 Anshu Rastogi1,2*
Anshu Rastogi1,2* Marek Zivcak2
Marek Zivcak2 Oksana Sytar2,3
Oksana Sytar2,3 Hazem M. Kalaji4,5
Hazem M. Kalaji4,5 Xiaolan He6
Xiaolan He6 Sonia Mbarki7
Sonia Mbarki7 Marian Brestic2*
Marian Brestic2*- 1Department of Meteorology, Poznan University of Life Sciences, Poznan, Poland
- 2Department of Plant Physiology, Slovak University of Agriculture, Nitra, Slovakia
- 3SRL “Physiological Bases of Plant Productivity,” Educational and Scientific Center “Institute of Biology and Medicine,” Taras Shevchenko National University of Kyiv, Kiev, Ukraine
- 4SI Technology, Warsaw, Poland
- 5Department of Plant Physiology, Faculty of Agriculture and Biology, Warsaw University of Life Science—SGGW, Warsaw, Poland
- 6Jiangsu Academy of Agricultural Sciences, Nanjing, China
- 7National Research Institute of Rural Engineering, Water and Forests, Aryanah, Tunisia
An increasing need of nanotechnology in various industries may cause a huge environment dispersion of nanoparticles in coming years. A concern about nanoparticles interaction with flora and fauna is raised due to a growing load of it in the environment. In recent years, several investigators have shown impact of nanoparticles on plant growth and their accumulation in food source. This review examines the research performed in the last decade to show how metal and metal oxide nanoparticles are influencing the plant metabolism. We addressed here, the impact of nanoparticle on plant in relation to its size, concentration, and exposure methodology. Based on the available reports, we proposed oxidative burst as a general mechanism through which the toxic effects of nanoparticles are spread in plants. This review summarizes the current understanding and the future possibilities of plant-nanoparticle research.
Introduction
Nanoparticles are classified as a material in which at least one dimension is <100 nm in diameter (Auffan et al., 2009). Nanoparticles are not new to the environment and occur naturally in the form of minerals, clays, and products of bacteria. It has been used since ancient times as a colorant for metals, but the systematic design and engineering of nanoparticles for various uses has started only in the last few decades (Maurer-Jones et al., 2013). Engineered nanoparticles are designed to have the properties which are not present in bulk samples of the same materials (Auffan et al., 2009). Engineered nanoparticles are composed of a variety of materials and occur in different sizes and shapes with a suite of synthetic surface molecules, which makes them distinct from naturally occurring materials (Radad et al., 2012; Maurer-Jones et al., 2013). Metal and metal oxides nanoparticles exhibit different physiochemical properties and are different than their native bulk compounds in several respects which includes its surface, optical, thermal, and electrical properties. Metal and metal oxide nanoparticles are manufactured by addition of reducing or oxidizing/precipitating agents during their synthesis, respectively (Sanchez-Dominguez et al., 2009). Several factors are responsible for nanoparticles reactivity with biomolecules which includes nanoparticles size, core composition, shape, surface properties, purity, stability, and method of manufacturing (Teske and Detweiler, 2015; Wang P. et al., 2016). There is a good chance that nanoparticles may retain the major characteristic of their bulk material, therefore, it is needed to consider the impact of bulk material while the study of nanoparticles interaction in environment, for example, heavy metals are toxic to plants whereas silicon as a metalloid was observed to be beneficial for plants (Yadav, 2010; Tubana et al., 2016; Helaly et al., 2017).
In the last decades, nanoparticles have been used in various household and industrial products. Due to the increasing use of nanoparticles in commercial products, different industries are developing novel nanoparticles for the improvement of their services and products. Some of the industries with an intensive use of nanoparticles, in which can be expected the release of nanoparticles to the environment, are indicated in Figure 1. A few of the many nanoparticles are used on a very large scale and have the potential for making its way into the environment. The nanoparticles can contaminate the environment through various processes such as, the improper management of industrial waste and improper disposal of products by the users. Several mathematical models are being developed to estimate the release of nanoparticles to the environment (Keller and Lazareva, 2014; Dumont et al., 2015). According to consumption of Silver nanoparticle (AgNP), and Zinc oxide nanoparticles in Europe per person, their release has been assumed to be significant and broadly distributed in European territory (Dumont et al., 2015). Keller and Lazareva (2014) have also estimated a significant release of different nanoparticles to the environment. In addition nanoparticles are susceptible to environmental conditions and can change their aggregation state, oxidation state, precipitation of secondary phases etc., in different environmental condition (Levard et al., 2012). The physical parameters and chemicals presence in different environment influence the stability of nanoparticles. Therefore, nanoparticles may behave differently in different condition (Levard et al., 2012), and thus their availability and reactivity in ecosystem is affected. The composition of nanoparticles may also change their properties and therefore their reactivity, penetration and translocation inside the plant which may leads to different responses of plants to the same nanoparticle, for example, Barrios et al. (2016) has shown that capping of nanoparticles influences the plant responses compared to exposure to bare nanoparticle. Plants are in continuous interaction with air, soil, and water, all of which may contain engineered nanoparticles. As the plants are also consumed by animals, the nanoparticles may be transferred to them. There is a risk that nanoparticles could invade the food chain and become dangerous to humans (the last link in the food chain). This is especially important, as the excessive usage of nanoparticles and their abundance in the environment would increase and, as a result, both plants and animals may become the source of nanoparticles for humans. Few studies in recent year confirm the trophic transfer of different nanoparticles through a terrestrial or aquatic food chain (Judy et al., 2012; Unrine et al., 2012; Hawthorne et al., 2014; De la Torre Roche et al., 2015; Tangaa et al., 2016).
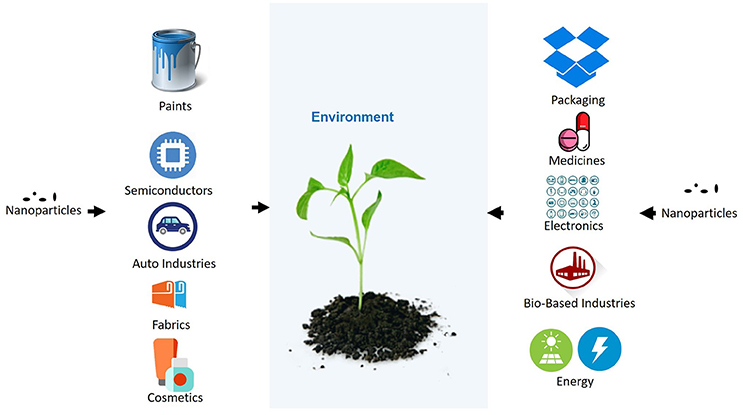
Figure 1. Uses of nanoparticles in different industries and its leakage to environment.
Despite the plants are producers and play a major role in the ecosystem, the impact of nanoparticles upon them is not well studied (Rico et al., 2011; Feng et al., 2013; Zuverza-Mena et al., 2017). The lack of proper detection methods for nanoparticles from environments makes the study of the nanoparticles complicated (Navratilova et al., 2015; Mahdi et al., 2017). Among different possible techniques, inductively coupled plasma mass spectroscopy (ICP-MS) is one of the most reliable techniques for the detection of nanoparticles (Hadioui et al., 2015; Navratilova et al., 2015; Mahdi et al., 2017). The research performed on different plants has shown that the nanoparticles may have both a positive and a negative impact on plants, depending on size, concentration, chemical composition, zeta potential, stability, and the shape of nanoparticles (Mirzajani et al., 2013; Rafique et al., 2014; Nhan et al., 2015; Tripathi et al., 2015, 2017; Costa and Sharma, 2016; Wang Z. et al., 2016). Several studies have depicted a negative impact of nanoparticles on plants in the form of decrease in plant growth, productivity and pigments (Landa et al., 2016; Tripathi et al., 2017). On the other hand, smartly designed nanoparticles are also used for the betterment of agricultural crop production, as growth stimulators, nanopesticides, nanofertilizers, soil improving agents, or sensors for monitoring different agricultural parameters in the field (Fraceto et al., 2016; Wang P. et al., 2016). Due to the increased interest in the area, most of the research depicting the influence of industrial nanoparticles on plants has been performed in recent years. Therefore, the purpose of this review is to systematically present and analyze the research performed in the last 10 years to give an overview of the recent advancement in the field.
In the following sections, we will discuss the presence of different types of nanoparticles in the environment, the impact of different nanoparticles on plants, and a concise discussion and a general mechanism through which nanoparticles may cause an impact on the plant.
Effect of Nanoparticles on Plants
Nanoparticles cover a heterogeneous range of materials (Santos et al., 2015), but only a few of them are extensively used and at present, the environment is at risk to be exposed to them. Metal and metal oxide nanoparticles of titanium dioxide (TiO2), silver, zinc oxide, cerium dioxide, copper, copper oxide, aluminum, nickel, and iron are most commonly used in industries and therefore are mostly studied for their impacts on different plants. Some non-metal nanoparticles, such as, single-walled carbon nanotubes and fullerene have been well studied to reveal their nanotoxicity mechanisms (Joner et al., 2008). From another side, enhancing growth together with an acceleration of seed germination for different organs of corn, tomato, rice, and soybean has been observed under exposure to single-walled carbon nanohorns (SWCNHs) (Lahiani et al., 2015). In recent years, nanoparticles have been developed to be used in agriculture as nanopesticides and nanofertilizers (which include the use of nanoparticles as nanocarrier for pesticides, fertilizers; Fraceto et al., 2016; Wang P. et al., 2016). Nanoparticles of chitosan were used to encapsulated herbicide, due to which the efficiency of herbicide was observed to be enhanced significantly (Maruyama et al., 2016). Mesoporous silicon nanoparticles as a metalloid nanoparticles have also been used to deliver DNA, proteins, and other chemicals in plants (Torney et al., 2007; Martin-Ortigosa et al., 2014). Use in agriculture includes nanoparticles, such as, nanozeolites (basic building blocks of silicate [SiO4]− and aluminates [AlO4]− tetrahedrons) as well as the hydrogels (consisting of different polymers such as, chitosan and alginate), which helps in the improvement of soil quality, and nanosensors (for monitoring plant and soil health; Fraceto et al., 2016). Silica nanoparticles were observed to be nontoxic to plant (Slomberg and Schoenfisch, 2012), but some authors observed the toxic effect due to decrees in pH of the media after addition of nanoparticles. Tripathi et al. (2015) have studied that Silica nanoparticle was able to alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings (Tripathi et al., 2015). Several studies on the impact of metal and metal oxide nanoparticles on plant have shown a toxic impact on plants, whereas few studies also indicated their beneficial role in the form of enhancing plant growth parameters and productivity (Castiglione et al., 2011; Clément et al., 2013; Dimkpa et al., 2013; Jaberzadeh et al., 2013; Jiang et al., 2014; Rafique et al., 2014; Raliya et al., 2015; Okupnik and Pflugmacher, 2016; Cvjetko et al., 2017; Tripathi et al., 2017).
