 Yan Su
Yan Su Youhe Gao
Youhe Gao- Gene Engineering Drug and Biotechnology Beijing Key Laboratory, College of Life Sciences, Beijing Normal University, Beijing, China
Objective: To investigate the effects of different edible oils on the rat body, we analyzed alterations in the urinary proteome and post-translational modifications (PTMs) following a one-week intake of olive oil, butter, lard, hydrogenated vegetable oil, and rapeseed oil.
Methods: Thirty male Wistar rats (7 weeks old, ~200 g) were randomly allocated into six groups (n = 5 per group). Group A served as the control group, while groups B-F were administered different edible oils. The daily intakes were calculated, respectively, according to the “2015–2020 Dietary Guidelines for Americans” and the “Dietary Guidelines for Chinese Residents.” Urine samples collected after 1 week were analyzed using label-free quantitative proteomics via LC–MS/MS. Differentially expressed proteins and differential post-translational modifications in the urinary proteome were screened for functional analysis.
Results: All edible oil groups exhibited significant alterations in metabolic pathways, with distinct proteomic profiles observed across oil types, but there were few common differentially expressed proteins among different groups. In addition, the olive oil group and the butter group were enriched with many biological pathways related to the nervous system, and the rapeseed oil group produced more differentially expressed proteins and biological pathways related to immunity.
Conclusion: The urinary proteome of rats showed significant changes after one-week intake of edible oils, and the effects of various edible oils on the rat urinary proteome were different from each other. This effect is comprehensive and multi-dimensional at the level of the rat body. The changes in post-translational modifications of the proteome were relatively small.
1 Introduction
Edible oils, as a major global dietary component, provide essential fatty acids and energy while maintaining physiological homeostasis (1). With the increasing attention to health, the nutritional values and potential health impacts of different edible oils have gradually become research hotspots.
Previous research indicates that diverse edible oils exhibit distinct physiological functions and health impacts owing to their unique fatty acid profiles and nutritional compositions (2). For example, olive oil, the main source of fat in the Mediterranean diet, is rich in monounsaturated fatty acids and is considered to have beneficial effects such as reducing the risk of cardiovascular diseases (3). Butter contains a relatively high amount of saturated fatty acids. Some studies have shown that excessive intake of saturated fatty acids in butter may be associated with an increased risk of cardiovascular diseases (4). However, a considerable number of research findings have indicated that there is insufficient evidence to support the relationship between them, or the relationship is not significant (5, 6). Lard is an animal oil consumed frequently by people in Asian regions. Its fatty acid composition is relatively balanced, including 52.1% saturated fatty acids, 35.8% monounsaturated fatty acids, and 11.6% polyunsaturated fatty acids, and its impact on health is controversial (7, 8). Hydrogenated vegetable oil, containing trans-fatty acids, is associated with adverse cardiovascular effects (9). Rapeseed oil, as a vegetable oil, has a relatively balanced fatty acid ratio and contains nutritional components such as antioxidants (10).
As a non-invasive detection technique, urinary proteomics can sensitively reflect the pathophysiological changes of the body by analyzing the proteins and their modification states in urine, and has become an important tool for discovering early disease biomarkers (11, 12). In recent years, this technique has demonstrated unique advantages in the mechanistic studies of metabolic syndrome (13), neurodegenerative diseases (14), and immune-related diseases (15). It is worth noting that post-translational modifications (PTMs), as a core mechanism for regulating protein function, still have many unknown aspects regarding their dynamic change patterns and biological significance in dietary interventions. PTMs significantly expand the functional diversity of proteins by adding chemical groups such as phosphorylation, glycosylation, and acetylation to amino acid residues, and are deeply involved in key physiological processes such as cell signaling, energy metabolism, and stress responses (16). For example, tyrosine phosphorylation modification plays a pivotal role in the insulin signaling pathway (17), while lysine acetylation is closely related to the regulation of liver lipid metabolism (18). Recent studies have shown that dietary interventions can affect metabolic phenotypes by altering the PTMs profile: N-glycosylation changes occur in patients with type 2 diabetes, and N-glycans can even identify individuals with an increased risk of disease development (19); polyphenolic components in olive oil can alleviate oxidative stress damage by regulating PTMs (20). However, currently, there is a lack of systematic research on the effects of different edible oils on the urinary proteome and PTMs. In particular, the changes in the urinary proteome and PTMs of the body after short-term intake of different oils have not been reported yet.
Given the significant differences in fatty acid compositions and nutritional characteristics among different edible oils, as well as their common consumption in daily diets, this study selected several representative edible oils, including olive oil, butter, lard, hydrogenated vegetable oil, and rapeseed oil, for research. To avoid the interference of short-term growth and development of rats on the experimental results, we chose to conduct parallel comparisons between different experimental groups and the control group. By comparing the differences in the urinary proteomes of rats, we preliminarily explored the possible impacts of short-term consumption of different edible oils on the metabolic processes in rats, providing certain theoretical references and experimental bases for a deeper understanding of their potential impacts on human health.
2 Materials and methods
2.1 Experimental animals and model establishment
Thirty 7-week-old male Wistar rats (weighing approximately 200 g) were purchased from Beijing Vital River Laboratory Animal Technology Co., Ltd. All rats were raised in a standard environment (room temperature: (22 ± 1) °C, humidity: 65–70%). The experiment started after the rats were raised in the new environment for 3 days. All experimental operations were reviewed and approved by the Ethics Committee of the College of Life Sciences, Beijing Normal University, with the approval number CLS-AWEC-B-2022-003.
In this experiment, group A was designated as the control group, receiving no additional edible oil. The other groups were fed with different types of edible oils, namely olive oil (group B), butter (group C), lard (group D), hydrogenated vegetable oil (group E), and rapeseed oil (group F), with 5 rats in each group. According to the “2015–2020 Dietary Guidelines for Americans” and the “Dietary Guidelines for Chinese Residents,” the ratio of the daily intake of edible oil to body weight for adults was calculated. Then, based on the equivalent dose ratio table for humans and animals converted according to body surface area, the dosage for rats was calculated as approximately 6.25-fold the human equivalent based on body surface area conversion. The daily intake of edible oil for rats was calculated as shown in Table 1.
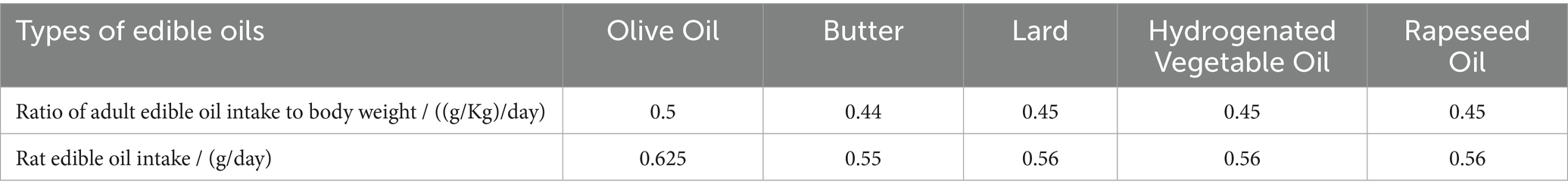
Table 1. Daily intake of edible oil for rats.
After the adaptive feeding period ended, at 17:00 every day, rats in different groups were fed with the corresponding edible oils for 7 consecutive days. In this experiment, the following feeding method was employed to ensure accurate intake of the specified dose of edible oil by rats: At a fixed time daily during the experiment, rats were gently restrained in a quiet environment to ensure their safety and minimize stress. For edible oils of different physical states, a needleless syringe (for liquid oil) or sterile forceps (for solid oil) was used to gently administer the daily dose of oil to the base of the rat’s tongue. The procedure was performed with care to avoid stimulating the pharynx and inducing a vomiting reflex. After feeding, rats were returned to their cages, and their spontaneous licking/swallowing behavior and subsequent physiological responses (such as choking or abnormal restlessness) were observed. On the 7th day, after the rats consumed the edible oils, they were placed in metabolic cages to metabolize. After 12 h overnight, urine was collected. Finally, the collected urine samples were centrifuged at 3,000 r/min for 30 min and then stored in a-80°C refrigerator.
2.2 Processing of urine samples
Urine Protein Extraction Procedure: First, thaw the urine samples, then centrifuge at 12,000 × g for 30 min at 4°C. Transfer the supernatant to a new EP tube. After that, add four-fold volume of absolute ethanol, mix well, and place it in a-20°C refrigerator for one-day precipitation. The next day, centrifuge the mixture at 12,000 × g for 30 min at 4°C, discard the supernatant, resuspend the protein precipitate in lysis buffer (8 mol/L urea, 2 mol/L thiourea, 25 mmol/L dithiothreitol, 50 mmol/L Tris–HCl, pH 8.5), centrifuge again at 12,000 × g for 30 min at 4°C, and retain the supernatant. Finally, determine the protein concentration using the Bradford method.
Protease Digestion Procedure: Take 100 μg of urine protein sample and put it into an EP tube. Add 25 mmol/L NH₄HCO₃ solution to make the total volume 200 μL as a standby sample. Then add dithiothreitol solution (Dithiothreitol, DTT, Sigma) to the tube until its final concentration is 20 mM. Place it in a 95°C metal bath for 10 min of heating, then cool it to room temperature. Next, add iodoacetamide (Iodoacetamide, IAA, Sigma) to a concentration of 50 mM, and let it stand in the dark at room temperature for 40 min. Subsequently, wash the membrane of a 10-kDa ultrafiltration tube (Pall, Port Washington, NY, United States): First, add 200 μL of UA solution (8 mol/L urea, 0.1 mol/L Tris–HCl, pH 8.5), and centrifuge at 14,000 × g for 10 min at 18°C, repeat twice. Add the prepared sample to the ultrafiltration tube and centrifuge at 14,000 × g for 40 min at 18°C. Then add 200 μL of UA solution and centrifuge at 14,000 × g for 30 min at 18°C, repeat twice. Subsequently, add 25 mmol/L NH₄HCO₃ solution and centrifuge at 14,000 × g for 30 min at 18°C, repeat twice. Finally, add trypsin (Trypsin Gold, Promega, Fitchburg, WI, United States) at a trypsin:protein ratio of 1:50, and incubate overnight at 37°C. After overnight incubation, centrifuge to collect the digested filtrate, desalt it using an Oasis HLB solid-phase extraction column, and vacuum-dry it to obtain freeze-dried peptides, which are stored at-80°C.
2.3 LC–MS/MS tandem mass spectrometry analysis
Peptides were reconstituted in 0.1% formic acid (FA) and diluted to a final concentration of 0.5 μg/μL. A total of 2 μL of peptide sample was loaded and separated using a Thermo Scientific Easy-nLC 1,200 liquid chromatography system under the following parameters: 90-min elution time; mobile phase A: 0.1% FA (Thermo Scientific); mobile phase B: 80% acetonitrile (ACN). The separated peptides were analyzed using an Orbitrap Fusion Lumos Tribrid mass spectrometer (Thermo Scientific) in data-independent acquisition (DIA) mode.
For analysis, 1 μg of peptides from each sample was loaded onto a C18 trap column and subsequently separated on a reverse-phase analytical column at a flow rate of 1 μL/min with a 120-min gradient elution (Buffer B: 2 to 6% in 1 min, 6 to 10% in 23 min, 10 to 20% in 67 min, 20 to 28% in 7 min, 28 to 95% in 20 min, and 95 to 5% in 2 min). identical LC parameters were used for both data-dependent (DDA-MS) and data-independent (DIA-MS) acquisition modes.
In DDA mode, 10 fractions obtained from centrifugal column separation were analyzed by mass spectrometry to generate a spectral library. The MS data were acquired in high-sensitivity mode. Full MS scans were acquired in the m/z range of 350–1,500 with a resolution of 120,000, while MS/MS scans were performed in the Orbitrap at a resolution of 30,000. The HCD (Higher-energy Collisional Dissociation) energy was set to 30%, the AGC (Automatic Gain Control) target was set to 5E-04, and the maximum injection time was 45 ms. Subsequently, individual urine samples were analyzed in DIA-MS mode. The DIA method employed 26 variable isolation windows for acquisition. Positive ion mode was set at 3,000 V, with full-scan resolution at 120,000 (m/z range: 350–1,200) and DIA scan resolution at 30,000. The HCD energy was set to 32%, the automatic gain control (AGC) target was set to 1E-06, and the maximum injection time was 50 ms.
