 Xuyang Pu1,2
Xuyang Pu1,2 Xueqiang Wu1,2,3,4*
Xueqiang Wu1,2,3,4*- 1Affiliated Meizhou Hospital of Shantou University Medical College, Shantou University, Meizhou, China
- 2Institute of Basic Medical Sciences, Meizhou People’s Hospital, Meizhou, China
- 3Guangdong Engineering Technological Research Center of Clinical Molecular Diagnosis and Antibody Drugs, Meizhou Academy of Medical Sciences, Meizhou, China
- 4Breast Center, MeiZhou People’s Hospital, Meizhou, China
A fundamental characteristic of gene mutations is the permanent alteration of the DNA sequence, including point mutations, deletions, inversions, and translocations. Among these, DNA point mutation detection has consistently remained a central focus of research across multiple disciplines due to its close association with a range of diseases, such as sickle cell anemia and β-thalassemia. However, the typically low abundance of such mutations presents a significant technical challenge. Due to technical limitations in detection sensitivity, increasing research efforts have been directed toward nucleic acid probe-based strategies to enhance the efficiency and accuracy of point mutation identification. This review summarizes the developments in nucleic acid probe-based techniques for detecting gene point mutations, with an emphasis on strategies involving pure nucleic acid probes as well as the synergistic use of enzymes, nucleic acid analogs, and nanotechnology. The principles, advantages, and limitations of the above technologies are also described and summarized. In addition, we also explored the application of AI technology in nucleic acid probes and the potential future challenges.
1 Introduction
Point mutation refers to an alteration that occurs at a single nucleotide of a DNA molecule, characterized by the substitution, insertion, or deletion of only one nucleotide in the DNA sequence. Mutated genes typically trigger a cascade of detectable biological alterations, including changes in protein expression and function, changes in RNA expression, epigenetic modifications (e.g., DNA methylation, histone modifications), and variations in metabolite profiles (Eriksson et al., 2017; Ho et al., 2008). Numerous genetic disorders and cancers originate from gene mutations that cause dysregulation of functional proteins. Among the most frequently mutated genes, EGFR, TP53, and KRAS are associated with diverse malignancies including non-small cell lung cancer (NSCLC) (Bai et al., 2024; Mao et al., 2021), colorectal carcinoma (Adorisio et al., 2025; Kim et al., 2020), and pancreatic cancer (Hashimoto et al., 2016; McCubrey et al., 2022). Detection of point mutations facilitates the early diagnosis of genetic diseases and the screening of carriers, while also contributing to the molecular subtyping of tumor-related diseases and guiding precision medicine. In addition, resistance in bacteria, viruses, and parasites is often mediated by point mutations (Carter et al., 2015; Menard and Dondorp, 2017), and identifying these mutations is valuable for clinical treatment and epidemiological prevention and control. Nevertheless, this process confronts significant technical challenges, notably the low concentration level of the target and interference from complex sample matrices.
The introduction of Sanger sequencing in 1977 revolutionized the field of genetic testing (Sanger et al., 1977). Nevertheless, Sanger sequencing has many limitations, such as a time-consuming process and high cost, which make it difficult to meet the demands for large-scale testing. Next-Generation Sequencing (NGS) has dramatically increased throughput through massive parallel sequencing, but it also has some drawbacks, such as shorter read lengths and higher requirements for sample and data analysis. Third-Generation Sequencing (TGS) overcomes some bottlenecks of NGS due to its ultra-long read length and non-reliance on PCR amplification (Van Dijk et al., 2018). However, TGS is still in the stage of rapid development, and its detection cost and high error rate need to be further optimized.
In recent years, nucleic acid probe-based mutation detection technologies have made significant progress. Nucleic acid probes play a central role in distinguishing perfectly matched sequences from single-base mismatches. Some research teams have developed pure nucleic acid probes that distinguish mismatches based on kinetic differences. However, the thermodynamic and kinetic differences caused by a single nucleotide variation are often subtle, requiring additional strategies to improve specificity and sensitivity. From a mechanistic perspective, existing approaches can be broadly divided into three categories. The first is enzyme-assisted methods, which exploit the high selectivity of ligases and exonucleases toward base pairing states, thereby achieving mismatch discrimination and signal amplification through catalytic reactions. The second is nanoparticle-based methods, which take advantage of the unique optical, electrical, or surface properties of nanomaterials to enable signal transduction and amplification. The third is nucleic acid analogue-based methods, in which chemically modified probes such as peptide nucleic acids (PNA) are employed to modulate hybridization stability and enhance single-base discrimination at the molecular level (Figure 1).
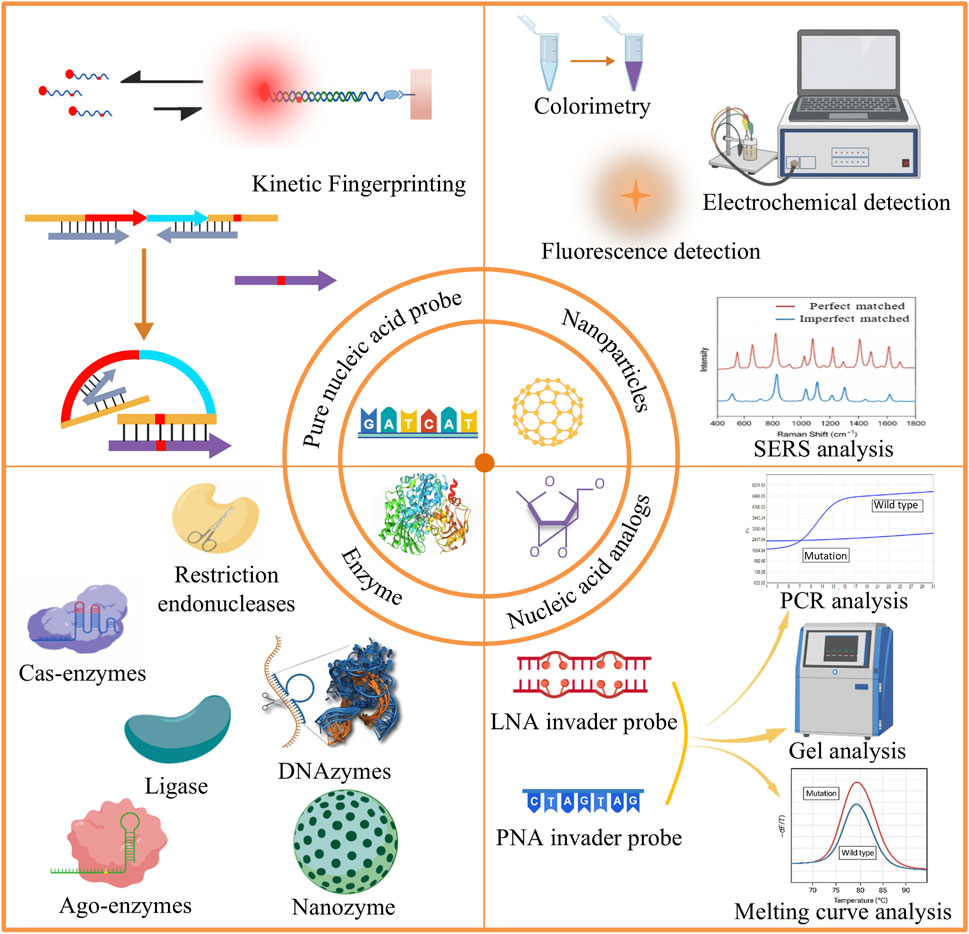
Figure 1. Schematic overview of different strategies for detecting gene point mutations: (1) Direct sequencing; (2) Nanoparticle-based methods; (3) Enzyme-based methods; (4) Nucleic acid analog-based methods.
In this review, we will start with the above technologies and systematically summarize the progress of nucleic acid probe-based point mutation detection methods, analyze the advantages and disadvantages of different technologies, as well as explore the potential directions for optimization and development.
2 Enzyme-mediated point mutation detection
Enzymes are widely available biocatalysts in biological systems, and their catalytic activity and high specificity make them valuable tools for gene mutation detection. Enzyme-based mutation detection methods typically require no complex instrumentation and are suitable for high-throughput and on-site detection. However, enzymes also present some inherent limitations. They are often highly dependent on specific sequences and may exhibit insufficient sensitivity when detecting rare mutations in complex samples. Nevertheless, with the development of enzyme engineering technology, enzyme-mediated nucleic acid probe-based mutation detection continues to hold significant promise.
2.1 Guided enzymes
Restriction endonucleases (REs) typically enable mutation identification and screening by directly recognizing specific nucleic acid sequences and cleaving wild-type sequences. The sequence with point mutations cannot be cleaved by RE, it can be normally amplified and enriched by PCR for subsequent detection (Evans et al., 2018), as shown in Figure 2A. However, the conventional RE-based methods suffer from limited sensitivity and require complex designs to enable the detection of low-abundance mutations (Smith et al., 2020). RNA-guided nucleases are a class of enzymes that can bind to specific RNA molecules, recognize and act on target nucleic acids. The principle is that specifically designed RNA probe molecules serve as guides to precisely direct enzymes to recognize and cleave or degrade target DNA or RNA sequences. The most representative of these enzymes are Cas enzymes in the CRISPR-Cas system and the Argonaute (Ago) enzymes in the RNA interference system.
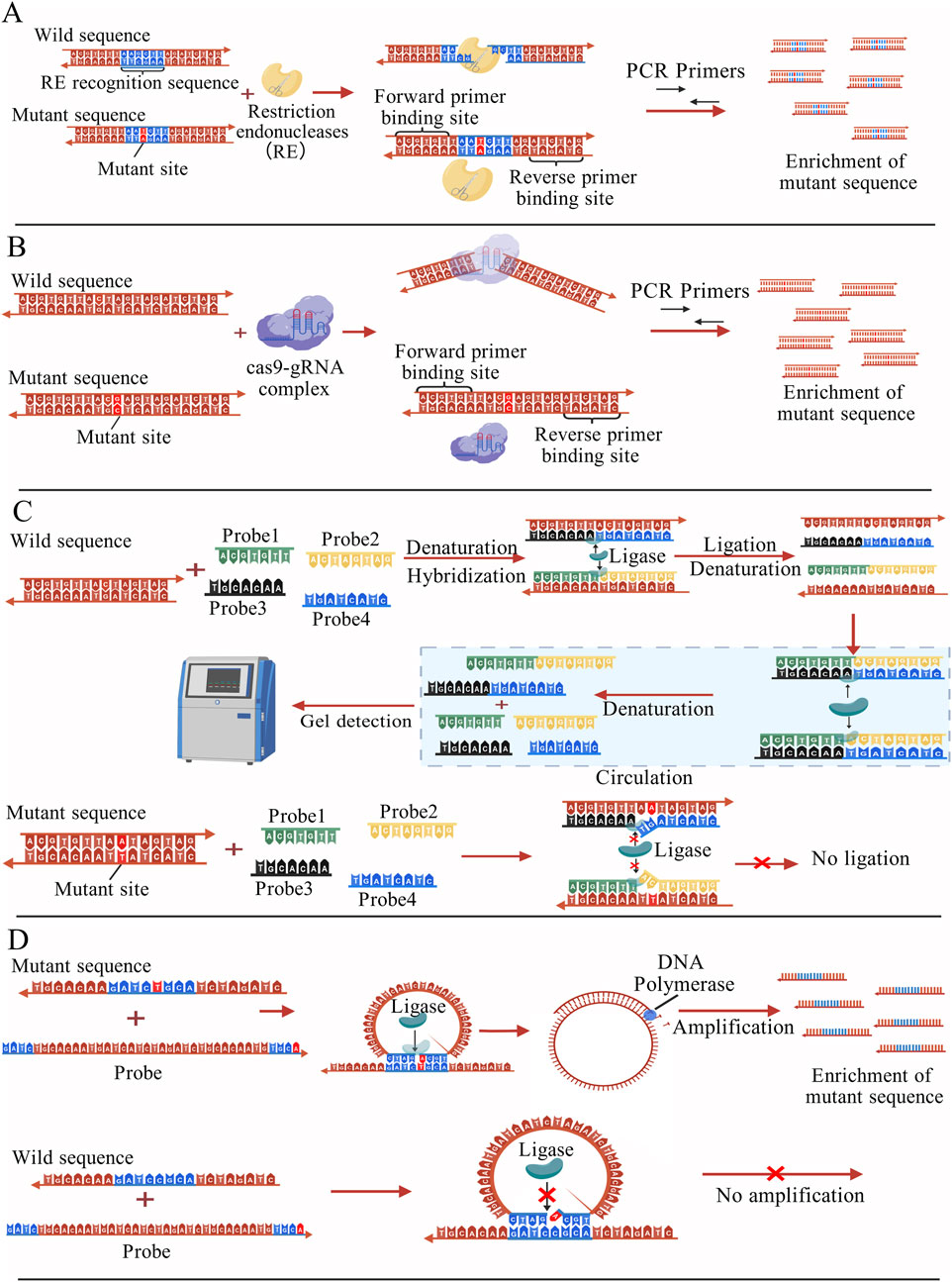
Figure 2. (A) Nucleic acid fragments with specific sequences are cleaved by REs and cannot be amplified by PCR, whereas fragments carrying mutation sites are not cleaved and can be selectively amplified and enriched for subsequent detection (Evans et al., 2018); (B) Under the guidance of sgRNA, the Cas9 protein cleaves the wild-type sequence, while the mutant sequence remains intact and can undergo normal PCR amplification, enrichment, and subsequent detection (Bae et al., 2019); (C) Two pairs of probes can only hybridize with perfectly matched sequences, and under the action of ligase, they are joined to form two new complementary nucleic acid strands. Through cyclic reactions, more probe pairs are ligated into new strands, which can then be analyzed by gel detection. When a point mutation is present, the probes cannot pair correctly, leading to ligation failure (Ruiz et al., 2020); (D) The probe perfectly matches the mutant sequence and is ligated into a new circular nucleic acid, which is then subjected to RCA amplification. The probe cannot fully match the wild-type sequence, resulting in ligation failure. (Ma and Gao, 2018); The figures are created with BioGDP.com (Jiang et al., 2025).
2.1.1 Cas-enzymes-based detection
Compared with conventional methods, single-stranded guide RNAs (sgRNAs) based on PAM sequences can significantly improve the editing efficiency of mutant alleles, with a relative specificity capable of reaching 15-fold (Bae et al., 2019). One such method is DASH (depletion of abundant sequences by hybridization), which increases detection sensitivity by designing specific sgRNAs and mediating Cas9 protein cleavage. Guided by the sgRNA, the Cas protein cleaves and removes the unwanted wild-type sequences, thereby allowing the mutant sequences to be preserved and subsequently amplified by PCR for detection (Figure 2B). Because the mutation site is altered to cause loss of the PAM site (NGG), Cas9 cannot cleave sequences containing this mutation, which enables the removal of wild sequences (Gu et al., 2016). The CRISPR/Cas9 system can also be used in conjunction with technologies such as Amplified Fragment Length Polymorphism Analysis (AFLPA) and Sanger sequencing for the enrichment of mutant sequences. When used in conjunction with PCR, it can achieve a sensitivity of 0.1% (Jia et al., 2018).
Although CRISPR/Cas9 is a powerful tool, it has many shortcomings such as operational complexity and the limitation of relying on specific PAM sites. The use of Cas9 variants with relaxed PAM sequence requirements, such as xCas9, Cas9-NG can effectively address this issue, but at the cost of reduced DNA targeting and cleavage capabilities (Legut et al., 2020). In addition, off-target effects are one of the most urgent problems to be solved using CRISPR/Cas9 (Guo et al., 2023; Kim et al., 2015; Kimberland et al., 2018). Even for highly specific sgRNAs, Cas9 can generate cuts in DNA sequences similar to the target sequence, triggering unwanted gene mutations or expression. Some software has been developed to predict the off-target effects of CRISPR/Cas9 using machine learning methods, but the accuracy of the assay still needs to be improved (Fan and Xu, 2021).
2.1.2 Ago-enzymes-based detection
The Ago family is a group of proteins widely involved in the regulation of gene expression, especially in RNA interference and microRNA-mediated gene silencing. The Ago proteins inhibit the expression of specific genes by binding to small RNA molecules, and subsequently recognizing and cleaving target mRNA. Different Ago family members can be categorized into different types according to their function. Such as PfAgo, TtAgo, and HsAgo2. Ago proteins from different species have different thermal stabilities, catalytic activities, and gene-editing potentials.
The Pyrococcus furiosus Argonaute (PfAgo) protein was used to propose a novel detection technique called PfAgo-mediated Nucleic Acid Detection (PAND) (He et al., 2019). The PfAgo protein can specifically recognize and cleave target DNA under the guidance of nucleic acid molecules, simultaneously generating short single-stranded DNA (ssDNA). The produced ssDNA is then able to bind to the corresponding fluorescent probes and guide the PfAgo protein for a second round of cleavage. The presence of the target DNA can be confirmed by detecting changes in fluorescence. Notably, by designing multiple pairs of guide sequences and fluorescent probes simultaneously, the PAND system enables simultaneous detection of multiple mutations in a single experiment. Experimental results demonstrate that PAND can successfully detect single nucleotide polymorphisms (SNPs) in the BRCA1 gene, as well as KRAS G12D and EGFR T790M mutations, with a low background noise level of 0.1%.
Ago proteins can also be integrated with various materials and molecules to enhance their functionality. A current example is the graphene field effect transistor (GFET) platform mediated by Ago (Kong et al., 2024). Ago proteins provided nanoscale binding channels, allowing the organization and directed formation of DNA probes that rapidly recognize target mutations. GFET is electrochemical sensor that allows real-time monitoring of electrical changes resulting from binding of DNA probe to target sequences. An array comprising multiple independent Ago-GFET sensing channels can be integrated on a single microchip via photolithography, enabling simultaneous modular and multiplex detection of tumor-associated miRNAs, circulating tumor DNA, and viral RNA from a large number of biological samples. Furthermore, by leveraging the cleavage and guiding properties of TtAgo, NAVIGATER (Nucleic Acid enrichment Via DNA Guided Argonaute from Thermus thermophilus) was demonstrated by Song et al. (2020). First, an excess of short DNA guide strands fully complementary to the wild-type sequence are designed and pre-bound with TtAgo to form a complex. This complex precisely cleaves perfectly matched wild-type DNA and RNA, while Mutant nucleic acid sequences remain intact. Following downstream Xenonucleic Acid PCR (XNA-PCR), NAVIGATOR enables ultra-sensitive detection of mutations as low as 0.01% in pancreatic cancer blood samples. More importantly, the system allows for the design of multiple DNA guide strands targeting different mutation sites, enabling simultaneous detection of multiple mutations in a single reaction system.
2.1.3 Limitations and optimization of guided enzymes
Although guided enzymes have been widely applied in gene detection and gene editing, there are still many limitations. One major challenge lies in improving delivery efficiency (Finn et al., 2018) and it is hoped that new viral vectors or non-viral delivery systems can be developed in the future to optimize this process (Sun et al., 2025; Tabebordbar et al., 2016). Moreover, the presence of pre-existing antibodies and T-cell immunity against bacterially derived Cas proteins in humans poses the risk of undesired immune responses to therapeutic vectors (Shen et al., 2022). With the continued maturation of protein engineering strategies, these immunological barriers may be progressively mitigated, and potentially even eliminated (Crudele and Chamberlain, 2018).
2.2 Ligase
2.2.1 Ligase chain reaction (LCR)
As a classical nucleic acid amplification technique, ligase chain reaction (LCR) has been used for a variety of gene mutation detection methods (Ruiz et al., 2020). The core principle is to design two pairs of oligonucleotide probes, each pair of probes targeting two complementary strands of mutated DNA. When the thermally denatured ssDNA hybridizes correctly with the probes, the ligase catalyzed the formation of phosphodiester bonds adjacent to the probes, and the two pairs of probes were ligated into two complete DNA strands. The number of intact probe strands increased exponentially through subsequent amplification cycles. Conversely, the probes do not completely hybridize with wild-type DNA sequences. The principle of LCR technology is illustrated in the schematic diagram in Figure 2C. In practical experiments, LCR is typically used in conjunction with other signal readout techniques (Liu Z.-J. et al., 2020).
2.2.2 Combining LCR with fluorescence
The combination of ligases and fluorescence is a promising strategy for detecting nucleic acid mutations. Sawamura et al. reported a method based on ligase-mediated branching hybridization chain reaction (LDR) combined with fluorescence quenching technology for the rapid detection of point mutations in mixed samples. This method detects mutations through the use of both differentiated and universal primers, enables specific detection within PCR amplification products, facilitates the formation of MBs via primer ligation reactions, and achieves fluorescence quenching via Förster Resonance Energy Transfer (FRET). Experimental results have demonstrated that this approach is capable of detecting mutant DNA at levels as low as 5% (Sawamura and Hashimoto, 2017). In order to increase the detection limit, a branching hybridisation chain reaction has been applied to ligase mutation detection, referred to as the ligation-mediated branching hybridisation chain reaction (ligation-bHCR). This method combines the ligase discriminatory ability and the signal amplification function of bHCR that is capable of resolving single-nucleotide variants in mRNAs at single-molecule resolution (Tang et al., 2017).
2.2.3 Combining ligase with RCA
Rolling Circle Amplification (RCA) is a common nucleic acid amplification technique that generates large numbers of DNA copies using circular DNA templates and enzyme-driven amplification reactions. The combination of RCA and ligase has been widely used for point mutation detection (Ma and Gao, 2018; Wu et al., 2010; Zou et al., 2014). This approach relies on the use of specially designed primer probes to guide the amplification of a circular DNA template. When the target DNA contains a mutation, the probe perfectly matches it, and the ligase catalyzes the formation of a phosphodiester bond. The probe is ligated into a new circular nucleic acid, which serves as a template for subsequent RCA amplification. In contrast, the probe cannot fully match the wild-type sequence and therefore cannot be efficiently ligated. The principle of DNA ligase-assisted RCA is shown in Figure 2D. Furthermore, RCA can also be combined with other molecules, such as nucleases and nanoparticles, to further enhance the detection sensitivity (Li et al., 2010; Shi et al., 2022).
