 Nazia Zaffar1*
Nazia Zaffar1* Viktoriia Lovynska2,3
Viktoriia Lovynska2,3 Alla Samarska4
Alla Samarska4 Tobias Arnstadt4Olivier Pourret5
Tobias Arnstadt4Olivier Pourret5 Stéphan Firmin5Petr Baroš6
Stéphan Firmin5Petr Baroš6 Eliška Lyko Vachková6Martin Palušák7
Eliška Lyko Vachková6Martin Palušák7 Stanisław Wacławek7
Stanisław Wacławek7 Edgar Peiter8
Edgar Peiter8 Oliver Wiche4,5
Oliver Wiche4,5- 1Institute for Biosciences, Biology/Ecology Group, TU Bergakademie Freiberg, Freiberg, Germany
- 2Institute of Bio- and Geosciences: Agrosphere (IBG-3), Forschungszentrum Jülich GmbH, Jülich, Germany
- 3Laboratory of Forestry, Dnipro State Agrarian and Economic University, Dnipro, Ukraine
- 4Applied Geoecology Group, Faculty of Natural and Environmental Sciences, University of Applied Sciences Zittau/Görlitz, Zittau, Germany
- 5Institut Polytechnique UniLaSalle, AGHYLE, Beauvais, France
- 6Department of Sustainable Fuels and Green Chemistry, University of Chemistry and Technology Prague, Prague, Czechia
- 7Institute for Nanomaterials, Advanced Technologies and Innovation, Technical University of Liberec, Liberec, Czechia
- 8Plant Nutrition Laboratory, Institute of Agricultural and Nutritional Sciences, Faculty of Natural Sciences III, Martin Luther University Halle-Wittenberg, Halle, Germany
This study investigates how sewage sludge and liquid digestate, as biosolid amendments, affect the mobility of cadmium (Cd), gallium (Ga), germanium (Ge), and rare earth elements (REEs) in soil, as well as their uptake by plants with differing nutritional strategies. Four species Alyssum murale, Lupinus albus, Fagopyrum esculentum, and Carthamus tinctorius were cultivated on unamended soil or soil amended with either sewage sludge or digestate. Shoot uptake of the essential elements P, Fe, Mn, Ni and of non-essential elements was evaluated alongside changes in ammonium-acetate-extractable (labile) element concentrations. For three species, root carboxylate exudation and rhizosphere acidification were also measured under variable phosphorus (P) supply conditions induced by the amendments. Both biosolids improved micronutrient availability across all species. However, increased shoot P concentrations were only observed in plants treated with sewage sludge. Digestate addition elevated total Ge (14.4%), labile Ga (178%), and labile REE (22%) concentrations in soil, while sewage sludge increased labile Cd (31%) and decreased labile REEs (18%) concentration. Neither amendment enhanced Ge uptake by plants. A higher proportion of labile Cd correlated with a higher Cd uptake in all tested plant species. However, the shoot net uptake of Ga and REE did not reflect their mobility in soil. More specifically, a higher Ga mobility in soil only increased Ga uptake in F. esculentum. F. esculentum acidified the rhizosphere and released fewer carboxylates under conditions of high P supply. Despite lower labile REEs concentrations in sewage sludge amended soil, L. albus and F. esculentum accumulated more REEs when the P supply was increased due to biosolids addition. The findings highlight that while Cd transfer is predictably linked to its labile soil pool, the uptake of Ge, Ga, and REEs depends on complex interactions between soil chemistry and plant-specific physiological traits responses to biosolid-derived nutrient inputs.
1 Introduction
The global production of sewage sludge and digestate from anaerobic fermentation is rising due to increasing urban populations (Rékási et al., 2019). As the main solid by-products from urban wastewater treatment and anaerobic digestion, these biosolids contain high concentrations of essential plant nutrients. Hence, biosolids represent valuable secondary raw materials for fertilizer production and are widely applied as organic fertilizers or soil conditioners (Pecorini et al., 2020; Jama-Rodzeńska et al., 2021; Kanteraki et al., 2022; Marchuk et al., 2023). However, biosolids also contain a diverse range of potentially toxic metal (loid)s (Marotrao et al., 2021; Kowalik et al., 2022; Zaffar et al., 2023) that can accumulate in soil and crops, often limiting their use in agriculture (Sinha et al., 2023; Alengebawy et al., 2021; Marchuk et al., 2023). Among these, cadmium (Cd), lead (Pb), chromium (Cr), and mercury (Hg) are the most extensively studied (Marotrao et al., 2021; Kowalik et al., 2022; Marchuk et al., 2023). In contrast, less attention has been paid to germanium (Ge), gallium (Ga), and rare earth elements (REEs: La-Lu, Sc,Y) despite their increasing detection in soils and plant biomass (Wiche et al., 2017; Okoroafor et al., 2022). Given their widespread occurrence and growing recognition as emerging environmental pollutants (Turcotte et al., 2022; Akarsu et al., 2023), as well as the potential role of REEs as beneficial elements (Qvarforth et al., 2025), it is important to assess how biosolid applications affect the plant availability not only of nutrients and commonly monitored metal (loid)s, but also of Ge, Ga, and REEs.
The plant availability of a given element depends on the complex interplay between soil-associated factors and plant-associated factors (Wiche et al., 2018). Soil-associated factors govern the solubility and chemical speciation of elements in soil, including soil pH, Eh, organic matter content, cation exchange capacity, and the distribution of elements in labile mineral and organic soil phases (Sheoran et al., 2016). In this regard, biosolid application may increase the concentrations of mobile and labile-bound elements in the soil when the elements contained are released from the organic matrix and/or when soil-born elements are mobilized through changes in pH and the increase in dissolved organic carbon following biosolid application (Badewa et al., 2023). Besides morphological root traits, plant-associated factors include root-derived chemical changes in the rhizosphere and the plant’s capacity for element uptake, transport and sequestration. The literature indicates large differences in the ability of plants to alter rhizosphere chemistry, to utilize a specific element pool (Lambers et al., 2015; Lu et al., 2020), and to cope with non-essential elements in the soil (Dillon and Courtney, 2023). With regard to metalloid accumulation and tolerance, the functional adaptations of plants include two basic physiological strategies: accumulators and excluders (Noor et al., 2022). The majority of plant species tolerate metals in the substrate by excluding the elements at the sites of uptake through restricting influx, promoting efflux, or extracellular complexation with organic ligands (Akram et al., 2024). In contrast, accumulators efficiently acquire/utilize elements from the soil and avoid element toxicity in the roots by rapidly transporting the elements to the shoots, where they are sequestrated in the leaf tissue. Specialized (hyper-) accumulators that evolved in metalliferous environments, of which Thlaspi caerulenscens and Alyssum murale are profoundly studied, accumulate more than 1,000 mg of Cd, Ni, and Zn in their shoot dry matter (van der Ent et al., 2013) but only when they are growing on soils with high metal mobility. These species rely on highly efficient transport and cellular hypertolerance mechanisms rather than alteration of rhizosphere chemistry (Pollard, 2023). In accumulator species from non-metalliferous soils, however, metal accumulation may appear as a side effect of functional traits related to P, Fe, and Mn acquisition (Monei et al., 2022), especially the release of carboxylates and acidification of the rhizosphere. Indeed, rhizosphere acidification has been linked with metal accumulation in different taxa from the Phytolaccaceae, Polygonaceae, Brassicaceae and Proteaceae (Kikis et al., 2024). Of these species, Fagopyrum esculentum unspecifically accumulates metals even when it is growing on soils with relatively low metal solubility, which has been attributed to rhizosphere acidification (Kreuzeder et al., 2018). In Lupinus albus and P-efficient species from the Proteaceae, Mn hyperaccumulation coincides with enhanced carboxylate release under conditions of P deficiency (Lambers et al., 2015; Lambers, 2022; Olt et al., 2022). Concomitantly, these species typically show low concentrations of non-essential elements (Martínez-Alcalá et al., 2013), likely as a consequence of element exclusion through extracellular complexation. Recently, relationships between carboxylate release and metal exclusion have been demonstrated for Al, Cd, Pb, and REEs (Wiche and Heilmeier, 2016; Wiche et al., 2023; Wiche and Pourret, 2023). Given that carboxylate release and rhizosphere acidification are regulated by the plant’s nutrient status (Lambers, 2022; Wiche et al., 2023), it is reasonable that soil amendment with digestate or sewage sludge impacts the uptake of non-essential elements not only through a direct increase in potentially available element pools in the soil but also indirectly through the alteration of plant nutrition.
To date, studies on Ga uptake in plants are scarce, and no research has addressed how sewage sludge or digestate amendments influence the availability and plant uptake of Ge, Ga, and REEs specially when the nutritional status of the plants is altered through the addition of amendments. Therefore, the objectives of this study were to: i) determine the impact of digestate and sewage sludge amendment on ammonium acetate-extractable (labile) essential (P, Fe, Mn, Ni) and non-essential elements (Cd, Ge, Ga, REEs) in soil, ii) evaluate the uptake of Cd, Ge, Ga, and REEs in four plants with contrasting element acquisition strategies using the P-efficient Mn accumulator L. albus (Pearse et al., 2006), the phosphophile unspecific metal accumulator F. esculentum, the element excluder Carthamus tinctorius Ciaramella et al., 2022), and the specialized Ni hyperaccumulator A. murale (Wiche and Pourret, 2023) and iii) investigate how changes in essential and non-essential element pools in the soil following biosolid amendment influence the soil-plant transfer of Cd, Ge, Ga, and REEs in these species.