To determine whether the metal and metal oxide nanoparticles represent a risk to plant organisms and the environment, analytical selection of information is needed regarding size, concentration, zeta potential, uptake by a certain type of plants and effects on the plant. In particular, the zeta potential represents an important reliable indicator of nanoparticle coagulation and reactivity in solution. Zeta potential indicates total electric potential of all particles and ions in solution, and thus get effected by changes in pH or ionic strength (Teske and Detweiler, 2015). The effect of nanoparticles on plants occurs in several physiological, morphological, and genotoxic changes. Therefore, for the effective use of nanotechnology in agriculture, it is important to know the role of certain nanoparticle (Nair, 2016). Effect of different metal and metal oxide nanoparticles on different plants is observed to be variable and ranges from their positive impact to the lethal impact in plants (Tables 1–4). To clearly compare the different studies, it is divided into four following sections.
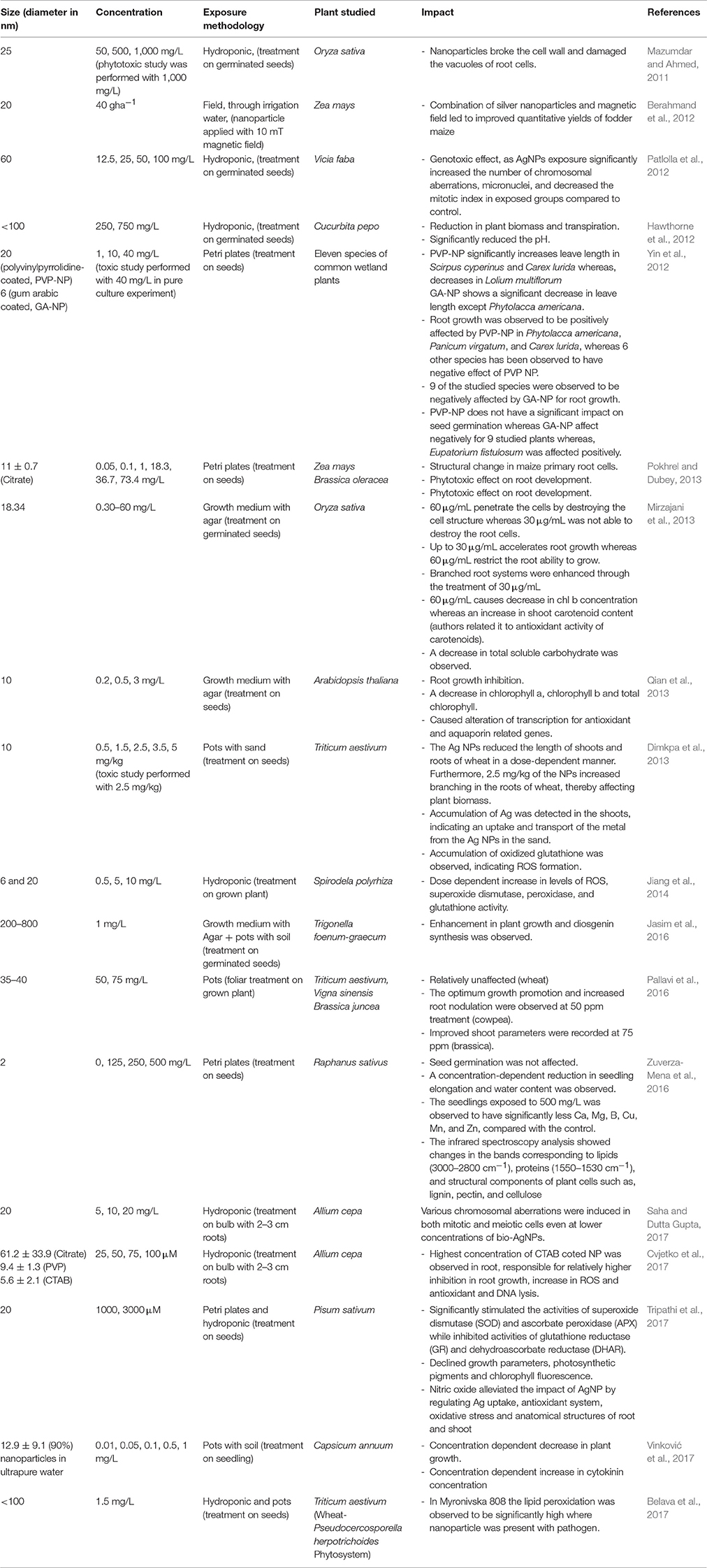
Table 1. Impact of AgNP on plants.
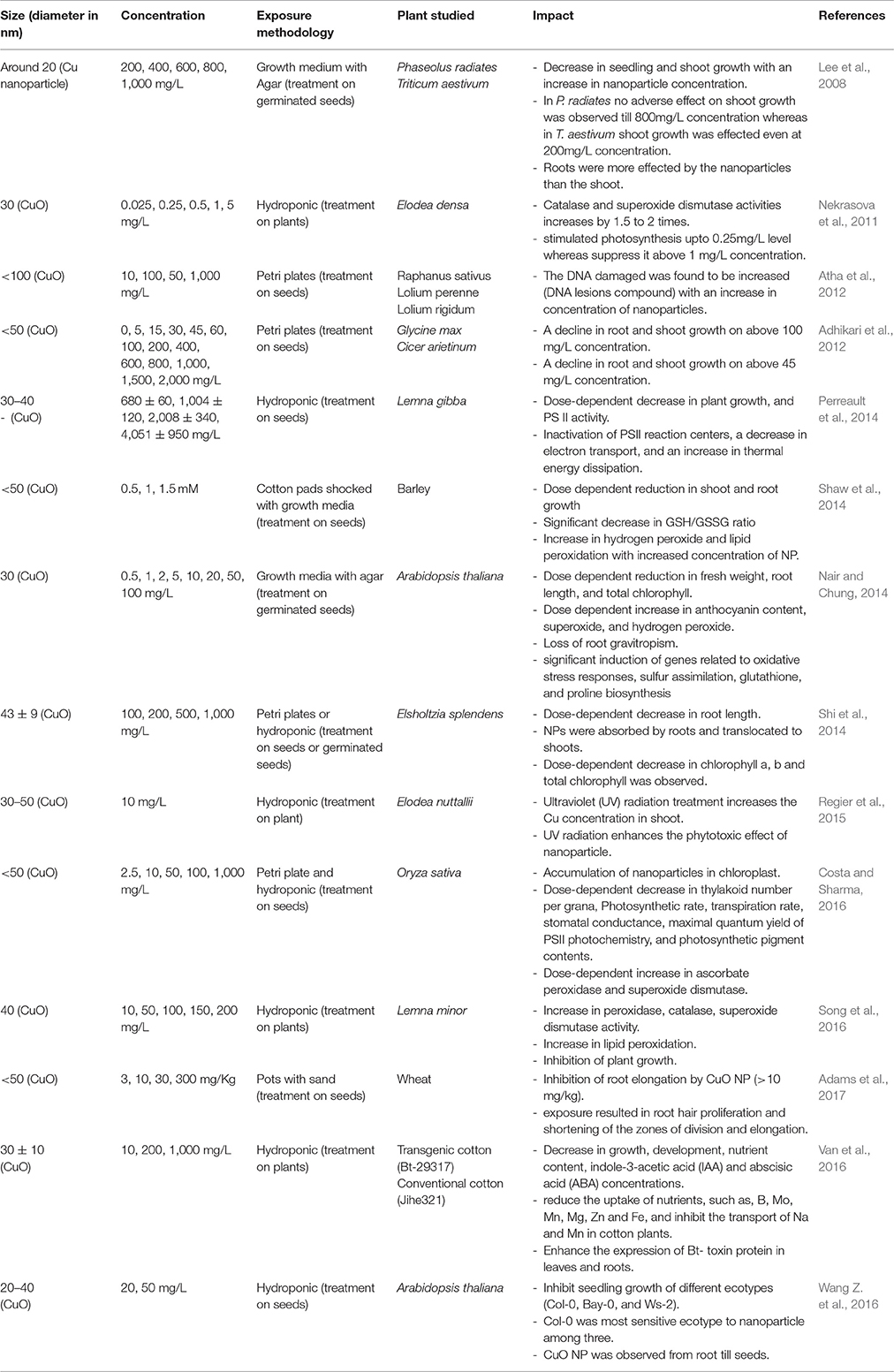
Table 2. Impact of Cu and CuO NP on plants.
Impact of Silver Nanoparticles (AgNP)
Among different nanoparticles, AgNPs are fetching more attention because of their intensive uses in various products, which includes their uses as antimicrobial agents, shampoo, soap, toothpaste, waste water treatment, food packaging materials, food storage containers, fabrics, room sprays, detergents, paint, etc. (Boxall et al., 2008; Rai et al., 2009; Wijnhoven et al., 2009). Due to its extensive use, the production of nanoparticles is increasing rapidly, among which the United States itself has been reported to produce 2,500 tons/year of AgNP, of which around 150 tons end up in sewage sludge and 80 tons in surface waters (Khaydarov et al., 2009; El-Temsah and Joner, 2012). Through sludge and surface water the AgNP may easily reach to the plants.
The AgNPs of 25 nm at high concentration was observed to break the cell wall and damage the vacuoles of root cells of Oryza sativa, thus causing a toxic effect (Mazumdar and Ahmed, 2011). Mirzajani et al. (2013) observed that the AgNP was unable to penetrate the root cells of O. sativa when present in low concentration (up to 30 μg/mL), whereas the higher concentration was able to destroy the cell structure and cause the toxic effect. The authors also reported that, the 30 μg/mL accelerates root growth, whereas 60 μg/mL restrict the ability of root to grow. The observations indicate that the penetration of AgNP is necessary to cause a toxic effect, whereas when present in surrounding, it may have a positive impact on plants. Krishnaraj et al. (2012) observed a mild or no effect of biologically synthesized AgNP on Bacopa monnieri. The synthesized nanoparticle induced the protein and carbohydrate synthesis and decreased the total phenol contents, which can be considered as a positive effect, it may be due to the presence of different size (2–50 nm) of nanoparticle with different penetration capacity in the highest applied concentration (100 ppm), or the different chemical property of biologically synthesized NP. The AgNP of 200–800 nm size was observed to enhance the plant growth (Jasim et al., 2016), whereas 35–40 nm of AgNP was observed to positively influence the root and shoot growth of different plant (Pallavi et al., 2016), which may be due to the inability of the penetration of large nanoparticles in studied low concentration as reported by Mirzajani et al. (2013). Different size of AgNP used in the various studies shows a clear correlation between the size and toxic relation of NP to the plant, the NP with lower size was always observed to have higher toxicity to the plant compared to larger NP (Yin et al., 2012; Jiang et al., 2014; Cvjetko et al., 2017).
