 Erhan Keyvan1*
Erhan Keyvan1* Elizabeth Adesemoye2Marie-Christine Champomier-Vergès3Emilie Chanséaume-Bussiere4
Elizabeth Adesemoye2Marie-Christine Champomier-Vergès3Emilie Chanséaume-Bussiere4 Julie Mardon5
Julie Mardon5 Daniela Nikolovska Nedelkoska6Recep Palamutoglu7
Daniela Nikolovska Nedelkoska6Recep Palamutoglu7 Pasquale Russo8Inga Sarand9Laurencia Songre-Ouattara10
Pasquale Russo8Inga Sarand9Laurencia Songre-Ouattara10 Biljana Trajkovska11
Biljana Trajkovska11 Sibel Karakaya12
Sibel Karakaya12 Michail Syrpas13
Michail Syrpas13 Christophe Chassard14
Christophe Chassard14 Smilja Pracer15
Smilja Pracer15 Guy Vergères16
Guy Vergères16 Daniel Heine17
Daniel Heine17 Christèle Humblot18,19*
Christèle Humblot18,19*- 1Department of Food Hygiene and Technology, Faculty of Veterinary Medicine, Burdur Mehmet Akif Ersoy University, Burdur, Türkiye
- 2Department of Microbiology, Faculty of Life Science, Federal University Oye Ekiti, Ekiti State, Nigeria
- 3Université Paris-Saclay, INRAE, AgroParisTech, Micalis Institute, Jouy-en-Josas, France
- 4Nutrifizz, Clermont-Ferrand, France
- 5Université Clermont Auvergne, INRAE, VetAgro Sup, UMR 0545 Fromage, Lempdes, France
- 6Faculty of Technology and Technical Sciences Veles, University St. Kliment Ohridski, Bitola, North Macedonia
- 7Department of Nutrition and Dietetics, Faculty of Health Sciences, Afyonkarahisar Health Sciences University, Afyon, Türkiye
- 8Department of Food, Environmental and Nutritional Sciences (DeFENS), University of Milan, Milan, Italy
- 9Department of Chemistry and Biotechnology, Tallinn University of Technology, Tallin, Estonia
- 10Centre National de la Recherche Scientifique et Technologique, Ouagadougou, Burkina Faso
- 11Faculty of Biotechnical Sciences, University St. Kliment Ohridski, Bitola, North Macedonia
- 12Department of Food Engineering, Faculty of Engineering, Ege University, Izmir, Türkiye
- 13Department of Food Science and Technology, Kaunas University of Technology, Kaunas, Lithuania
- 14Université Clermont Auvergne, INRAE, VetAgro Sup, UMR 0545 Fromage, Aurillac, France
- 15Institute for Biological Research" Siniša Stanković", National Institute of the Republic of Serbia, University of Belgrade, Belgrade, Serbia
- 16Agroscope, Bern, Switzerland
- 17School of Agricultural, Forest, and Food Sciences, Bern University of Applied Sciences, Bern, Switzerland
- 18Avignon Université, CIRAD, Institut Agro, IRD, Université de La Réunion, Montpellier, France
- 19French National Research Institute for Sustainable Development (IRD), Montpellier, France
Introduction: Vitamin deficiencies remain a global health issue, particularly among vulnerable populations. As microorganisms also produce vitamins, this has led to considering Fermented Foods (FF) as potential vehicles for improving vitamin intake. This systematic narrative review, which exclusively relies on human studies, aims to assess the extent to which the consumption of vitamin-rich FF contributes to the maintenance or enhancement of vitamin status in healthy or deficient populations.
Methods: A comprehensive literature search (1970–2024) was conducted following the protocols of EFSA and the COST Action PIMENTO to identify interventional and observational studies investigating the influence of FF on biomarkers of vitamin status.
Results: Findings confirm that certain microorganisms, including Bacillus subtilis, Propionibacterium freudenreichii, and some lactic acid bacteria, can increase the levels of vitamins K2, B2, B9, and B12 in FF. Evidence of bioavailability and physiological effects is reported. Notably, folate (vitamin B9) bioavailability was enhanced in some cases following the consumption of Camembert cheese naturally rich in folate, while vitamin K2 status was effectively improved in several studies on natto (fermented soy) and in one study on Jarlsberg cheese. However, evidence for other B vitamins (B1, B2, B3, B5, B6 and B12) is limited or inconsistent, and no human evidence exists for other vitamins. Vitamin bioavailability was found to be significantly influenced by the food matrix, fermentation type, microbial strain, and the form of the vitamin (vitamers). Effects may also be influenced by interactions with gut microbiota, including microbial vitamin synthesis and modulation of absorption.
Discussion: Despite encouraging data, there is a lack of well-controlled, large-scale human studies to validate FF as a sustainable strategy to improve vitamin status. Future human studies research should investigate strain-specific effects, food matrix interactions, and long-term health outcomes.
1 Introduction
Vitamin deficiencies have severe health consequences worldwide (1). Vitamins can act as cofactors, signaling molecules, or gene regulators, thus affecting metabolic pathways, enzymatic reactions, or cellular homeostasis. They are categorized into water-soluble and fat-soluble vitamins, and each has a specific role, which is presented in this review.
Vitamin requirements are higher for young children, pregnant and lactating women as well as women of reproductive age, adolescents, and older adults, who are therefore particularly at risk of deficiencies (2). The prevalence of individual vitamin deficiencies is poorly quantified, due to undiagnosed cases resulting from unclear symptoms and the non-inclusion of biomarkers for measuring vitamin status in population studies (1). Micronutrient malnutrition affects approximately from 2 billion up to 4–5 billion people worldwide (3, 4). Aside from specific populations with malabsorption related to disease or treatment, the primary cause of vitamin deficiencies is inadequate intake (5).
Common strategies to combat deficiencies include dietary diversification, fortification of staple foods, and supplementation (1). Although the effectiveness of these strategies is well established, the global prevalence of micronutrient deficiencies is still high and varies depending on contextual factors (1, 4). Dietary diversification involves consuming a variety of nutritionally rich foods, but this may be limited for some populations due to cost or dietary choices, such as excluding animal-based foods, which are the primary source of vitamin B12 (6). Fortification of staple foods and supplementation using capsules, tablets or drops relies on synthetic forms of vitamins, which may be unaffordable or inaccessible to vulnerable populations in resource-limited regions (7). In addition, the consumption of synthetic forms of vitamin B9 can mask symptoms of vitamin B12 deficiency and lead to irreversible neuropathy (2). Therefore, exploring additional ways to improve vitamin intake to complement existing strategies is essential.
Fermented foods (FF) are a particular category of foods, which are produced from various materials, through microbial action. FF are estimated to account for around 30% of the foods consumed worldwide (8). FF are highly diverse and are prepared from all types of substrates (milk, cereals, fruits, etc.), using different processes, different microorganisms (spontaneous fermentation, back-slopping, or inoculation with defined starter strains), and different fermentation types (lactic, acetic, etc.) resulting in a wide variety of food products consumed as staples, beverages or condiments (9). They may therefore significantly contribute to consumers’ vitamin intake.
Among microorganisms responsible for fermentation, some have been demonstrated to possess vitamin-forming capabilities and have been successfully utilized to enhance the vitamin content of raw materials (10). However, the vitamin production capacity drastically differs, and not all vitamins are present in FF; with their formation depending on the microbial strain and fermentation conditions. Among the fat-soluble vitamins, microbial synthesis of vitamin K2 (menaquinones) in FF is the most studied. Vitamin K2, which differs structurally and functionally from vitamin K1 (a plant-derived phylloquinone), plays a key role in blood coagulation and bone metabolism by activating vitamin K-dependent proteins. Various FF such as fermented soybeans (natto, cheonggukjang) and cheeses are valuable dietary sources of vitamin K2, with microorganisms such as Bacillus subtilis responsible for its formation (11).
Among water-soluble vitamins, the ability of lactic acid bacteria (LAB) to synthesize different B-group vitamins have been studied, with particular emphasis on folate (vitamin B9) and riboflavin (vitamin B2) and to a lesser extent on cobalamin (vitamin B12), which is primarily associated with Propionibacterium freudenreichii (12, 13). B-group vitamins are essential for energy metabolism, DNA synthesis, neurotransmitter function, and the regulation of homocysteine. They act as enzyme cofactors in metabolic pathways. They are found in various FF, including dairy products (14).
Other vitamins, such as the fat-soluble vitamins A, D and E, as well as the water soluble vitamin C (ascorbic acid), are rarely studied in the context of FF. Vitamin A is involved in vision, immune function, and cellular differentiation through gene regulation (15). Vitamin D, regulates calcium and phosphate metabolism, essential for bone health and immune function through nuclear receptor activation (16). Vitamin E functions as an antioxidant, protecting cell membranes from oxidative damage and influencing lipid metabolism (17). Similarly, vitamin C (ascorbic acid), which also functions as an antioxidant, enhances iron absorption, supports collagen synthesis, and plays a role in immune regulation (18).
Some microorganisms are capable of producing precursors to vitamins A, D, and E, or industrially relevant forms of vitamin C; however, these pathways are uncommon or absent in bacteria present in traditional FF (19, 20). Vitamin C and E are major representatives of industrially produced vitamins (21). The production of vitamin E is chemical and/or using engineered microorganisms and little is known about the localization of the tocopherol biosynthetic enzymes in bacteria. Photosynthetic bacteria such as Synechocystis spp. have contributed to the identification of several tocopherol pathway genes (22). Vitamin C can be produced by some bacteria, but not usually those involved in food fermentation, and often by genetically engineered strains. Therefore, its formation is generally not applicable in FF (23). In fermented vegetables such as sauerkraut, vitamin C is present in meaningful amounts, but it originates primarily from the raw cabbage and not from microbial synthesis (114). In some commercial products, acerola powder or other fruit extracts may be added to boost vitamin C content, but this should not be attributed to the fermentation process itself.
The incorporation of vitamin-producing microorganisms in food fermentation can enhance the nutritional quality and offers a sustainable, environmentally friendly alternative to chemical synthesis. Ongoing research on novel microbial sources and optimization strategies indicates that microbial vitamin production could play a crucial role in enhancing global nutrition security (24). Many articles explore the use of microorganisms to increase vitaminic content of various raw materials and food, with several reviews comprehensively summarizing their findings (11, 12, 25). However, the absorption and physiological effects of naturally occurring forms of vitamins (vitamers) differ depending on factors such as food matrix and fermentation conditions. Whether FF can help maintain adequate vitamin status in healthy individuals, or improve status in deficient populations, remains insufficiently studied.
Regarding hosts, microorganisms from the gut microbiota may also contribute to the host’s vitamin status through direct vitamin synthesis in the digestive tract or by improving vitamin bioavailability (26). FF can modulate gut microbiota composition and function, potentially enhancing its contribution to vitamin metabolism. In addition, FF can serve as a source of microorganisms with vitamin-synthesizing capabilities, which may continue to produce vitamins directly in the gut during gastrointestinal transit (26). The contribution of gut microbes to vitamin status is insufficiently studied and the effect of FF consumption on the contribution of the gut microbes is inexistent. Nevertheless, some hypothesis have been formulated by several authors, and will be presented in the manuscript.
The objective of this systematic narrative review was to assess whether the consumption of FF contributes to the vitamin status of both healthy and vitamin-deficient populations. This review is part of a series of articles of the COST Action CA20128—Promoting Innovation of ferMENTed fOods (PIMENTO) on the health benefits and risks of FF (27). A unique aspect of this work is that it includes only human studies, evaluating the effects of FF in which vitamins were formed through fermentation, based on changes in vitamin levels or related biomarkers in the body. In addition, all types of FF and all types of vitamins were considered, ranging from well-studied dairy products to fermented soybeans and cereal grains. The potential link between FF consumption, gut microbiota modulation, and vitamin status is briefly discussed, though not systematically addressed in the included studies due to scarcity of related literature.
2 Methods
2.1 Study design and protocol
This systematic narrative review was conducted in accordance with the indicative guidance of the European Food Safety Authority (EFSA) for evaluating mechanisms of action and follows a predefined protocol aligned with the PROSPERO format, the PIMENTO Study Protocol, and the 24-step methodological framework proposed by Muka et al. (27, 28). The protocol was registered in the Open Science Framework (OSF) using the Open-Ended Registration modus (29) and is available at https://osf.io/8psdz/. The timeline for the review extends from September 1, 2023, to December 31, 2024.
2.2 Literature search
A systematic search was conducted across PubMed, Scopus, and the Cochrane Library covering publications. In two phases: first from January 1, 1970, to August 31, 2023, in English, then, an extra search that extend August 2023 to December 2024. The search utilized a generic string developed by the University of Zurich Library for PIMENTO, covering the maximum diversity of FF groups, human studies, and dietary intake terms. Functional terms related to vitamin metabolism were added to the generic PIMENTO search string. In addition to the PIMENTO search strings, search terms specific to the review were created to encompass the functional aspects of the study, including clinical indications and biological activities associated with vitamin status. Studies involving both healthy and vitamin-deficient populations (including children) were included, as vitamin deficiency is particularly relevant in younger populations. The systematic review encompasses human intervention studies, including randomized controlled trials, non-randomized controlled studies, and various other intervention studies, as well as observational studies such as cohort studies, case–control studies, and cross-sectional studies, in accordance with the EFSA guidance (30). Systematic reviews, whether including meta-analysis or not, were screened to identify potentially relevant studies. Animal and in vitro studies were excluded from the systematic review process. The selection of studies and extraction of data adhered to the procedures specified by Muka et al. and the Cochrane Handbook (28, 31). The CADIMA software assisted the organization and screening of references, thereby ensuring a systematic and reproducible selection process (32). The search and study selection strategy were documented with a PRISMA flow diagram to illustrate the inclusion and exclusion process (Figure 1).
