation assembly is contactless and label-free, but requires specialized equipment and currently can be achieved in modest scale (Hahn et al., 2025). Organ-on-chip (microfluidic) platforms recreate barrier interfaces and perfusion on PDMS or thermoplastics with defined ECM coatings, trading physiological fidelity for lower throughput and fabrication standardization (Ellis et al., 2017). Thus, the choice of system should be guided by application-specific needs—such as scalability, reproducibility, and clinical compatibility—as the field advances toward standardized, GMP-compliant hPSC manufacturing.
2.4 Recent advances in automating the techniques for culturing hPSCs
Reprogramming hiPSCs is a multi-step, time-consuming process requiring strict sterility and continuous manual monitoring. This process is inherently labor-intensive and prone to variability. Cellular heterogeneity resulting from lineage reprogramming complicates the isolation of monoclonal hiPSC lines. These challenges underscore the need for scalable, standardized cell manufacturing platforms to enable efficient clinical and research applications (Celiz et al., 2014a). Manual culture techniques are incompatible with high-throughput production, driving the demand for automation; therefore, advances in automated technologies have led to the development of several platforms for hiPSC reprogramming and maintenance.
Table 8 summarizes key technological advancements in automating the generation and cultivation of hiPSCs. It highlights pre-automation and early machine-vision monitoring for continuous, noninvasive observation (Narkilahti et al., 2007), the first bench-scale liquid-handling platforms for standardized maintenance and differentiation (Thomas et al., 2009), automated imaging with colony picking to improve clonality (Haupt et al., 2012), machine-learning quality control based on colony and nuclear morphology (Tokunaga et al., 2014), deep-learning segmentation enabling objective masks (Zhang et al., 2019), and time-lapse AI evaluation for label-free QC (Kato et al., 2016), CNN-based colony classification for rapid triage (Witmer and Bhanu, 2018), and modular robotics for closed, GMP-oriented manufacturing (Tissue Factory, StemCellFactory, and high-throughput robotic lines) (Tristan et al., 2021; Kikuchi et al., 2018; Zhang et al., 2019; Elanzew et al., 2020). Recent U-Net–guided clone picking further reduces operator bias while improving clonality (Powell et al., 2023). Together, these systems improve reproducibility, reduce variability, and support GMP-compatible scaling from reprogramming through expansion and directed differentiation.

Table 8. Key technological advancements in automating the generation and cultivation of hiPSC.
2.5 Validation of pluripotency under animal-free conditions
A key concern in adopting animal-free hPSC culture systems is whether they can robustly support self-renewal and multilineage differentiation. Multiple studies have demonstrated that hPSCs maintained under feeder-free, xeno-free conditions retain hallmark pluripotency markers—such as OCT4, NANOG, SSEA4, and TRA-1-60—in over 90%–95% of cells, as confirmed by immunostaining and flow cytometry (Villa-Diaz et al., 2010; Badenes et al., 2014; Shimizu et al., 2022). These cultures also preserve a stable diploid karyotype across extended passages in defined media like Essential 8 and TeSR-E8. Furthermore, gene expression analyses show sustained activation of core pluripotency genes (e.g., SOX2, OCT4, NANOG), confirming genomic stability and undifferentiated status (Timilsina et al., 2023). Critically, hPSCs expanded under animal-free conditions retain full developmental potential, as evidenced by their ability to differentiate into all three germ layers in vitro and in vivo, including through embryoid body and teratoma assays (Villa-Diaz et al., 2010; Chen et al., 2017). Collectively, these findings validate that animal-free culture systems are suitable for maintaining both molecular and functional pluripotency, supporting their use in clinical and translational applications.
2.6 Clinical and translational implications of animal-free hPSC culture
The transition to animal-free hPSC culture is driven by the increasing demand for regulatory compliance and clinical-grade manufacturing. Regulatory agencies such as the U.S. Food and Drug Administration (FDA), European Medicines Agency (EMA), and Japan’s Pharmaceuticals and Medical Devices Agency (PMDA) mandate the use of xeno-free, traceable, and well-characterized reagents to mitigate risks associated with animal-derived components (Ozawa et al., 2023; Tanaka et al., 2023). In alignment with current GMP standards, defined and animal-free systems— such as Essential 8 medium and recombinant laminins— are now integral to the production of clinical-grade hPSC derivatives. These platforms have already supported early-phase clinical trials, including transplantation of hESC-derived retinal pigment epithelial (RPE) cells for macular degeneration without culture-related adverse events (Da Cruz et al., 2018). Beyond safety, animal-free systems offer enhanced scalability and integration with bioreactor-based bioprocesses. Unlike feeder-dependent systems or Matrigel, defined synthetic substrates and microcarriers can be incorporated into automated workflows for large-scale stem cell production. Demonstrations of successful hPSC expansion in stirred-tank bioreactors using xeno-free media underscore the translational feasibility of these systems (Krawetz et al., 2010). Furthermore, regulatory submissions now require comprehensive Chemistry, Manufacturing, and Controls (CMC) documentation, including the provenance of all culture components. In addition, animal-free systems simplify CMC dossiers by minimizing immunogenic risks and offering consistent manufacturing inputs (Herland et al., 2020). The 2024 FDA guidance further mandates full disclosure and justification of all culture materials in Investigational New Drug (IND) applications, positioning recombinant human albumin and other synthetic substitutes as preferred alternatives. Collectively, these advances ensure that hPSCs cultured under animal-free conditions can meet the stringent quality, safety, and scalability requirements of regenerative therapies.
2.7 Toward safe, scalable, and regulatory-compliant hPSC culture systems
The transition to animal-free, chemically defined culture systems marks a pivotal evolution in hPSC research, offering enhanced consistency, safety, and regulatory readiness. As demonstrated across various platforms—from xeno-free media and synthetic substrates to three-dimensional bioreactor-compatible matrices—these systems maintain the core properties of pluripotency while enabling scalable and GMP-compliant manufacturing. Comparative analysis reveals that while each system has distinct advantages and limitations, collectively they represent a robust foundation for clinical grade hPSC expansion and differentiation. Ongoing validation of genomic stability, lineage specificity, and therapeutic efficacy will be essential to fully realize the translational potential of these innovations. Ultimately, the adoption of animal-free systems is not only a technical refinement but also a necessary step toward ethical, standardized, and clinically viable stem cell-based therapies.
3 hiPSC technology in drug discovery and regenerative medicine
hiPSC technology has advanced the clinical and pharmaceutical research landscapes. By generating human disease models with unprecedented accuracy, hiPSCs facilitate in-depth clinical investigations that were previously unattainable. In pharmaceutical research, hiPSC-derived models enable high-throughput screening and toxicity studies, revolutionizing drug discovery by accelerating the identification of potential therapeutics. In the future, hiPSC technology is expected to enable clinical applications—including tissue and organ replacement and personalized medicine via disease-in-a-dish models—providing new avenues for tailored treatments and improved patient outcomes. This convergence of scientific innovation holds immense promise for reshaping medical research paradigms and improving healthcare delivery worldwide.
3.1 hiPSC-based disease modeling
hiPSC-based disease modeling has revolutionized biomedical research by enabling the generation of patient-specific cells that retain the genetic background of the donor. Since their introduction in 2006, hiPSCs have provided a renewable source of pluripotent cells capable of differentiating into disease-relevant lineages, thereby allowing mechanistic studies in a human genetic context (Takahashi et al., 2007; Vo et al., 2024). Compared to hESCs, hiPSCs bypass ethical concerns and are more accessible, facilitating the study of a broader range of inherited and sporadic diseases. The integration of genome editing tools such as CRISPR/Cas9 further enhances model fidelity by enabling the generation of isogenic controls, which help distinguish pathogenic variants from background genetic noise (Hockemeyer and Jaenisch, 2016). Together, these capabilities position hiPSCs as a powerful system for modeling complex human diseases with unprecedented precision.
As summarized in Tables 9–11, diverse organoid systems have been established for diseases of endodermal, mesodermal, and ectodermal origins, including lung, liver, gastric, intestinal, pancreatic organoids, cardiac, renal-nephrotic, muscular organoids/spheroid, brain, cerebral, retinal, spinal organoids. In addition to organoids following are examples of popular disease modeling: ARPKD disease models, Polycystic Kidney Disease, Seckel syndrome, Miller-Dieker lissencephaly, Alzheimer’s disease, Fragile X syndrome, Leigh syndrome, Parkinson’s disease, Rett syndrome, FTD, Spinal muscular atrophy, Glaucoma, RP, FRD, altogether demonstrating the versatility and precision of this modeling approach.
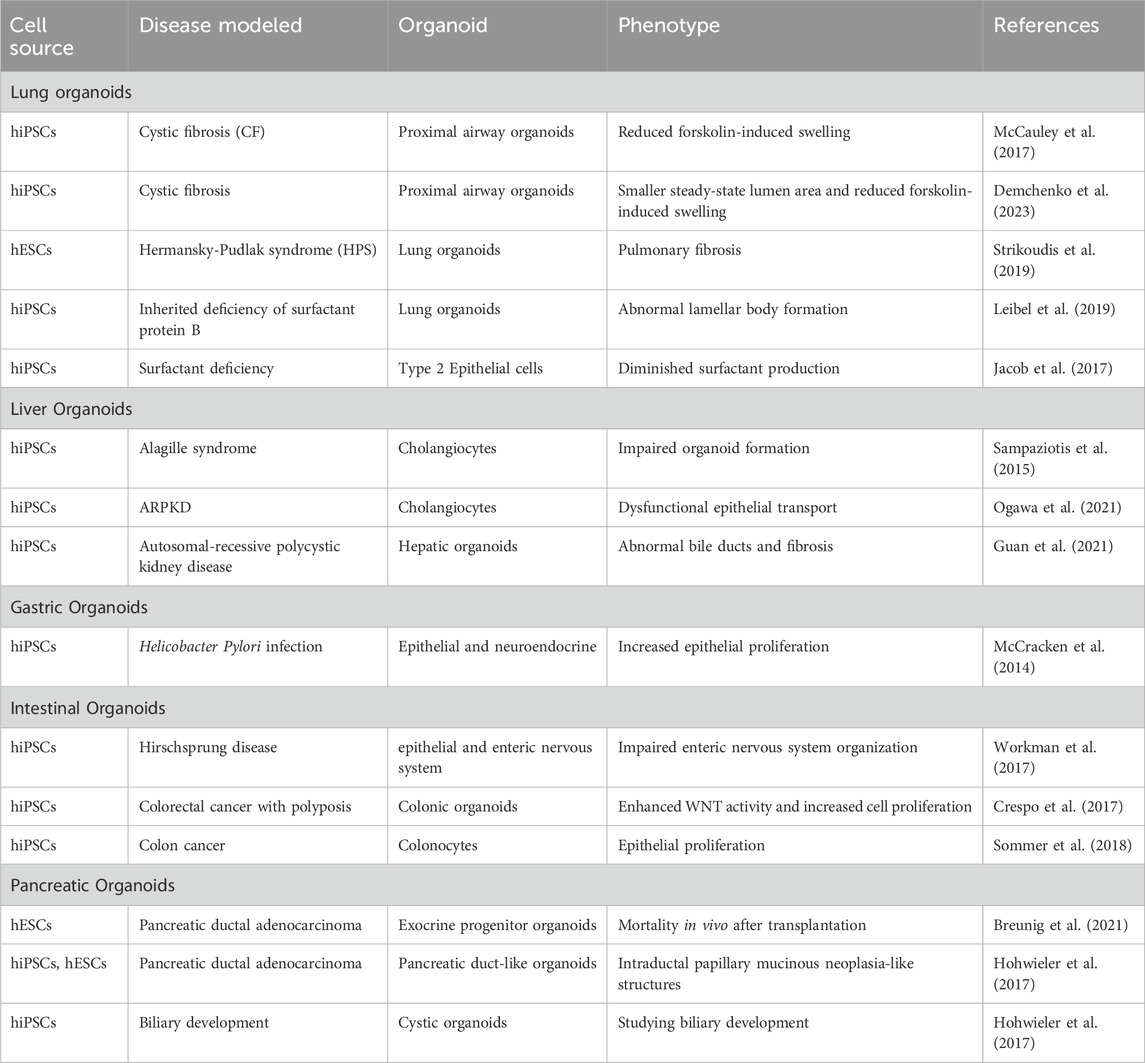
Table 9. Overview of key hPSC-derived organoids in disease research and modeling with endoderm origin.