AgNPs (of comparatively small size i.e., <30 nm) when applied in high concentration were observed to inhibit the root and shoot growth in different plant studied (Dimkpa et al., 2013; Qian et al., 2013; Tripathi et al., 2017; Vinković et al., 2017). As a response to AgNP stress an enhancement in reactive oxygen species (ROS) was observed, which also leads to the enhanced production of antioxidant enzymes and molecules as an adaptive mechanism (Dimkpa et al., 2013; Jiang et al., 2014; Cvjetko et al., 2017; Tripathi et al., 2017). The AgNPs were also observed to cause an impact on DNA and influences the gene expression in several plants (Patlolla et al., 2012; Qian et al., 2013; Cvjetko et al., 2017; Saha and Dutta Gupta, 2017). Physiological impacts of AgNP was observed in the form of a decrease in transpiration (Hawthorne et al., 2012), chlorophyll concentration (Mirzajani et al., 2013; Qian et al., 2013; Tripathi et al., 2017); and chlorophyll fluorescence (Tripathi et al., 2017). A significant alteration in different macromolecules, lipids, proteins, lignin, pectin and cellulose were observed in Raphanus sativus when treated with 2 nm AgNP with 500 mg/L concentration (Zuverza-Mena et al., 2016). Plant hormones such as, cytokinin and auxin were also observed to be affected by the AgNP (Yin et al., 2012; Vinković et al., 2017).
Recent studies have shown that when AgNP was combined with different treatment/compounds, it may have a different impact on plants (Berahmand et al., 2012; Belava et al., 2017; Tripathi et al., 2017). This can be explained by the influence of other phenomena/compound on AgNP. AgNP treatment in combination with magnetic field was observed to improve quantitative yields in Zea mays (Berahmand et al., 2012), whereas the nitric oxide was observed to alleviate the impact of AgNP by regulating Ag uptake, an antioxidant system, oxidative stress, and anatomical structure (Tripathi et al., 2017). In the wheat-pathogen phytosystem, an enhancement of lipid peroxidation was observed, when compared with NP or pathogen alone (Belava et al., 2017). Due to its fungicidal activity, AgNP have been tested against few plant-pathogenic fungi, and their impact was found to be significant in eliminating the fungi (Jo et al., 2009). But their use in agriculture is still questionable as AgNP is known to release silver ions with its age, moreover, they can affect the biomass accumulation in soil (Johansson et al., 1998; Liu and Hurt, 2010).
The study clearly indicates AgNP exhibit an impact on different aspect of plant morphology, physiology, and biochemistry, which depends on the size, properties, and concentration of the NP in use. On the basis of the indicated studies, it can be hypothesized that for exhibiting a toxic effects AgNP need to penetrate the plant tissue and interfere with different metabolic activities. For better understanding, the influence of different AgNP on a plant is summarized in Table 1.
Impact of Copper and Copper Oxide Nanoparticles
Copper is an essential micronutrient, which is incorporated in many proteins and enzymes, therefore, playing a significant role in plant health and nutrition. Copper nanoparticles (Cu NP) are widely used in different commercial applications such as, an antimicrobial agent, catalysts, gas sensors, electronics, batteries, heat transfer fluids, etc. (Kasana et al., 2017). Due to its oxidative property, copper oxide nanoparticles (CuO NP) are assumed to have a higher toxic effect than Cu NP. CuO NP was observed to have a positive impact on Elodea densa (waterweed) and stimulate photosynthesis at low concentration (<0.25 mg/L), but the impact scenario completely changes with higher doses and at 1 mg/L concentration a clear suppression in photosynthesis was observed (Nekrasova et al., 2011). The root morphology was reported to be adversely affected with Cu and CuO NP, with almost complete inhibition with a high dose of NP (Lee et al., 2008; Adhikari et al., 2012; Perreault et al., 2014; Shaw et al., 2014; Song et al., 2016; Adams et al., 2017). CuO NP was observed to enhance the production of ROS in plants (Nair and Chung, 2014; Shaw et al., 2014). Different antioxidant compounds were observed to be significantly increased in plants treated with NP indicating the activation of the protective mechanism by plants (Shaw et al., 2014; Song et al., 2016). The genetic level study on Arabidopsis thaliana has shown that at 0.2 mg/L concentration CuO NP does not cause any impact on the expression of genes related to oxidative stress responses, sulfur assimilation, glutathione, and proline biosynthesis (ATPS, APR, CS, GCL, P5CS1, and P5GS2), whereas, the gene expression was observed to be upregulated at higher concentrations (Nair and Chung, 2014). Atha et al. (2012) have reported a significant accumulation of oxidatively modified, mutagenic DNA lesions in different plants indicating the DNA damage as a response to CuO NP treatment. CuO NP was also observed to negatively affect the photosynthetic activity by inactivating PS II reaction centers, and causing a decrease in electron transport, thylakoid number per grana, photosynthetic rate, photosynthetic pigments, transpiration rate, stomatal conductance (Perreault et al., 2014; Costa and Sharma, 2016). Phytohormones were also observed to be altered as a response to CuO NP (Nair and Chung, 2014; Van et al., 2016). When different varieties or plants were studied together, the influence of CuO NP was observed to be different in genetically diverse plants (Lee et al., 2008; Adhikari et al., 2012; Atha et al., 2012; Van et al., 2016; Wang Z. et al., 2016). A combined treatment of the plant with ultraviolet radiation and CuO NP were observed to significantly enhance the phytotoxic effect of CuO NP (Regier et al., 2015).
The study indicates that the Cu and CuO NPs are toxic to plants when present in concentrations higher than 0.2 mg/L and it influence the growth, physiology, and biochemistry of plants.
Impact of Titanium Dioxide Nanoparticles (TiO2 NP)
TiO2 NP belong to the most used nanoparticles, which are used in cosmetic and skin care products, antibacterial and cleaning air products, paints, and for decomposing organic matter in wastewater (Castiglione et al., 2011; Clément et al., 2013). Few studies have been performed to indicate the influence of TiO2 NP shows that the TiO2 NP may influence plants in positive and negative ways (Table 3).
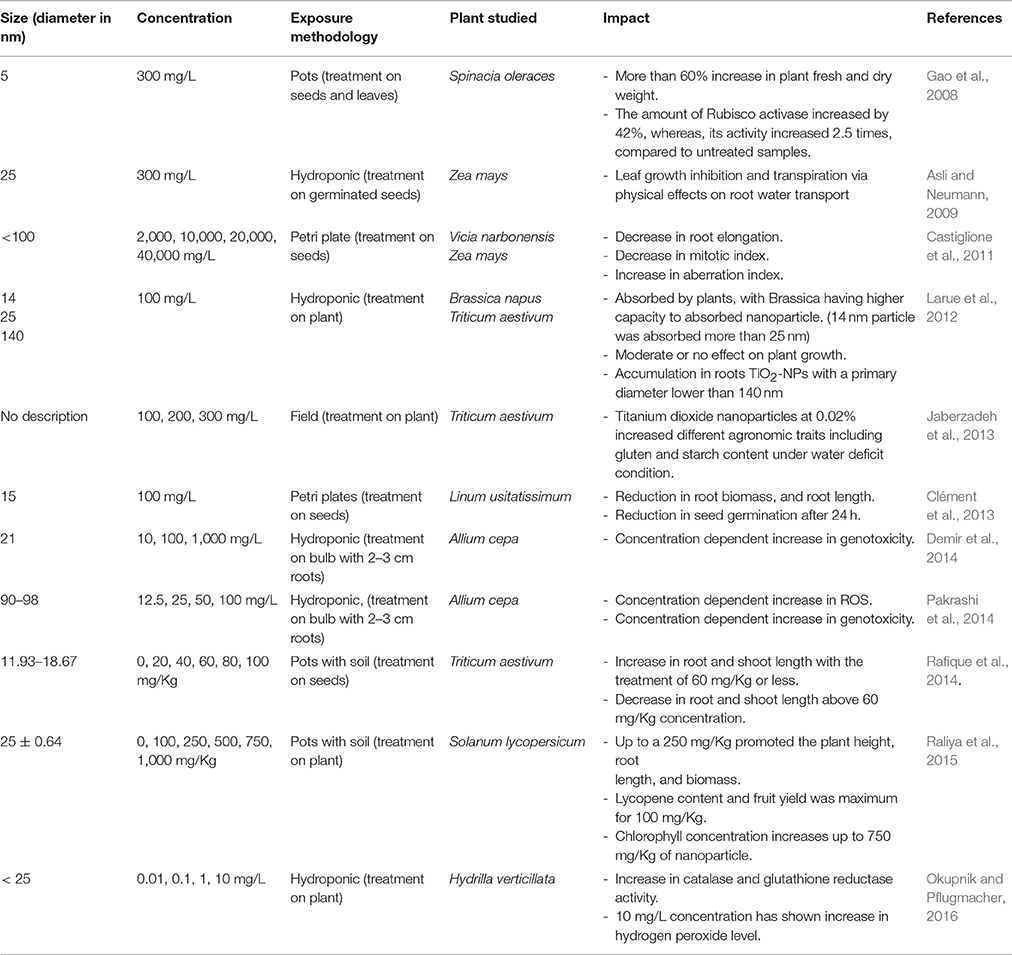
Table 3. Impact of TiO2 NP on plants.
Due to photocatalytic properties of titanium nanoparticles, most of the studies where TiO2 NP was used at foliar level has shown a positive impacts on plant (Table 3; Gao et al., 2008; Jaberzadeh et al., 2013; Raliya et al., 2015). Spinach was observed to have more than 60% increase in the fresh weight and dry weight under the influence of TiO2 NPs (Gao et al., 2008). The author also observed the increase in amount and activity of Rubisco activase in photosynthesis. The foliar treatment of TiO2 NP also showed a better growth of the plant, increase in fruit yield, and chlorophyll concentration in Solanum lycopersicum (Raliya et al., 2015). Jaberzadeh et al. (2013) reported that TiO2 NP counteracts the water stress in Triticum aestivum by improving agronomic traits.
The smaller TiO2 NP was also observed to be transported by roots, it was suggested that above diameter 140 nm, TiO2 NPs are no longer accumulated in roots. TiO2 NPs with a diameter above 36 nm was observed to be accumulated in wheat root parenchyma but did not reach the stele and therefore, do not translocate to the shoot (Larue et al., 2012). A plant response to hydroponics exposure to TiO2 NPs may differ from the response to TiO2 NP-contaminated soil exposure. Colloidal suspensions of nanoparticles were observed to inhibit the leaf growth and transpiration via physical effects on root-water transport system (Asli and Neumann, 2009). At high concentration, TiO2 NP was observed to be toxic to plants, even in soil system (Rafique et al., 2014). The phytotoxic response was found to be similar to AgNP or CuO NP, with a decrease in plant growth, mitotic index, and an increase in ROS, antioxidant activity, and genotoxicity (Table 3). The study indicates that the impact of TiO2 NP on different plants depends on the concentration, ways of treatments, and size of NP. The study also indicated that availability of TiO2 NP for a plant is different when provided through foliar treatment than through soil or water solution. The positive influence of TiO2 NP was correlated with the photocatalytic activity of Ti, but the mechanism behind this interaction is so far not understood.
