 Gabriela Ibieta1,2*
Gabriela Ibieta1,2* Jimena Ortiz-Sempértegui1,2
Jimena Ortiz-Sempértegui1,2 Carl Grey1
Carl Grey1 J. Mauricio Peñarrieta2
J. Mauricio Peñarrieta2 Javier A. Linares-Pastén1*
Javier A. Linares-Pastén1*- 1Biotechnology and Applied Microbiology, Faculty of Engineering LTH, Lund University, Lund, Sweden
- 2Instituto de Investigaciones Químicas IIQ, Universidad Mayor de San Andrés UMSA, La Paz, Bolivia
High nutritional value Andean seeds—tarwi (Lupinus mutabilis), cañihua (Chenopodium pallidicaule), and quinoa (Chenopodium quinoa)—were subjected to solid-state fermentation with Levilactobacillus brevis DSM 1269. This strain can convert L-glutamic acid into the neurotransmitter GABA. Fermented tarwi exhibited the highest GABA production, at 4 mg/g sample, which correlates with its higher protein content compared to fermented quinoa and cañihua, at 1 mg/g and 0.3 mg/g, respectively. Seeds kept at room temperature before fermentation produced higher concentrations of GABA compared to seeds kept at 4 °C. Autoclaving, a mandatory step for fermentation, resulted in a decrease in L-glutamic acid in tarwi seeds and an increase in quinoa and cañihua seeds. Additionally, fermentation produced lactic acid and acetic acid, together with an increase in the content of free essential amino acids, including threonine, histidine, methionine, isoleucine, leucine, valine, and lysine. This work demonstrated, for the first time, the functional valorisation of tarwi, cañihua, and quinoa through the production of bioactive metabolites and the enhancement of essential free amino acids via fermentation with L. brevis.
1 Introduction
Fermentation is an ancient process originally developed to extend the shelf life and improve the sensory properties of food (1). Today, fermentation is a biotechnological tool used for developing new food products with enhanced nutritional and functional properties (2). Lactic acid bacteria (LAB) strains are of interest in food matrices fermentation because they can improve the physicochemical and sensory properties, also can produce enzymes and bioactive metabolites such as peptides, exopolysaccharides, bacteriocins, vitamins, short-chain fatty acids (SCFAs), γ-amino butyric acid (GABA), and other metabolites (3–5). Additionally, many LAB strains are considered probiotics, conferring beneficial effects on human health.
Fermentation also affects the proteins and amino acid profile (6). It is particularly interesting for improving the nutritional and functional value of plant proteins. Some LAB strains express proteases that can release peptides and free amino acids from the food matrices, increasing the content of free amino acids. On the other hand, the content of free amino acids can also decrease if the strain utilises them in its own metabolism (7). Therefore, quantifying the free amino acid profile before and after fermentation provides insights into the microorganism’s activity towards proteins. A desirable amino acid profile can include the increment of essential amino acids or precursors of bioactive metabolites, for example, tryptophan that later can be converted into serotonin, or glutamic acid, which can be converted into GABA.
GABA is a non-protein amino acid that has gained interest due to its numerous health benefits for humans (8). GABA is the primary inhibitory neurotransmitter in the mature central nervous system (9). It has also been proven to prevent and treat diseases, including diabetes, depression, anxiety, and chronic insomnia (10–13). Another important property of GABA is its role as a mediator in the gut–brain axis (14).
Levilactobacillus brevis has been studied by many authors as an efficient GABA-producing strain. It produces GABA primarily as a mechanism to tolerate and survive acidic conditions within its environment (15). For example, L. brevis strains isolated from Chinese pickles (LBG-29 and LBG-24) could be utilised to enhance beverage functionality due to their high GABA production (>300 mg/L) (8). Another study demostrated the production of GABA in soybean via fermentation with Levilactobacillus brevis NPS-QW 145, resulting in 2.3 g/L of GABA, which can be used to produce a yoghurt-like product with enhanced GABA levels (16). Studies have also shown that L. brevis CRL 2013 does not possess antibiotic resistance genes and virulence markers in its genome, making it safe for probiotic applications (17).
Seeds, such as beans, rice, and sesame, are natural sources of GABA (18). However, in seeds with low or undetectable GABA levels, its content can be increased through two primary approaches: germination or fermentation. In this study, three Andean seeds with undetectable GABA levels —tarwi (Lupinus mutabilis), cañihua (Chenopodium pallidicaule), and quinoa (Chenopodium quinoa) — were selected for GABA enhancement via fermentation with L. brevis. These seeds are attractive due to their high nutritional value and sustainable production conditions, including high protein content and diverse amino acid profile (19, 20). For these seeds, very little literature is found; for example, quinoa was fermented with L. plantarum and subjected to an additional cold stress treatment, which improved GABA production (21). In comparison, cañihua and tarwi have only been reported to produce GABA by germination (22).
Solid-state fermentation (SSF) is considered a sustainable biotechnology that utilises low amounts of water to transform solid plant-based substrates into bioactive compounds (23). SSF is well-known in the food industry as a method for producing metabolites, enzymes, and organoleptic properties such as flavour and colour (24). SSF has also been studied for GABA production; for example, a recent study showed that SSF of hemp by Lactobacillus casei and Bacillus subtilis can produce a GABA-enhanced drink, yielding a product with 102.45 mg/100 mL of GABA, compared to 79.84 mg/100 mL of GABA produced by germination (25). Recently, GABA was produced in tarwi, cañihua, and quinoa Real seeds by enzymatic treatment with L. brevis glutamate decarboxylase (GAD), GadA, and GadB, where the highest yield was obtained in tarwi seeds by GadB; 2.46 mg/g of dry sample in 1 mL reactions (26).
In this context, the production of GABA by fermentation of tarwi, cañihua, and quinoa with L. brevis is reported for the first time. Alongside GABA, short-chain fatty acids (SCFAs) were also quantified as an indication of fermentation extent. Additionally, the quantification of soluble free amino acids was performed both before and after fermentation, considering the autoclavation step.
2 Materials and methods
2.1 Chemicals
De Man, Rogosa and Sharpe (MRS) Broth media, MRS agar, NaClO, L-glutamic acid, γ-aminobutyric acid (GABA), dansyl chloride, acetic acid, sulphuric acid, sodium phosphate dibasic heptahydrate, sodium phosphate monobasic monohydrate, ammonium acetate (LC–MS and HPLC grade), acetonitrile (LC–MS and HPLC grade), short-chain fatty acids standards; acetic acid, formic acid, propionic acid and butyric acid, lactic acid, amino acid standard AAS18 (Supelco). All reagents were purchased from Sigma Aldrich at analytical grade.
2.2 Bacterial strain
Levilactobacillus brevis DSM 1269 was purchased from Leibniz Institute DSMZ—German Collection of Microorganisms and Cell Cultures GmbH.
2.3 Bacterial strain stock preparation
The MRS broth was prepared in 51 g/L distilled water according to the manufacturer (Sigma-Aldrich). Subsequently, 25 mL of the medium was dispensed into a 40 mL Wheaton serum bottle. The medium was then deprived of oxygen by bubbling it with N2 for 5 min, and the bottle was closed with rubber stoppers. The bottle was sterilised in an autoclave.
Under sterile conditions, 1 mL of the medium was transferred from the Wheaton serum bottle to a 1.5 mL Eppendorf tube. A freeze-dried bacterium strain was resuspended in the medium and then transferred to the Wheaton serum bottle using another syringe. Following the recommendations for handling freeze-dried bacterial strains from Leibniz Institute DSMZ—German Collection of Microorganisms and Cell Cultures GmbH.
The bacterium was incubated at 30 °C for 24 h. Then, using a sterile syringe on the sterile bench, 100 μL of a bacterial sample was taken for cultivation on an agar plate. The plate was incubated at 30 °C for 24 h in an anaerobic chamber. Several colonies were observed on the plate and kept at 4 °C.
2.4 Seeds sampling
Cañihua (Chenopodium pallidicaule) seeds were collected from Chojñacota, Gualberto Villarroel Province (coordinates: 17°29′41.469″S 68°03′53.368″W), with an altitude of 3,770 (m.a.s.l.). Tarwi (Lupinus mutabilis) seeds were collected from Carabuco—Puerto Mayor de Carabuco, Camacho Province (coordinates: 15°44′00″S 69°01′00″W), with an altitude of 3,904 (m.a.s.l.). Both collection sites are within the Department of La Paz, Bolivia. Finally, a sample of commercial quinoa available in Swedish supermarkets was added to the study.
2.5 Seeds disinfection
Before disinfection, the moisture of the seeds was determined in triplicate, using a moisture analyser (Radwag MAC 110/WH, Poland).
For disinfection, 20 g of each seed was washed with 60 mL of 1% NaClO (v/v) for 30 min. It was washed three times with ultra-pure water (Milli-Q) and left overnight in 200 mL of Milli-Q water. The next day, the seeds were rewashed twice with Milli-Q water.