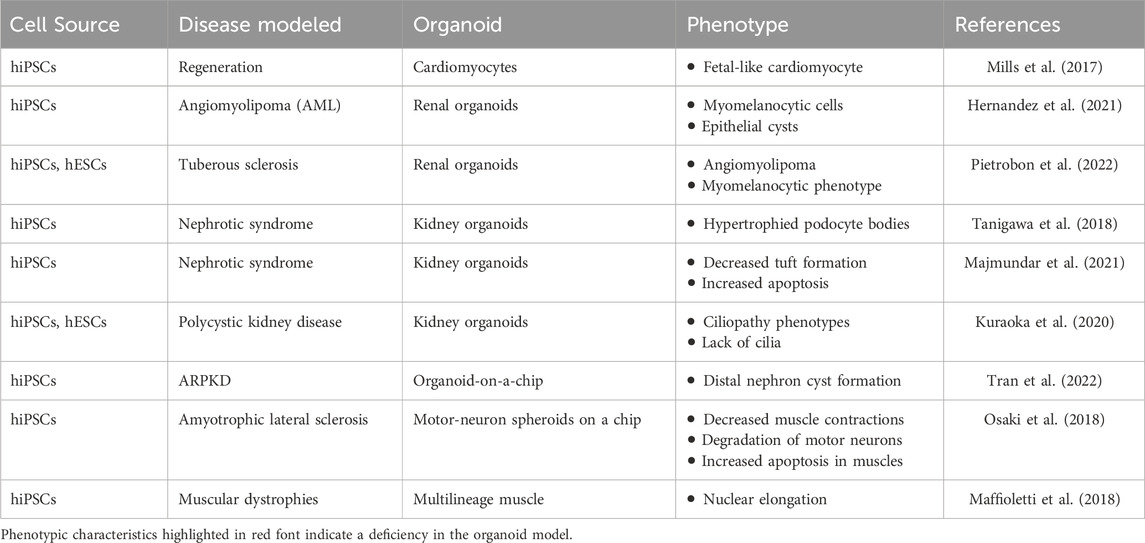
Table 10. Overview of key hPSC-derived organoids in disease research and modeling of mesoderm origin.

Table 11. Overview of key hPSC-derived organoids in disease research and modeling of ectoderm origin.
3.2 hiPSC-based drug discovery and development
hiPSCs have transformed drug discovery by enabling high-throughput screening (HTS) platforms that are both physiologically relevant and scalable. By differentiating into disease-relevant cell types, hiPSCs support mechanistic studies, phenotypic screening, and safety profiling across diverse disease models.
Table 12 summarizes hiPSC-based high-throughput screening strategies across platforms, assay modalities, and applications. Isogenic disease modeling uses genome-edited pairs to isolate genotype–phenotype relationships and quantify drug responses (Soldner et al., 2011). High-content imaging enables automated, scalable morphology and multiparameter phenotyping (Bray et al., 2016). Electrophysiological assays, including microelectrode arrays and calcium imaging, measure neural and cardiac function and detect proarrhythmic or neurotoxic effects (Sala et al., 2018). Chemogenomic phenotypic screening applies annotated compound libraries to map pathway dependencies and support target validation (Grafton et al., 2021). Organotypic co-cultures—organoids and microfluidic systems—reconstitute tissue architecture and cell–cell interactions for context-aware screening (Huh et al., 2010). Multi-omics integration aggregates transcriptomic and proteomic readouts to discover biomarkers and stratify patient-relevant responses (Volpato et al., 2018). Together, these modalities provide quantitative phenotypes, strengthen mechanism-of-action inference, and increase translational value for preclinical decision-making.

Table 12. Summary of iPSC-based high-throughput screening (HTS) strategies and their applications.
3.3 The future of regenerative medicine: advancements and bottlenecks
Regenerative medicine has advanced significantly since the first derivation of hESCs in 1998 and the generation of hiPSCs in 2007. These cell types offer the ability to repair or replace damaged tissues, with hiPSCs providing additional advantages due to their patient-specific origin and ethical acceptability. The integration of hiPSC technology with precise genome editing and 3D organoid culture has expanded the potential for developing personalized cell-based therapies. Despite these breakthroughs, several critical challenges hinder clinical translation. Long-term culture of pluripotent cells can lead to tumorigenicity and genetic instability, necessitating rigorous quality control measures. hiPSC-derived therapeutic products must be evaluated for genomic integrity, purity, and sterility under cGMP (Liang et al., 2013). These requirements include the use of xeno-free, chemically defined media and the routine verification of normal karyotypes and pluripotency markers. Producing cell therapy products that comply with GMP standards can cost more than $800,000 per application. In addition, the cost to generate a single iPSC cell line can be as high as $20,000 (or $0.1 per well in a 96-well plate) using currently most selected substrates like Matrigel and Synthemax (Shi et al., 2017). Additional hurdles include establishing effective immunological tolerance, especially for hESC-based interventions. Addressing these technical, regulatory, and economic barriers is vital to unlock the full therapeutic promise of hPSCs in clinical regenerative medicine.
4 Current applications of hiPSC technology in toxicity screening
4.1 Application of hiPSC technology in toxicity screening
Traditional toxicity screening has long relied on animal models and immortalized cell lines, which often fail to predict accurate human responses to drugs and environmental toxins. This limitation has contributed to the high failure rates in drug development, with approximately 30% of potential therapeutics terminated due to unforeseen toxicities, particularly cardiotoxicity (Fritsche et al., 2021). This high attrition rate is largely attributed to the poor predictive capacity of conventional preclinical models, which fail to forecast human-specific toxicological outcomes reliably. The emergence of hiPSC technology has offered a transformative solution to these challenges by providing scalable, human-relevant cellular models for toxicity assessment. hiPSC-derived cells offer several advantages over conventional screening methods, including their human origin, ability to differentiate into disease-relevant cell types, and capacity to model patient-specific responses (Fritsche et al., 2020). The technology enables the development of predictive assays that can identify safety concerns early in the drug development process, potentially reducing costly late-stage failures.
Table 13 summarizes the advantages and limitations of both conventional and computational preclinical models. Conventional models each add value yet leave key translational gaps. Cancer cell lines are scalable and low-cost for target-based assays but drift genomically and lack tissue specificity and human predictivity (Trastulla et al., 2022). Primary human cells benchmark physiology but are short-lived, donor-variable, and hard to scale (Bulutoglu et al., 2020). Animal models provide systemic biology and PK/PD context yet face interspecies differences, ethics, and high cost/time (Yang and Papoian, 2018). Ex vivo tissues preserve native architecture but have short viability and low throughput (Jin et al., 2021). Two-dimensional cultures are reproducible but are missing matrix, mechanical, and paracrine cues needed for microenvironment fidelity (Ware et al., 2015). In silico approaches are rapid and inexpensive but depend on high-quality training data, require experimental validation, and have limited dynamical realism (Fritsche et al., 2020). Human iPSC-derived models address these limitations by generating renewable, patient-specific, lineage-accurate tissues; enabling isogenic controls and population genetics; and supporting advanced formats— organoids and micro-physiological chips— that restore microenvironmental mechanics, multicellular interactions, and cross-tissue crosstalk, thereby improving mechanistic insight and human predictivity.
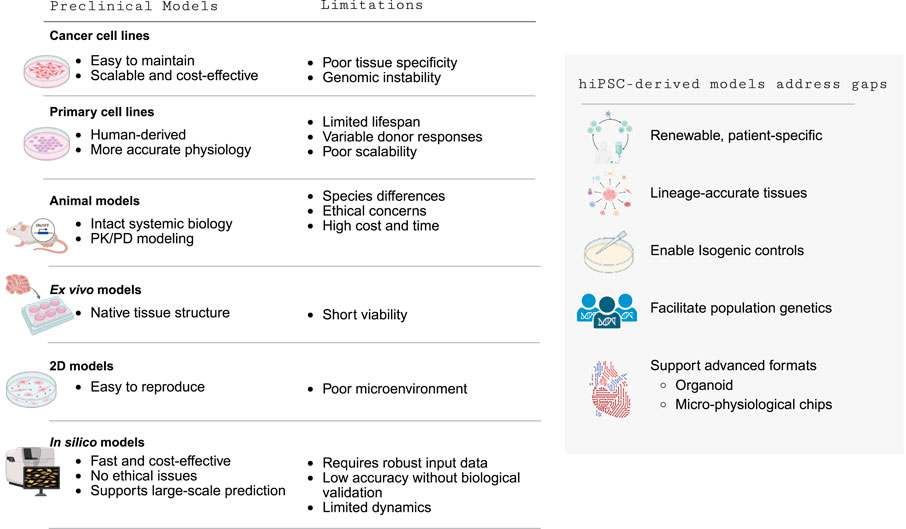
Table 13. Advantages and limitations of conventional and computational preclinical models.
4.2 hiPSC in toxicology: advancing beyond traditional models
hiPSC-based toxicity screening offers compelling advantages over traditional methods, addressing fundamental limitations in current safety assessment paradigms. The human relevance of iPSC-derived models represents a critical breakthrough, overcoming species differences inherent in animal models that often fail to predict human responses (Fritsche et al., 2020). Mouse models, for instance, poorly mimic human inflammatory diseases and drug responses, leading to the need for human-based systems (Seok et al., 2013). hiPSCs can be differentiated into diverse cell types, including hepatocytes, cardiomyocytes, neurons, and other disease-relevant cells, enabling comprehensive toxicity assessment across multiple organ systems. The technology’s capacity for patient-specific modeling allows personalized toxicity testing and pharmacogenomics studies, enabling researchers to assess how genetic variations influence individual susceptibility to toxic compounds (Yildirim et al., 2025). This approach facilitates disease modeling by testing compounds in disease-affected cell lines, demonstrating how pathological conditions may alter toxicity profiles. hiPSC platforms have high-throughput potential and are compatible with automation and multi-well formats for large-scale screening applications (Tristan et al., 2021). Additionally, these systems offer significant ethical advantages by reducing animal usage, aligning with the 3Rs principle (replacement, reduction, and refinement), and supporting regulatory initiatives like the FDA Modernization Act 2.0 (Yildirim et al., 2025; Tannenbaum and Bennett, 2015).
4.3 Integrated toxicity screening using iPSC-derived human cells
Human iPSC-derived cell types are increasingly utilized across a broad range of toxicological contexts, providing organ-specific models that enhance the detection and characterization of adverse effects.
Figure 2 summarizes the current applications of hiPSC-derived cells in modeling toxicity across various organ systems. Specifically, hiPSC cardiomyocytes quantify proarrhythmic risk, including QT prolongation, hERG blockade, contractility deficits, and drug-induced arrhythmias (Visone et al., 2023). Moreover, hiPSC hepatocyte-like cells enable DILI assessment by capturing mitochondrial dysfunction, bile-acid transport defects, and broader metabolic toxicities (Tamargo-Rubio et al., 2023). Likewise, neural hPSC models (neurons and astrocytes) reveal seizure liability, neurodevelopmental perturbations, neurodegeneration, and synapto-toxicity (Tukker et al., 2019). In addition, kidney models composed of proximal tubule and podocyte lineages resolve filtration and transporter-mediated injuries typical of agents such as cisplatin and aminoglycosides (Lawrence et al., 2022). Similarly, hematopoietic progenitor systems detect marrow suppression, genotoxicity, and cytopenias (Fransen and Leonard, 2023). Finally, integrated platforms that combine organoids, micro-physiological chips, automation, and high-content phenotyping extend these assays to scalable safety pharmacology and compound prioritization (Vandana et al., 2023). Collectively, these human-relevant models increase mechanistic resolution and improve translational decision-making.