2 Materials and methods
2.1 Greenhouse experiment on the effect of biosolids
Alyssum murale (yellow tuft), Lupinus albus (white lupin), Carthamus tinctorius (safflower) and Fagopyrum esculentum (buckwheat) were cultivated on soil (unamended), and soil mixed with sewage sludge or digestate from anaerobic fermentation in the greenhouse. Each treatment within each plant species was fivefold replicated. The soil was collected from the campus of TU Bergakademie Freiberg and characterized as luvisol. The vegetation was stripped off, and 200 kg of topsoil (0–20 cm depth) was collected, homogenized and sieved (2 mm). The sewage sludge was obtained from the sewage treatment plant in Muldental, Germany. The catchment area of the sewage plant includes industrial areas. Hence, the concentrations of toxic elements frequently exceed the thresholds for sewage sludge and limits the use of the material for thermal treatment. The digestate was obtained from the biogas lab of the Institute of Thermodynamics, TU Freiberg and derived from mesophilic anaerobic fermentation batch experiments with cow dung and grass biomass. The digestate and sewage sludge were thoroughly mixed. Of each material, the soil and the biosolids, ten homogeneous samples were collected and stored in centrifuge tubes at 4°C before being analyzed. In total, sixty pots (volume 2 L) were filled with either 3 kg of a homogenous mixture of 1800 g soil and 1,200 g sewage sludge (40:60% sewage sludge: soil, fw), 2,550 g soil and 450 g digestate (15:85% digestate: soil fw), or unamended soil as a reference. The water contents of soil, sewage sludge and digestate were 16%, 77% and 94%, respectively. Thus, the amount of digestate and sewage sludge added corresponded to 3% and 24% on a dry matter basis. Here, a higher portion of sewage sludge was added due to the substantially higher labile P concentrations in digestate compared to the sewage sludge (Supplementary Material S1). The substrates were allowed to settle for 4 weeks. Shortly before the transplantation of seedlings, soil samples (5 g of soil, sampling depth 10 cm) were collected from five randomly selected pots within each treatment. The samples were stored in centrifuge tubes at 4°C before being analyzed.
Seeds of A. murale (origin: Ankara, Kizilcahamam) were provided by the Botanical Garden and Botanical Museum Berlin, and seeds of C. tinctorius (cv CT05 Calin) were provided by the Exsemine Company. Seeds of F. esculentum (cv Bamby) and L. albus (cv Feodora) were obtained from Bornträger GmbH. Seeds of L. albus, F. esculentum, A. murale and C. tinctorius were surface-sterilized (H2O2) and germinated on a wet filter paper in a Petri dish. One individual of the three-day-old seedlings was placed in the middle of the pots to obtain each species growing on each substrate in fivefold replication. The pots were incubated in a growth chamber with 65% relative humidity, 25°C average temperature and 600 μmol/m2 s Photosynthetically active radiation (PAR) in a fully randomized design. The plants did not receive additional fertilizer; all pots were watered weekly with 200 mL of tap water over 6 weeks. After 6 weeks of plant growth, all plants were cut 1 cm above the soil surface. The shoot biomass was washed with deionized water, dried at 60°C, ground to a fine powder using an ultra-centrifugal mill (type ZM 1000, Retsch, Germany), and stored in centrifuge tubes at 4°C until being analyzed.
2.2 Collection of root exudates
A separate greenhouse experiment was designed for the determination of root exudates in cultivars of L. albus (cv. Feodora), F. esculentum (cv. Bamby), and C. tinctorius (cv. CT05 Calin) depending on P status. Seeds were surface-sterilized by washing with 0.5% sodium hypochlorite (NaOCl) for 3 min, followed by carefully rinsing with deionized water, and then allowed to germinate in Petri dishes in a growth chamber at 20°C. After germination, the seedlings of each plant species (one seedling per pot) were planted in 10 plastic pots (2 L total volume) filled with acid (HNO3) washed quartz sand. The pots were incubated for 5 weeks with a 15 h photoperiod, 18°C–30°C, relative humidity of 65%, and an average photosynthetically active photon flux density of 600 μmol/m2 s. During 5 weeks, all plants received weekly 200 mL of a 1/5 strength Hoaglands solution (Arnon and Stout, 1939) but with differing P concentrations. Specifically, half of the plants received a solution containing 100 μM KHPO4 together with the other nutrients (P100), while the other plants received 20 μM P (P20). After a cultivation period of 4 weeks, the mature plants were carefully removed from the sand by washing with tap water and transferred into glass beakers containing 100 mL of a 2.5 μM CaCl2 solution, where they were let to stay for 30 min under the growth lamp and allowed to release carboxylates into the collection solutions (Neumann et al., 2009). Immediately after the collection of root exudates, the pH was measured, and 1 mL L-1 Micropur was added to prevent microbial decomposition of carboxylates (Oburger et al., 2013). Thereafter, the shoots and roots were separated, weighed, and dried for 24 h at 60°C.
2.3 Chemical characterization of biosolids and soils
Each sample of the unamended soil, digestate, sewage sludge, and soil amended with the biosolids was homogenized and split into two subsamples. One-half was dried at 105°C to determine total element concentrations and water content. The other half was left fresh for the determination of labile-bound elements, mineral nitrogen (Nmin), soil pH, and DL-extractable phosphate. The dried samples were powdered in a boron carbide mortar. The ground samples (100 mg) were fully digested in a laboratory microwave (Ethos Plus MLS) with a mixture of HNO3 and HF, according to Krachler et al. (2002). In all sample processing steps, certified reference soil samples GBW 07406 and GBW 07407 were used for quality control. The resulting solutions were stored at 4°C before being analyzed. For the determination of mobile/exchangeable and acid-soluble elements, the samples were extracted with 1 M NH4+-acetate, pH 5, for 24 h, according to Wiche et al. (2017). The resulting solutions were centrifuged, filtered (200 nm), and stored at 4°C before being analyzed. For analysis of mineral N (NO3−, and NH4+), the substrate samples were extracted with deionized water and 1 mol/L KCl (1:10 extracts) and photometrically analyzed according to Bolleter et al., 1961 and Hartley and Asai (1963).
2.4 Determination of element concentrations and carboxylates
Ground plant samples (100 mg) were digested in a laboratory microwave (Ethos plus) with 1.9 mL of nitric acid (65% supra) and 0.6 mL of hydrofluoric acid (4.9% supra), according to Krachler et al. (2002). In all sample processing steps, certified reference plant samples NCS ZC73032 and NCS ZC73030 were used for quality control. The concentrations of P, Fe, Mn, Zn, Ni, Cd, Ga, Ge and REE in soil extracts and digestion solutions were determined by Inductively Coupled Plasma Mass Spectrometry (ICP-MS, xseries 2, Thermo Scientific and NexION 300D, Perkin Elmer), using 10 μg/L rhodium and rhenium as internal standards (Krachler et al. 2002; Monei et al., 2022). Calibration solutions ranging from 0.01 to 100 μg/L were prepared through suitable dilution of a multi-element stock standard solution (Merck). Accuracy verification was conducted by analyzing the certified reference materials GBW 07406, GBW 07407, NCS ZC73032, and NCS ZC73030 (LGC Standards); the results from soil analysis deviated by less than 16%, and the results from plant analysis by less than 12% from the certified values. Concentrations of acetate, malonate, fumarate, glutarate, malate, and citrate in the collection solutions were determined by ion chromatography equipped with suppressed conductivity detection (ICS-5000, Thermo Scientific). Organic anions were separated at 30°C on an IonPac® AS11-HC column (Thermo Scientific) using gradient elution with sodium hydroxide as eluent and a flow rate of 1.0 mL/min.
2.5 Data processing and statistics
Concentrations of light rare earth elements (LREEs) and heavy rare earth elements (HREEs) in the plant and soil samples were calculated as sums of La, Ce, Pr, Nd, Pm, Sm, Eu (LREEs) and Gd, Tb, Y, Ho, Er, Yb, Tm, Lu (HREEs) according to Tyler (2004). Based on LREE and HREE concentration the LREE/HREE-ratio was calculated to explore changes in REE fractionation depending on pant species and soil treatment (Wiche et al., 2023; Wiche and Pourret, 2023). All element concentrations reported were calculated on a dry-weight basis. Variance homogeneity, a model requirement of ANOVA, was checked using the Brown-Forsythe test. In the case of variance inhomogeneity, the data were transformed using log transformation to achieve variance homogeneity. Differences between element concentrations and contents in plants and soil were tested using a one-factor analysis of variance (ANOVA) followed by a Bonferroni post-hoc test (α = 5%). All statistical analyses were performed using SAS OnDemand for Academics.
3 Results
3.1 Physicochemical properties of the sewage sludge, digestate, and biosolids-amended soil
Sewage sludge and digestate had similar organic matter contents, whereas digestate had a higher pH and higher Nmin concentrations (Table 1). The sewage sludge contained 137% higher total P concentrations than digestate, but most of P (99%) was present in relatively insoluble forms and could not be dissolved by NH4+-acetate (pH 5) (Supplementary Material S1). In comparison, 42% of P in the digestate was present in the mobile, exchangeable, and acid-soluble forms. In addition, the digestate contained higher total and labile concentrations of Mn, Zn, and Ni as well as of total Fe (Supplementary Material S1).
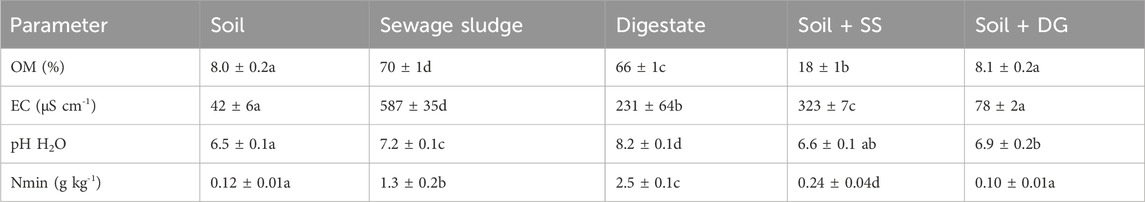
Table 1. Physicochemical parameters of soil (unamended), sewage sludge, digestate, and mixtures of soil and sewage sludge (soil + SS) or soil and digestate (soil + DG), respectively. Mean ± standard deviation (n = 4–5). Organic matter content (OM), electrical conductivity (EC), pH in aqueous solution (pH H2O), and mineral nitrogen (Nmin). Differences between means were identified by a one-way analysis of variance followed by a Bonferroni post-hoc test. Means with different letters are significantly different at (α = 5%).