Some of the Notable Studies on the Impact of Other Metal and Metal Oxide Nanoparticles
Several other nanoparticles are used in different industries to improve the product/service quality. Some of these nanoparticles are used in plant study to observe their effects on plants (Table 4). Cerium dioxide nanoparticles (CeO2 NP) are mainly used in automotive industries and semiconductor industry, and can interfere with the cell metabolism due to its oxidative properties. CeO2 NP was observed to behave differently in tomato plant when coated with citric acid, in comparison to bare CeO2 NP (Barrios et al., 2016). Coated NP was observed to increase chlorophyll concentration, stem length, and catalase activity. The change in behavior of NP can be attributed to different chemical properties and size of the nanoparticle due to the presence of the cap. A different study using CeO2 in the presence of soil organic matter confirms that the surrounding and composition influences the behavior of nanoparticle for plant (Majumdar et al., 2016). Field and soil experiments with wheat and rice have shown that application of CeO2 NP compromised the quality of grain (Rico et al., 2013; Du et al., 2015). The hydroponic experiment performed on cotton shows that CeO2 NP destroys the vascular bundles in cotton with a decrease in indole-3-acetic acid and abscisic acid (Nhan et al., 2015). The authors also observed that conventional cotton was more sustainable to CeO2 NP stress in comparison to transgenic cotton. Nickel oxide nanoparticle (NiO NP) was observed to induce ROS, and antioxidant molecules whereas, it was observed to reduce the concentration of plant pigments (Faisal et al., 2013; Soares et al., 2016). Zinc (Zn) and Zinc oxide nanoparticles (ZnO NP) affected plant germination and had negative effects on root elongation (Lin and Xing, 2007, 2008). ZnO NP was also observed to reduce photosynthetic efficiency and antioxidant activity whereas it induces the ROS production in the wheat plant (Tripathi et al., 2017). DNA fragmentation was also observed due to ZnO NP toxicity in Allium cepa (Ghosh et al., 2016). Treatment of ZnO NP at 1,000 ppm concentration was observed to promote seed germination and seedling vigor, and in turn showed early establishment in soil manifested by early flowering and higher leaf chlorophyll content, but the higher concentration of ZnO NP at 2,000 ppm was observed to have negative and toxic effect on the growth and yield of peanut (Prasad et al., 2012). In turn, Stampoulis et al. (2009) did not observe any significant effects of ZnO on Cucurbita pepo in studied concentration. Burklew et al. (2012) observed an increase in the expression of different stress related micro RNA in tobacco plant when exposed to Aluminum oxide nanoparticle (Al2O3 NP). NiO NP was observed to induce the apoptosis and promote the release of caspase-3 proteases from mitochondria (Faisal et al., 2013). The authors also observed that nitric oxide ameliorates the toxic effect of nanoparticle. The Iron oxide nanoparticles (Fe3O4 NP) at lower concentrations were observed to have beneficiary impact on plant and improves germination (Iannone et al., 2016; Li et al., 2016), whereas cadmium oxide nanoparticles (CdO NP) at low concentration were found to increase total amino acid production without influencing photosynthetic parameters (Vecerova et al., 2016)
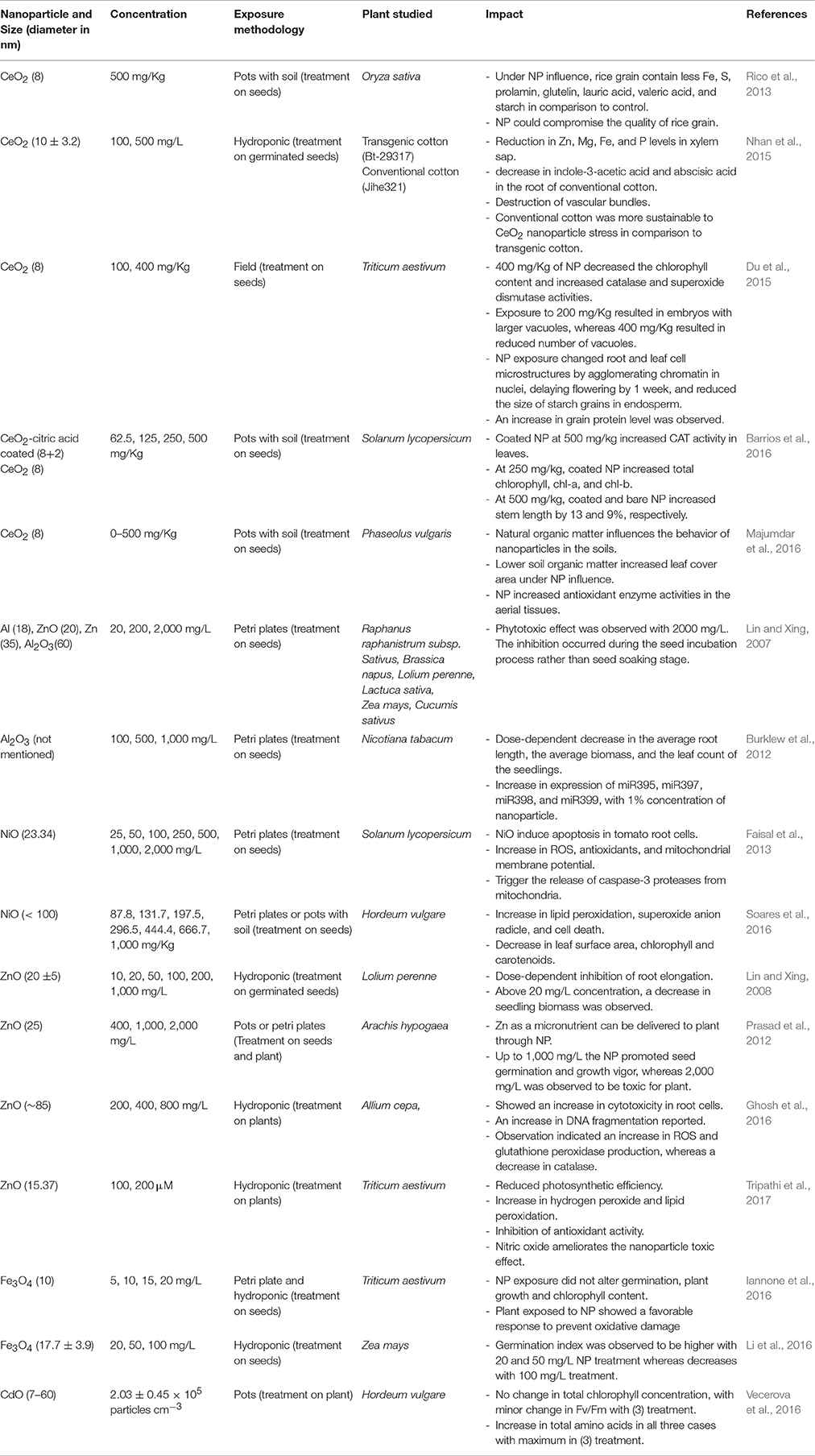
Table 4. Impact of few other important metal and metal oxide NPs on plants.
The study clearly indicates that the presence of industrial nanoparticles in the environment influence the plants. Despite the positive effects of some NPs, the studies clearly indicate that all kinds of nanoparticles represent the possible environmental risk.
Mechanism of Nanoparticle-Plant Interaction
Based on the scientific works performed, it is evident that most of the nanoparticles are toxic to the plants in high concentration. It is hypothesized that, for exhibiting the toxic effect the uptake of nanoparticles by plant and their translocation into different tissues is needed. Further, based on their transportation, properties, and reactivity, the nanoparticles may interfere with different metabolic activity to produce an impact on plants.
Nanoparticle Uptake
The nanoparticles when present in higher concentrations are observed to damage the plant cell wall and plasma membrane, thus penetrating it and interacting with the different plant's processes (Mazumdar and Ahmed, 2011; Mirzajani et al., 2013). Nanoparticles can enter plant tissue either through root or the above ground parts including root junctions and wounds. For uptake and translocation nanoparticles has to go through various chemical and physiological barriers. When nanoparticles interact with plant, cell wall is the first barrier it has to cross. Plant cell walls are a structure which is composed of cellulose which permits the entry of small particles and restricting the larger one, therefore smaller nanoparticles can go through this layer in a comparatively easy way in respect to larger nanoparticles. The size exclusion limit for the plant cell wall is between 5 and 20 nm (Dietz and Herth, 2011). Some of the nanoparticles have been reported to induce the formation of larger pores in cell wall which further facilitate the entry of large nanoparticles (Navarro et al., 2008; Kurepa et al., 2010). From the cell wall the nanoparticles may move through endocytosis (Etxeberria et al., 2006), and further, through the symplastic transport, it may travel to different plant tissues (Ma et al., 2010). Recently, Wong et al. (2016) have proposed a mathematical model which indicates lipid exchange mechanism for nanoparticle transport inside the plant cells. The study indicated that size, magnitude, and zeta potentials are key in determining the transport of nanoparticle inside the plant.
Nanoparticle-Plant Interaction Pathways
Nanoparticles may interfere with plant metabolism in several ways, such as, by providing micronutrients (Liu and Lal, 2015), regulation of genes (Nair and Chung, 2014), or interfering with different oxidative processes in plants which results in oxidative burst (Figure 2; Hossain et al., 2015). From the previous part of this article, it is clear that several nanoparticles when present in excess results into ROS production, and interfere with the oxidative mechanism, whereas other types of interactive pathways are not deciphered and there is still much work needed to understand the other pathways. Therefore, the oxidative part is elaborated in Figure 2 and further discussed here.