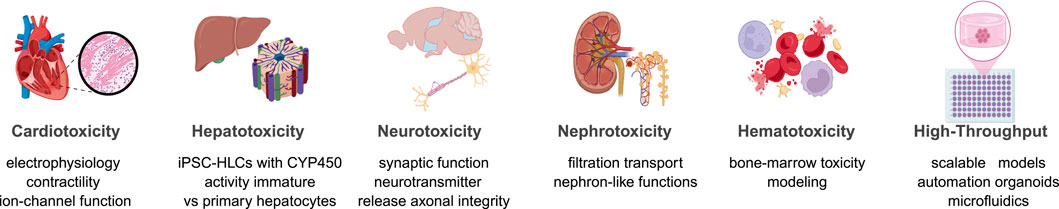
Figure 2. Current applications of iPSC technology in toxicity screening.
The power of hiPSC-based toxicity screening is significantly enhanced by integrating advanced analytical technologies that enable detailed and high-throughput assessment of cellular responses.
Table 14 summarizes key technologies, their applications, and associated methodological features in current hiPSC-based toxicology platforms. High-content imaging quantifies morphology, apoptosis, and mitochondrial integrity in hiPSC-derived cardiomyocytes, hepatocytes, and neurons, enabling scalable phenotypic screening (Tolosa et al., 2015). Microelectrode arrays and calcium assays in hiPSC-cardiomyocytes provide electrophysiological endpoints aligned with pro-arrhythmia risk assessment and CiPA frameworks (Vandana et al., 2023). Brain, liver, and kidney organoids supply tissue-level architecture and paracrine signaling for systems-level toxicity readouts, albeit with variability that requires rigorous QC (Grimm et al., 2015). Multi-omics layers (transcriptomics, proteomics, metabolomics) map mechanism-of-action and furnish candidate biomarkers for stratification and translation (Nabi et al., 2022). CRISPR-engineered isogenic hiPSC lines resolve gene–environment and pharmacogenomic interactions, strengthening causal inference and target validation (Kim et al., 2020; Sayed et al., 2020). Together, these integrated modalities deliver human-relevant, multiparametric endpoints that improve predictive validity and support regulatory-grade decision making.

Table 14. Integrated technologies supporting hiPSC-based toxicity screening: applications, capabilities, and limitations.
4.4 Future directions in hiPSC-based toxicity screening
The field of hiPSC-based toxicity screening is undergoing rapid transformation, driven by advances that aim to enhance physiological relevance, scalability, and predictive accuracy. Efforts are focused on developing mature, multicellular organoid systems that better replicate adult tissue functionality through strategies such as mechanical stimulation, extended culture, and co-differentiation. Concurrently, the integration of hiPSC-derived tissues with organ-on-a-chip platforms is enabling dynamic modeling of tissue-tissue interfaces, fluidic flow, and multi-organ interactions—key for capturing systemic toxicity (Park et al., 2019). AI is increasingly applied to extract meaningful patterns from high-dimensional datasets, including omics, imaging, and electrophysiology, thereby improving early toxicity prediction (Vo et al., 2024; Tran et al., 2023). Furthermore, the expansion of hiPSC models into environmental and precision toxicology is enabling the study of gene-environment interactions and personalized risk profiles.
These innovations (Figure 3) collectively position hiPSC-based systems as next-generation platforms for mechanistic toxicology, regulatory testing, and individualized safety assessment. Mature multicellular organoids, organ-on-chip integration, AI-driven prediction, and environmental/precision toxicology are converging to increase physiological fidelity and decision utility. Organoid maturation by prolonged culture, defined mechanical cues, and strategic co-culture improves adult-like and age-relevant responses, yet immaturity, variability, and limited standardization persist (Fritsche et al., 2021). Coupling hiPSC-derived tissues to microfluidic chips enables controlled flow, tissue–tissue interfaces, and multi-organ coupling for dynamic, real-time readouts; however, complexity, cost, and scalability on these systems remain as barriers (Ahmed and Moreira Teixeira, 2022). Machine-learning models that mine high-content imaging, electrophysiology, and multi-omics can reveal subtle injury signatures and improve prediction, but they require large, high-quality datasets, rigorous model development and validation (Tran et al., 2023). Patient-specific hiPSC models support quantification of genotype–exposure interactions for individualized risk assessment and public-health protection; robust data integration, external validation, and regulatory uptake are still needed to validate this individual models (Chandy et al., 2022). Collectively, these trajectories position hiPSC platforms for next-generation mechanistic toxicology, regulatory testing, and individualized safety assessment, contingent on harmonized methods and regulatory alignment (Tran et al., 2023; Ahmed and Moreira Teixeira, 2022; Chandy et al., 2022; Fritsche et al., 2021).

Figure 3. Advancing iPSC-based toxicity screening-technologies, applications, and translational considerations.
4.5 Barriers to implementation of hiPSC-based toxicology platforms
Figure 4 illustrates barriers to implementing hiPSC-based toxicology platforms, which include persistent cellular immaturity across lineages—such as low IK1 and glycolytic bias in cardiomyocytes (Yang et al., 2014), fetal CYP profiles in hepatocytes (Gurevich et al., 2020), and incomplete synaptic/myelination features in neural cultures (James et al., 2021), which limits modeling of adult and chronic toxicities. Other barriers include: limited scalability and reproducibility driven by donor, clone, and batch effects and by matrix, media, and passage-dependent drift, reducing inter-laboratory concordance and HTS readiness (Volpato and Webber, 2020); lack of harmonized protocols, standardized endpoints, and qualified QC materials, which impedes cross-study comparison and assay qualification (OECD, 2018); restricted functional complexity due to absent vasculature, immune components, and organ-level microanatomy (e.g., sinusoid, glomerulus, neurovascular unit), diminishing physiological fidelity (Yip et al., 2023); and lagging regulatory acceptance because contexts of use, reference compound sets, performance standards, and formal validation remain incomplete despite enabling policies (e.g., FDA Modernization Act 2.0) (Sewell et al., 2024). Active mitigation actions includes maturation strategies (prolonged culture, metabolic/electrical/mechanical conditioning, co-culture), automated standardized SOPs with in-process QC and reference panels, interlaboratory ring trials, and bioengineered, vascularized, immune-competent, 3D, and micro-physiological systems advanced with regulators (Sewell et al., 2024). Addressing these bottlenecks will transition hiPSC assays from research tools to validated human-relevant tests that support regulatory safety decision-making.
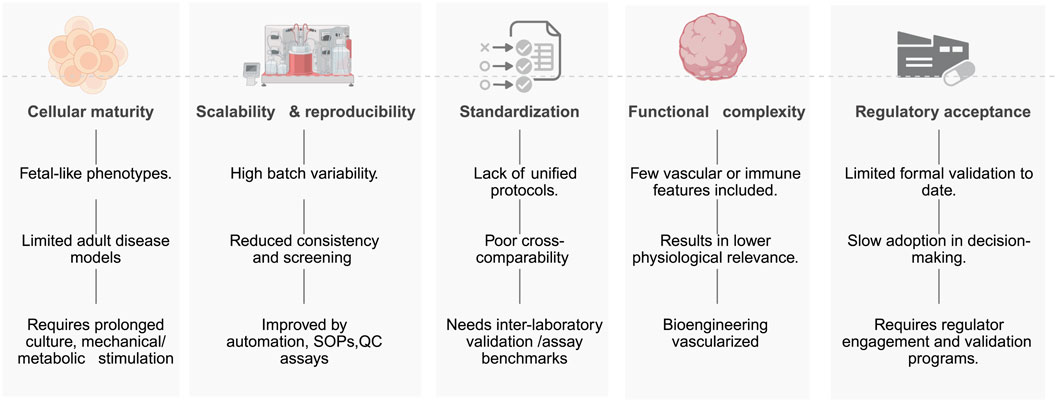
Figure 4. Barriers to implementation in iPSC-Based Toxicity Screening.
5 Conclusion
The advent of hiPSCs represents a revolutionary advancement in the field of preclinical safety assessment and pharmacological research. By enabling the reprogramming of human somatic cells into pluripotent stem cells, hiPSCs provide an ethically sound, renewable, and genetically matched cellular resource for drug discovery, toxicity testing, and disease modeling (Kleiman and Engle, 2021). This innovation significantly improves the predictive accuracy of organ-specific toxicities and individual patient responses, surpassing traditional animal models, which frequently fail to replicate human-specific biological processes and adverse reactions accurately (Tran et al., 2023).
Current hiPSC-based models have notably enhanced the efficiency and accuracy of high-throughput drug screening by offering the capability to differentiate into diverse cell lineages, including cardiomyocytes, hepatocytes, neurons, endothelial cells, and renal cells (Tristan et al., 2021). These cell models facilitate detailed mechanistic investigations and more precise efficacy and safety evaluations, thus streamlining the drug development pipeline and substantially reducing costs and development timelines. Particularly in toxicology, hiPSC-derived cellular systems have demonstrated their potential to identify toxic effects missed by conventional animal studies, thereby significantly reducing attrition rates in later stages of clinical trials (Olson et al., 2000).
Moreover, the integration of hiPSC technology with advanced complementary platforms such as 3D organoids, microfluidic organ-on-a-chip systems, and AI-driven analytics has dramatically broadened the scope and depth of preclinical studies (Yildirim et al., 2025). These technological synergies enable a closer simulation of complex human physiological environments, improved modeling of dynamic tissue-tissue interactions, and more accurate prediction of drug metabolism, pharmacokinetics, and pharmacodynamics (Jin et al., 2021). Such advancements are crucial for improving precision medicine and personalizing therapeutic interventions, highlighting the transformative potential of iPSC technology in both pharmaceutical innovation and personalized patient care.
Nevertheless, despite these substantial advancements, the full translational and clinical potential of hiPSC-based platforms faces persistent challenges. Key issues include inherent variability between hiPSC lines due to differences in genetic background, reprogramming methods, and clonal selection processes. Additionally, standardized differentiation and assay protocols remain limited, complicating cross-study comparisons and regulatory acceptance. Another critical obstacle is the immature phenotype of hiPSC-derived cells, which often exhibit fetal or neonatal characteristics rather than fully mature adult functionalities, thereby limiting their predictive capabilities for chronic toxicities and adult-onset diseases (Volpato et al., 2018; Yang et al., 2014; OECD, 2018; Yip et al., 2023; Sewell et al., 2024).
Addressing these challenges requires ongoing interdisciplinary collaboration, robust quality control measures, and the development of standardized, reproducible methodologies. Advances in automation, biomanufacturing, and real-time monitoring technologies are anticipated to alleviate issues related to scalability and reproducibility, further facilitating the broader adoption of hiPSC-derived platforms across academic and industrial research environments (Fritsche et al., 2021).
Regulatory frameworks must also evolve concurrently to fully integrate hiPSC-based models into standard drug safety assessment practices. Encouragingly, regulatory agencies like the FDA have already begun initiatives, such as the FDA Modernization Act 2.0, which advocate for the inclusion of advanced human-relevant methods to reduce animal testing (Yildirim et al., 2025; Ahmed et al., 2023). This regulatory support underscores the growing recognition of hiPSC technology as a scientifically credible and ethically preferable alternative within toxicology and drug discovery.
Building on this regulatory momentum, there is broad consensus that human-relevant, xeno-free alternatives are needed to address the poor translation of animal models and to operationalize the 3Rs (Tannenbaum and Bennett, 2015; Tanner, 2018). Accordingly, xeno-free hPSC and organoid systems are prioritized to reduce biosafety and variability and to meet GMP expectations for clinical translation (Rodríguez-Pizà et al., 2010). International bodies have established acceptance pathways for non-animal testing methods. Organization for Economic Co-operation and Development (OECD) Test Guidelines now include human-relevant alternatives, while European Union Reference Laboratory (EURL) -Reference Laboratory for Alternatives to Animal Testing (ECVAM) provides validation frameworks (Stephens et al., 2013). Collectively, these positions justify and accelerate adoption of xeno-free, human-based platforms for preclinical safety assessment and drug discovery.
In summary, hiPSC technology has become an indispensable asset in modern pharmacological research and toxicological evaluation, poised to significantly transform preclinical drug testing paradigms. Continued advancements in differentiation protocols, standardization efforts, and regulatory harmonization are vital to overcoming existing barriers and fully realizing the transformative potential of hiPSCs. Ultimately, the integration of hiPSC technology with emerging advanced technologies promises safer, more effective, and more personalized medical therapies, profoundly impacting public health outcomes and revolutionizing the pharmaceutical industry.