Concerning the potentially toxic elements, there were no differences in the total concentrations of Cd, Ge, LREE and HREE in sewage sludge and digestate (Supplementary Material S1). However, the concentration of Ga was roughly 5-fold higher in digestate than in sewage sludge. Concomitantly, the sewage sludge contained significantly higher concentrations of labile Cd, Ge, LREE, and HREE and was characterized by a higher LREE/HREE ratio (Supplementary Material S1). When added to the soil, sewage sludge increased soil organic matter contents, EC, and Nmin significantly, whereas there was no effect of the addition of digestate (Table 1). We emphasize that the digestate had a higher water content than the sewage sludge (77% sewage sludge and 90% digestate) and was added at lower application rates, which impairs a direct comparison between the two treatments.
Nevertheless, compared to the unamended soil, the application of digestate increased soil pH, but this was not the case when sewage sludge was added (Table 1). The addition of both biosolids, the digestate and sewage sludge, respectively, did not alter the total concentrations of Cd and did not change the LREE/HREE ratio of the soil (Table 3). Moreover, the addition of digestate did not alter the total concentrations of the plant nutrients P, Fe, Mn and Zn (Table 2) or of the non-essential elements Ge, LREE, and HREE (Table 3). However, compared to the unamended soil, Ni concentrations were 53%, and Ga concentrations were 17% significantly higher when digestate was added (Table 2). In comparison, in the soil amended with sewage sludge, the total concentrations of P, Zn, and Ni were significantly higher than in unamended soil. At the same time, total Fe and Mn, Ge, Ga, LREE, and HREE concentrations were lower, whereas Cd concentrations, as well as LREE/HREE ratios, remained unchanged (Tables 2, 3).
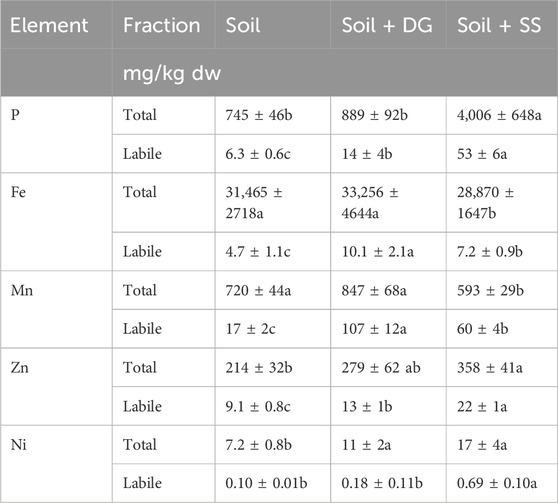
Table 2. Total and NH4-acetate extractable concentrations (mg/kg dw) of essential elements in soil (unamended), soil amended with digestate (soil + DG) and soil amended with sewage sludge (soil + SS). Mean ± standard error (n = 4–5). Concentrations within the same element fraction between different substrates were compared by a one-way analysis of variance followed by a Bonferroni post-hoc test. Means with different letters are significantly different at (α = 5%).
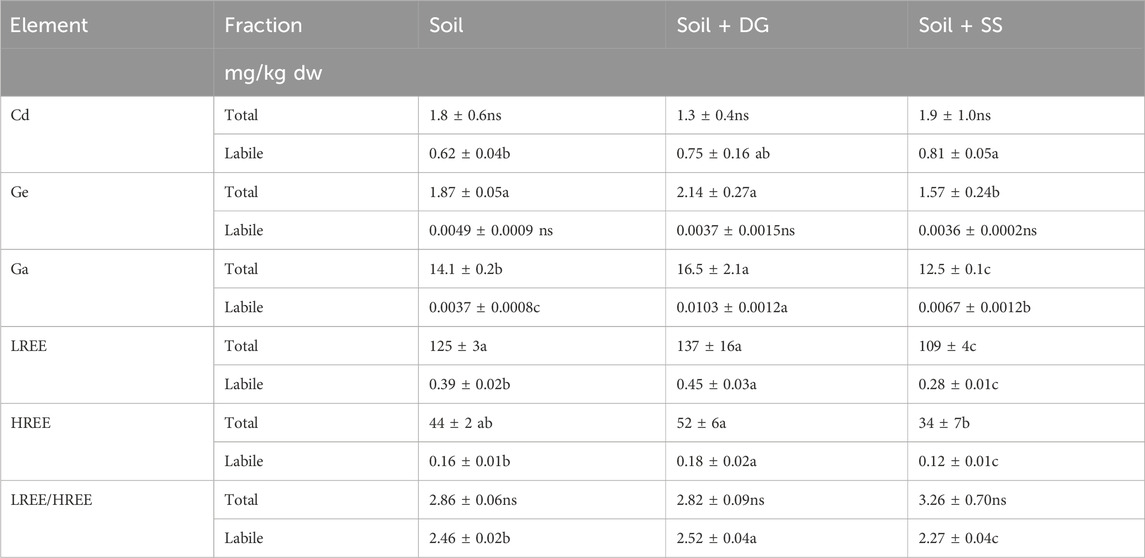
Table 3. Total and NH4-acetate extractable concentrations (mg/kg dw) of essential elements in soil (unamended), soil amended with digestate (soil + DG), and soil amended with sewage sludge (soil + SS). Mean ± standard error (n = 4–5). Concentrations within the same element fraction between different substrates were compared by a one-way analysis of variance followed by a Bonferroni post-hoc test. Means with different letters are significantly different at (α = 5%).
Although total concentrations of P, Fe, Mn, and Zn remained unchanged, the addition of digestate led to a higher mobility of these elements in the soil (by 122, 115, 529, and 44%). The increase in total Ni did not correspond with a higher Ni mobility. Similarly, the addition of digestate did not increase the mobility of Cd and Ge. However, the digestate significantly increased the mobility of Ga, LREE, and HREE, and it led to a higher LREE/HREE ratio in the NH4+-acetate-extractable element fraction. The sewage sludge increased the mobility of all considered plant nutrients, above all P, which showed a 741% higher mobility when sewage sludge was added. Moreover, the addition of sewage sludge increased the concentrations of NH4+-acetate-extractable Cd and Ga while the concentrations of mobile LREE, HREE, and the LREE/HREE-ratio decreased, resulting in LREE/HREE-ratios that were the lowest compared to all treatments (Table 3).
3.2 Carboxylate release in response to P-supply
Compared to plants growing under conditions of low P supply (20 µM P), all plants with high P supply (100 µM P) responded with increased shoot P concentrations (Table 4). Shoot P increased by 99%, 171%, and 195% in L. albus, F. esculentum, and C. tinctorius, respectively, indicating a more strongly pronounced effect in F. esculentum and C. tinctorius than in L. albus. All plants responded to increased shoot P supply with higher shoot biomass, except F. esculentum, which was characterized by marginal differences in shoot mass between the P treatments. The root mass of L. albus was higher when P supply was low, whereas the root mass of C. tinctorius declined. Neither L. albus nor C. tinctorius altered the pH of the exudate collection solutions. Irrespective of P supply, the collection solutions of F. esculentum showed the lowest pH values of all plant species tested and had significantly lower pH values under conditions of high P supply (Table 4). Low P supply increased the release of malonate and citrate in F. esculentum but did not alter the release of other carboxylates. Thus, there were no significant differences in the sum of carboxylates in this species. Similarly, in C. tinctorius, total carboxylate release remained unchanged in response to P supply. Here, only single components were affected in divergent ways, showing a lower release of malate, but a higher release of citrate when P supply was high. In contrast, L. albus responded to a high P supply with decreased malate release, whereas release of other compounds remained unchanged, leading to a net reduction of total carboxylate release by 60% in high P-supplied plants compared to P-deficient plants (Table 4).
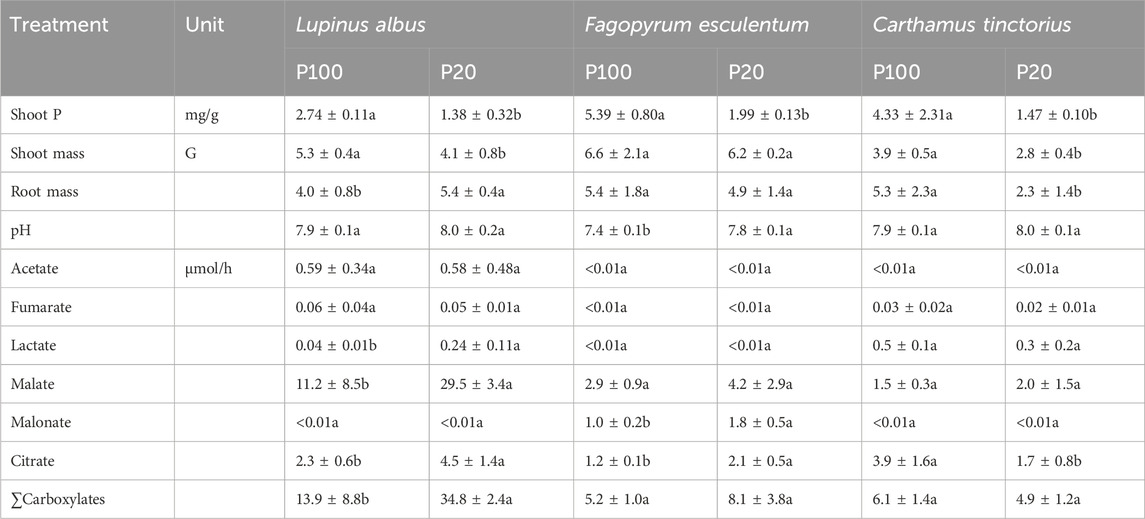
Table 4. Shoot P concentrations shoot mass, root mass, root carboxylate release (µmol/h), and pH of the collection solutions after 30 min of exudation time of plants cultivated in quartz sand treated with nutrient solutions containing 100 μM P or 20 μM P. Differences between P treatments within a specific plant species were compared by t-tests with Bonferroni adjustment. Means with different letters are significantly different at (α = 5%).