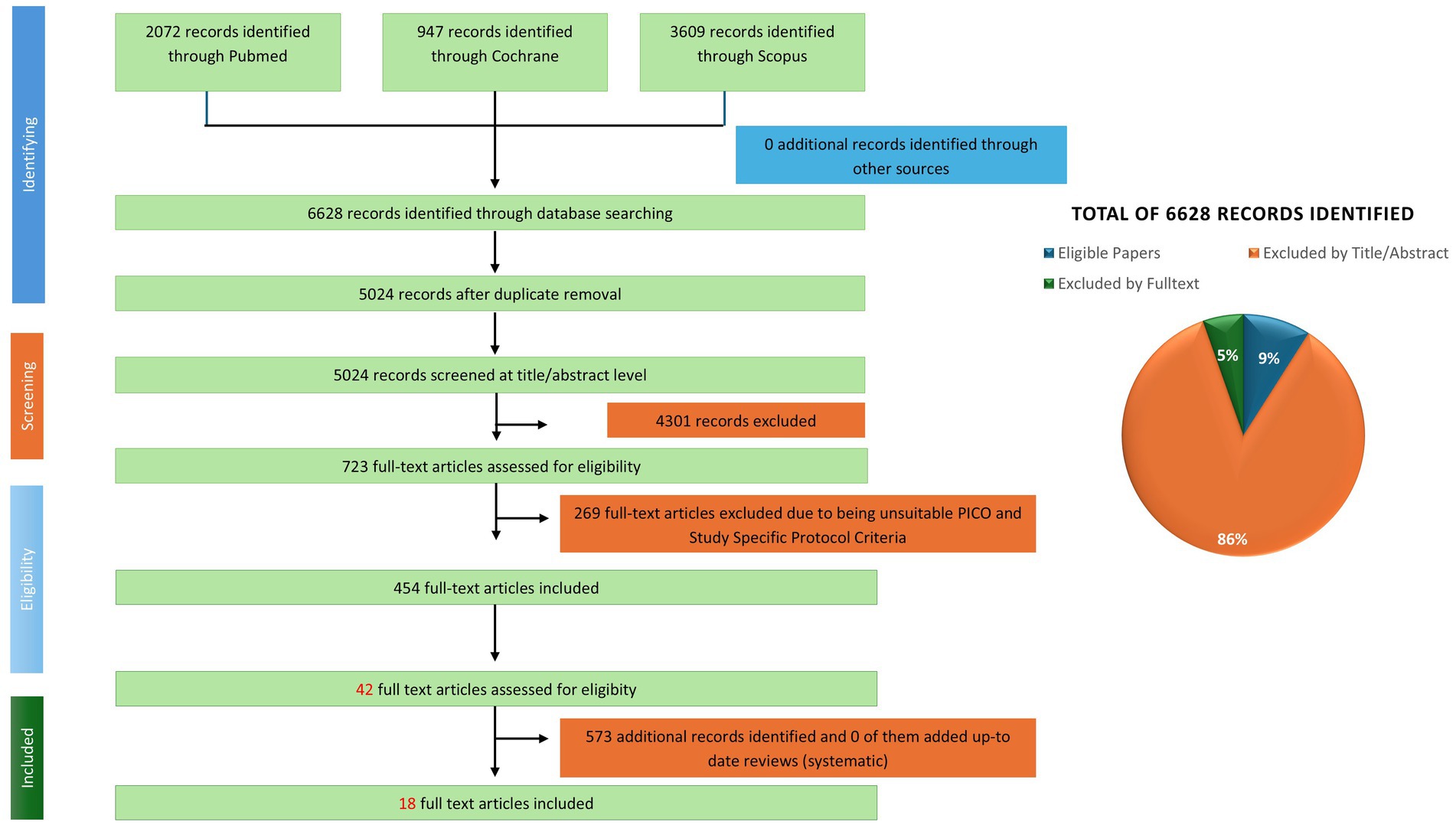
Figure 1. PRISMA flow diagram illustrating the study selection process and results of the systematic literature search.
2.3 Study selection criteria
The selection of studies was based on predefined inclusion and exclusion criteria, aligned with the PIMENTO Study Protocol. The study population, intervention/exposure, and comparator conditions were established using the PI(E)COS framework to ensure the relevance and quality of the studies included.
2.3.1 Inclusion and exclusion criteria
2.3.1.1 Population
The review encompassed individuals of all ages, both healthy and those with vitamin deficiencies. Studies involving non-healthy populations, with the exception of those specifically exhibiting vitamin deficiencies, were excluded. Individuals aged 18 and older were classified as adults; however, children were also included due to their increased susceptibility to vitamin deficiencies.
2.3.1.2 Intervention or exposure
This review analyses the intervention/exposure of FF consumption, as stated in the PIMENTO search string, encompassing the following food groups: dairy products, meat and fish, fruits and vegetables, beverages, legumes, cereals, and grains.
Exclusion criteria: FF fortified with synthetic forms of vitamins (e.g., fermented milk fortified with vitamin D). Alcoholic beverages containing more than 1.25% alcohol by volume. Research on probiotics as supplements suggests that their effectiveness is contingent upon their addition at the onset of the fermentation process, where they contribute to the fermentation process. Studies contain prebiotic fibers or additional bioactive compounds as potential confounding variables. Applications of FF beyond nutrition, such as topical or nasal uses. Interventions could be standalone or combined with other dietary components, provided that non-fermented control conditions were appropriately managed.
2.3.1.3 Comparator or control
The control groups included: Non-consumption or lower consumption of the FF in question. Consumption of the non-fermented version of the food(s). Any suitable non-fermented placebo or control.
A two-step approach for study selection was employed: Studies were first evaluated according to the PI(E)O criteria, which encompass Population, Intervention/Exposure, and Outcome. The comparator conditions were subsequently evaluated independently. This method facilitated a thorough assessment of the control groups in the chosen studies, while also highlighting deficiencies in the quality of control measures across the studies.
2.3.1.4 Outcomes
The primary outcome of this review is an increase in vitamin levels in the studied population after consuming FF. The evaluation of vitamin status was be conducted using specific biomarkers for each vitamin. For example, serum folate concentration was used as a biomarker for folate status.
Furthermore, safety outcomes were evaluated, encompassing any adverse effects or risk indicators linked to the intake of FF, as detailed in Appendix B of the EFSA guidance (30).
P = All healthy or vitamin deficient population, all ages
I = Any FF
C = Non-consumption or lower consumption of the FF in question. Consumption of the non-fermented version of the food(s). Any suitable non-fermented placebo or control
O = Increase in vitamin levels.
2.4 Study selection and data extraction
The data extraction process followed the methodology outlined in the PIMENTO Study Protocol and was conducted in accordance with the systematic review guidelines by Muka et al. (28). The following steps were implemented to ensure accuracy and consistency:
2.4.1 Study selection
Initial screening of titles and abstracts by at least two independent reviewers. Retrieval of full-text articles and application of inclusion/exclusion criteria. Resolution of discrepancies through discussion or consultation with a third reviewer.
2.4.2 Data extraction and organization
The specific data extraction form, a modified version of the PIMENTO WG3 data extraction form was developed using guidance from the Cochrane Handbook for Systematic Reviews of Interventions (31), the EFSA guidance on health claims (30) (Appendix B), and the STROBE statement for observational studies (33). These forms were used to collect structured data from each eligible study, including study design, population demographics, health and vitamin status, intervention characteristics (type, dosage, frequency), comparator details, primary and secondary outcomes (e.g., vitamin biomarkers), and indicators of bias and study quality.
2.4.3 Systematic selection and screening software
Screening and reference management were conducted using the CADIMA software, where a consistency test was applied to a subset of the dataset (32). In cases of inconsistency, the study protocol and selection strategy were refined, including the use of a third reviewer when necessary, to improve reproducibility and systematic accuracy. The PRISMA flow diagram was used to document the literature selection process transparently (Figure 1).
2.5 Non-systematic review components
In line with the PIMENTO S2 Study Protocol, this review includes a non-systematic component to complement the systematic findings. It summarizes the characteristics of the FF, including their microbial strains and food matrices. Additional supportive evidence from the literature addresses the bioavailability of vitamins in FF, their mechanisms of action such as microbial biosynthesis and gut microbiota modulation, and the safety aspects, including microbial safety and potential adverse effects.
3 Results and discussion
This review analyzed 18 studies on health responses in humans following the consumption of FF (Figure 1). Table 1 provides an overview of the vitamins present in FF, their effects, biomarkers and role of food and gut microbiota. Acting as cofactors, signaling molecules, or gene regulators, vitamins influence metabolic pathways, enzymatic reactions, and cellular homeostasis.
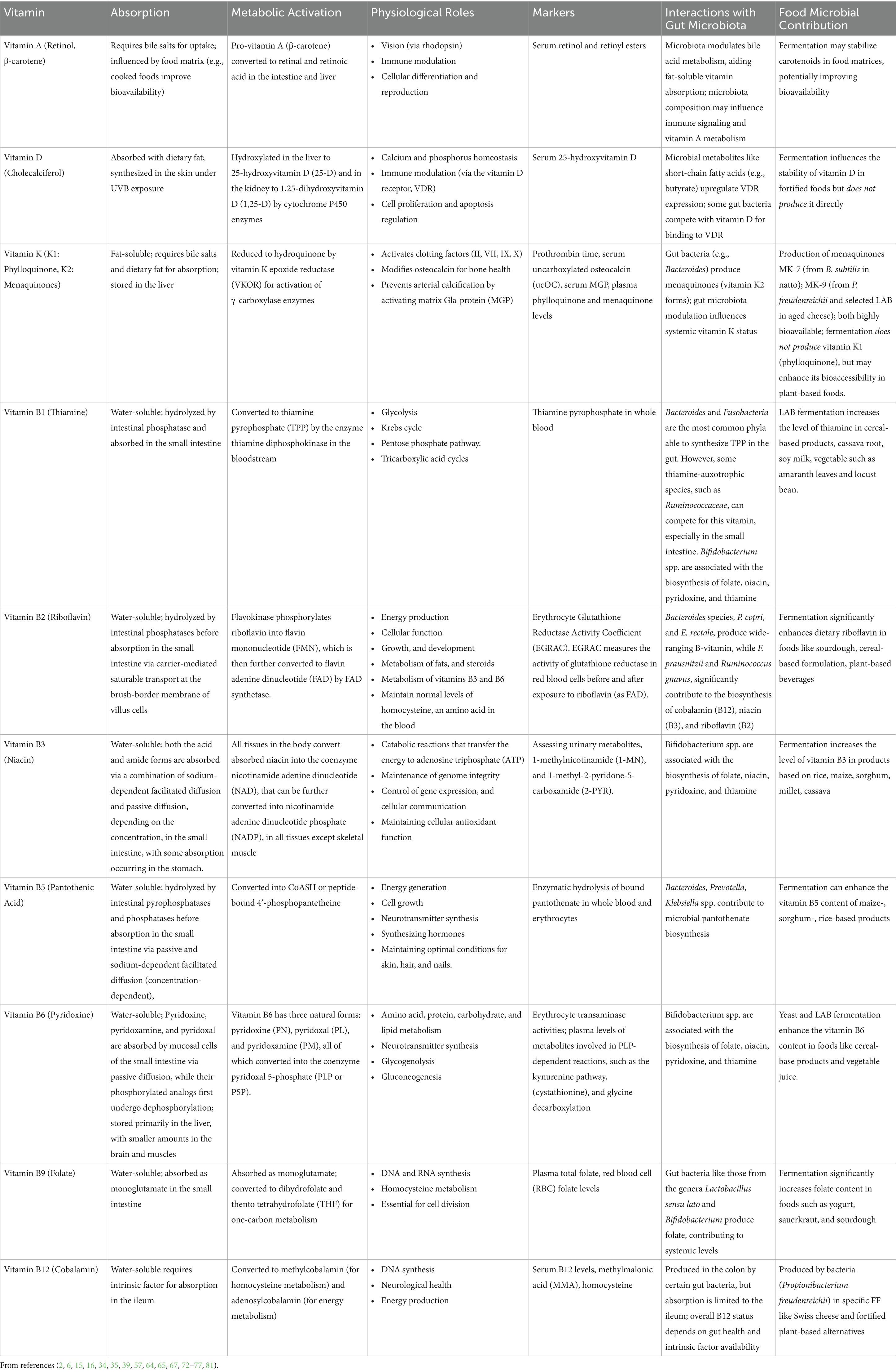
Table 1. Overview of water- and fat-soluble vitamins relevant in FF, (A, D, K, B1, B2, B3, B5, B6, B9, B12), including their absorption, metabolic activation, physiological roles, biomarkers, gut microbiota interactions, and microbial contributions.