Author contributions
NC: Conceptualization, Data curation, Investigation, Formal Analysis, Writing – original draft, Writing – review and editing. LV-D: Conceptualization, Funding acquisition, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported with funding from NSF Awards 2026049, 2234541, and NIH Award 1R15CA254006-01A1.
Acknowledgements
Zotero reference manager (Roy Rosenzweig Center for History and New Media) was used to curate and import records from PubMed (NCBI, U.S. NLM/NIH), Google Scholar (Google LLC), and Scopus (Elsevier). Anara (formerly Unriddle; Anara Labs, Inc.) was used to support document analysis and integrate searches across the library. Figures were created and edited in BioRender (Science Suite Inc., dba BioRender). Generative AI tools—Gemini 2.5 Pro (Google DeepMind), ChatGPT Pro (OpenAI; GPT-5 access), and Quill Bot (Learneo)— were used for language editing and visual concept drafting. All AI-assisted outputs were reviewed by the authors for factual accuracy, originality, and adherence to ethical standards.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. Anara AI was used partially to obtain references.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahmed, HMMAM, and Moreira Teixeira, L. S. (2022). New Endeavors of (Micro)Tissue Engineering: Cells Tissues Organs on-Chip and Communication Thereof. Cells Tissues Organs 211 (6), 721–735. doi:10.1159/000516356
Ahmed, S. M., Shivnaraine, R. V., and Wu, J. C. (2023). FDA Modernization Act 2.0 Paves the Way to Computational Biology and Clinical Trials in a Dish. Circulation 148 (4), 309–311. doi:10.1161/CIRCULATIONAHA.123.065585
Badenes, S. M., Fernandes, T. G., Rodrigues, C. A. V., Diogo, M. M., and Cabral, J. M. S. (2014). “Scalable Expansion of Human-Induced Pluripotent Stem Cells in Xeno-Free Microcarriers,” in Stem Cells and Good Manufacturing Practices. Editor K. Turksen (New York, NY: Springer New York), 23–29. doi:10.1007/7651_2014_106
Badenes, S. M., Fernandes, T. G., Cordeiro, C. S. M., Boucher, S., Kuninger, D., Vemuri, M. C., et al. (2016). Defined Essential 8TM Medium and Vitronectin Efficiently Support Scalable Xeno-Free Expansion of Human Induced Pluripotent Stem Cells in Stirred Microcarrier Culture Systems. PLOS ONE 11 (3), e0151264. doi:10.1371/journal.pone.0151264
Bershteyn, M., Nowakowski, T. J., Pollen, A. A., Lullo, E. D., Nene, A., Wynshaw-Boris, A., et al. (2017). Human iPSC-Derived Cerebral Organoids Model Cellular Features of Lissencephaly and Reveal Prolonged Mitosis of Outer Radial Glia. Cell Stem Cell 20 (4), 435–449.e4. doi:10.1016/j.stem.2016.12.007
Bian, S., Repic, M., Guo, Z., Kavirayani, A., Burkard, T., Bagley, J. A., et al. (2018). Genetically engineered cerebral organoids model brain tumor formation. Nat. Methods 15 (8), 631–639. doi:10.1038/s41592-018-0070-7
Biological Industries. Press Release (2016). Biological Industries Announces FDA Drug Master File Acceptance For Nutristem hPSC XF Medium. Available online at: https://www.biospace.com/b-biological-industries-b-announces-fda-drug-master-file-acceptance-for-nutristem-hpsc-xf-medium.
Bowles, K. R., Silva, M. C., Whitney, K., Bertucci, T., Berlind, J. E., Lai, J. D., et al. (2021). ELAVL4, splicing, and glutamatergic dysfunction precede neuron loss in MAPT mutation cerebral organoids. Cell 184 (17), 4547–4563.e17. doi:10.1016/j.cell.2021.07.003
Braam, S. R., Zeinstra, L., Litjens, S., Ward-van Oostwaard, D., van den Brink, S., van Laake, L., et al. (2008). Recombinant vitronectin is a functionally defined substrate that supports human embryonic stem cell self-renewal via alphavbeta5 integrin. Stem Cells 26 (9), 2257–2265. doi:10.1634/stemcells.2008-0291
Brafman, D. A., Chang, C. W., Fernandez, A., Willert, K., Varghese, S., and Chien, S. (2010). Long-term human pluripotent stem cell self-renewal on synthetic polymer surfaces. Biomaterials 31 (34), 9135–9144. doi:10.1016/j.biomaterials.2010.08.007
Bray, M. A., Singh, S., Han, H., Davis, C. T., Borgeson, B., Hartland, C., et al. (2016). Cell Painting, a high-content image-based assay for morphological profiling using multiplexed fluorescent dyes. Nat. Protoc. 11 (9), 1757–1774. doi:10.1038/nprot.2016.105
Breunig, M., Merkle, J., Wagner, M., Melzer, M. K., Barth, T. F. E., Engleitner, T., et al. (2021). Modeling plasticity and dysplasia of pancreatic ductal organoids derived from human pluripotent stem cells. Cell Stem Cell 28 (6), 1105–1124.e19. doi:10.1016/j.stem.2021.03.005
Bulutoglu, B., Rey-Bedón, C., Mert, S., Tian, L., Jang, Y. Y., Yarmush, M. L., et al. (2020). “A comparison of hepato-cellular in vitro platforms to study CYP3A4 induction,”. PLOS ONE. Editor K. Aalto-Setala, 15.
Celiz, A. D., Smith, J. G. W., Langer, R., Anderson, D. G., Winkler, D. A., Barrett, D. A., et al. (2014a). Materials for stem cell factories of the future. Nat. Mater 13 (6), 570–579. doi:10.1038/nmat3972
Celiz, A. D., Smith, J. G. W., Patel, A. K., Langer, R., Anderson, D. G., Barrett, D. A., et al. (2014b). Chemically diverse polymer microarrays and high throughput surface characterisation: a method for discovery of materials for stem cell culture†Electronic supplementary information (ESI). Biomater. Sci. 2 (11), 1604–1611. doi:10.1039/c4bm00054d
Chandy, M., Obal, D., and Wu, J. C. (2022). Elucidating effects of environmental exposure using human-induced pluripotent stem cell disease modeling. EMBO Mol. Med. 14 (11), e13260. doi:10.15252/emmm.202013260
Chen, G., Gulbranson, D. R., Hou, Z., Bolin, J. M., Ruotti, V., Probasco, M. D., et al. (2011). Chemically defined conditions for human iPSC derivation and culture. Nat. Methods 8 (5), 424–429. doi:10.1038/nmeth.1593
Chen, Y. M., Chen, L. H., Li, M. P., Li, H. F., Higuchi, A., Kumar, S. S., et al. (2017). Xeno-free culture of human pluripotent stem cells on oligopeptide-grafted hydrogels with various molecular designs. Sci. Rep. 7 (1), 45146. doi:10.1038/srep45146
Cho, Y., Lee, H., Jeong, W., Jung, K. B., Lee, S. Y., Park, S., et al. (2024). Long-Term Culture of Human Pluripotent Stem Cells in Xeno-Free Condition Using Functional Polymer Films. Adv. Mater 17, 2403952. doi:10.1002/adma.202403952
Cobo, F., Stacey, G. N., Hunt, C., Cabrera, C., Nieto, A., Montes, R., et al. (2005). Microbiological control in stem cell banks: approaches to standardisation. Appl. Microbiol. Biotechnol. 68 (4), 456–466. doi:10.1007/s00253-005-0062-2
Crespo, M., Vilar, E., Tsai, S. Y., Chang, K., Amin, S., Srinivasan, T., et al. (2017). Colonic organoids derived from human induced pluripotent stem cells for modeling colorectal cancer and drug testing. Nat. Med. 23 (7), 878–884. doi:10.1038/nm.4355
Da Cruz, L., Fynes, K., Georgiadis, O., Kerby, J., Luo, Y. H., Ahmado, A., et al. (2018). Phase 1 clinical study of an embryonic stem cell–derived retinal pigment epithelium patch in age-related macular degeneration. Nat. Biotechnol. 36 (4), 328–337. doi:10.1038/nbt.4114
Dakhore, S., Nayer, B., and Hasegawa, K. (2018). Human Pluripotent Stem Cell Culture: Current Status, Challenges, and Advancement. Stem Cells Int. 2018, 7396905–7396917. doi:10.1155/2018/7396905
Demchenko, A., Kondrateva, E., Tabakov, V., Efremova, A., Salikhova, D., Bukharova, T., et al. (2023). Airway and Lung Organoids from Human-Induced Pluripotent Stem Cells Can Be Used to Assess CFTR Conductance. Int. J. Mol. Sci. 24 (7), 6293. doi:10.3390/ijms24076293
Deng, Y., Zhang, X., Zhao, X., Li, Q., Ye, Z., Li, Z., et al. (2013). Long-term self-renewal of human pluripotent stem cells on peptide-decorated poly(OEGMA-co-HEMA) brushes under fully defined conditions. Acta Biomater. 9 (11), 8840–8850. doi:10.1016/j.actbio.2013.07.017
Eiselleova, L., Peterkova, I., Neradil, J., Slaninova, I., Hampl, A., and Dvorak, P. (2008). Comparative study of mouse and human feeder cells for human embryonic stem cells. Int. J. Dev. Biol. 52 (4), 353–363. doi:10.1387/ijdb.082590le
Elanzew, A., Nießing, B., Langendoerfer, D., Rippel, O., Piotrowski, T., Schenk, F., et al. (2020). The StemCellFactory: A Modular System Integration for Automated Generation and Expansion of Human Induced Pluripotent Stem Cells. Front. Bioeng. Biotechnol. 8, 580352. doi:10.3389/fbioe.2020.580352
Ellis, B. W., Acun, A., Can, U. I., and Zorlutuna, P. (2017). Human iPSC-derived myocardium-on-chip with capillary-like flow for personalized medicine. Biomicrofluidics 11 (2), 024105. doi:10.1063/1.4978468
Englund, M. C. O., Caisander, G., Noaksson, K., Emanuelsson, K., Lundin, K., Bergh, C., et al. (2010). The establishment of 20 different human embryonic stem cell lines and subclones; a report on derivation, culture, characterisation and banking. Vitro Cell Dev. Biol. - Anim. 46 (3–4), 217–230. doi:10.1007/s11626-010-9289-z
Faulkner-Jones, A., Fyfe, C., Cornelissen, D. J., Gardner, J., King, J., Courtney, A., et al. (2015). Bioprinting of human pluripotent stem cells and their directed differentiation into hepatocyte-like cells for the generation of mini-livers in 3D. Biofabrication 7 (4), 044102. doi:10.1088/1758-5090/7/4/044102
Forghani, A., Kriegh, L., Hogan, K., Chen, C., Brewer, G., Tighe, T. B., et al. (2017). Fabrication and characterization of cell sheets using methylcellulose and PNIPAAm thermoresponsive polymers: A comparison Study. J. Biomed. Mater Res. A 105 (5), 1346–1354. doi:10.1002/jbm.a.36014