To ensure data quality, a pooled peptide sample from all specimens was used for instrument calibration. A DIA analysis of the pooled sample was performed every 7–9 sample runs as a technical replicate for quality control.
2.4 Database search
The Spectronaut Pulsar software (Biognosys AG, Switzerland, Accessed in 2024) was used to search the database for the raw files of the mass spectrometer acquisition results, and compare them with the SwissProt Human database. Calculate the peptide abundance by adding the peak areas of the respective fragment ions in MS2. Protein intensity was calculated by summing the respective peptide intensity.
2.5 Open-pFind unrestricted modification search
Use the pFind Studio software (version 3.2.1, Institute of Computing Technology, Chinese Academy of Sciences) to perform an unrestricted modification search on the three technical replicates of each sample. Use the default parameter settings during the search. The database used is the Rattus norvegicus database downloaded from UniProt, which has been updated to September 2024. The instrument type is set as Higher-energy Collisional Dissociation-Fourier Transform Mass Spectrometry (HCD-FTMS), the selected enzyme is trypsin with full enzyme specificity, allowing a maximum of 2 missed cleavage sites. The precursor mass tolerance is set to ±20 ppm, the fragment ion mass tolerance was also set to ±20 ppm, and the open-search mode is selected. The screening condition clearly stipulates that the false discovery rate (FDR) at the peptide level should be less than 1%.
2.6 Bioinformatics analysis of protein data
Each sample undergoes three technical replicates. The obtained data are averaged and used for statistical analysis. In this experiment, group comparisons were carried out between the edible oil groups (groups B, C, D, E, F) and the control group (group A) to screen for differentially expressed proteins. The screening criteria for differentially expressed proteins are: fold-change (FC) between groups ≥ 1.5 or ≤ 0.67, and a two-tailed paired t-test with a p-value < 0.05. Given the exploratory nature of this study and the limited sample size (n = 5 per group), we prioritized minimizing false negative results over performing strict multiple testing corrections (such as false discovery rate adjustment) to retain potential biological signals. However, we enhanced the reliability of the research findings through the following complementary strategies: First, we conducted random grouping verification. Second, we carried out biological validation by performing functional annotation of differentially expressed proteins based on the literature. The names and functions of the screened differentially expressed proteins are queried through the Uniprot website,1 and biological function enrichment analysis is performed through the DAVID database.2 Also, retrieve reported literature in the Pubmed database3 to conduct functional analysis of the differentially expressed proteins.
2.7 Bioinformatics analysis of protein post-translational modifications
Use Open-pFind to carry out an unrestricted modification search to obtain the post-translational modification PROTEIN file for each sample. Subsequently, download the Python script ‘pFind_protein_contrast_script’ from the GitHub platform (the website is https://github.com/daheitu/scripts_for_pFind3_protocol.io), and use this script to summarize the post-translational modification identification results (i.e., the PROTEIN files) of different samples. Similarly, group comparisons were carried out between the edible oil groups (groups B, C, D, E, F) and the control group (group A) to screen for differential modifications. The screening criteria for differential modifications are: FC ≥ 1.5 or ≤ 0.67, and the p-value of two-tailed paired t-test analysis < 0.05. Query the names and functions of the proteins where the screened differential modifications are located through the Uniprot website (See Text footnote 1).
3 Results
3.1 Behavioral observation of rats
The body weights of rats were measured before and after the experiment (Figure 1a). Comparative analysis revealed that experimental groups (excluding group E) exhibited greater weight gain than the control group. The food intake of rats during the experiment was counted (Figure 1b), and it was found that the food intake of rats in the experimental groups was higher than that of the control group. Behavioral observations indicated that rats in groups C and D preferentially consumed butter and lard, whereas group E rats showed aversion to hydrogenated vegetable oil.
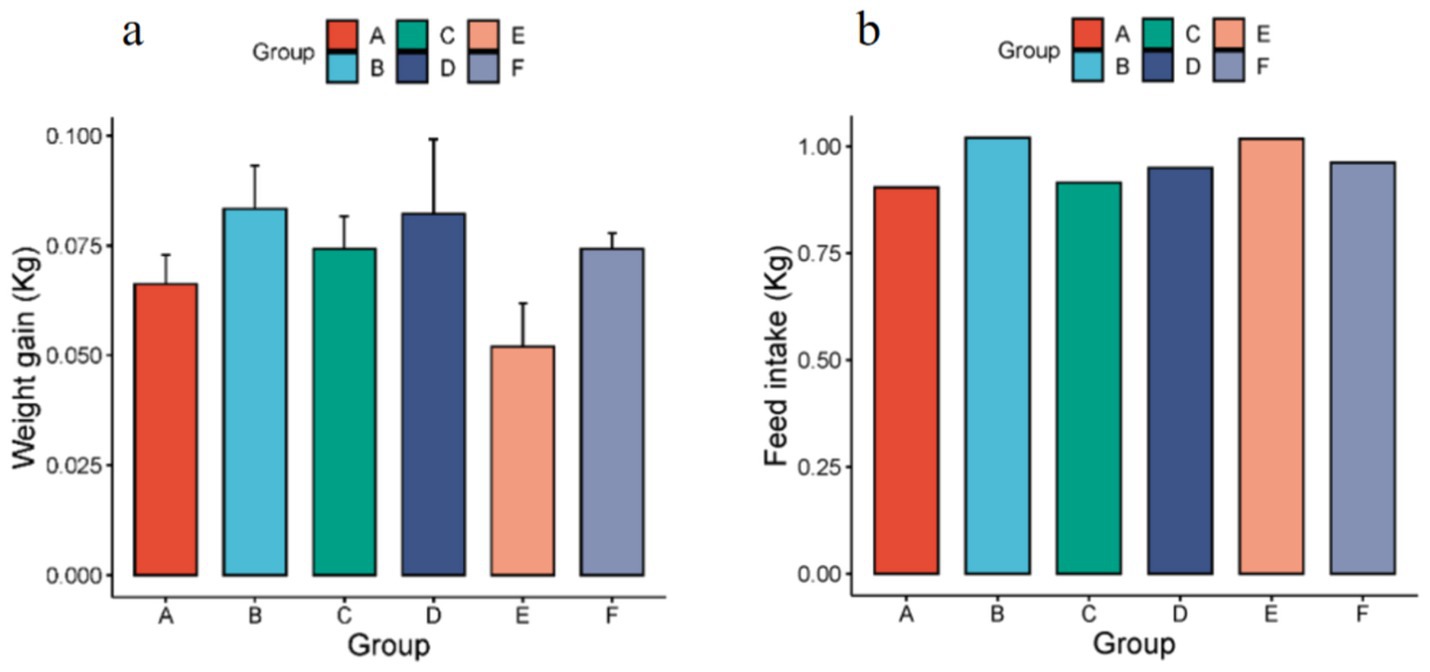
Figure 1. (a) Body weight changes in rats before and after dietary intervention. Data are presented as mean ± SD (n = 5 per group). (b). The total food consumption of rats in each group during the experimental process (n = 5 per group, A-the control group, B-the olive oil group, C-the butter group, D-the lard group, E-the hydrogenated vegetable oil group, F-the rapeseed oil group).
3.2 Group analysis of urinary protein composition
3.2.1 Random grouping
The control group samples (n = 5) and the experimental group samples (n = 5) were randomly divided into two groups, with a total of 126 grouping types. Among all the random combination types, the average number of differentially expressed proteins for all random times was calculated according to the same screening criteria. The ratio of the average number of differentially expressed proteins to the number of differentially expressed proteins obtained under normal grouping is the proportion of randomly generated differentially expressed proteins, as listed in Table 2. These results indicate that the number of randomly generated differentially expressed proteins is relatively small, and the reliability of screening differentially expressed proteins is high. It shows that most of the differentially expressed proteins we obtained are not randomly generated but are the result of the impact of short-term intake of different edible oils.

Table 2. Proportion of randomly generated differentially expressed proteins obtained from random grouping.
3.2.2 Group analysis of urinary protein composition in the olive oil group
3.2.2.1 Differentially expressed proteins
The urinary proteins of the olive oil group (group B) and the control group (group A) were compared. The screening criteria for differentially expressed proteins were: fold-change FC ≥ 1.5 or ≤ 0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 94 differentially expressed proteins were identified in the olive oil group, among which 17 were down-regulated and 77 were up-regulated. The differentially expressed proteins were arranged in ascending order of FC, and retrieved through Uniprot. The detailed information is listed in Appendix Table 1.
3.2.2.2 Functional analysis of differentially expressed proteins
The identified differentially expressed proteins were searched in the Pubmed database for relevant literature.
Among the differentially expressed proteins, the one with the largest down-regulation amplitude and a change from “present to absent” was Alpha-1,3-mannosyl-glycoprotein 4-beta-N-acetylglucosaminyltransferase A (FC = 0, p = 3.81E-02). Research shows that N-acetylglucosaminyltransferase IV is responsible for transferring uridine diphosphate-N-acetylglucosamine to specific glycoprotein structures and is involved in the glycosylation modification of tumor-related N-glycans. This process may be associated with the pathogenesis of diabetes (21).
In addition, some proteins have been shown in existing literature to be related to olive oil, oxidative stress, or metabolism:
IQ motif containing GTPase activating protein 2 (FC = 0.094, p = 2.22E-02) plays an important role in regulating liver fuel storage, especially glycogen levels. In female mice lacking this protein, a significant reduction in glycogen accumulation was observed (22).
Selenoprotein P (FC = 0.213, p = 1.71E-03), as the main selenium-transporting protein, plays a crucial role in maintaining selenium homeostasis in the body and the hierarchical structure of selenoprotein expression. Adequate selenium supply is essential for the normal development and function of the brain through SELENOP, and is crucial for male fertility, proper neurological function, and selenium metabolism (23).
Heat shock protein HSP 90-beta (FC = 0.318, p = 8.62E-03) is abundantly expressed in cardiomyocytes. By maintaining the level of glutathione (a major redox mediator), it is involved in maintaining the redox homeostasis of the cardiovascular system (24).
Glutathione peroxidase (FC = 3.773, p = 2.25E-03) is an important antioxidant enzyme that plays a key role in protecting cells from oxidative stress damage. Studies have found that ligstroside, a polyphenolic compound in olive oil, can increase the expression of Glutathione peroxidase, which may help improve the antioxidant capacity of cells and thus protect cells from damage caused by oxidative stress (25).
Angiopoietin-like 2 (FC = 7.891, p = 1.18E-02) is an important angiogenic factor. In recent years, it has also been found to be involved in mediating the inflammatory process. Its expression is up-regulated in various inflammatory diseases and is directly related to the regulation of inflammatory-related signaling pathways (26).
Leucine-rich repeat transmembrane protein FLRT2 (FC = 8.891, p = 5.15E-03) promotes lipid peroxidation by increasing the expression of acyl-CoA synthetase long-chain family member 4, thereby triggering ferroptosis and inhibiting the malignant phenotype of human bladder cancer cells. It has been identified as a tumor-suppressor gene (27).
Oxidized purine nucleoside triphosphate hydrolase (FC = 10.262, p = 2.83E-03) hydrolyzes oxidized purine nucleoside triphosphates (such as 8-oxo-dGTP and 2-hydroxy-dATP) to monophosphates, thus preventing the mis-incorporation of these oxidized nucleotides during replication (28).
3.2.2.3 Enrichment analysis of biological processes of differentially expressed proteins
The DAVID database was used to perform enrichment analysis of biological processes (BP) on the 94 differentially expressed proteins identified by group analysis. The results showed that a total of 29 BP pathways were significantly enriched (p < 0.05), and the detailed information is presented in Figure 2.