As important tools in nucleic acid research, both LCR and RCA rely heavily on the catalytic function of ligase. It has been shown that the sensitivity of the assay can be further improved by combining the two research methods (Cheng et al., 2013). However, the complexity of ligase-based experimental procedures demands significant technical skills.
2.2.4 Limitations and optimization of ligases
Theoretically, DNA ligases catalyze ligation events exclusively in the presence of perfect base pairing. However, the complexity of ligase-based experimental procedures demands significant technical skills (Wang and Zhang, 2014; Xu et al., 2016; Zhang et al., 2015). Furthermore, the most commonly used T4 DNA ligase is unstable at high temperatures and generally exhibits lower catalytic efficiency than DNA polymerases, resulting in an overall slower reaction rate compared with PCR (Lohman et al., 2014). These limitations may be addressed through advances in protein engineering. In addition, the continued exploration of thermophilic and extremophilic microorganisms is expected to facilitate the discovery and development of novel ligases (Tanabe et al., 2015).
2.3 DNAzyme
To overcome the limitations of traditional enzymes, such as poor stability, stringent storage requirements, and strict experimental conditions, increasing numbers of research groups have been dedicated to finding alternatives to conventional enzymes in recent years. DNAzymes represent a promising option. Unlike traditional protein enzymes, DNAzymes are obtained through in vitro selection methods (SELEX) and are composed of ssDNA molecules. Compared with natural protein enzymes, DNAzymes exhibit greater tolerance and stability (Li et al., 2025). They have already been widely applied in point mutation detection, where rational design allows them to be activated only under fully complementary conditions, thereby enabling highly specific recognition of single-base mismatches (Song et al., 2012).
2.3.1 DNAzymes-based detection
DNAzyme-based mutation detection is generally integrated with other techniques or molecular components (Zhang et al., 2021), rather than relying solely on the DNAzyme itself, in order to achieve enhanced sensitivity and reliability in experimental results (Wang H.-Q. et al., 2011). To date, most methods developed involve complex probe labeling. Li et al. introduced a G-quadruplex DNAzyme into the Gap-LCR technique to achieve a detection strategy without chemically modified probes (Zhou et al., 2013). After Gap-LCR amplification, the DNAzyme exhibited peroxidase-like catalytic activity in the presence of hemin, catalyzing substrates (ABTS, tyramine) to produce colorimetric or fluorescent signals, thereby converting the presence of mutant DNA into visible or measurable outputs. Experimental results showed that this method was able to detect mutations in the presence of a 105-fold excess of wild-type DNA.
Although detection based on G-quadruplex DNAzymes has many advantages over traditional enzymatic methods, the low catalytic activity of DNAzymes also limits their widespread application. Therefore, how to amplify DNAzymes has become a hotspot in analytical research. Xu et al. chose to amplify DNAzymes and enhance signals via target-catalyzed hairpin assembly (CHA), developing an enzyme-free and label-free DNAzyme sensor for SNP genotyping. This sensor successfully detected A-G single-nucleotide mutations on human chromosomes associated with Alzheimer’s disease in real saliva samples (Xu et al., 2017). Similarly, Li et al. adopted a comparable strategy, using flap endonuclease 1 (FEN1)-invader-triggered CHA to induce DNAzyme amplification, and developed a fluorescence amplification strategy based on multi-DNAzyme-linked structures. This method was successfully applied to the detection of point mutations in the KARS gene, achieving a detection limit as low as 4.23 fM (Li et al., 2022). Moreover, it performed excellently in complex serum samples, providing a new approach for isothermal and highly sensitive SNP detection.
2.3.2 Limitations and future optimization of DNAzymes
Although DNAzymes possess multiple advantages compared with traditional enzymes, their catalytic activity still requires improvement (Wang et al., 2021). Most DNAzymes depend on specific metal ions (such as Mg2+ or Pb2+) as cofactors, and their activity may be reduced in the ionic environments of complex biological samples (Rosenbach et al., 2020). In addition, DNAzyme kinetic simulation experiments still show certain limitations in detecting DNA point mutations (Kenward and Dorfman, 2009). In the future, high-throughput SELEX and machine learning-assisted optimization of selection processes are expected to yield more efficient and specific DNAzymes. Similarly, DNAzymes are anticipated to be combined with tools such as MB and CRISPR-Cas systems to construct new composite detection platforms.
2.4 Nanozyme
Nanozymes refer to nanomaterials that possess catalytic activities similar to those of natural enzymes. Unlike natural enzymes or artificially recombined enzymes, nanozymes are nanoparticles or nanostructures with unique surface architectures and catalytic sites (Liang and Yan, 2019), but they can mimic the functions of natural enzymes such as peroxidases and oxidases (Hamed et al., 2024; Wang X. et al., 2023). Nanozymes exhibit several advantages, including resistance to inactivation, high thermal stability, protease tolerance, and low cost, making them excellent alternatives to traditional enzymes (Zandieh and Liu, 2024).
2.4.1 Nanozyme-based detection
Through π-π stacking interactions, Guo et al. synthesized hemin-graphene hybrid nanosheets (H-GNs). Due to the presence of hemin, the H-GNs possess intrinsic peroxidase-like activity. In addition, H-GNs exhibit distinct affinities toward ssDNA and dsDNA. Based on these properties, the researchers developed a label-free colorimetric detection system for identifying single-base mutations in related DNA. Probe DNA adsorbs onto the H-GNs in single-stranded form and detaches only in the presence of fully complementary target DNA. Utilizing the catalytic activity of H-GNs, the TMB-H2O2 reaction is catalyzed to produce a color change indicating the presence or absence of a mutation. This detection system has been successfully applied for SNP detection in the HBV virus gene (Guo et al., 2011). Nanozymes composed of mesoporous silica and platinum nanoparticles (MS-PtNPs) synthesized by Chen et al. also exhibit peroxidase-like activity, which can be controlled by loading ssDNA. The adsorption of excessive DNA probes on MS blocks the nucleation sites of PtNP. Resulting in decreased peroxidase activity. Activity was restored when complementary target DNA was introduced. They combined it with a 3,3,5,5-tetramethylbenzidine/H2O2 color system to develop a rapid method for colorimetric determination of DNA. The method can detect relevant DNA at a minimum of 2.6 nM and can distinguish single base mismatches (Chen et al., 2018). Some research teams have constructed novel nanozymes by integrating existing enzymes onto a nanoscale platform (Petree et al., 2018). This approach enables the nanozymes to exhibit multiple enzymatic activities, which holds promise for broader and more diverse applications in future genetic detection.
2.4.2 Limitations and optimization of nanozyme
Nanozymes, as exogenous nanomaterials, may contain heavy metals such as cadmium and mercury. Their metabolic processes within the body are not yet fully understood, and they may potentially trigger immune responses or cause organ accumulation and toxicity (Li et al., 2020). It is hoped that nanozymes composed of biodegradable materials, such as certain metal-organic frameworks (MOFs) or natural polymers, can be developed in the future (Liu Y. et al., 2020; Liu Y. et al., 2023). Additionally, the catalytic activity of nanozymes is highly dependent on their size, surface chemistry, and exposed surface area. It is extremely challenging to mass-produce nanozymes with entirely consistent structures in a highly reproducible manner. Developing synthesis methods that are precise, reproducible, and capable of ensuring batch-to-batch consistency represents a critical direction for future nanozyme optimization (Wang H. et al., 2024).
3 Nanomaterials-based point mutation detection
Gene mutation detection techniques have become increasingly diverse in recent years. Nanoparticles have attracted significant attention owing to their unique physicochemical properties, such as distinct optical behavior, surface modifiability, and superior electrochemical characteristics. The advent of nanomaterials has created new opportunities for signal transduction and amplification in gene mutation detection. The advantages and limitations of common nanoparticles are shown in Table 1. Their signal outputs are usually combined with different methods, such as colorimetry, electrochemistry, and surface-enhanced Raman scattering (SERS). In this subsection, nanoparticle-assisted nucleic acid probe-based mutation detection methods are introduced according to the different signal output strategies employed.
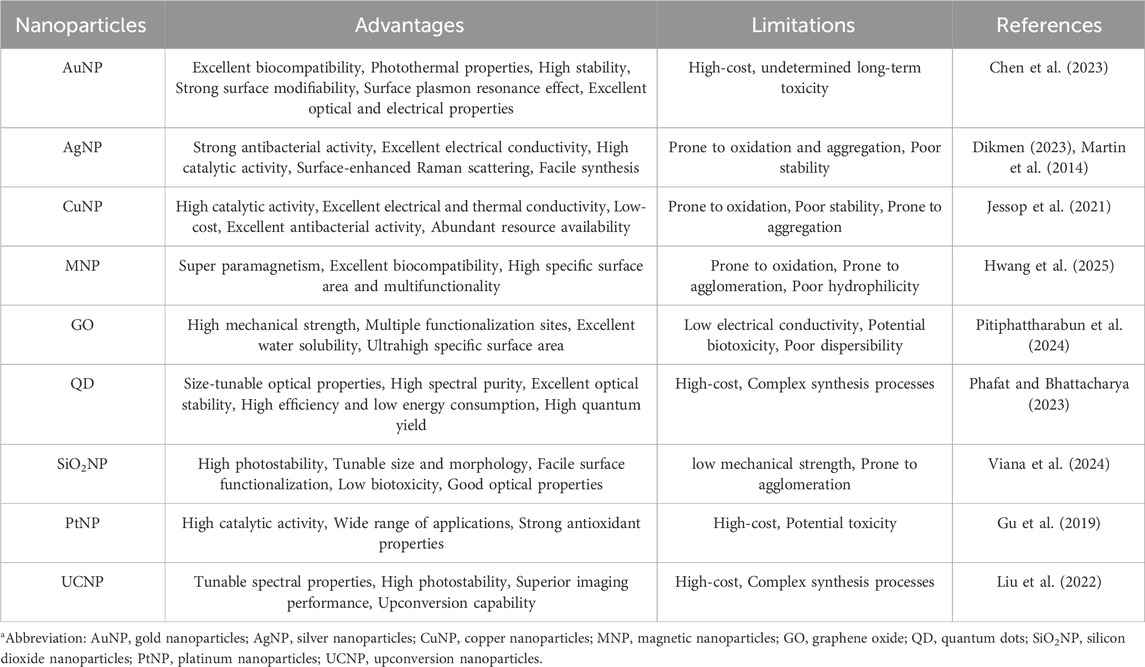
Table 1. Advantages and limitations of common nanoparticlesa.
3.1 Colorimetry
Colorimetry is an analytical method based on the visible color change of a substance after its reaction with a reagent. It is widely used for substance detection and clinical diagnosis because of its simplicity and cost-effectiveness. However, colorimetric methods often suffer from background interference and low sensitivity when detecting targets at low concentrations. Nanomaterials can significantly enhance the efficiency of colorimetric assays because of their unique physicochemical properties.
3.1.1 AuNP-based colorimetry
In colorimetric assays based on nanoparticles, AuNPs are preferred because of their excellent biocompatibility, strong surface modification ability and high stability compared with other particles. As early as 1997, a study was conducted using AuNPs to differentiate target single-stranded oligonucleotides by colorimetric assay and obtained good detection efficiency (Elghanian et al., 1997). The changes in the aggregation behavior of DNA-modified AuNPs induced by salt allowed the distinction between fully complementary and terminal single-base mismatched double-stranded DNA (dsDNA). When the designed probe perfectly matches the target sequence, AuNPs aggregate and produce a color change (Sato, 2005). Since then, colorimetric methods have been increasingly used to detect small nucleic acid molecules and proteins. One example is the detection of EGFR mutations, which can be applied to the early diagnosis of NSCLC (Ramanathan et al., 2019). This study designed probe sequences complementary to the mutated sequences, forming a rigid double-stranded structure. In the presence of an appropriate concentration of NaCl, the dsDNA could not adsorb onto the surface of the AuNPs. Under high salt conditions, the AuNPs lost their protective layer and aggregated, causing a color change to purple. This successfully indicated the presence of EGFR mutations. The double-stranded structure formed between the wild-type sequence and the probe was unstable, and part of the ssDNA could still adsorb onto the surface of the AuNPs, which inhibited aggregation, as shown in Figure 3. Similar methods have also been applied to detect SNPs and other mutation sites in various diseases, such as psoriasis, breast cancer (Bai et al., 2020; Oh and Lee, 2011), and KRAS mutations (Valentini et al., 2013).
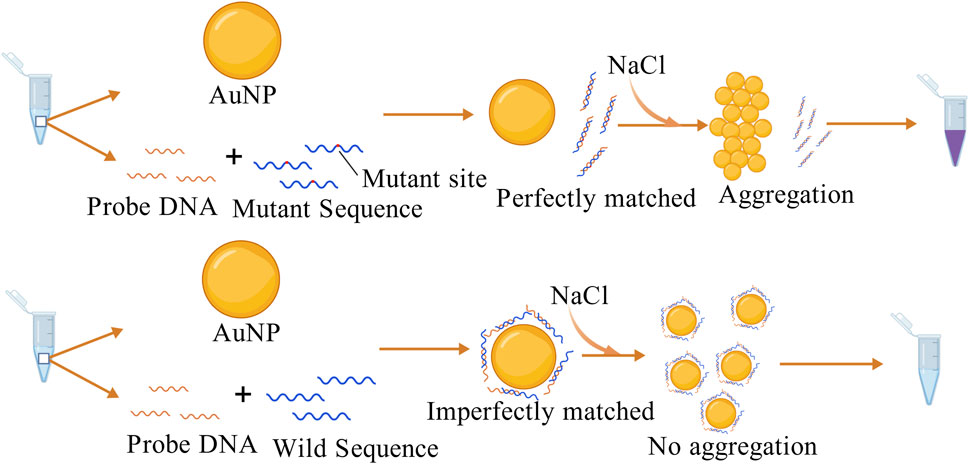
Figure 3. The probe perfectly matches the mutant sequence and cannot adsorb onto the AuNP surface, causing the AuNPs to lose their protective layer and aggregate with a color change in the presence of NaCl. In contrast, the probe partially complements the wild-type sequence, leaving some ssDNA able to adsorb onto the AuNP surface, inhibiting AuNP aggregation (Ramanathan et al., 2019). The figure is created with BioGDP.com.
3.1.2 Combining AuNP with other components-based colorimetry
Because of the excellent biocompatibility of AuNPs, researchers often combine them with other reaction components to construct systems with better performance (Zou et al., 2015). TEMPO-cellulose nanocrystals (TC-CNC)-based stabilized AuNPs probe (TC-AuNPs) was developed by Ganguly et al. The nanoparticles were induced to aggregate using electrostatic interactions between the pathogen DNA and the probe, with a sensitivity capable of reaching 20 fM (Ganguly et al., 2021). Furthermore, Cho and his colleagues explored a molecular mechanism-guided strategy by utilizing the mismatch recognition protein Mutator S (MutS) to specifically identify mismatch sites within double strands on the surface of AuNPs. Different mutation types (e.g., GT, CT, and CC mismatches) were quantitatively analyzed based on the melting temperature (Tm) differences of nanoparticle assemblies. The Tm shift reached 14.7 °C, demonstrating the feasibility of protein-nanoparticle complexes in mutation-specific recognition (Cho et al., 2008).
3.1.3 Limitations and future optimization of colorimetry
Though AuNPs provide excellent detection sensitivity for optical colorimetric analysis, however, there are also many limitations (Zhang et al., 2020). Detection signals are more prone to interference in sequences with high GC content or complex secondary structures (Chou et al., 2019). Moreover, most colorimetric assays merely provide binary (“yes/no”) or semi-quantitative strong/weak signals, making precise quantification challenging. Future strategies may involve integrating nucleic acid amplification techniques or CRISPR/Cas systems with colorimetric readouts to improve single-base resolution. Additionally, the selection of salts is regarded as one of the critical factors influencing reaction conditions (Yu et al., 2009). Unlike conventionally used sodium chloride, the application of sodium fluoride has increased sensitivity by 2.3-fold. This enhancement is attributed to the weak adsorption of fluoride ions on gold nanoparticles. Notably, polyadenine sequences provide stronger protection for gold nanoparticles than poly-thymine sequences, suggesting the potential value of sequence design in sensor optimization (Hu et al., 2020).
3.2 Fluorescence methods
Compared to traditional fluorescent dyes, nanoparticles (e.g., QDs, metal nanoparticles) offer higher fluorescence quantum yields and lower background noise, making them highly sensitive for detecting low-concentration mutations. QDs stand out for their high photostability and surface tunability, which allow them to bind to specific DNA probes and detect mutations by changes in fluorescence signals (Hosseini et al., 2017; Kim and Gang, 2018).
3.2.1 QD-based fluorescence detection
In 2005, Zhang et al. proposed a DNA nanosensor based on single QD and FRET. When the target DNA is present, the biotin-labeled capture probe and the Cy5-labeled reporter probe “sandwich” the target. The resulting complex binds to QDs modified with streptavidin on the surface, triggering a FRET effect. Under excitation with a 488 nm laser, both QD and Cy5 emissions can be detected. In the absence of target DNA, only the QD signal is observed. And the sensitivity of the single molecule detection is less than 50 copies, as shown in Figure 4A, which is a hundred-fold increase in sensitivity over standard MB (Zhang C.-Y. et al., 2005). Subsequent research teams developed QDs of different colors to label probes specific to mutant and wild-type alleles independently, enabling fluorescent signals to distinguish DNA types (Yeh, 2006). In a dispersed state, QDs generally have narrow bandwidth, high color purity in fluorescence spectra and large fluorescence quantum yield. When QDs aggregate, their fluorescence can be reduced to the point of becoming completely quenched. The mechanisms and factors contributing to fluorescence quenching may include FRET, Dexter energy transfer, QD concentration, and external environmental conditions (Kim et al., 2010; Liang et al., 2016; Zhou et al., 2022). Although fluorescence quenching is generally considered an unfavorable factor, in certain cases, it can be leveraged to develop highly sensitive detection methods. Utilizing the fluorescence quenching phenomenon of QDs aggregation, an assay has been proposed to simultaneously detect multiple mutations in the EGFR gene. The technique utilizes the spectrally resolved properties of QDs of different sizes to couple with specific molecular beacon probes targeting common EGFR mutations (L858R). The probes are designed for different mutation sites, and they are conjugated to distinct QDs. In the absence of mutation sequences, the QDs remain dispersed and continuously emit fluorescence. When a specific mutation is present, the mutation sequence hybridizes with the corresponding probe on the QD to form a double-stranded structure, leading to QD aggregation and subsequent fluorescence quenching. This method enables simultaneous detection of two mutations in clinical samples at 10 ng/μL (Figure 4B), with a detection limit as low as 0.01% (Kang et al., 2012).
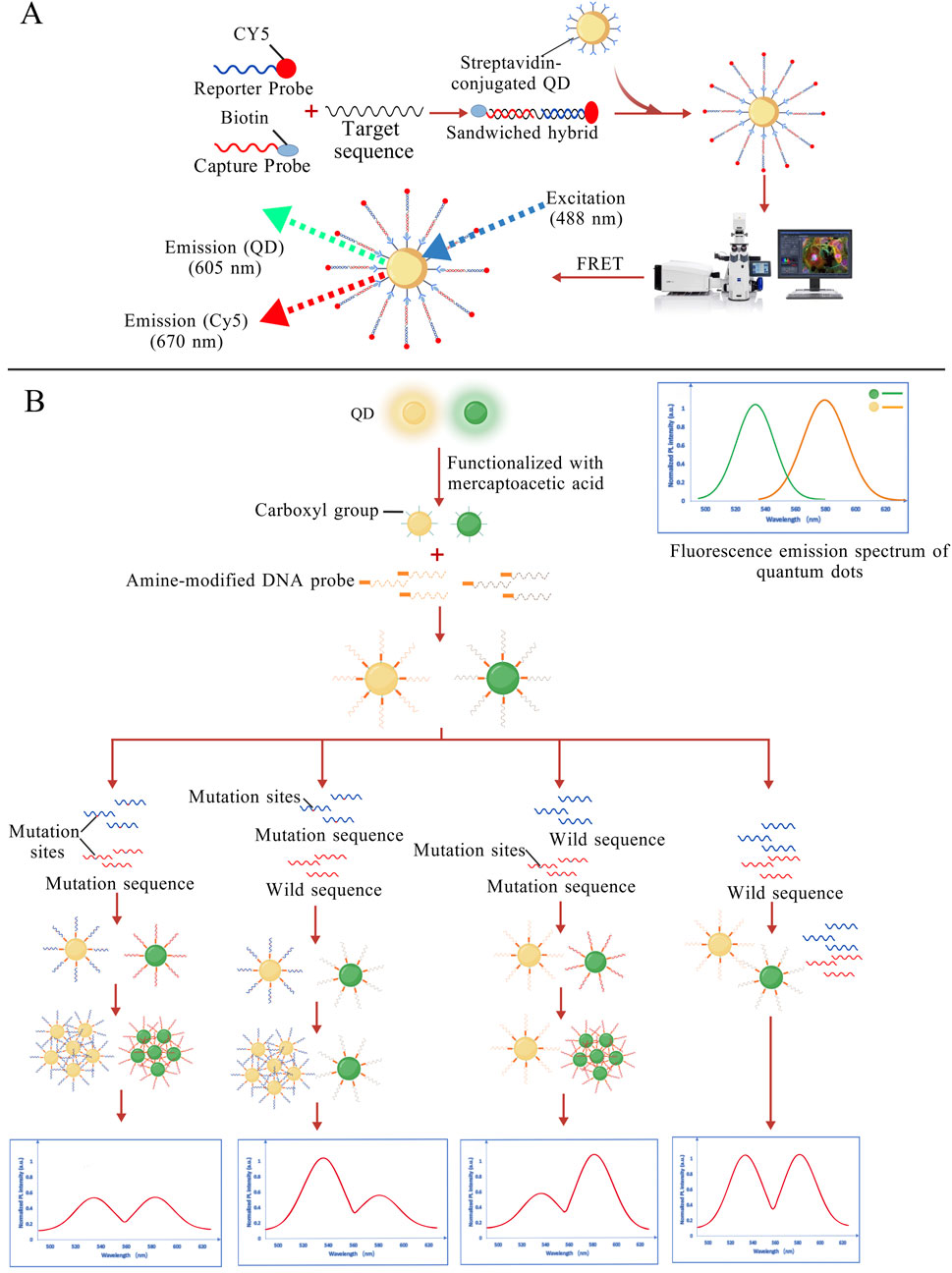
Figure 4. (A) The reporter probe and capture probe sandwich the target DNA and are conjugated to QDs. Under 488 nm laser excitation, signals from the QDs and Cy5 can be detected (Zhang R.-Y. et al., 2005); (B) Probes designed for different mutation sites are conjugated to distinct QDs. When the mutation sequence is present, the corresponding QDs aggregate, leading to fluorescence quenching, thereby enabling mutation detection. (Kang et al., 2012). The figures are created with BioGDP.com.