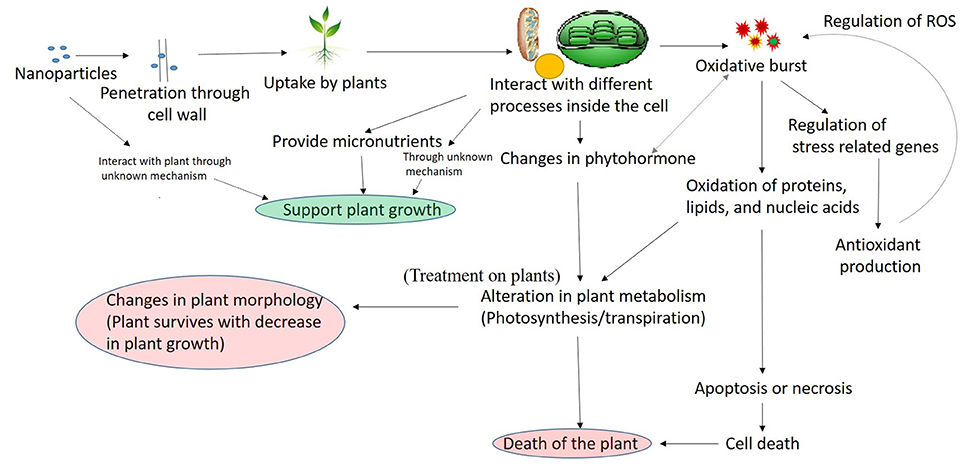
Figure 2. A general mechanism of nanoparticles interaction with plant.
The entered nanoparticles can interfere with electron transport chain of mitochondria and chloroplast, which may results into the oxidative burst, observed by the increase in ROS concentration (Dimkpa et al., 2013; Faisal et al., 2013; Jiang et al., 2014; Pakrashi et al., 2014; Cvjetko et al., 2017). It has been previously reported that under influence of different stress factors, the rate of carbon fixation is limited, which causes an increase in photoinhibition potentially steering the photosystem toward overproduction of superoxide anion radicals and H2O (Foyer and Noctor, 2005). Once the ROS is produced as the result of nanoparticle interaction, it is known that it interacts with almost all cellular components producing protein modifications, lipid peroxidation, and damage to DNA (Van Breusegem and Dat, 2006). Several reports have shown an increase in lipid peroxidation and DNA damage in plants-nanoparticle interaction (Atha et al., 2012; Belava et al., 2017; Cvjetko et al., 2017; Saha and Dutta Gupta, 2017), which confirms that plant interaction with nanoparticles leads into lipid peroxidation. The increased production of ROS can either induce apoptosis or necrosis (Van Breusegem and Dat, 2006; Rastogi and Pospíšil, 2012; Faisal et al., 2013), which results in plant cell death. Despite their destructive activity, ROS are also known to have a signaling role in a variety of cellular processes including tolerance to environmental stresses (Sharma et al., 2012). The destructive or signaling role of ROS depends on the equilibrium between ROS production and scavenging. Due to the multifunctional roles of ROS, the cells have developed a strong antioxidant mechanism to precisely control the level of ROS. The antioxidant mechanism contains the production of enzymatic (superoxide dismutase, catalase, and guaiacol peroxidase) and non-enzymatic (Ascorbate, glutathione, carotenoids, tocopherols, and phenolics) molecules (Sharma et al., 2012). To cope up with the stress plants increases the production of antioxidant molecules (Rastogi and Pospíšil, 2010; Sharma et al., 2012). Several reports have shown the increased production of the antioxidant molecule in the plant under the influence of nanoparticles (Faisal et al., 2013; Jiang et al., 2014; Costa and Sharma, 2016), which confirms the regulation of antioxidant system as a response to nanoparticle interaction with plant. If the antioxidant produced are unable to control the ROS, The ROS oxidized the cell macromolecules and results into the death of the cell by apoptosis or necrosis (Sharma et al., 2012), which ultimately results in the death of the plant. The recent reports have also shown that phytohormone plays an important role in plant stress response signaling (O'Brien and Benková, 2013). The hormonal control of plant development and stress adaptation is considered to be the outcome of a complex network of synergistic and antagonistic interactions between various hormones. The ROS are also linked to hormonal signaling in a complex manner and influence each other activity (Kwak et al., 2006). Different hormonal pathways are known to be upregulated or downregulated in response to different type of stresses (O'Brien and Benková, 2013). The observed increase in cytokinin level in Capsicum annuum as a response to AgNP stress, and a decreases in IAA and ABA in a cotton plant in response to CuO NP indicates that nanoparticle influence the hormonal balance in plants, thus affecting the plant metabolism. Therefore, it can be said that the toxic effect of nanoparticles in the plant is mainly mediated through ROS.
The electron transport chain in mitochondria and chloroplast operates in aerobic environment, and thus the excess production of ROS impact the processes (Foyer and Shigeoka, 2011). Photosynthesis is also considered to be a good measure of overall performance of plants (Kalaji et al., 2014). It is the only energy input in plants and thereby impacts all aspects of plant metabolism and physiology. Thus, the measurement of photosynthetic pigment and activity is a good measure to access the impact of stress factors. Different reports have shown that the nanoparticles influence the photosynthetic pigment concentration and its activity in plants (Qian et al., 2013; Perreault et al., 2014; Tripathi et al., 2017). A very high concentration of nanoparticles may severely affect the photosynthesis which may result in plant growth suppression or plant death. Several reports have observed significant decrease in plant growth as the result of nanoparticle exposure (For reference see Tables 1–4). Root is the primary organ for up taking nanoparticles from soil/water are adversely affected in comparison to shoot in some plants (Pokhrel and Dubey, 2013; Qian et al., 2013; Shaw et al., 2014; Tripathi et al., 2017; Vinković et al., 2017).
Conclusion and Future Perspective
In modern age, nanoparticles are used intensively and becoming a part of the human life. But due to the need of present and modern life, environment cannot be neglected. It is evident from the studies that metal and metal oxides nanoparticles in excess are harmful to plants, whereas, when present in traces it can be beneficial for plants. Therefore, the increasing concentration of nanoparticles in the environment may cause a serious impact on agriculture in future. This review took out clear information from known literature to shows an influence of metal and metal oxide nanoparticle on the plant, but there are needs of research to understand the molecular mechanism of plant nanoparticle interaction. There are few research showing the beneficial role of metal and metal oxide nanoparticles in agriculture, but the mechanism at large extant are not understood, and the studies are in its primitive stage. Therefore, a lot of study is needed before bringing the nanoparticles to the field. Most of the study performed shows morphological variation caused to plants due to metal and metal oxide nanoparticles. The study also shows a clear lack of standardization for nanoparticles phytotoxic assay. Therefore, research is needed to be done in the area to understand the impact of metal and metal oxides nanoparticle on plant physiology and molecular biology.
Nanoscience is attracting lot of research funding, some of which need to be diverted for the awareness of the people about the proper disposal of nanoparticle products. The research is also needed to be performed in the area of remediation of nanoparticle from agriculture soil and wastewater.
Author Contributions
AR, MB, and MZ discussed the idea. AR, MZ, OS, MB, and XH prepared the manuscript. HK and SM read and improved the manuscript, also helped AR in preparing the tables. AR prepared the figures. All authors read and worked on the scientific language of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer, ASP, and handling Editor declared their shared affiliation.
Acknowledgments
This work was supported by the research project of the Slovak Research and Development Agency under the project APVV-15-0721, VEGA 1/0923/16, and National Science Centre Poland, under the project UMO-2016/21/B/ST10/02271. AR thanks, Slovak Academic Information Agency (SAIA) for providing National Scholarship for research in the year 2017.
References
Adams, J., Wright, M., Wagner, H., Valiente, J., Britt, D., and Anderson, A. (2017). Cu from dissolution of CuO nanoparticles signals changes in root morphology. Plant Physiol. Biochem. 110, 108–117. doi: 10.1016/j.plaphy.2016.08.005
Adhikari, T., Kundu, S., Biswas, A. K., Tarafdar, J. C., and Rao, A. S. (2012). Effect of copper oxide nano particle on seed germination of selected crops. J. Agric. Sci. Technol. A 2, 815–823. doi: 10.17265/2161-6256/2012.06A.009
Asli, S., and Neumann, P. M. (2009). Colloidal suspensions of clay or titanium dioxide nanoparticles can inhibit leaf growth and transpiration via physical effects on root water transport. Plant Cell Environ. 32, 577–584. doi: 10.1111/j.1365-3040.2009.01952.x
Atha, D. H., Wang, H., Petersen, E. J., Cleveland, D., Holbrook, R. D., Jaruga, P., et al. (2012). Copper oxide nanoparticle mediated DNA damage in terrestrial plant models. Environ. Sci. Technol. 46, 1819–1827. doi: 10.1021/es202660k
Auffan, M., Rose, J., Bottero, J. Y., Lowry, G. V., Jolivet, J. P., and Wiesner, M. R. (2009). Towards a definition of inorganic nanoparticles from an environmental, health and safety perspective. Nat. Nanotechnol. 4, 634–641. doi: 10.1038/nnano.2009.242
Barrios, A. C., Rico, C. M., Trujillo-Reyes, J., Medina-Velo, I. A., Peralta-Videa, J. R., and Gardea-Torresdey, J. L. (2016). Effects of uncoated and citric acid coated cerium oxide nanoparticles, bulk cerium oxide, cerium acetate, and citric acid on tomato plants. Sci. Total Environ. 563–564, 956–964. doi: 10.1016/j.scitotenv.2015.11.143
Belava, V. N., Panyuta, O. O., Yakovleva, G. M., Pysmenna, Y. M., and Volkogon, M. V. (2017). The effect of silver and copper nanoparticles on the Wheat-Pseudocercosporella herpotrichoides pathosystem. Nanoscale Res. Lett. 12:250. doi: 10.1186/s11671-017-2028-6