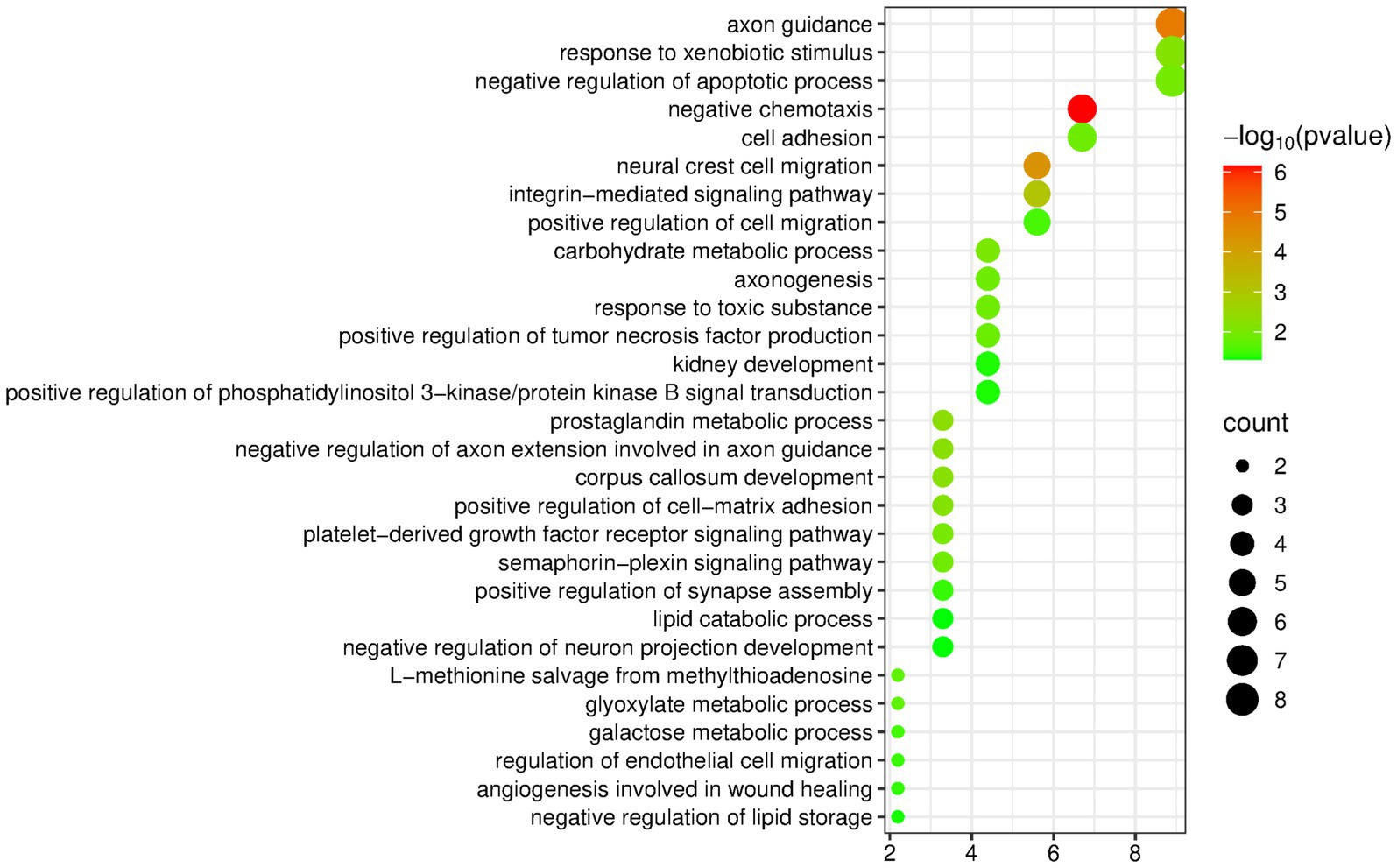
Figure 2. Biological processes enriched by differentially expressed proteins between the olive oil group and the control group (Horizontal axis corresponds to count value, which represents the number of proteins enriched in this pathway. The color of the circle corresponds to the −log10(p) value).
Many of the enriched pathways are related to the nervous system, including axon guidance, neural crest cell migration, negative regulation of axon extension involved in axon guidance, corpus callosum development, axogenesis, positive regulation of synapse assembly, and negative regulation of neuron projection development. Multiple studies have shown that extra-virgin olive oil has important neuroprotective activities, which are closely related to its specific phenolic components. Therefore, nervous-system-related pathways may be enriched (29–31).
The pathway “regulation of endothelial cell migration” has also been mentioned in olive oil research: Hydroxytyrosol and secoiridoids are both components of olive oil, and they play a role in regulating proteins related to endothelial cell proliferation and migration and in regulating proteins related to heart failure in cardiac tissue (20).
In addition, the negative regulation of lipid storage, lipid catabolic process, and negative regulation of lipid biosynthetic process are three pathways directly related to fat intake.
3.2.3 Group analysis of urinary protein composition in the butter group
3.2.3.1 Differentially expressed proteins
The urinary proteins of the butter group (group C) and the control group (group A) were compared. The screening criteria for differentially expressed proteins were: fold-change FC ≥ 1.5 or ≤ 0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 89 differentially expressed proteins were identified in the butter group, among which 15 were down-regulated and 74 were up-regulated. The differentially expressed proteins were arranged in ascending order of FC, and retrieved through Uniprot. The detailed information is listed in Appendix Table 2.
3.2.3.2 Functional analysis of differentially expressed proteins
The identified differentially expressed proteins were searched in the Pubmed database for relevant literature.
Some of the differentially expressed proteins in the butter group were the same as those in the olive oil group, such as Heat shock protein HSP 90-beta, Glutathione peroxidase, Oxidized purine nucleoside triphosphate hydrolase, etc.
Among the other down-regulated differentially expressed proteins in the butter group: C-C motif chemokine 2 (FC = 0.190, p = 2.18E-02) plays a major role in the pathogenesis of cardiovascular diseases. Its down-regulation is considered one of the pleiotropic properties of statins (32). Neuronal membrane glycoprotein M6-a (FC = 0.277, p = 2.56E-02) plays an important role in activating the Src/MAPK/ERK, PKC, and PI3K/AKT, Rufy3-Rap2-STEF/Tiam2 signaling pathways, promoting neurite outgrowth and neuronal polarization respectively, and is also involved in the formation and maturation of dendritic spines (33). Selenoprotein P (FC = 0.340, p = 2.72E-04), as the main selenium-transporting protein, maintains an adequate selenium supply in the brain, which is crucial for the normal development and function of the brain. It may also be related to the pathological processes of the central nervous system and has antioxidant activity (34).
Among the up-regulated proteins: Metalloproteinase inhibitor 3 (FC = 306.177, p = 1.05E-02) had the largest up-regulation amplitude. Studies have found that overexpression of TIMP3 inhibited pathways related to metabolic inflammation and stress, including the activation of Jun NH2-terminal kinase and p38 kinase, and reduced the activation of oxidative stress signals, which were related to lipid peroxidation, protein carbonylation, and nitration (35). An increase in the activity of Prenylcysteine oxidase 1 (FC = 17.347, p = 4.97E-02) may lead to an increase in hydrogen peroxide, thus increasing the oxidative burden during the propagation of low-density lipoproteins. Therefore, this protein can serve as a potential drug target and a new biomarker for cardiovascular diseases (36). Cellular repressor of E1A-stimulated genes 1 (FC = 6.517, p = 3.59E-02) may regulate the homeostasis of vascular wall cells and inhibit the inflammation of vascular tissue cells and macrophages, and has a potential protective effect against inflammation (37). Palmitoyl-protein thioesterase 1 (FC = 3.266, p = 1.42E-02) was up-regulated in rats on a high-fat diet. The increase in its expression may be harmful to the function of Sertoli cells during spermatogenesis (38).
3.2.3.3 Enrichment analysis of biological processes of differentially expressed proteins
The DAVID database was used to perform enrichment analysis of biological processes (BP) on the 89 differentially expressed proteins identified by group analysis. The results showed that a total of 40 BP pathways were significantly enriched (p < 0.05), and the detailed information is presented in Figure 3.
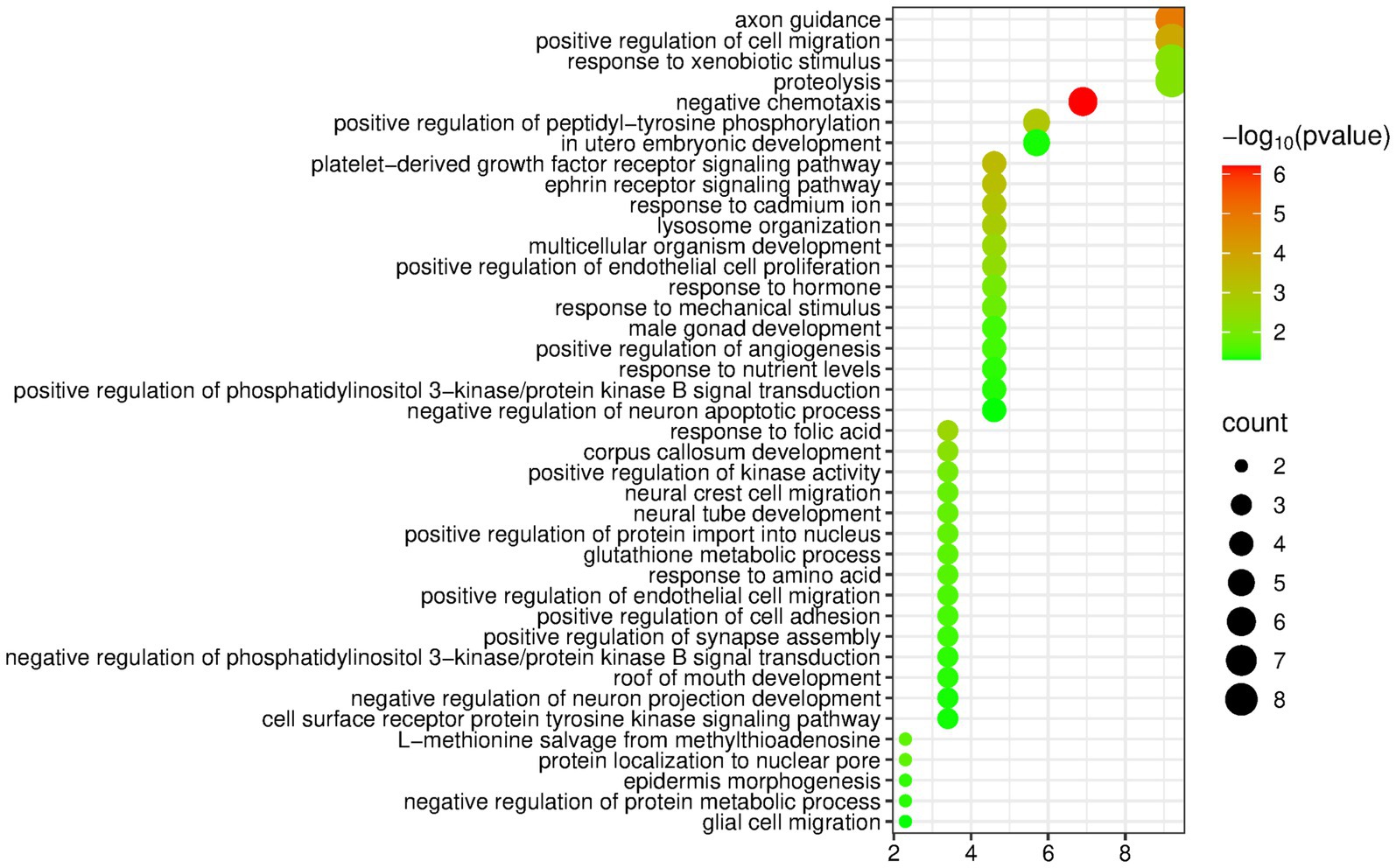
Figure 3. Biological processes enriched by differentially expressed proteins between the butter group and the control group (Horizontal axis corresponds to count value, which represents the number of proteins enriched in this pathway. The color of the circle corresponds to the −log10(p) value).