CdTe QDs are known for their exceptional robustness (Farahmandzadeh et al., 2024; Yang et al., 2019). CdTe QDs also exhibit tunable fluorescence properties upon interaction with different nucleotides, including emission wavelength shifts (red- or blue-shift) and fluorescence quenching (Siegberg and Herten, 2011). These spectral changes allow for the differentiation of specific DNA sequences. Building on this principle, Moulick et al. developed an innovative approach for detecting DNA presence, damage, and mutations. They successfully identified the target DNA in extracts of human cancer cells (PC3) and normal cells (PNT1A) at concentrations as low as 500 pM (Moulick et al., 2017).
3.2.2 UCNP-based fluorescence detection
Upconversion nanomaterials (UCNPs), a class of nanomaterials capable of converting low-energy light (infrared) into higher-energy light (visible or ultraviolet), have gained significant attention in bioimaging and spectral analysis (Mohan and Poddar, 2021). Kumar et al. designed an oligonucleotide sensor based on UCNPs (donor) and TAMRA fluorescent dye (acceptor) to detect the AT single-nucleotide mutation associated with sickle cell disease. This sensor captures the target DNA through a sandwich hybridization approach and triggers energy transfer based on the luminescence resonance energy transfer (LRET) mechanism. The method enables highly sensitive detection of the pathogenic AT mutation in sickle cell disease, achieving a detection limit as low as 120 fM (Kumar et al., 2009). Sensitive detection of DNA can also be achieved by tuning the excitation and emission wavelengths of UCNPs. In this approach, a near-infrared (NIR) fluorescent nanoprobe is constructed through complementary pairing of surface-modified DNA between UCNPs and silver nanoclusters (AgNCs). Experimental results demonstrated that the probe could detect KRAS gene mutations within a concentration range of 5 pM–1000 pM (Wang P. et al., 2014).
3.2.3 Limitations and future optimization of fluorescence methods
Although fluorescence detection is more sensitive than colorimetric methods, background autofluorescence and nonspecific binding in complex samples still affect the signal-to-noise ratio (Izumi et al., 2011; Wang S. et al., 2023). In addition, quantitative results may be influenced by photobleaching, fluorescence quenching, and probe concentration, leading to insufficient reproducibility. By employing appropriate background suppression strategies, such as the use of MB or dual-probe recognition, nonspecific signals can be effectively reduced (Lee et al., 2024).
3.3 Electrochemical methods
Electrochemical methods usually affect the electrochemical signals through redox reactions, ion migration, or conductivity changes to achieve the detection of target molecules. Nanomaterials show great potential for use in electrochemical methods owing to their excellent electrical conductivity, large specific surface area, and good surface functionalization.
3.3.1 Design and optimization of AuNP-based electrochemical methods
AuNPs have become a popular choice in building electrochemical biosensors. By coating electrode surfaces with AuNPs, these sensors can boost DNA hybridization signals, making them more effective at detecting base mismatches (Zhang and Zhang, 2010), especially AG mismatches. Compared to conventional sensors, AuNP-modified sensors exhibit significantly enhanced sensitivity, achieving detection limits in the nanomolar range (Wang Q. et al., 2011; Zhang R.-Y. et al., 2005). In AuNP-based mutation detection systems, the immobilization density of DNA probes critically governs both sensitivity and specificity. Conventional methodologies relying on thiol-Au covalent bonding are constrained by steric hindrance and inter-probe electrostatic repulsion, leading to suboptimal probe loading density and heterogeneous distribution. To enhance DNA probe attachment density, recent advancements incorporate vertically aligned multi-walled carbon nanotubes (MWCNTs) synergistically with AuNPs on modified electrodes. Experimental validation has demonstrated favorable detection sensitivity for sequence-specific hybridization events related to TP53 gene detection, monitored via electrochemical impedance spectroscopy (EIS) (Fayazfar et al., 2014). In addition, current strategies predominantly immobilize AuNPs onto electrode surfaces via chemical bonding or physical adsorption. These strong interactions often result in permanent occupation of surface-active sites and irreversible alterations to the crystalline structure of AuNPs, severely compromising their reusability. To overcome this limitation, Chen’s team has proposed an electrode modification-free detection system in which AuNPs are directly used as the electrolyte. In this system, AuNPs act as catalysts whose activity is suppressed by the adsorption of ssDNA. The introduction of target DNA induces hybridization with the probe, resulting in desorption from the AuNP surface. The restored AuNPs catalyze the oxidation of glucose, initiating a cascade of redox reactions involving horseradish peroxidase (HRP) and the redox indicator thionine. The electrochemical signal is subsequently measured using a glassy carbon electrode (GCE) (Figure 5A). The system allows direct reuse by simply replacing the electrolyte solution for subsequent experiments (Chen et al., 2014).
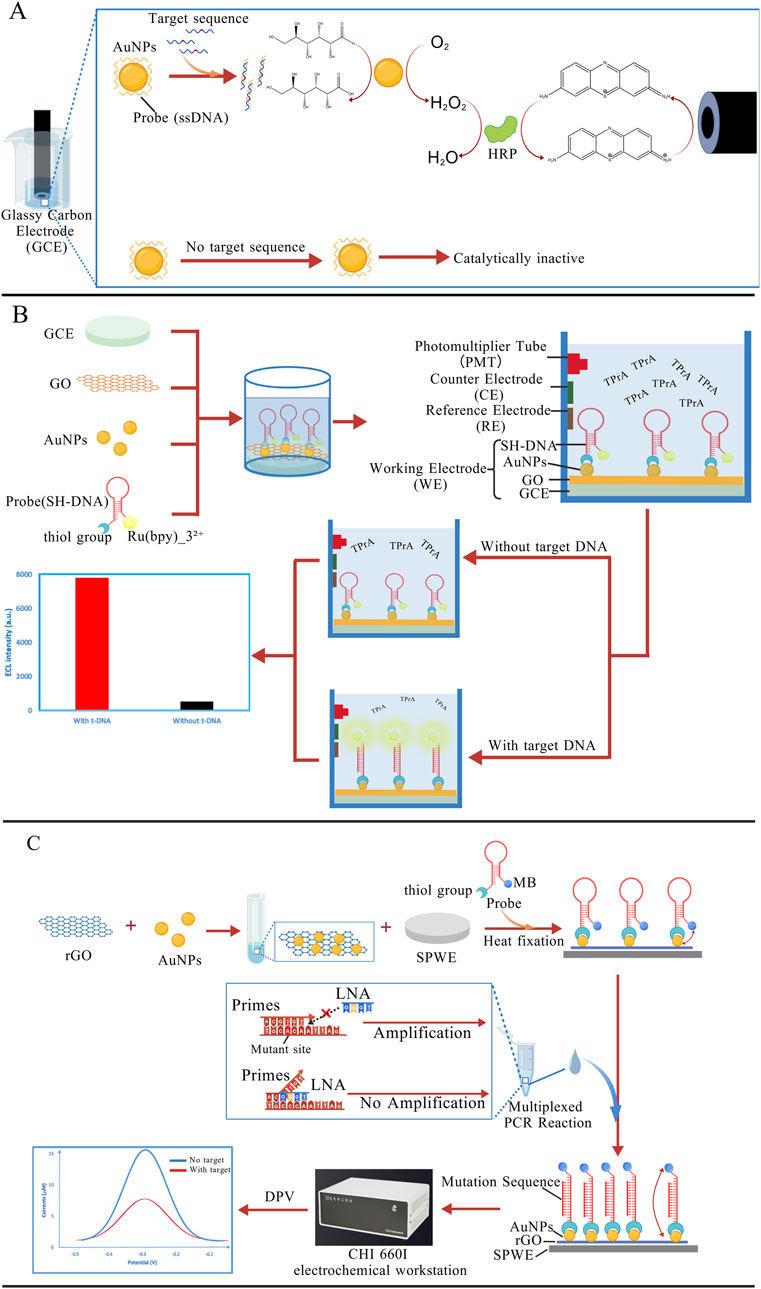
Figure 5. (A) AuNPs act as catalysts, and the catalytic activity is suppressed by the probes adsorbed on the surface. When the target DNA is present, the probes dissociate from the AuNP surface, thereby allowing the AuNPs to initiate a redox cascade reaction (Chen et al., 2014); (B) On the GCE, GO can quench the ECL of the Ru (bpy)32+/tripropylamine (TPrA) system. In the presence of the target sequence, the hairpin probe structure unfolds, leading to the removal of GO from the electrode surface. As a result, the ECL signal is recovered (Huang et al., 2015); (C) Under the action of the LNA probe, the mutant sequence is selectively amplified. When the PCR products are introduced into the electrochemical sensor, the probe hairpin structure is opened, moving the MB molecule away from the electrode surface and resulting in a decrease in the electrochemical signal (Wang et al., 2022). The figures are created with BioGDP.com.
3.3.2 GO-based electrochemical methods
Other nanomaterials have also demonstrated comparable performance in electrochemical DNA sensing platforms (Hu et al., 2011; Mehrgardi and Ahangar, 2011; Muti et al., 2010; Skotadis et al., 2016). Graphene oxide (GO), one of the most versatile nanomaterials, has found widespread use in sensor development, electrochemical systems, and energy storage applications. Notably, GO efficiently and stably quenches the electrochemiluminescence (ECL) of the tris(2,2′-bipyridine)-ruthenium (II) (Ru (bpy)32+)/tripropylamine (TPrA) system when deposited onto GCEs. This quenching behavior is primarily attributed to GO’s oxygen-rich surface functional groups, limited electrical conductivity, and the two-dimensional structure of the GO sheets (McCall et al., 1999). Subsequently, Huang et al. exploited this phenomenon to engineer a novel biosensing platform by incorporating GO onto AuNP-functionalized GCEs via DNA probe self-assembly. The GO-mediated ECL quenching served as the core detection mechanism. The DNA probes maintained intimate contact with GO surfaces without target DNA, resulting in substantial ECL suppression. Hybridization with complementary targets induced probe detachment from GO, thereby restoring ECL signals (Figure 5B). Systematic characterization revealed a linear response to target DNA concentrations spanning six orders of magnitude (100 a.m.-10 pM) (Huang et al., 2015).
3.3.3 QD-based electrochemical methods
QDs exhibit not only excellent optical properties but also favorable electron transport and storage capabilities, and even the fundamental property of improving electrode interfaces. Therefore, some research groups have combined QD with electrochemical methods for detection. In the electrochemical sensor designed by Mazloum-Ardakani et al., CdS QDs were utilized as the primary functional material to enhance the electroactivity of the electrode interface. This sensor demonstrated the successful application of high-sensitivity detection for the ApoB100 R3500Q mutation (Mazloum-Ardakani et al., 2015). Yang et al. developed a highly sensitive ECL biosensor by employing CdS QDs labeled with thioglycolic acid (TGA) and AuNP-labeled hairpin DNA probes. In the absence of target DNA, FRET occurs when AuNPs are in close proximity to CdS QDs, quenching the ECL signal of the CdS QDs. In the presence of target DNA, the hairpin structure is opened, increasing the distance between AuNPs and CdS QDs, thereby reducing energy transfer and enhancing the ECL signal. This biosensor ultimately achieved precise detection of the EGFR T790M mutation, with a detection limit as low as 3.4 a.m (Yang et al., 2023). This approach presents a novel strategy for electrochemical processes incorporating QDs, which could similarly be extended to other QDs (Yang et al., 2021).
3.3.4 Combining nanoparticles with amplification techniques
As electrochemical detection techniques improve, researchers are exploring novel strategies that integrate selective nucleic acid amplification techniques with nanoparticles (Lu et al., 2017; Yang et al., 2016). As an example, Wang et al. developed a detection platform that combines electrochemical sensing with LNA-mediated multiplex PCR. The system enables selective amplification of mutant EGFR genes (L858R, T790M), followed by detection of the amplified products using a screen-printed working electrode (SPWE) modified with reduced graphene oxide-gold nanoparticles (rGO-AuNPs). In the absence of target DNA, the probe folds into a hairpin structure that brings the methylene blue (MB) molecule close to the electrode surface. As an electroactive molecule, MB enables efficient electron transfer between the electrode and itself, generating a strong current signal. In the presence of the mutant sequence, the hairpin structure of the probe unfolds, causing the MB label at its end to move away from the electrode surface. This structural change leads to a decrease in the electrochemical signal (Figure 5C). Experimental results showed that this method could accurately detect rare mutations at just 0.05%, offering a sensitivity improvement of two orders of magnitude compared to traditional electrochemical methods (Wang et al., 2022). Owing to the signal amplification ability of nanoparticles and their applicability under different experimental conditions, nanoparticles are promising for improved diagnostics of several diseases.
3.3.5 Limitations and future optimization of electrochemical methods
Electrochemical gene detection usually requires electrode modification with nanomaterials or functionalization (such as AuNPs or carbon nanotubes), which involves complex processes and high equipment demands. In addition, conformational changes of immobilized probes on the electrode surface and nonspecific adsorption can interfere with signal interpretation. By introducing novel electrocatalytic materials such as nanozymes or MOFs, the signal intensity and stability can be improved, thereby partially addressing these issues (Liu et al., 2024; Wang Y. et al., 2023).
3.4 Surface-enhanced Raman scattering (SERS)
Raman scattering signals arise from the inelastic scattering of light with matter. Although Raman signal intensity constitutes only about one-millionth of the incident light, when molecules are adsorbed onto nanoscale rough surfaces (such as AuNP, CuNP, or nanostructures), the Raman scattering signal is greatly enhanced, and even single-molecule detection can be achieved. The enhancement effect mainly originates from Localized Surface Plasmon Resonance (LSPR) and the chemical enhancement mechanism between the molecule and the metal surface. Owing to the unique molecular fingerprint characteristics of SERS, this technique enables highly sensitive detection of specific molecules at extremely low concentrations (Lyu et al., 2024).
Detection of target molecules with SERS can be accomplished by using two main approaches. SERS can directly obtain structural information of molecules through the enhanced Raman signal of molecules that come close to the metal surface (Xie and Schlücker, 2013). Alternatively, SERS labels or nanolabels conjugated to recognition elements (e.g., antibodies) produce strong Raman signals in proximity to the target, enabling both qualitative and quantitative analysis in complex biological environments (De Silva Indrasekara and Fabris, 2015). Accordingly, the design and selection of nanomaterials are crucial in SERS experiments. In nucleic acid detection, SERS mainly utilizes the design of specific nucleic acid probes, which hybridize with target DNA or RNA. Probes designed on the metal surface form perfect matches with mutant sequences and imperfect matches with wild-type sequences, generating different SERS signals. This difference in signal constitutes the key principle of using SERS to detect point mutations, as shown in Figure 6. The proper design of nanostructured surfaces critically affects both the detection rate and the detection limit of mutation sequences.
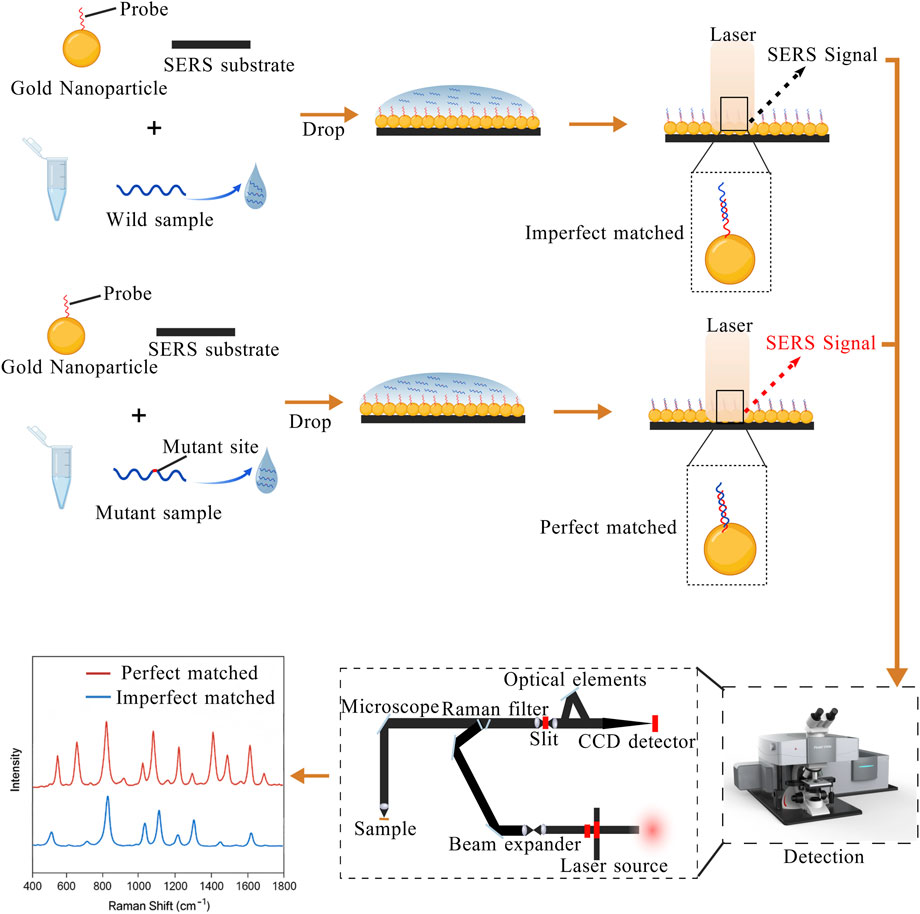
Figure 6. The probes on gold nanoparticles are partially complementary to the wild-type sequence but perfectly complementary to the mutant sequence, generating distinguishable SERS signals. The figure is created with BioGDP.com.
3.4.1 Combining SERS with PCR
Li et al. employed a modified sodium citrate reduction method to synthesize AgNP colloids with irregular surface roughness, which facilitated the formation of “hot spots”. Subsequently, the addition of spermine acted as both an aggregating agent and a neutralizer of the DNA backbone, leading to further aggregation of AgNP and the formation of additional hot spots, thereby significantly enhancing the SERS signal. In addition, they combined SERS with multiplex PCR to simultaneously amplify multiple mutation types in a single reaction. The specifically designed nucleic acid probes produced distinct SERS signals when hybridized with different sequences, and the relative peak intensities in the SERS spectra enabled discrimination among various nucleic acid sequences. Using this approach, they successfully achieved simultaneous detection of multiple hotspot point mutations in genes such as KRAS and PIK3CA (Li et al., 2017; Li et al., 2018). In contrast to conventional PCR, asymmetric PCR (Asy-PCR) adjusts the ratio of forward and reverse primers to preferentially generate ssDNA. A research team developed a highly specific Asy-PCR/SERS method to detect KRAS G12 V mutation. The researchers chose to functionalize AuNPs and then conjugated them with the Raman reporter molecule DTNB and mutation-specific DNA probes to form SERS nanotags. Subsequently, PCR products were hybridized with the tags via heating and annealing, the target complexes were enriched using streptavidin-coated magnetic beads, and characteristic peaks were detected with a Raman spectrometer. This method can be used for the detection of KRAS G12V mutations at just 0.1% in the presence of non-target sequences (Lyu et al., 2021). However, a limitation of this approach lies in the potential self-folding of ssDNA products generated by Asy-PCR, which may form unintended secondary structures (e.g., hairpins), thereby reducing hybridization efficiency with specific probes. Although optimized PCR designs allow for the simultaneous amplification of multiple DNA targets and improve detection throughput, they remain limited by the intrinsic complexity of the amplification process. To address this, Wu et al. proposed an amplification-free SERS-based strategy. They used Au@Ag core-shell nanorods (Au@Ag NRs) to prepare a highly sensitive and uniform SERS substrate, yielding an enhancement factor of 1.85 × 105. By this approach, they successfully distinguished mutant from wild-type KRAS genes with a detection limit of 50 fM (Wu et al., 2019).
3.4.2 Limitations and optimization of SERS-based point mutation detection
Although SERS has shown great promise in boosting sensitivity, especially combined with other techniques, traditional approaches still face a few notable hurdles in DNA analysis. For instance, signal instability frequently results from interference caused by impurities and nanomaterials with low surface enhancement factors, with thymine (T)-derived signals being particularly susceptible (Wang H. et al., 2014). To address these issues, researchers have introduced thiosulfate-modified AgNPs, which not only eliminate the unwanted effects of impurities but also help maintain the integrity of DNA signals (Zeng et al., 2021; Zhang et al., 2022). Another limitation arises from the intrinsically small scattering cross-sections of conventional Raman markers. Li et al. engineered a hybrid sensing platform which integrates silver nanorice antennae with gold triangular nanoprism arrays (SNA-GTNA) for the detection of HBV DNA. This nanoarray-coupled architecture demonstrated a remarkable SERS signal amplification exceeding four orders of magnitude compared to planar gold film substrates, accompanied by an extended electromagnetic field enhancement spanning hundreds of nanometers (Li et al., 2013). Notably, while classical theory posits that Raman signal intensity diminishes with increasing distance between reporter molecules and metallic substrates, emerging evidence reveals counterintuitive enhancement phenomena in SERS-based sandwich-structured DNA sensors. Specifically, certain configurations exhibit stronger signals when reporters are positioned farther from the metallic surface. This spatial dependency introduces new design considerations for optimizing SERS-based nucleic acid detection architectures (Pyrak et al., 2023).