Fransen, L. F. H., and Leonard, M. O. (2023). Induced pluripotent and CD34+ stem cell derived myeloid cells display differential responses to particle and dust mite exposure. Sci. Rep. 13 (1), 9375. doi:10.1038/s41598-023-36508-3
Fritsche, E., Tigges, J., Hartmann, J., Kapr, J., Serafini, M. M., and Viviani, B. (2020). “Neural In Vitro Models for Studying Substances Acting on the Central Nervous System,” in Organotypic Models in Drug Development. Editors M. Schäfer-Korting, S. Stuchi Maria-Engler, and R. Landsiedel (Cham: Springer International Publishing), 111–141. doi:10.1007/164_2020_367
Fritsche, E., Haarmann-Stemmann, T., Kapr, J., Galanjuk, S., Hartmann, J., Mertens, P. R., et al. (2021). Stem Cells for Next Level Toxicity Testing in the 21st Century. Small 17 (15), 2006252. doi:10.1002/smll.202006252
Gabriel, E., Wason, A., Ramani, A., Gooi, L. M., Keller, P., Pozniakovsky, A., et al. (2016). CPAP promotes timely cilium disassembly to maintain neural progenitor pool. EMBO J. 35 (8), 803–819. doi:10.15252/embj.201593679
Garcez, P. P., Loiola, E. C., Madeiro Da Costa, R., Higa, L. M., Trindade, P., Delvecchio, R., et al. (2016). Zika virus impairs growth in human neurospheres and brain organoids. Science. 352 (6287), 816–818. doi:10.1126/science.aaf6116
Grafton, F., Ho, J., Ranjbarvaziri, S., Farshidfar, F., Budan, A., Steltzer, S., et al. (2021). Deep learning detects cardiotoxicity in a high-content screen with induced pluripotent stem cell-derived cardiomyocytes. eLife 10, e68714. doi:10.7554/eLife.68714
Grimm, F. A., Iwata, Y., Sirenko, O., Bittner, M., and Rusyn, I. (2015). High-Content Assay Multiplexing for Toxicity Screening in Induced Pluripotent Stem Cell-Derived Cardiomyocytes and Hepatocytes. Assay. Drug Dev. Technol. 13 (9), 529–546. doi:10.1089/adt.2015.659
Guan, Y., Enejder, A., Wang, M., Fang, Z., Cui, L., Chen, S. Y., et al. (2021). A human multi-lineage hepatic organoid model for liver fibrosis. Nat. Commun. 12 (1), 6138. doi:10.1038/s41467-021-26410-9
Gurevich, I., Burton, S. A., Munn, C., Ohshima, M., Goedland, M. E., Czysz, K., et al. (2020). iPSC-derived hepatocytes generated from NASH donors provide a valuable platform for disease modeling and drug discovery. Biol. Open 1, 055087. doi:10.1242/bio.055087
Hahn, J., Aksoy, E., Hamad, S., Kuckelkorn, C., Gómez Montoya, A., Ritter, M., et al. (2025). Mass Production of Uniform Embryoid Bodies by Acoustic Standing Waves. Small Methods 11, e01283. doi:10.1002/smtd.202501283
Hanna, J., Cheng, A. W., Saha, K., Kim, J., Lengner, C. J., Soldner, F., et al. (2010). Human embryonic stem cells with biological and epigenetic characteristics similar to those of mouse ESCs. Proc. Natl. Acad. Sci. 107 (20), 9222–9227. doi:10.1073/pnas.1004584107
Hatton, I. A., Galbraith, E. D., Merleau, N. S. C., Miettinen, T. P., Smith, B. M., and Shander, J. A. (2025). The human cell count and size distribution. Proc. Natl. Acad. Sci. U. S. A. 120 (39), e2303077120. doi:10.1073/pnas.2303077120
Haupt, S., Grützner, J., Thier, M., Kallweit, T., Rath, B. H., Laufenberg, I., et al. (2012). Automated selection and harvesting of pluripotent stem cell colonies. Biotechnol. Appl. Biochem. 59 (2), 77–87. doi:10.1002/bab.1014
Herland, A., Maoz, B. M., Das, D., Somayaji, M. R., Prantil-Baun, R., Novak, R., et al. (2020). Quantitative prediction of human pharmacokinetic responses to drugs via fluidically coupled vascularized organ chips. Nat. Biomed. Eng. 4 (4), 421–436. doi:10.1038/s41551-019-0498-9
Hernandez, J. O. R., Wang, X., Vazquez-Segoviano, M., Lopez-Marfil, M., Sobral-Reyes, M. F., Moran-Horowich, A., et al. (2021). A tissue-bioengineering strategy for modeling rare human kidney diseases in vivo. Nat. Commun. 12 (1), 6496. doi:10.1038/s41467-021-26596-y
Higuchi, A., Kao, S. H., Ling, Q. D., Chen, Y. M., Li, H. F., Alarfaj, A. A., et al. (2015). Long-term xeno-free culture of human pluripotent stem cells on hydrogels with optimal elasticity. Sci. Rep. 5 (1), 18136. doi:10.1038/srep18136
Hockemeyer, D., and Jaenisch, R. (2016). Induced Pluripotent Stem Cells Meet Genome Editing. Cell Stem Cell 18 (5), 573–586. doi:10.1016/j.stem.2016.04.013
Hohwieler, M., Illing, A., Hermann, P. C., Mayer, T., Stockmann, M., Perkhofer, L., et al. (2017). Human pluripotent stem cell-derived acinar/ductal organoids generate human pancreas upon orthotopic transplantation and allow disease modelling. Gut 66 (3), 473–486. doi:10.1136/gutjnl-2016-312423
Hor, J. H., Soh, E. S. Y., Tan, L. Y., Lim, V. J. W., Santosa, M. M., Ho, B. X., et al. (2018). Cell cycle inhibitors protect motor neurons in an organoid model of Spinal Muscular Atrophy. Cell Death Dis. 9 (11), 1100–1112. doi:10.1038/s41419-018-1081-0
Hughes, C. S., Postovit, L. M., and Lajoie, G. A. (2010). Matrigel: A complex protein mixture required for optimal growth of cell culture. PROTEOMICS 10 (9), 1886–1890. doi:10.1002/pmic.200900758
Huh, D., Matthews, B. D., Mammoto, A., Montoya-Zavala, M., Hsin, H. Y., and Ingber, D. E. (2010). Reconstituting Organ-Level Lung Functions on a Chip. Science 328 (5986), 1662–1668. doi:10.1126/science.1188302
Ichisima, J., Suzuki, N. M., Samata, B., Awaya, T., Takahashi, J., Hagiwara, M., et al. (2019). Verification and rectification of cell type-specific splicing of a Seckel syndrome-associated ATR mutation using iPS cell model. J. Hum. Genet. 64 (5), 445–458. doi:10.1038/s10038-019-0574-8
Ieyasu, A., Ishida, R., Kimura, T., Morita, M., Wilkinson, A. C., Sudo, K., et al. (2017). An All-Recombinant Protein-Based Culture System Specifically Identifies Hematopoietic Stem Cell Maintenance Factors. Stem Cell Rep. 8 (3), 500–508. doi:10.1016/j.stemcr.2017.01.015
Irwin, E. F., Gupta, R., Dashti, D. C., and Healy, K. E. (2011). Engineered polymer-media interfaces for the long-term self-renewal of human embryonic stem cells. Biomaterials 32 (29), 6912–6919. doi:10.1016/j.biomaterials.2011.05.058
Jacob, A., Morley, M., Hawkins, F., McCauley, K. B., Jean, J. C., Heins, H., et al. (2017). Differentiation of Human Pluripotent Stem Cells into Functional Lung Alveolar Epithelial Cells. Cell Stem Cell 21 (4), 472–488.e10. doi:10.1016/j.stem.2017.08.014
James, D., Levine, A. J., Besser, D., and Hemmati-Brivanlou, A. (2005). TGFbeta/activin/nodal signaling is necessary for the maintenance of pluripotency in human embryonic stem cells. Development 132 (6), 1273–1282. doi:10.1242/dev.01706
James, O. G., Selvaraj, B. T., Magnani, D., Burr, K., Connick, P., Barton, S. K., et al. (2021). iPSC-derived myelinoids to study myelin biology of humans. Dev. Cell 56 (9), 1346–1358.e6. doi:10.1016/j.devcel.2021.04.006
Jin, M., Yi, X., Liao, W., Chen, Q., Yang, W., Li, Y., et al. (2021). Advancements in stem cell-derived hepatocyte-like cell models for hepatotoxicity testing. Stem Cell Res. Ther. 12 (1), 84. doi:10.1186/s13287-021-02152-9
Joutsijoki, H., Haponen, M., Baldin, I., Rasku, J., Gizatdinova, Y., Paci, M., et al. (2014). “Histogram-based classification of iPSC colony images using machine learning methods,” in 2014 IEEE International Conference on Systems, Man, and Cybernetics (SMC) (San Diego, CA, USA: IEEE), 2611–2617. Available online at: https://ieeexplore.ieee.org/document/6974321.
Kadoshima, T., Sakaguchi, H., Nakano, T., Soen, M., Ando, S., Eiraku, M., et al. (2013). Self-organization of axial polarity, inside-out layer pattern, and species-specific progenitor dynamics in human ES cell–derived neocortex. Proc. Natl. Acad. Sci. 110 (50), 20284–20289. doi:10.1073/pnas.1315710110
Kang, Y., Zhou, Y., Li, Y., Han, Y., Xu, J., Niu, W., et al. (2021). A human forebrain organoid model of fragile X syndrome exhibits altered neurogenesis and highlights new treatment strategies. Nat. Neurosci. 24 (10), 1377–1391. doi:10.1038/s41593-021-00913-6
Kanton, S., Boyle, M. J., He, Z., Santel, M., Weigert, A., Sanchís-Calleja, F., et al. (2019). Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature 574 (7778), 418–422. doi:10.1038/s41586-019-1654-9
Kato, R., Matsumoto, M., Sasaki, H., Joto, R., Okada, M., Ikeda, Y., et al. (2016). Parametric analysis of colony morphology of non-labelled live human pluripotent stem cells for cell quality control. Sci. Rep. 6 (1), 34009. doi:10.1038/srep34009
Kawase, E., and Nakatsuji, N. (2023). Development of substrates for the culture of human pluripotent stem cells. Biomater. Sci. 11 (9), 2974–2987. doi:10.1039/d2bm01473d
Kikuchi, T., Kino-oka, M., Wada, M., Kobayashi, T., Kato, M., Takeda, S., et al. (2018). A novel, flexible and automated manufacturing facility for cell-based health care products: Tissue Factory. Regen. Ther. 9, 89–99. doi:10.1016/j.reth.2018.08.004
Kim, J., Sachdev, P., and Sidhu, K. (2013). Alginate microcapsule as a 3D platform for the efficient differentiation of human embryonic stem cells to dopamine neurons. Stem Cell Res. 11 (3), 978–989. doi:10.1016/j.scr.2013.06.005
Kim, J., Im, I., Kim, H., Jeon, J. S., Kang, E., Jo, S., et al. (2020). Live-cell screening platform using human-induced pluripotent stem cells expressing fluorescence-tagged cytochrome P450 1A1. FASEB J. 34 (7), 9141–9155. doi:10.1096/fj.201903110R