Similar to the olive oil group, the butter group was also enriched with many pathways related to the nervous system, including axon guidance, corpus callosum development, neural crest cell migration, etc. In addition, the butter group was enriched with some metabolism-related pathways, such as proteolysis, glutathione metabolic process, response to nutrient levels, and negative regulation of protein metabolic process.
3.2.4 Group analysis of urinary protein composition in the lard group
3.2.4.1 Differentially expressed proteins
The urinary proteins of the lard group (group D) and the control group (group A) were compared. The screening criteria for differentially expressed proteins were: fold-change FC ≥ 1.5 or ≤ 0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 104 differentially expressed proteins were identified in the lard group, among which 25 were down-regulated and 79 were up-regulated. The differentially expressed proteins were arranged in ascending order of FC, and retrieved through Uniprot. The detailed information is listed in Appendix Table 3.
3.2.4.2 Functional analysis of differentially expressed proteins
The identified differentially expressed proteins were searched in the Pubmed database for relevant literature.
Among the down-regulated differentially expressed proteins in the lard group: Mitogen-activated protein kinase 3 (FC = 0, p = 2.67E-02) may play an important role in the functional regulation of neurons. Its regulation of the mGlu5 receptor is involved in various neurobiological processes, such as synaptic plasticity, learning, and memory (39). Protein S100-A1 (FC = 0.111, p = 2.99E-02) undergoes a large conformational change when binding to calcium to interact with numerous protein targets, including proteins involved in calcium signaling, neurotransmitter release (synapsins I and II), and the cytoskeleton (40). Dickkopf WNT signaling pathway inhibitor 3 (FC = 0.120, p = 4.74E-02) is significantly down-expressed in the brain tissues of Alzheimer’s disease patients and transgenic mouse models of Alzheimer’s disease (41).
Among the up-regulated differentially expressed proteins: The protein with the largest up-regulation amplitude was Metalloproteinase inhibitor 4 (FC = 85.055, p = 3.60E-02), which belongs to the family of extracellular matrix metalloproteinase inhibitors and is over-expressed in various cancers. Currently, there is no research on its association with lard intake (42). Retinol-binding protein 1 (FC = 38.811, p = 3.04E-02), as a chaperone protein, regulates the uptake, subsequent esterification, and bioavailability of retinol and can deliver vitamin A to cells by interacting with cell-membrane receptors (43, 44). Hyaluronan and proteoglycan link protein 1 (FC = 13.005, p = 5.51E-03) inhibits the NLRP3 inflammasome by stimulating the Nrf2/ARE pathway, thereby suppressing neuroinflammation, enhancing motor neuron survival, and improving neurological functional recovery after spinal cord injury (45). Prominin-2 (FC = 8.125, p = 4.53E-02) is induced by ferroptosis stimulation and can play a role in resisting ferroptosis (46). Protein disulfide-isomerase A6 (FC = 8.080, p = 4.80E-02) is related to spinal cord injury repair and has also been found to promote the repair of damaged neurons (47). 2-amino-3-carboxymuconate-6-semialdehyde decarboxylase (FC = 7.322, p = 1.31E-03) plays a key role in tryptophan catabolism and is an attractive therapeutic target for treating diseases associated with elevated levels of tryptophan metabolites (48).
3.2.4.3 Enrichment analysis of biological processes of differentially expressed proteins
The DAVID database was used to perform enrichment analysis of biological processes (BP) on the 104 differentially expressed proteins identified by group analysis. The results showed that a total of 32 BP pathways were significantly enriched (p < 0.05), and the detailed information is presented in Figure 4.
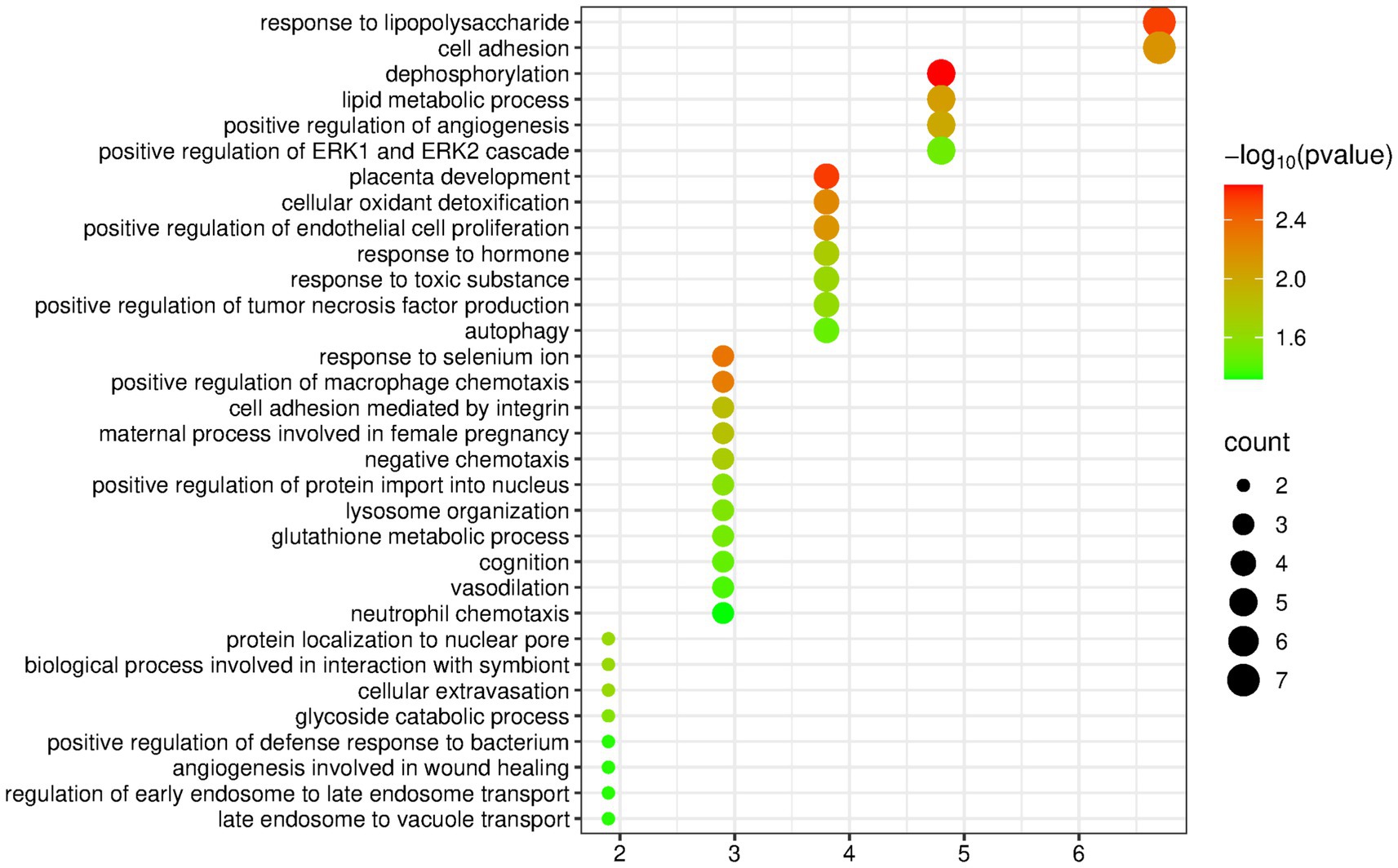
Figure 4. Biological processes enriched by differentially expressed proteins between the lard group and the control group (Horizontal axis corresponds to count value, which represents the number of proteins enriched in this pathway. The color of the circle corresponds to the −log10(p) value).
Among the enriched pathways, there were some metabolism-related pathways, including the response to lipopolysaccharide, lipid metabolic process, glycoside catabolic process, and glutathione metabolic process. The nervous-system-related pathways that appeared frequently in the olive oil group and the butter group did not appear.
3.2.5 Group analysis of urinary protein composition in the hydrogenated vegetable oil group
3.2.5.1 Differentially expressed proteins
The urinary proteins of the hydrogenated vegetable oil group (group E) and the control group (group A) were compared. The screening criteria for differentially expressed proteins were: fold-change FC ≥ 1.5 or ≤ 0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 63 differentially expressed proteins were identified in the hydrogenated vegetable oil group, among which 34 were down-regulated and 29 were up-regulated. The differentially expressed proteins were arranged in ascending order of FC, and retrieved through Uniprot. The detailed information is listed in Appendix Table 4.
3.2.5.2 Functional analysis of differentially expressed proteins
The identified differentially expressed proteins were searched in the Pubmed database for relevant literature.
Among the down-regulated differentially expressed proteins in the hydrogenated vegetable oil group: Disabled homolog 2 (FC = 0, p = 4.72E-02) plays a crucial role in controlling macrophage phenotypic polarization and adipose tissue inflammation. Its deficiency leads to exacerbated adipose tissue inflammation and insulin resistance (49). Muscles with knockout of Calmodulin-like protein 3 (FC = 0.011, p = 4.55E-02) gene show weakened calcium release, reduced calmodulin kinase signaling, and impaired muscle adaptation to exercise (50). Integrin alpha-1 (FC = 0.036, p = 2.67E-02) can promote hepatic insulin action and lipid accumulation simultaneously under high-fat diet challenge, and plays a role in liver metabolism. Its deficiency can alter fatty acid metabolism in high-fat diet mice and improve fatty liver conditions (51). Alpha-2-HS-glycoprotein (FC = 0.116, p = 4.48E-02) is a multifunctional plasma glycoprotein mainly synthesized in the liver. It is considered an important part of various normal and pathological processes, including bone metabolism regulation, vascular calcification, insulin resistance, and protease activity control. During obesity and related complications such as type 2 diabetes, metabolic syndrome, and non-alcoholic fatty liver disease, the circulating level of Alpha-2-HS-glycoprotein is significantly increased (52). Eppin (FC = 0.192, p = 2.36E-02) is a biomarker of androgen action in human semen. Research has found that it is closely related to semen quality, sperm motility, etc. Down-regulation of Eppin expression significantly reduces sperm motility (53).
Among the up-regulated differentially expressed proteins: Phosphoserine phosphatase (FC = 14.636, p = 4.44E-02) is the protein with the largest up-regulation amplitude. No literature research has shown its correlation with metabolic activities. Catechol O-methyltransferase (FC = 3.841, p = 2.40E-02) plays a key role in metabolizing dopamine. Common functional polymorphisms in the gene of this protein affect cognitive functions related to the prefrontal cortex, sleep–wake regulation, and may affect sleep pathology (54).
3.2.5.3 Enrichment analysis of biological processes of differentially expressed proteins
The DAVID database was used to perform enrichment analysis of biological processes (BP) on the 63 differentially expressed proteins identified by group analysis. The results showed that a total of 16 BP pathways were significantly enriched (p < 0.05), and the detailed information is presented in Figure 5.