The seeds were then divided into two groups: one kept at room temperature (approximately 20 °C) for 24 h and the second at 4 °C for 24 h. Then, both groups were freeze-dried and ground into flour. The samples were named cañihua treated at 4 °C (C-4), cañihua treated at room temperature (C-RT), and the same for tarwi and quinoa: T-4, T-RT, Q-4, and Q-RT, respectively. These prior treatments of the seeds were tested because a previous work determined that a cold stress treatment on the seeds enhances GABA production through fermentation of quinoa seeds (21).
2.6 Fermentation
Each fermentation was conducted in duplicate due to the limited availability of raw samples and the high analytical demand of LC–MS/HPLC quantification. However, we highlight that all analytical measurements were performed in duplicate for each replicate, and the results showed high consistency between duplicates, indicating good reproducibility.
The fermentation setting consisted of using 2 g of each previously freeze-dried sample in Wheaton serum bottles with 25 mL of Milli-Q water. Then, the bottles were deprived of oxygen by bubbling them with N2 for 5 min and then closed with rubber stoppers. The bottles were sterilised in an autoclave.
Then, 200 μL of the bacterial stock (L. brevis, 25 CFU/μL) was introduced to the sterile bottles. The fermentation took place for 48 h at 30 °C. These parameters were determined as optimal for GABA production by L. brevis fermentation in the literature (21, 27).
To determine the impact of autoclaving on the conversion of L-glutamic acid into GABA, controls were performed using the same amounts, both with and without autoclaving. Then, both control groups were left for 48 h at 30 °C.
Given that solid-state fermentation was performed, the OD does not directly demonstrate the bacterial growth. It can demonstrate the presence of particles in solution (metabolites), which are products of fermentation. Therefore, the fermentation was determined by quantifying SCFAs, lactic acid, and GABA, which were measured in the aqueous fraction.
2.7 Derivatisation of amino acids and GABA for HPLC analysis
To quantify free amino acids and GABA by HPLC, performing derivatization before analysis is essential. The method was used previously in the group (26). In Eppendorf tubes, an aliquot of 150 μL of each sample was taken. Then, 300 μL of 0.1 M sodium bicarbonate buffer (pH 9.8), 150 μL of 10 mg/mL dansyl chloride in acetonitrile, and 900 μL of water were added. The tubes were incubated for 40 min at 80 °C. Finally, 150 μL of 20 μL/mL acetic acid was added to stop the reaction, and the tubes were centrifuged for 5 min at 12,000 rpm.
The supernatants were filtered through 0.2 μm pore size polytetrafluoroethylene (PTFE) membrane filters and stored at −20 °C until analysis by LC–MS and HPLC.
2.8 Identification of amino acids and GABA by liquid chromatography-mass spectrometry (LC–MS)
A liquid chromatography-mass spectrometry system, Dionex Ultimate 3,000 UHPLC (Thermo Fisher Scientific, Rockford, Illinois, United States), LTQ-Velos pro mass spectrometer using ESI in positive mode, connected to a UV detector at a wavelength of 275 nm equipped with a Kinetex 2.6 μm EVO C18 100 Å column 100 × 2.1 mm (Phenomenex, Torrance, California, United States), was used to identify amino acids and GABA. Two mobile phases were used: 30 mM ammonium acetate (A) and acetonitrile (B). A gradient between the solvents was employed; the separation starts with 18% of B. After 13 min, it increases to 22%, then when the total time reaches 16.5 min, B increases to 16.5%, and at a total time of 20.5 min, B increases to 30%. At a total time of 24 min, B decreases to 18%, and finally, at a total time of 27 min, B decreases to 10%. The method was based on a previous work (26) with modifications. Ten microliters of the sample were injected, the flow was maintained at 0.3 mL/min during the analysis, and the column temperature was set at 30 °C.
2.9 Quantification of amino acids and GABA with HPLC
After the identification of all amino acids and GABA by LC–MS, their quantification was performed using the same method described in section 2.8 in a high-performance liquid chromatography (HPLC) Vanquish (Thermo Fisher Scientific, Waltham, Massachusetts, United States) system connected to a UV detector at a wavelength of 275 nm equipped with the Kinetex 2.6 μm EVO C18 100 Å column 100 × 2.1 mm (Phenomenex, Torrance, California, United States).
2.10 Statistical analysis of amino acid profile and GABA production
Principal component analysis (PCA), heat maps, and dendrograms were performed on the soluble amino acid profiles of samples, including control groups and fermented samples, using Python (version 3.11) within a Jupyter Notebook environment. Cohen’s d test was applied to GABA production in fermented samples. Visualizations of the PCA, heat maps, and dendrograms were created using matplotlib (version 3.8.0) and seaborn (version 0.13.0).
2.11 Short-chain fatty acids (SCFAs) analysis
A high-performance liquid chromatography (HPLC) Vanquish (Thermo Fisher Scientific, Waltham, Massachusetts, United States) system connected to an RI detector, equipped with an Aminex HPX-87H column 300 mm × 7.8 mm (Bio-Rad), was used to quantify short-chain fatty acids (SCFAs), lactic acid, and ethanol. One millilitre of each sample was mixed with 20 μL of 20% (v/v) sulphuric acid and incubated for 15 min at 4 °C. The samples were filtered through 0.2 μm pore size PTFE membrane filters and stored at 4 °C. Lactic acid and SCFAs were separated using 5 mM sulphuric acid at a flow rate of 0.5 mL/min, while the column compartment was maintained at 40 °C (28). Standards of lactic acid, acetic acid, propionic acid, and butyric acid in concentrations of 0.1, 1, 2, 5, and 10 mg/mL were prepared in Milli-Q water and analysed under the same conditions as the samples.
3 Results
The initial moisture content of the samples was: tarwi, 5.8% ± 0.08; cañihua, 7.4% ± 0.3; and quinoa, 9.28% ± 0.61.
3.1 Identification of amino acids and GABA by LC–MS
The identification of amino acids and GABA was based on their derivatization with dansyl chloride, and the molecular weight of the derivatised amino acids was determined using LC–MS. Table 1 shows the molecular weights identified.
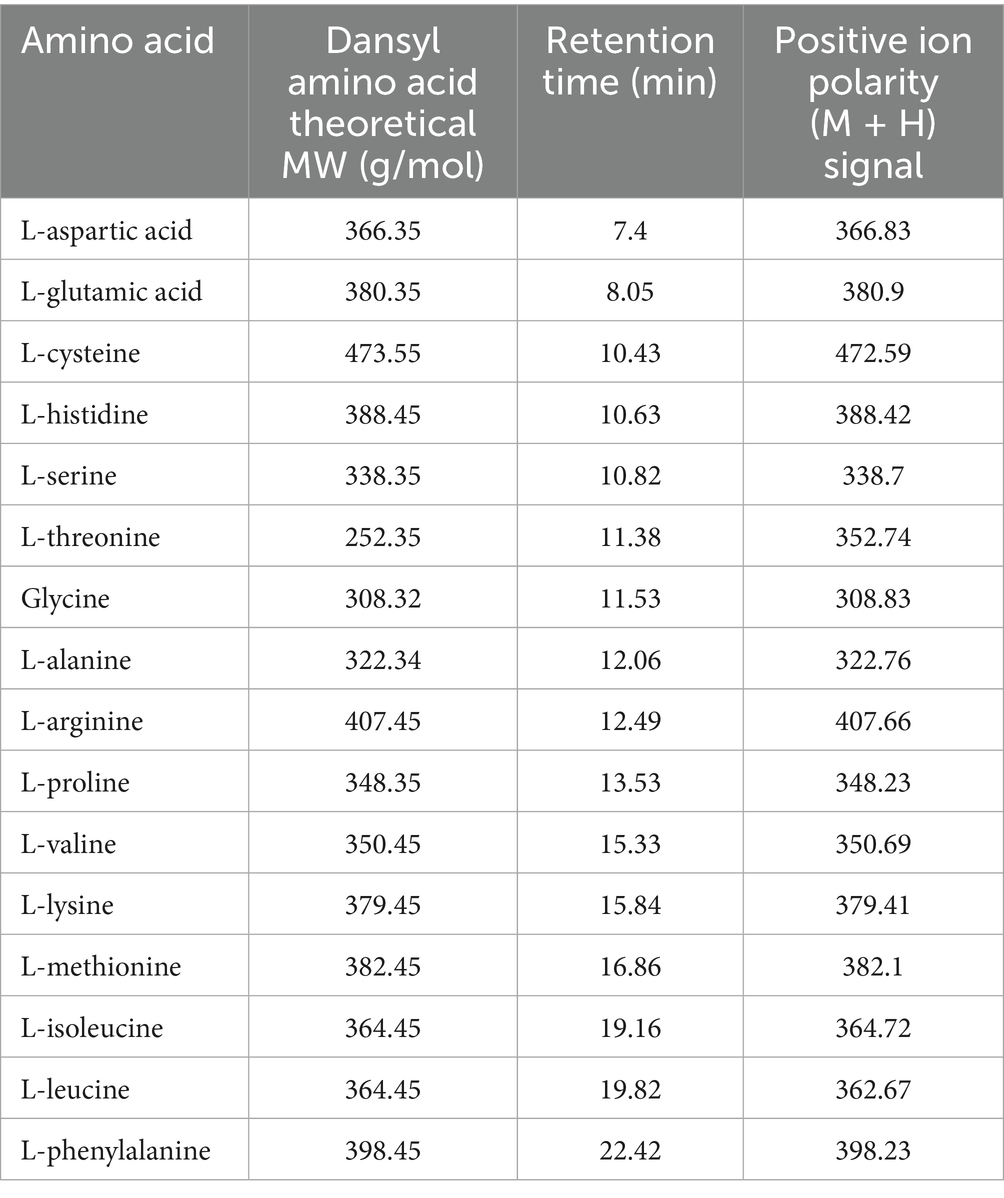
Table 1. Dasyl-derivatised amino acids detected by LC–MS.