SERS technology demonstrates exceptional sensitivity and resolution not only in genetic-level analyses but also in disease subtyping, pathogen identification, and viral infection detection (AlSafadi et al., 2024; Chen et al., 2025; He et al., 2024; Ye et al., 2022). Although nanoparticle-based detection technologies possess great potential for sensitivity and specificity, the toxicity of such particles is still nonnegligible. The exploration of nanoparticle biocompatibility and biodegradability is of priority to future research. Thus, analytical performance must be balanced against potential biological hazards to ensure safe diagnostic applications (Brunelli et al., 2024; Ruijter et al., 2023).
4 Nucleic acid analogs-based mutation detection
4.1 Peptide nucleic acid (PNA)
PNA is a synthetically engineered molecule. Unlike DNA, which features a negatively charged phosphodiester backbone, PNA possesses a neutral pseudo-peptide backbone. This structural distinction endows PNA with several unique properties. On one hand, the peptide backbone renders PNA electrically neutral. The lack of electrostatic repulsion enables PNA to exhibit significantly higher binding affinity to complementary DNA than conventional DNA/DNA duplexes (Almarsson and Bruice, 1993; Nielsen, 1997). On the other hand, PNA demonstrates high resistance to common nucleases and proteases (Yagita et al., 2020), maintaining excellent stability both in vitro and in vivo (Rose et al., 2019). Owing to its high binding specificity and affinity for target DNA or RNA, PNA has been widely applied in nucleic acid detection platforms.
4.1.1 Combining PNA with PCR
Due to its resistance to DNA polymerase, PNA has been combined with PCR to develop PNA-mediated PCR clamping technology (Fouz and Appella, 2020). Its principle is shown in Figure 7A, PNA probes can invade wild-type dsDNA and bind tightly to it, thereby preventing subsequent PCR amplification. In contrast, due to the presence of the mutation site, the probe cannot invade mutant dsDNA, allowing the mutant sequence to be normally amplified. A research team employed this method to evaluate the feasibility of plasma cell-free DNA (cfDNA) as an alternative specimen for detecting EGFR mutations in NSCLC patients. Among 40 patients with confirmed tumor tissue mutation status, 35 were identified as carrying EGFR-activating mutations (Kim et al., 2013). Although this approach successfully achieved partial non-invasive detection, its sensitivity remains suboptimal and requires further optimization. Rosso and co-workers integrated fluorescence in situ hybridization (FISH) with PNA-PCR by employing fluorescence-labeled PNA probes to detect BCR-ABL T315I mutations in two subtypes of chronic myeloid leukemia (CML). This approach achieved a detection sensitivity of 0.5% mutant alleles in mixed samples (Rosso et al., 2015).
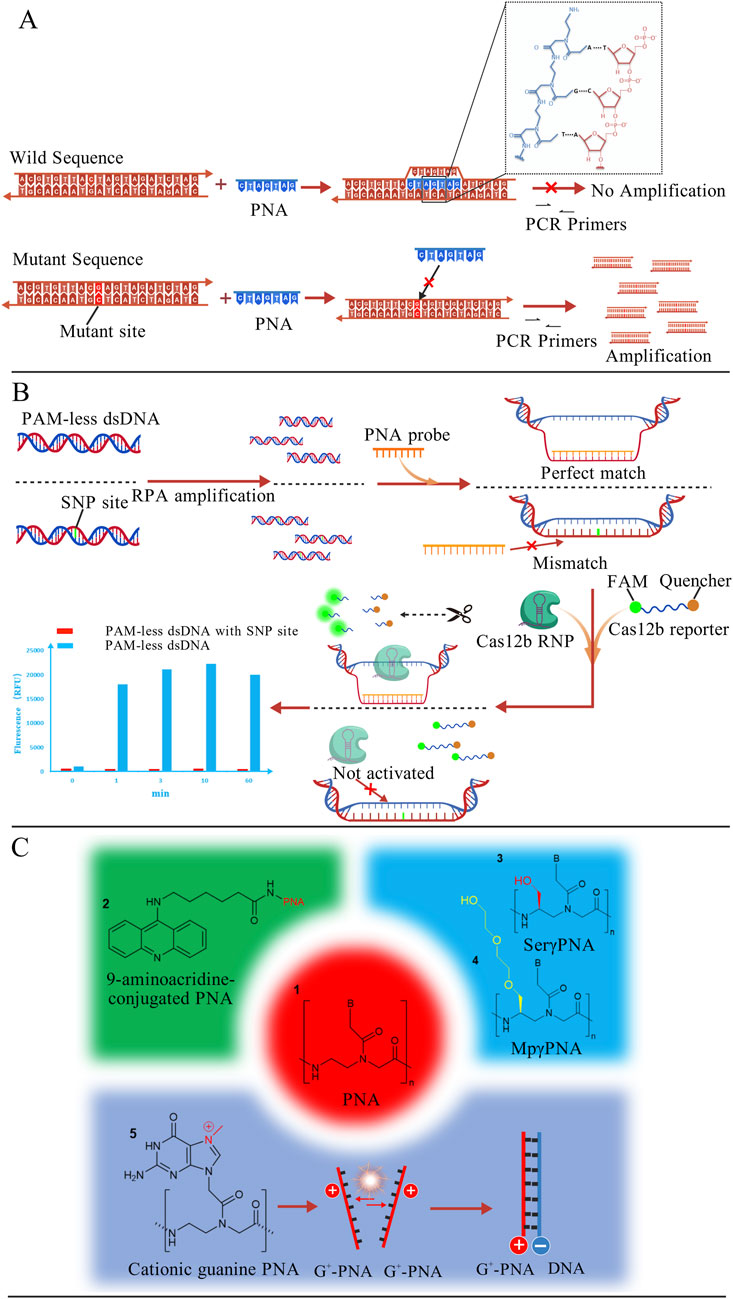
Figure 7. (A) PNA probes invade wild-type dsDNA, preventing its subsequent amplification. Mutant DNA, which cannot be invaded, can thus be amplified and detected. The binding mode of PNA with DNA is shown in the figure (Fouz and Appella, 2020); (B) PNA probes invade PAM-less dsDNA and generate a ssDNA window. Subsequently, Cas12b RNP and reporter molecules are introduced. The exposed ssDNA activates the trans-cleavage activity of Cas12b, which cleaves the reporter molecule and generates a strong fluorescence signal. PAM-less dsDNA with SNP site cannot be invaded and fails to activate Cas12b cleavage activity (Jiang et al., 2023); (C) Structural formula of PNA with specific molecular modifications (Bentin and Nielsen, 2003; Hibino et al., 2020; Oyaghire et al., 2023). The figures are created with BioGDP.com.
4.1.2 Combining PNA with protease
Compared to DNA, PNA exhibits greater structural flexibility (Chhetri et al., 2022), allowing it to adopt multiple conformations. This property enables PNA to interact with a broad range of biomolecules, including enzymes (Saikia et al., 2021). Jiang et al. developed a PNA-based universal detection toolkit (PNA-Pdx) that integrates the high specificity of PNA probes with CRISPR-Cas12b systems, as shown in Figure 7B. Relying on its high affinity, PNA invades and locally unwinds wild-type dsDNA, forming a ssDNA window. Subsequently, the Cas12b-sgRNA complex and fluorescent reporter molecules labeled with a fluorophore and quencher are introduced. If the exposed single-stranded sequence is fully complementary to the sgRNA, the trans-cleavage activity of Cas12b is activated, cleaving the reporter and generating a strong fluorescence signal. In the presence of an SNP mismatch, however, the probe cannot effectively invade, resulting in markedly reduced Cas12b activity and quenched fluorescence, thereby enabling precise SNP discrimination. This platform successfully detected diverse mutation types including AG, GC, and TA substitutions with a sensitivity of 300 pM. The methodology not only enhanced detection sensitivity but also overcame the PAM dependency constraints inherent to conventional CRISPR technologies (Jiang et al., 2023).
4.1.3 Limitations and optimization of PNA probe
Although conventional PNA is theoretically capable of sequence-specific binding to DNA or RNA, its practical binding efficiency is often suboptimal. Researchers typically address this by modifying PNA with auxiliary molecules at specific sites to enhance binding affinity (Figure 7C). For instance, acridine-conjugated PNAs exhibit binding affinities 20–150-fold higher than those of unmodified PNAs (Bentin and Nielsen, 2003). Further structural optimizations, such as γ-position modifications with N7-methylguanine (G+), hydroxymethyl and diethylene glycol, have been demonstrated to enhance PNA binding capacity to varying degrees (Hibino et al., 2020; Oyaghire et al., 2023). Additionally, conventional PNAs exhibit inherent challenges including poor water solubility and inefficient cellular internalization (Gasparello et al., 2019). To overcome these barriers, Aiba et al. conjugated nuclear localization signal (NLS) peptides to pseudo-complementary PNA (pcPNA) (Aiba et al., 2015). Experimental validation revealed that NLS-pcPNA rapidly formed invasion complexes with dsDNA even at low concentrations and under high-salt conditions (100 mM NaCl). Moreover, the predominant synthesis of PNA via solid-phase peptide synthesis (SPPS) incurs high production costs, restricting its widespread adoption. Researchers have reduced the cost of PNA synthesis by employing physicochemical strategies such as utilizing low-cost commercial fluid-handling components (Kallmyer et al., 2020), incorporating safety-catch protecting groups (Nandhini et al., 2023), and implementing ultrasound-assisted synthesis (Del Bene et al., 2023). With continued advancements in synthetic technologies, it is anticipated that more cost-effective synthesis approaches will emerge in the future.
4.2 Locked nucleic acids (LNA)
LNA is a chemically modified nucleic acid analog characterized by a unique structure. This structure is formed by a methylene bridge linking the 2′-oxygen and 4′-carbon atoms of the ribose ring. This modification locks the ribose in a C3′-endo conformation, significantly enhancing its binding affinity for complementary DNA or RNA strands (Egli et al., 2001; Petersen et al., 2002). Compared to regular DNA, LNA provides a 2 °C–8 °C increase in thermal stability (ΔTm) for each incorporated monomer (Owczarzy et al., 2011), along with better discrimination against base mismatches (Mishra et al., 2013). Owing to these properties, LNA has emerged as an optimal candidate for the design of high-sensitivity nucleic acid probes.
4.2.1 Design and optimization of LNA probes
Traditional invader probes are short, blunt-ended dsDNA. By incorporating intercalating agents such as 2′-O-(pyren-1-yl)methyl-RNA monomers arranged in a specific zipper configuration, these probes generate “energetic hotspots”. These hotspots weaken the stability of the DNA duplex by disrupting base-pair stacking. However, conventional Invader probes exhibit poor efficiency for long DNA fragments and GC-rich sequences, and show dependency on specific ionic strength and pH conditions. Building upon this framework, researchers developed a novel dsDNA-targeting probe, termed Invader LNA, by incorporating 2′-N-(pyren-1-yl)methyl-2′-amino-α-L-LNA monomers. This probe maintains robust mismatch discrimination capability even under high-salt (710 mM Na+) and physiologically relevant ionic strength conditions (Sau et al., 2010). Moreover, incorporating a 3-6 nucleotide single-stranded overhang at the 5′ end of the probe shortens the duplex region, further promoting strand separation. The new designed invasive LNA probes can invade mutant sequences, and be detected by gel analysis (Figure 8A). Experimental results demonstrated that this design further enhances both the sensitivity and specificity of the Invader LNA probes (Adhikari et al., 2021; Adhikari et al., 2022).
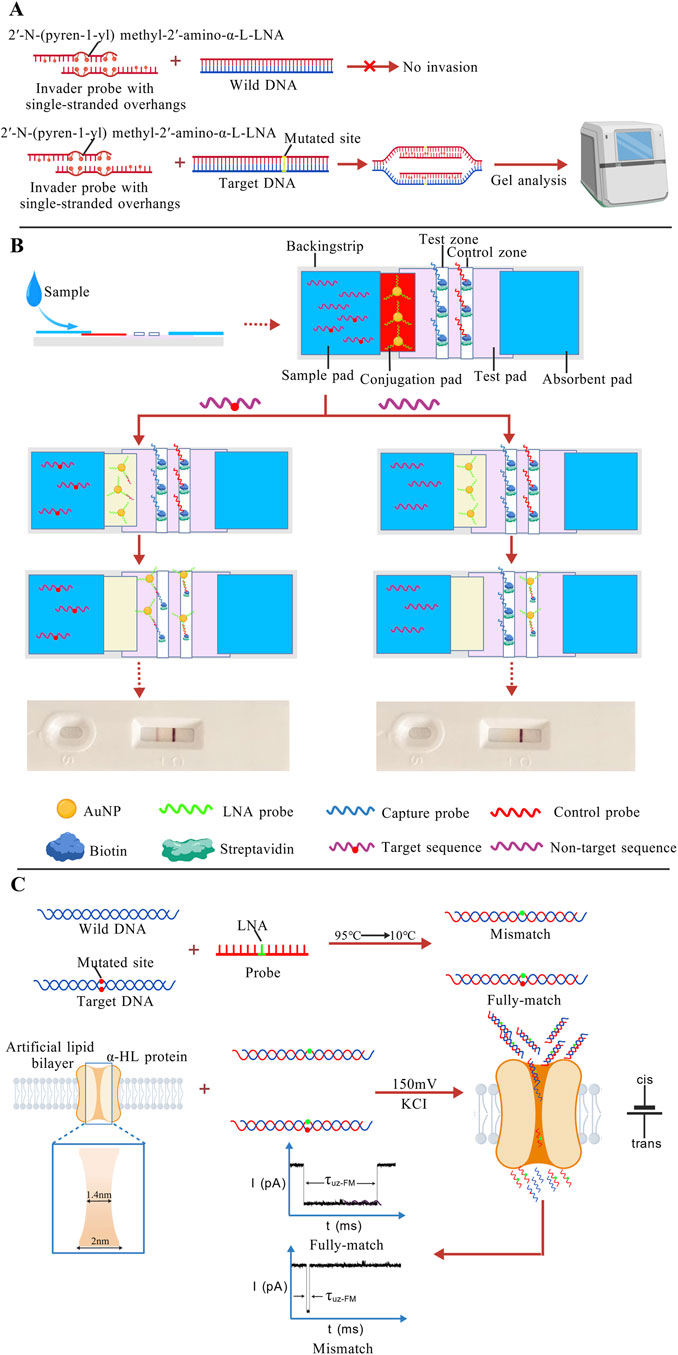
Figure 8. (A) Probes with single-stranded overhangs can exclusively invade mutant DNA without affecting wild-type DNA, ultimately allowing differentiation through gel electrophoresis results (Adhikari et al., 2022); (B) The target sequence migrates from the sample pad to the conjugation pad via capillary action, where it binds to LNA-modified AuNP probes to form a complex. A part of the complex is captured by the capture probe. Excess gold nanoparticle probes are captured by complementary probes at the C line, resulting in color development at the T line and C line. The appearance of two lines indicates a positive detection result. When the target sequence is absent in the sample, no complex is formed. Only the LNA probes are captured and visualized at the C line, producing a single pink band, which indicates a negative detection result (Rastogi et al., 2012); (C) The LNA probe is fully complementary to the mutant DNA but mismatched with the wild-type DNA. Under an applied voltage, mismatched duplexes dissociate, and translocate rapidly through the nanopore, whereas the perfectly matched duplex passes through more slowly (Tian et al., 2018). The figures are created with BioGDP.com.
4.2.2 Combining LNA with PCR
Similar to PNA, the high binding affinity and specificity of LNA probes enable the selective suppression of wild-type DNA amplification during PCR, allowing for the detection of mutations at frequencies as low as 0.1% (Ishige et al., 2016; Warshawsky and Mularo, 2011). Furthermore, a synergistic approach combining both PNA and LNA has been developed. In this strategy, fully complementary PNA probes stably bind to and block the amplification of wild-type DNA. Meanwhile, LNA-modified primers complementary to mutant sequences facilitate selective amplification of mutant DNA. This method achieved rapid KRAS G12D and G12V mutation detection by loop-mediated isothermal amplification (LAMP) at 65 °C. The assay demonstrated a detection sensitivity exceeding 0.1%, surpassing that of conventional sequencing and traditional PNA-clamping PCR (Itonaga et al., 2016). The biocomplex formed by the joint application of PNA and LNA interacting with target DNA exhibits enhanced stability and specificity (Lundin et al., 2005).
4.2.3 Combining LNA with nanotechnology
Among various nanomaterials, AuNPs are the most commonly used in combination with LNA (Lin et al., 2013a; 2013b). The excellent mismatch discrimination capability of LNA, together with the stability and signal amplification properties of AuNPs, often enables lower detection limits. For instance, Shiva et al. developed a lateral flow test strip that integrates LNA-modified probes with gold nanoparticles for the detection of DNA specific to the foodborne pathogen E. coli O157:H7. After the target DNA sample binds with the LNA-modified AuNP probes on the conjugation pad to form a complex. A part of the target DNA is specifically captured by the immobilized capture probe in this zone, thereby aggregating the AuNPs on the test line (T line) to form a visible pink band. Excess AuNP probes continue to flow further to the control line (C line), where they are captured by another complementary probe, forming a second pink band-resulting in the appearance of two lines. If the target DNA is absent, the AuNP probes cannot form the complex. However, these free probes continue to flow to the C line, where they are immobilized, forming a single pink band (Figure 8B). This method achieved a detection limit as low as 0.4 nM (Rastogi et al., 2012). LNA can not only work synergistically with traditional inorganic nanomaterials such as AuNPs and MNPs (Wang K.-H. et al., 2024), but also be involved in the applications of biological nanosensors. In one study, researchers embedded α-hemolysin (α-HL) protein nanopores into an artificial phospholipid membrane to construct a nanopore-based biosensor. The narrowest region of the nanopore channel measures approximately 1.4 nm in diameter, allowing only ssDNA to pass through. The designed LNA probes partially hybridize with the wild-type sequence and fully complementary with the sequence containing a mutation site. When a specific voltage is applied to the sensor, the partially complementary DNA sequences rapidly dissociate due to reduced double-strand stability, and the dissociated ssDNA can pass through the nanopore within a short time. In contrast, the fully matched sequences require more time to pass (Figure 8C). By analyzing the current signals and translocation duration, the system can distinguish between wild-type and mutant sequences (Tian et al., 2018).
4.2.4 Limitations and future optimization of LNA probes
However, LNA remains more costly than conventional probes (Hansen et al., 2003; Pfundheller et al., 2005). The incorporation positions of LNA also require precise optimization (Laxton et al., 2011), and extensive LNA modifications may induce structural rigidity in the probes (Crinelli, 2002; Ferreira et al., 2021). Additionally, the potential toxicity risks associated with LNA probes warrant further investigation, as high concentrations of LNA-modified oligonucleotides might trigger immune responses and hepatotoxic effects (Romero-Palomo et al., 2021; Swayze et al., 2007). Researchers have developed LNA structure-toxicity prediction tools to assist in the optimization of LNA design and reduce the risk of toxicity (Burdick et al., 2014).
5 Pure nucleic acid probe-based detection
In recent years, an increasing number of studies have focused on point mutation detection by exploiting the intrinsic higher-order structures and dynamic intermolecular reactions of nucleic acids themselves (Lahiri et al., 2019; Zhang et al., 2023). This approach does not rely on exogenous components such as enzymes or nanomaterials, but rather achieves highly sensitive discrimination of single-base differences through conformational transitions of nucleic acids at the secondary or higher structural level.
Based on nucleic acid kinetics, Wu et al. proposed and validated a novel terminal self-competitive nucleic acid probe (TSCP). The entire probe consists of one long strand and two short inhibitory strands. The long strand contains complementary sequences to both the wild-type and mutant genes, as well as two distinct HCR initiators. In the absence of the target, the probe tightly binds to the inhibitory strands, thereby “locking” the HCR initiators. When the sample is introduced, the wild-type and mutant sequences replace their respective inhibitory strands and hybridize with the probe, simultaneously exposing the HCR initiators (Figure 9A). Subsequent fluorescence signals indicate the presence of the corresponding sequences, enabling accurate detection at concentrations as low as the nM level (Wu et al., 2025). However, in traditional competitive probe systems, it is often difficult to simultaneously achieve high sensitivity and specificity. The length and concentration of the blocking strand usually exhibit a negative correlation, making the optimization process cumbersome. Chen et al. proposed the 4-way Strand Exchange LEd Competitive DNA Testing (SELECT) system, which effectively addresses this issue by allowing the probe to bind to the target sequence and generate a signal without requiring complete dissociation from the inhibitory strand. This system successfully detected KRAS G12D and G13D mutations in ovarian cancer patient samples, with a detection limit of 0.1% (Chen et al., 2019). Conventional nucleic acid probes are generally designed to be long, since shorter probes tend to destabilize probe-target duplexes and cause transient dissociation. Interestingly, Su et al. turned this inherent limitation into an advantage by establishing a detection method that leverages the transient binding and dissociation of short DNA probes (Su et al., 2017). In high-salt conditions, short fluorescent probes are driven to associate with the target sequence, producing detectable fluorescence upon hybridization. However, even a single-base mismatch is strongly penalized under such high-salt environments. By monitoring the fluorescence duration with total internal reflection fluorescence microscopy (TIRFM), mutant and wild-type sequences can be effectively distinguished. A schematic diagram of this method is shown in Figure 9B. Such methods that differentiate molecules based on kinetic differences are sometimes referred to as “Kinetic Fingerprinting”.