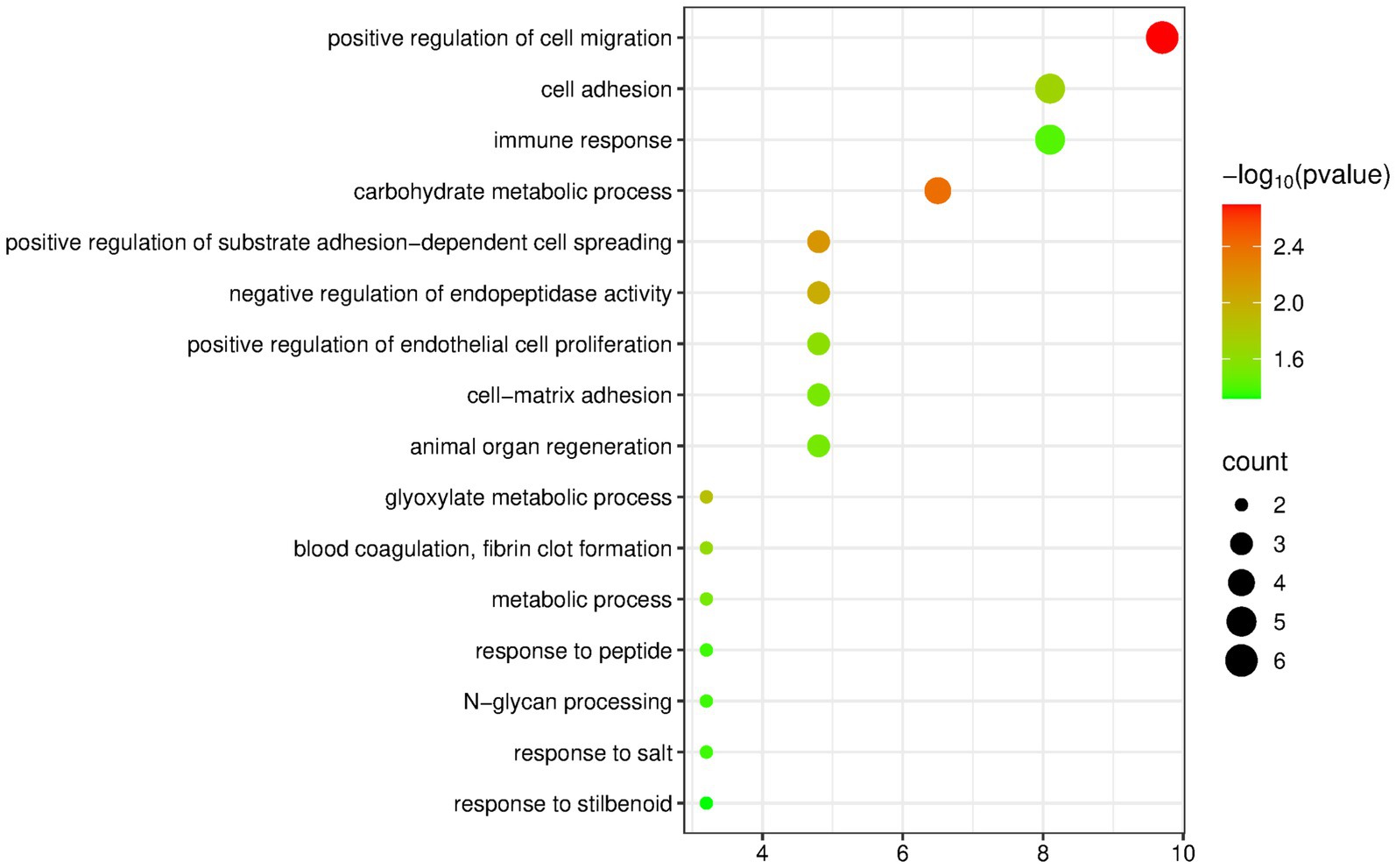
Figure 5. Biological processes enriched by differentially expressed proteins between the hydrogenated vegetable oil group and the control group (Horizontal axis corresponds to count value, which represents the number of proteins enriched in this pathway. The color of the circle corresponds to the −log10(p) value).
Among the enriched pathways, there were some metabolism-related pathways, including carbohydrate metabolic process, glyoxylate metabolic process, and metabolic process. Also, the nervous-system-related pathways that appeared frequently in the olive oil group and the butter group were not present.
3.2.6 Group analysis of urinary protein composition in the rapeseed oil group
3.2.6.1 Differentially expressed proteins
The urinary proteins of the rapeseed oil group (group F) and the control group (group A) were compared. The screening criteria for differentially expressed proteins were: fold-change FC ≥ 1.5 or ≤ 0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 105 differentially expressed proteins were identified in the rapeseed oil group, among which 53 were down-regulated and 52 were up-regulated. The differentially expressed proteins were arranged in ascending order of FC, and retrieved through Uniprot. The detailed information is listed in Appendix Table 5.
3.2.6.2 Functional analysis of differentially expressed proteins
The identified differentially expressed proteins were searched in the Pubmed database for relevant literature.
Among the down-regulated differentially expressed proteins in the rapeseed oil group, a total of 20 differentially expressed proteins had the largest down-regulation amplitude with a change from “present to absent.” Existing literature has proven that multiple of these proteins are related to immune-inflammation, oxidative stress, etc. Thrombospondin 4 (FC = 0, p = 4.46E-02) is a stress-inducible secreted glycoprotein that plays a crucial role in tissue injury and healing. It can activate the adaptive endoplasmic reticulum stress response and enhance sarcolemma stability in the heart and skeletal muscles. Defective flux of Thrombospondin 4 through the secretory pathway impairs the stability of the cardiomyocyte membrane and leads to cardiomyopathy (55). Interleukin 10 receptor subunit beta (FC = 0, p = 2.63E-02) is one of the components of the Interleukin 10 receptor, which is common to all members of the IL-10 cytokine family. Interleukin-10 is a type 2 T-helper cell cytokine with extensive anti-inflammatory effects (56, 57). Ring finger protein 150 (FC = 0, p = 2.83E-02) may be involved in regulating the antioxidant stress signaling pathway in neurons, enhancing the resistance of neurons to oxidative stress, and thus reducing neuronal damage and death (58). Interleukin 17 receptor A (FC = 0, p = 2.50E-02) is the receptor for interleukin-17A, which is a pro-inflammatory cytokine related to the rapid malignant progression and treatment resistance of colorectal cancer (59). Endothelial protein C receptor (FC = 0, p = 3.81E-02) plays a positive role in normal homeostasis, anticoagulant pathways, inflammation, and cell stemness, and is considered a potential effector or mediator of inflammatory diseases (60). The only known functional ligand of Cell surface glycoprotein CD200 receptor 1 (CD200R) (FC = 0, p = 4.12E-02) is CD200, and their interaction leads to the activation of anti-inflammatory signals in CD200R-expressing cells. When this interaction becomes insufficient due to aging or disease, chronic inflammation occurs (61). In addition, Protein O-linked-mannose beta-1,2-N-acetylglucosaminyltransferase 1 (POMGnT1) (FC = 0, p = 3.26E-02) is mainly expressed in neurons and also to a certain extent in glial cells. The expression of POMGnT1 decreases in mouse and cell models of Alzheimer’s disease (62).
Among the up-regulated differentially expressed proteins, Serine peptidase inhibitor Kunitz type 3 (FC = 43.172, p = 4.78E-02) had the largest fold-change. It is involved in processes such as blood coagulation and fibrinolysis, tumor immunity, inflammation regulation, and resistance to bacterial and fungal infections (63). CCN family member 1 (FC = 9.008, p = 3.96E-02) is an extracellular matrix protein that has a potential role in wound healing, accelerating re-epithelialization by promoting keratinocyte migration and proliferation (64).
3.2.6.3 Enrichment analysis of biological processes of differentially expressed proteins
The DAVID database was used to perform enrichment analysis of biological processes (BP) on the 105 differentially expressed proteins identified by group analysis. The results showed that a total of 51 BP pathways were significantly enriched (p < 0.05), and the detailed information is presented in Figure 6.
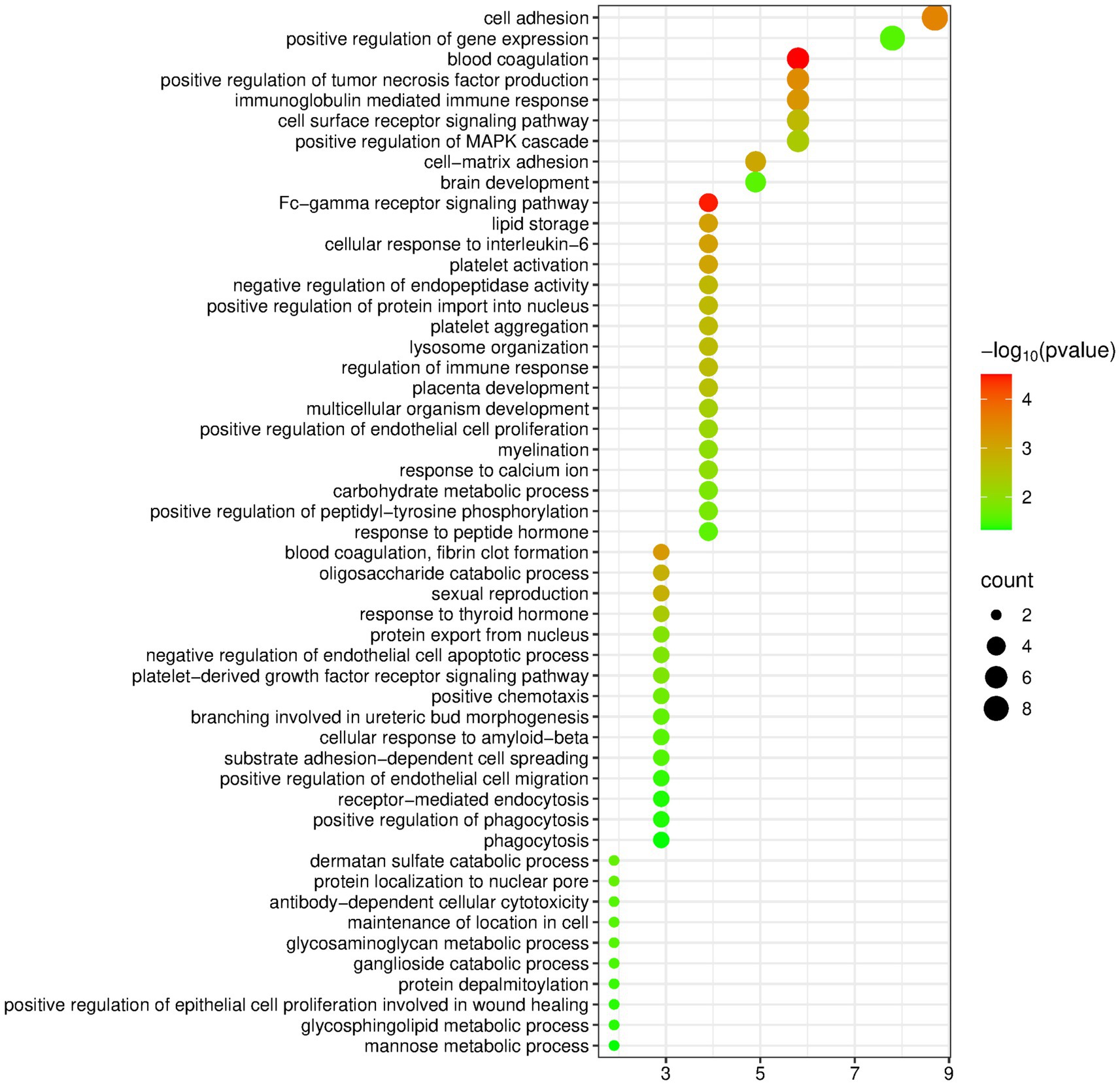
Figure 6. Biological processes enriched by differentially expressed proteins between the rapeseed oil group and the control group (Horizontal axis corresponds to count value, which represents the number of proteins enriched in this pathway. The color of the circle corresponds to the −log10(p) value).
Compared with other edible oil groups, the pathways enriched in the rapeseed oil group contained a large number of immune-related pathways, including the Fc-γ receptor signaling pathway, immunoglobulin-mediated immune response, cellular response to interleukin 6, regulation of the immune response, positive regulation of endothelial cell proliferation, negative regulation of the endothelial cell apoptotic process, antibody-dependent cell-mediated cytotoxicity, and positive regulation of endothelial cell migration. Studies have found that rapeseed oil shows certain positive effects in supporting the acquired immune capacity of weaned mice, especially in promoting the antibody response (65).
In addition, many metabolism-related pathways were also enriched, such as lipid storage, oligosaccharide catabolic process, carbohydrate metabolic process, etc. It is worth noting the pathway “cellular response to beta-amyloid protein.” Amyloid-beta aggregates in the brain play a central role in the pathogenesis of Alzheimer’s disease, and research has shown that consumption of vegetable oil can be used as a preventive or adjuvant strategy to slow down or prevent the progression of neurodegenerative diseases (66, 67).