Kleiman, R. J., and Engle, S. J. (2021). Human inducible pluripotent stem cells: Realization of initial promise in drug discovery. Cell Stem Cell 28 (9), 1507–1515. doi:10.1016/j.stem.2021.08.002
Kleinman, H. K., McGarvey, M. L., Hassell, J. R., Star, V. L., Cannon, F. B., Laurie, G. W., et al. (1986). Basement membrane complexes with biological activity. Biochemistry 25 (2), 312–318. doi:10.1021/bi00350a005
Klim, J. R., Li, L., Wrighton, P. J., Piekarczyk, M. S., and Kiessling, L. L. (2010). A defined glycosaminoglycan-binding substratum for human pluripotent stem cells. Nat. Methods. 7 (12), 989–994. doi:10.1038/nmeth.1532
Krawetz, R., Taiani, J. T., Liu, S., Meng, G., Li, X., Kallos, M. S., et al. (2010). Large-Scale Expansion of Pluripotent Human Embryonic Stem Cells in Stirred-Suspension Bioreactors. Tissue Eng. Part C Methods 16 (4), 573–582. doi:10.1089/ten.TEC.2009.0228
Kuraoka, S., Tanigawa, S., Taguchi, A., Hotta, A., Nakazato, H., Osafune, K., et al. (2020). PKD1-Dependent Renal Cystogenesis in Human Induced Pluripotent Stem Cell-Derived Ureteric Bud/Collecting Duct Organoids. J. Am. Soc. Nephrol. 31 (10), 2355–2371. doi:10.1681/ASN.2020030378
Lancaster, M. A., Renner, M., Martin, C. A., Wenzel, D., Bicknell, L. S., Hurles, M. E., et al. (2013). Cerebral organoids model human brain development and microcephaly. Nature 501 (7467), 373–379. doi:10.1038/nature12517
Lane, A., Jovanovic, K., Shortall, C., Ottaviani, D., Panes, A. B., Schwarz, N., et al. (2020). Modeling and Rescue of RP2 Retinitis Pigmentosa Using iPSC-Derived Retinal Organoids. Stem Cell Rep. 15 (1), 67–79. doi:10.1016/j.stemcr.2020.05.007
Lawrence, M. L., Elhendawi, M., Morlock, M., Liu, W., Liu, S., Palakkan, A., et al. (2022). Human iPSC-derived renal organoids engineered to report oxidative stress can predict drug-induced toxicity. iScience 25 (3), 103884. doi:10.1016/j.isci.2022.103884
Lee, Y., Lee, H. J., Ham, S., Jeong, D., Lee, M., Lee, U., et al. (2022). Plant-derived human recombinant growth factors and serum albumin maintain stemness of human-induced pluripotent stem cells. Cell Biol. Int. 46 (1), 139–147. doi:10.1002/cbin.11715
Leibel, S. L., Winquist, A., Tseu, I., Wang, J., Luo, D., Shojaie, S., et al. (2019). Reversal of Surfactant Protein B Deficiency in Patient Specific Human Induced Pluripotent Stem Cell Derived Lung Organoids by Gene Therapy. Sci. Rep. 9 (1), 13450. doi:10.1038/s41598-019-49696-8
Liang, Y., Zhang, H., Feng, Q. S., Cai, M. B., Deng, W., Qin, D., et al. (2013). The propensity for tumorigenesis in human induced pluripotent stem cells is related with genomic instability. Chin. J. Cancer 32 (4), 205–212. doi:10.5732/cjc.012.10065
Liu, G., David, B. T., Trawczynski, M., and Fessler, R. G. (2020). Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem Cell Rev. Rep. 16 (1), 3–32. doi:10.1007/s12015-019-09935-x
Ludwig, T. E., Levenstein, M. E., Jones, J. M., Berggren, W. T., Mitchen, E. R., Frane, J. L., et al. (2006a). Derivation of human embryonic stem cells in defined conditions. Nat. Biotechnol. 24 (2), 185–187. doi:10.1038/nbt1177
Ludwig, T. E., Bergendahl, V., Levenstein, M. E., Yu, J., Probasco, M. D., and Thomson, J. A. (2006b). Feeder-independent culture of human embryonic stem cells. Nat. Methods 3 (8), 637–646. doi:10.1038/nmeth902
Maffioletti, S. M., Sarcar, S., Henderson, A. B. H., Mannhardt, I., Pinton, L., Moyle, L. A., et al. (2018). Three-Dimensional Human iPSC-Derived Artificial Skeletal Muscles Model Muscular Dystrophies and Enable Multilineage Tissue Engineering. Cell Rep. 23 (3), 899–908. doi:10.1016/j.celrep.2018.03.091
Majmundar, A. J., Buerger, F., Forbes, T. A., Klämbt, V., Schneider, R., Deutsch, K., et al. (2021). Recessive NOS1AP variants impair actin remodeling and cause glomerulopathy in humans and mice. Sci. Adv. 7 (1), eabe1386. doi:10.1126/sciadv.abe1386
Mariani, J., Coppola, G., Zhang, P., Abyzov, A., Provini, L., Tomasini, L., et al. (2015). FOXG1-Dependent Dysregulation of GABA/Glutamate Neuron Differentiation in Autism Spectrum Disorders. Cell 162 (2), 375–390. doi:10.1016/j.cell.2015.06.034
Martin, M. J., Muotri, A., Gage, F., and Varki, A. (2005). Human embryonic stem cells express an immunogenic nonhuman sialic acid. Nat. Med. 11 (2), 228–232. doi:10.1038/nm1181
Mazzara, P. G., Muggeo, S., Luoni, M., Massimino, L., Zaghi, M., Valverde, P. T. T., et al. (2020). Frataxin gene editing rescues Friedreich’s ataxia pathology in dorsal root ganglia organoid-derived sensory neurons. Nat. Commun. 11 (1), 4178. doi:10.1038/s41467-020-17954-3
McCauley, K. B., Hawkins, F., Serra, M., Thomas, D. C., Jacob, A., and Kotton, D. N. (2017). Efficient Derivation of Functional Human Airway Epithelium from Pluripotent Stem Cells via Temporal Regulation of Wnt Signaling. Cell Stem Cell 20 (6), 844–857.e6. doi:10.1016/j.stem.2017.03.001
McCracken, K. W., Catá, E. M., Crawford, C. M., Sinagoga, K. L., Schumacher, M., Rockich, B. E., et al. (2014). Modelling human development and disease in pluripotent stem-cell-derived gastric organoids. Nature 516 (7531), 400–404. doi:10.1038/nature13863
Mei, Y., Saha, K., Bogatyrev, S. R., Yang, J., Hook, A. L., Kalcioglu, Z. I., et al. (2010). Combinatorial development of biomaterials for clonal growth of human pluripotent stem cells. Nat. Mater. 9 (9), 768–778. doi:10.1038/nmat2812
Melkoumian, Z., Weber, J. L., Weber, D. M., Fadeev, A. G., Zhou, Y., Dolley-Sonneville, P., et al. (2010). Synthetic peptide-acrylate surfaces for long-term self-renewal and cardiomyocyte differentiation of human embryonic stem cells. Nat. Biotechnol. 28 (6), 606–610. doi:10.1038/nbt.1629
Mellios, N., Feldman, D. A., Sheridan, S. D., Ip, J. P. K., Kwok, S., Amoah, S. K., et al. (2018). MeCP2-regulated miRNAs control early human neurogenesis through differential effects on ERK and AKT signaling. Mol. Psychiatry 23 (4), 1051–1065. doi:10.1038/mp.2017.86
Meyer, K., Feldman, H. M., Lu, T., Drake, D., Lim, E. T., Ling, K. H., et al. (2019). REST and Neural Gene Network Dysregulation in iPSC Models of Alzheimer’s Disease. Cell Rep. 26 (5), 1112–1127.e9. doi:10.1016/j.celrep.2019.01.023
Mikkola, M., Olsson, C., Palgi, J., Ustinov, J., Palomaki, T., Horelli-Kuitunen, N., et al. (2006). Distinct differentiation characteristics of individual human embryonic stem cell lines. BMC Dev. Biol. 6 (1), 40. doi:10.1186/1471-213X-6-40
Mills, R. J., Titmarsh, D. M., Koenig, X., Parker, B. L., Ryall, J. G., Quaife-Ryan, G. A., et al. (2017). Functional screening in human cardiac organoids reveals a metabolic mechanism for cardiomyocyte cell cycle arrest. Proc. Natl. Acad. Sci. 114 (40), E8372–E8381. doi:10.1073/pnas.1707316114
Miyazaki, T., Futaki, S., Hasegawa, K., Kawasaki, M., Sanzen, N., Hayashi, M., et al. (2008). Recombinant human laminin isoforms can support the undifferentiated growth of human embryonic stem cells. Biochem. Biophys. Res. Commun. 375 (1), 27–32. doi:10.1016/j.bbrc.2008.07.111
Miyazaki, T., Futaki, S., Suemori, H., Taniguchi, Y., Yamada, M., Kawasaki, M., et al. (2012). Laminin E8 fragments support efficient adhesion and expansion of dissociated human pluripotent stem cells. Nat. Commun. 3 (1), 1236. doi:10.1038/ncomms2231
Mohr, J. C., De Pablo, J. J., and Palecek, S. P. (2006). 3-D microwell culture of human embryonic stem cells. Biomaterials 27 (36), 6032–6042. doi:10.1016/j.biomaterials.2006.07.012
Mohr, J. C., Zhang, J., Azarin, S. M., Soerens, A. G., De Pablo, J. J., Thomson, J. A., et al. (2010). The microwell control of embryoid body size in order to regulate cardiac differentiation of human embryonic stem cells. Biomaterials 31 (7), 1885–1893. doi:10.1016/j.biomaterials.2009.11.033
Nabi, S. U., Ali, S. I., Rather, M. A., Sheikh, W. M., Altaf, M., Singh, H., et al. (2022). Organoids: A new approach in toxicity testing of nanotherapeutics. J. Appl. Toxicol. 42 (1), 52–72. doi:10.1002/jat.4206
Nagaoka, M., Si-Tayeb, K., Akaike, T., and Duncan, S. A. (2010). Culture of human pluripotent stem cells using completely defined conditions on a recombinant E-cadherin substratum. BMC Dev. Biol. 10 (1), 60. doi:10.1186/1471-213X-10-60
Nakagawa, M., Taniguchi, Y., Senda, S., Takizawa, N., Ichisaka, T., Asano, K., et al. (2014). A novel efficient feeder-free culture system for the derivation of human induced pluripotent stem cells. Sci. Rep. 4 (1), 3594. doi:10.1038/srep03594
Narkilahti, S., Rajala, K., Pihlajamäki, H., Suuronen, R., Hovatta, O., and Skottman, H. (2007). Monitoring and analysis of dynamic growth of human embryonic stem cells: comparison of automated instrumentation and conventional culturing methods. Biomed. Eng. OnLine 6 (1), 11. doi:10.1186/1475-925X-6-11
Nasir, A., Thorpe, J., Burroughs, L., Meurs, J., Pijuan-Galito, S., Irvine, D. J., et al. (2021). Discovery of a Novel Polymer for Xeno-Free, Long-Term Culture of Human Pluripotent Stem Cell Expansion. Adv. Healthc. Mater 10 (6), 2001448. doi:10.1002/adhm.202001448
OECD (2018). Guidance Document on Good In Vitro Method Practices (GIVIMP). OECD. Available online at: https://www.oecd.org/en/publications/guidance-document-on-good-in-vitro-method-practices-givimp_9789264304796-en.html.