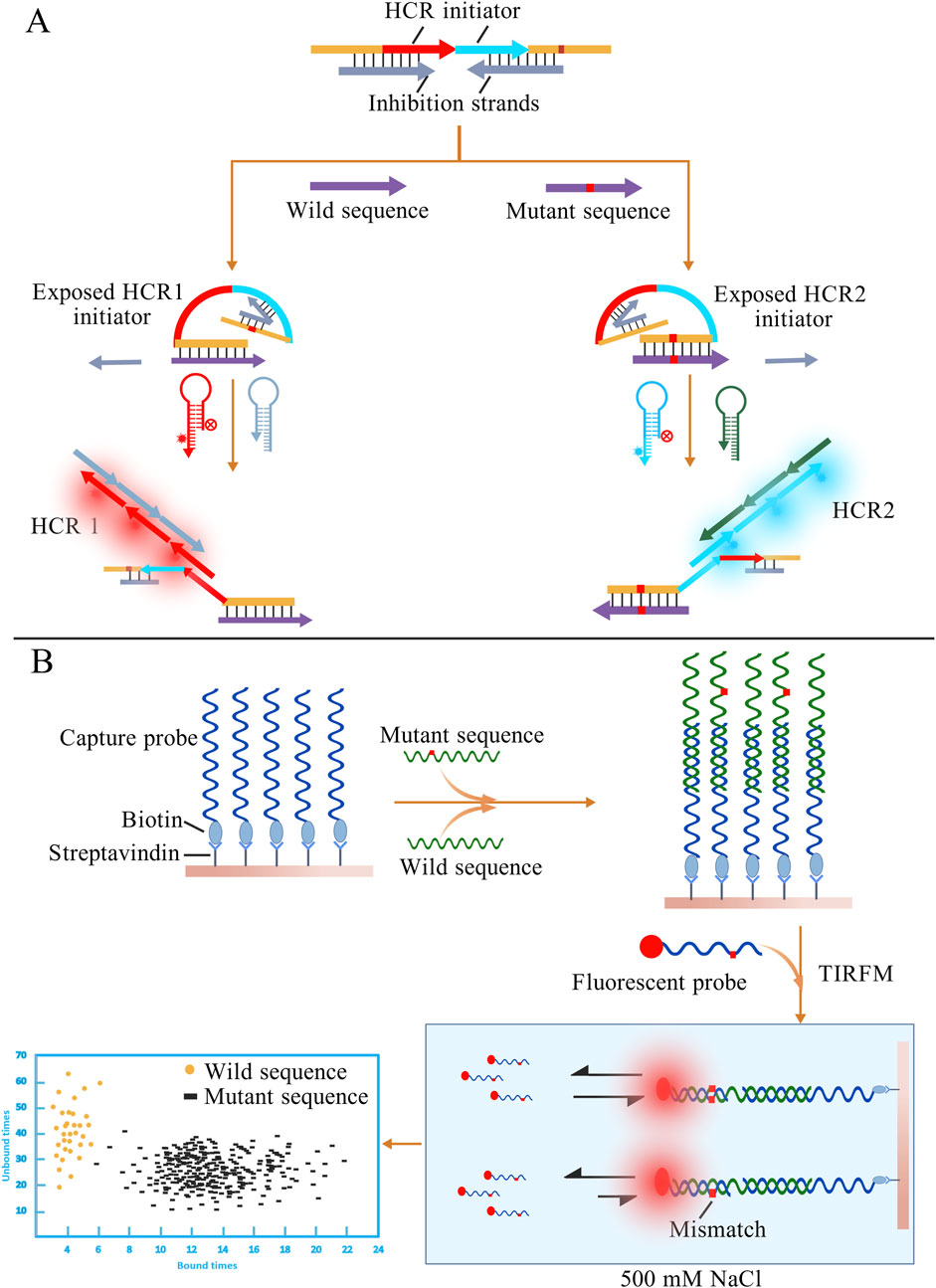
Figure 9. (A) Different target sequences displace the corresponding inhibitory strands, thereby exposing distinct HCR initiators. The presence of wild-type or mutant sequences can then be determined based on the resulting HCR fluorescence signals. (B) Excess capture probes first hybridize with the nucleic acid sequences in the sample. After adding fluorescent probes, fully matched sequences exhibit longer binding durations with the probes. The presence of mutant sequences is determined by observing the duration of fluorescent signals under TIRFM. The figures are created with BioGDP.com.
Beyond reliance on intrinsic nucleic acid kinetics, certain research groups have introduced chemical modifications (such as pyrene and 2′-O-methyl RNA) into probe design to enhance structural stability and detection sensitivity (Kholodar et al., 2013; Zhou et al., 2018). Meanwhile, other groups have sought to integrate nucleic acid probes with complementary analytical technologies. Zhang et al. combined catalytic hairpin assembly (CHA), FRET, and toehold-mediated strand displacement (TMSD) to develop a dual-base mismatch strategy. By introducing an extra mismatch, the system could distinguish WT from MT with high sensitivity, reaching a detection limit of 4.3 fM (Zhang et al., 2024).
In addition to the limitations mentioned above, pure nucleic acid probe-based detection also suffers from limited anti-interference capability and insufficient dynamic regulation, which lead to reduced detection efficiency in complex biological samples. However, with a deeper understanding of nucleic acid kinetics, energy differences, and free energy landscapes, it is believed that highly specific and selective enzyme-free nucleic acid probes will be designed in the future for the direct detection of point mutations.
6 From laboratory to clinic
Validation using clinical samples represents the next critical step for most nucleic acid probe-based detection methods, which will establish a solid foundation for their comprehensive advancement into clinical testing. However, the vast majority of the experimental strategies mentioned above remain confined to the laboratory stage, utilizing either genetically extracted materials from self-cultured cells or artificially synthesized nucleotide fragments. Substantial differences exist between real clinical samples and these idealized specimens, as complex background factors may significantly compromise detection efficiency. Thus, transitioning success in experimental models to success in clinical samples constitutes a major challenge for current nucleic acid probe-based detection technologies.
However, several studies have successfully achieved validation using real clinical samples. Examples include the NAVIGATER system proposed by Song et al. (2020), the AuNP-based colorimetric method employed by Bai et al. (2020), and so on (Table 2). Although the detection sensitivity was slightly lower than that observed with synthetic ssDNA, these approaches still achieved considerable detection limits. Nevertheless, critical issues must be addressed before large-scale clinical application and integration into clinical decision-making can be realized. These include the inherent stability of enzymes, the biotoxicity of nanomaterials, the water solubility of PNA probes, and challenges related to their delivery.
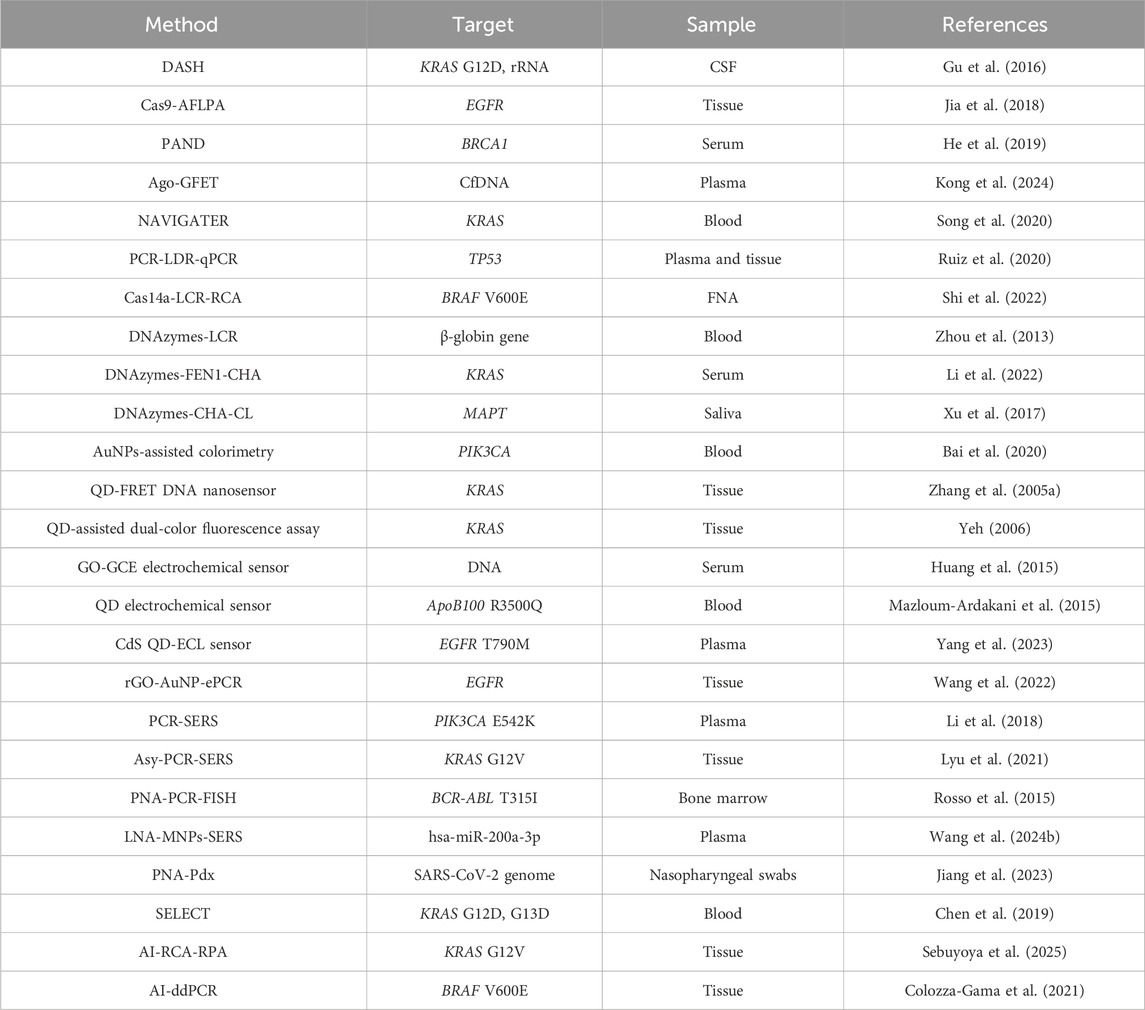
Table 2. A summary of technologies applicable to clinical sample detection.
Currently, nucleic acid probe-based methods that have been truly successfully applied to clinical testing are still predominantly PCR-based detection systems, such as fluorescent PCR and digital PCR (Ryncarz et al., 1999; Stamey et al., 2001). Commercial PCR kits targeting common mutation sites are readily available on the market and have achieved successful commercialization. For patients, this eliminates the need to endure lengthy and complex processes like Sanger sequencing or NGS. Moreover, compared to sequencing, precise PCR detection offers significantly reduced costs. The commercial success of PCR-based assays is also helping drive the development of more nucleic acid probe-based detection technologies, though several limitations remain. PCR-based methods require complicated thermal cycling procedures, during which enzymes are still indispensable. The development of simpler and faster approaches, ideally operable at room temperature, remains a key direction for future research. Nucleic acid probe-based detection also has a major advantage: once a simple and efficient detection strategy is established, it can be readily adapted to other rare sites simply by altering the probe sequence. Nevertheless, specific kinetics and GC content still need to be taken into consideration.
While nucleic acid probe-based approaches hold considerable promise at the experimental level, their transition toward large-scale commercialization and clinical adoption is constrained by stringent regulatory requirements. Chief among these is the need to rigorously establish clinical efficacy across heterogeneous populations, including variations in age, sex, and ethnicity. Addressing this demand entails large-scale clinical studies, which impose substantial temporal and economic burdens.
Although nucleic acid probe-based strategies have demonstrated great potential in the laboratory, their large-scale commercialization and clinical translation still face significant regulatory barriers. One key challenge is the requirement to demonstrate the clinical efficacy of the probes in the target population, ensuring reliable detection limits across patients of different ages, genders, and ethnic backgrounds. This necessitates large sample sizes and extensive clinical trials, which usually demand substantial time and financial resources. Furthermore, rigorous data must be provided to demonstrate the sensitivity and specificity of the strategy. What is the LOD of this method in patient cohorts known to carry the target point mutation compared to the gold standard, Can it reliably avoid false positives in healthy populations. In addition, it is essential to prove its ability to distinguish single-base differences without cross-reactivity, which requires testing with various potentially interfering mutant sequences. At the sample level, different types of specimens (blood, tissue, FFPE samples) may require distinct detection standards, necessitating additional validation data. Moreover, factors such as the type of collection tubes, storage temperature, and maximum storage duration must be standardized.
7 Integration of AI in nucleic acid probe-based mutation detection
Artificial intelligence (AI) has developed rapidly in recent years, covering fields such as medicine, engineering, and education, and has brought tremendous convenience to the world. In the medical domain, AI technology is widely used in disease diagnosis and precision medicine, demonstrating remarkable capabilities particularly in image recognition and genetic analysis. Although the integration of AI with nucleic acid probe-based detection is not yet mainstream, studies have shown that AI can effectively support such detection methods.
7.1 AI-assisted probe design
The design of nucleic acid probes involves potential secondary structures, which may lead to unexpected binding modes during hybridization with the target. Machine learning and AI technologies can predict these possible secondary structures. Combined with free energy analysis, they assist in designing more stable probes (Calonaci et al., 2020; Singh et al., 2019). Additionally, probe design must also consider factors such as complementarity to the target sequence and GC content. Deep learning-based AI approaches can further predict the binding efficiency between the probe and the target, facilitating the development of probes with higher specificity. Li et al. proposed a deep learning-based method named Deep DNAshape, which accurately accounts for the influence of extended flanking regions. It enables high-throughput prediction of DNA structural features for sequences of any length and quantity, thereby elucidating the impact of flanking regions on the structural properties of target DNA sequences (Li et al., 2024). This approach not only provides detailed structural analysis of probe-target binding but also offers valuable insights for the design of nucleic acid probes.
As early as 2009, studies had already embedded machine learning algorithms into thermodynamics-based primer design tools to accelerate the screening of efficient and specific primer pairs (Mann et al., 2009). In recent years, an increasing number of AI tools have been applied to the evaluation and optimization of primers and probes. For example, Dwivedi-Yu et al. introduced swga2.0, which combines active and machine learning to assess the amplification efficacy of individual primers and primer sets, enabling the design of primers for selective whole-genome amplification (Dwivedi-Yu et al., 2023). Similarly, Lin et al. developed a machine learning platform named BioInnovate AI. Compared to traditional tools such as Primer3 and Primer-BLAST, BioInnovate AI integrates key thermodynamic parameters to more accurately predict amplification success rates. By leveraging machine learning algorithms to predict the effectiveness of PCR primers and probes, the platform reduced PCR reagent development time by approximately 90%, significantly shortening the development timeline for diagnostic reagents targeting emerging infectious diseases (Lin et al., 2025).
7.2 AI-assisted signal analysis
The vast majority of nucleic acid probe-based detection methods rely on various forms of signal output, such as the fluorescence, electrochemical signals, and SERS signals mentioned above. However, when confronted with diverse and complex signals-such as background noise in fluorescence or intricate spectral profiles in SERS-manual interpretation or conventional algorithmic analysis tends to be time-consuming and susceptible to subjective bias. The development of AI models offers a promising new approach to signal interpretation. Through large-scale training, many AI-driven deep learning models can accurately decode complex signals, thereby improving both sensitivity and specificity (Zhou et al., 2024).
The optical filtering assembly, an essential component of fluorescence microscopy, helps suppress background noise while highlighting specific fluorescence signals. However, its use also increases the complexity of the fluorescence microscopy imaging system. To address this limitation, Dai et al. proposed a deep learning-based, filter-free fluorescence imaging method that leverages digital spectral filtering. This approach enables automatic selection of fluorescence channels and accurate prediction of fluorescence signals after image acquisition. Experimental results demonstrate that the method achieves excellent sensitivity and specificity across varying magnification levels (Dai et al., 2025). In addition, to address the problem of misjudgment caused by signal heterogeneity in conventional SERS detection, Zhao et al. developed an AI-driven SERS-LFIA automated detection system. This system innovatively integrates large-area SERS scanning imaging around the T-line with a deep learning model, enabling result interpretation based on the distribution patterns of nanoprobe signals rather than solely on signal intensity. The method achieved an accuracy of 94.52% in the test dataset (Zhao et al., 2025). Such examples of AI-based signal analysis pave the way for future advancements in nucleic acid probe signal interpretation.
7.3 Examples of AI-assisted mutation detection
With the rapid advancement of AI technology, an increasing number of models are being integrated into molecular diagnostics through deep learning applications in signal processing, pattern recognition, and predictive modeling. Although in most fields AI models still serve as auxiliary diagnostic tools, in several studies that have successfully incorporated AI into mutation detection, they play a significant role in assisting with result interpretation, signal analysis, and system optimization.
Sebuyoya and colleagues integrated an AI model into an electrochemical sensing platform employing RCA and recombinase polymerase amplification (RPA). This sensor is capable of detecting the KRAS G12V mutation at a frequency as low as 1% amidst a high background of wild-type sequences. Furthermore, when validated across seven cancer cell lines and 11 clinical samples from colorectal cancer patients, the results showed complete concordance with next-generation sequencing (NGS). The AI model enabled fully automated interpretation within this system and demonstrated the ability to perfectly distinguish complex cases-including samples with other mutations such as G12S and G13D intentionally introduced by researchers-a task that can be challenging for manual interpretation (Sebuyoya et al., 2025). Similarly, another research team employed a trained machine learning model to rapidly interpret results from CRISPR-Cas13-based lateral flow assays (LFA). As a widely used detection method, LFA offers advantages such as rapidity and low cost. However, visual interpretation by humans is prone to errors-especially when the test line is faint or under poor lighting conditions-potentially leading to false positives or false negatives. In contrast, the machine learning model can determine results within 0.2 s with an accuracy of 96.5%. More importantly, this high performance was consistently achieved across photos taken under diverse backgrounds (such as sofas, carpets, and beds), varying lighting conditions, and different smartphone angles and models (Xue et al., 2025).
Beyond conventional result interpretation, AI technology is also influencing other aspects of nucleic acid detection. In the AI-assisted droplet digital PCR (ddPCR) system developed by Colozza-Gama, a machine learning algorithm was employed to address a core challenge in ddPCR data analysis: automated droplet classification. Traditional methods often require manual classification by researchers, which introduces subjective variability between different operators. The optimized ddPCR platform ultimately achieved BRAF V600E detection comparable to the gold standard (pyrosequencing), and the correlation between automated classification results and the gold standard (R = 0.66) was significantly higher than that of manual classification (R = 0.55) (Colozza-Gama et al., 2021). This strategy not only offers a new approach to optimizing ddPCR, but also provides a foundation for integrating machine learning algorithms into ddPCR systems in the future. These advancements not only enhance the efficiency of nucleic acid probe-based detection in terms of time and economic cost but also elevate the overall standard of detection to a new level.
7.4 Challenges at the convergence of AI and laboratory
Although AI tools demonstrate significant potential in medical testing and disease diagnosis, their integration into laboratory settings and eventual application in clinical diagnostics face considerable challenges. The development of every AI model requires a large volume of high-quality samples. However, differences in instruments and environmental conditions across laboratories may lead to data inconsistencies. A well-trained model may exhibit suboptimal outcomes when applied in other locations. Additionally, many complex AI models suffer from the “black box” problem-lacking interpretable biological explanations-which hinders their adoption in clinical practice. This represents one of the major obstacles that current AI models must overcome. Moreover, for a mature AI tool to be deployed in clinical settings, it must undergo rigorous clinical trials to demonstrate its efficacy and robustness, and obtain approval from regulatory agencies such as the FDA in the United States. This is a time-consuming process. Given the rapid pace of AI development, a more advanced model may emerge before its predecessor even completes regulatory review. Finally, the development of AI systems demands substantial expertise in computer science and often requires a team of specialized professionals. For medical researchers and conventional laboratories, this presents a significant barrier to entry.
8 Summary and outlook
Mutations are closely associated with disease susceptibility, pathogenesis, and therapeutic outcomes. The detection of relevant point mutations is crucial for early disease prevention and diagnosis. In recent years, DNA point mutation detection has undergone rapid development, evolving from traditional methods such as Sanger sequencing and PCR to more advanced approaches like NGS and TGS, all of which have achieved remarkable success. However, these methods are often limited either by low throughput and suboptimal performance in identifying low-frequency mutations, or by high costs and complex data analysis. In the current global healthcare context, there is a growing demand for rapid and accurate screening of diseases at early stages. Therefore, there is an urgent need to develop detection technologies with higher sensitivity, enhanced specificity, and simplified signal readouts. This review summarizes the design strategies for point mutation detection using nucleic acid probes, with a focus on the applications of enzymatic reactions, nanotechnology, nucleic acid derivatives, and pure nucleic acid probes. Additionally, we discuss emerging AI-assisted detection technologies at the end of the article, The advantages and disadvantages of each approach are summarized in Table 3. A comparative analysis of each specific method is provided in Table 4.
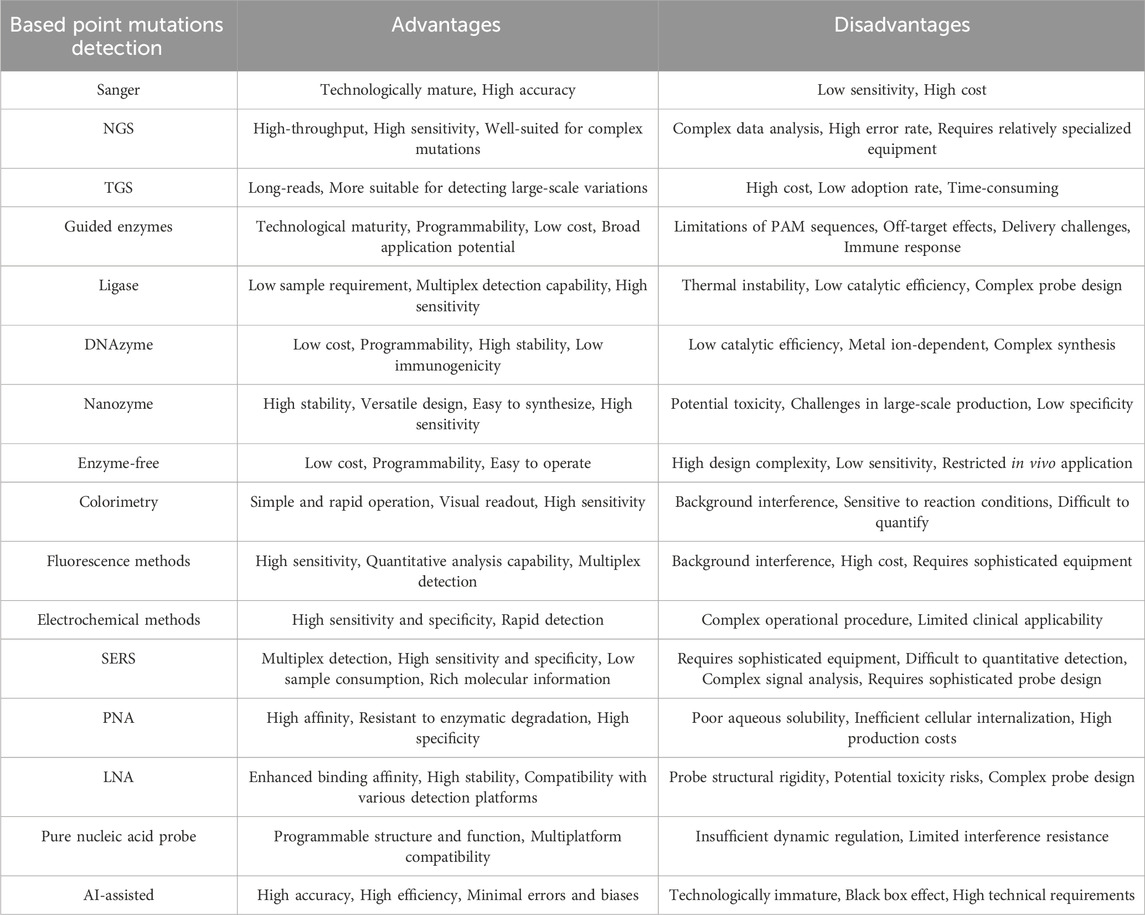
Table 3. Advantages and disadvantages of various point mutation detection techniques.