3.2.7 Common differentially expressed proteins in the group analysis of different edible oils
The common differentially expressed proteins in the five-group experiments were counted (Figure 7). It was found that there were few common differentially expressed proteins, indicating that different edible oils have different impacts on the body. However, two proteins, Amyloid P component serum and 5′-3′ exonuclease PLD3, were up-regulated in the urine of all experimental groups. Amyloid P component serum is believed to potentially promote the progression of neurodegenerative diseases, including Alzheimer’s disease, by binding to β-amyloid protein to increase its stability and inducing neuronal apoptosis (68). 5′-3′ exonuclease PLD3 is highly expressed in brain neurons. Its level is down-regulated in the brains of Alzheimer’s disease patients and is negatively correlated with the levels of amyloid-precursor protein and amyloid-β protein. PLD3 may be involved in the pathogenesis of Alzheimer’s disease through the processing of amyloid-precursor protein (69).
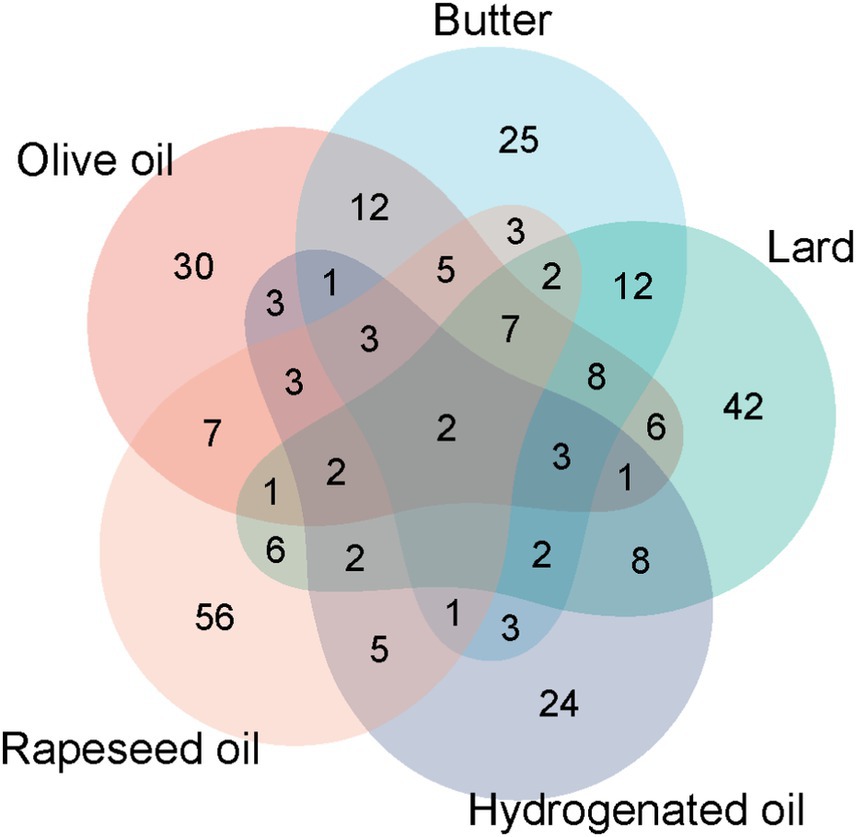
Figure 7. Common differentially expressed proteins in the urinary proteomes of the olive oil group, butter group, lard group, hydrogenated vegetable oil group, and rapeseed oil group.
3.3 Group analysis of urinary post-translational modifications
3.3.1 Random grouping
The control group samples (n = 5) and the experimental group samples (n = 5) were randomly divided into two groups, with a total of 126 grouping types. Among all the random combination types, the average number of post-translational modifications for all random times was calculated according to the same screening criteria. The ratio of the average number of post-translational modifications to the number of post-translational modifications obtained under normal grouping is the proportion of randomly generated post-translational modifications, as listed in Table 3. These results indicate that the proportion of randomly generated post-translational modifications is relatively large, suggesting that short-term intake of different edible oils has a relatively small impact on post-translational modifications in the rat body.

Table 3. Proportion of randomly generated differential modifications obtained from random grouping.
3.3.2 Group analysis of post-translational modifications in the olive oil group
The post-translational modifications of the olive oil group and the control group were compared. The screening criteria for differential modifications were: FC ≥ 1.5 or ≤0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 41 differential modifications were identified in the olive oil group, among which 17 were down-regulated and 24 were up-regulated. The differentially modified proteins were arranged in ascending order of FC, and the detailed information is listed in Appendix Table 6.
Among the proteins with differential modifications showing an up- or down-regulation of more than 10-fold: P97574, namely Stanniocalcin-1 (FC = 0, p = 4.59E-02), may be involved in the digestion and absorption in the gastrointestinal tract and kidneys of those parts with active digestion and absorption functions (70).
3.3.3 Group analysis of post-translational modifications in the butter group
The post-translational modifications of the butter group and the control group were compared. The screening criteria for differential modifications were: FC ≥ 1.5 or ≤0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 49 differential modifications were identified in the butter group, among which 4 were down-regulated and 45 were up-regulated. The differentially modified proteins were arranged in ascending order of FC, and the detailed information is listed in Appendix Table 7.
Among the proteins with differential modifications showing an up- or down-regulation of more than 10-fold: P02770, namely Albumin (FC = 37, p = 1.87E-02), plays a crucial role in transporting various endogenous and exogenous molecules and maintaining the colloid osmotic pressure of blood. It has many enzymatic activities, and its free thiol groups determine that this protein can participate in redox reactions (71). Q5FVF9, namely Biotinidase (FC = 24, p = 4.78E-02), is an enzyme that helps the body reuse and recycle biotin in food (72).
3.3.4 Group analysis of post-translational modifications in the lard group
The post-translational modifications of the lard group and the control group were compared. The screening criteria for differential modifications were: FC ≥ 1.5 or ≤0.67, and two-tailed paired t-test p < 0.05. The results showed that, compared with the control group, a total of 99 differential modifications were identified in the lard group, among which 23 were down-regulated and 76 were up-regulated. The differentially modified proteins were arranged in ascending order of FC, and the detailed information is listed in Appendix Table 8.
Among the proteins with differential modifications showing an up- or down-regulation of more than 10-fold: P07151, namely Beta-2-microglobulin (FC = 15, p = 8.64E-03), may play a role in the oxidative stress state of the elderly and can be used as a new biomarker of oxidative stress (73). P11232, namely Thioredoxin (FC = 0.091, p = 3.41E-02), plays an important role in maintaining the redox state of cells and regulating redox signal transduction. Together with glutathione, it plays a central role in combating oxidative stress (74).
3.3.5 Group analysis of post-translational modifications in the hydrogenated vegetable oil group
The post-translational modifications of the hydrogenated vegetable oil group were compared with those of the control group. The screening criteria for differential modifications were set as FC ≥ 1.5 or ≤ 0.67, and a two-tailed paired t-test with p < 0.05. The results indicated that, compared to the control group, a total of 50 differential modifications were identified in the hydrogenated vegetable oil group, among which 38 were down-regulated and 12 were up-regulated. The differentially modified proteins were arranged in ascending order of FC, and the detailed information is presented in Appendix Table 9.
Among the proteins with differential modifications showing an up- or down-regulation of more than 10-fold, P27590, namely Uromodulin (FC = 0.053, p = 4.91E-02), has multiple functions in kidney physiology, renal tubular transport, and mineral metabolism. A higher serum concentration of Uromodulin is associated with higher renal function and preserved renal reserve. It is also related to a lower risk of cardiovascular diseases and diabetes (75).
3.3.6 Group analysis of post-translational modifications in the rapeseed oil group
The post-translational modifications of the rapeseed oil group were compared with those of the control group. The differential modification screening criteria were FC ≥ 1.5 or ≤ 0.67, and a two-tailed paired t-test with p < 0.05. The results showed that, compared to the control group, a total of 50 differential modifications were identified in the rapeseed oil group, with 38 down-regulated and 12 up-regulated. The differentially modified proteins were arranged in ascending order of FC, and the detailed information is listed in Appendix Table 10.
Among the proteins with differential modifications showing an up- or down-regulation of more than 10-fold: P07632, namely Superoxide dismutase (FC = 0, p = 2.95E-02), can neutralize superoxide free radicals and protect organisms from oxidative stress, playing an important role in the antioxidant process (76). P47853, namely Biglycan (FC = 0.045, p = 4.47E-02), can reduce high-fat-diet-induced obesity and improve glucose tolerance in mice (77).
4 Discussion
This study systematically characterized the differential impacts of short-term consumption of various edible oils on the rat urinary proteome using label-free quantitative proteomics. The results showed that all edible oil groups triggered significant changes in metabolic-related protein pathways. In addition, different oils had specific effects on biological pathways: the olive oil and butter groups were significantly enriched in nervous system-related pathways (such as axon guidance and synaptic assembly regulation), while the rapeseed oil group was more involved in immune regulation pathways (such as Fc-γ receptor signaling and cellular response to interleukin-6). These differential effects likely reflect the unique bioactive compounds in each oil, such as hydroxytyrosol in olive oil and phytosterols in rapeseed oil. For example, hydroxytyrosol in olive oil has been proven to have a neuroprotective effect. It can prevent the aggregation of β-amyloid protein and oxidative stress in Alzheimer’s disease through the overexpression of SKN-1/NRF2 transcription factors and HSP-16.2 (78, 79); the phytosterols rich in rapeseed oil may enhance immune homeostasis by regulating T cell function (80).
In addition, the consistent upregulation of two proteins—serum amyloid P component and 5′-3′ exonuclease PLD3—in all experimental groups warrants further investigation. The former, as a stabilizer of β-amyloid protein, may suggest the potential impact of edible oil intake on the risk of neurodegeneration (68); the downregulation of 5′-3′ exonuclease PLD3 in the Alzheimer’s disease model is in contrast to its upregulation in this study, suggesting that dietary components may be involved in neuroprotection or pathological processes by regulating the processing of amyloid precursor protein. The causal relationship needs to be verified through long-term intervention experiments (69).
At the level of post-translational modifications, the results of random grouping showed that a large proportion of post-translational modifications were randomly generated. The limited overall modification changes may reflect the weak direct impact of short-term intake of different edible oils on the protein structure in the rat body. It is worth noting that currently, there is still a lack of a systematic understanding of the specific biological significance of most post-translational modifications in the context of this study, which makes it impossible for us to conduct in-depth discussions on some modification events for the time being. This limitation indicates that in future follow-up studies, it is necessary to combine targeted modification proteomics with functional verification methods to clarify the regulatory mechanism of key modification sites on metabolic pathways and disease risks.
In addition, this study has certain limitations. Firstly, the absence of multiple testing correction may increase the risk of false positives. However, in this exploratory study with a small sample size, this choice was made after careful consideration, aiming to avoid an excessive number of false negative results. In this study, the biological significance and reliability of the research results were strengthened through random grouping, functional annotation, and cross-validation with existing literature. Future validation in a larger cohort and the adoption of more stringent statistical adjustment methods will further confirm the reliability of these preliminary results. Secondly, we did not precisely quantify the changes in energy intake after adding edible oils to the diet of rats, and failed to calculate the percentage increase in daily energy intake. However, it was actually observed that compared with the control group, the food intake of rats increased after they consumed edible oils, and their total energy intake was higher than that of the control group. Excessive energy intake is highly likely to interfere with the urinary proteome, which serves as the observation index in this study, and thus affects the experimental results.
5 Conclusion
This study for the first time revealed the regulatory effect of short-term dietary oil intervention on the multi-dimensional physiological network of the body from the perspective of the urinary proteome, providing a molecular basis for personalized dietary recommendations. The urinary proteome of rats showed obvious changes after 1 week of edible oil intake, and there were significant differences in the effects of different types of edible oils on the urinary proteome of rats, and their effects on the rat body were comprehensive. Compared with the urinary proteome itself, the changes in post-translational modifications of the proteome were relatively small, indicating that the intake of edible oils has a limited impact on post-translational modifications of proteins in the short term. However, the limitations of extrapolating from animal models to humans, the relatively short intervention period, and the lack of mechanistic research still need to be improved through long-term human cohort studies and cell experiments.