3.2 Pretreatment influence on free glutamic acid concentration
Tarwi samples behave differently from cañihua and quinoa when autoclaved (Figure 1). In tarwi seeds, a higher concentration of glutamic acid is observed when the samples are not autoclaved in both RT and 4 °C pretreatments: 2.71 mg/g and 3.69 mg/g, respectively, compared to the autoclaved control samples: 0.9 mg/g at RT and 2.07 mg/g at 4 °C. Indicating a loss of glutamic acid in the autoclaving process. However, cañihua and quinoa exhibit higher concentrations of free glutamic acid after autoclaving.
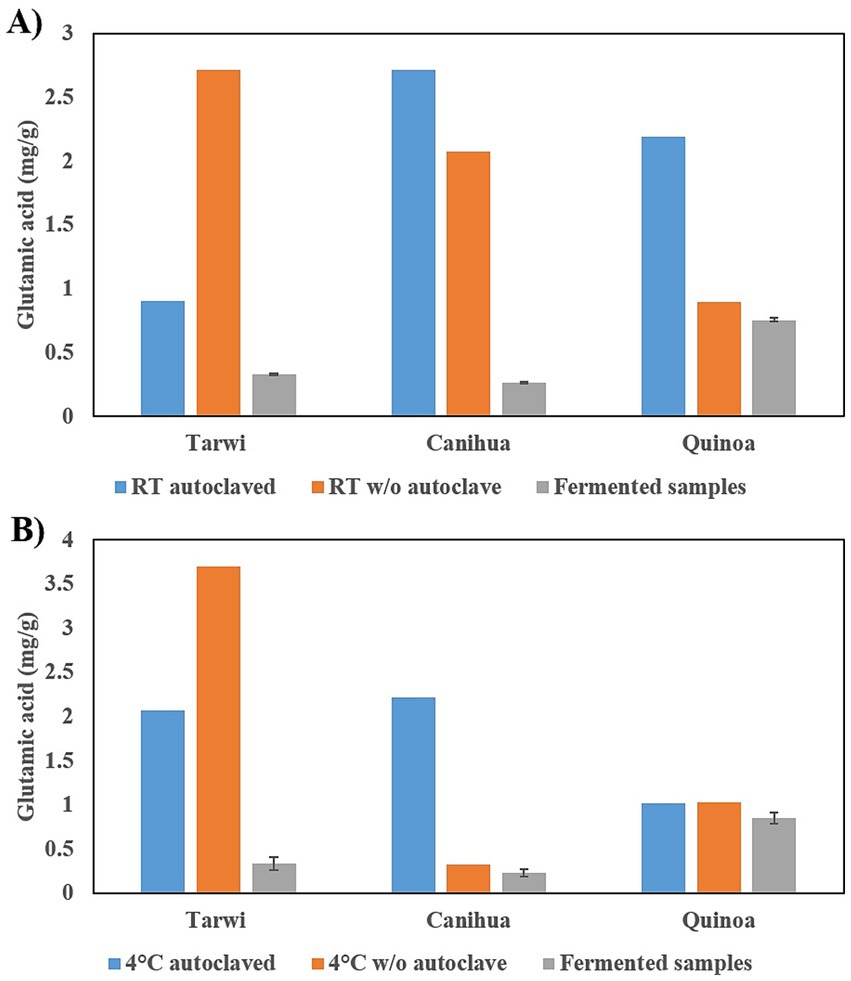
Figure 1. Effect of autoclaving on soluble free glutamic acid content. (A) Pretreatment at room temperature, concentration of glutamic acid in mg/g in controls with and without autoclave, and fermented samples. (B) Pretreatment at 4 °C, glutamic acid content in controls with and without autoclave and fermented samples. Results from one replicate of controls and duplicates of fermentation, expressed as average ± standard deviation.
Comparing the concentration of free glutamic acid with both pretreatments, for tarwi, the pretreatment at 4 °C produces a higher concentration of free glutamic acid, 3.69 mg/g. In contrast, cañihua and quinoa have higher free glutamic acid concentrations when pretreated at RT (around 20 °C) and autoclaved: 0.26 mg/g and 0.75 mg/g, respectively.
Finally, after fermentation, the concentration of free glutamic acid decreases in all cases, indicating that glutamic acid could be transformed into GABA; fermented tarwi produced 4.03 mg/g at RT and 3.64 mg/g at 4 °C, fermented cañihua produced 0.97 mg/g at RT and 0.89 mg/g at 4 °C. Finally, quinoa produced 0.3 mg/g of GABA with RT pretreatment, but no GABA was produced in fermented samples pretreated at 4 °C (Figure 2).
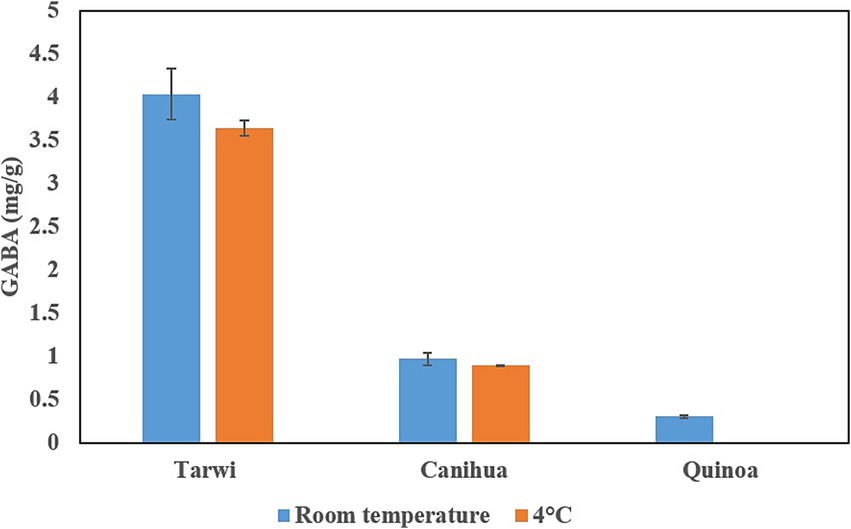
Figure 2. Production of GABA in three Andean seeds by fermentation with L. brevis. Two treatments, RT and 4 °C, were compared before the fermentation. Results from duplicates of fermentation.
3.3 Production of GABA by fermentation with L. brevis
GABA was produced in all fermentation products except for quinoa samples pretreated at 4 °C. Cohen’s d test shows that GABA production in tarwi and cañihua with room temperature prior treatment has a large effect size (d > 0.8), compared to the 4 °C previous treatment of the seeds (Figure 2).
3.4 Change in amino acid profile after fermentation with L. brevis
The amino acid profile was determined in all samples, with both prior treatments (RT or at 4 °C).
Fermentation had different effects on the amino acid composition of every seed. For example, in fermented tarwi, most free amino acids are present in similar amounts regardless of the prior thermal treatment; only a few show differences: cysteine, histidine, threonine, glycine, and proline were higher with prior RT treatment, and aspartic acid and valine were higher in seeds kept at 4 °C before fermentation. The control samples (autoclaved and non-autoclaved) also had the highest concentrations when kept at 4 °C compared to RT (Figure 3). It is also noted that autoclaving decreases the concentration of amino acids compared to non-autoclaved samples. While in cañihua after fermentation, when the seeds were kept at 4 °C previously, aspartic acid, cysteine, and isoleucine were found in higher amounts, while for histidine, glycine, valine, phenylalanine, and leucine, with a previous treatment at RT, were found in higher amounts (Figure 4). For quinoa, alanine is completely consumed during fermentation. Seeds previously treated at 4 °C showed higher amounts of aspartic acid, glutamic acid, cysteine, valine, methionine, and isoleucine. Glycine and proline were higher in seeds kept at RT before fermentation (Figure 5).