The detailed presentation of the respective observational or interventional studies is presented in the relevant sections of this review, organized by individual vitamin (Table 2). To provide context for the observed effects, the following section introduces the food source of the vitamin, the underlying mechanisms of action of the vitamins being investigated and a presentation of human studies.
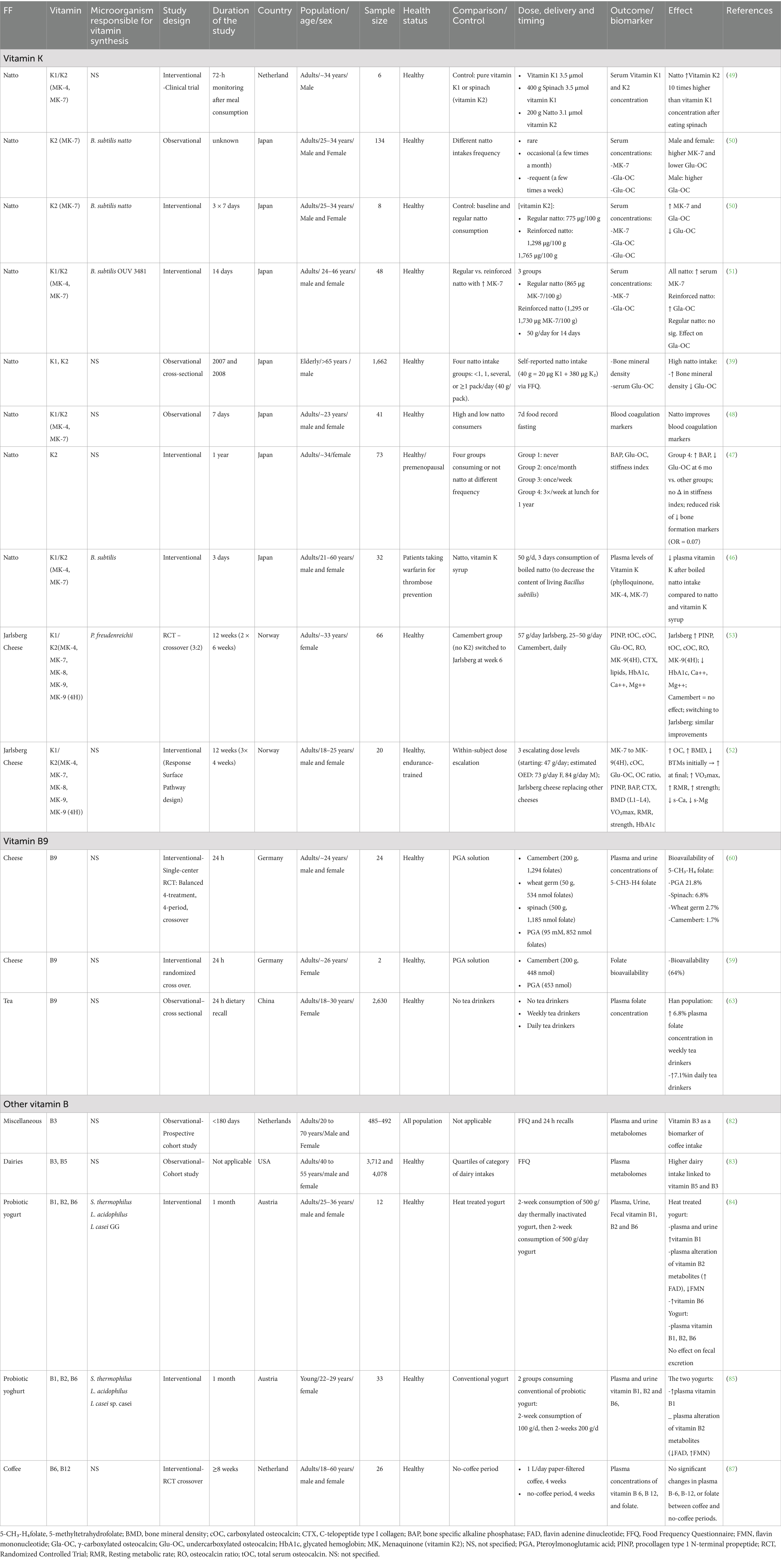
Table 2. Effect of the consumption of FF fortified in vitamin K and B through fermentation on vitamin status and related health outcomes in human.
3.1 Fermented soybeans and cheese are valuable sources of vitamin K
3.1.1 Food sources of vitamin K
Vitamin K exists as phylloquinone (K1) and menaquinones (K2, MK-4 to MK − 13), which differ in dietary sources, bioavailability, and function. Vitamin K1, primarily found in leafy greens and plant oils, accounts for the majority of dietary intake, but has limited bioavailability (34, 35). In contrast, vitamin K2, produced by bacterial fermentation, dairy products, and gut microbiota, exhibits longer half-life and enhanced extrahepatic distribution, making it particularly relevant for cardiovascular and bone health (35–38). Compared to vitamin K1, MK-7 and MK-9 exhibit prolonged half-lives, enhancing their bioavailability and ensuring sustained activation of vitamin K-dependent pathways (34, 39). Among menaquinones, MK-7, abundant in natto, is distinguished by its superior bioavailability and tissue retention, which is attributed to its longer unsaturated side chain (34). Swiss-type cheeses, such as Emmentaler and Jarlsberg, are rich in MK-9 and MK-9(4H), primarily due to the activity of Propionibacterium freudenreichii during cheese fermentation. Notably, Jarlsberg cheese was developed in Norway in the 1950s based on Emmental cheesemaking techniques, with researchers at the Agricultural University of Norway successfully adapting bacterial fermentation processes to create the cheese’s distinctive nutty flavor and characteristic holes via P. freudenreichii metabolism (40).
3.1.2 Role of vitamin K and mechanism of action
The Adequate Intake (AI) for vitamin K, as established by the U.S. National Institutes of Health, is 120 μg/day for adult men and 90 μg/day for adult women (41). Vitamin K is essential for post-translational γ-carboxylation of glutamate residues in vitamin K-dependent proteins (VKDPs), a reaction catalyzed by γ-glutamyl carboxylase (GGCX) (Table 1) (34, 42, 43). This modification enables the activation of proteins involved in coagulation (factors II, VII, IX, and X), bone metabolism (osteocalcin), and vascular health (matrix Gla-protein, MGP) (44, 45). The vitamin K cycle, driven by vitamin K epoxide reductase (VKOR) and vitamin K reductase (VKR), ensures continuous recycling of vitamin K to maintain γ-carboxylation activity (42, 43). Beyond coagulation, vitamin K plays an integral role in bone metabolism, where it regulates osteocalcin carboxylation, a key factor in bone mineralization and skeletal integrity (44). The primary consequence of vitamin K deficiency is a bleeding tendency due to the relative inactivity of procoagulant proteins (2).
Vitamin K deficiency results in uncarboxylated osteocalcin, which impairs bone formation and increases fracture risk. Similarly, in the vascular system, MGP serves as a potent inhibitor of arterial calcification, preventing calcium deposition in soft tissues. Insufficient vitamin K leads to an accumulation of uncarboxylated MGP, which has been linked to vascular calcification and elevated cardiovascular disease risk (45). Studies indicate that K2 supplementation helps mitigate vascular calcification, reinforcing its importance in cardiovascular health (35). Emerging research suggests that vitamin K also plays roles in mitochondrial function, inflammation modulation, and neuroprotection, though further studies are needed to clarify its broader physiological impact (34, 35).
3.1.3 Health effects of natto and cheese consumption
FF such as natto and Jarlsberg cheese are valuable dietary sources of vitamin K2. This review synthesizes findings from seven primary studies on natto (39, 46–51) and two on Jarlsberg cheese (52, 53), focusing on elderly individuals, premenopausal and postmenopausal women, and individuals with varied vitamin K status (Table 2).
While natto has been extensively studied, particularly in Japan, recent findings suggest Jarlsberg cheese – fermented with P. freudenreichii—may serve as an alternative vitamin K2 source in Western diets. However, comparative studies between these foods remain scarce, and the extent to which their different menaquinones [MK-7 vs. MK-9(4-H)] exert similar physiological effects remains unclear.
Natto is particularly rich in menaquinone-7 (MK-7), and serum MK-7 concentrations following natto intake have been shown to increase significantly and remain detectable for over 72 h, indicating prolonged bioactivity (49, 50). Tsukamoto et al. (50, 51) demonstrated that habitual natto consumption leads to a significant increase in serum MK-7 concentrations in a dose-dependent manner, with higher intake correlating with elevated γ-carboxylated osteocalcin (cOC) levels, particularly in men.
Sakamoto et al. (48) did not measure MK-7 levels but provided evidence that individuals with higher natto intake had increased plasma vitamin K concentrations and improved blood coagulation markers. Fujita et al. (39) found that elderly men with higher natto intake exhibited higher bone mineral density (BMD) at the total hip and femoral neck, with these effects likely mediated by vitamin K’s role in osteocalcin activation.
Several studies have established a dose-dependent relationship between natto intake and undercarboxylated osteocalcin (ucOC), a biomarker of vitamin K deficiency. Katsuyama et al. (47) reported that premenopausal women who consumed natto three times a week had significantly lower ucOC levels and higher bone-specific alkaline phosphatase (BAP) levels compared to those with lower natto intake. These findings suggest that natto promotes bone formation and may help maintain skeletal integrity before menopause.
Natto has also been linked to improved vascular health and regulation of coagulation. Schurgers and Vermeer (49) found that MK-7 from natto is associated with enhanced MGP activation. Tsukamoto et al. (50) showed that natto intake reduces plasma protein induced by vitamin K absence-II (PIVKA-II) levels, a marker of vitamin K deficiency, and lowers urinary calcium excretion, suggesting improved calcium retention.
For individuals taking the anticoagulant warfarin, natto consumption may pose potential negative interactions. Indeed, Homma et al. (46) demonstrated that consuming natto significantly increased plasma vitamin K concentrations, which could counteract warfarin’s anticoagulant effects. However, boiling natto markedly reduces its vitamin K bioavailability, potentially offering a method for patients on anticoagulation therapy to consume natto with a lower risk of drug interaction.
Jarlsberg cheese, a natural source of MK-9 (4H), has gained attention for its potential benefits to bone health and metabolism. A randomized controlled trial in premenopausal women demonstrated a significant increase in serum MK-9 levels and bone formation markers, particularly procollagen type 1 N-terminal propeptide (PINP), following regular consumption of Jarlsberg. When participants switched from Camembert (control) to Jarlsberg, their vitamin K2 levels and calcium absorption improved, suggesting a positive effect on bone metabolism (53). A follow-up study by the same group extended the intervention to 12 weeks, confirming sustained increases in PINP, total osteocalcin (tOC), and cOC, without elevating the bone resorption marker CTX. These findings support an anabolic effect on bone, possibly mediated by improved vitamin K2 status (52).
Jarlsberg cheese may also influence metabolism more widely. Indeed, in the same study, a reduction in glycated hemoglobin (HbA1c) suggests potential benefits for glucose regulation, though the underlying mechanisms are not yet clear (53). According to data by Walther et al. (54), Swiss cheeses, such as Emmentaler and Raclette, also contain notable amounts of long-chain menaquinones, with up to 549 μg/kg of MK-9(4H) in Emmentaler and 826 μg/kg of MK-7 in Raclette. However, to reach the dosages applied in clinical trials (180–200 μg/day), daily consumption would need to exceed 100 g, which may not be feasible for all populations due to high fat and salt content.
One possible factor is the presence of P. freudenreichii, a bacterial strain commonly found in cheese, which produces MK-9 and may contribute to vitamin K2 availability. However, unlike natto, the effect of MK-9 from cheese on blood clotting markers remains unclear due to a lack of direct intervention studies. More research is needed to determine whether MK-9 from dairy products offers the same cardiovascular and coagulation benefits as the menaquinones found in fermented soy.
The extended presence in circulation may explain why MK-7-rich natto and MK-9-rich cheese provide lasting physiological effects, particularly in populations at risk for osteoporosis and arterial calcification.
3.1.4 Differences in responses and most relevant target groups
Variability in response to vitamin K2-rich foods has been observed across demographic groups. Men appear to exhibit a stronger correlation between MK-7 intake and γ-carboxylated osteocalcin activation, whereas older adults show greater improvements in bone density following natto consumption compared to younger individuals (39, 50). Individuals with low habitual vitamin K intake, particularly those consuming minimal FF, may experience more pronounced benefits from dietary interventions (34).
The strongest evidence for benefits exists among elderly individuals at risk for osteoporosis and vascular calcification, postmenopausal women experiencing accelerated bone loss, and individuals with vitamin K deficiency indicated by elevated PIVKA-II levels. Emerging research suggests potential applications for individuals with cardiovascular risk factors, as well as those with metabolic disorders such as diabetes, where reductions in HbA1c have been noted following Jarlsberg cheese consumption (53).