Berahmand, A. A., Ghafariyan-Panahi, A., Sahabi, H., Feizi, H., Rezvani-Moghaddam, P., Shahtahmassebi, N., et al. (2012). Effects silver nanoparticles and magnetic field on growth of fodder maize (Zea mays L.). Biol. Trace Elem. Res. 149, 419–424. doi: 10.1007/s12011-012-9434-5
Boxall, P., Purcell, J., and Wright, P.M. (eds.). (2008). “Human resource management: scope, analysis and significance” in The Oxford Handbook of Human Resource Management (Oxford: Oxford University Press), 1–18. doi: 10.1093/oxfordhb/9780199547029.003.0001
Burklew, C. E., Ashlock, J., Winfrey, W. B., and Zhang, B. (2012). Effects of aluminum oxide nanoparticles on the growth, development, and microRNA expression of tobacco (Nicotiana tabacum). PLoS ONE 7:e34783. doi: 10.1371/journal.pone.0034783
Castiglione, M. R., Giorgetti, L., Geri, C., and Cremonini, R. (2011). The effects of nano-TiO2 on seed germination, development and mitosis of root tip cells of Vicia narbonensis L. and Zea mays L. J. Nanoparticle Res. 13, 2443–2449. doi: 10.1007/s11051-010-0135-8
Clément, L., Hurel, C., and Marmier, N. (2013). Toxicity of TiO2 nanoparticles to cladocerans, algae, rotifers and plants – effects of size and crystalline structure. Chemosphere 90, 1083–1090. doi: 10.1016/j.chemosphere.2012.09.013
Costa, M. V. J. D., and Sharma, P. K. (2016). Effect of copper oxide nanoparticles on growth, morphology, photosynthesis, and antioxidant response in Oryza sativa. Photosynthetica 54, 110–119. doi: 10.1007/s11099-015-0167-5
Cvjetko, P., Milošić, A., Domijan, A. M., Vinković-Vrček, I., Tolić, S., Peharec-Štefanić, P., et al. (2017). Toxicity of silver ions and differently coated silver nanoparticles in Allium cepa roots. Ecotoxicol. Environ. Saf. 137, 18–28. doi: 10.1016/j.ecoenv.2016.11.009
De la Torre Roche, R., Servin, A., Hawthorne, J., Xing, B., Newman, L. A., Ma, X., et al. (2015). Terrestrial trophic transfer of bulk and nanoparticle La2O3 does not depend on particle size. Environ. Sci. Technol. 49, 11866–11874. doi: 10.1021/acs.est.5b02583
Demir, E., Kaya, N., and Kaya, B. (2014). Genotoxic effects of zinc oxide and titanium dioxide nanoparticles on root meristem cells of Allium cepa by comet assay. Turk. J. Biol. 38, 31–39. doi: 10.3906/biy-1306-11
Dietz, K.-J., and Herth, S. (2011). Plant nanotoxicology. Trends Plant Sci. 16, 582–589. doi: 10.1016/j.tplants.2011.08.003
Dimkpa, C. O., McLean, J. E., Martineau, N., Britt, D. W., Haverkamp, R., and Anderson, A. J. (2013). Silver nanoparticles disrupt wheat (Triticum aestivum L.) growth in a sand matrix. Environ. Sci. Technol. 47, 1082–1090. doi: 10.1021/es302973y
Du, W., Gardea-Torresdey, J. L., Ji, R., Yin, Y., Zhu, J., Peralta-Videa, J. R., et al. (2015). Physiological and biochemical changes imposed by CeO2 nanoparticles on wheat: a life cycle field study. Environ. Sci. Technol. 49, 11884–11893. doi: 10.1021/acs.est.5b03055
Dumont, E., Johnson, A. C., Keller, V. D., and Williams, R. J. (2015). Nano silver and nano zinc-oxide in surface waters - exposure estimation for Europe at high spatial and temporal resolution. Environ. Pollut. 196, 341–349. doi: 10.1016/j.envpol.2014.10.022
El-Temsah, Y. S., and Joner, E. J. (2012). Ecotoxicological effects on earthworms of fresh and aged nano-sized zero-valent iron (nZVI) in soil. Chemosphere 89, 76–82. doi: 10.1016/j.chemosphere.2012.04.020
Etxeberria, E., Gonzalez, P., Pozueta-Romero, J., and Romero, J. P. (2006). Fluid phase endocytic uptake of artificial nano-spheres and fluorescent quantum dots by sycamore cultured cells: evidence for the distribution of solutes to different intracellular compartments. Plant Signal. Behav. 1, 196–200. doi: 10.4161/psb.1.4.3142
Faisal, M., Saquib, Q., Alatar, A. A., Al-Khedhairy, A. A., Hegazy, A. K., and Musarrat, J. (2013). Phytotoxic hazards of NiO-nanoparticles in tomato: a study on mechanism of cell death. J. Hazard. Mater. 250–251, 318–332. doi: 10.1016/j.jhazmat.2013.01.063
Feng, Y., Cui, X., He, S., Dong, G., Chen, M., Wang, J., et al. (2013). The role of metal nanoparticles in influencing arbuscular mycorrhizal fungi effects on plant growth. Environ. Sci. Technol. 47, 9496–9504. doi: 10.1021/es402109n
Foyer, C. H., and Noctor, G. (2005). Redox homeostasis and antioxidant signaling: a metabolic interface between stress perception and physiological responses. Plant Cell 17, 1866–1875. doi: 10.1105/tpc.105.033589
Foyer, C. H., and Shigeoka, S. (2011). Understanding oxidative stress and antioxidant functions to enhance photosynthesis. Plant Physiol. 155, 93–100. doi: 10.1104/pp.110.166181
Fraceto, L. F., Grillo, R., de Medeiros, G. A., Scognamiglio, V., Rea, G., and Bartolucci, C. (2016). Nanotechnology in agriculture: which innovation potential does it have? Front. Environ. Sci. 4:20. doi: 10.3389/fenvs.2016.00020
Gao, F., Liu, C., Qu, C., Zheng, L., Yang, F., Su, M., et al. (2008). Was improvement of spinach growth by nano-TiO(2) treatment related to the changes of Rubisco activase? Biometals 21, 211–217. doi: 10.1007/s10534-007-9110-y
Ghosh, M., Jana, A., Sinha, S., Jothiramajayam, M., Nag, A., Chakraborty, A., et al. (2016). Effects of ZnO nanoparticles in plants: cytotoxicity, genotoxicity, deregulation of antioxidant defenses, and cell-cycle arrest. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 807, 25–32. doi: 10.1016/j.mrgentox.2016.07.006
Hadioui, M., Merdzan, V., and Wilkinson, K. J. (2015). Detection and characterization of ZnO nanoparticles in surface and waste waters using single particle ICPMS. Environ. Sci. Technol. 49, 6141–6148. doi: 10.1021/acs.est.5b00681
Hawthorne, J., De la Torre Roche, R., Xing, B., Newman, L. A., Ma, X., Majumdar, S., et al. (2014). Particle-size dependent accumulation and trophic transfer of cerium oxide through a terrestrial food chain. Environ. Sci. Technol. 48, 13102–13109. doi: 10.1021/es503792f
Hawthorne, J., Musante, C., Sinha, S. K., and White, J. C. (2012). Accumulation and phytotoxicity of engineered nanoparticles to Cucurbita pepo. Int. J. Phytoremediation 14, 429–442. doi: 10.1080/15226514.2011.620903
Helaly, M. N., El-Hoseiny, H., El-Sheery, N. I., Rastogi, A., and Kalaji, H. M. (2017). Regulation and physiological role of silicon in alleviating drought stress of mango. Plant Physiol. Biochem. 118, 31–44. doi: 10.1016/j.plaphy.2017.05.021
Hossain, Z., Mustafa, G., and Komatsu, S. (2015). Plant responses to nanoparticle stress. Int. J. Mol. Sci. 16, 26644–26653. doi: 10.3390/ijms161125980
Iannone, M. F., Groppa, M. D., de Sousa, M. E., van Raap, M. B. F., and Benavides, M. P. (2016). Impact of magnetite iron oxide nanoparticles on wheat (Triticum aestivum L.) development: evaluation of oxidative damage. Environ. Exp. Bot. 131, 77–88. doi: 10.1016/j.envexpbot.2016.07.004
Jaberzadeh, A., Moaveni, P., Moghadam, H. R. T., and Zahedi, H. (2013). Influence of bulk and nanoparticles titanium foliar application on some agronomic traits, seed gluten and starch contents of wheat subjected to water deficit stress. Not. Bot. Horti Agrobo. 41, 201–207. doi: 10.15835/nbha4119093
Jasim, B., Thomas, R., Mathew, J., and Radhakrishnan, E. K. (2016). Plant growth and diosgenin enhancement effect of silver nanoparticles in Fenugreek (Trigonella foenum-graecum L.). Saudi Pharm. J. 25, 443–447. doi: 10.1016/j.jsps.2016.09.012
Jiang, H. S., Qiu, X. N., Li, G. B., Li, W., and Yin, L. Y. (2014). Silver nanoparticles induced accumulation of reactive oxygen species and alteration of antioxidant systems in the aquatic plant Spirodela polyrhiza. Environ. Toxicol. Chem. 33, 1398–1405. doi: 10.1002/etc.2577
Jo, Y. K., Kim, B. H., and Jung, G. (2009). Antifungal activity of silver ions and nanoparticles on phytopathogenic fungi. Plant Dis. 93, 1037–1043. doi: 10.1094/PDIS-93-10-1037
Johansson, M., Pell, M., and Stenström, J. (1998). Kinetics of SubstrateInduced Respiration (SIR) and denitrification: applications to a soil amended with silver. Ambio 27, 40–44.
Joner, E. J., Hartnik, T., and Amundsen, C. E. (2008). Norwegian Pollution Control Authority Report no. TA 2304/2007. Bioforsk; Environmental fate and ecotoxicity of engineered nanoparticles, 1–64.
Judy, J. D., Unrine, J. M., Rao, W., and Bertsch, P. M. (2012). Bioaccumulation of gold nanomaterials by Manduca sexta through dietary uptake of surface contaminated plant tissue. Environ. Sci. Technol. 46, 12672–12678. doi: 10.1021/es303333w
Kalaji, H. M., Schansker, G., Ladle, R. J., Goltsev, V., Bosa, K., Allakhverdiev, S. I., et al. (2014). Frequently asked questions about in vivo chlorophyll fluorescence: practical issues. Photosyn. Res. 122, 121–158. doi: 10.1007/s11120-014-0024-6
Kasana, R. C., Panwar, N. R., Kaul, R. K., and Kumar, P. (2017). Biosynthesis and effects of copper nanoparticles on plants. Environ. Chem. Lett. 15, 233–240. doi: 10.1007/s10311-017-0615-5
Keller, A. A., and Lazareva, A. (2014). Predicted releases of engineered nanomaterials: from global to regional to local. Environ. Sci. Technol. Lett. 1, 65–70. doi: 10.1021/ez400106t
Khaydarov, R. R., Khaydarov, R. A., Gapurova, O., Estrin, Y., Evgrafova, S., Scheper, T., et al. (2009). “Antimicrobial effects of silver nanoparticles synthesized by an electrochemical method,” in Nanostructured Material for Advanced Technological Applications, NATO Science for Peace and Security Series B: Physics and Biophysics, Part 5, eds J. P. Reithmaier, P. Petkov, W. Kulisch, and C. Popov (Dordrecht: Springer), 215–218.