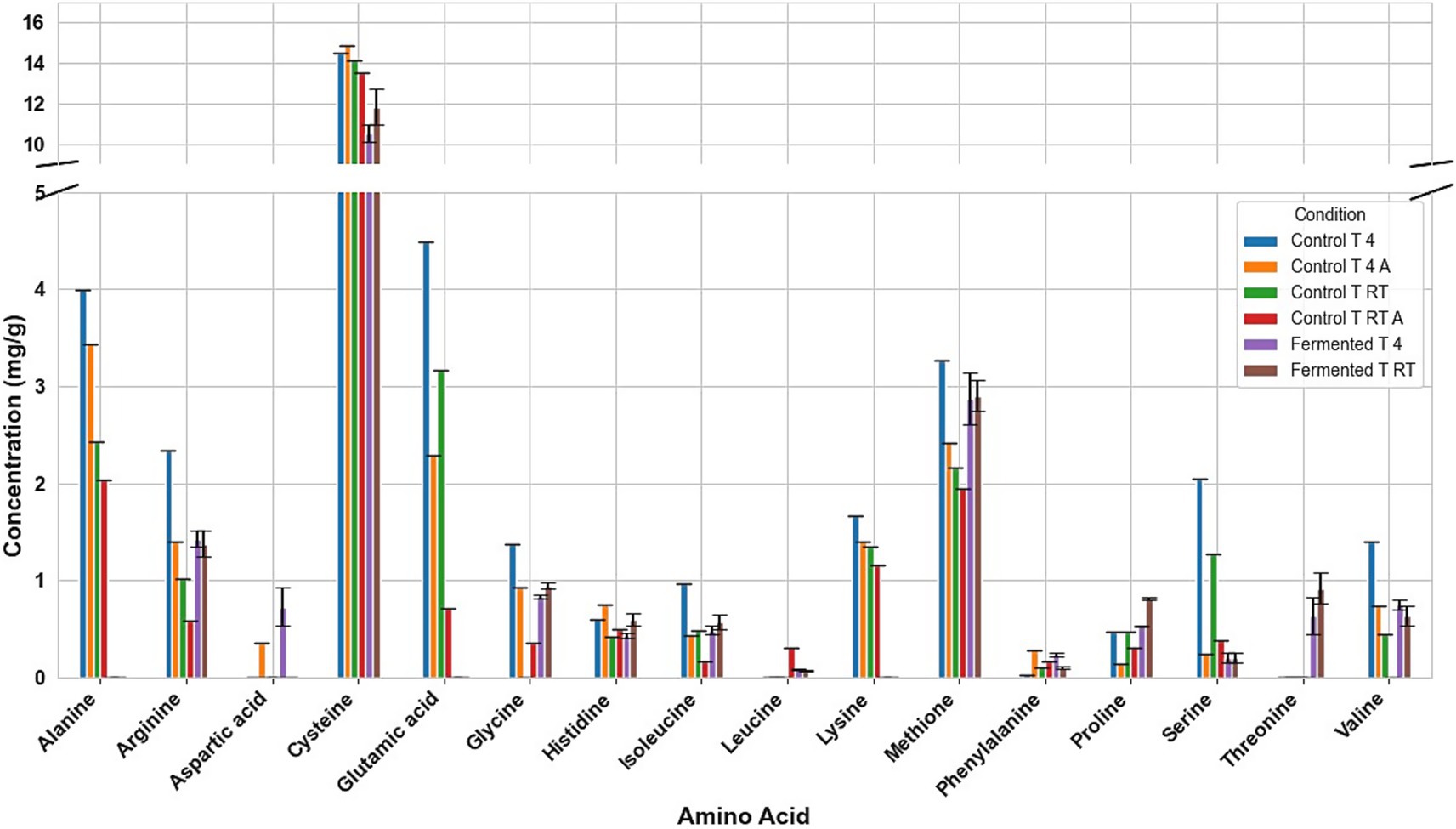
Figure 3. Amino acid profile of Tarwi samples. T RT A, room temperature pretreatment autoclaved; T RT, room temperature pretreatment w/o autoclaving; T 4 A, 4 °C pretreatment autoclaved; T 4, 4 °C pretreatment w/o autoclaving. Results from one replicate of controls and duplicates of fermentation samples, expressed as average ± standard deviation.
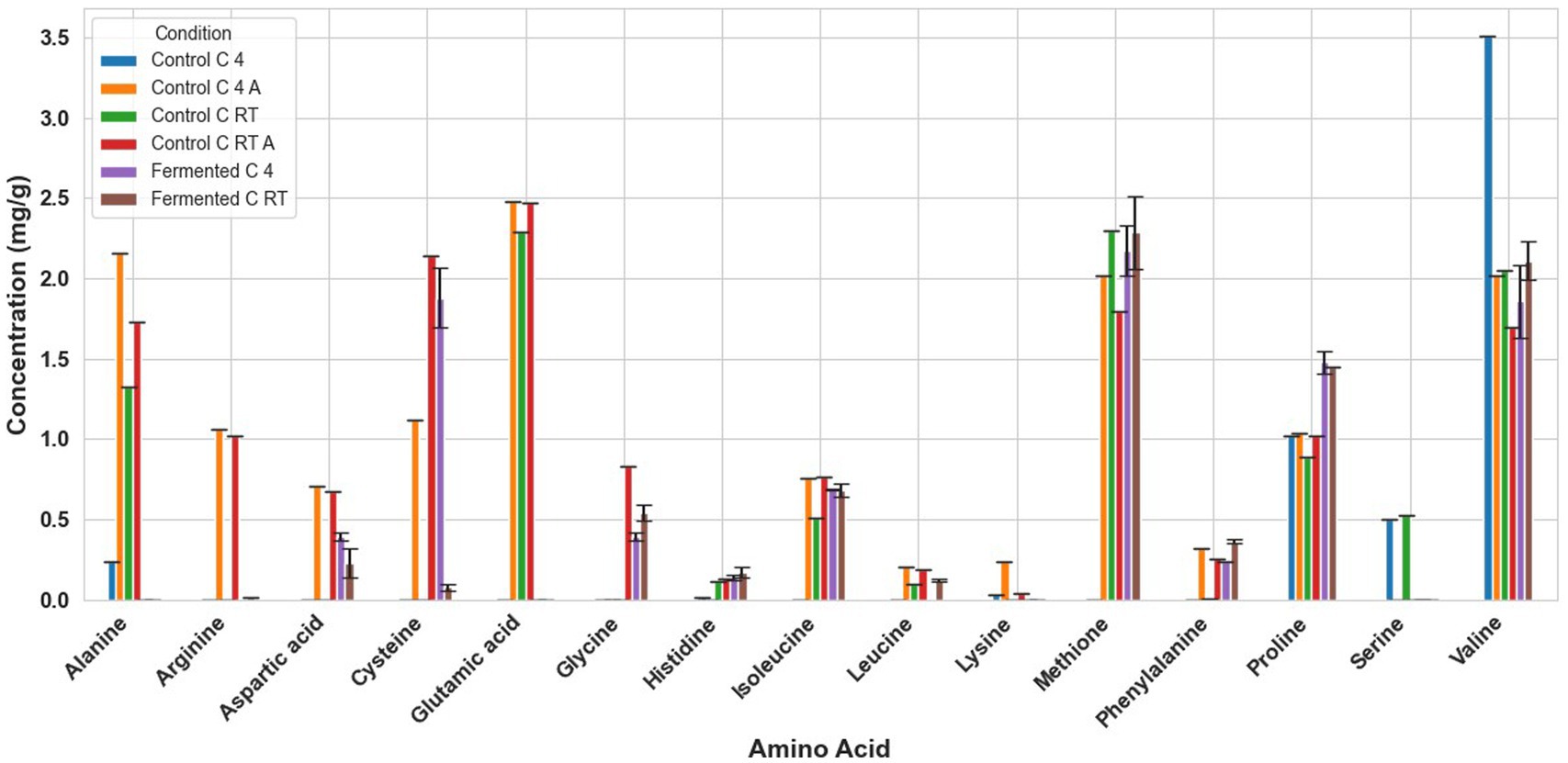
Figure 4. Amino acid profile of cañihua samples. C RT A, room temperature pretreatment autoclaved; C RT, room temperature pretreatment w/o autoclaving; C 4 A, 4 °C pretreatment autoclaved; C 4, 4 °C pretreatment w/o autoclaving. Results from one replicate of controls and duplicates of fermentation samples, expressed as average ± standard deviation.