Despite strong evidence supporting the health benefits of natto and Jarlsberg cheese, several limitations remain. Most studies have focused on elderly individuals and postmenopausal women, limiting generalizability to younger populations (39, 47). Additionally, differences in dietary habits and gut microbiota may influence vitamin K metabolism and bioavailability (34).
While vitamin K2 is endogenously produced by gut bacteria, its systemic contribution to vitamin status is limited due to inefficient colonic absorption (34). Nonetheless, the gut microbiota plays an indirect role in vitamin K metabolism by modulating bile acid turnover and menaquinone production (35). Menaquinone profiles differ by microbial species: P. freudenreichii in Jarlsberg cheese synthesize MK-9 (4H), whereas B. subtilis in natto produces MK-7 (49, 53). These microbial contributions highlight the importance of FF as external sources of bioavailable vitamin K2.
Existing research has predominantly examined Japanese and European cohorts, leaving a gap in understanding how regional dietary patterns and microbiota-driven metabolism affect vitamin K bioavailability (35).
Further studies should investigate whether MK-7 from natto and MK-9 from cheese exert equivalent effects on bone and cardiovascular health, given their structural differences (49). Long-term intervention trials are needed to evaluate whether routine intake of vitamin K2-rich FF reduces fracture risk and cardiovascular disease incidence (44). Moreover, vitamin K2’s role in glucose metabolism and insulin sensitivity warrants further exploration (53).
Lastly, the potential of plant-based FF as alternative vitamin K2 sources should be investigated to meet the dietary needs of vegetarians and vegans, given their lower menaquinone intake, mainly present in animal-based products (34).
3.2 Folate (vitamin B9) produced during fermentation is able to contribute to an adequate folate status
3.2.1 Food sources and forms of folates
Folate, also known as vitamin B9, is a generic term used for a family of compounds known as vitamers, differing by their oxidation state and the length of their glutamate residue. The chemical lability of all naturally-occurring folates results in a significant loss of biochemical activity during food storage and processing. In contrast, the synthetic form of this vitamin, folic acid, is a fully oxidized monoglutamate and the most chemically stable form (55). The bioavailability of natural folates compared to the synthetic form, folic acid, is variable and estimated to be between 10 and 98% (56). Folates are found in a variety of foods, including meat liver, green leafy vegetables, some other fruits and vegetables as well as legumes and cereals (as wholegrain products) (2). The majority of the population can meet the RDA with a daily intake of 400 μg of folate (2).
3.2.2 Role and function of folates
Folate functions as a coenzyme or co-substrate in one-carbon transfers in the synthesis of nucleic acids (DNA and RNA) and metabolism of amino acids (16). Folate is also essential for the conversion of homocysteine to methionine and the synthesis of S-adenosyl-methionine, an important methyl donor for more than 100 methyltransferases for a wide range of substrates such as DNA, hormones, proteins, neurotransmitters and membrane phospholipids, all of which being regulators of important physiological processes (55). Folate deficiency impairs DNA replication and cell division, that can lead to megaloblastic anemia, or neural tube defects (Table 1) (57).
3.2.3 FF sources of folates in human studies
Despite the high number of articles dealing with the increase in folate content of food using folate-producing microorganisms, only three human studies were identified in the literature, two on camembert cheese, and one on tea (Table 2).
The definition of dietary folate equivalents is generally based on the assumption of 50% bioavailability (58). The aim of Mönch et al. (59) work was to assess the relative bioavailability of folates in several folate-rich foods, including low-fat camembert cheese. They conducted a short-term human study measuring plasma folate levels after ingestion of the foods using an isotopic dilution assay. This demonstrated that folate from the tested foods including camembert was absorbed with matrix-dependent bioavailability and with high interindividual variations, thus showing that total folate concentration in food does not always correlate with its bioavailability. The authors also measured the folate concentration of the most abundant vitamers in low-fat camembert from different brands, to investigate the potential variation in folate bioavailability within the same food. Folate concentration in camembert ranged from 224 to 647 nmol/100 g (59, 60), which is 5 to 15 times higher than in milk (61) if calculated for dry weight. Moreover, a significant variation in folate bioavailability has been reported depending on the cheese brand. Notably, Mönch et al. (59) observed that low-fat camembert in their study exhibited substantially higher folate bioavailability (65–71%) compared to the 8.8% reported in a previous study on a different brand (60). Despite containing a lower total folate content (224 nmol/100 g) compared to the previously studied camembert (647 nmol/100 g), the cheese analyzed in this study presented a distinct folate profile. Specifically, 5-formyltetrahydrofolate was the predominant vitamer, as opposed to tetrahydrofolate in the higher-folate cheese. An intraindividual comparison of participants who participated in both studies further confirmed the striking difference in bioavailability between the two cheese brands. These findings highlight that fermentation processes, cheese composition, and folate vitamer distribution significantly influence folate bioavailability, reinforcing that a food product (e.g., camembert cheese) cannot be considered a homogeneous dietary standard with predictable folate bioavailability (59).
Several hypotheses have been proposed regarding the differences in folate bioavailability, including differences in the kinetics and bioavailability of folate vitamers, particularly polyglutamate forms; the presence of deconjugase inhibitors and entrapment of folates within the food matrix (60).
Tea can also be a source of folates, reaching up to 7 μg/100 mL of brewed tea (62). A positive correlation has been observed between tea consumption and plasma folate concentration in China, with specificity varying by ethnic group and tea type. Han women, drinking unfermented tea weekly, and Uighur women, drinking fully fermented tea weekly, exhibited this positive correlation; however, the role of fermentation was unclear and deserves further research (63).
Although many articles are dedicated to the study of fermentation as a way to improve folate content of FF, the lack of human studies together with the large differences in the bioavailability of vitamin B9 due to a food matrix effect or the vitamers present in the FF, leads to under- or overestimated the impact of FF-enriched food on the folate status of populations. Further human studies would help to better understand how these FF could be used to improve vitamin B9 status, especially for vulnerable populations at risk of deficiency, such as young children and women of childbearing age.
3.3 FF as source of the other vitamin B (B1, B2, B3, B5, B6, B12)
In addition to vitamin B9, other vitamin Bs can be produced by different microorganisms during the fermentation of various foods. However, this capability is strain-specific, as microorganisms can either synthesize or consume them. Five studies were dedicated to the effect of FF consumption on vitamin B status (Table 2).
3.3.1 Food source and roles of the other vitamin B
Thiamine (vitamin B1) is found in whole grains, legumes, nuts, and meat. The RDA is 1.6 mg for adult males. A deficiency can result in conditions such as beriberi or Wernicke-Korsakoff syndrome, which affect the nervous system (64). Vitamin B1 is an essential cofactor involved in carbohydrate metabolism, the respiratory chain, amino acid catabolism, and nucleic acid biosynthesis. In addition, thiamine plays a neuro-modulatory role by supporting acetylcholine neurotransmission and helps maintain neuronal and cellular membrane integrity (65). Recently, LAB strains have been screened for their ability to produce this vitamin (Table 1) (66).
Food sources of riboflavin (vitamin B2) include liver, eggs, dark green vegetables, legumes, whole and enriched grain products, milk, and fermented products (67). In recent years, different lactic acid bacteria selected for their ability to overproduce this vitamin have been proposed for the in situ biofortification of various FF (68–71). The RDA is 1.4 mg for adult males. Ariboflavinosis can cause symptoms such as cracked lips, a sore throat, and skin disorders. Vitamin B2 is the precursor of the coenzymes flavin mononucleotide (FMN) and flavin adenine dinucleotide (FAD) that are involved in critical metabolic pathways such as the cycle of Krebs, redox reactions, as well as in the biosynthesis of other vitamins (Table 1).
Dietary niacin (vitamin B3) is primarily found in meat, poultry, fish, eggs, dairy products, nuts, seeds, whole grains, legumes, and certain vegetables. The RDA is 16 mg/day for adults. A deficiency of niacin leads to pellagra, a condition marked by dermatitis, diarrhea, and dementia (72). Vitamin B3 act as precursor of nicotinamide adenine dinucleotide (NAD) and NAD phosphate (NADP), both of which play crucial roles in energy production and redox reactions. Recent studies highlighted its anti-inflammatory and antioxidant properties. Additionally, this vitamin helps regulate blood cholesterol levels, particularly by boosting high-density lipoprotein (HDL) cholesterol (Table 1) (73, 74).
Pantothenic acid (vitamin B5) is primarily found in meat, eggs, dairy, whole grains, legumes, avocados, some vegetables and mushrooms. The RDA is considered to be between 5 and 7 mg for adults. Vitamin B5 plays a crucial role in metabolizing carbohydrates, proteins, fatty acids. In recent years, D-pantothenic acid has been investigated for its potential in treating neurodegenerative diseases (Table 1) (75).
The richest sources of pyridoxine, pyridoxal and pyridoxamine (vitamin B6) include fish, beef liver and other organ meats, potatoes and other starchy vegetables, and fruit. The RDA is about 2.0 mg/day for adults. Vitamin B6 deficiency is linked to several pathological conditions, including end-stage renal disease, chronic renal insufficiency, and other kidney-related disorders. Moreover, an insufficient intake of this vitamin can lead to malabsorption syndromes, such as celiac disease and inflammatory bowel diseases, including Crohn’s disease and ulcerative colitis (76). Vitamin B6 plays a role in various biological processes, including amino acid, sugar, and lipid metabolism (Table 1) (77).
Cobalamin (vitamin B12) is the only vitamin exclusively from microbial origin, since synthesized by some bacteria and archaea. Cobalamin is present in foods of animal origin, due to the production by microorganisms present in the digestive tract, which is absorbed and incorporated into animal tissues. Thus, animal-sourced products such as meat, fish, eggs and dairy products are main dietary source of cobalamin (WHO, 2004). RDA between 3.8 and 20.7 μg is required to maintain adequate levels and prevent deficiency in healthy adults and older individuals (6). Among bacteria of food interest, Propionibacterium spp. is well known for producing cobalamin to varying extents (78). This capability has been reported also in some strains of lactic acid bacteria (LAB) (79). However, this property remains controversial, as LAB often synthesize corrinoid compounds (pseudo-cobalamin) that are biologically unavailable to humans (80). A deficiency in this vital nutrient can lead to both hematological and neurological disorders, including the serious condition known as pernicious anemia (81). Vitamin B12 is essential in cellular metabolism, especially in DNA synthesis, methylation and mitochondrial metabolism (Table 1).
3.3.2 FF consumption contributes to the vitamin B status of different population
Five studies were identified as addressing the effect of consuming some FF on the nutritional status of these metabolites, and all of them tackled different vitamin B deficiencies simultaneously. Plasma or urine metabolomes analysis have been recently used to detect biomarkers from FF consumption (Table 2).
In a prospective cohort study of Dutch men and women (20 to 70 years), non-targeted metabolomics analyses were performed on plasma and urine samples (n = 485 and n = 492, respectively) of a subcohort of participants (n = 531) (82). Urinary niacin (vitamin B3) has been reported as the unique vitamin B candidate biomarker of habitual coffee intake and the other vitamin B (B2 and B5) also present in coffee were not identified (82). Whether the fermentation of coffee is responsible for its vitamin B content remains to be clarified. A recent cohort study conducted within the Framingham Heart Study Offspring (3,712 participants, average age ~55 years) and third generation cohorts (4,078 participants, average age ~40 years) utilized both targeted and untargeted biomarkers to analyze plasma metabolomes associated with quantity and type of dairy intake (83). A link between dairy consumption and the circulating levels of pantothenate (vitamin B5) and niacin (vitamin B3) was established. Specifically, pantothenate concentrations demonstrated a statistically significant association with overall dairy intake in the third-generation cohort, aligning with observations in the offspring cohort. However, correlation with milk only was not significant, suggesting that fermentation processes might contribute to increased vitamin levels. Notably, plasma pantothenate displayed a statistically significant and directionally consistent association with cream and butter in the offspring cohort (83).