Krishnaraj, C., Ramachandran, R., Mohan, K., and Kalaichelvan, P. T. (2012). Optimization for rapid synthesis of silver nanoparticles and its effect on phytopathogenic fungi. Spectrochim. Acta A Mol. Biomol. Spectrosc. 93, 95–99. doi: 10.1016/j.saa.2012.03.002
Kurepa, J., Paunesku, T., Vogt, S., Arora, H., Rabatic, B. M., Lu, J., et al. (2010). Uptake and distribution of ultrasmall anatase TiO2 Alizarin red S nanoconjugates in Arabidopsis thaliana. Nano Lett. 10, 2296–2302. doi: 10.1021/nl903518f
Kwak, J. M., Nguyen, V., and Schroeder, J. I. (2006). The role of reactive oxygen species in hormonal responses. Plant Physiol. 141, 323–329. doi: 10.1104/pp.106.079004
Lahiani, M. H., Chen, J., Irin, F., Puretzky, A. A., Green, M. J., and Khodakovskaya, M. V. (2015). Interaction of carbon nanohorns with plants: uptake and biological effects. Carbon N. Y. 81, 607–619. doi: 10.1016/j.carbon.2014.09.095
Landa, P., Cyrusova, T., Jerabkova, J., Drabek, O., Vanek, T., and Podlipna, R. (2016). Effect of metal oxides on plant germination: phytotoxicity of nanoparticles, bulk materials, and metal ions. Water Air Soil 227, 448. doi: 10.1007/s11270-016-3156-9
Larue, C., Laurette, J., Herlin-Boime, N., Khodja, H., Fayard, B., Flank, A. M., et al. (2012). Accumulation, translocation and impact of TiO2 nanoparticles in wheat (Triticum aestivum spp.): influence of diameter and crystal phase. Sci. Total Environ. 431, 197–208. doi: 10.1016/j.scitotenv.2012.04.073
Lee, W. M., An, Y. J., Yoon, H., and Kweon, H. S. (2008). Toxicity and bioavailability of copper nanoparticles to the terrestrial plants mung bean (Phaseolus radiatus) and wheat (Triticum aestivum): plant agar test for water-insoluble nanoparticles. Environ. Toxicol. Chem. 27, 1915–1921. doi: 10.1897/07-481.1
Levard, C., Hotze, E. M., Lowry, G. V., and Brown, G. E. Jr. (2012). Environmental transformations of silver nanoparticles: impact on stability and toxicity. Environ. Sci. Technol. 46, 6900–6914. doi: 10.1021/es2037405
Li, J., Hu, J., Ma, C., Wang, Y., Wu, C., Huang, J., et al. (2016). Uptake, translocation and physiological effects of magnetic iron oxide (γ-Fe2O3) nanoparticles in corn (Zea mays L.). Chemosphere 159, 326–334. doi: 10.1016/j.chemosphere.2016.05.083
Lin, D., and Xing, B. (2007). Phytotoxicity of nanoparticles: inhibition of seed germination and root growth. Environ. Pollut. 150, 243–250. doi: 10.1016/j.envpol.2007.01.016
Lin, D., and Xing, B. (2008). Root uptake and phytotoxicity of ZnO nanoparticles. Environ. Sci. Technol. 42, 5580–5585. doi: 10.1021/es800422x
Liu, J., and Hurt, R. H. (2010). Ion release kinetics and particle persistence in aqueous nano-silver colloids. Environ. Sci. Technol. 44, 2169–2175. doi: 10.1021/es9035557
Liu, R., and Lal, R. (2015). Potentials of engineered nanoparticles as fertilizers for increasing agronomic productions. Sci. Total Environ. 514, 131–139. doi: 10.1016/j.scitotenv.2015.01.104
Ma, X., Geisler-Lee, J., Deng, Y., and Kolmakov, A. (2010). Interactions between engineered nanoparticles (ENPs) and plants: phytotoxicity, uptake and accumulation. Sci. Total Environ. 408, 3053–3061. doi: 10.1016/j.scitotenv.2010.03.031
Mahdi, K. N. M., Petersc, R. J. B., Klumpp, E., Bohme, S., Ploeg, M. V. D., Ritsema, C., et al. (2017). Silver nanoparticles in soil: aqueous extraction combined with single-particle ICP-MS for detection and characterization. Environ. Nanotechnol. Monit. Manage 7, 24–33. doi: 10.1016/j.enmm.2016.12.002
Majumdar, S., Peralta-Videa, J. R., Trujillo-Reyes, J., Sun, Y., Barrios, A. C., Niu, G., et al. (2016). Soil organic matter influences cerium translocation and physiological processes in kidney bean plants exposed to cerium oxide nanoparticles. Sci. Total Environ. 569–570, 201–211. doi: 10.1016/j.scitotenv.2016.06.087
Martin-Ortigosa, S., Peterson, D. J., Valenstein, J. S., Lin, V. S., Trewyn, B. G., Lyznik, L. A., et al. (2014). Mesoporous silica nanoparticle-mediated intracellular cre protein delivery for maize genome editing via loxP site excision. Plant Physiol. 164, 537–547. doi: 10.1104/pp.113.233650
Maruyama, C. R., Guilger, M., Pascoli, M., Bileshy-José, N., Abhilash, P. C., Fraceto, L. F., et al. (2016). Nanoparticles based on chitosan as carriers for the combined herbicides imazapic and imazapyr. Sci. Rep. 6:19768. doi: 10.1038/srep19768
Maurer-Jones, M. A., Gunsolus, I. L., Murphy, C. J., and Haynes, C. L. (2013). Toxicity of engineered nanoparticles in the environment. Anal. Chem. 85, 3036–3049. doi: 10.1021/ac303636s
Mazumdar, H., and Ahmed, G. U. (2011). Phytotoxicity effect of silver nanoparticles on Oryza sativa. Int. J. ChemTech. Res. 3, 1494–1500. Available online at: http://sphinxsai.com/Vol.3No.3/Chem/chVol.3No.3J_S11_8.htm
Mirzajani, F., Askari, H., Hamzelou, S., Farzaneh, M., and Ghassempour, A. (2013). Effect of silver nanoparticles on Oryza sativa L. and its rhizosphere bacteria. Ecotoxicol. Environ. Saf. 88, 48–54. doi: 10.1016/j.ecoenv.2012.10.018
Nair, P. M., and Chung, I. M. (2014). Impact of copper oxide nanoparticles exposure on Arabidopsis thaliana growth, root system development, root lignificaion, and molecular level changes. Environ. Sci. Pollut. Res. Int. 21, 12709–12022. doi: 10.1007/s11356-014-3210-3
Nair, R. (2016). “Effects of nanoparticles on plant growth and development,” in Plant Nanotechnology, eds C. Kole, D. S. Kumar, and M. V. Khodakovskaya (Cham: Springer), 95–118.
Navarro, E., Piccapietra, F., Wagner, B., Marconi, F., Kaegi, R., Odzak, N., et al. (2008). Toxicity of silver nanoparticles to Chlamydomonas reinhardtii. Environ. Sci. Technol. 42, 8959–8964. doi: 10.1021/es801785m
Navratilova, J., Praetorius, A., Gondikas, A., Fabienke, W., von-der Kammer, F., and Hofmann, T. (2015). Detection of engineered copper nanoparticles in soil using single particle ICP-MS. Int. J. Environ. Res. Public Health 12, 15756–15768. doi: 10.3390/ijerph121215020
Nekrasova, G. F., Ushakova, O. S., Ermakov, A. E., and Uimin, M. A. (2011). Effects of copper (II) ions and copper oxide nanoparticles on Elodea densa Planch. Russ. J. Ecol. 42, 458–463. doi: 10.1134/S1067413611060117
Nhan, L. V., Ma, C., Rui, Y., Liu, S., Li, X., Xing, B., et al. (2015). Phytotoxic mechanism of nanoparticles: destruction of chloroplasts and vascular bundles and alteration of nutrient absorption. Sci. Rep. 5:11618. doi: 10.1038/srep11618
O'Brien, J. A., and Benková, E. (2013). Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 4:451. doi: 10.3389/fpls.2013.00451
Okupnik, A., and Pflugmacher, S. (2016). Oxidative stress response of the aquatic macrophyte Hydrilla verticillata exposed to TiO2 nanoparticles. Environ. Toxicol. Chem. 35, 2859–2866. doi: 10.1002/etc.3469
Pakrashi, S., Jain, N., Dalai, S., Jayakumar, J., Chandrasekaran, P. T., Raichur, A. M., et al. (2014). In vivo genotoxicity assessment of titanium dioxide nanoparticles by Allium cepa root tip assay at high exposure concentrations. PLoS ONE 9:e87789. doi: 10.1371/journal.pone.0087789
Pallavi Mehta, C. M., Srivastava, R., Arora, S., and Sharma, A. K. (2016). Impact assessment of silver nanoparticles on plant growth and soil bacterial diversity. Biotech 6:254. doi: 10.1007/s13205-016-0567-7
Patlolla, A. K., Berry, A., May, L., and Tchounwou, P. B. (2012). Genotoxicity of silver nanoparticles in Vicia faba: a pilot study on the environmental monitoring of nanoparticles. Int. J. Environ. Res. Public Health 9, 1649–1662. doi: 10.3390/ijerph9051649
Perreault, F., Samadani, M., and Dewez, D. (2014). Effect of soluble copper released from copper oxide nanoparticles solubilisation on growth and photosynthetic processes of Lemna gibba L. Nanotoxicology 8, 374–382. doi: 10.3109/17435390.2013.789936
Pokhrel, L. R., and Dubey, B. (2013). Evaluation of developmental responses of two crop plants exposed to silver and zinc oxide nanoparticles. Sci. Total Environ. 452–453, 321–332. doi: 10.1016/j.scitotenv.2013.02.059
Prasad, T. N. V. K. V., Sudhakar, P., Sreenivasulu, Y., Latha, P., Munaswamy, V., Raja Reddy, K., et al. (2012). Effect of nanoscale zinc oxide particles on the germination, growth and yield of peanut. J. Plant Nutr. 35, 906–927. doi: 10.1080/01904167.2012.663443
Qian, H., Peng, X., Han, X., Ren, J., Sun, L., and Fu, Z. (2013). Comparison of the toxicity of silver nanoparticles and silver ions on the growth of terrestrial plant model Arabidopsis thaliana. J. Environ. Sci. 25, 1947–1955. doi: 10.1016/S1001-0742(12)60301-5
Radad, K., Al-Shraim, M., Moldzio, R., and Rausch, W. D. (2012). Recent advances in benefits and hazards of engineered nanoparticles. Environ. Toxicol. Pharmacol. 34, 661–672. doi: 10.1016/j.etap.2012.07.011
Rafique, R., Arshad, M., Khokhar, M. F., Qazi, I. A., Hamza, A., and Virk, N. (2014). Growth response of wheat to titania nanoparticles application. NUST J. Engin. Sci. 7, 42–46. Available online at: http://journals.nust.edu.pk/index.php/njes/article/view/133
Rai, M., Yadav, A., and Gade, A. (2009). Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 27, 76–83. doi: 10.1016/j.biotechadv.2008.09.002