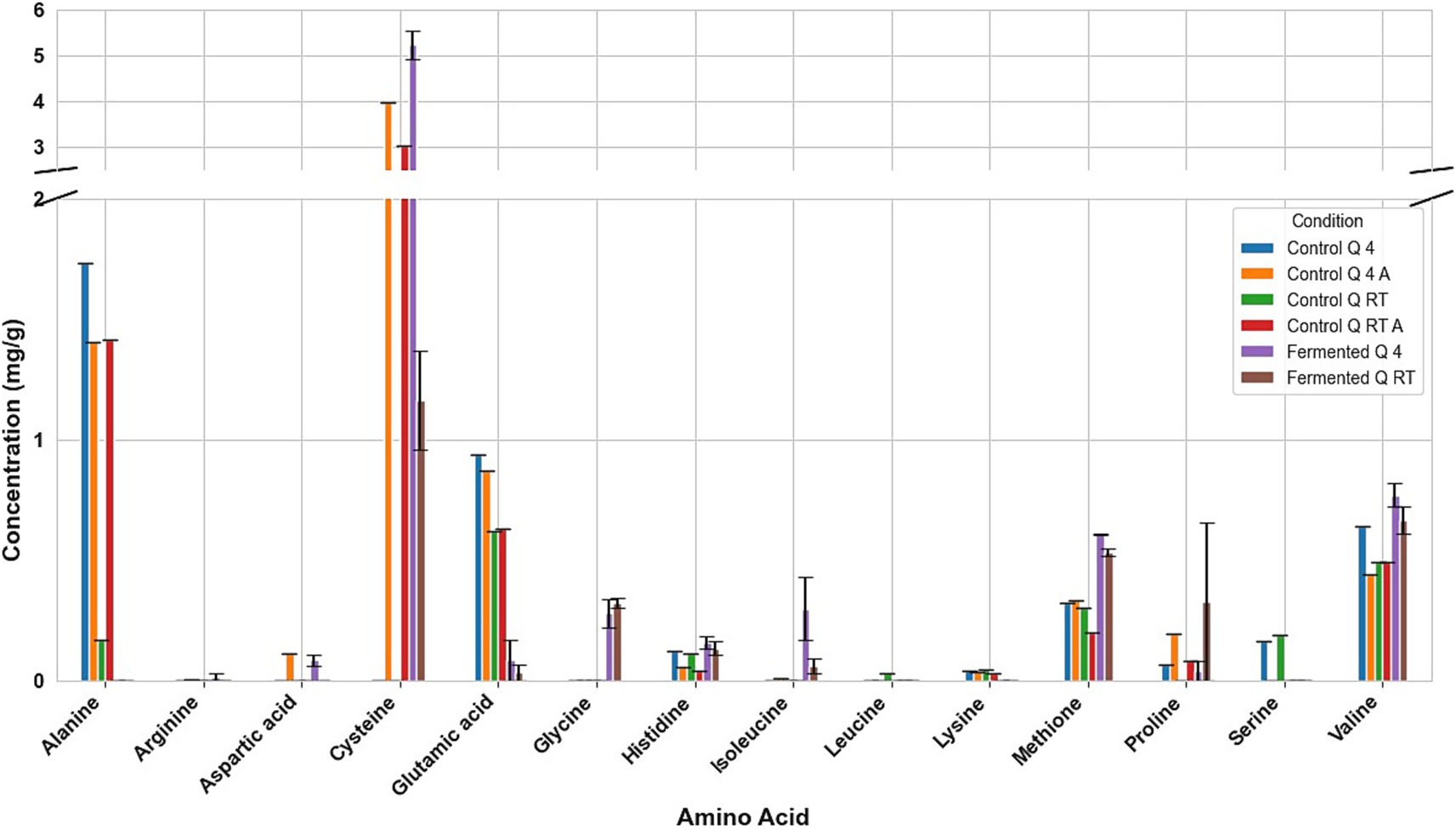
Figure 5. Amino acid profile of quinoa samples. Q RT A, room temperature pretreatment autoclaved; Q RT, room temperature pretreatment w/o autoclaving; Q 4 A, 4 °C pretreatment autoclaved; Q 4, 4 °C pretreatment w/o autoclaving. Results from one replicate of controls and duplicates of fermentation samples, expressed as average ± standard deviation.
The conversion of glutamic acid into GABA was clear. In tarwi, there is almost no glutamic acid after fermentation, and the highest amount of GABA produced by fermentation was 4 mg/g. The same behaviour was observed in cañihua, where almost all glutamic acid was consumed during fermentation, and GABA is produced. On the other hand, a considerable amount of glutamic acid remained in quinoa samples after fermentation, and GABA production was very low. However, no GABA is present in the samples fermented (T 4).
Another interesting result is the influence of autoclaving on the amino acid composition of the seeds. In tarwi seeds, autoclaving resulted in a decrease in the concentration of most amino acids. In cañihua seeds, the opposite effect was observed; most of the amino acids increased in concentration after autoclaving. Finally, in quinoa seeds, there was no clear tendency, unlike in the other seeds; almost half of the amino acids decreased in concentration, while the other half increased in concentration.
PCA was used to explore the amino acid profiles with prior thermal treatments and fermentation (Figure 6). For tarwi (Figure 6A), PC1 was affected mainly by glutamic acid, serine, alanine, arginine, valine, isoleucine, leucine, and phenylalanine. PC2 captured additional variation, with methionine, threonine, proline, and glycine showing a strong influence, while cysteine and lysine showed a lesser extent. The heat map (Figure 7A) supports that the main increase with prior RT treatment is in threonine and proline, as well as aspartic acid, when the seeds are treated at 4 °C. Fermented samples (T RT and T 4) formed distinct clusters separate from control groups; this was also confirmed in the dendrogram (Figure 8A). The dendrogram also shows that autoclaving affected the controls when a prior 4 °C treatment was applied.
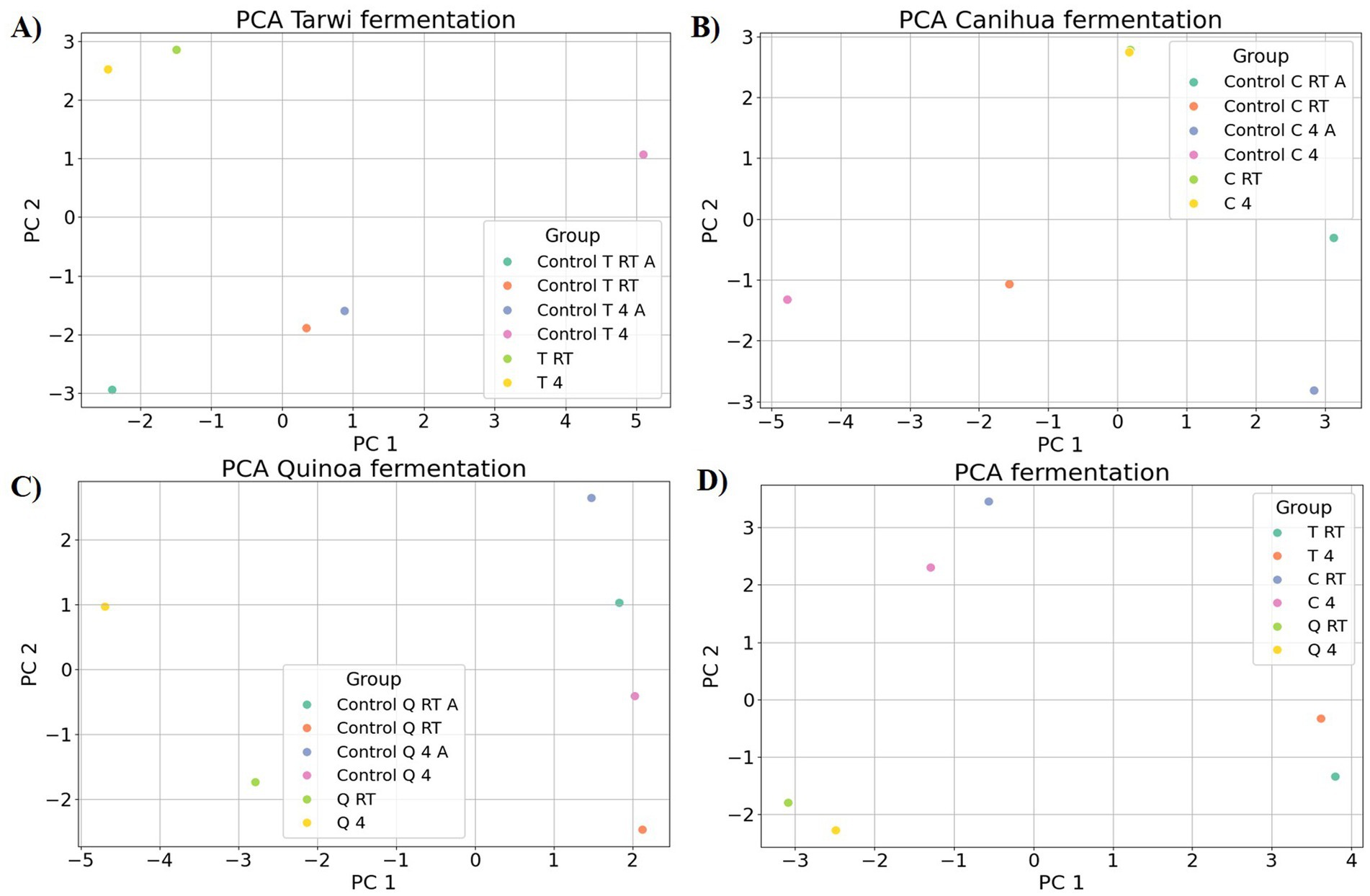
Figure 6. Principal component analysis (PCA) of amino acid profiles across fermentation treatments. (A) tarwi, (B) cañihua, (C) quinoa, and (D) all fermented samples. All plots were made in Python (v3.11) using matplotlib (v3.8.0) and seaborn (v0.13.0).