Two interventional studies investigated the effect of either the consumption of traditional yogurt fermented with Streptococcus thermophilus and Lactobacillus acidophilus or probiotic yoghurt fermented with Lacticaseibacillus casei GG in addition, on thiamine (vitamin B1), riboflavin (vitamin B2), and pyridoxine (vitamin B6) status in healthy adults (84, 85). The first study aimed to determine whether viable probiotics in yoghurt could influence the status of B1, B2, and B6 vitamins in healthy adult humans; either by using or producing these vitamins (84). Thus, 12 volunteers (six healthy males and 6 healthy females, aged 25–36 years) consumed 500 g of yoghurt daily during 4 weeks. They received yoghurts containing thermally inactivated cultures during the first 2-week period and yoghurts not submitted to heat treatment during the second 2-week period. Thiamin levels in plasma significantly declined after the first 2 weeks and continued to decrease during the second period,. Thiamine urinary excretion and B2 metabolites were also affected. Indeed, FAD increased significantly after consuming heat-treated yogurt but dropped after switching to untreated yogurt. Conversely, FMN levels decreased initially but rose later, though not significantly. Free riboflavin in plasma and urine showed a slight, non-significant increase. Pyridoxal-5-phosphate (vitamin B6) levels increased with heat-treated yogurt but decreased afterwards, with no significant differences observed. Therefore, these results suggest that the presence of Lacticaseibacillus casei GG as a probiotic in yoghurt would reduce the bioavailability of vitamins B1, B2 and B6, despite the fact that it was known to produce thiamine, riboflavin and folate in culture medium (84, 86). Investigation of the potential vitamin B-producing capabilities of the bacteria used in yogurt preparation was not shown. In a similar approach by the same research team with lower amounts of yogurt consumption and another probiotic strain (Lacticaseibacillus casei subsp. casei) were used. This intervention study was conducted in young healthy Austrian women (n = 33) with the aim to investigate whether probiotic and/or traditional yoghurt bacteria were able to influence the status of B1, B2 and B6 vitamins (85). The results indicated that daily consumption of 200 g of both probiotic and conventional yoghurt for 2 weeks resulted in the increase levels of plasma thiamine and free riboflavin, while the urinary excretion of these vitamins remained unaffected. Interestingly, the plasma concentration of FAD decreased significantly after consuming 100 g of yoghurt/day, while, conversely, the plasma levels of FMN and free riboflavin increased significantly, suggesting a different physiological role of these flavins. The average status of vitamin B6 was unaffected by yoghurt intake in both plasma and urine. Overall, the authors concluded that the reported variations in plasma vitamins levels were more likely consistent with yoghurt consumption as a fermented dairy product rather than by the specific ingestion of probiotic bacteria (85). These two studies on a limited number of volunteers, differing in sex, the amount of FF and the bacterial strains used, gave different results, underlying the need to evaluate the effect of FF consumption enriched in vitamins by fermentation in human on a larger scale.
One study investigated the link between coffee consumption and levels of certain B-vitamins in the blood. The impact of daily consumption of 1 L of paper-filtered coffee on plasma total homocysteine concentrations was assessed in healthy subjects through a randomized, controlled, and crossover intervention (87). Briefly, 26 healthy participants (male and female, aged 18–60 years) were involved in a 4-week study (coffee consumption and no-coffee period) with a crossover design. This study also monitored plasma concentrations of vitamin B6, vitamin B12, and folate, as their deficiencies might be related to elevated homocysteine concentrations. However, no significant differences were found between the coffee and no-coffee periods (87). Nevertheless, no information on the vitamin B6 and B12 content of coffee were provided.
Surprisingly, no studies were found on the effect of FF enriched in vitamin B12 by fermentation in humans. Vitamin B12 is absent from non-animal-based products, and many people in the world consume no or little of this food category due to affordability or personal choices; therefore, this topic deserves specific attention. In addition, different microorganisms encountered in FF may produce pseudo-cobalamin, which is inactive in humans. Intervention studies using FF fortified in vitamin B12 are obviously required.
3.4 Vitamin A and D, the left aside of vitamin production through fermentation
3.4.1 Vitamin A is produced by a few microorganisms not present in FF
Vitamin A is naturally present only in animal-derived foods, while many fruits and vegetables can provide carotenoids that can be converted into vitamin A at intestinal level (15). The main dietary sources of vitamin A include liver, dairy products, and fish, while provitamin A carotenoids are abundant in colorful fruits and vegetables, such as carrots, sweet potatoes, and spinach. The RDA of vitamin A is 900 and 700 μg/day for adult men and women, respectively. Vitamin A is an essential nutrient best known for supporting vision and regulating cell function, proliferation, and tissue homeostasis. A deficiency can result in poor nutritional status, increased mortality risk, weakened overall health, reduced tissue growth, slower metabolic processes, and heightened susceptibility to infections (15).
Different microorganisms belonging to Erwinia or Agrobacterium genera among others, can also synthesize vitamin A or provitamin A, but only a few of them are present in FF (20). Microorganisms may also modulate carotenoids bioavailability (26). Only one study dealt with the effect of ingestion of fermented orange juice or its non-fermented counterpart on carotenoid profile on blood carotenoids. Bioavailability of carotenoids was higher when orange juice was fermented. However, carotenoids were most probably coming from the orange juice itself, and not from the action of microorganisms since carotenoid profiles were similar in the two juices. The yeast species used for fermentation, Pichia kluyveri var. kluyveriis is not known for its carotenoid producing capability (88).
3.4.2 Vitamin D can be produced by Saccharomyces cerevisiae, but is not used to increase vitamin D content in FF
Vitamin D, also referred to as calciferol, is a fat-soluble vitamin that can either be synthesized endogenously in the skin following exposure to UV-B irradiation or can be provided from food sources and dietary supplements. The version made in the skin is referred to as cholecalciferol (vitamin D3) whereas the dietary form can be vitamin D3 or a plant-origin compound known as ergocalciferol (vitamin D2). Vitamin D obtained from sunlight, food, and supplements is initially biologically inactive and must undergo two hydroxylation processes in the body to be converted into its active form, 1,25-dihydroxyvitamin D [1,25-(OH)2D], also known as calcitriol. The RDA of vitamin D is 400 and 800 μg/day for adult male and elderly, respectively (2, 16).
Vitamin D, through its hormonal form calcitriol is a central regulator of calcium and phosphate homeostasis, which is necessary for normal bone mineralization, muscle function, nerve conduction, and overall cellular activity in the body (2, 16). Vitamin D also plays various other roles in the body, such as reducing inflammation and modulating processes like cell growth, neuromuscular function, immune responses, and glucose metabolism (16). Additionally, vitamin D has immunomodulatory effects that may alter responses to infections in vivo. These properties, which include promoting cell differentiation and modulating immune function, are why vitamin D derivatives are effectively used in the treatment of various skin conditions. Adequate vitamin D levels prevent rickets in children and osteomalacia in adults, while also helping protect older individuals from osteoporosis (16).
Literature reports the capability of yeasts, particularly Saccharomyces cerevisiae, to synthesize ergosterol, one of the precursors of vitamin D, for industrial production, but literature on the use of yeast to increase vitamin D content of FF and their possible contribution to vitamin D intakes is not available (19). The few studies dealing with the ingestion of vitamin D from FF are from dairy products, where the vitamin D is added through fortification (89, 90). Whether fermentation can be used to increase the vitamin D content in food and improve dietary intake of vitamin D remains to be explored.
3.5 Detailed FF characterization is key for essentiality of nutrients
FF characterization is necessary to be able to ascertain the scientific evidence of health claims based on FF enriched in vitamin by fermentation (30). Different factors may influence the physiological effect of vitmains, including batch-to-batch variability, their concentration in FF as well as difference in chemical structure (vitamers, precursors) and bioavailability. The study on camembert cheese is one of the rare examples where the major vitamin B9 vitamers were quantified in the FF and in the plasma after ingestion of the FF (59, 60). The different bioavailability patterns and the important individual observed variation underscore the need to implement similar studies in different food matrixes, with various fermentation types, in order to verify the real contribution of the FF enriched in vitamin by fermentation at a large scale.
In addition, the formulation of the FF should be specified and this is particularly relevant since the food matrix may influence the absorption of the vitamin. Indeed, in the same example, folate from spinach was better absorbed than folate from cheese and different hypotheses were raised to explain this result (see corresponding paragraph). This example highlights the need to evaluate many FF enriched in vitamins through fermentation to ascertain the efficiency of the strategy in different contexts.
The variability between individual foods, which may influence the vitamin content of the FF should also be characterized. Of the 18 studies discussed in the present paper, only two, on cheese and vitamin B9, investigated the possible differences between two brands of the same cheese and highlighted significant differences. This variability would be even more important to assess in the case of FF with non-controlled fermentation (spontaneous, backslopping, initial high endogenous microbiota), since the microorganisms involved differ from one fermentation to another. As microorganisms may produce or consume vitamin B9, the overall production/consumption balance should be assessed. Indeed, many traditional foods prepared using spontaneous or backslopping fermentation at room temperature exhibit almost at total disappearance of folate initially present in the raw material or a huge increase in the vitamin B9 content, able theoretically to cover a large part of the RNI (30%) (91, 92).
Where controlled fermentation with known microorganisms is used, sufficient information must be provided, in addition to the identification of the microbial species, to enable the control authorities to carry out their own monitoring (30). On the 18 studies presented here, only five specified the species used for fermentation.
More generally, while the FF category is almost always indicated (dairy products, fermented legumes, etc.), the precise description of the food is almost never sufficient. For example, ingredients (and even less their composition) are not necessarily mentioned, the process is not always described. Although, the food safety of the FF presented in this review is never mentioned, we can hypothesize that in interventional studies, the ethical approval included this aspect and that it is difficult to assess it from observational studies.
3.6 Role of microbiota to improve vitamin status
Microorganisms in the gut can influence the host’s vitaminic status by synthesizing vitamin or regulating their absorption. Notably, bioavailability appears to be enhanced by the production of short-chain fatty acids, which are the end products of microbial polysaccharide fermentation (93). In turn, dietary vitamin intake can modulate the composition and functionality of the microbiome, which in its turn influences bioavailability of vitamins. Microorganisms in FF could also synthesize vitamins while transiting through the digestive tract. If the role of gut microbiota is well documented for vitamin K (see corresponding section), it is not the case for all B vitamin.
3.6.1 Synthesis of vitamin by microorganisms in the gut
While most vitamins are obtained from food, some can be synthesized by gut bacteria and absorbed in the colon (94). The role of intestinal bacteria in vitamin K production has been recognized for decades, and their contribution to other vitamins synthesis has also been documented (26). Bacterial synthesis may provide 10–50% of the body’s vitamin K requirements, though the exact proportion remains uncertain (11). Most menaquinones (MK-n) are produced in the colon without bile salts, raising questions about their absorption, as bile salts are typically required for vitamin K uptake in the small intestine. If absorbed, MK-n from colonic bacteria likely follows a bile-salt-independent route, whereas those produced in the ileum—a smaller fraction—may be absorbed via a bile-dependent pathway (95). However, the bioavailability of MK-n from gut bacteria is limited, as they remain within bacterial membranes rather than being freely available (96).
Data on the role of intestinal bacteria from the gut on vitamin B productions is less documented. Metagenomic analyses have revealed that some bacteria, such as Bacteroidetes and Actinobacteria, possess the genetic potential to synthesize vitamin B1, whereas most Firmicutes do not (97). Lactobacillaceae and Bifidobacteriaceae produce riboflavin, which may be utilized by colonic epithelial cells, though its exact contribution in vivo remains unclear (98, 99). While riboflavin biosynthesis has been well studied in B. subtilis and Escherichia coli, a systematic genome analysis of 256 human gut strains found that 40–65% could theoretically produce one or more B vitamins (B1, B2, B3, B5, B6, B7, B9, and B12) (100). Complete folate biosynthesis pathways were detected in nearly all Bacteroidetes, Fusobacteriota (syn. Fusobacteria), and Proteobacteria genomes, whereas Actinobacteria and Firmicutes often lacked the para-aminobenzoic acid biosynthesis pathway (26).
The de novo synthesis of cobalamin (vitamin B12) is an intricate process that involves over 30 enzymatic steps and more than 60 gene families (101). This capability is exclusive to certain bacteria and archaea. Transporters of vitamin B12 are present only in the upper parts of the digestive tract. Therefore, the eventual synthesis by bacteria from the colon should not contribute significantly to the vitamin B12 status of the host (102). LAB are present in higher relative abundance in the small intestine, but to our knowledge, no studies have investigated their eventual role in contributing to cobalamin absorption.
3.6.2 Does vitamin consumption enhance gut microbiota–mediated vitamin biosynthesis/ absorption?
Within the human gut microbiota, increased folate intake has been positively correlated with a higher abundance of Faecalibacterium, Akkermansia, and Roseburia genera, as observed in mucosal and colonic biopsy samples (103). Furthermore, carotenoid intake and status have been linked to increased microbial diversity and a rise in beneficial bacteria. The most relevant studies on the impact of carotenoids on gut microbiota have been reported by Bernabeu et al. (104). There is a strong evidence that carotenoids exert a prebiotic effect, promoting the growth of specific bacterial species such as Lachnospiraceae, Bifidobacterium, and Prevotellaceae. Park et al. (105) demonstrated that vitamin B1 significantly influences the survival and competition of bacteria within the gut microbiota. They also showed that vitamin B1 plays a role in butyrate production and contributes to the regulation of acetate levels in the intestinal environment. Additionally, microbial riboflavin production is influenced by a fiber-rich diet (106). Bacteria require folate for growth: some are prototrophic and can synthesize it from environmental precursors, while others, known as auxotrophic bacteria, must acquire it directly from their surroundings. These findings suggest that the gut microbiota is a key source of folates and that changes in its composition—driven by factors such as fiber consumption—may affect folate requirements (107, 108).
Considering that microorganisms can produce and use vitamin, an interplay between microorganism for vitamin occurs. This has been demonstrated, for example, in a synthetic co-culture experiment using bacteria from a human fecal sample, where differences in cross-feeding were observed (109). The bacteria in the gut are also able to capture and differentiate vitamin B12 analogs, competing in the gut (110).
On the whole, the contribution of gut microbes to vitamin status and the effect of FF consumption on vitamin synthesis in the gut remains to be studied more specifically.