Data availability statement
The original contributions presented in the study are publicly available. The mass spectrometry proteomics data are available at the ProteomeXchange Consortium via the iProX partner with the dataset identifier IPX0011315000 (https://www.iprox.cn//page/SCV017.html?query=IPX0011315000).
Ethics statement
The animal study was approved by the Ethics Committee of the College of Life Sciences, Beijing Normal University. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
YS: Conceptualization, Formal analysis, Investigation, Methodology, Validation, Visualization, Writing – original draft. YG: Conceptualization, Data curation, Funding acquisition, Methodology, Project administration, Resources, Software, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the National Key R&D Program of China (2023YFA1801900), Beijing Natural Science Foundation (L246002), Beijing Normal University (11100704).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2025.1571846/full#supplementary-material
Footnotes
References
1. DiNicolantonio, JJ, and O'Keefe, JH. Importance of maintaining a low omega-6/omega-3 ratio for reducing inflammation. Open Heart. (2018) 5:e000946. doi: 10.1136/openhrt-2018-000946
2. Petersen, KS, Maki, KC, Calder, PC, Belury, MA, Messina, M, Kirkpatrick, CF, et al. Perspective on the health effects of unsaturated fatty acids and commonly consumed plant oils high in unsaturated fat - CORRIGENDUM. Br J Nutr. (2024) 132:1051. doi: 10.1017/S0007114524002915
3. Xia, M, Zhong, Y, Peng, Y, and Qian, C. Olive oil consumption and risk of cardiovascular disease and all-cause mortality: a meta-analysis of prospective cohort studies. Front Nutr. (2022) 9:1041203. doi: 10.3389/fnut.2022.1041203
4. Hooper, L, Martin, N, Jimoh, OF, Kirk, C, Foster, E, and Abdelhamid, AS. Reduction in saturated fat intake for cardiovascular disease. Cochrane Database Syst Rev. (2020) 5:CD011737. doi: 10.1002/14651858.CD011737.pub2
5. Siri-Tarino, PW, Sun, Q, Hu, FB, and Krauss, RM. Meta-analysis of prospective cohort studies evaluating the association of saturated fat with cardiovascular disease. Am J Clin Nutr. (2010) 91:535–46. doi: 10.3945/ajcn.2009.27725
6. Zhu, Y, Bo, Y, and Liu, Y. Dietary total fat, fatty acids intake, and risk of cardiovascular disease: a dose-response meta-analysis of cohort studies. Lipids Health Dis. (2019) 18:91. doi: 10.1186/s12944-019-1035-2
7. Yan, S, Liu, S, Qu, J, Li, X, Hu, J, Zhang, L, et al. A lard and soybean oil mixture alleviates low-fat-high-carbohydrate diet-induced nonalcoholic fatty liver disease in mice. Nutrients. (2022) 14:560. doi: 10.3390/nu14030560
8. Li, H, Zhu, Y, Zhao, F, Song, S, Li, Y, Xu, X, et al. Fish oil, lard and soybean oil differentially shape gut microbiota of middle-aged rats. Sci Rep. (2017) 7:826. doi: 10.1038/s41598-017-00969-0
9. Mozaffarian, D, Katan, MB, Ascherio, A, Stampfer, MJ, and Willett, WC. Trans fatty acids and cardiovascular disease. N Engl J Med. (2006) 354:1601–13. doi: 10.1056/NEJMra054035
10. Shen, J, Liu, Y, Wang, X, Bai, J, Lin, L, Luo, F, et al. A comprehensive review of health-benefiting components in rapeseed oil. Nutrients. (2023) 15:999. doi: 10.3390/nu15040999
11. Pisitkun, T, Johnstone, R, and Knepper, MA. Discovery of urinary biomarkers. Mol Cell Proteomics. (2006) 5:1760–71. doi: 10.1074/mcp.R600004-MCP200
12. Wei, J, and Gao, Y. Early disease biomarkers can be found using animal models urine proteomics. Expert Rev Proteomics. (2021) 18:363–78. doi: 10.1080/14789450.2021.1937133
13. Liu, CH, Zheng, S, Wang, S, Wu, D, Jiang, W, Zeng, Q, et al. Urine proteome in distinguishing hepatic steatosis in patients with metabolic-associated fatty liver disease. Diagnostics (Basel). (2022) 12:1412. doi: 10.3390/diagnostics12061412
14. Seol, W, Kim, H, and Son, I. Urinary biomarkers for neurodegenerative diseases. Exp Neurobiol. (2020) 29:325–33. doi: 10.5607/en20042
15. Bi, X, Liu, W, Ding, X, Liang, S, Zheng, Y, Zhu, X, et al. Proteomic and metabolomic profiling of urine uncovers immune responses in patients with COVID-19. Cell Rep. (2022) 38:110271. doi: 10.1016/j.celrep.2021.110271
16. Walsh, CT, Garneau-Tsodikova, S, and Gatto, GJ Jr. Protein posttranslational modifications: the chemistry of proteome diversifications. Angew Chem Int Ed Engl. (2005) 44:7342–72. doi: 10.1002/anie.200501023
17. Tabernero, L, Aricescu, AR, Jones, EY, and Szedlacsek, SE. Protein tyrosine phosphatases: structure-function relationships. FEBS J. (2008) 275:867–82. doi: 10.1111/j.1742-4658.2008.06251.x
18. Choudhary, C, Kumar, C, Gnad, F, Nielsen, ML, Rehman, M, Walther, TC, et al. Lysine acetylation targets protein complexes and co-regulates major cellular functions. Science. (2009) 325:834–40. doi: 10.1126/science.1175371
19. Štambuk, T, and Gornik, O. Protein glycosylation in diabetes. Adv Exp Med Biol. (2021) 1325:285–305. doi: 10.1007/978-3-030-70115-4_14
20. Catalán, Ú, Rubió, L, de Las, L, Hazas, MC, Herrero, P, Nadal, P, et al. Hydroxytyrosol and its complex forms (secoiridoids) modulate aorta and heart proteome in healthy rats: potential cardio-protective effects. Mol Nutr Food Res. (2016) 60:2114–29. doi: 10.1002/mnfr.201600052
21. Rudman, N, Gornik, O, and Lauc, G. Altered N-glycosylation profiles as potential biomarkers and drug targets in diabetes. FEBS Lett. (2019) 593:1598–615. doi: 10.1002/1873-3468.13495
22. Sen, A. Scaffolding protein IQ motif containing GTPase activating protein 2 regulates liver metabolic homeostasis. FASEB J. (2020) 34:1. doi: 10.1096/fasebj.2020.34.s1.00664
23. Shetty, S, and Copeland, PR. Molecular mechanism of selenoprotein P synthesis. Biochim Biophys Acta Gen Subj. (2018) 1862:2506–10. doi: 10.1016/j.bbagen.2018.04.011
24. Christians, ES, Ishiwata, T, and Benjamin, IJ. Small heat shock proteins in redox metabolism: implications for cardiovascular diseases. Int J Biochem Cell Biol. (2012) 44:1632–45. doi: 10.1016/j.biocel.2012.06.006
25. Grewal, R, Reutzel, M, Dilberger, B, Hein, H, Zotzel, J, Marx, S, et al. Purified oleocanthal and ligstroside protect against mitochondrial dysfunction in models of early Alzheimer's disease and brain ageing. Exp Neurol. (2020) 328:113248. doi: 10.1016/j.expneurol.2020.113248
26. Scholz, A, Plate, KH, and Reiss, Y. Angiopoietin-2: a multifaceted cytokine that functions in both angiogenesis and inflammation. Ann N Y Acad Sci. (2015) 1347:45–51. doi: 10.1111/nyas.12726
27. Jiang, P, Ning, J, Yu, W, Rao, T, Ruan, Y, and Cheng, F. FLRT2 suppresses bladder cancer progression through inducing ferroptosis. J Cell Mol Med. (2024) 28:e17855. doi: 10.1111/jcmm.17855
28. Mishima, M, Sakai, Y, Itoh, N, Kamiya, H, Furuichi, M, Takahashi, M, et al. Structure of human MTH1, a Nudix family hydrolase that selectively degrades oxidized purine nucleoside triphosphates. J Biol Chem. (2004) 279:33806–15. doi: 10.1074/jbc.M402393200
29. Barbalace, MC, Zallocco, L, Beghelli, D, Ronci, M, Scortichini, S, Digiacomo, M, et al. Antioxidant and neuroprotective activity of extra virgin olive oil extracts obtained from Quercetano cultivar trees grown in different areas of the Tuscany region (Italy). Antioxidants. (2021) 10:421. doi: 10.3390/antiox10030421
30. Villareal, MO, Sasaki, K, Margout, D, Savry, C, Almaksour, Z, Larroque, M, et al. Neuroprotective effect of Picholine virgin olive oil and its hydroxycinnamic acids component against β-amyloid-induced toxicity in SH-SY5Y neurotypic cells. Cytotechnology. (2016) 68:2567–78. doi: 10.1007/s10616-016-9980-3
31. Lambert de Malezieu, M, Courtel, P, Sleno, L, Abasq, ML, and Ramassamy, C. Synergistic properties of bioavailable phenolic compounds from olive oil: electron transfer and neuroprotective properties. Nutr Neurosci. (2021) 24:660–73. doi: 10.1080/1028415X.2019.1666480
32. Gholamalizadeh, H, Ensan, B, Karav, S, Jamialahmadi, T, and Sahebkar, A. Regulatory effects of statins on CCL2/CCR2 axis in cardiovascular diseases: new insight into pleiotropic effects of statins. J Inflamm. (2024) 21:51. doi: 10.1186/s12950-024-00420-y
33. León, A, Aparicio, GI, and Scorticati, C. Neuronal glycoprotein M6a: an emerging molecule in chemical synapse formation and dysfunction. Front Synaptic Neurosci. (2021) 13:661681. doi: 10.3389/fnsyn.2021.661681
34. Solovyev, N. Selenoprotein P and its potential role in Alzheimer's disease. Hormones (Athens). (2020) 19:73–9. doi: 10.1007/s42000-019-00112-w
35. Menghini, R, Casagrande, V, Menini, S, Marino, A, Marzano, V, Hribal, ML, et al. TIMP3 overexpression in macrophages protects from insulin resistance, adipose inflammation, and nonalcoholic fatty liver disease in mice. Diabetes. (2012) 61:454–62. doi: 10.2337/db11-0613
36. Herrera-Marcos, LV, Lou-Bonafonte, JM, Martinez-Gracia, MV, Arnal, C, Navarro, MA, and Osada, J. Prenylcysteine oxidase 1, a pro-oxidant enzyme of low density lipoproteins. Front Biosci. (2018) 23:1020–37. doi: 10.2741/4631
37. Tian, X, Yan, C, and Han, Y. Cellular repressor of E1A-stimulated genes, a new potential therapeutic target for atherosclerosis. Curr Drug Targets. (2017) 18:1800–4. doi: 10.2174/1389450117666161026111250
38. Liu, Y, Zhao, W, Gu, G, Lu, L, Feng, J, Guo, Q, et al. Palmitoyl-protein thioesterase 1 (PPT1): an obesity-induced rat testicular marker of reduced fertility. Mol Reprod Dev. (2014) 81:55–65. doi: 10.1002/mrd.22281
39. Jin, DZ, Mao, LM, and Wang, JQ. The role of extracellular signal-regulated kinases (ERK) in the regulation of mGlu5 receptors in neurons. J Mol Neurosci. (2018) 66:629–38. doi: 10.1007/s12031-018-1193-0
40. Wright, NT, Cannon, BR, Zimmer, DB, and Weber, DJ. S100A1: structure, function, and therapeutic potential. Curr Chem Biol. (2009) 3:138–45. doi: 10.2174/187231309788166460