Ogawa, M., Jiang, J. X., Xia, S., Yang, D., Ding, A., Laselva, O., et al. (2021). Generation of functional ciliated cholangiocytes from human pluripotent stem cells. Nat. Commun. 12 (1), 6504. doi:10.1038/s41467-021-26764-0
Oh, S. K. W., Chen, A. K., Mok, Y., Chen, X., Lim, U. M., Chin, A., et al. (2009). Long-term microcarrier suspension cultures of human embryonic stem cells. Stem Cell Res. 2 (3), 219–230. doi:10.1016/j.scr.2009.02.005
Olson, H., Betton, G., Robinson, D., Thomas, K., Monro, A., Kolaja, G., et al. (2000). Concordance of the Toxicity of Pharmaceuticals in Humans and in Animals. Regul. Toxicol. Pharmacol. 32 (1), 56–67. doi:10.1006/rtph.2000.1399
Osaki, T., Uzel, S. G. M., and Kamm, R. D. (2018). Microphysiological 3D model of amyotrophic lateral sclerosis (ALS) from human iPS-derived muscle cells and optogenetic motor neurons. Sci. Adv. 4 (10), eaat5847. doi:10.1126/sciadv.aat5847
Otsuji, T. G., Bin, J., Yoshimura, A., Tomura, M., Tateyama, D., Minami, I., et al. (2014). A 3D Sphere Culture System Containing Functional Polymers for Large-Scale Human Pluripotent Stem Cell Production. Stem Cell Rep. 2 (5), 734–745. doi:10.1016/j.stemcr.2014.03.012
Ozawa, H., Matsumoto, T., and Nakagawa, M. (2023). Culturing human pluripotent stem cells for regenerative medicine. Expert Opin. Biol. Ther. 23 (6), 479–489. doi:10.1080/14712598.2023.2225701
Park, H. J., Yang, K., Kim, M. J., Jang, J., Lee, M., Kim, D. W., et al. (2015). Bio-inspired oligovitronectin-grafted surface for enhanced self-renewal and long-term maintenance of human pluripotent stem cells under feeder-free conditions. Biomaterials 50, 127–139. doi:10.1016/j.biomaterials.2015.01.015
Park, S. E., Georgescu, A., and Huh, D. (2019). Organoids-on-a-chip. Science 364 (6444), 960–965. doi:10.1126/science.aaw7894
Park, S., Gwon, Y., Khan, S. A., Jang, K. J., and Kim, J. (2023). Engineering considerations of iPSC-based personalized medicine. Biomater. Res. 27 (1), 67. doi:10.1186/s40824-023-00382-x
Paşca, A. M., Sloan, S. A., Clarke, L. E., Tian, Y., Makinson, C. D., Huber, N., et al. (2015). Functional cortical neurons and astrocytes from human pluripotent stem cells in 3D culture. Nat. Methods 12 (7), 671–678. doi:10.1038/nmeth.3415
Pellegrini, L., Bonfio, C., Chadwick, J., Begum, F., Skehel, M., and Lancaster, M. A. (2020). Human CNS barrier-forming organoids with cerebrospinal fluid production. Science 369 (6500), eaaz5626. doi:10.1126/science.aaz5626
Peng, B. Y., Singh, A. K., Tsai, C. Y., Chan, C. H., Deng, Y. H., Wu, C. M., et al. (2023). Platelet-derived biomaterial with hyaluronic acid alleviates temporal-mandibular joint osteoarthritis: clinical trial from dish to human. J. Biomed. Sci. 30 (1), 77. doi:10.1186/s12929-023-00962-y
Pietrobon, A., Yockell-Lelièvre, J., Flood, T. A., and Stanford, W. L. (2022). Renal organoid modeling of tuberous sclerosis complex reveals lesion features arise from diverse developmental processes. Cell Rep. 40 (1), 111048. doi:10.1016/j.celrep.2022.111048
Powell, K. A., Bohrer, L. R., Stone, N. E., Hittle, B., Anfinson, K. R., Luangphakdy, V., et al. (2023). Automated human induced pluripotent stem cell colony segmentation for use in cell culture automation applications. SLAS Technol. 28 (6), 416–422. doi:10.1016/j.slast.2023.07.004
Rivera-Ordaz, A., Peli, V., Manzini, P., Barilani, M., and Lazzari, L. (2021). Critical Analysis of cGMP Large-Scale Expansion Process in Bioreactors of Human Induced Pluripotent Stem Cells in the Framework of Quality by Design. BioDrugs 35 (6), 693–714. doi:10.1007/s40259-021-00503-9
Rodin, S., Domogatskaya, A., Ström, S., Hansson, E. M., Chien, K. R., Inzunza, J., et al. (2010). Long-term self-renewal of human pluripotent stem cells on human recombinant laminin-511. Nat. Biotechnol. 28 (6), 611–615. doi:10.1038/nbt.1620
Rodin, S., Antonsson, L., Hovatta, O., and Tryggvason, K. (2014). Monolayer culturing and cloning of human pluripotent stem cells on laminin-521–based matrices under xeno-free and chemically defined conditions. Nat. Protoc. 9 (10), 2354–2368. doi:10.1038/nprot.2014.159
Rodrigues, A., Slembrouck-Brec, A., Nanteau, C., Terray, A., Tymoshenko, Y., Zagar, Y., et al. (2022). Modeling PRPF31 retinitis pigmentosa using retinal pigment epithelium and organoids combined with gene augmentation rescue. Npj Regen. Med. 7 (1), 39. doi:10.1038/s41536-022-00235-6
Rodríguez-Pizà, I., Richaud-Patin, Y., Vassena, R., González, F., Barrero, M. J., Veiga, A., et al. (2010). Reprogramming of Human Fibroblasts to Induced Pluripotent Stem Cells under Xeno-free Conditions. Stem Cells 28 (1), 36–44. doi:10.1002/stem.248
Romero-Morales, A. I., Robertson, G. L., Rastogi, A., Rasmussen, M. L., Temuri, H., McElroy, G. S., et al. (2022). Human iPSC-derived cerebral organoids model features of Leigh syndrome and reveal abnormal corticogenesis. Development 149 (20), dev199914. doi:10.1242/dev.199914
Ronneberger, O., Fischer, P., and Brox, T. (2015). “U-Net: Convolutional Networks for Biomedical Image Segmentation,” in Medical Image Computing and Computer-Assisted Intervention – MICCAI 2015. Editors N. Navab, J. Hornegger, W. M. Wells, and A. F. Frangi (Cham: Springer International Publishing), 234–241.
Sala, L., Van Meer, B. J., Tertoolen, L. G. J., Bakkers, J., Bellin, M., Davis, R. P., et al. (2018). MUSCLEMOTION: A Versatile Open Software Tool to Quantify Cardiomyocyte and Cardiac Muscle Contraction In Vitro and In Vivo. Circ. Res. 122 (3), e5–e16. doi:10.1161/CIRCRESAHA.117.312067
Sampaziotis, F., Cardoso de Brito, M., Madrigal, P., Bertero, A., Saeb-Parsy, K., Soares, F. A. C., et al. (2015). Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation. Nat. Biotechnol. 33 (8), 845–852. doi:10.1038/nbt.3275
Sayed, N., Liu, C., Ameen, M., Himmati, F., Zhang, J. Z., Khanamiri, S., et al. (2020). Clinical trial in a dish using iPSCs shows lovastatin improves endothelial dysfunction and cellular cross-talk in LMNA cardiomyopathy. Sci. Transl. Med. 12 (554), eaax9276. doi:10.1126/scitranslmed.aax9276
Schwedhelm, I., Zdzieblo, D., Appelt-Menzel, A., Berger, C., Schmitz, T., Schuldt, B., et al. (2019). Automated real-time monitoring of human pluripotent stem cell aggregation in stirred tank reactors. Sci. Rep. 9 (1), 12297. doi:10.1038/s41598-019-48814-w
Sen, A., Kallos, M. S., and Behie, L. A. (2002). Expansion of mammalian neural stem cells in bioreactors: effect of power input and medium viscosity. Dev. Brain Res. 134 (1–2), 103–113. doi:10.1016/s0165-3806(01)00328-5
Seok, J., Warren, H. S., Cuenca, A. G., Mindrinos, M. N., Baker, H. V., Xu, W., et al. (2013). Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. 110 (9), 3507–3512. doi:10.1073/pnas.1222878110
Sewell, F., Alexander-White, C., Brescia, S., Currie, R. A., Roberts, R., Roper, C., et al. (2024). New approach methodologies (NAMs): identifying and overcoming hurdles to accelerated adoption. Toxicol. Res. 13 (2), tfae044. doi:10.1093/toxres/tfae044
Shi, Y., Inoue, H., Wu, J. C., and Yamanaka, S. (2017). Induced pluripotent stem cell technology: a decade of progress. Nat. Rev. Drug Discov. 16 (2), 115–130. doi:10.1038/nrd.2016.245
Shimizu, E., Iguchi, H., Le, M. N. T., Nakamura, Y., Kobayashi, D., Arai, Y., et al. (2022). A chemically-defined plastic scaffold for the xeno-free production of human pluripotent stem cells. Sci. Rep. 12 (1), 2516. doi:10.1038/s41598-022-06356-8
Shoji, M., Minato, H., Ogaki, S., Seki, M., Suzuki, Y., Kume, S., et al. (2018). Different murine-derived feeder cells alter the definitive endoderm differentiation of human induced pluripotent stem cells. PLOS ONE 13 (7), e0201239. doi:10.1371/journal.pone.0201239
Skorik, C., Mullin, N. K., Shi, M., Zhang, Y., Hunter, P., Tang, Y., et al. (2020). Xeno-Free Reprogramming of Peripheral Blood Mononuclear Erythroblasts on Laminin-521. Curr. Protoc. Stem Cell Biol. 52 (1), e103. doi:10.1002/cpsc.103
Soares, F. A. C., Chandra, A., Thomas, R. J., Pedersen, R. A., Vallier, L., and Williams, D. J. (2014). Investigating the feasibility of scale up and automation of human induced pluripotent stem cells cultured in aggregates in feeder free conditions. J. Biotechnol. 173, 53–58. doi:10.1016/j.jbiotec.2013.12.009
Soldner, F., Laganière, J., Cheng, A. W., Hockemeyer, D., Gao, Q., Alagappan, R., et al. (2011). Generation of Isogenic Pluripotent Stem Cells Differing Exclusively at Two Early Onset Parkinson Point Mutations. Cell 146 (2), 318–331. doi:10.1016/j.cell.2011.06.019
Sommer, C. A., Capilla, A., Molina-Estevez, F. J., Gianotti-Sommer, A., Skvir, N., Caballero, I., et al. (2018). Modeling APC mutagenesis and familial adenomatous polyposis using human iPS cells. PLOS ONE 13 (7), e0200657. doi:10.1371/journal.pone.0200657
Stephens, M. L., and Mak, N. S. (2013). “History of the 3Rs in Toxicity Testing: From Russell and Burch to 21st Century Toxicology,” in Reducing, Refining and Replacing the Use of Animals in Toxicity Testing. Editors D. Allen, and M. D. Waters (The Royal Society of Chemistry), 1–43. Available online at: https://books.rsc.org/books/book/1868/chapter/2313119/History-of-the-3Rs-in-Toxicity-Testing-From.
Strikoudis, A., Cieślak, A., Loffredo, L., Chen, Y. W., Patel, N., Saqi, A., et al. (2019). Modeling of Fibrotic Lung Disease Using 3D Organoids Derived from Human Pluripotent Stem Cells. Cell Rep. 27 (12), 3709–3723.e5. doi:10.1016/j.celrep.2019.05.077
Takahashi, K., Tanabe, K., Ohnuki, M., Narita, M., Ichisaka, T., Tomoda, K., et al. (2007). Induction of Pluripotent Stem Cells from Adult Human Fibroblasts by Defined Factors. Cell 131 (5), 861–872. doi:10.1016/j.cell.2007.11.019
Tamargo-Rubio, I., Simpson, A. B., Hoogerland, J. A., and Fu, J. (2023). Human induced pluripotent stem cell–derived liver-on-a-chip for studying drug metabolism: the challenge of the cytochrome P450 family. Front. Pharmacol. 14, 1223108. doi:10.3389/fphar.2023.1223108
Tanaka, T., Yoshimura, K., Chang, R., Choi, B., Gai, Y., Gupta, P. K., et al. (2023). Comparison of guidelines for biological ancillary materials used for the manufacture of gene and cellular therapy products in Asia. Cytotherapy 25 (2), 220–228. doi:10.1016/j.jcyt.2022.09.005
Tanigawa, S., Islam, M., Sharmin, S., Naganuma, H., Yoshimura, Y., Haque, F., et al. (2018). Organoids from Nephrotic Disease-Derived iPSCs Identify Impaired NEPHRIN Localization and Slit Diaphragm Formation in Kidney Podocytes. Stem Cell Rep. 11 (3), 727–740. doi:10.1016/j.stemcr.2018.08.003
Tannenbaum, J., and Bennett, B. T. (2015). Russell and Burch’s 3Rs then and now: the need for clarity in definition and purpose. J. Am. Assoc. Lab. Anim. Sci. JAALAS 54 (2), 120–132.
Tanner, R. (2018). The 3Rs: What are Medical Scientists Doing about Animal Testing? Front Young Minds. 5, 44.