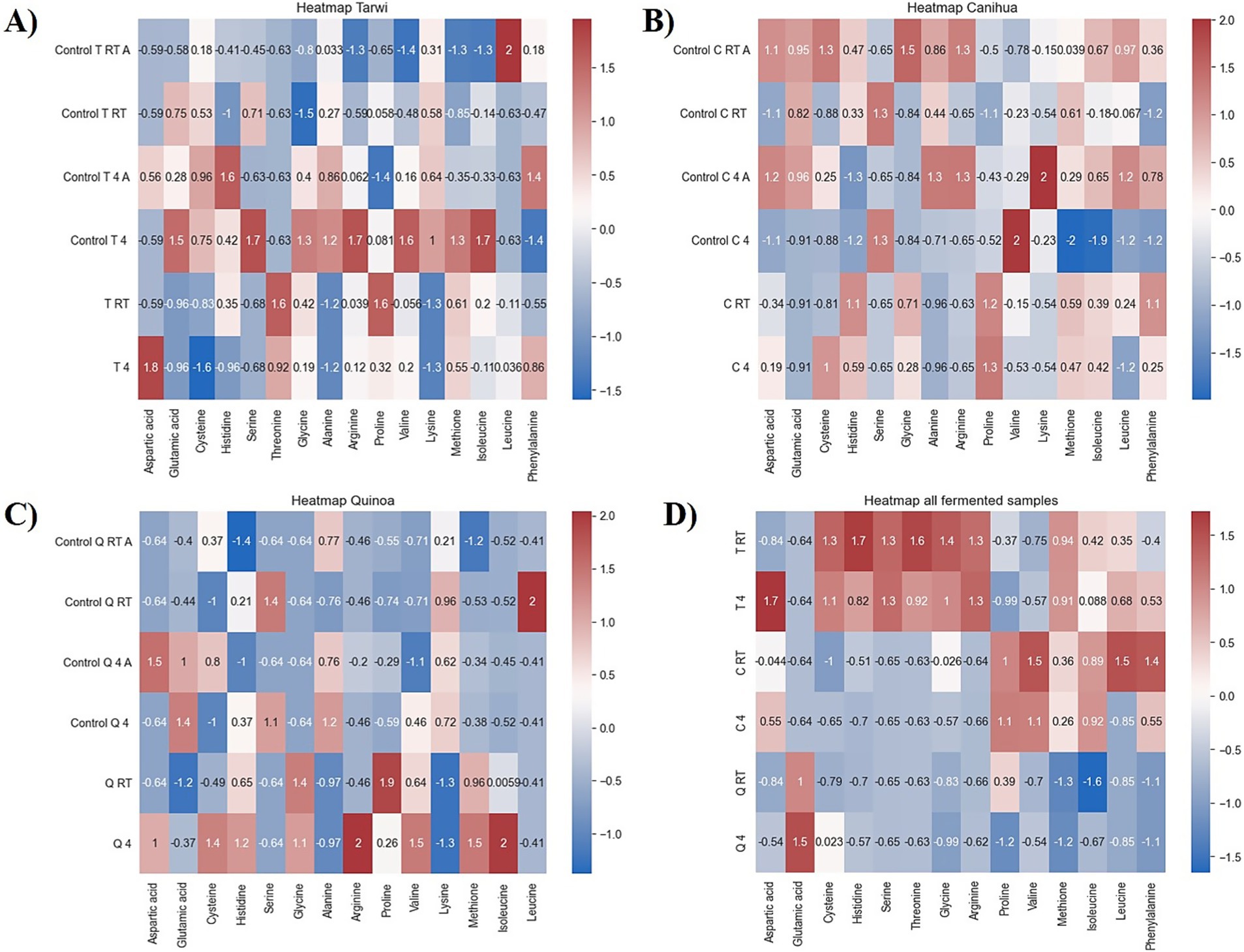
Figure 7. Heat maps of controls. RT, room temperature prior treatment; 4, 4 °C; A, autoclaved control; (A) Tarwi, (B) Cañihua, (C) quinoa, and (D) all fermented samples. All plots were made in Python (v3.11) using matplotlib (v3.8.0) and seaborn (v0.13.0).
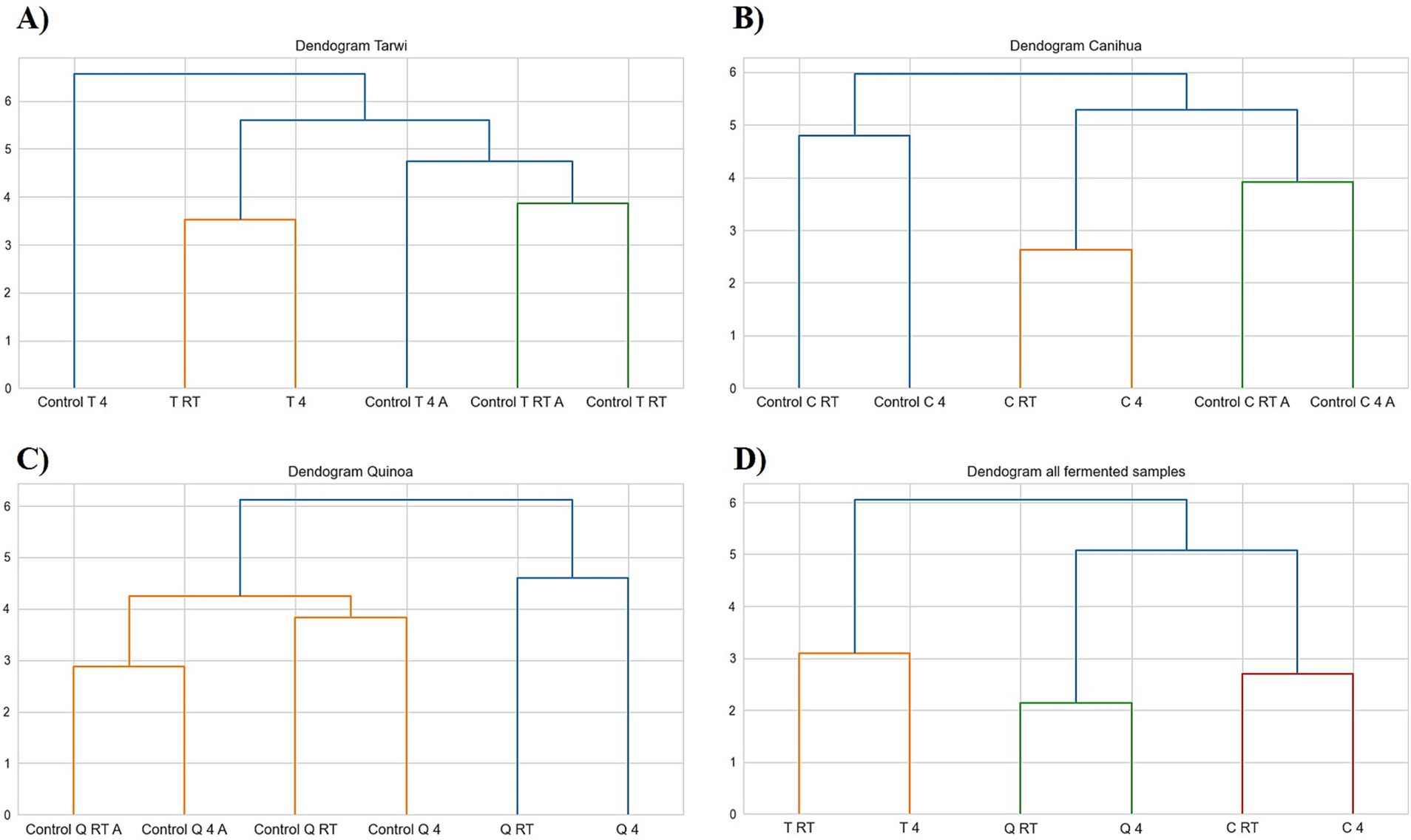
Figure 8. Dendrograms of controls; RT, room temperature prior treatment; 4, 4 °C; A, autoclaved control; (A) Tarwi, (B) Cañihua, (C) Quinoa, and (D) all fermented samples. All plots were made in Python (v3.11) using matplotlib (v3.8.0) and seaborn (v0.13.0).
For cañihua, PC1 was primarily influenced by hydrophobic and branched-chain amino acids, including isoleucine, aspartic acid, leucine, valine, and serine. This axis separates samples based on the polarity and hydrophobicity of their amino acids. In the PCA plot (Figure 6B), this separation is evident along PC1, where both controls (RT prior thermal treatment) cluster distinctly from fermented samples, suggesting that fermentation alters the abundance of these amino acids. PC2 was highly influenced by proline, histidine, alanine, and lysine. These amino acids may reflect structural or charge-related shifts during fermentation. PC2 distinguishes both controls (kept at 4 °C) from the other groups, indicating that it may enhance amino acids associated with protein rigidity and aromaticity. These observations are confirmed with the dendrogram (Figure 8B), where autoclaved controls cluster differently from the non-autoclaved ones (independently of the prior treatment). The heat map (Figure 7B) supports that the main increase resulting from fermentation is in proline.
PCA analysis of quinoa fermentation reveals that PC1 is influenced by lysine, alanine, serine, glycine, methionine, and isoleucine. In the PCA plot (Figure 6C), both control groups (RT) cluster at the positive end of PC1, suggesting higher levels of lysine, alanine, and serine. In contrast, both fermented samples shift toward the negative end, indicating fermentation-induced changes in the hydrophobic amino acid content. PC2 is dominated by cysteine, aspartic acid, leucine, and serine. These patterns suggest that PC2 captures variation in sulphur-containing and acidic amino acids. Then, the heat map (Figure 7C) confirms that arginine and isoleucine are the amino acids that increase the most with fermentation after a 4 °C treatment, and proline when the seeds have undergone prior RT treatment. The dendrogram (Figure 8C) suggests that the fermented samples cluster differently from all controls, but the controls are slightly affected by autoclavation, as they form a separate cluster that is still very similar to the non-autoclaved samples.