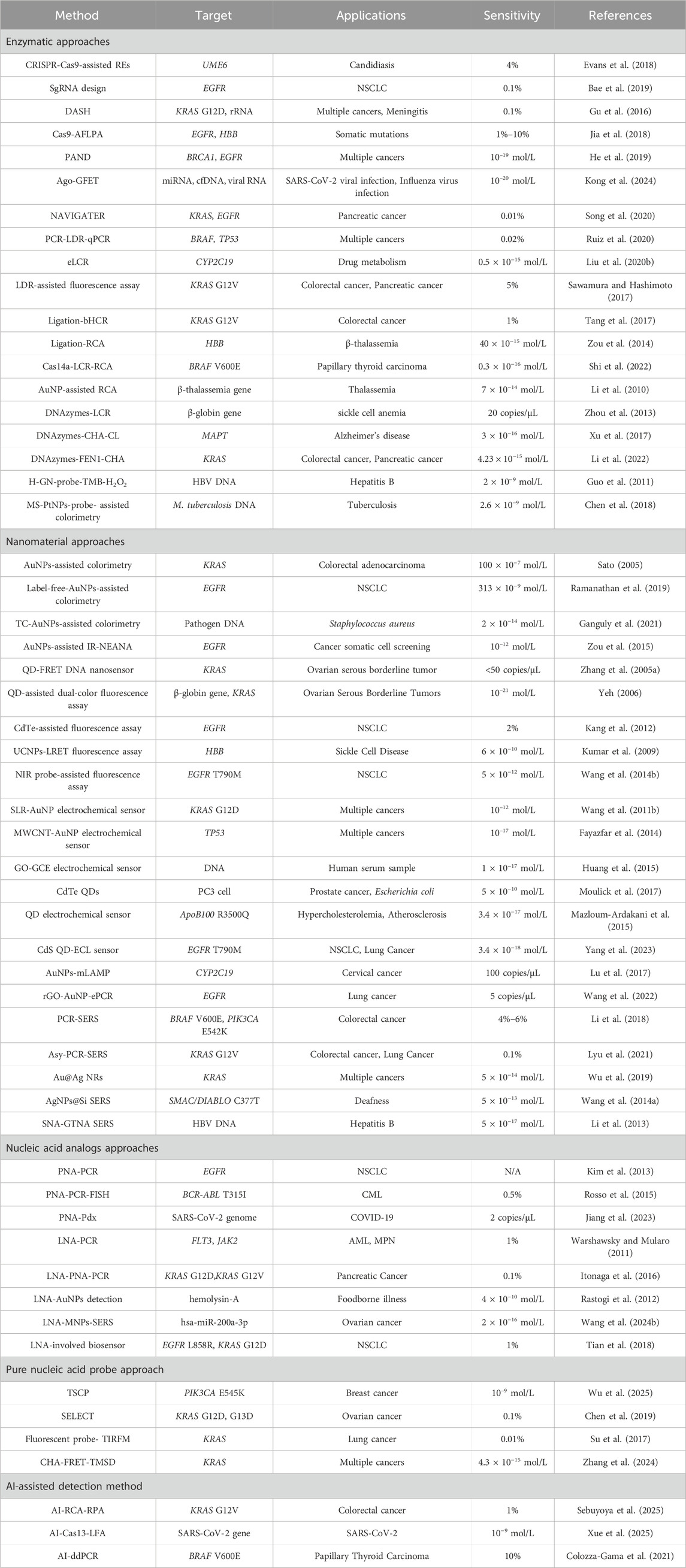
Table 4. Comparison of techniques for detecting gene mutations.
As sequencing technologies continue to mature, it is expected that numerous detection strategies compatible with TGS will be developed. Furthermore, with the rapid advancement of AI, many novel probe design concepts and analytical strategies are likely to emerge in the field of mutation detection. Additionally, the establishment of large-scale datasets and continuous deep learning will drive the development of more comprehensive and efficient models. These advances may help address unresolved biological problems and unpredictable kinetic processes. In the future, AI models specifically tailored for mutation detection-rather than merely serving auxiliary roles-could be developed and even integrated with clinical diagnostics, thereby driving progress in precision medicine.
Author contributions
XP: Conceptualization, Writing – original draft, Writing – review and editing. XW: Formal Analysis, Funding acquisition, Supervision, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by GuangDong Basic and Applied Basic Research Foundation (2023A1515220132), Applied Science and Technology Special Fund Project of Meizhou City (2021B0204002) and Scientific Research and Cultivation Project of Meizhou People’s Hospital (PY-C2021048).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adhikari, S. P., Vukelich, P., Guenther, D. C., Karmakar, S., and Hrdlicka, P. J. (2021). Recognition of double-stranded DNA using LNA-modified toehold Invader probes. Org. Biomol. Chem. 19, 9276–9290. doi:10.1039/D1OB01888D
Adhikari, S. P., Karmakar, S., and Hrdlicka, P. J. (2022). Nicked Invader probes: multistranded and sequence-unrestricted recognition of double-stranded DNA. Org. Biomol. Chem. 20, 1019–1030. doi:10.1039/D1OB02019F
Adorisio, R., Ciardiello, D., Rappa, A., Gervaso, L., Pelizzari, G., Marinucci, L., et al. (2025). Investigating the pathogenicity of uncommon KRAS mutations and their association with clinicopathologic characteristics in patients with colorectal cancer. J. Mol. Diagn 27, 130–138. doi:10.1016/j.jmoldx.2024.11.007
Aiba, Y., Honda, Y., and Komiyama, M. (2015). Promotion of double-duplex invasion of peptide nucleic acids through conjugation with nuclear localization signal peptide. Chem. Eur. J. 21, 4021–4026. doi:10.1002/chem.201406085
Almarsson, O., and Bruice, T. C. (1993). Peptide nucleic acid (PNA) conformation and polymorphism in PNA-DNA and PNA-RNA hybrids. Proc. Natl. Acad. Sci. U.S.A. 90, 9542–9546. doi:10.1073/pnas.90.20.9542
AlSafadi, A., Ramachandran, K., Columbus, S., Tlili, A., Daoudi, K., and Gaidi, M. (2024). Highly efficient, label free, ultrafast plasmonic SERS biosensor (silver nanoarrays/Si) to detect GJB2 gene expressed deafness mutations in real time validated with PCR studies. Int. J. Biol. Macromol. 259, 129381. doi:10.1016/j.ijbiomac.2024.129381
Bae, T., Kim, H., Kim, J. H., Kim, Y. J., Lee, S. H., Ham, B.-J., et al. (2019). Specificity assessment of CRISPR genome editing of oncogenic EGFR point mutation with single-base differences. Molecules 25, 52. doi:10.3390/molecules25010052
Bai, Y., Li, H., Xu, J., Huang, Y., Zhang, X., Weng, J., et al. (2020). Ultrasensitive colorimetric biosensor for BRCA1 mutation based on multiple signal amplification strategy. Biosens. Bioelectron. 166, 112424. doi:10.1016/j.bios.2020.112424
Bai, B., An, X., Qu, Q., Liu, X., Liu, Y., and Wei, L. (2024). The clinical features and prognostic implications of the co-mutated TP53 gene in advanced non-small cell lung cancer. Clin. Transl. Oncol. 26, 3236–3245. doi:10.1007/s12094-024-03533-1
Bentin, T., and Nielsen, P. E. (2003). Superior duplex DNA strand invasion by acridine conjugated peptide nucleic acids. J. Am. Chem. Soc. 125, 6378–6379. doi:10.1021/ja029936t
Brunelli, A., Cazzagon, V., Faraggiana, E., Bettiol, C., Picone, M., Marcomini, A., et al. (2024). An overview on dispersion procedures and testing methods for the ecotoxicity testing of nanomaterials in the marine environment. Sci. Total Environ. 921, 171132. doi:10.1016/j.scitotenv.2024.171132
Burdick, A. D., Sciabola, S., Mantena, S. R., Hollingshead, B. D., Stanton, R., Warneke, J. A., et al. (2014). Sequence motifs associated with hepatotoxicity of locked nucleic acid—modified antisense oligonucleotides. Nucleic Acids Res. 42, 4882–4891. doi:10.1093/nar/gku142
Calonaci, N., Jones, A., Cuturello, F., Sattler, M., and Bussi, G. (2020). Machine learning a model for RNA structure prediction. Nar. Genom. Bioinform. 2, lqaa090. doi:10.1093/nargab/lqaa090
Carter, T. E., Boulter, A., Existe, A., Romain, J. R., St Victor, J. Y., Mulligan, C. J., et al. (2015). Artemisinin resistance-associated polymorphisms at the K13-propeller locus are absent in Plasmodium falciparum isolates from Haiti. Am. J. Trop. Med. Hyg. 92, 552–554. doi:10.4269/ajtmh.14-0664
Chen, G., Tong, H., Gao, T., Chen, Y., and Li, G. (2014). Direct application of gold nanoparticles to one-pot electrochemical biosensors. Anal. Chim. Acta. 849, 1–6. doi:10.1016/j.aca.2014.08.011
Chen, W., Fang, X., Ye, X., Wang, X., and Kong, J. (2018). Colorimetric DNA assay by exploiting the DNA-controlled peroxidase mimicking activity of mesoporous silica loaded with platinum nanoparticles. Mikrochim. Acta 185, 544. doi:10.1007/s00604-018-3026-9
Chen, X., Liu, N., Liu, L., Chen, W., Chen, N., Lin, M., et al. (2019). Thermodynamics and kinetics guided probe design for uniformly sensitive and specific DNA hybridization without optimization. Nat. Commun. 10, 4675. doi:10.1038/s41467-019-12593-9
Chen, W.-T., Chiu, P.-Y., and Chen, C.-F. (2023). A flash signal amplification approach for ultrasensitive and rapid detection of single nucleotide polymorphisms in tuberculosis. Biosens. Bioelectron. 237, 115514. doi:10.1016/j.bios.2023.115514
Chen, L., Zeng, X., Yang, F., Yang, T., Chen, Y., Zhao, Y., et al. (2025). Rapid and sensitive detection of mutations in SARS-CoV-2 by surface-enhanced Raman spectroscopy. Langmuir 41, 3583–3591. doi:10.1021/acs.langmuir.4c04725
Cheng, Y., Zhao, J., Jia, H., Yuan, Z., and Li, Z. (2013). Ligase chain reaction coupled with rolling circle amplification for high sensitivity detection of single nucleotide polymorphisms. Analyst 138, 2958. doi:10.1039/c3an36920j
Chhetri, K. B., Sharma, A., Naskar, S., and Maiti, P. K. (2022). Nanoscale structures and mechanics of peptide nucleic acids. Nanoscale 14, 6620–6635. doi:10.1039/D1NR04239D
Cho, M., Han, M. S., and Ban, C. (2008). Detection of mismatched DNAs via the binding affinity of MutS using a gold nanoparticle-based competitive colorimetric method. Chem. Commun. 4573, 4573–4575. doi:10.1039/b811346g
Chou, W.-C., Hu, W.-P., Yang, Y.-S., Chan, H. W.-H., and Chen, W.-Y. (2019). Neutralized chimeric DNA probe for the improvement of GC-rich RNA detection specificity on the nanowire field-effect transistor. Sci. Rep. 9, 11056. doi:10.1038/s41598-019-47522-9
Colozza-Gama, G. A., Callegari, F., Bešič, N., Paniza, A. C. de J., and Cerutti, J. M. (2021). Machine learning algorithm improved automated droplet classification of ddPCR for detection of BRAF V600E in paraffin-embedded samples. Sci. Rep. 11, 12648. doi:10.1038/s41598-021-92014-4
Crinelli, R. (2002). Design and characterization of decoy oligonucleotides containing locked nucleic acids. Nucleic Acids Res. 30, 2435–2443. doi:10.1093/nar/30.11.2435
Crudele, J. M., and Chamberlain, J. S. (2018). Cas9 immunity creates challenges for CRISPR gene editing therapies. Nat. Commun. 9, 3497. doi:10.1038/s41467-018-05843-9
Dai, B., You, S., Wang, K., Long, Y., Chen, J., Upreti, N., et al. (2025). Deep learning-enabled filter-free fluorescence microscope. Sci. Adv. 11, eadq2494. doi:10.1126/sciadv.adq2494
De Silva Indrasekara, A. S., and Fabris, L. (2015). Sers-based approaches toward genetic profiling. Bioanalysis 7, 263–278. doi:10.4155/bio.14.295
Del Bene, A., D’Aniello, A., Tomassi, S., Merlino, F., Mazzarella, V., Russo, R., et al. (2023). Ultrasound-assisted peptide nucleic acids synthesis (US-pnas). Ultrason. Sonochem. 95, 106360. doi:10.1016/j.ultsonch.2023.106360
Dikmen, G. (2023). Surface enhanced Raman spectroscopy sensor based on silver nanoparticles/multi wall carbon nanotubes for ultrasensitive detection of cholesterol. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 303, 123235. doi:10.1016/j.saa.2023.123235
Dwivedi-Yu, J. A., Oppler, Z. J., Mitchell, M. W., Song, Y. S., and Brisson, D. (2023). A fast machine-learning-guided primer design pipeline for selective whole genome amplification. PLoS Comput. Biol. 19, e1010137. doi:10.1371/journal.pcbi.1010137
Egli, M., Teplova, M., Minasov, G., Kumar, R., and Wengel, J. (2001). X-ray crystal structure of a locked nucleic acid (LNA) duplex composed of a palindromic 10-mer DNA strand containing one LNA thymine monomer. Chem. Commun., 651–652. doi:10.1039/b009447l
Elghanian, R., Storhoff, J. J., Mucic, R. C., Letsinger, R. L., and Mirkin, C. A. (1997). Selective colorimetric detection of polynucleotides based on the distance-dependent optical properties of gold nanoparticles. Science 277, 1078–1081. doi:10.1126/science.277.5329.1078
Eriksson, M., Ambroise, G., Ouchida, A. T., Lima Queiroz, A., Smith, D., Gimenez-Cassina, A., et al. (2017). Effect of mutant p53 proteins on glycolysis and mitochondrial metabolism. Mol. Cell. Biol. 37, e00328-17. doi:10.1128/MCB.00328-17
Evans, B. A., Smith, O. L., Pickerill, E. S., York, M. K., Buenconsejo, K. J. P., Chambers, A. E., et al. (2018). Restriction digest screening facilitates efficient detection of site-directed mutations introduced by CRISPR in C. albicans UME6. PeerJ 6, e4920. doi:10.7717/peerj.4920
Fan, Y., and Xu, H. (2021). Prediction of off-target effects in CRISPR/Cas9 system by ensemble learning. CBIO 16, 1169–1178. doi:10.2174/1574893616666210811100938
Farahmandzadeh, F., Molahosseini, E., Portakaloo, S. N., Molaei, M., and Khanzadeh, M. (2024). Efficient gamma ray detection using CdTe/CdS core/shell quantum dots: a simple and rapid fluorescence approach. J. Fluoresc. 35, 1539–1547. doi:10.1007/s10895-024-03630-y
Fayazfar, H., Afshar, A., Dolati, M., and Dolati, A. (2014). DNA impedance biosensor for detection of cancer, TP53 gene mutation, based on gold nanoparticles/aligned carbon nanotubes modified electrode. Anal. Chim. Acta. 836, 34–44. doi:10.1016/j.aca.2014.05.029
Ferreira, I., Slott, S., Astakhova, K., and Weber, G. (2021). Complete mesoscopic parameterization of single LNA modifications in DNA applied to oncogene probe design. J. Chem. Inf. Model. 61, 3615–3624. doi:10.1021/acs.jcim.1c00470
Finn, J. D., Smith, A. R., Patel, M. C., Shaw, L., Youniss, M. R., van Heteren, J., et al. (2018). A single administration of CRISPR/Cas9 lipid nanoparticles achieves robust and persistent in vivo genome editing. Cell Rep. 22, 2227–2235. doi:10.1016/j.celrep.2018.02.014
Fouz, M. F., and Appella, D. H. (2020). PNA clamping in nucleic acid amplification protocols to detect single nucleotide mutations related to cancer. Molecules 25, 786. doi:10.3390/molecules25040786
Ganguly, K., Patel, D. K., Dutta, S. D., and Lim, K.-T. (2021). TEMPO-cellulose nanocrystal-capped gold nanoparticles for colorimetric detection of pathogenic DNA. ACS Omega 6, 12424–12431. doi:10.1021/acsomega.1c00359
Gasparello, J., Manicardi, A., Casnati, A., Corradini, R., Gambari, R., Finotti, A., et al. (2019). Efficient cell penetration and delivery of peptide nucleic acids by an argininocalix[4]arene. Sci. Rep. 9, 3036. doi:10.1038/s41598-019-39211-4
Gu, W., Crawford, E. D., O’Donovan, B. D., Wilson, M. R., Chow, E. D., Retallack, H., et al. (2016). Depletion of Abundant Sequences by Hybridization (DASH): using Cas9 to remove unwanted high-abundance species in sequencing libraries and molecular counting applications. Genome Biol. 17, 41. doi:10.1186/s13059-016-0904-5
Gu, T., Wang, Y., Lu, Y., Cheng, L., Feng, L., Zhang, H., et al. (2019). Platinum nanoparticles to enable electrodynamic therapy for effective cancer treatment. Adv. Mater. 31, 1806803. doi:10.1002/adma.201806803
Guo, Y., Deng, L., Li, J., Guo, S., Wang, E., and Dong, S. (2011). Hemin-graphene hybrid nanosheets with intrinsic peroxidase-like activity for label-free colorimetric detection of single-nucleotide polymorphism. ACS Nano 5, 1282–1290. doi:10.1021/nn1029586
Guo, C., Ma, X., Gao, F., and Guo, Y. (2023). Off-target effects in CRISPR/Cas9 gene editing. Front. Bioeng. Biotechnol. 11, 1143157. doi:10.3389/fbioe.2023.1143157
Hamed, E. M., He, L., Rai, V., Hu, S., and Li, S. F. Y. (2024). Copper single-atom nanozyme mimicking galactose oxidase with superior catalytic activity and selectivity. Small 20, e2405986. doi:10.1002/smll.202405986
Hansen, H. F., Olsen, O., and Koch, T. (2003). New standards in LNA synthesis. Nucleos. Nucleotid. Nucleic Acids 22, 1273–1275. doi:10.1081/NCN-120022944
Hashimoto, D., Arima, K., Yokoyama, N., Chikamoto, A., Taki, K., Inoue, R., et al. (2016). Heterogeneity of KRAS mutations in pancreatic ductal adenocarcinoma. Pancreas 45, 1111–1114. doi:10.1097/MPA.0000000000000624
He, R., Wang, L., Wang, F., Li, W., Liu, Y., Li, A., et al. (2019). Pyrococcus furiosus Argonaute-mediated nucleic acid detection. Chem. Commun. 55, 13219–13222. doi:10.1039/C9CC07339F
He, M., Jin, L., Wang, F., Wang, X., You, Y., and He, H. (2024). Simple, ultrasensitive detection of superoxide anion radical mutations in melanoma mice with SERS microneedles. Spectrochim. Acta A Mol. Biomol. Spectrosc. 316, 124292. doi:10.1016/j.saa.2024.124292
Hibino, M., Aiba, Y., and Shoji, O. (2020). Cationic guanine: positively charged nucleobase with improved DNA affinity inhibits self-duplex formation. Chem. Commun. 56, 2546–2549. doi:10.1039/D0CC00169D
Ho, P., Huang, M.-Z., Fwu, V. T., Lin, S.-C., Hsiao, K.-J., and Su, T.-S. (2008). Simultaneous assessment of the effects of exonic mutations on RNA splicing and protein functions. Biochem. Biophys. Res. Commun. 373, 515–520. doi:10.1016/j.bbrc.2008.06.072
Hosseini, M., Mohammadi, S., Borghei, Y.-S., and Ganjali, M. R. (2017). Detection of p53 gene mutation (Single-Base mismatch) using a fluorescent silver nanoclusters. J. Fluoresc. 27, 1443–1448. doi:10.1007/s10895-017-2083-5
Hu, R., Wu, Z.-S., Zhang, S., Shen, G.-L., and Yu, R. (2011). Robust electrochemical system for screening single nucleotide polymorphisms. Chem. Commun. 47, 1294–1296. doi:10.1039/C0CC03884A
Hu, S., Huang, P.-J. J., Wang, J., and Liu, J. (2020). Dissecting the effect of salt for more sensitive label-free colorimetric detection of DNA using gold nanoparticles. Anal. Chem. 92, 13354–13360. doi:10.1021/acs.analchem.0c02688
Huang, X., Huang, X., Zhang, A., Zhuo, B., Lu, F., Chen, Y., et al. (2015). Quenching of the electrochemiluminescence of RU-complex tagged shared-stem hairpin probes by graphene oxide and its application to quantitative turn-on detection of DNA. Biosens. Bioelectron. 70, 441–446. doi:10.1016/j.bios.2015.03.072
Hwang, K. Y., Brown, D., Attanayake, S. B., Luu, D., Nguyen, M. D., Lee, T. R., et al. (2025). Signal differentiation of moving magnetic nanoparticles for enhanced biodetection and diagnostics. Biosensors 15, 116. doi:10.3390/bios15020116
Ishige, T., Itoga, S., Matsushita, K., and Nomura, F. (2016). Locked nucleic acid probe enhances Sanger sequencing sensitivity and improves diagnostic accuracy of high-resolution melting-based KRAS mutational analysis. Clin. Chim. Acta 457, 75–80. doi:10.1016/j.cca.2016.04.005
Itonaga, M., Matsuzaki, I., Warigaya, K., Tamura, T., Shimizu, Y., Fujimoto, M., et al. (2016). Novel methodology for rapid detection of KRAS mutation using PNA-LNA mediated loop-mediated isothermal amplification. PLoS ONE 11, e0151654. doi:10.1371/journal.pone.0151654
Izumi, H., Ishimoto, T., Yamamoto, H., Nishijo, H., and Mori, H. (2011). Bioluminescence imaging of Arc expression enables detection of activity-dependent and plastic changes in the visual cortex of adult mice. Brain Struct. Funct. 216, 91–104. doi:10.1007/s00429-010-0297-2
Jessop, I. A., Pérez, Y. P., Jachura, A., Nuñez, H., Saldías, C., Isaacs, M., et al. (2021). New hybrid copper nanoparticles/conjugated polyelectrolyte composite with antibacterial activity. Polymers 13, 401. doi:10.3390/polym13030401
Jia, C., Huai, C., Ding, J., Hu, L., Su, B., Chen, H., et al. (2018). New applications of CRISPR/Cas9 system on mutant DNA detection. Gene 641, 55–62. doi:10.1016/j.gene.2017.10.023
Jiang, W., Aman, R., Ali, Z., Rao, G. S., and Mahfouz, M. (2023). PNA-pdx: versatile peptide nucleic acid-based detection of nucleic acids and SNPs. Anal. Chem. 95, 14209–14218. doi:10.1021/acs.analchem.3c01809
Jiang, S., Li, H., Zhang, L., Mu, W., Zhang, Y., Chen, T., et al. (2025). Generic Diagramming Platform (GDP): a comprehensive database of high-quality biomedical graphics. Nucleic Acids Res. 53, D1670–D1676. doi:10.1093/nar/gkae973
Kallmyer, N. E., Rider, N. E., and Reuel, N. F. (2020). Design and validation of a frugal, automated, solid-phase peptide synthesizer. PLoS ONE 15, e0237473. doi:10.1371/journal.pone.0237473