4 Perspectives, gaps and conclusion
This systematic narrative review highlights the potential of FF as a natural way for enhancing vitamin intake via microbial production. However, significant gaps remain in understanding the effectiveness and reliability of this approach (Table 3). The bioavailability of vitamins from FF varies depending on the microbial strains, fermentation conditions, vitamers produced, and the food matrix involved. There is a notable lack of human studies on several essential vitamins, including B12, which is particularly important for population consuming no or low animal-based products. These sources of variation are often overlooked when vitamin intakes are estimated through food consumption surveys, which can lead to a significant misestimation of vitamin intakes. An example from Ethiopia showed that the sampled population had totally inadequate dietary intakes of vitamin B12, but only 25% of the population had low serum cobalamin levels (111). One hypothesis is that high consumption of FF and the role of fermentation, which has been estimated but not experimentally validated (112).
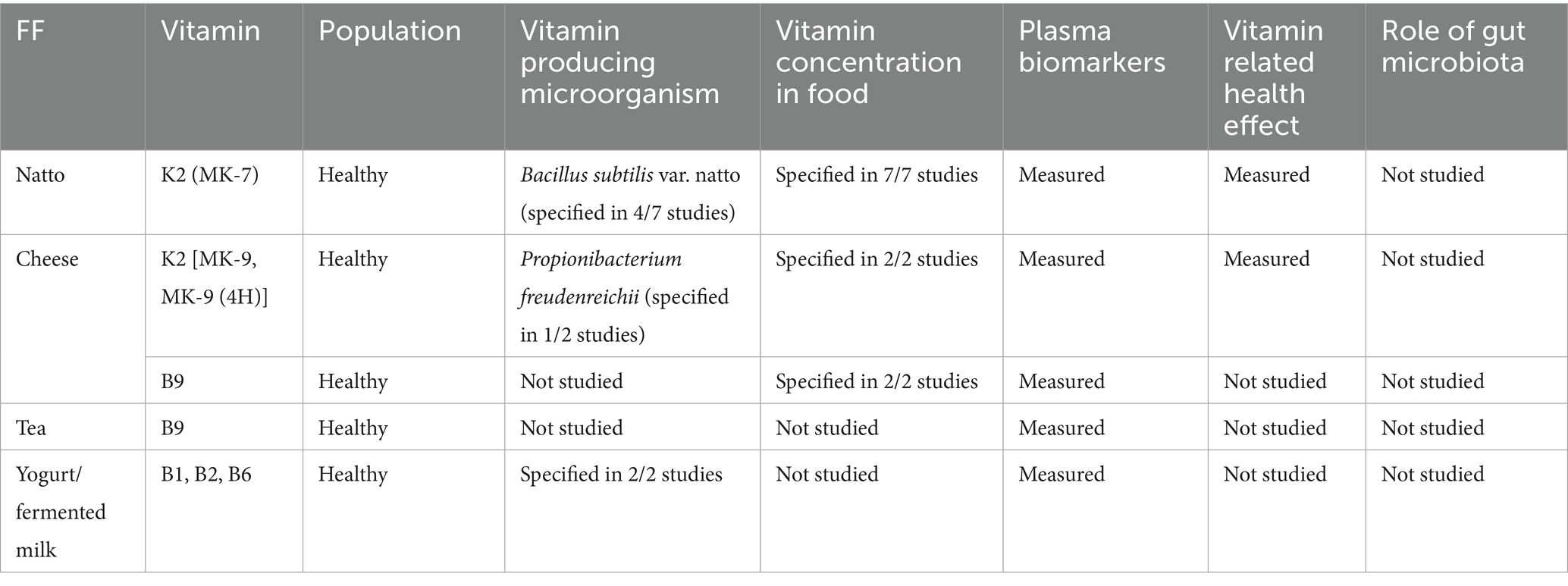
Table 3. Overview of the FF, microbiota and summary of the findings.
The wide variety of FF around the world, the microorganisms responsible for their fermentation and their potential contribution to vitamin intakes has also been underexplored. Many microorganisms very well known for their vitamin synthesis capabilities, such as Saccharomyces cerevisiae, which can produce vitamin B9 in large amount, have not been studied in humans. All the data presented in this review are from Europe, Asia, and the USA, excluding other parts of the world that also consume various FF, with potentially different vitamin profiles (9). Populations at risk of deficiencies include young children, women of childbearing age and the elderly, and future studies should include these different categories. This wide variety of FF may a serve as a source of inspiration for one country to learn from another, utilizing the microbial diversity encountered in these traditional FF. These FF can be used as single strains but also combined to explore cumulative vitamin production by using microorganisms with different vitamin potentials together. Indeed, examples of the use of combination of strains with synthesis vitamin capabilities are rare, and not always efficient (113).
Numerous studies lack comprehensive information regarding the composition, safety, and microbiological content of the examined FF. To endorse FF-based vitamin fortification as a public health strategy, future research should concentrate on well-designed human intervention studies, thorough evaluations of vitamin variants and food matrices, and uniform documentation of fermentation methodologies and microbial strains. Moreover, the impact of the gut microbiota on vitamin status, encompassing vitamin synthesis, absorption, and microbial cross-feeding, remains inadequately defined. Addressing these gaps will enable better assessment of the nutritional impact of FF and guide the development of evidence-based dietary recommendations.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding authors.
Author contributions
EK: Conceptualization, Supervision, Writing – original draft, Writing – review & editing. EA: Writing – original draft, Conceptualization, Writing – review & editing. M-CC-V: Writing – original draft, Conceptualization, Writing – review & editing. EC-B: Writing – original draft, Conceptualization, Writing – review & editing. JM: Writing – original draft, Conceptualization, Writing – review & editing. DNN: Writing – original draft, Conceptualization, Writing – review & editing. RP: Conceptualization, Writing – review & editing. PR: Writing – original draft, Conceptualization, Writing – review & editing. IS: Conceptualization, Writing – review & editing. LS-O: Writing – original draft, Conceptualization, Writing – review & editing. BT: Writing – original draft, Conceptualization, Writing – review & editing. SK: Writing – review & editing. MS: Writing – review & editing. CC: Funding acquisition, Supervision, Writing – review & editing. SP: Conceptualization, Writing – review & editing. GV: Conceptualization, Writing – review & editing. DH: Writing – original draft, Conceptualization, Writing – review & editing. CH: Conceptualization, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. PIMENTO CA20128 was supported by COST (European Cooperation in Science and Technology; www.cost.eu).
Acknowledgments
This article/publication is based upon work from COST Action PIMENTO CA20128, supported by COST (European Cooperation in Science and Technology).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The handling editor BY declared a past co-authorship with the authors EK, SK, MS, CC, GV, and CH.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Stevens, GA, Beal, T, Mbuya, MNN, Luo, H, Neufeld, LM, Addo, OY, et al. Micronutrient deficiencies among preschool-aged children and women of reproductive age worldwide: a pooled analysis of individual-level data from population-representative surveys. Lancet Glob Health. (2022) 10:e1590–9. doi: 10.1016/s2214-109x(22)00367-9
3. Passarelli, S, Free, CM, Shepon, A, Beal, T, Batis, C, and Golden, CD. Global estimation of dietary micronutrient inadequacies: a modelling analysis. Lancet Glob Health. (2024) 12:e1590–9. doi: 10.1016/S2214-109X(24)00276-6
4. WHO. National strategies for overcoming micronutrient malnutrition. Montreal, France: WHO (1991).
5. Ramakrishnan, U. Prevalence of micronutrient malnutrition worldwide. Nutr Rev. (2002) 60:S46–52. doi: 10.1301/00296640260130731
6. Sobczyńska-Malefora, A, Delvin, E, McCaddon, A, Ahmadi, KR, and Harrington, DJ. Vitamin B12 status in health and disease: a critical review. Diagnosis of deficiency and insufficiency – clinical and laboratory pitfalls. Crit Rev Clin Lab Sci. (2021) 58:399–429. doi: 10.1080/10408363.2021.1885339
7. Pasricha, S-R, Tye-Din, J, Muckenthaler, MU, and Swinkels, DW. Iron deficiency. Lancet. (2021) 397:233–48. doi: 10.1016/S0140-6736(20)32594-0
8. Hutkins, S. “Microbiology and Technology of Fermented Foods ” Oxford, Blackwell publishing. (2008)
9. Gänzle, M. The periodic table of fermented foods: limitations and opportunities. Appl Microbiol Biotechnol. Oxford, Blackwell publishing. (2022) 106:2815–26. doi: 10.1007/s00253-022-11909-y
10. Adesemoye, ET, Sanni, AI, Spano, G, Capozzi, V, and Fragasso, M. Lactic acid bacteria diversity in fermented foods as potential bio-resources contributing to alleviate malnutrition in developing countries: Nigeria as a case study. Fermentation. (2025) 11:103. doi: 10.3390/fermentation11020103
11. Bonaldo, F, and Leroy, F. Review: bacterially produced vitamin K2 and its potential to generate health benefits in humans. Trends Food Sci Technol. (2024) 147:104461. doi: 10.1016/j.tifs.2024.104461
12. Capozzi, V, Russo, P, Dueñas, MT, López, P, and Spano, G. Lactic acid bacteria producing B-group vitamins: a great potential for functional cereals products. Appl Microbiol Biotechnol. (2012) 96:1383–94. doi: 10.1007/s00253-012-4440-2
13. LeBlanc, JG, Laiño, JE, Valle, MJ, Vannini, V, Sinderen, D, Taranto, MP, et al. B-group vitamin production by lactic acid bacteria—current knowledge and potential applications: vitamin production by LAB. J Appl Microbiol. (2011) 111:1297–309. doi: 10.1111/j.1365-2672.2011.05157.x
14. Colinas, M, and Fitzpatrick, TB. Coenzymes and the primary and specialized metabolism interface. Curr Opin Plant Biol. (2022) 66:102170. doi: 10.1016/j.pbi.2021.102170
15. Blomhoff, R, and Blomhoff, HK. Overview of retinoid metabolism and function. J Neurobiol. (2006) 66:606–30. doi: 10.1002/neu.20242
16. Ross, AC. Modern nutrition in health and disease. 11th ed. Philadelphia: Wolters Kluwer Health/Lippincott Williams & Wilkins (2014). 1616 p.
17. Traber, MG, and Atkinson, J. Vitamin E, antioxidant and nothing more. Free Radic Biol Med. (2007) 43:4–15. doi: 10.1016/j.freeradbiomed.2007.03.024
18. Ross, AC. Modern nutrition in health and disease. LWW: Place of publication not identified (2012).
19. Jordá, T, and Puig, S. Regulation of ergosterol biosynthesis in Saccharomyces cerevisiae. Genes. (2020) 11:795. doi: 10.3390/genes11070795
20. Liang, M-H, Zhu, J, and Jiang, J-G. Carotenoids biosynthesis and cleavage related genes from bacteria to plants. Crit Rev Food Sci Nutr. (2018) 58:2314–33. doi: 10.1080/10408398.2017.1322552
21. Ye, Z, Shi, B, Huang, Y, Ma, T, Xiang, Z, Hu, B, et al. Revolution of vitamin E production by starting from microbial fermented farnesene to isophytol. Innovation. (2022) 3:100228. doi: 10.1016/j.xinn.2022.100228
22. Valentin, HE, and Qi, Q. Biotechnological production and application of vitamin E: current state and prospects. Appl Microbiol Biotechnol. (2005) 68:436–44. doi: 10.1007/s00253-005-0017-7
23. Bremus, C, Herrmann, U, Bringer-Meyer, S, and Sahm, H. The use of microorganisms in l-ascorbic acid production. J Biotechnol. (2006) 124:196–205. doi: 10.1016/j.jbiotec.2006.01.010
24. Averianova, LA, Balabanova, LA, Son, OM, Podvolotskaya, AB, and Tekutyeva, LA. Production of Vitamin B2 (riboflavin) by microorganisms: an overview. Front Bioeng Biotechnol. (2020) 8:570828. doi: 10.3389/fbioe.2020.570828
25. Olanbiwoninu, A, Greppi, A, Awotundun, T, Adebayo, EA, Spano, G, Mora, D, et al. Microbial-based biofortification to mitigate African micronutrients deficiency: a focus on plant-based fermentation as source of B-group vitamins. Food Biosci. (2023) 55:102996. doi: 10.1016/j.fbio.2023.102996
26. Dje Kouadio, DK, Wieringa, F, Greffeuille, V, and Humblot, C. Bacteria from the gut influence the host micronutrient status. Crit Rev Food Sci Nutr. (2024) 64:10714–29. doi: 10.1080/10408398.2023.2227888
27. Todorovic, S, Akpinar, A, Assunção, R, Bär, C, Bavaro, SL, Berkel Kasikci, M, et al. Health benefits and risks of fermented foods—the PIMENTO initiative. Front Nutr. (2024) 11:1458536. doi: 10.3389/fnut.2024.1458536
28. Muka, T, Glisic, M, Milic, J, Verhoog, S, Bohlius, J, Bramer, W, et al. A 24-step guide on how to design, conduct, and successfully publish a systematic review and meta-analysis in medical research. Eur J Epidemiol. (2020) 35:49–60. doi: 10.1007/s10654-019-00576-5
29. Bakker, M, Veldkamp, CLS, Van Assen, MALM, Crompvoets, EAV, Ong, HH, Nosek, BA, et al. Ensuring the quality and specificity of preregistrations. PLoS Biol. (2020) 18:e3000937. doi: 10.1371/journal.pbio.3000937
30. EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific and technical guidance for the preparation and presentation of a health claim application (revision 3)1. EFSA. (2021) 19:554. doi: 10.2903/j.efsa.2021.6554
31. Higgins, J, Thomas, J, Chandler, J, Cumpston, M, Li, T, and Page, M. Cochrane handbook for systematic reviews of interventions. (2022). Available online at: www.training.cochrane.org/handbook (Accessed December 06, 2025).