3.3 Plant growth and shoot nutrient concentrations in plants treated with biosolids
Plants of F. esculentum developed the highest biomass, and those of A. murale the lowest (Table 5). The addition of digestate tended to decrease plant growth of all species except C. tinctorius; however, this effect was not statistically significant at α = 5%. In contrast, sewage sludge-treated plants of F. esculentum, C. tinctorius, and L. albus developed 88%, 158% and 82% higher biomass, respectively. The addition of digestate did not alter the shoot P concentrations of the tested plants. There were apparent effects on the concentrations of the trace nutrients Fe and Mn, which varied among plant species, whereas Zn and Ni remained unchanged. In L. albus, trace nutrient concentrations were unaffected by the addition of digestate. Notably, L. albus showed the highest shoot Mn concentrations (more than 1,000 mg/kg Mn) in both digestate-treated and unamended soil, (Table 5). Shoot Fe and Mn of F. esculentum tended to increase; however, this was not statistically significant due to high data variability. In the other species, the addition of digestate led to higher shoot Fe and Mn concentrations (except Fe in C. tinctorius, which were lower), indicating an improved metal nutrient supply (Table 5).
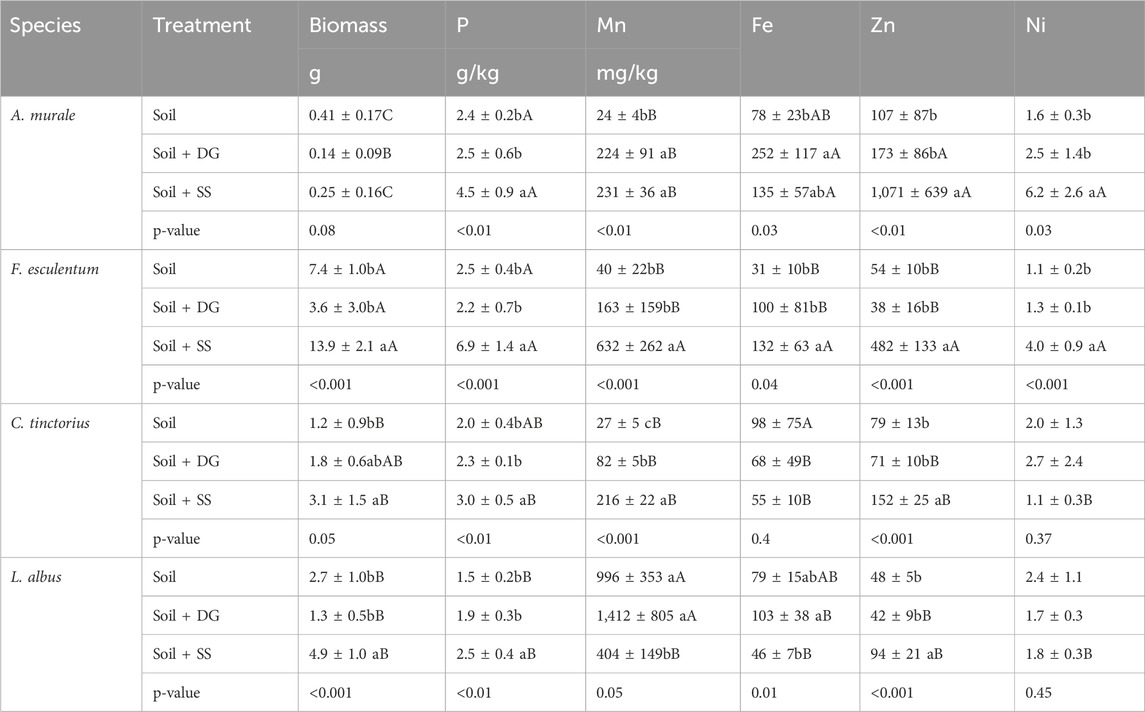
Table 5. Shoot biomass and concentrations of nutrients in plants cultivated on soil (unamended), soil amended with digestate (Soil + DG), and soil amended with sewage sludge (Soil + SS); mean ± sd, n = 4–5. Small letters show concentration differences among different substrates for the same plant species. Capital letters denote accumulation differences between plant species within same substrate. Significant differences were identified by one-factor analysis of variance (ANOVA) followed by a Bonferroni post-hoc test. Means with different letters are statistically significantly different at (ɑ = 5%).
Compared to the plants grown in unamended soil, the addition of sewage sludge strongly increased the concentrations of shoot P in all species. Shoot Mn concentrations were increased in A. murale, F. esculentum and C. tinctorius by 863%, 1,480%, and 700%, but not in L. albus, which exhibited a lower Mn concentration than plants grown in unamended soil (Table 5). In addition, the sewage sludge led to a higher Zn concentration in all species. In particular, sewage sludge-treated A. murale showed the highest shoot Zn concentrations of all plant species tested (more than 1,000 mg/kg Zn). Moreover, A. murale and F. esculentum responded to the sewage sludge treatment with 287% and 263% higher Ni concentrations compared to the plants grown in unamended soil. In contrast, shoot Ni remained unchanged in C. tinctorius and L. albus. Iron concentrations remained relatively unaffected in sewage sludge-treated plants, except in F. esculentum and A. murale, where Fe increased by 325% and 73%, respectively (Table 5).
3.4 Shoot accumulation of non-essential elements
Considering all treatments, A. murale showed the highest concentrations of the studied non-essential elements and L. albus the lowest (Table 6). The addition of digestate did not significantly alter Cd concentrations in the investigated species (Table 6). Germanium concentrations were generally very low and rarely exceeded 10 μg/kg. In F. esculentum, Ge was not detectable, irrespective of the treatment. Moreover, the digestate did not alter the concentrations of Ga, LREE and HREE in F. esculentum and C. tinctorius. However, in A. murale, Ge, Ga, LREE, and HREE concentrations were higher when digestate was added (Table 6). Lupinus albus did not show altered concentrations of Ge, Ga and HREE but exclusively showed a higher concentration of LREE.
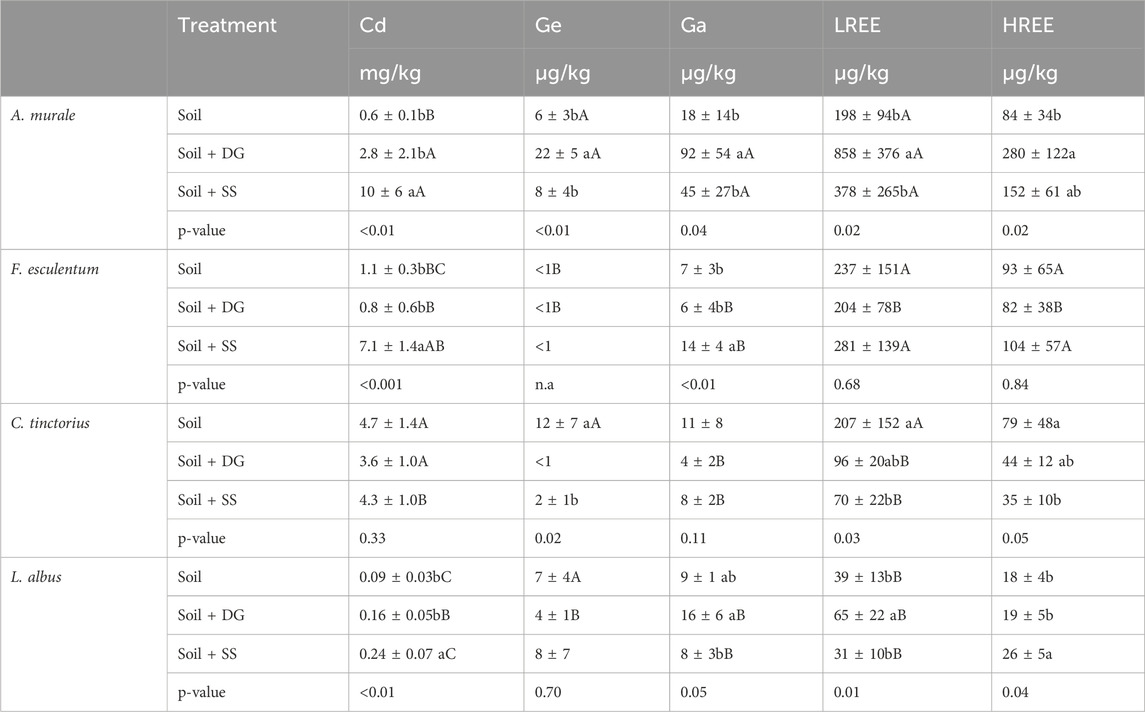
Table 6. Concentrations of non-essential elements in plants cultivated on soil (unamended) soil amended with digestate (Soil + DG) and soil amended with sewage sludge (Soil + SS); mean ± sd, n = 4–5). Significant differences within the substrate were identified by one-factor analysis of variance (ANOVA) followed by a Bonferroni post hoc test. Small letters show concentration differences among different substrates for the a same plant species. Capital letters denote accumulation differences between plant species within a same substrate. Means with different letters are statistically significantly different at (ɑ = 5%).
When sewage sludge was added, all plants except C. tinctorius responded with substantially higher Cd concentrations. Further, C. tinctorius showed declining concentrations of all other investigated elements, while the concentrations remained unchanged in A. murale. Fagopyrum esculentum showed a higher concentration of Ga but did not show any changes in LREE and HREE concentrations. Finally, L. albus did not show changes in Ge, Ga and LREE concentrations; however, this plant species showed higher HREE concentrations when sewage sludge was added, indicating significant changes in LREE/HREE ratios (Figure 1).