Kang, T., Lee, H., Choe, D., Joo, S.-W., Lee, S. Y., Yoon, K.-A., et al. (2012). Simultaneous detection of multiple mutations in epidermal growth factor receptor based on fluorescence quenching of quantum dots. Biosens. Bioelectron. 31, 558–561. doi:10.1016/j.bios.2011.10.025
Kenward, M., and Dorfman, K. D. (2009). Coarse-grained brownian dynamics simulations of the 10-23 DNAzyme. Biophys. J. 97, 2785–2793. doi:10.1016/j.bpj.2009.09.003
Kholodar, S. A., Novopashina, D. S., Meschaninova, M. I., and Venyaminova, A. G. (2013). Multipyrene tandem probes for point mutations detection in DNA. J. Nucleic Acids 2013, 1–12. doi:10.1155/2013/860457
Kim, S., and Gang, J. (2018). The detection of a mismatched DNA by using hairpin DNA-templated silver nanoclusters. Anal. Biochem. 549, 171–173. doi:10.1016/j.ab.2018.03.026
Kim, T., Choe, D., Joo, S.-W., Lee, S. Y., and Lee, K. (2010). Mathematical modeling of DNA-mediated selective aggregation of CdS quantum dots. J. Colloid Interface Sci. 347, 209–214. doi:10.1016/j.jcis.2010.03.057
Kim, H.-R., Lee, S. Y., Hyun, D.-S., Lee, M. K., Lee, H.-K., Choi, C.-M., et al. (2013). Detection of EGFR mutations in circulating free DNA by PNA-mediated PCR clamping. J. Exp. Clin. Cancer Res. 32, 50. doi:10.1186/1756-9966-32-50
Kim, D., Bae, S., Park, J., Kim, E., Kim, S., Yu, H. R., et al. (2015). Digenome-seq: genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nat. Methods 12, 237–243. doi:10.1038/nmeth.3284
Kim, N., Cho, D., Kim, H., Kim, S., Cha, Y., Greulich, H., et al. (2020). Colorectal adenocarcinoma-derived EGFR mutants are oncogenic and sensitive to EGFR-targeted monoclonal antibodies, cetuximab and panitumumab. Int. J. Cancer 146, 2194–2200. doi:10.1002/ijc.32499
Kimberland, M. L., Hou, W., Alfonso-Pecchio, A., Wilson, S., Rao, Y., Zhang, S., et al. (2018). Strategies for controlling CRISPR/Cas9 off-target effects and biological variations in mammalian genome editing experiments. J. Biotechnol. 284, 91–101. doi:10.1016/j.jbiotec.2018.08.007
Kong, D., Zhang, S., Guo, M., Li, S., Wang, Q., Gou, J., et al. (2024). Ultra-Fast single-nucleotide-variation detection enabled by argonaute-mediated transistor platform. Adv. Mater. 36, 2307366. doi:10.1002/adma.202307366
Kumar, M., Guo, Y., and Zhang, P. (2009). Highly sensitive and selective oligonucleotide sensor for sickle cell disease gene using photon upconverting nanoparticles. Biosens. Bioelectron. 24, 1522–1526. doi:10.1016/j.bios.2008.08.023
Lahiri, H., Banerjee, S., and Mukhopadhyay, R. (2019). Free-energy-based gene mutation detection using LNA probes. ACS Sens. 4, 2688–2696. doi:10.1021/acssensors.9b01115
Laxton, C., Brady, K., Moschos, S., Turnpenny, P., Rawal, J., Pryde, D. C., et al. (2011). Selection, optimization, and pharmacokinetic properties of a novel, potent antiviral locked nucleic acid-based antisense oligomer targeting hepatitis C virus internal ribosome entry site. Antimicrob. Agents Chemother. 55, 3105–3114. doi:10.1128/AAC.00222-11
Lee, E. H., Jeong, M., Park, K., Lee, D. G., Lee, E. J., Lee, H., et al. (2024). Detection of miR-133a-5p using a molecular beacon probe for investigating postmortem intervals. Noncoding RNA 10, 58. doi:10.3390/ncrna10060058
Legut, M., Daniloski, Z., Xue, X., McKenzie, D., Guo, X., Wessels, H.-H., et al. (2020). High-throughput screens of PAM-flexible Cas9 variants for gene knockout and transcriptional modulation. Cell Rep. 30, 2859–2868.e5. doi:10.1016/j.celrep.2020.02.010
Li, J., Deng, T., Chu, X., Yang, R., Jiang, J., Shen, G., et al. (2010). Rolling circle amplification combined with gold nanoparticle aggregates for highly sensitive identification of single-nucleotide polymorphisms. Anal. Chem. 82, 2811–2816. doi:10.1021/ac100336n
Li, M., Cushing, S. K., Liang, H., Suri, S., Ma, D., and Wu, N. (2013). Plasmonic nanorice antenna on triangle nanoarray for surface-enhanced Raman scattering detection of hepatitis B virus DNA. Anal. Chem. 85, 2072–2078. doi:10.1021/ac303387a
Li, X., Yang, T., Li, C. S., Wang, D., Song, Y., and Jin, L. (2017). Detection of EGFR mutation in plasma using multiplex allele-specific PCR (MAS-PCR) and surface enhanced Raman spectroscopy. Sci. Rep. 7, 4771. doi:10.1038/s41598-017-05050-4
Li, X., Yang, T., Li, C. S., Song, Y., Lou, H., Guan, D., et al. (2018). Surface enhanced Raman spectroscopy (SERS) for the multiplex detection of braf, kras, and Pik3ca mutations in plasma of colorectal cancer patients. Theranostics 8, 1678–1689. doi:10.7150/thno.22502
Li, H., Huang, T., Wang, Y., Pan, B., Zhang, L., Zhang, Q., et al. (2020). Toxicity of alumina nanoparticles in the immune system of mice. Nanomedicine (Lond) 15, 927–946. doi:10.2217/nnm-2020-0009
Li, X., Liao, L., Jiang, B., Yuan, R., and Xiang, Y. (2022). Invader assay-induced catalytic assembly of multi-DNAzyme junctions for sensitive detection of single nucleotide polymorphisms. Anal. Chim. Acta 1224, 340225. doi:10.1016/j.aca.2022.340225
Li, J., Chiu, T.-P., and Rohs, R. (2024). Predicting DNA structure using a deep learning method. Nat. Commun. 15, 1243. doi:10.1038/s41467-024-45191-5
Li, J., Jiang, L., Wu, H., Zou, Y., Zhu, S., Huang, Y., et al. (2025). Self-contained G-quadruplex/hemin DNAzyme: a superior ready-made catalyst for in situ imaging analysis. Nucleic Acids Res. 53, gkaf227. doi:10.1093/nar/gkaf227
Liang, M., and Yan, X. (2019). Nanozymes: from new concepts, mechanisms, and standards to applications. Acc. Chem. Res. 52, 2190–2200. doi:10.1021/acs.accounts.9b00140
Liang, Z., Kang, M., Payne, G. F., Wang, X., and Sun, R. (2016). Probing energy and electron transfer mechanisms in fluorescence quenching of biomass carbon quantum dots. ACS Appl. Mater. Interfaces 8, 17478–17488. doi:10.1021/acsami.6b04826
Lin, L., Liu, A., Zhao, C., Weng, S., Lei, Y., Liu, Q., et al. (2013a). A chronocoulometric LNA sensor for amplified detection of K-ras mutation based on site-specific DNA cleavage of restriction endonuclease. Biosens. Bioelectron. 42, 409–414. doi:10.1016/j.bios.2012.09.063
Lin, L., Weng, S., Zhao, C., Liu, Q., Liu, A., and Lin, X. (2013b). Hairpin LNA biosensor with enzyme tagged AuNPs as tracer for amperometric detection of K-ras mutation gene. Electrochim. Acta 108, 808–813. doi:10.1016/j.electacta.2013.07.042
Lin, H.-H., Chung, H.-Y., Lin, T.-H., Chang, C.-K., Perng, C.-L., Hung, K.-S., et al. (2025). BioInnovate AI: a machine learning platform for rapid PCR assay design in emerging infectious disease diagnostics. Diagn. (Basel) 15, 1445. doi:10.3390/diagnostics15121445
Liu, Y., Cheng, Y., Zhang, H., Zhou, M., Yu, Y., Lin, S., et al. (2020a). Integrated cascade nanozyme catalyzes in vivo ROS scavenging for anti-inflammatory therapy. Sci. Adv. 6, eabb2695. doi:10.1126/sciadv.abb2695
Liu, Z.-J., Yang, L.-Y., Wei, Q.-X., Ye, C.-L., Xu, X.-W., Zhong, G.-X., et al. (2020b). A novel ligase chain reaction-based electrochemical biosensing strategy for highly sensitive point mutation detection from human whole blood. Talanta 216, 120966. doi:10.1016/j.talanta.2020.120966
Liu, C., Zheng, X., Dai, T., Wang, H., Chen, X., Chen, B., et al. (2022). Reversibly photoswitching upconversion nanoparticles for super-sensitive photoacoustic molecular imaging. Angew. Chem. Int. Ed. 61, e202116802. doi:10.1002/anie.202116802
Liu, C., Zhao, X., Wang, Z., Zhao, Y., Li, R., Chen, X., et al. (2023). Metal-organic framework-modulated Fe3O4 composite au nanoparticles for antibacterial wound healing via synergistic peroxidase-like nanozymatic catalysis. J. Nanobiotechnol. 21, 427. doi:10.1186/s12951-023-02186-6
Liu, Y., Cheng, C., Zhao, Z., Liu, W., and Qi, L. (2024). MOF-polymer composites with well-distributed gold nanoparticles for visual monitoring of homocysteine. Analyst 149, 1658–1664. doi:10.1039/d3an01934a
Lohman, G. J. S., Zhang, Y., Zhelkovsky, A. M., Cantor, E. J., and Evans, T. C. (2014). Efficient DNA ligation in DNA-RNA hybrid helices by Chlorella virus DNA ligase. Nucleic Acids Res. 42, 1831–1844. doi:10.1093/nar/gkt1032
Lu, Y., Ma, X., Wang, J., Sheng, N., Dong, T., Song, Q., et al. (2017). Visualized detection of single-base difference in multiplexed loop-mediated isothermal amplification amplicons by invasive reaction coupled with oligonucleotide probe-modified gold nanoparticles. Biosens. Bioelectron. 90, 388–393. doi:10.1016/j.bios.2016.12.015
Lundin, K. E., Hasan, M., Moreno, P. M., Törnquist, E., Oprea, I., Svahn, M. G., et al. (2005). Increased stability and specificity through combined hybridization of peptide nucleic acid (PNA) and locked nucleic acid (LNA) to supercoiled plasmids for PNA-anchored “Bioplex” formation. Biomol. Eng. 22, 185–192. doi:10.1016/j.bioeng.2005.07.003
Lyu, N., Rajendran, V. K., Li, J., Engel, A., Molloy, M. P., and Wang, Y. (2021). Highly specific detection of KRAS single nucleotide polymorphism by asymmetric PCR/SERS assay. Analyst 146, 5714–5721. doi:10.1039/D1AN01108A
Lyu, N., Hassanzadeh-Barforoushi, A., Rey Gomez, L. M., Zhang, W., and Wang, Y. (2024). SERS biosensors for liquid biopsy towards cancer diagnosis by detection of various circulating biomarkers: current progress and perspectives. Nano Converg. 11, 22. doi:10.1186/s40580-024-00428-3
Ma, Q., and Gao, Z. (2018). A simple and ultrasensitive fluorescence assay for single-nucleotide polymorphism. Anal. Bioanal. Chem. 410, 3093–3100. doi:10.1007/s00216-018-0874-4
Mann, T., Humbert, R., Dorschner, M., Stamatoyannopoulos, J., and Noble, W. S. (2009). A thermodynamic approach to PCR primer design. Nucleic Acids Res. 37, e95. doi:10.1093/nar/gkp443
Mao, L., Zhao, W., Li, X., Zhang, S., Zhou, C., Zhou, D., et al. (2021). Mutation spectrum of EGFR from 21,324 Chinese patients with non-small cell lung cancer (NSCLC) successfully tested by multiple methods in a CAP-accredited laboratory. Pathol. Oncol. Res. 27, 602726. doi:10.3389/pore.2021.602726
Martin, M. N., Allen, A. J., MacCuspie, R. I., and Hackley, V. A. (2014). Dissolution, agglomerate morphology, and stability limits of protein-coated silver nanoparticles. Langmuir 30, 11442–11452. doi:10.1021/la502973z
Mazloum-Ardakani, M., Aghaei, R., and Heidari, M. M. (2015). Quantum-dot biosensor for hybridization and detection of R3500Q mutation of apolipoprotein B-100 gene. Biosens. Bioelectron. 72, 362–369. doi:10.1016/j.bios.2015.05.014
McCall, J., Alexander, C., and Richter, M. M. (1999). Quenching of electrogenerated chemiluminescence by phenols, hydroquinones, catechols, and benzoquinones. Anal. Chem. 71, 2523–2527. doi:10.1021/ac981322c
McCubrey, J. A., Abrams, S. L., Steelman, L. S., Cocco, L., Ratti, S., Martelli, A. M., et al. (2022). APR-246—the mutant TP53 reactivator—increases the effectiveness of berberine and modified berberines to inhibit the proliferation of pancreatic cancer cells. Biomolecules 12, 276. doi:10.3390/biom12020276
Mehrgardi, M. A., and Ahangar, L. E. (2011). Silver nanoparticles as redox reporters for the amplified electrochemical detection of the single base mismatches. Biosens. Bioelectron. 26, 4308–4313. doi:10.1016/j.bios.2011.04.020
Menard, D., and Dondorp, A. (2017). Antimalarial Drug resistance: a Threat to Malaria elimination. Cold Spring Harb. Perspect. Med. 7, a025619. doi:10.1101/cshperspect.a025619
Mishra, S., Ghosh, S., and Mukhopadhyay, R. (2013). Maximizing mismatch discrimination by surface-Tethered locked nucleic acid probes via ionic tuning. Anal. Chem. 85, 1615–1623. doi:10.1021/ac3028382
Mohan, M., and Poddar, R. (2021). Polymerically engineered upconversion nanoparticles (UCNPs) as contrast agent for functionally modified optical coherence tomography (OCT). Mater. Sci. Eng. C 121, 111841. doi:10.1016/j.msec.2020.111841
Moulick, A., Milosavljevic, V., Vlachova, J., Podgajny, R., Hynek, D., Kopel, P., et al. (2017). Using CdTe/ZnSe core/shell quantum dots to detect DNA and damage to DNA. IJN 12, 1277–1291. doi:10.2147/IJN.S121840
Muti, M., Kuralay, F., Erdem, A., Abaci, S., Yumak, T., and Sinağ, A. (2010). Tin oxide nanoparticles-polymer modified single-use sensors for electrochemical monitoring of label-free DNA hybridization. Talanta 82, 1680–1686. doi:10.1016/j.talanta.2010.07.040
Nandhini, K. P., Noki, S., Brasil, E., Albericio, F., and De La Torre, B. G. (2023). A safety-catch protecting group strategy compatible with Boc-chemistry for the synthesis of peptide nucleic acids (PNAs). Org. Biomol. Chem. 21, 8125–8135. doi:10.1039/D3OB01348K
Nielsen, P. E. (1997). Peptide nucleic acid (PNA) from DNA recognition to antisense and DNA structure. Biophys. Chem. 68, 103–108. doi:10.1016/S0301-4622(97)00019-7
Oh, J.-H., and Lee, J.-S. (2011). Designed hybridization properties of DNA–gold nanoparticle conjugates for the Ultraselective detection of a single-base mutation in the breast cancer gene BRCA1. Anal. Chem. 83, 7364–7370. doi:10.1021/ac201291y
Owczarzy, R., You, Y., Groth, C. L., and Tataurov, A. V. (2011). Stability and mismatch discrimination of locked nucleic acid–DNA duplexes. Biochemistry 50, 9352–9367. doi:10.1021/bi200904e
Oyaghire, S. N., Quijano, E., Perera, J. D. R., Mandl, H. K., Saltzman, W. M., Bahal, R., et al. (2023). DNA recognition and induced genome modification by a hydroxymethyl-γ tail-clamp peptide nucleic acid. Cell Rep. Phys. Sci. 4, 101635. doi:10.1016/j.xcrp.2023.101635
Petersen, M., Bondensgaard, K., Wengel, J., and Jacobsen, J. P. (2002). Locked nucleic acid (LNA) recognition of RNA: NMR solution structures of LNA:RNA hybrids. J. Am. Chem. Soc. 124, 5974–5982. doi:10.1021/ja012288d
Petree, J. R., Yehl, K., Galior, K., Glazier, R., Deal, B., and Salaita, K. (2018). Site-selective RNA splicing nanozyme: DNAzyme and RtcB conjugates on a gold nanoparticle. Acs. Chem. Biol. 13, 215–224. doi:10.1021/acschembio.7b00437
Pfundheller, H. M., Sørensen, A. M., Lomholt, C., Johansen, A. M., Koch, T., and Wengel, J. (2005). Locked nucleic acid synthesis. Methods Mol. Biol. 288, 127–146. doi:10.1385/1-59259-823-4:127
Phafat, B., and Bhattacharya, S. (2023). Quantum dots as Theranostic agents: recent advancements, SurfaceModifications, and future applications. MRMC 23, 1257–1272. doi:10.2174/1389557522666220405202222
Pitiphattharabun, S., Auewattanapun, K., Htet, T. L., Thu, M. M., Panomsuwan, G., Techapiesancharoenkij, R., et al. (2024). Reduced graphene oxide/zinc oxide composite as an electrochemical sensor for acetylcholine detection. Sci. Rep. 14, 14224. doi:10.1038/s41598-024-64238-7
Pyrak, E., Kowalczyk, A., Weyher, J. L., Nowicka, A. M., and Kudelski, A. (2023). Influence of sandwich-type DNA construction strategy and plasmonic metal on signal generated by SERS DNA sensors. Spectrochim. Acta A Mol. Biomol. Spectrosc. 295, 122606. doi:10.1016/j.saa.2023.122606
Ramanathan, S., Gopinath, S. C. B., Arshad, M. K.Md., Poopalan, P., and Anbu, P. (2019). A DNA based visual and colorimetric aggregation assay for the early growth factor receptor (EGFR) mutation by using unmodified gold nanoparticles. Microchim. Acta 186, 546. doi:10.1007/s00604-019-3696-y
Rastogi, S. K., Gibson, C. M., Branen, J. R., Eric Aston, D., Larry Branen, A., and Hrdlicka, P. J. (2012). DNA detection on lateral flow test strips: enhanced signal sensitivity using LNA-conjugated gold nanoparticles. Chem. Commun. 48, 7714. doi:10.1039/c2cc33430e
Romero-Palomo, F., Festag, M., Lenz, B., Schadt, S., Brink, A., Kipar, A., et al. (2021). Safety, tissue distribution, and metabolism of LNA-containing antisense oligonucleotides in Rats. Toxicol. Pathol. 49, 1174–1192. doi:10.1177/01926233211011615
Rose, M., Lapuebla, A., Landman, D., and Quale, J. (2019). In vitro and in vivo activity of a novel antisense peptide nucleic acid Compound against Multidrug-Resistant Acinetobacter baumannii Microb. Drug resist. Microb. Drug Resist. 25, 961–965. doi:10.1089/mdr.2018.0179
Rosenbach, H., Victor, J., Etzkorn, M., Steger, G., Riesner, D., and Span, I. (2020). Molecular features and metal ions that influence 10-23 DNAzyme activity. Molecules 25, 3100. doi:10.3390/molecules25133100
Rosso, V., Bracco, E., Pedrola, R., Carturan, S., Signorino, E., Petiti, J., et al. (2015). Detection of BCR-ABL T315I mutation by peptide nucleic acid directed PCR clamping and by peptide nucleic acid FISH. Biomark. Res. 3, 15. doi:10.1186/s40364-015-0039-y
Ruijter, N., Soeteman-Hernández, L. G., Carrière, M., Boyles, M., McLean, P., Catalán, J., et al. (2023). The state of the Art and challenges of in vitro methods for human hazard assessment of nanomaterials in the context of safe-by-design. Nanomaterials 13, 472. doi:10.3390/nano13030472
Ruiz, C., Huang, J., Giardina, S. F., Feinberg, P. B., Mirza, A. H., Bacolod, M. D., et al. (2020). Single-molecule detection of cancer mutations using a novel PCR-LDR-qPCR assay. Hum. Mutat. 41, 1051–1068. doi:10.1002/humu.23987
Ryncarz, A. J., Goddard, J., Wald, A., Huang, M. L., Roizman, B., and Corey, L. (1999). Development of a high-throughput quantitative assay for detecting herpes simplex virus DNA in clinical samples. J. Clin. Microbiol. 37, 1941–1947. doi:10.1128/JCM.37.6.1941-1947.1999
Saikia, N., Taha, M., and Pandey, R. (2021). Molecular insights on the dynamic stability of peptide nucleic acid functionalized carbon and boron nitride nanotubes. Phys. Chem. Chem. Phys. 23, 219–228. doi:10.1039/D0CP05759B