Finally, the PCA of tarwi, cañihua, and quinoa fermented samples shows that PC1 was primarily driven by glycine, serine, arginine, and threonine, with a high reduction of glutamic acid, reinforcing its conversion into GABA. In the PCA plot (Figure 6D, Table 2), fermented samples kept at RT before fermentation cluster toward the positive end of PC1, showing higher glycine, serine, arginine, and threonine. While fermented samples kept at 4 °C before fermentation shifted leftward, suggesting a reduction in these components over time. PC2 was shaped by valine, phenylalanine, and proline, indicating that this axis captures variation in hydrophobic and aromatic amino acids. Glutamic acid again contributed highly, reinforcing its inverse relationship with fermentation-driven changes. Similarly, PCA (Figure 6D) and the dendrogram (Figure 8D) indicate that all fermented samples cluster differently, depending on the seeds.
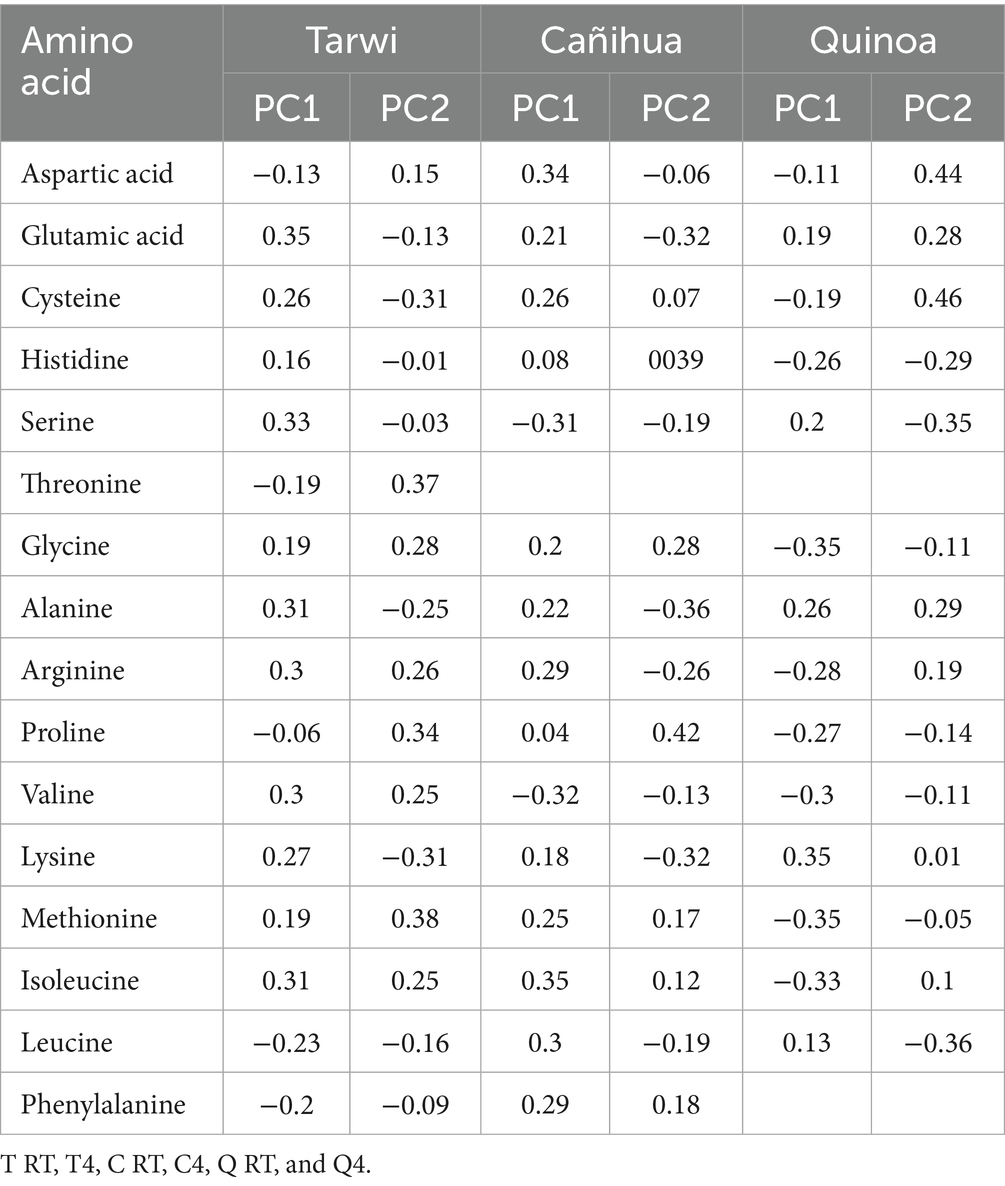
Table 2. Loadings for PCA analysis for each fermented sample
3.5 Production of short-chain fatty acids (SCFAs)
In all three seeds, lactic acid and acetic acid were produced. The product of fermentation from cañihua seeds shows the highest levels of those acids (Figure 9).
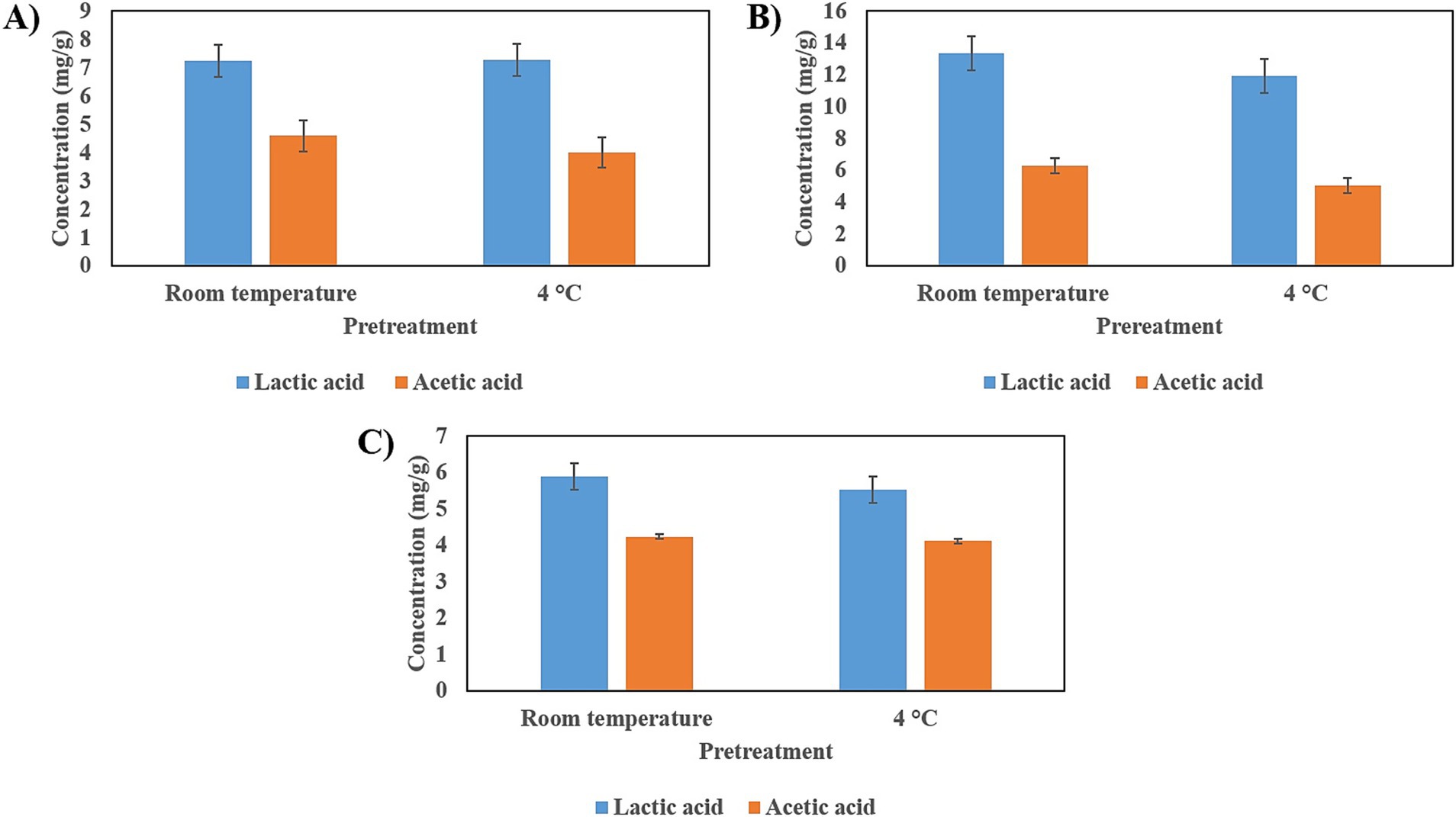
Figure 9. Production of short-chain fatty acids (SCFAs) by fermentation of seeds with L. brevis. (A) SCFAs produced by fermentation on tarwi seeds. (B) SCFAs produced by fermentation on cañihua seeds. (C) SCFAs produced by fermentation on quinoa seeds. Results expressed as average ± standard deviation.