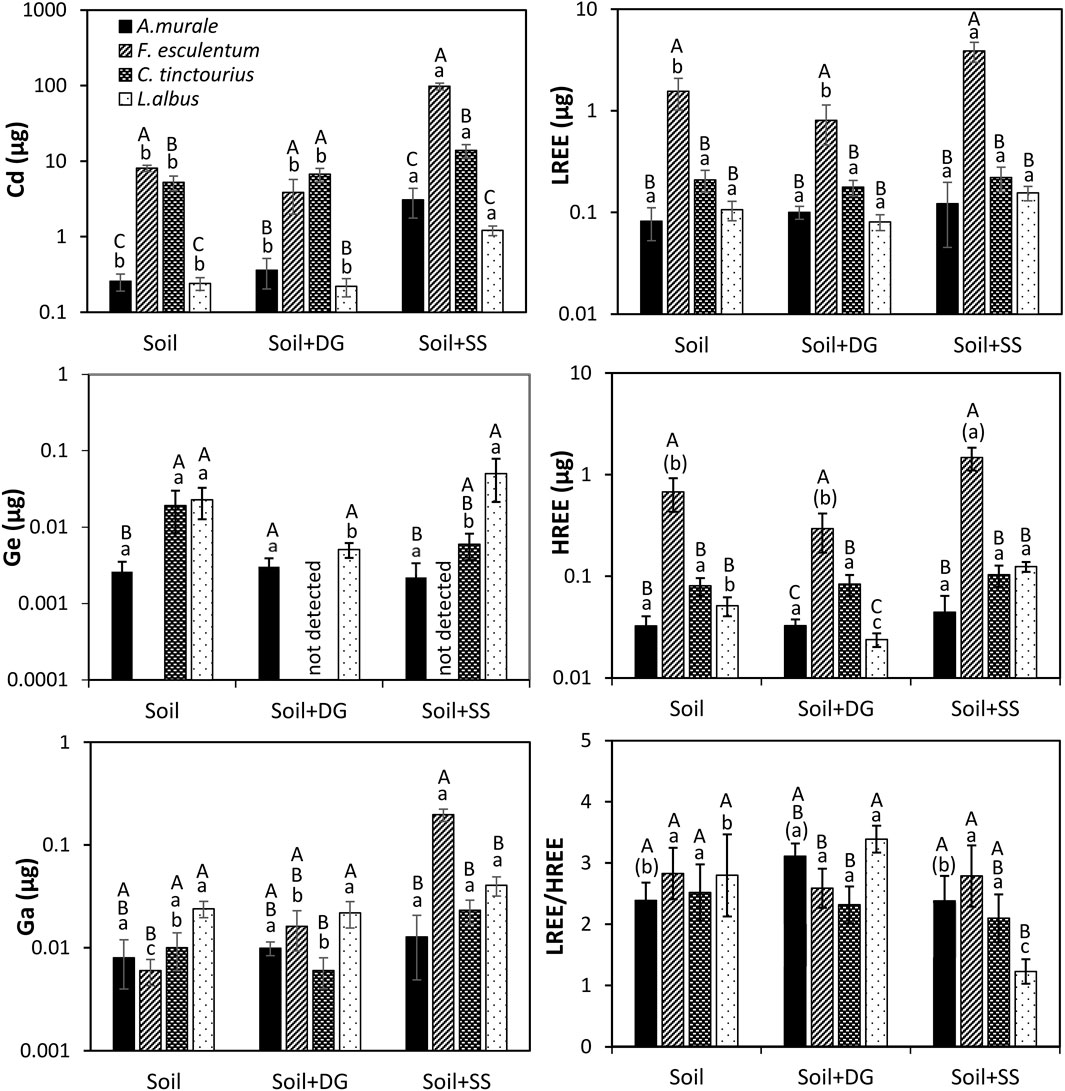
Figure 1. Effect of substrate properties on the accumulation of essential elements in the target plant species. Mean ± se (n = 4–5). Soil (S) unamended, soil + digestate (Soil + DG) and soil + sewage sludge (Soil + SS). Differences in shoot element contents were identified by one-way analysis of variance (ANOVA) followed by a Bonferroni test. Small letters show accumulation differences among different treatments within a specific plant species. Capital letters denote the accumulation differences of elements within a specific substrate among different plant species. Means with small letters (without bracket) different letters are statistically significantly different at (ɑ = 5%). Small letters in brackets denote a statistical significance at (ɑ = 10%).
Considering the shoot biomass and the herein quantified element concentrations, amounts of elements in the respective plant tissues and whole shoot contents were calculated (Figure 1). Plants grown in unamended soil of F. esculentum and C. tinctorius showed by far the highest net shoot uptake of Cd. Cadmium accumulation decreased in the order F. esculentum > C. tinctorius > L. albus, A. murale. A similar trend was observed when REE accumulation was compared among the species (Figure 1). Germanium was not detectable in F. esculentum, and there were no differences in Ga accumulation between A. murale, C. tinctorius and L. albus. Instead, the lowest Ga uptake was observed in F. esculentum.
The addition of digestate did not alter the accumulation of Cd in the investigated plant species, and A. murale did not show any changes in net shoot uptake of all investigated elements, including Cd. Similarly, F. esculentum did not show changes in LREE, HREE, and Ge uptake; however, a higher Ga uptake was observed (Figure 1). Conversely, in C. tinctorius, Ge and Ga accumulation declined following the addition of digestate. Lower Ge contents were also visible in L. albus, but Ga contents remained unchanged. In addition, the shoots of L. albus contained lower HREE contents, leading to significantly higher LREE/HREE ratios (Figure 1).
The addition of sewage sludge substantially increased the Cd uptake in all plants, regardless of the tested species (Figure 1). Germanium uptake was only affected in C. tinctorius, showing a lower content when sewage sludge was added, and Ga uptake was only affected in F. esculentum following the addition of sewage sludge. With regard to REEs, sewage sludge did not alter LREE and HREE accumulation in A. murale and C. tinctorius. However, F. esculentum responded to the addition of sewage sludge with a higher accumulation of both LREE and HREE, but there were no changes in the LREE/HREE ratios. In contrast, in L. albus, HREE concentrations were higher compared to the plants grown in unamended soil, leading to a substantially lower LREE/HREE ratio.
4 Discussion
4.1 Differences in shoot element uptake among the tested plant species
The soil used in our study was characterized by moderate mobility of phosphorus and micronutrients (Table 2), elevated concentrations of Cd (Kabata-Pendias, 2004) and average earth crust levels of Ga, Ge, and REE (Table 3) (Kabata-Pendias, 2004; Wiche et al., 2017; Wiche et al., 2018). Of these elements, 33% of total Cd was present in mobile and exchangeable forms that can be readily absorbed by plant roots (Bali et al., 2020). In contrast, the proportion of labile Ge, Ga, and REE did not exceed 0.5% of the total concentrations. Based on results from NH4+-acetate extracts, the potential availability decreased in the order Cd > LREE > HREE > Ge, Ga (Table 3). This is in accordance with the findings of Tyler and Olsson (2001), who demonstrated that compared to Cd and REE, the solubility of Ge and Ga in soils is low, and their mobilization requires substantial changes in physicochemical soil properties. Consequently, shoot element contents decreased in the order Cd > REE > Ga, Ge (Table 6; Figure 1). However, there were significant differences between plant species in handling the mobile elements, depending on the plant’s ability to take up and translocate the elements. The low accumulation of Ge (Figure 1) with equal (barely detectable) concentrations across all species (Table 6) can be attributed to its low solubility in the soil (Table 6) and/or inefficient uptake and root-shoot transport. The dicots tested do not have silicon transporters that are involved in Ge uptake (Nikolić et al., 2023; Kaiser et al., 2020). After Ge had been passively absorbed, the element was likely diluted by the biomass accumulation that decreased in the order F. esculentum > L. albus > C. tinctorius >> A. murale (Table 5). In contrast to Ge, the uptake of Cd, REE, and Ga is mediated by Zn, Fe, and Ca transporters (Shi et al., 2022), so differences in element uptake depend more strongly on processes related to plant nutrition. The shoot nutrient concentrations of plants grown in unamended soil (Table 5) indicated that all plants contained adequate P concentrations, except L. albus, which exhibited shoot P concentrations below the critical level of 2 g/kg (Lambers et al., 2013). Surely, the shoot P status might not sufficiently reflect P availability due to an altered P mobility within the plants (Zohar et al., 2024). Nevertheless, high Mn concentrations of L. albus (Table 5) clearly indicated that the plants released carboxylates under low P conditions (Lambers, 2022). Indeed, the lupin cultivar used in our study exhibited a substantially higher carboxylate release when P supply declined (Table 4).