Raliya, R., Nair, R., Chavalmane, S., Wang, W. N., and Biswas, P. (2015). Mechanistic evaluation of translocation and physiological impact of titanium dioxide and zinc oxide nanoparticles on the tomato (Solanum lycopersicum L.) plant. Metallomics 7, 1584–1594. doi: 10.1039/C5MT00168D
Rastogi, A., and Pospíšil, P. (2010). Effect of exogenous hydrogen peroxide on biophoton emission from radish root cells. Plant Physiol. Biochem. 48, 117–123. doi: 10.1016/j.plaphy.2009.12.011
Rastogi, A., and Pospíšil, P. (2012). Production of hydrogen peroxide and hydroxyl radical in potato tuber during the necrotrophic phase of hemibiotrophic pathogen Phytophthora infestans infection. J. Photochem. Photobiol. B Biol. 117, 202–206. doi: 10.1016/j.jphotobiol.2012.10.001
Regier, N., Cosio, C., von-Moos, N., and Slaveykova, V. I. (2015). Effects of copper-oxide nanoparticles, dissolved copper and ultraviolet radiation on copper bioaccumulation, photosynthesis and oxidative stress in the aquatic macrophyte Elodea nuttallii. Chemosphere 128, 56–61. doi: 10.1016/j.chemosphere.2014.12.078
Rico, C. M., Majumdar, S., Duarte-Gardea, M., Peralta-Videa, J. R., and Gardea-Torresdey, J. L. (2011). Interaction of nanoparticles with edible plants and their possible implications in the food chain. J. Agric. Food Chem. 59, 3485–3498. doi: 10.1021/jf104517j
Rico, C. M., Morales, M. I., Barrios, A. C., McCreary, R., Hong, J., Lee, W. Y., et al. (2013). Effect of cerium oxide nanoparticles on the quality of rice (Oryza sativa L.) grains. J. Agric. Food Chem. 61, 11278–11285. doi: 10.1021/jf404046v
Saha, N., and Dutta Gupta, S. (2017). Low-dose toxicity of biogenic silver nanoparticles fabricated by Swertia chirata on root tips and flower buds of Allium cepa. J. Hazard. Mater. 330, 18–28. doi: 10.1016/j.jhazmat.2017.01.021
Sanchez-Dominguez, M., Boutonnet, M., and Solans, C. (2009). A novel approach to metal and metal oxide nanoparticle synthesis: the oil-in-water microemulsion reaction method. J. Nanopart. Res. 11, 1823. doi: 10.1007/s11051-009-9660-8
Santos, C. S. C., Gabriel, B., Blanchy, M., Menes, O., García, D., Blanco, M., et al. (2015). Industrial applications of nanoparticles – a prospective overview. Mater. Today Proc. 2, 456–465. doi: 10.1016/j.matpr.2015.04.056
Sharma, P., Jha, A. B., Dubey, R. S., and Pessarakli, M. (2012). Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012:217037. doi: 10.1155/2012/217037
Shaw, A. K., Ghosh, S., Kalaji, H. M., Bosa, K., Brestic, M., Zivcak, M., et al. (2014). Nano-CuO stress induced modulation of antioxidative defense and photosynthetic performance of syrian barley (Hordeum vulgare L.). Environ. Exp. Bot. 102, 37–47. doi: 10.1016/j.envexpbot.2014.02.016
Shi, J., Peng, C., Yang, Y., Yang, J., Zhang, H., Yuan, X., et al. (2014). Phytotoxicity and accumulation of copper oxide nanoparticles to the Cu-tolerant plant Elsholtzia splendens. Nanotoxicology 8, 179–188. doi: 10.3109/17435390.2013.766768
Slomberg, D. L., and Schoenfisch, M. H. (2012). Silica nanoparticle phytotoxicity to Arabidopsis thaliana. Environ. Sci. Technol. 46, 10247–10254. doi: 10.1021/es300949f
Soares, C., Branco-Neves, S., de-Sousa, A., Pereira, R., and Fidalgo, F. (2016). Ecotoxicological relevance of nano-NiO and acetaminophen to Hordeum vulgare L.: combining standardized procedures and physiological endpoints. Chemosphere 165, 442–452. doi: 10.1016/j.chemosphere.2016.09.053
Song, G., Hou, W., Gao, Y., Wang, Y., Lin, L., Zhang, Z., et al. (2016). Effects of CuO nanoparticles on Lemna minor. Bot. Stud. 57:3. doi: 10.1186/s40529-016-0118-x
Stampoulis, D., Sinha, S. K., and White, J. C. (2009). Assay-dependent phytotoxicity of nanoparticles to plants. Environ. Sci. Technol. 43, 9473–9479. doi: 10.1021/es901695c
Tangaa, S. R., Selck, H., Winther-Nielsen, N., and Khan, F. R. (2016). Trophic transfer of metal-based nanoparticles in aquatic environments: a review and recommendations for future research focus. Environ. Sci. Nano. 3, 966–981. doi: 10.1039/C5EN00280J
Teske, S. S., and Detweiler, C. S. (2015). The biomechanisms of metal and metal-oxide nanoparticles' interactions with cells. Int. J. Environ. Res. Public Health 12, 1112–1134. doi: 10.3390/ijerph120201112
Torney, F., Trewyn, B. G., Lin, V. S., and Wang, K. (2007). Mesoporous silica nanoparticles deliver DNA and chemicals into plants. Nat. Nanotechnol. 2, 295–300. doi: 10.1038/nnano.2007.108
Tripathi, D. K., Singh, S., Singh, S., Srivastava, P. K., Singh, V. P., Singh, S., et al. (2017). Nitric oxide alleviates silver nanoparticles (AgNps)-induced phytotoxicity in Pisum sativum seedlings. Plant Physiol. Biochem. 110, 167–177. doi: 10.1016/j.plaphy.2016.06.015
Tripathi, D. K., Singh, V. P., Prasad, S. M., Chauhan, D. K., and Dubey, N. K. (2015). Silicon nanoparticles (SiNp) alleviate chromium (VI) phytotoxicity in Pisum sativum (L.) seedlings. Plant Physiol. Biochem. 96, 189–198. doi: 10.1016/j.plaphy.2015.07.026
Tubana, B. S., Babu, T., and Datnoff, L. E. (2016). A review of silicon in soils and plants and its role in US agriculture: history and future perspectives. Soil Sci. 181, 393–411. doi: 10.1097/SS.0000000000000179
Unrine, J. M., Shoults-Wilson, W. A., Zhurbich, O., Bertsch, P. M., and Tsyusko, O. V. (2012). Trophic transfer of Au nanoparticles from soil along a simulated terrestrial food chain. Environ. Sci. Technol. 46, 9753–9760. doi: 10.1021/es3025325
Van, N. L., Ma, C., Shang, J., Rui, Y., Liu, S., and Xing, B. (2016). Effects of CuO nanoparticles on insecticidal activity and phytotoxicity in conventional and transgenic cotton. Chemosphere 144, 661–670. doi: 10.1016/j.chemosphere.2015.09.028
Van Breusegem, F., and Dat, J. F. (2006). Reactive oxygen species in plant cell death. Plant Physiol. 141, 384–390. doi: 10.1104/pp.106.078295
Vecerova, K., Večeřa, Z., Dočekal, B., Oravec, M., Pompeiano, A., Tříska, J., et al. (2016). Changes of primary and secondary metabolites in barley plants exposed to CdO nanoparticles. Environ. Pollut. 218, 207–218. doi: 10.1016/j.envpol.2016.05.013
Vinković, T., Novák, O., Strnad, M., Goessler, W., Jurašin, D. D., Paradiković, N., et al. (2017). Cytokinin response in pepper plants (Capsicum annuum L.) exposed to silver nanoparticles. Environ. Res. 156, 10–18. doi: 10.1016/j.envres.2017.03.015
Wang, P., Lombi, E., Zhao, F. J., and Kopittke, P. M. (2016). Nanotechnology: a new opportunity in plant sciences. Trends Plant Sci. 21, 699–712. doi: 10.1016/j.tplants.2016.04.005
Wang, Z., Xu, L., Zhao, J., Wang, X., White, J. C., and Xing, B. (2016). CuO Nanoparticle interaction with Arabidopsis thaliana: toxicity, parent-progeny transfer, and gene expression. Environ. Sci. Technol. 50, 6008–6016. doi: 10.1021/acs.est.6b01017
Wijnhoven, S. W. P., Peijnenburg, W. J. G. M., Herberts, C. A., Hagens, W. I., Oomen, A. G., Heugens, E. H. W., et al. (2009). Nano-silver - a review of available data and knowledge gaps in human and environmental risk assessment. Nanotoxicology 3, 109–138. doi: 10.1080/17435390902725914
Wong, M. H., Misra, R. P., Giraldo, J. P., Kwak, S. Y., Son, Y., Landry, M. P., et al. (2016). Lipid Exchange Envelope Penetration (LEEP) of nanoparticles for plant engineering: a universal localization mechanism. Nano Lett. 16, 1161–1172. doi: 10.1021/acs.nanolett.5b04467
Yadav, S. K. (2010). Heavy metals toxicity in plants: an overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 76, 167–179. doi: 10.1016/j.sajb.2009.10.007
Yin, L., Colman, B. P., McGill, B. M., Wright, J. P., and Bernhardt, E. S. (2012). Effects of silver nanoparticle exposure on germination and early growth of eleven wetland plants. PLoS ONE 7:e47674. doi: 10.1371/journal.pone.0047674
Zuverza-Mena, N., Armendariz, R., Peralta-Videa, J. R., and Gardea-Torresdey, J. L. (2016). Effects of silver nanoparticles on radish sprouts: root growth reduction and modifications in the nutritional value. Front. Plant Sci. 7:90. doi: 10.3389/fpls.2016.00090
Zuverza-Mena, N., Martínez-Fernández, D., Du, W., Hernandez-Viezcas, J. A., Bonilla-Bird, N., López-Moreno, M. L., et al. (2017). Exposure of engineered nanomaterials to plants: insights into the physiological and biochemical responses-a review. Plant Physiol. Biochem. 110, 236–264. doi: 10.1016/j.plaphy.2016.05.037
Keywords: nanoparticles, nanotoxicology, oxidative stress, industrial pollutants, silver nanoparticles (Ag-NPs)
Citation: Rastogi A, Zivcak M, Sytar O, Kalaji HM, He X, Mbarki S and Brestic M (2017) Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 5:78. doi: 10.3389/fchem.2017.00078
Received: 03 July 2017; Accepted: 26 September 2017;
Published: 12 October 2017.
Edited by:
Leonardo Fernandes Fraceto, Universidade Estadual Paulista Júlio de Mesquita Filho (UNESP), BrazilReviewed by:
Anderson Espirito Santo Pereira, Universidade Estadual Paulista Júlio de Mesquita Filho (UNESP), BrazilJuliana Lischka Sampaio Mayer, Universidade Estadual de Campinas, Brazil
Vishal Tripathi, Banaras Hindu University, India
Copyright © 2017 Rastogi, Zivcak, Sytar, Kalaji, He, Mbarki and Brestic. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anshu Rastogi, YW5zaHVzbHNAZ21haWwuY29t
Marian Brestic, bWFyaWFuLmJyZXN0aWNAdW5pYWcuc2s=