32. Kohl, C, McIntosh, EJ, Unger, S, Haddaway, NR, Kecke, S, Schiemann, J, et al. Online tools supporting the conduct and reporting of systematic reviews and systematic maps: a case study on CADIMA and review of existing tools. Environ Evid. (2018) 7:8. doi: 10.1186/s13750-018-0115-5
33. Elm, EV, Altman, DG, Egger, M, Pocock, SJ, Gøtzsche, PC, and Vandenbroucke, JP. Strengthening the reporting of observational studies in epidemiology (STROBE) statement: guidelines for reporting observational studies. BMJ. (2007) 335:806–8. doi: 10.1136/bmj.39335.541782.AD
34. Mladěnka, P, Macáková, K, Kujovská Krčmová, L, Javorská, L, Mrštná, K, Carazo, A, et al. Vitamin K – sources, physiological role, kinetics, deficiency, detection, therapeutic use, and toxicity. Nutr Rev. (2022) 80:677–98. doi: 10.1093/nutrit/nuab061
35. Yan, Q, Zhang, T, O’Connor, C, Barlow, JW, Walsh, J, Scalabrino, G, et al. The biological responses of vitamin K2: a comprehensive review. Food Sci Nutr. (2023) 11:1634–56. doi: 10.1002/fsn3.3213
36. Knez, E, Kadac-Czapska, K, and Grembecka, M. Effect of fermentation on the nutritional quality of the selected vegetables and legumes and their health effects. Life. (2023) 13:655. doi: 10.3390/life13030655
37. Booth, L, and Vitamin, S. Vitamin K: food composition and dietary intakes. Food Nutr Res. (2012) 56:5505. doi: 10.3402/fnr.v56i0.5505
38. Tarvainen, M, Fabritius, M, and Yang, B. Determination of vitamin K composition of fermented food. Food Chem. (2019) 275:515–22. doi: 10.1016/j.foodchem.2018.09.136
39. Fujita, Y, Iki, M, Tamaki, J, Kouda, K, Yura, A, Kadowaki, E, et al. Association between vitamin K intake from fermented soybeans, natto, and bone mineral density in elderly Japanese men: the Fujiwara-kyo osteoporosis risk in men (FORMEN) study. Osteoporos Int. (2012) 23:705–14. doi: 10.1007/s00198-011-1594-1
40. Rehn, U, Vogensen, FK, Persson, S-E, Hallin Saedén, K, Nilsson, BF, and Ardö, Y. Influence of microflora on texture and contents of amino acids, organic acids, and volatiles in semi-hard cheese made with DL-starter and propionibacteria. J Dairy Sci. (2011) 94:1098–111. doi: 10.3168/jds.2010-3146
41. Institute of Medicine (U.S.). DRI: Dietary reference intakes for vitamin A, vitamin K, arsenic, boron, chromium, copper, iodine, iron, manganese, molybdenum, nickel, silicon, vanadium, and zinc: A report of the panel on micronutrients. And the standing committee on the scientific evaluation of dietary reference intakes, food and nutrition board, Institute of Medicine. Washington, D.C: National Academy Press (2010). 773 p.
42. Shearer, MJ, and Newman, P. Metabolism and cell biology of vitamin K. Thromb Haemost. (2008) 100:530–47. doi: 10.1160/TH08-03-0147
43. Tie, J-K, and Stafford, DW. Functional Study of the Vitamin K Cycle Enzymes in Live Cells. Methods Enzymol. (2017) 584:349–94. doi: 10.1016/bs.mie.2016.10.015
44. Akbari, S, and Rasouli-Ghahroudi, AA. Vitamin K and bone metabolism: A review of the latest evidence in preclinical studies. Biomed Res Int. (2018) 2018:1–8. doi: 10.1155/2018/4629383
45. Theuwissen, E, Smit, E, and Vermeer, C. The role of Vitamin K in soft-tissue calcification. Adv Nutr. (2012) 3:166–73. doi: 10.3945/an.111.001628
46. Homma, K, Wakana, N, Suzuki, Y, Nukui, M, Daimatsu, T, Tanaka, E, et al. Treatment of natto, a fermented soybean preparation, to prevent excessive plasma Vitamin K concentrations in patients taking warfarin. J Nutr Sci Vitaminol. (2006) 52:297–301. doi: 10.3177/jnsv.52.297
47. Katsuyama, H, Ideguchi, S, Fukunaga, M, Fukunaga, T, Saijoh, K, and Sunami, S. Promotion of bone formation by fermented soybean (natto) intake in premenopausal women. J Nutri Sci Vitaminol. (2004) 50:114–20. doi: 10.3177/jnsv.50.114
48. Sakamoto, N, Nishiike, T, Iguchi, H, and Sakamoto, K. The effect of diet on blood Vitamin K status and urinary mineral excretion assessed by a food questionnaire. Nutr Health. (1999) 13:1–10. doi: 10.1177/026010609901300101
49. Schurgers, LJ, and Vermeer, C. Determination of phylloquinone and menaquinones in food. Pathophysiol Haemos Thromb. (2000) 30:298–307. doi: 10.1159/000054147
50. Tsukamoto, Y, Ichise, H, Kakuda, H, and Yamaguchi, M. Intake of fermented soybean (natto) increases circulating vitamin K 2 (menaquinone-7) and γ-carboxylated osteocalcin concentration in normal individuals. J Bone Miner Metab. (2000) 18:216–22. doi: 10.1007/s007740070023
51. Tsukamoto, Y, Ichise, H, and Yamaguchi, M. Prolonged intake of dietary fermented soybeans (natto) with the reinforced vitamin K2 (menaquinone-7) enhances circulating.GAMMA.-carboxylated osteocalcin concentration in normal individuals. J Health Sci. (2000) 46:317–21. doi: 10.1248/jhs.46.317
52. Lundberg, HE, Sundgot-Borgen, J, Mathisen, TF, Holo, H, Whist, AC, and Larsen, S. Cheese positively affects serum osteocalcin levels, bone turnover markers and bone mineral density in cross-country skiers: a dose-response study. Int J Clin Trials. (2024) 11:91–100. doi: 10.18203/2349-3259.ijct20240961
53. Lundberg, HE, Glasø, M, Chhura, R, Shukla, AA, Austlid, T, Sarwar, Z, et al. Effect on bone anabolic markers of daily cheese intake with and without vitamin K2: a randomised clinical trial. BMJ Nutr Prev Health. (2022) 5:182–90. doi: 10.1136/bmjnph-2022-000424
54. Walther, B, Guggisberg, D, Schmidt, RS, Portmann, R, Risse, M-C, Badertscher, R, et al. Quantitative analysis of menaquinones (vitamin K2) in various types of cheese from Switzerland. Int Dairy J. (2021) 112:104853. doi: 10.1016/j.idairyj.2020.104853
55. European Food Safety Authority (EFSA). Dietary reference values for nutrients summary report. EFS3. (2017) 14:121. doi: 10.2903/sp.efsa.2017.e15121
56. Brouwer, IA, Van Dusseldorp, M, West, CE, Meyboom, S, Thomas, CMG, Duran, M, et al. Dietary folate from vegetables and Citrus fruit decreases plasma homocysteine concentrations in humans in a dietary controlled trial. J Nutr. (1999) 129:1135–9. doi: 10.1093/jn/129.6.1135
57. Hariz, A, and Bhattacharya, PT. Megaloblastic Anemia. StatPearls. Treasure Island (FL): StatPearls Publishing (2025).
58. Instyitute of Medicine. Dietary reference intakes for Thiamin, riboflavin, niacin, Vitamin B6, folate, Vitamin B12, pantothenic acid, biotin, and choline. Washington: National Academies Press (2000). 1 p.
59. Mönch, S, Netzel, M, Netzel, G, Ott, U, Frank, T, and Rychlik, M. Pilot study on folate bioavailability from a camembert cheese reveals contradictory findings to recent results from a human short-term study. Front Nutr. (2016) 3:9. doi: 10.3389/fnut.2016.00009
60. Mönch, S, Netzel, M, Netzel, G, Ott, U, Frank, T, and Rychlik, M. Folate bioavailability from foods rich in folates assessed in a short term human study using stable isotope dilution assays. Food Funct. (2015) 6:241–7. doi: 10.1039/C4FO00658E
61. Forssén, KM, Jägerstad, MI, Wigertz, K, and Witthöft, CM. Folates and dairy products: A critical update. J Am Coll Nutr. (2000) 19:100S–10S. doi: 10.1080/07315724.2000.10718071
62. Chen, TS, Lui, CK, and Smith, CH. Folacin content of tea. J Am Diet Assoc. (1983) 82:627–32. doi: 10.1016/S0002-8223(21)00141-3
63. Jia, X, Ren, M, Zhang, Y, Ye, R, Zhang, L, and Li, Z. Association between tea drinking and plasma folate concentration among women aged 18–30 years in China. Public Health Nutr. (2021) 24:4929–36. doi: 10.1017/S1368980020004851
64. Singleton, C, and Martin, P. Molecular mechanisms of thiamine utilization. Curr Mol Med. (2001) 1:197–207. doi: 10.2174/1566524013363870
65. Dhir, S, Tarasenko, M, Napoli, E, and Giulivi, C. Neurological, psychiatric, and biochemical aspects of thiamine deficiency in children and adults. Front Psych. (2019) 10:207. doi: 10.3389/fpsyt.2019.00207
66. Teran, MDM, LeBlanc, M, and LeBlanc, JG. Thiamine-producing lactic acid bacteria and their potential use in the prevention of neurodegenerative diseases. Appl Microbiol Biotechnol. (2021) 105:2097–107. doi: 10.1007/s00253-021-11148-7
67. EFSA. Scientific opinion on the substantiation of health claims related to riboflavin (vitamin B2) and contribution to normal energy-yielding metabolism (ID 29, 35, 36, 42), contribution to normal metabolism of iron (ID 30, 37), maintenance of normal skin and mucous membranes (ID 31, 33), contribution to normal psychological functions (ID 32), maintenance of normal bone (ID 33), maintenance of normal teeth (ID 33), maintenance of normal hair (ID 33), maintenance of normal nails (ID 33), maintenance of normal vision (ID 39), maintenance of normal red blood cells (ID 40), reduction of tiredness and fatigue (ID 41), protection of DNA, proteins and lipids from oxidative damage (ID 207), and maintenance of the normal function of the nervous system (ID 213) pursuant to article 13(1) of regulation (EC) no 1924/2006. EFSA J. (2020) 8:1814. doi: 10.2903/j.efsa.2010.1814
68. Diez-Ozaeta, I, Berasarte, I, Zeid, AF, Fernández, M, Russo, P, López, P, et al. Functional characterization of the riboflavin-overproducing and dextran-producing Weissella cibaria BAL3C-5 C120T strain for the development of biofortified plant-based beverages. Int J Food Microbiol. (2025) 426:110908. doi: 10.1016/j.ijfoodmicro.2024.110908
69. Russo, P, Capozzi, V, Arena, MP, Spadaccino, G, Dueñas, MT, López, P, et al. Riboflavin-overproducing strains of Lactobacillus fermentum for riboflavin-enriched bread. Appl Microbiol Biotechnol. (2014) 98:3691–700. doi: 10.1007/s00253-013-5484-7
70. Spacova, I, Ahannach, S, Breynaert, A, Erreygers, I, Wittouck, S, Bron, PA, et al. Spontaneous riboflavin-overproducing Limosilactobacillus reuteri for biofortification of fermented foods. Front Nutr. (2022) 9:916607. doi: 10.3389/fnut.2022.916607
71. Yépez, A, Russo, P, Spano, G, Khomenko, I, Biasioli, F, Capozzi, V, et al. In situ riboflavin fortification of different kefir-like cereal-based beverages using selected Andean LAB strains. Food Microbiol. (2019) 77:61–8. doi: 10.1016/j.fm.2018.08.008
72. Gaby, SK, Bendich, A, Singh, VN, and Machlin, LJ. Vitamin intake and health: A scientific review. Boca Raton: CRC Press (1991).