Carboxylates and protons released by plant roots not only mobilize P and micronutrients, but also increase the solubility of an array of non-essential elements (Andresen et al., 2018) through dissolution, complexation, and ligand exchange (Kang and Peña, 2023; Wiche and Pourret, 2023). However, the mobile elements are not necessarily available for uptake when the elements are present as organo-metal complexes (Lee et al., 2021). Uptake systems predominantly shuttle ions through membranes, as demonstrated for Al, Cd, and REE (Wiche et al., 2023). Consequently, rhizosphere acidification, which predominantly mobilizes elements through dissolution and cation exchange, is likely an important plant trait related to accumulating non-essential elements in non-metalliferous environments, unlike carboxylate release. Overall, this explains the relatively low contents of non-essential elements in L. albus and the high contents in F. esculentum (Table 6; Figure 1). Possibly, the lupins efficiently mobilized the elements in the root zone, but chelation by carboxylates excluded them from uptake, whereas F. esculentum acidified the rhizosphere and translocated the absorbed elements to the shoots. High shoot Ga contents in L. albus could be a side effect of Mn hyperaccumulation and the upregulation of transition metal transporters under high Mn availability inflicted by P deficiency (Olt et al., 2022), given that Ga shares chemical similarities with Fe (Yandem and Jabłońska-Czapla, 2024). Concomitantly, Ga mobilization in the soil requires substantial changes in pH and/or the presence of high concentrations of carboxylates (Tyler and Olsson, 2001). We emphasize that information on differences in Ga accumulation in plant species is very scarce in the literature, and the elucidation of processes is fundamental in the face of soil pollution with this emerging pollutant (Shtangeeva, 2023). Carthamus tinctorius released minor amounts of carboxylates and did not acidify the rhizosphere (Table 4) as it has been previously described for P-inefficient phosphophilic species of the Brassicaceae (Lambers, 2022). Hence, this plant species accumulated only high amounts of Cd (Figure 1), which exhibited the highest mobility in the substrate (Table 3). Unfortunately, we did not analyze the carboxylate release of A. murale. Nonetheless, the literature indicates that A. murale is adapted to environments with high metal solubility (Bani et al., 2010) and relies on effective metal transport and internal detoxification rather than rhizosphere processes related to element mobilization and/or exclusion (Wiche and Pourret, 2023). Therefore, it is not surprising that the plants grown in unamended soil in our study exhibited relatively low Zn, Mn, and Ni concentrations (Table 5). The concentrations observed were two orders of magnitude lower than in plants from metalliferous environments (Bani et al., 2015; Van der Ent et al., 2021). Possibly, the plants suffered Ni deficiency, which in turn may have affected biomass development (Bani et al., 2015). In this plant species, high shoot concentrations of Ga, Ge, and REE might be related to processes of metal tolerance, but the resulting net element uptake was low due to the low biomass of this species.
4.2 Effect of digestate on plant nutrition and element accumulation in plants
All plants except C. tinctorius responded to the addition of digestate by accumulating less biomass. However, all plants showed higher micronutrient concentrations (Table 5), likely resulting from a higher proportion of labile nutrients in the digestate-amended soil (Table 2). A higher portion of labile P (Table 2) did not improve shoot P supply (Table 5), indicating metabolic changes or changes in root activity. Fedeli et al. (2023) investigated the effect of different forms of digestate on plant growth and demonstrated that solid digestate may have adverse effects, while liquid digestate improved plant growth. In our study, we observed growth inhibition following the addition of liquid digestate, except for C. tinctorius, where plant growth remained unchanged. Possibly, the digestate detrimentally altered the soil microbiome (Karimi et al., 2022), led to soil compaction (Caracciolo et al., 2022) or altered other physicochemical soil properties (Przygocka-Cyna and Grzebisz, 2018). Future studies should consider microbial communities changes depending on digestate composition. In this regard, the unaffected growth of C. tinctorius is particularly interesting (Table 4). Carthamus tinctorius develops extensive root systems (Montiel et al., 2020) and is described as a suitable species for crop production on marginal soils (Rosero et al., 2020), highlighting the role of species-specific traits in plant responses to soil additives like digestate. Given that concentrations do not necessarily reflect element uptake due to enrichment or dilution of elements in varying biomass, plant availability was evaluated by net shoot uptake (Figure 1). Changes in aboveground biomass are typically accompanied by changes in root biomass and altered metabolic activity as a stress response that, in turn, can affect element absorption (Shtangeeva, 2023). The present work concentrates on shoot content. In a future study, root content will be considered to clarify the total net uptake of plant biomass.
Nevertheless, we clearly observed that the digestate did neither affect labile Cd in the soil (Table 3) nor Cd uptake in the tested plants (Figure 1). A lower Ge uptake (Figure 1) but unchanged element mobility could derive from a lower Ge diffusion in the soil when the pH and soil organic matter (OM) raised following digestate addition (Table 1). Moreover, slightly higher concentrations of labile REEs in the substrate did not alter LREE and HREE uptake in A. murale, C. tinctorius and F. esculentum. The LREE/HREE ratios in A. murale reflected the higher LREE/HREE ratio of the digestate-treated soil (Table 3; Figure 1), suggesting that A. murale utilizes this element pool for uptake and translocates the elements to the shoots without discrimination. Although not statistically significant at α = 5%, the LREE/HREE ratio of L. albus exceeded the LREE/HREE ratio of A. murale, suggesting a discrimination of HREE relative to LREE through extracellular complexation with carboxylates (Wiche and Pourret, 2023) (Figure 1). In fact, L. albus showed a significantly lower HREE uptake, which cannot be explained by altered soil properties (Figure 1). Still, the digestate-treated lupins were P-deficient (Table 5) and released large amounts of carboxylates, as indicated by the high shoot Mn concentrations (Table 5). Non-significant results might be from low sample size so the validation remains field for future studies. Similarly, a higher Ga mobility in the digestate treatment increased Ga uptake only in F. esculentum, while A. murale, L. albus, and C. tinctorius did not respond to a higher portion of labile Ga in the substrate. Overall, this suggests that Ga and REE uptake and accumulation are predominantly controlled by plant physiological traits related to nutrition acquisition (Zohar et al., 2024) rather than by element mobility in the soil.
4.3 Effect of sewage sludge on plant nutrition and element accumulation in the plants
The sewage sludge-treated soil was characterized by a substantially higher mobility of all measured essential nutrients, as well as Cd and Ga (Tables 2, 3). Ge mobility was not affected by sewage sludge, and Ge uptake in plants tended to decrease (Figure 1), suggesting that the impact of biosolids on the soil–plant transfer of Ge is low. All plants except A. murale responded to the sewage sludge treatment with substantially higher biomass and all plants exhibited a luxury supply of P and micronutrients (Table 5). High Mn and Zn concentrations in A. murale, F. esculentum, and C. tinctorius likely derive from a higher portion of labile elements in the substrate in concert with higher root activity. All plants responded to the higher Cd mobility in the substrate with a higher Cd uptake (Figure 1). Still, the plants exhibited the same pattern of shoot Cd contents as the plants grown in unamended soil, supporting the application of NH4+-acetate-extracts for estimating overall Cd mobility in soil (Wang et al., 2024). However, species-specific physiological traits clearly govern the degree to which a plant can utilize this mobile element pool in the soil. Similarly, to the treatment with digestate, a higher Ga mobility in the substrate only affected shoot accumulation in F. esculentum. Moreover, the addition of sewage sludge resulted in unchanged LREE and HREE accumulation in A. murale, F. esculentum, and C. tinctorius. Possibly, the nutrients contained in the sewage sludge improved root growth, leading to a larger soil volume accessed by the roots. At the same time, L. albus exhibited a significantly lower LREE/HREE ratio that clearly resulted from a higher shoot uptake of HREE relative to LREE, which cannot be solely explained by altered soil chemistry (Table 3; Figure 1). Instead, it seems that the changes in nutrient supply following sewage sludge amendment altered root activity and element acquisition processes. In L. albus, Mn concentrations declined, suggesting a lower carboxylate release under conditions of high P supply (Lambers et al., 2015; Tables 4, 5). Given that HREEs form more stable complexes with carboxylates than LREEs (Wiche and Pourret, 2023), reduced carboxylate concentrations in the rhizosphere and apoplast may enhance HREE uptake and thus decrease the LREE/HREE ratio (Wiche et al., 2023). This demonstrates that the assessment of the effects of biosolids on the soil-plant transfer of Ge, Ga, and REE should include not only altered element concentrations in the soil but also physiological responses of the plant to altered nutritional status.
5 Conclusion
We demonstrated that both digestate and sewage sludge influence not only the mobility of nutrients in the soil but also the occurrence of Ge, Ga, and REE in plant-available forms. The physico-chemical properties and the spectrum of elements in sewage sludge and digestate may vary depending on their origin and the treatment technology. Moreover a longer experimental time would be necessary to explore the fate of elements in soil plant system in detail. Nevertheless, changes in total concentrations and element mobility assessed by NH4+-acetate extraction do not sufficiently explain element availability to plants, especially when it comes to the assessment of soil-plant transfer of Ga and REE. Of the investigated elements, Ge mobility and its uptake by plants were the least affected by sewage sludge and digestate, suggesting that the risk of soil-plant transfer is relatively low. We emphasize that digestate amendment increases the total Ge in the soil, which may accumulate in the soil over time and pose a risk once the element is released from the organic matrix. In contrast, mobility and plant-availability of Cd, Ga, and REEs were clearly affected depending on soil amendment and plant species. A higher portion of NH4+-acetate-extractable Cd in the soil resulting from the sewage sludge treatment increased Cd accumulation in all species, indicating that the plants utilized this mobile element pool during uptake. Consequently, the assessment of mobile/exchangeable elements seems to be a good proxy of soil-plant transfer for this element. However, plant-availability of Ga and REE was not directly reflected by the elements’ mobility in the soil. Instead, it seems that the soil-plant transfer of Ga is governed by physiological traits involved in uptake, while the soil-plant transfer of REE additionally depends on the plant’s nutritional status and below-ground functional traits related to phosphorus acquisition. This highlights the necessity for the evaluation of soil-plant transfer of Ge, Ga, and REEs in long-term field experiments, in which changes in soil chemistry are monitored in addition to physiological responses of species and genotypes to altered nutrient supply.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
NZ: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Software, Visualization, Writing – original draft, Writing – review and editing. VL: Data curation, Formal Analysis, Methodology, Writing – original draft. AS: Writing – original draft. TA: Data curation, Formal Analysis, Writing – review and editing. OP: Data curation, Writing – review and editing. SF: Data curation, Writing – review and editing. PB: Data curation, Writing – review and editing. EV: Data curation, Writing – review and editing. MP: Data curation, Formal Analysis, Writing – review and editing. SW: Data curation, Formal Analysis, Writing – review and editing. EP: Writing – original draft. OW: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Software, Supervision, Validation, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. OW received a research scholarship granted by the OECD in the framework of the Co-operative Research Programme: Sustainable Agricultural and Food Systems. NZ was funded in the framework of the PhaNoMix project, financed by the Federal Ministry of Food and Agriculture (BMEL) on behalf of the FNR (grant number 2220NR074B). This work was partially supported by the European Union’s HORIZON EUROPE WIDERA 2021 program under the SURRI project, Grant Agreement No. 101079345.