Sanger, F., Nicklen, S., and Coulson, A. R. (1977). DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74, 5463–5467. doi:10.1073/pnas.74.12.5463
Sato, K. (2005). Non-cross-linking gold nanoparticle aggregation as a detection method for single-base substitutions. Nucleic Acids Res. 33, e4. doi:10.1093/nar/gni007
Sau, S. P., Kumar, T. S., and Hrdlicka, P. J. (2010). Invader LNA: efficient targeting of short double stranded DNA. Org. Biomol. Chem. 8, 2028. doi:10.1039/b923465a
Sawamura, K., and Hashimoto, M. (2017). A fluorescence quenching assay based on molecular beacon formation through a ligase detection reaction for facile and rapid detection of point mutations. Anal. Sci. 33, 1457–1460. doi:10.2116/analsci.33.1457
Sebuyoya, R., Sevcikova, S., Yusuf, B., and Bartosik, M. (2025). Integrating isothermal amplification techniques and LNA-based AI-assisted electrochemical bioassay for analysis of KRAS G12V point mutation. Talanta 288, 127709. doi:10.1016/j.talanta.2025.127709
Shen, X., Lin, Q., Liang, Z., Wang, J., Yang, X., Liang, Y., et al. (2022). Reduction of pre-existing adaptive immune responses against SaCas9 in humans using Epitope Mapping and identification. CRISPR J. 5, 445–456. doi:10.1089/crispr.2021.0142
Shi, W., Gong, Y., Zhang, D., Yang, T., Yi, M., Tan, J., et al. (2022). A dual identification strategy based on padlock ligation and CRISPR/Cas14a for highly specific detection of BRAF V600E mutation in clinical samples. Anal. Methods 14, 1913–1921. doi:10.1039/D2AY00319H
Siegberg, D., and Herten, D.-P. (2011). Fluorescence quenching of quantum dots by DNA nucleotides and amino acids. Aust. J. Chem. 64, 512. doi:10.1071/CH10293
Singh, J., Hanson, J., Paliwal, K., and Zhou, Y. (2019). RNA secondary structure prediction using an ensemble of two-dimensional deep neural networks and transfer learning. Nat. Commun. 10, 5407. doi:10.1038/s41467-019-13395-9
Skotadis, E., Voutyras, K., Chatzipetrou, M., Tsekenis, G., Patsiouras, L., Madianos, L., et al. (2016). Label-free DNA biosensor based on resistance change of platinum nanoparticles assemblies. Biosens. Bioelectron. 81, 388–394. doi:10.1016/j.bios.2016.03.028
Smith, M., Smith, K., Olstein, A., Oleinikov, A., and Ghindilis, A. (2020). Restriction endonuclease-based assays for DNA detection and isothermal exponential signal amplification. Sensors 20, 3873. doi:10.3390/s20143873
Song, Y., Zhang, W., An, Y., Cui, L., Yu, C., Zhu, Z., et al. (2012). Label-free visual detection of nucleic acids in biological samples with single-base mismatch detection capability. Chem. Commun. (Camb) 48, 576–578. doi:10.1039/c1cc15777a
Song, J., Hegge, J. W., Mauk, M. G., Chen, J., Till, J. E., Bhagwat, N., et al. (2020). Highly specific enrichment of rare nucleic acid fractions using Thermus thermophilus argonaute with applications in cancer diagnostics. Nucleic Acids Res. 48, e19. doi:10.1093/nar/gkz1165
Stamey, F. R., Patel, M. M., Holloway, B. P., and Pellett, P. E. (2001). Quantitative, fluorogenic probe PCR assay for detection of human herpesvirus 8 DNA in clinical specimens. J. Clin. Microbiol. 39, 3537–3540. doi:10.1128/JCM.39.10.3537-3540.2001
Su, X., Li, L., Wang, S., Hao, D., Wang, L., and Yu, C. (2017). Single-molecule counting of point mutations by transient DNA binding. Sci. Rep. 7, 43824. doi:10.1038/srep43824
Sun, P., Wang, S., Yan, Q., Zeng, J., Wu, Z., and Qi, X. (2025). Non-nuclear localization signal-guided CRISPR/Cas9 ribonucleoproteins for translocation and gene editing via apoferritin delivery vectors. Nanoscale 17, 6660–6675. doi:10.1039/d4nr04762a
Swayze, E. E., Siwkowski, A. M., Wancewicz, E. V., Migawa, M. T., Wyrzykiewicz, T. K., Hung, G., et al. (2007). Antisense oligonucleotides containing locked nucleic acid improve potency but cause significant hepatotoxicity in animals. Nucleic Acids Res. 35, 687–700. doi:10.1093/nar/gkl1071
Tabebordbar, M., Zhu, K., Cheng, J. K. W., Chew, W. L., Widrick, J. J., Yan, W. X., et al. (2016). In vivo gene editing in dystrophic mouse muscle and muscle stem cells. Science 351, 407–411. doi:10.1126/science.aad5177
Tanabe, M., Ishino, Y., and Nishida, H. (2015). From structure-function analyses to protein engineering for practical applications of DNA ligase. Archaea 2015, 1–20. doi:10.1155/2015/267570
Tang, Y., Zhang, X.-L., Tang, L.-J., Yu, R.-Q., and Jiang, J.-H. (2017). In situ imaging of individual mRNA mutation in single cells using ligation-mediated branched hybridization chain reaction (Ligation-bHCR). Anal. Chem. 89, 3445–3451. doi:10.1021/acs.analchem.6b04312
Tian, K., Chen, X., Luan, B., Singh, P., Yang, Z., Gates, K. S., et al. (2018). Single locked nucleic acid-enhanced nanopore genetic discrimination of pathogenic Serotypes and cancer driver mutations. ACS Nano 12, 4194–4205. doi:10.1021/acsnano.8b01198
Valentini, P., Fiammengo, R., Sabella, S., Gariboldi, M., Maiorano, G., Cingolani, R., et al. (2013). Gold-nanoparticle-based colorimetric discrimination of cancer-related point mutations with Picomolar sensitivity. ACS Nano 7, 5530–5538. doi:10.1021/nn401757w
Van Dijk, E. L., Jaszczyszyn, Y., Naquin, D., and Thermes, C. (2018). The third revolution in sequencing technology. Trends Genet. 34, 666–681. doi:10.1016/j.tig.2018.05.008
Viana, M., Souza, T., Bueno-Silva, B., Gonçalves, F., Braga, R., Nascimento, F., et al. (2024). Antimicrobial and optical properties of a new biogenic silica-coated silver nanoparticles incorporated into experimental resin. J. Clin. Exp. Dent. 16, e151–e158. doi:10.4317/jced.61004
Wang, P., and Zhang, P. (2014). Ligase-assisted, upconversion luminescence resonance energy transfer-based method for specific and sensitive detection of V600E mutation in the BRAF gene. RSC Adv. 4, 56235–56240. doi:10.1039/C4RA10181B
Wang, H.-Q., Liu, W.-Y., Wu, Z., Tang, L.-J., Xu, X.-M., Yu, R.-Q., et al. (2011a). Homogeneous label-free genotyping of single nucleotide polymorphism using ligation-mediated strand displacement amplification with DNAzyme-based chemiluminescence detection. Anal. Chem. 83, 1883–1889. doi:10.1021/ac200138v
Wang, Q., Yang, L., Yang, X., Wang, K., He, L., and Zhu, J. (2011b). Electrochemical biosensors for detection of point mutation based on surface ligation reaction and oligonucleotides modified gold nanoparticles. Anal. Chim. Acta 688, 163–167. doi:10.1016/j.aca.2011.01.004
Wang, H., Jiang, X., Wang, X., Wei, X., Zhu, Y., Sun, B., et al. (2014a). Hairpin DNA-assisted Silicon/silver-based surface-enhanced Raman scattering sensing platform for Ultrahighly sensitive and specific discrimination of deafness mutations in a real system. Anal. Chem. 86, 7368–7376. doi:10.1021/ac501675d
Wang, P., Joshi, P., Alazemi, A., and Zhang, P. (2014b). Upconversion nanoparticle-based ligase-assisted method for specific and sensitive detection of T790M mutation in epidermal growth factor receptor. Biosens. Bioelectron. 62, 120–126. doi:10.1016/j.bios.2014.06.037
Wang, Y., Nguyen, K., Spitale, R. C., and Chaput, J. C. (2021). A biologically stable DNAzyme that efficiently silences gene expression in cells. Nat. Chem. 13, 319–326. doi:10.1038/s41557-021-00645-x
Wang, Y., Sun, H., Xu, G., Guan, M., Zhang, Q., Wang, Z., et al. (2022). A multiplexed electrochemical quantitative polymerase chain reaction platform for single-base mutation analysis. Biosens. Bioelectron. 214, 114496. doi:10.1016/j.bios.2022.114496
Wang, S., Ren, X., Wang, J., Peng, Q., Niu, X., Song, C., et al. (2023a). Blocking autofluorescence in brain tissues affected by ischemic stroke, hemorrhagic stroke, or traumatic brain injury. Front. Immunol. 14, 1168292. doi:10.3389/fimmu.2023.1168292
Wang, X., Wei, G., Liu, W., Zhang, Y., Zhu, C., Sun, Q., et al. (2023b). Platinum-nickel nanoparticles with enhanced Oxidase-like activity for total Antioxidant capacity bioassay. Anal. Chem. 95, 5937–5945. doi:10.1021/acs.analchem.2c05425
Wang, Y., Qian, Y., Zhang, L., Zhang, Z., Chen, S., Liu, J., et al. (2023c). Conductive metal-organic framework Microelectrodes regulated by conjugated molecular Wires for monitoring of Dopamine in the mouse brain. J. Am. Chem. Soc. 145, 2118–2126. doi:10.1021/jacs.2c07053
Wang, H., Jian, M., Fan, J., He, Y., and Wang, Z. (2024a). Scalable synthesis of Au@CeO2 nanozyme for development of colorimetric lateral flow immunochromatographic assay to sensitively detect heart-type fatty acid binding protein. Talanta 273, 125852. doi:10.1016/j.talanta.2024.125852
Wang, K.-H., Chen, Y.-Y., Wang, C.-H., Hsu, K.-F., Chau, L.-K., Wang, S.-C., et al. (2024b). Ultrasensitive amplification-free detection of circulating miRNA via droplet-based processing of SERS tag–miRNA–magnetic nanoparticle sandwich nanocomplexes on a paper-based electrowetting-on-dielectric platform. Analyst 149, 1981–1987. doi:10.1039/D3AN01429K
Warshawsky, I., and Mularo, F. (2011). Locked nucleic acid probes for enhanced detection of FLT3 D835/I836, JAK2 V617F and NPM1 mutations. J. Clin. Pathol. 64, 905–910. doi:10.1136/jclinpath-2011-200086
Wu, L., Liu, Q., Wu, Z., and Lu, Z. (2010). Detection of HIV cDNA point mutations with rolling-circle amplification arrays. Mol 15, 619–626. doi:10.3390/molecules15020619
Wu, L., Garrido-Maestu, A., Guerreiro, J. R. L., Carvalho, S., Abalde-Cela, S., Prado, M., et al. (2019). Amplification-free SERS analysis of DNA mutation in cancer cells with single-base sensitivity. Nanoscale 11, 7781–7789. doi:10.1039/C9NR00501C
Wu, X., Pu, X., Zhang, Y., Wu, H., Yu, Z., He, W., et al. (2025). A nucleic acid-based strategy for highly specific discrimination between mutant and wild-type sequences. Anal. Biochem. 705, 115930. doi:10.1016/j.ab.2025.115930
Xie, W., and Schlücker, S. (2013). Medical applications of surface-enhanced Raman scattering. Phys. Chem. Chem. Phys. 15, 5329. doi:10.1039/c3cp43858a
Xu, Q., Huang, S., Ma, F., Tang, B., and Zhang, C. (2016). Controllable mismatched ligation for Bioluminescence screening of known and Unknown mutations. Anal. Chem. 88, 2431–2439. doi:10.1021/acs.analchem.5b04540
Xu, H., Wu, Q., and Shen, H. (2017). A DNAzyme sensor based on target-catalyzed hairpin assembly for enzyme-free and non-label single nucleotide polymorphism genotyping. Talanta 167, 630–637. doi:10.1016/j.talanta.2017.03.001
Xue, M., Gonzalez, D. H., Osikpa, E., Gao, X., and Lillehoj, P. B. (2025). Rapid and automated interpretation of CRISPR-Cas13-based lateral flow assay test results using machine learning. Sens. Diagn 4, 171–181. doi:10.1039/d4sd00314d
Yagita, R., Murakami, K., Ikeda, H., and Irie, K. (2020). Synthesis and physicochemical properties of 20-mer peptide nucleic acid conjugates with testosterone 17β-carboxylic acid. Tetrahedron Lett. 61, 151781. doi:10.1016/j.tetlet.2020.151781
Yang, C., Dou, B., Yang, J., Yuan, R., and Xiang, Y. (2016). Cross-triggered and cascaded recycling amplification for ultrasensitive electrochemical sensing of the mutant human p53 gene. Chem. Commun. 52, 8707–8710. doi:10.1039/C6CC04587A
Yang, B., Zhang, S., Fang, X., and Kong, J. (2019). Double signal amplification strategy for ultrasensitive electrochemical biosensor based on nuclease and quantum dot-DNA nanocomposites in the detection of breast cancer 1 gene mutation. Biosens. Bioelectron. 142, 111544. doi:10.1016/j.bios.2019.111544
Yang, X., Liao, M., Zhang, H., Gong, J., Yang, F., Xu, M., et al. (2021). An electrochemiluminescence resonance energy transfer biosensor for the detection of circulating tumor DNA from blood plasma. iScience 24, 103019. doi:10.1016/j.isci.2021.103019
Yang, F., Gong, J., Li, M., Jiang, X., Zhang, J., Liao, M., et al. (2023). Electrochemiluminescent CdS quantum dots biosensor for cancer mutation detection at different positions on linear DNA Analytes. Anal. Chem. 95, 14016–14024. doi:10.1021/acs.analchem.3c02649
Ye, M., Chen, Y., Wang, Y., Xiao, L., Lin, Q., Lin, H., et al. (2022). Subtype discrimination of acute myeloid leukemia based on plasma SERS technique. Spectrochim. Acta A Mol. Biomol. Spectrosc. 271, 120865. doi:10.1016/j.saa.2022.120865
Yeh, H.-C. (2006). Homogeneous point mutation detection by quantum dot-mediated two-color fluorescence coincidence analysis. Nucleic Acids Res. 34, e35. doi:10.1093/nar/gkl021
Yu, C.-J., Cheng, T.-L., and Tseng, W.-L. (2009). Effects of Mn2+ on oligonucleotide-gold nanoparticle hybrids for colorimetric sensing of Hg2+: improving colorimetric sensitivity and accelerating color change. Biosens. Bioelectron. 25, 204–210. doi:10.1016/j.bios.2009.06.038
Zandieh, M., and Liu, J. (2024). Nanozymes: definition, activity, and mechanisms. Adv. Mater. 36, e2211041. doi:10.1002/adma.202211041
Zeng, J., Dong, M., Zhu, B., Gao, X., Chen, D., and Li, Y. (2021). Label-free detection of C–T mutations by surface-enhanced Raman spectroscopy using thiosulfate-modified nanoparticles. Anal. Chem. 93, 1951–1956. doi:10.1021/acs.analchem.0c04052
Zhang, K., and Zhang, Y. (2010). Lable-free electrochemical DNA sensor based on gold nanoparticles/poly(neutral red) modified electrode. Electroanalysis 22, 673–679. doi:10.1002/elan.200900453
Zhang, C.-Y., Yeh, H.-C., Kuroki, M. T., and Wang, T.-H. (2005a). Single-quantum-dot-based DNA nanosensor. Nat. Mater. 4, 826–831. doi:10.1038/nmat1508
Zhang, R.-Y., Wang, X.-M., Gong, S.-J., and He, N.-Y. (2005b). Electrochemical detection of single A-G mismatch using biosensing surface based on gold nanoparticles. Genom. Proteom. Bioinform 3, 47–51. doi:10.1016/S1672-0229(05)03007-X
Zhang, L., Wang, J., Coetzer, M., Angione, S., Kantor, R., and Tripathi, A. (2015). One-step ligation on RNA amplification for the detection of point mutations. J. Mol. Diagn. 17, 679–688. doi:10.1016/j.jmoldx.2015.07.001
Zhang, X., Fan, X., Wang, Y., Lei, F., Li, L., Liu, J., et al. (2020). Highly stable colorimetric sensing by assembly of gold nanoparticles with SYBR green I: from charge screening to charge neutralization. Anal. Chem. 92, 1455–1462. doi:10.1021/acs.analchem.9b04660
Zhang, Z., Hu, Y., Yuan, W., Hu, M., Deng, Y., Xiao, X., et al. (2021). Endonuclease IV-regulated DNAzyme motor for universal single-nucleotide variation discrimination. Anal. Chem. 93, 9939–9948. doi:10.1021/acs.analchem.1c02230
Zhang, Y., Zeng, J., Huang, C., Zhu, B., Zhang, Q., and Chen, D. (2022). Label-free detection of ssDNA base insertion and deletion mutations by surface-enhanced Raman spectroscopy. Anal. Bioanal. Chem. 414, 1461–1468. doi:10.1007/s00216-021-03799-2
Zhang, X., Li, Q., Chao, Q., Zhang, Y., Sun, X., Fan, G.-C., et al. (2023). A protein enzyme-free strategy for fluorescence detection of single nucleotide polymorphisms using asymmetric MNAzymes. Anal. Chim. Acta 1243, 340811. doi:10.1016/j.aca.2023.340811
Zhang, Y., Wang, L., Ye, J., Chen, J., Xu, S., Bu, S., et al. (2024). Rationally designed dual base pair mismatch enables toehold-mediated strand displacement to efficiently recognize single-nucleotide polymorphism without enzymes. Anal. Chem. 96, 554–563. doi:10.1021/acs.analchem.3c04778
Zhao, S., Xu, M., Lin, C., Zhang, W., Li, D., Peng, Y., et al. (2025). AI-driven comprehensive SERS-LFIA system: improving virus automated diagnostics through SERS image recognition and deep learning. Biosens. (Basel) 15, 458. doi:10.3390/bios15070458
Zhou, L., Du, F., Zhao, Y., Yameen, A., Chen, H., and Tang, Z. (2013). DNAzyme based gap-LCR detection of single-nucleotide polymorphism. Biosens. Bioelectron. 45, 141–147. doi:10.1016/j.bios.2013.01.061
Zhou, Q.-Y., Yuan, F., Zhang, X.-H., Zhou, Y.-L., and Zhang, X.-X. (2018). Simultaneous multiple single nucleotide polymorphism detection based on click chemistry combined with DNA-encoded probes. Chem. Sci. 9, 3335–3340. doi:10.1039/c8sc00307f
Zhou, H., Yang, C., Liao, M., Li, M., Diao, N., Wu, S., et al. (2022). Exploring the mechanism of CdTe quantum dots as fluorescent probe to detect Hg(II) ion from the perspectives of fluorescence polarization and light scattering. Chem. Phys. Lett. 792, 139415. doi:10.1016/j.cplett.2022.139415
Zhou, S., Miao, Y., Qiu, H., Yao, Y., Wang, W., and Chen, C. (2024). Deep learning based local feature classification to automatically identify single molecule fluorescence events. Commun. Biol. 7, 1404. doi:10.1038/s42003-024-07122-4
Zou, Z., Qing, Z., He, X., Wang, K., He, D., Shi, H., et al. (2014). Ligation-rolling circle amplification combined with γ-cyclodextrin mediated stemless molecular beacon for sensitive and specific genotyping of single-nucleotide polymorphism. Talanta 125, 306–312. doi:10.1016/j.talanta.2014.03.014
Keywords: mutation detection, point mutations, low-level mutation, enzymes, nucleic acid analogs, nanomaterials
Citation: Pu X and Wu X (2025) Advances in nucleic acid probe-based detection of gene point mutations: a review. Front. Chem. 13:1672155. doi: 10.3389/fchem.2025.1672155
Received: 24 July 2025; Accepted: 15 September 2025;
Published: 25 September 2025.
Edited by:
Serge Cosnier, UMR5250 Département de Chimie Moléculaire (DCM), FranceCopyright © 2025 Pu and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xueqiang Wu, d3V4dWVxaWFuZ0BtenJteXkuY29t