41. Zhang, L, Sun, C, Jin, Y, Gao, K, Shi, X, Qiu, W, et al. Dickkopf 3 (Dkk3) improves amyloid-β pathology, cognitive dysfunction, and cerebral glucose metabolism in a transgenic mouse model of Alzheimer's disease. J Alzheimers Dis. (2017) 60:733–46. doi: 10.3233/JAD-161254
42. Lizarraga, F, Espinosa, M, Ceballos-Cancino, G, Vazquez-Santillan, K, Bahena-Ocampo, I, Schwarz-Cruz, Y, et al. Tissue inhibitor of metalloproteinases-4 (TIMP-4) regulates stemness in cervical cancer cells. Mol Carcinog. (2016) 55:1952–61. doi: 10.1002/mc.22442
43. Guo, Z, Zhao, Y, Wu, Y, Zhang, Y, Wang, R, Liu, W, et al. Cellular retinol-binding protein 1: a therapeutic and diagnostic tumor marker. Mol Biol Rep. (2023) 50:1885–94. doi: 10.1007/s11033-022-08179-2
44. Zhong, M, Kawaguchi, R, Ter-Stepanian, M, Kassai, M, and Sun, H. Vitamin a transport and the transmembrane pore in the cell-surface receptor for plasma retinol binding protein. PLoS One. (2013) 8:e73838. doi: 10.1371/journal.pone.0073838
45. Yang, H, Hu, B, Wang, X, Chen, W, and Zhou, H. The effects of hyaluronan and proteoglycan link protein 1 (HAPLN1) in ameliorating spinal cord injury mediated by Nrf2. Biotechnol Appl Biochem. (2024) 71:929–39. doi: 10.1002/bab.2587
46. Brown, CW, Chhoy, P, Mukhopadhyay, D, Karner, ER, and Mercurio, AM. Targeting prominin2 transcription to overcome ferroptosis resistance in cancer. EMBO Mol Med. (2021) 13:e13792. doi: 10.15252/emmm.202013792
47. Luo, J, Xie, M, Peng, C, Ma, Y, Wang, K, Lin, G, et al. Protein disulfide isomerase A6 promotes the repair of injured nerve through interactions with spastin. Front Mol Neurosci. (2022) 15:950586. doi: 10.3389/fnmol.2022.950586
48. Pucci, L, Perozzi, S, Cimadamore, F, Orsomando, G, and Raffaelli, N. Tissue expression and biochemical characterization of human 2-amino 3-carboxymuconate 6-semialdehyde decarboxylase, a key enzyme in tryptophan catabolism. FEBS J. (2007) 274:827–40. doi: 10.1111/j.1742-4658.2007.05635.x
49. Adamson, SE, Griffiths, R, Moravec, R, Senthivinayagam, S, Montgomery, G, Chen, W, et al. Disabled homolog 2 controls macrophage phenotypic polarization and adipose tissue inflammation. J Clin Invest. (2016) 126:1311–22. doi: 10.1172/JCI79590
50. Ermolova, N, Kramerova, I, and Spencer, MJ. Autolytic activation of calpain 3 proteinase is facilitated by calmodulin protein. J Biol Chem. (2015) 290:996–1004. doi: 10.1074/jbc.M114.588780
51. Williams, AS, Kang, L, Zheng, J, Grueter, C, Bracy, DP, James, FD, et al. Integrin α1-null mice exhibit improved fatty liver when fed a high fat diet despite severe hepatic insulin resistance. J Biol Chem. (2015) 290:6546–57. doi: 10.1074/jbc.M114.615716
52. Bourebaba, L, and Marycz, K. Pathophysiological implication of Fetuin-a glycoprotein in the development of metabolic disorders: a concise review. J Clin Med. (2019) 8:2033. doi: 10.3390/jcm8122033
53. Xu, J, He, M, Wang, W, Hou, J, Chen, X, Ding, X, et al. siRNA-mediated Eppin testicular silencing causes changes in sperm motility and calcium currents in mice. Reprod Biol. (2021) 21:100485. doi: 10.1016/j.repbio.2021.100485
54. Dauvilliers, Y, Tafti, M, and Landolt, HP. Catechol-O-methyltransferase, dopamine, and sleep-wake regulation. Sleep Med Rev. (2015) 22:47–53. doi: 10.1016/j.smrv.2014.10.006
55. Brody, MJ, Vanhoutte, D, Schips, TG, Boyer, JG, Bakshi, CV, Sargent, MA, et al. Defective flux of Thrombospondin-4 through the secretory pathway impairs cardiomyocyte membrane stability and causes cardiomyopathy. Mol Cell Biol. (2018) 38:e00114–8. doi: 10.1128/MCB.00114-18
56. Geginat, J. Introduction to the special issue: Interleukin-10 "the surprising twists and turns of an anti-inflammatory cytokine on its way to the clinic". Semin Immunol. (2019) 44:101343. doi: 10.1016/j.smim.2019.101343
57. He, JQ, Shumansky, K, Zhang, X, Connett, JE, Anthonisen, NR, and Sandford, AJ. Polymorphisms of interleukin-10 and its receptor and lung function in COPD. Eur Respir J. (2007) 29:1120–6. doi: 10.1183/09031936.00002907
58. Zhao, C, Rispe, C, and Nabity, PD. Secretory RING finger proteins function as effectors in a grapevine galling insect. BMC Genomics. (2019) 20:923. doi: 10.1186/s12864-019-6313-x
59. Wang, K, Kim, MK, Di Caro, G, Wong, J, Shalapour, S, Wan, J, et al. Interleukin-17 receptor a signaling in transformed enterocytes promotes early colorectal tumorigenesis. Immunity. (2014) 41:1052–63. doi: 10.1016/j.immuni.2014.11.009
60. O'Hehir, ZD, Lynch, T, O'Neill, S, March, L, and Xue, M. Endothelial protein C receptor and its impact on rheumatic disease. J Clin Med. (2024) 13:2030. doi: 10.3390/jcm13072030
61. Walker, DG, and Lue, LF. Understanding the neurobiology of CD200 and the CD200 receptor: a therapeutic target for controlling inflammation in human brains? Future Neurol. (2013) 8:321–32. doi: 10.2217/fnl.13.14
62. Feng, Y, Jiang, H, Li, G, He, G, and Li, X. Decreased expression of protein O-linked mannose β-1,2-N-acetylglucosaminyltransferase 1 contributes to Alzheimer's disease-like pathologies. J Neurophysiol. (2022) 127:1067–74. doi: 10.1152/jn.00362.2021
63. Liu, Y, Jiang, S, Li, Q, and Kong, Y. Advances of Kunitz-type serine protease inhibitors. Sheng Wu Gong Cheng Xue Bao. (2021) 37:3988–4000. doi: 10.13345/j.cjb.200802
64. Du, H, Zhou, Y, Suo, Y, Liang, X, Chai, B, Duan, R, et al. CCN1 accelerates re-epithelialization by promoting keratinocyte migration and proliferation during cutaneous wound healing. Biochem Biophys Res Commun. (2018) 505:966–72. doi: 10.1016/j.bbrc.2018.09.001
65. Hillyer, LM, and Woodward, B. A comparison of the capacity of six cold-pressed plant oils to support development of acquired immune competence in the weanling mouse: superiority of low-linoleic-acid oils. Br J Nutr. (2002) 88:171–81. doi: 10.1079/BJNBJN2002602
66. Tiwari, S, Atluri, V, Kaushik, A, Yndart, A, and Nair, M. Alzheimer's disease: pathogenesis, diagnostics, and therapeutics. Int J Nanomedicine. (2019) 14:5541–54. doi: 10.2147/IJN.S200490
67. Hashempour-Baltork, F, Farshi, P, Mirza Alizadeh, A, Eskandarzadeh, S, Abedinzadeh, S, Azadmard-Damirchi, S, et al. Effect of refined edible oils on neurodegenerative disorders. Adv Pharm Bull. (2023) 13:461–8. doi: 10.34172/apb.2023.060
68. Urbányi, Z, Forrai, E, Sárvári, M, Likó, I, Illés, J, and Pázmány, T. Glycosaminoglycans inhibit neurodegenerative effects of serum amyloid P component in vitro. Neurochem Int. (2005) 46:471–7. doi: 10.1016/j.neuint.2004.12.001
69. Wang, J, Yu, JT, and Tan, L. PLD3 in Alzheimer's disease. Mol Neurobiol. (2015) 51:480–6. doi: 10.1007/s12035-014-8779-5
70. Kobayashi, R, Nakagomi, Y, Shimura, Y, Mochizuki, M, Kobayashi, K, Sugita, K, et al. Expression of stanniocalcin-1 in gastrointestinal tracts of neonatal and mature rats. Biochem Biophys Res Commun. (2009) 389:478–83. doi: 10.1016/j.bbrc.2009.08.169
71. Bocedi, A, Cattani, G, Stella, L, Massoud, R, and Ricci, G. Thiol disulfide exchange reactions in human serum albumin: the apparent paradox of the redox transitions of Cys34. FEBS J. (2018) 285:3225–37. doi: 10.1111/febs.14609
72. Procter, M, Wolf, B, Crockett, DK, and Mao, R. The biotinidase gene variants registry: a paradigm public database. G3. (2013) 3:727–31. doi: 10.1534/g3.113.005835
73. Althubiti, M, Elzubier, M, Alotaibi, GS, Althubaiti, MA, Alsadi, HH, Alhazmi, ZA, et al. Beta 2 microglobulin correlates with oxidative stress in elderly. Exp Gerontol. (2021) 150:111359. doi: 10.1016/j.exger.2021.111359
74. Jaganjac, M, Milkovic, L, Sunjic, SB, and Zarkovic, N. The NRF2, Thioredoxin, and glutathione system in tumorigenesis and anticancer therapies. Antioxidants. (2020) 9:1151. doi: 10.3390/antiox9111151
75. Wolf, MTF, Zhang, J, and Nie, M. Uromodulin in mineral metabolism. Curr Opin Nephrol Hypertens. (2019) 28:481–9. doi: 10.1097/MNH.0000000000000522
76. Saxena, P, Selvaraj, K, Khare, SK, and Chaudhary, N. Superoxide dismutase as multipotent therapeutic antioxidant enzyme: role in human diseases. Biotechnol Lett. (2022) 44:1–22. doi: 10.1007/s10529-021-03200-3
77. Chung, I, Kim, SA, Kim, S, Lee, JO, Park, CY, Lee, J, et al. Biglycan reduces body weight by regulating food intake in mice and improves glucose metabolism through AMPK/AKT dual pathways in skeletal muscle. FASEB J. (2021) 35:e21794. doi: 10.1096/fj.202002039RR
78. De La Cruz, JP, Ruiz-Moreno, MI, Guerrero, A, Reyes, JJ, Benitez-Guerrero, A, Espartero, JL, et al. Differences in the neuroprotective effect of orally administered virgin olive oil (Olea europaea) polyphenols tyrosol and hydroxytyrosol in rats. J Agric Food Chem. (2015) 63:5957–63. doi: 10.1021/acs.jafc.5b00627
79. Romero-Márquez, JM, Navarro-Hortal, MD, Jiménez-Trigo, V, Muñoz-Ollero, P, Forbes-Hernández, TY, Esteban-Muñoz, A, et al. An olive-derived extract 20% rich in Hydroxytyrosol prevents β-amyloid aggregation and oxidative stress, two features of Alzheimer disease, via SKN-1/NRF2 and HSP-16.2 in Caenorhabditis elegans. Antioxidants. (2022) 11:629. doi: 10.3390/antiox11040629
Keywords: urinary proteome, olive oil, butter, lard, hydrogenated vegetable oil, rapeseed oil, post-translational modification
Citation: Su Y and Gao Y (2025) Effects of one-week intake of different edible oils on the urinary proteome of rats. Front. Nutr. 12:1571846. doi: 10.3389/fnut.2025.1571846
Edited by:
B. Gowda Siddabasave Gowda, Hokkaido University, JapanReviewed by:
Wantao Ying, Beijing Proteome Research Center, ChinaYumi Watanabe, Niigata University, Japan
Ken-Ichi Kobayashi, Notre Dame Seishin University, Japan
Copyright © 2025 Su and Gao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Youhe Gao, Z2FveW91aGVAYm51LmVkdS5jbg==