Acknowledgments
Numerous students were involved in the experiments; the authors are indebted to them for their activity. We are grateful to Hermann Heilmeier, Roland Achtziger, Elke Richert and Michel-Pierre Faucon for discussions and support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fenvs.2025.1628175/full#supplementary-material
References
Akarsu, C., Sönmez, V. Z., and Sivri, N. (2023). An overview of the occurrence and distribution of plastics in wastewater treatment plants and the necessity of developing up-to-date management strategies. Camb. Prisms Plast. 1, e23. doi:10.1017/plc.2023.23
Akram, U., Shahzadi, I., Peerzada, S. M. S., Sajal, F., and Farooq, A. (2024). “Metalloids accumulation and translocation in plants,” in Metals and metalloids in plant signaling. Signaling and communication in plants. Editor T. Aftab (Cham: Springer). doi:10.1007/978-3-031-59024-5_2
Alengebawy, A., Abdelkhalek, S. T., Qureshi, S. R., and Wang, M. Q. (2021). Heavy metals and pesticides toxicity in agricultural soil and plants: ecological risks and human health implications. Toxics 9, 42–34. doi:10.3390/toxics9030042
Andresen, E., Peiter, E., and Küpper, H. (2018). Trace metal metabolism in plants. J. Exp. Bot. 69, 909–954. doi:10.1093/jxb/erx465
Arnon, D., and Stout, P. (1939). The essentiality of certain elements in. Plant Physiol. 14, 1460–1470. doi:10.1104/pp.14.2.371
Badewa, E. A., Yeung, C. C., Whalen, J. K., and Oelbermann, M. (2023). Compost and biosolids increase long-term soil organic carbon stocks. Can. J. Soil Sci. 103, 483–492. doi:10.1139/cjss-2022-0104
Bali, A. S., Sidhu, G. P. S., and Kumar, V. (2020). Root exudates ameliorate cadmium tolerance in plants: a review. Springer International Publishing.
Bani, A., Echevarria, G., Sulçe, S., and Morel, J. L. (2015). Improving the agronomy of Alyssum murale for extensive phytomining: a five-year field study. Int. J. Phytoremediation 17, 117–127. doi:10.1080/15226514.2013.862204
Bani, A., Pavlova, D., Echevarria, G., Mullaj, A., Reeves, R. D., Morel, J. L., et al. (2010). Nickel hyperaccumulation by the species of alyssum and thlaspi (brassicaceae) from the ultramafic soils of the balkans. Bot. Serbica 34, 3–14.
Bolleter, W. T., Bushman, C. J., and Tidwell, P. W. (1961). Spectrophotometric determination of ammonia as indophenol. Anal. Chem. 33 (4), 592–594. doi:10.1021/ac60172a034
Caracciolo, A. B., Visca, A., Rauseo, J., Spataro, F., Garbini, G. L., Grenni, P., et al. (2022). Bioaccumulation of antibiotics and resistance genes in lettuce following cattle manure and digestate fertilization and their effects on soil and phyllosphere microbial communities. Environ. Pollut. 315, 120413. doi:10.1016/j.envpol.2022.120413
Ciaramella, B. R., Corinzia, S. A., Cosentino, S. L., and Testa, G. (2022). Phytoremediation of heavy metal contaminated soils using safflower. Agronomy 12, 2302. doi:10.3390/agronomy12102302
Dillon, L., and Courtney, R. (2023). Availability and transfer of non-essential elements in a rehabilitated bauxite residue grassland system. Plant Soil 497, 105–118. doi:10.1007/s11104-023-06213-1
Fedeli, R., Celletti, S., Loppi, S., and Vannini, A. (2023). Comparison of the effect of solid and liquid digestate on the growth of lettuce (Lactuca sativa L.) plants. Agronomy 13 (3), 782. doi:10.3390/agronomy13030782
Hartley, A. M., and Asai, R. I. (1963). Spectrophotometric determination of nitrate with 2, 6-Xylenol reagent. Anal. Chem. 35 (9), 1207–1213. doi:10.1021/ac60202a001
Jama-Rodzeńska, A., Sowiński, J., Koziel, J. A., and Białowiec, A. (2021). Phosphorus recovery from sewage sludge ash based on cradle-to-cradle approach—Mini-review. Minerals 11, 985. doi:10.3390/min11090985
Kabata-Pendias, A. (2004). Soil-plant transfer of trace elements - an environmental issue. Geoderma 122, 143–149. doi:10.1016/j.geoderma.2004.01.004
Kaiser, S., Wagner, S., Moschner, C., Funke, C., and Wiche, O. (2020). Accumulation of germanium (Ge) in plant tissues of grasses is not solely driven by its incorporation in phytoliths. Biogeochemistry 148, 49–68. doi:10.1007/s10533-020-00646-x
Kang, K., and Peña, J. (2023). Siderophore-mediated mobilization of manganese limits iron solubility in mixed mineral systems. ACS Earth Sp. Chem. 7, 662–675. doi:10.1021/acsearthspacechem.2c00271
Kanteraki, A. E., Isari, E. A., Svarnas, P., and Kalavrouziotis, I. K. (2022). Biosolids: the trojan horse or the beautiful helen for soil fertilization? Sci. Total Environ. 839, 156270. doi:10.1016/j.scitotenv.2022.156270
Karimi, B., Sadet-Bourgeteau, S., Cannavacciuolo, M., Chauvin, C., Flamin, C., Haumont, A., et al. (2022). Impact of biogas digestates on soil microbiota in agriculture: a review. Environ. Chem. Lett. 20 (5), 3265–3288. doi:10.1007/s10311-022-01451-8
Kikis, C., Thalassinos, G., and Antoniadis, V. (2024). Soil phytomining: recent developments—A review. Soil Syst. 8, 8. doi:10.3390/soilsystems8010008
Krachler, M., Mohl, C., Emons, H., and Shotyk, W. (2002). Influence of digestion procedures on the determination of rare earth elements in peat and plant samples by USN-ICP-MS. J. Anal. At. Spectrom. 17 (8), 844–851. doi:10.1039/b200780k
Kowalik, R., Gawdzik, J., Bąk-Patyna, P., Ramiączek, P., and Jurišević, N. (2022). Risk analysis of heavy metals migration from sewage sludge of wastewater treatment plants. Int. J. Environ. Res. Public Health 19 (18), 11829. doi:10.3390/ijerph191811829
Kreuzeder, A., Santner, J., Scharsching, V., Oburger, E., Hoefer, C., Hann, S., et al. (2018). In situ observation of localized, sub-mm scale changes of phosphorus biogeochemistry in the rhizosphere. Plant Soil 424, 573–589. doi:10.1007/s11104-017-3542-0
Lambers, H. (2022). Phosphorus acquisition and utilization in plants. Annu. Rev. Plant Biol. 73, 17–42. doi:10.1146/annurev-arplant-102720-125738
Lambers, H., Clements, J. C., and Nelson, M. N. (2013). How a phosphorus-acquisition strategy based on carboxylate exudation powers the success and agronomic potential of lupines (lupinus, fabaceae). Am. J. Bot. 100 (2), 263–288. doi:10.3732/ajb.1200474
Lambers, H., Hayes, P. E., Laliberté, E., Oliveira, R. S., and Turner, B. L. (2015). Leaf manganese accumulation and phosphorus-acquisition efficiency. Trends Plant Sci. 20, 83–90. doi:10.1016/j.tplants.2014.10.007
Lee, M., Seo, B., and Kim, K. R. (2021). Effects of free metal ions and organo-metal complexes on the absorption of lead and cadmium by plants. J. Appl. Biol. Chem. 64 (2), 159–164. doi:10.3839/jabc.2021.023
Lu, H. l., Nkoh, J. N., Abdulaha-Al, B. M., Dong, G., Li, J. y., and Xu, R. k. (2020). Plants alter surface charge and functional groups of their roots to adapt to acidic soil conditions. Environ. Pollut. 267, 115590. doi:10.1016/j.envpol.2020.115590
Marchuk, S., Tait, S., Sinha, P., Harris, P., Antille, D. L., and McCabe, B. K. (2023). Biosolids-derived fertilisers: a review of challenges and opportunities. Sci. Total Environ. 875, 162555. doi:10.1016/j.scitotenv.2023.162555
Marotrao, A. L., Singh, S. K., Patra, A., Kumar, O., Jatav, S. S., and Yadav, S. N. (2021). Assessing heavy metal accumulation in plants and soil with sewage sludge application under rice–wheat system in an indo-gangetic inceptisol. Arabian J. Geosciences 14, 2391–11. doi:10.1007/s12517-021-08659-y
Martínez-Alcalá, I., Hernández, L. E., Esteban, E., Walker, D. J., and Pilar Bernal, M. (2013). Responses of Noccaea caerulescens and Lupinus albus in trace elements-contaminated soils. Plant Physiol. Biochem. 66, 47–55. doi:10.1016/j.plaphy.2013.01.017
Monei, N., Hitch, M., Heim, J., Pourret, O., Heilmeier, H., and Wiche, O. (2022). Effect of substrate properties and phosphorus supply on facilitating the uptake of rare earth elements (REE) in mixed culture cropping systems of Hordeum vulgare, Lupinus albus and Lupinus angustifolius. Environ. Sci. Pollut. Res. 29, 57172–57189. doi:10.1007/s11356-022-19775-x
Montiel, C. B., Secco, D., Marins, A. C., Junior, L. A. Z., Deggerone, J. R., and Bassegio, D. (2020). Root and shoot growth in safflower as affected by soil compaction. Aust. J. Crop Sci. 14, 1443–1448. doi:10.21475/ajcs.20.14.09.p2466
Neumann, G., George, T. S., and Plassard, C. (2009). Strategies and methods for studying the Rhizosphere—the plant science toolbox. Plant Soil 321, 431–456. doi:10.1007/s11104-009-9953-9