Thomas, R. J., Anderson, D., Chandra, A., Smith, N. M., Young, L. E., Williams, D., et al. (2009). Automated, scalable culture of human embryonic stem cells in feeder-free conditions. Biotechnol. Bioeng. 102 (6), 1636–1644. doi:10.1002/bit.22187
Thomson, J. A., Itskovitz-Eldor, J., Shapiro, S. S., Waknitz, M. A., Swiergiel, J. J., Marshall, V. S., et al. (1998). Embryonic Stem Cell Lines Derived from Human Blastocysts. Science 282 (5391), 1145–1147. doi:10.1126/science.282.5391.1145
Timilsina, S., McCandliss, K. F., Trivedi, E., and Villa-Diaz, L. G. (2023). Enhanced Expansion of Human Pluripotent Stem Cells and Somatic Cell Reprogramming Using Defined and Xeno-Free Culture Conditions. Bioengineering 10 (9), 999. doi:10.3390/bioengineering10090999
Tokunaga, K., Saitoh, N., Goldberg, I. G., Sakamoto, C., Yasuda, Y., Yoshida, Y., et al. (2014). Computational image analysis of colony and nuclear morphology to evaluate human induced pluripotent stem cells. Sci. Rep. 4 (1), 6996. doi:10.1038/srep06996
Tolosa, L., Carmona, A., Castell, J. V., Gómez-Lechón, M. J., and Donato, M. T. (2015). High-content screening of drug-induced mitochondrial impairment in hepatic cells: effects of statins. Arch. Toxicol. 89 (10), 1847–1860. doi:10.1007/s00204-014-1334-3
Torizal, F. G., Kim, S. M., Horiguchi, I., Inamura, K., Suzuki, I., Morimura, T., et al. (2022). Production of homogenous size-controlled human induced pluripotent stem cell aggregates using ring-shaped culture vessel. J. Tissue Eng. Regen. Med. 16 (3), 254–266. doi:10.1002/term.3278
Tran, T., Song, C. J., Nguyen, T., Cheng, S. Y., McMahon, J. A., Yang, R., et al. (2022). A scalable organoid model of human autosomal dominant polycystic kidney disease for disease mechanism and drug discovery. Cell Stem Cell 29 (7), 1083–1101.e7. doi:10.1016/j.stem.2022.06.005
Tran, T. T. V., Surya Wibowo, A., Tayara, H., and Chong, K. T. (2023). Artificial Intelligence in Drug Toxicity Prediction: Recent Advances, Challenges, and Future Perspectives. J. Chem. Inf. Model 63 (9), 2628–2643. doi:10.1021/acs.jcim.3c00200
Trastulla, L., Noorbakhsh, J., Vazquez, F., McFarland, J., and Iorio, F. (2022). Computational estimation of quality and clinical relevance of cancer cell lines. Mol. Syst. Biol. 18 (7), e11017. doi:10.15252/msb.202211017
Tristan, C. A., Ormanoglu, P., Slamecka, J., Malley, C., Chu, P. H., Jovanovic, V. M., et al. (2021). Robotic high-throughput biomanufacturing and functional differentiation of human pluripotent stem cells. Stem Cell Rep. 16 (12), 3076–3092. doi:10.1016/j.stemcr.2021.11.004
Tukker, A., Van Kleef, R. G. D. M., Wijnolts, F. M. J., De Groot, A., and Westerink, R. H. S. (2019). Towards animal-free neurotoxicity screening: Applicability of hiPSC-derived neuronal models for in vitro seizure liability assessment. ALTEX 37, 121–135. doi:10.14573/altex.1907121
Unger, C., Skottman, H., Blomberg, P., Sirac Dilber, M., and Hovatta, O. (2008). Good manufacturing practice and clinical-grade human embryonic stem cell lines. Hum. Mol. Genet. 17 (R1), R48–R53. doi:10.1093/hmg/ddn079
Vandana, J. J., Manrique, C., Lacko, L. A., and Chen, S. (2023). Human pluripotent-stem-cell-derived organoids for drug discovery and evaluation. Cell Stem Cell 30 (5), 571–591. doi:10.1016/j.stem.2023.04.011
VanderWall, K. B., Huang, K. C., Pan, Y., Lavekar, S. S., Fligor, C. M., Allsop, A. R., et al. (2020). Retinal Ganglion Cells With a Glaucoma OPTN(E50K) Mutation Exhibit Neurodegenerative Phenotypes when Derived from Three-Dimensional Retinal Organoids. Stem Cell Rep. 15 (1), 52–66. doi:10.1016/j.stemcr.2020.05.009
Villa-Diaz, L. G., Pacut, C., Slawny, N. A., Ding, J., O’Shea, K. S., and Smith, G. D. (2009). Analysis of the Factors that Limit the Ability of Feeder Cells to Maintain the Undifferentiated State of Human Embryonic Stem Cells. Stem Cells Dev. 18 (4), 641–651. doi:10.1089/scd.2008.0010
Villa-Diaz, L. G., Nandivada, H., Ding, J., Nogueira-de-Souza, N. C., Krebsbach, P. H., O’Shea, K. S., et al. (2010). Synthetic polymer coatings for long-term growth of human embryonic stem cells. Nat. Biotechnol. 28 (6), 581–583. doi:10.1038/nbt.1631
Villa-Diaz, L. G., Ross, A. M., Lahann, J., and Krebsbach, P. H. (2013). Concise Review: The Evolution of human pluripotent stem cell culture: From feeder cells to synthetic coatings. Stem Cells 31 (1), 1–7. doi:10.1002/stem.1260
Villa-Diaz, L. G., Kim, J. K., Laperle, A., Palecek, S. P., and Krebsbach, P. H. (2016). Inhibition of Focal Adhesion Kinase Signaling by Integrin α6β1 Supports Human Pluripotent Stem Cell Self-Renewal. Stem Cells 34 (7), 1753–1764. doi:10.1002/stem.2349
Visone, R., Lozano-Juan, F., Marzorati, S., Rivolta, M. W., Pesenti, E., Redaelli, A., et al. (2023). Predicting human cardiac QT alterations and pro-arrhythmic effects of compounds with a 3D beating heart-on-chip platform. Toxicol. Sci. 191 (1), 47–60. doi:10.1093/toxsci/kfac108
Vo, Q. D., Saito, Y., Ida, T., Nakamura, K., and Yuasa, S. (2024). The use of artificial intelligence in induced pluripotent stem cell-based technology over 10-year period: A systematic scoping review. PLoS One 19, e0302537. doi:10.1371/journal.pone.0302537
Volpato, V., and Webber, C. (2020). Addressing variability in iPSC-derived models of human disease: guidelines to promote reproducibility. Dis. Model Mech. 13 (1), dmm042317. doi:10.1242/dmm.042317
Volpato, V., Smith, J., Sandor, C., Ried, J. S., Baud, A., Handel, A., et al. (2018). Reproducibility of Molecular Phenotypes after Long-Term Differentiation to Human iPSC-Derived Neurons: A Multi-Site Omics Study. Stem Cell Rep. 11 (4), 897–911. doi:10.1016/j.stemcr.2018.08.013
Wan, H., Fu, R., Tong, M., Wang, Y., Wang, L., Wang, S., et al. (2022). Influence of feeder cells on transcriptomic analysis of pluripotent stem cells. Cell Prolif. 55 (2), e13189. doi:10.1111/cpr.13189
Wang, G., Zhang, H., Zhao, Y., Li, J., Cai, J., Wang, P., et al. (2005). Noggin and bFGF cooperate to maintain the pluripotency of human embryonic stem cells in the absence of feeder layers. Biochem. Biophys. Res. Commun. 330 (3), 934–942. doi:10.1016/j.bbrc.2005.03.058
Ware, B. R., Berger, D. R., and Khetani, S. R. (2015). Prediction of Drug-Induced Liver Injury in Micropatterned Co-cultures Containing iPSC-Derived Human Hepatocytes. Toxicol. Sci. 145 (2), 252–262. doi:10.1093/toxsci/kfv048
Winanto, K. Z. J., Soh, B. S., Fan, Y., and Ng, S. Y. (2020). Organoid cultures of MELAS neural cells reveal hyperactive Notch signaling that impacts neurodevelopment. Cell Death Dis. 11 (3), 182. doi:10.1038/s41419-020-2383-6
Witmer, A., and Bhanu, B. (2018). “HESCNET: A Synthetically Pre-Trained Convolutional Neural Network for Human Embryonic Stem Cell Colony Classification,” in 2018 25th IEEE International Conference on Image Processing (ICIP) (Athens: IEEE), 2441–2445. Available online at: https://ieeexplore.ieee.org/document/8451624/.
Workman, M. J., Mahe, M. M., Trisno, S., Poling, H. M., Watson, C. L., Sundaram, N., et al. (2017). Engineered human pluripotent-stem-cell-derived intestinal tissues with a functional enteric nervous system. Nat. Med. 23 (1), 49–59. doi:10.1038/nm.4233
Wu, J., Rostami, M. R., Cadavid Olaya, D. P., and Tzanakakis, E. S. (2014). Oxygen Transport and Stem Cell Aggregation in Stirred-Suspension Bioreactor Cultures. PLoS ONE, 9, e102486. doi:10.1371/journal.pone.0102486
Wulansari, N., Darsono, W. H. W., Woo, H. J., Chang, M. Y., Kim, J., Bae, E. J., et al. (2021). Neurodevelopmental defects and neurodegenerative phenotypes in human brain organoids carrying Parkinson’s disease-linked DNAJC6 mutations. Sci. Adv. 7 (8), eabb1540. doi:10.1126/sciadv.abb1540
Xu, C., Inokuma, M. S., Denham, J., Golds, K., Kundu, P., Gold, J. D., et al. (2001). Feeder-free growth of undifferentiated human embryonic stem cells. Nat. Biotechnol. 19 (10), 971–974. doi:10.1038/nbt1001-971
Yang, X., and Papoian, T. (2018). Moving beyond the comprehensive in vitro proarrhythmia assay: Use of human-induced pluripotent stem cell-derived cardiomyocytes to assess contractile effects associated with drug-induced structural cardiotoxicity. J. Appl. Toxicol. 38 (9), 1166–1176. doi:10.1002/jat.3611
Yang, X., Pabon, L., and Murry, C. E. (2014). Engineering Adolescence: Maturation of Human Pluripotent Stem Cell–Derived Cardiomyocytes. Circ. Res. 114 (3), 511–523. doi:10.1161/CIRCRESAHA.114.300558
Yildirim, Z., Swanson, K., Wu, X., Zou, J., and Wu, J. (2025). Next-Gen Therapeutics: Pioneering Drug Discovery with iPSCs, Genomics, AI, and Clinical Trials in a Dish. Annu. Rev. Pharmacol. Toxicol. 65 (1), 71–90. doi:10.1146/annurev-pharmtox-022724-095035
Yip, S., Wang, N., and Sugimura, R. (2023). Give them vasculature and immune cells – how to fill the gap of organoids. Cells Tissues Organs 212, 369–382. doi:10.1159/000529431
Yuan, F., Fang, K. H., Cao, S. Y., Qu, Z. Y., Li, Q., Krencik, R., et al. (2015). Efficient generation of region-specific forebrain neurons from human pluripotent stem cells under highly defined condition. Sci. Rep. 5 (1), 18550. doi:10.1038/srep18550
Zhang, H., Shao, X., Peng, Y., Teng, Y., Saravanan, K. M., Zhang, H., et al. (2019). A novel machine learning based approach for iPS progenitor cell identification. PLOS Comput. Biol. 15, e1007351. doi:10.1371/journal.pcbi.1007351
Zhao, J., Fu, Y., Yamazaki, Y., Ren, Y., Davis, M. D., Liu, C. C., et al. (2020). APOE4 exacerbates synapse loss and neurodegeneration in Alzheimer’s disease patient iPSC-derived cerebral organoids. Nat. Commun. 11 (1), 5540. doi:10.1038/s41467-020-19264-0
Zhou, P., Yin, B., Zhang, R., Xu, Z., Liu, Y., Yan, Y., et al. (2018). Molecular basis for RGD-containing peptides supporting adhesion and self-renewal of human pluripotent stem cells on synthetic surface. Colloids Surf. B Biointerfaces 171, 451–460. doi:10.1016/j.colsurfb.2018.07.050
Keywords: human pluripotent stem cells, feeder-free culture, xeno-free substrates, drug discovery, toxicology
Citation: Chaudhary N and Villa-Diaz LG (2025) Advancements in the in vitro culture of human pluripotent stem cells: progress, challenges, and future directions: comprehensive review. Front. Toxicol. 7:1667573. doi: 10.3389/ftox.2025.1667573
Received: 16 July 2025; Accepted: 20 October 2025;
Published: 02 December 2025.
Edited by:
Tilo Weber, Animal Welfare Academy of the German Welfare Federation, GermanyReviewed by:
Lincoln G. Moro, University of São Paulo, BrazilCecilia Benazzato, Department of Health, Brazil
Copyright © 2025 Chaudhary and Villa-Diaz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis G. Villa-Diaz, bHVpc3ZpbGxhZGlhekBvYWtsYW5kLmVkdQ==