73. Rad, EY, Saboori, S, Tammam, J, Thondre, PS, and Coe, S. The effect of niacin on inflammatory markers and adipokines: a systematic review and meta-analysis of interventional studies. Eur J Nutr. (2024) 63:2011–24. doi: 10.1007/s00394-024-03425-8
74. Wu, J. Association between dietary niacin intake and kidney stones in American adults. Sci Rep. (2025) 15:5666. doi: 10.1038/s41598-025-87227-w
75. Xu, J, Patassini, S, Begley, P, Church, S, Waldvogel, HJ, Faull, RLM, et al. Cerebral deficiency of vitamin B5 (d-pantothenic acid; pantothenate) as a potentially-reversible cause of neurodegeneration and dementia in sporadic Alzheimer’s disease. Biochem Biophys Res Commun. (2020) 527:676–81. doi: 10.1016/j.bbrc.2020.05.015
76. Contestabile, R, Di Salvo, ML, Bunik, V, Tramonti, A, and Vernì, F. The multifaceted role of vitamin B6 in cancer: Drosophila as a model system to investigate DNA damage. Open Biol. (2020) 10:200034. doi: 10.1098/rsob.200034
77. Hoegl, A, Nodwell, MB, Kirsch, VC, Bach, NC, Pfanzelt, M, Stahl, M, et al. Mining the cellular inventory of pyridoxal phosphate-dependent enzymes with functionalized cofactor mimics. Nat Chem. (2018) 10:1234–45. doi: 10.1038/s41557-018-0144-2
78. Xie, C, Coda, R, Chamlagain, B, Edelmann, M, Varmanen, P, Piironen, V, et al. Fermentation of cereal, pseudo-cereal and legume materials with Propionibacterium freudenreichii and Levilactobacillus brevis for vitamin B12 fortification. LWT. (2021) 137:110431. doi: 10.1016/j.lwt.2020.110431
79. Torres, AC, Vannini, V, Bonacina, J, Font, G, Saavedra, L, and Taranto, MP. Cobalamin production by Lactobacillus coryniformis: biochemical identification of the synthetized corrinoid and genomic analysis of the biosynthetic cluster. BMC Microbiol. (2016) 16:240. doi: 10.1186/s12866-016-0854-9
80. Varmanen, P, Deptula, P, Chamlagain, B, and Piironen, V. Letter to the editor on ‘enhancing vitamin B12 content in soy-yogurt by Lactobacillus reuteri, IJFM. 206:56–59.’. Int J Food Microbiol. (2016) 228:33. doi: 10.1016/j.ijfoodmicro.2016.03.029
81. Green, R, Allen, LH, Bjørke-Monsen, A-L, Brito, A, Guéant, J-L, Miller, JW, et al. Vitamin B12 deficiency. Nat Rev Dis Primers. (2017) 3:17040. doi: 10.1038/nrdp.2017.40
82. Li, KJ, Burton-Pimentel, KJ, Brouwer-Brolsma, EM, Blaser, C, Badertscher, R, Pimentel, G, et al. Identifying plasma and urinary biomarkers of fermented food intake and their associations with cardiometabolic health in a Dutch observational cohort. J Agric Food Chem. (2023) 71:4426–39. doi: 10.1021/acs.jafc.2c05669
83. Hruby, A, Dennis, C, and Jacques, PF. Dairy intake in 2 American adult cohorts associates with novel and known targeted and nontargeted circulating metabolites. J Nutr. (2020) 150:1272–83. doi: 10.1093/jn/nxaa021
84. Elmadfa, I, Heinzle, C, Majchrzak, D, and Foissy, H. Influence of a probiotic yoghurt on the status of vitamins B1, B2 and B6 in the healthy adult human. Ann Nutr Metab. (2001) 45:13–8. doi: 10.1159/000046700
85. Fabian, E, Majchrzak, D, Dieminger, B, Meyer, E, and Elmadfa, I. Influence of probiotic and conventional yoghurt on the status of vitamins B1, B2 and B6 in Young healthy women. Ann Nutr Metab. (2008) 52:29–36. doi: 10.1159/000114408
86. LeBlanc, JG, Chain, F, Martín, R, Bermúdez-Humarán, LG, Courau, S, and Langella, P. Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb Cell Factories. (2017) 16:79. doi: 10.1186/s12934-017-0691-z
87. Urgert, R, Van Vliet, T, Zock, PL, and Katan, MB. Heavy coffee consumption and plasma homocysteine: a randomized controlled trial in healthy volunteers. Am J Clin Nutr. (2000) 72:1107–10. doi: 10.1093/ajcn/72.5.1107
88. Hornero-Méndez, D, Cerrillo, I, Ortega, Á, Rodríguez-Griñolo, M-R, Escudero-López, B, Martín, F, et al. β-Cryptoxanthin is more bioavailable in humans from fermented orange juice than from orange juice. Food Chem. (2018) 262:215–20. doi: 10.1016/j.foodchem.2018.04.083
89. Shahar, DR, Schwarzfuchs, D, Fraser, D, Vardi, H, Thiery, J, Fiedler, GM, et al. Dairy calcium intake, serum vitamin D, and successful weight loss. Am J Clin Nutr. (2010) 92:1017–22. doi: 10.3945/ajcn.2010.29355
90. Thams, L, Stounbjerg, NG, Hvid, LG, Mølgaard, C, Hansen, M, and Damsgaard, CT. Effects of high dairy protein intake and vitamin D supplementation on body composition and cardiometabolic markers in 6–8-y-old children—the D-pro trial. Am J Clin Nutr. (2022) 115:1080–91. doi: 10.1093/ajcn/nqab424
91. Ashagrie, H, Baye, K, Guibert, B, Seyoum, Y, Rochette, I, and Humblot, C. Cereal-based fermented foods as a source of folate and cobalamin: the role of endogenous microbiota. Food Res Int. (2023) 174:113625. doi: 10.1016/j.foodres.2023.113625
92. Bationo, F, Humblot, C, Songré-Ouattara, LT, Hama-Ba, F, Le Merrer, M, Chapron, M, et al. Total folate in West African cereal-based fermented foods: bioaccessibility and influence of processing. J Food Compos Anal. (2020) 85:103309. doi: 10.1016/j.jfca.2019.103309
93. Barone, M, D’Amico, F, Brigidi, P, and Turroni, S. Gut microbiome–micronutrient interaction: the key to controlling the bioavailability of minerals and vitamins? Biofactors. (2022) 48:307–14. doi: 10.1002/biof.1835
94. Voland, L, Le Roy, T, Debédat, J, and Clément, K. Gut microbiota and vitamin status in persons with obesity: A key interplay. Obes Rev. (2022) 23:e13377. doi: 10.1111/obr.13377
95. Groenen-Van Dooren, MMCL, Ronden, JE, Soute, BAM, and Vermeer, C. Bioavailability of phylloquinone and menaquinones after oral and colorectal administration in vitamin K-deficient rats. Biochem Pharmacol. (1995) 50:797–801. doi: 10.1016/0006-2952(95)00202-B
96. Kemp, JA, Alvarenga, L, Cardozo, LFMF, Dai, L, Stenvinkel, P, Shiels, PG, et al. Dysbiosis in patients with chronic kidney disease: let us talk about Vitamin K. Curr Nutr Rep. (2022) 11:765–79. doi: 10.1007/s13668-022-00438-9
97. Frank, RAW, Leeper, FJ, and Luisi, BF. Structure, mechanism and catalytic duality of thiamine-dependent enzymes. Cell Mol Life Sci. (2007) 64:892–905. doi: 10.1007/s00018-007-6423-5
98. LeBlanc, JG, Milani, C, De Giori, GS, Sesma, F, Van Sinderen, D, and Ventura, M. Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Curr Opin Biotechnol. (2013) 24:160–8. doi: 10.1016/j.copbio.2012.08.005
99. Thakur, K, Tomar, SK, and De, S. Lactic acid bacteria as a cell factory for riboflavin production. Microb Biotechnol. (2016) 9:441–51. doi: 10.1111/1751-7915.12335
100. Magnúsdóttir, S, Ravcheev, D, De Crécy-Lagard, V, and Thiele, I. Systematic genome assessment of B-vitamin biosynthesis suggests co-operation among gut microbes. Front Genet. (2015) 6:148. doi: 10.3389/fgene.2015.00148
101. Zhou, J, Yu, X, Liu, J, Qin, W, He, Z, Stahl, D, et al. VB12 path for accurate metagenomic profiling of microbially driven cobalamin synthesis pathways. mSystems. (2021) 6:e00497-21. doi: 10.1128/mSystems.00497-21
102. Degnan, PH, Taga, ME, and Goodman, AL. Vitamin B 12 as a modulator of gut microbial ecology. Cell Metab. (2014) 20:769–78. doi: 10.1016/j.cmet.2014.10.002
103. Gurwara, S, Ajami, NJ, Jang, A, Hessel, FC, Chen, L, Plew, S, et al. Dietary nutrients involved in one-carbon metabolism and colonic mucosa-associated gut microbiome in individuals with an endoscopically Normal Colon. Nutrients. (2019) 11:613. doi: 10.3390/nu11030613
104. Bernabeu, M, Gharibzahedi, SMT, Ganaie, AA, Macha, MA, Dar, BN, Castagnini, JM, et al. The potential modulation of gut microbiota and oxidative stress by dietary carotenoid pigments. Crit Rev Food Sci Nutr. (2024) 64:12555–73. doi: 10.1080/10408398.2023.2254383
105. Park, J, Hosomi, K, Kawashima, H, Chen, Y-A, Mohsen, A, Ohno, H, et al. Dietary Vitamin B1 intake influences gut microbial community and the consequent production of short-Chain fatty acids. Nutrients. (2022) 14:2078. doi: 10.3390/nu14102078
106. Bourgonje, AR. Microbial riboflavin biosynthesis associates with response to dietary Fiber consumption: time to personalize adjunct therapy in patients with inflammatory bowel disease? Gastroenterology. (2023) 165:515–6. doi: 10.1053/j.gastro.2022.10.016
107. Aufreiter, S, Kim, JH, and O’Connor, DL. Dietary oligosaccharides increase colonic weight and the amount but not concentration of bacterially synthesized folate in the colon of piglets. J Nutr. (2011) 141:366–72. doi: 10.3945/jn.110.135343
108. Sepehr, E, Peace, RW, Jee, P, Lampi, BJ, Brooks, SPJ, and Storey, KB. Folate derived from Cecal bacterial fermentation does not increase liver folate stores in 28-d folate-depleted male Sprague-Dawley rats. J Nutr. (2003) 133:1347–54. doi: 10.1093/jn/133.5.1347
109. Soto-Martin, EC, Warnke, I, Farquharson, FM, Christodoulou, M, Horgan, G, Derrien, M, et al. Vitamin biosynthesis by human gut butyrate-producing bacteria and cross-feeding in synthetic microbial communities. mBio. (2020) 11:e00886-20. doi: 10.1128/mBio.00886-20
110. Degnan, PH, Barry, NA, Mok, KC, Taga, ME, and Goodman, AL. Human gut microbes use multiple transporters to distinguish Vitamin B12 Analogs and compete in the gut. Cell Host Microbe. (2014) 15:47–57. doi: 10.1016/j.chom.2013.12.007
111. Andersen, CT, Tadesse, AW, Bromage, S, Fekadu, H, Hemler, EC, Passarelli, S, et al. Anemia Etiology in Ethiopia: assessment of nutritional, infectious disease, and other risk factors in a population-based cross-sectional survey of women, men, and children. J Nutr. (2022) 152:501–12. doi: 10.1093/jn/nxab366
112. Verger, EO, Fortin, S, Tamene, A, Ashagrie, H, Mouquet-Rivier, C, and Humblot, C. Ignoring the impact of fermentation could result in substantial misestimation of folate and cobalamin adequacy: A simulation study on injera consumption in the Ethiopian context. Curr Dev Nutr. (2025) 9:104581. doi: 10.1016/j.cdnut.2025.104581
113. Ashagrie, H, Baye, K, Guibert, B, Rochette, I, Tisseyre, P, and Humblot, C. The use of propionic and lactic acid bacteria to produce cobalamin and folate in injera, an Ethiopian cereal-based fermented food. Int J Food Microbiol. (2025) 426:110909. doi: 10.1016/j.ijfoodmicro.2024.110909
Keywords: B vitamins, fermented foods, human intervention studies, microbial vitamin formation, vitamin bioavailability, vitamin K2
Citation: Keyvan E, Adesemoye E, Champomier-Vergès M-C, Chanséaume-Bussiere E, Mardon J, Nikolovska Nedelkoska D, Palamutoglu R, Russo P, Sarand I, Songre-Ouattara L, Trajkovska B, Karakaya S, Syrpas M, Chassard C, Pracer S, Vergères G, Heine D and Humblot C (2025) Vitamins formed by microorganisms in fermented foods: effects on human vitamin status—a systematic narrative review. Front. Nutr. 12:1653666. doi: 10.3389/fnut.2025.1653666
Edited by:
Birsen Yilmaz, Çukurova University, TürkiyeReviewed by:
Tevfik Koçak, Gumushane University, TürkiyeSavio Fernandes, Lusofona University, Portugal
Copyright © 2025 Keyvan, Adesemoye, Champomier-Vergès, Chanséaume-Bussiere, Mardon, Nikolovska Nedelkoska, Palamutoglu, Russo, Sarand, Songre-Ouattara, Trajkovska, Karakaya, Syrpas, Chassard, Pracer, Vergères, Heine and Humblot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Erhan Keyvan, ZXJoYW5rZXl2YW5AbWVobWV0YWtpZi5lZHUudHI= Christèle Humblot, Y2hyaXN0ZWxlLmh1bWJsb3RAaXJkLmZy