Nikolić, D., Bosnić, D., and Samardžić, J. (2023). Silicon in action: between iron scarcity and excess copper. Front. Plant Sci. 14, 1039053. doi:10.3389/fpls.2023.1039053
Noor, I., Sohail, H., Sun, J., Nawaz, M. A., Li, G., Hasanuzzaman, M., et al. (2022). Heavy metal and metalloid toxicity in horticultural plants: tolerance mechanism and remediation strategies. Chemosphere 303, 135196. doi:10.1016/j.chemosphere.2022.135196
Oburger, E., Dell‘mour, M., Hann, S., Wieshammer, G., Puschenreiter, M., and Wenzel, W. W. (2013). Evaluation of a novel tool for sampling root exudates from soil-grown plants compared to conventional techniques. Environ. Exp. Bot. 87, 235–247. doi:10.1016/j.envexpbot.2012.11.007
Okoroafor, P. U., Ogunkunle, C. O., Heilmeier, H., and Wiche, O. (2022). Phytoaccumulation potential of nine plant species for selected nutrients, (REEs), germanium (Ge), and potentially toxic elements (PTEs) in soil. Int. J. Phytoremediation 0, 1–11. doi:10.1080/15226514.2021.2025207
Olt, P., Alejandro-Martinez, S., Fermum, J., Ramos, E., Peiter, E., and Ludewig, U. (2022). The vacuolar transporter LaMTP8.1 detoxifies manganese in leaves of Lupinus albus. Physiol. Plant 13807, e13807. doi:10.1111/ppl.13807
Pearse, S. J., Veneklaas, E. J., Cawthray, G., Bolland, M. D. A., and Lambers, H. (2006). Triticum aestivum shows a greater biomass response to a supply of aluminium phosphate than Lupinus albus, despite releasing fewer carboxylates into the rhizosphere. New Phytol. 169, 515–524. doi:10.1111/j.1469-8137.2005.01614.x
Pecorini, I., Peruzzi, E., Albini, E., Doni, S., Macci, C., Masciandaro, G., et al. (2020). Evaluation of MSW compost and digestate mixtures for a circular economy application. Sustain 12, 3042. doi:10.3390/su12073042
Pollard, A. J. (2023). Inadvertent uptake of trace elements and its role in the physiology and evolution of hyperaccumulators. Plant Soil 483, 711–719. doi:10.1007/s11104-022-05856-w
Przygocka-Cyna, K., and Grzebisz, W. (2018). Biogas digestate - benefits and risks for soil fertility and crop quality - an evaluation of grain maize response. Open Chem. 16, 258–271. doi:10.1515/chem-2018-0027
Qvarforth, A., Svensson, P. A., Lundgren, M., Rodushkin, I., Engström, E., Paulukat, C., et al. (2025). Geochemical insights into plant uptake of technology-critical elements: a case study on lettuce from European soils. Chemosphere 371, 144073. doi:10.1016/j.chemosphere.2025.144073
Rékási, M., Mazsu, N., Draskovits, E., Bernhardt, B., Szabó, A., Rivier, P. A., et al. (2019). Comparing the agrochemical properties of compost and vermicomposts produced from municipal sewage sludge digestate. Bioresour. Technol. 291, 121861. doi:10.1016/j.biortech.2019.121861
Rosero, A., Granda, L., Berdugo-Cely, J. A., Šamajová, O., Šamaj, J., and Cerkal, R. (2020). A dual strategy of breeding for drought tolerance and introducing drought-tolerant, underutilized crops into production systems to enhance their resilience to water deficiency. Plants 9, 1263. doi:10.3390/plants9101263
Sheoran, V., Sheoran, A. S., and Poonia, P. (2016). Factors affecting phytoextraction: a review. Pedosphere 26 (2), 148–166. doi:10.1016/S1002-0160(15)60032-7
Shi, Y., Guo, S., Zhao, X., Xu, M., Xu, J., Xing, G., et al. (2022). Comparative physiological and transcriptomics analysis revealed crucial mechanisms of silicon-mediated tolerance to iron deficiency in tomato. Front. Plant Sci. 13, 1094451–14. doi:10.3389/fpls.2022.1094451
Shtangeeva, I. (2023). Temporal variability of gallium in natural plants. Toxics 11 (8), 675. doi:10.3390/toxics11080675
Sinha, P., Marchuk, S., Harris, P., Antille, D. L., and McCabe, B. K. (2023). Potential for land application of biosolids-derived biochar in Australia. A Rev. 1–29. doi:10.3390/su151410909
Turcotte, P., Smyth, S. A., Gagné, F., and Gagnon, C. (2022). Lanthanides release and partitioning in municipal wastewater effluents. Toxics 10 (5), 254. doi:10.3390/toxics10050254
Tyler, G. (2004). Rare earth elements in soil and plant systems-a review. Plant and soil, 267, 191–206. doi:10.1007/s11104-005-4888-2
Tyler, G., and Olsson, T. (2001). Plant uptake of major and minor mineral elements as influenced by soil acidity and liming. Plant Soil 230, 307–321. doi:10.1023/A:1010314400976
Van der Ent, A., Baker, A. J., Reeves, R. D., Pollard, A. J., and Schat, H. (2013). Hyperaccumulators of metal and metalloid trace elements: facts and fiction. Plant soil 362, 319–334. doi:10.1007/s11104-012-1287-3
van der Ent, A., Joseph Pollard, A., Echevarria, G., Abubakari, F., Erskine, P. D., Baker, A. J. M., et al. (2021). Exceptional uptake and accumulation of chemical elements in plants: extending the hyperaccumulation paradigm. 99–131. doi:10.1007/978-3-030-58904-2_6
Wang, J. F., Liu, C., Xu, Z. M., Wang, F. P., Sun, Y. Y., Huang, J. W., et al. (2024). Microbial mechanisms in nitrogen fertilization: modulating the re-mobilization of clay mineral-bound cadmium in agricultural soils. Sci. Total Environ. 926, 171809. doi:10.1016/j.scitotenv.2024.171809
Wiche, O., Dittrich, C., Pourret, O., Monei, N., Heim, J., and Lambers, H. (2023). Relationships between carboxylate-based nutrient-acquisition strategies, phosphorus-nutritional status and rare earth element accumulation in plants. Plant Soil 489, 645–666. doi:10.1007/s11104-023-06049-9
Wiche, O., and Heilmeier, H. (2016). Germanium (Ge) and rare earth element (REE) accumulation in selected energy crops cultivated on two different soils. Min. Eng. 92, 208–215. doi:10.1016/j.mineng.2016.03.023
Wiche, O., and Pourret, O. (2023). The role of root carboxylate release on rare earth element (hyper)accumulation in plants – a biogeochemical perspective on rhizosphere chemistry. Plant Soil 492, 79–90. doi:10.1007/s11104-023-06177-2
Wiche, O., Székely, B., Moschner, C., and Heilmeier, H. (2018). Germanium in the soil-plant system—a review. Environ. Sci. Pollut. Res. 25, 31938–31956. doi:10.1007/s11356-018-3172-y
Wiche, O., Zertani, V., Hentschel, W., Achtziger, R., and Midula, P. (2017). Germanium and rare earth elements in topsoil and soil-grown plants on different land use types in the mining area of freiberg (Germany). J. Geochem. Explor 175, 120–129. doi:10.1016/j.gexplo.2017.01.008
Yandem, G., and Jabłońska-Czapla, M. (2024). Review of indium, gallium, and germanium as emerging contaminants: occurrence, speciation and evaluation of the potential environmental impact. Archives Environ. Prot., 84–99. doi:10.24425/aep.2024.151688
Zaffar, N., Ferchau, E., Heilmeier, H., Boldt, C., Salcedo, L. D. P., Reitz, T., et al. (2023). Enrichment and chemical fractionation of plant nutrients, potentially toxic and economically valuable elements in digestate from mesophilic and thermophilic fermentation. Biomass Bioenergy 173, 106779. doi:10.1016/j.biombioe.2023.106779
Zohar, I., Ganem, H. E., DiSegni, D. M., and Jonas-Levi, A. (2024). The impact of alternative recycled and synthetic phosphorus sources on plant growth and responses, soil interactions and sustainable agriculture-lettuce (Lactuca sativa) as a case model. Sci. Total Environ. 948, 174719. doi:10.1016/j.scitotenv.2024.174719
Keywords: soil amendment, heavy metal, plant availability, metalloids, phytoextraction
Citation: Zaffar N, Lovynska V, Samarska A, Arnstadt T, Pourret O, Firmin S, Baroš P, Vachková EL, Palušák M, Wacławek S, Peiter E and Wiche O (2025) Effect of sewage sludge and digestate from anaerobic fermentation on the accumulation of cadmium (Cd), gallium (Ga), germanium (Ge), and rare earth elements (REEs) in soil and uptake by plants with different nutrition strategies. Front. Environ. Sci. 13:1628175. doi: 10.3389/fenvs.2025.1628175
Received: 13 May 2025; Accepted: 26 June 2025;
Published: 09 July 2025.
Edited by:
Chamindra L. Vithana, Southern Cross University, AustraliaReviewed by:
Iris Zohar, Tel-Hai College, IsraelChuck Chuan N. G., Xiamen University Malaysia (XMUM), Malaysia
Obed N. Lense, State University of Papua, Indonesia
Copyright © 2025 Zaffar, Lovynska, Samarska, Arnstadt, Pourret, Firmin, Baroš, Vachková, Palušák, Wacławek, Peiter and Wiche. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nazia Zaffar, bmF6aWEuemFmZmFyQGlvZXoudHUtZnJlaWJlcmcuZGU=