4 Discussion
The production of fermented foods and beverages using LAB has become a significant area of interest for both research and industry, as the metabolites produced can enhance food quality and offer health benefits. One of these metabolites is γ-aminobutyric acid (GABA). Many LAB strains have been studied for their capacity to produce GABA, and several authors have concluded that L. brevis is the most efficient strain (29). GABA production can vary depending on conditions such as pH, temperature, and the composition of the medium (30).
Fermentation by L. brevis of various food matrices to produce GABA has been previously studied. For example, black soybean milk fermented by L. brevis FPA 3709 at 37 °C for 48 h achieved a final GABA concentration of 5.42 mg/mL (31). Black raspberry was also fermented by L. brevis GABA100 at 37 °C for 15 days, with a final GABA concentration of 2.42 mg/mL (32). Soybean and rice seem to be the most extensively studied food matrices for GABA enrichment (33). Other notable groups that appear to have potential for GABA enrichment include legumes and cereals. Heating and cooling have been tested as pretreatments to see their effect on GABA production by fermentation. Interestingly, on the one hand, a heat drying treatment at 40 °C resulted in increased GABA production in immature soybean seeds, reaching a level of up to 4.48 mg/g (34).
On the other hand, cold stress, if applied to the seeds before fermentation, is efficient in enhancing GABA production in quinoa seeds (21), these authors conclude that the higher GABA produced may be attributed to an adaptive response of the cells to cold, which leads to acidification of the cell membrane, due to the depletion of H+ caused by decarboxylation during the synthesis of GABA. However, our results show that GABA production in the three seeds, tarwi, cañihua, and quinoa, with a room temperature treatment of the seeds before fermentation, is higher than that of the prior treatment at 4 °C. However, no heating pretreatments were studied. Recently, in our research group, GABA was produced enzymatically from Andean seeds (26). The production of GABA in tarwi was higher through fermentation compared to the small-scale enzymatic reaction, but lower compared to the large-scale enzymatic reactions (Table 3). Fermented cañihua produced more GABA by enzymatic treatment than fermentation. However, keeping the seeds at 4 °C did show a positive effect, as evidenced by the increased concentration of certain amino acids after fermenting quinoa and canihua seeds. The most significant difference is observed in cysteine concentration after fermentation of both seeds with the pretreatment at 4 °C compared to RT pretreatment. In fermented tarwi seeds, the same effect is observed in almost all amino acids, but not for threonine, proline, and isoleucine.

Table 3. Comparison of enzymatic and fermentation production of GABA in Andean seeds with L. brevis.
Short-chain fatty acids (SCFAs) and lactic acid are also important products of LAB fermentation. In this study, only lactic and acetic acid were produced through fermentation by L. brevis, with higher concentrations of lactic acid than acetic acid being observed. Other studies have identified a group of sugars that, when present in the culture medium, consistently produce more lactic than acetic acid, regardless of the L. brevis strain (35). These sugars are D-glucose, D-galactose, D-mannose, and D-mannitol, suggesting that some of these sugars may be present in the seeds used in this study, which could lead to a higher production of lactic acid than acetic acid in all cases. Acetic acid has been found to enhance the production of anti-inflammatory mediators, including IL-10, transforming growth factor-β (TGF-β), and annexin A1 (36). Lactic acid has demonstrated various bioactive capacities, including its critical role in modulating inflammation in a model of small intestine injury caused by indomethacin (37). It has also been shown to contribute to maintaining intestinal barrier function (38).
Little is known about the effect of autoclaving on the free amino acid profile before fermentation, especially in Andean seeds. Early studies reported a 10% decrease in lysine after autoclaving chickpea and pulses but no change in other amino acids (39). Another study also found a decline in lysine when autoclaving blends of soybean, white beans, sesame, and peanut (40). However, recent work found no change in the amino acid profile when cheese whey permeates are autoclaved (41). In the present work, autoclaving affected the amino acid profiles of tarwi, cañihua, and quinoa in distinct ways.
Some free amino acids have increased in the soluble phase after the fermentation of the Andean seeds. Tarwi’s fermentation product shows that threonine was produced. This essential amino acid is a crucial component of gastrointestinal mucin and serves as a nutritional modulator that influences the intestinal immune system through complex signalling networks (42). Other essential amino acids increased in concentration after the fermentation of tarwi by L. brevis, including histidine, isoleucine, leucine, methionine, and valine. In fermented cañihua, glycine is not observed in the controls of soluble amino acids but is present after fermentation. This non-essential amino acid has also been suggested as a conditionally essential amino acid, as it has been observed that glycine supplementation is needed in metabolic disorders such as obesity and type 2 diabetes (43). Glycine is not only a building block of proteins, but it is also required for many metabolic processes, and it has a broad spectrum of anti-inflammatory, cytoprotective, and immunomodulatory properties (44). Other essential amino acids that increased in concentration after the fermentation of cañihua by L. brevis are histidine, phenylalanine, isoleucine, and methionine. In fermented quinoa, glycine and isoleucine are produced during the fermentation. Isoleucine is an essential amino acid and together with leucine and valine it comprises 17% of the human skeletal muscle and is used to support protein synthesis (45). Methionine, histidine, and valine are other essential amino acids that are increased in concentration by quinoa fermentation.
Besides the beneficial effects of free amino acids, they can also influence the taste of fermented food. They have been classified into four groups: amino acids that contribute umami taste (aspartic acid and glutamic acid), amino acids that contribute to sweetness (serine, alanine, glycine and threonine), amino acids that contribute to bitterness (arginine, histidine, tyrosine, leucine, valine, methionine, isoleucine, phenylalanine, lysine and proline), and finally cysteine is considered tasteless (46). In this sense, fermented tarwi exhibits an increase in the concentration of free amino acids that contribute to bitter taste, but only one that could contribute to its sweet taste. Fermented cañihua and quinoa have an increase in concentration of two bitter-contributor amino acids and one sweet-contributor. It can be concluded that the amino acid profile in fermented products may contribute to an increased bitterness.
In this exploratory study, a limitation is that fermentations were conducted in duplicate due to restricted seed availability and the high analytical demand of LC–MS/HPLC analyses. Nevertheless, all analytical measurements were performed in duplicate for each replicate, and results were consistent, indicating good reproducibility. Future work will include larger-scale fermentations with additional replicates to strengthen statistical robustness. In addition, the study of organoleptic properties of fermented Andean seeds aims to develop nutritional and functional fermented foods, extend their shelf life, and enhance their palatability.
5 Conclusion
This study explores the enhancement of the functional value of Andean seeds through fermentation with L. brevis DSM 1269. As an environmentally friendly technique, it enables the production of bioactive compounds such as lactic acid, acetic acid, and GABA, while also increasing the production of essential amino acids, including threonine, histidine, methionine, isoleucine, leucine, valine, and lysine. In this sense, these results could lead to scaling up the process and potentially applying it to formulate novel, fermented, plant-based foods with enhanced properties.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
GI: Writing – review & editing, Writing – original draft, Validation, Investigation, Formal analysis, Conceptualization. JO-S: Writing – review & editing, Investigation. CG: Writing review & editing, Investigation. JMP: Writing – review & editing, Resources, Project administration, Funding acquisition. JAL-P: Writing – review & editing, Supervision, Resources, Project administration, Conceptualization, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The Swedish International Development Cooperation Agency (SIDA), and Lund University provides open-access funding.
Acknowledgments
The authors thank the Swedish International Development Cooperation Agency (SIDA) for supporting the present study. The views expressed herein do not necessarily represent those of SIDA or its Board of Governors. Lund University provides open-access funding.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Gan, R-Y, Li, H-B, Gunaratne, A, Sui, Z-Q, and Corke, H. Effects of fermented edible seeds and their products on human health: bioactive components and bioactivities. Compr Rev Food Sci Food Saf. (2017) 16:489–531. doi: 10.1111/1541-4337.12257
2. Terefe, NS. 5 - recent developments in fermentation technology: toward the next revolution in food production In: P Juliano, R Buckow, MH Nguyen, K Knoerzer, and J Sellahewa, editors. Food engineering innovations across the food supply chain. Amsterdam, Netherlands: Elsevier (2022).
3. Abdul Hakim, BN, Xuan, NJ, and Oslan, SNH. A comprehensive review of bioactive compounds from lactic acid bacteria: potential functions as functional food in dietetics and the food industry. Foods. (2023) 12:2850. doi: 10.3390/foods12152850
4. Lu, Y, Tan, X, Lv, Y, Yang, G, Chi, Y, and He, Q. Physicochemical properties and microbial community dynamics during Chinese horse bean-chili-paste fermentation, revealed by culture-dependent and culture-independent approaches. Food Microbiol. (2020) 85:103309. doi: 10.1016/j.fm.2019.103309
5. Yu, J, Geng, Y, Xia, H, Ma, D, Liu, C, Wu, R, et al. Lab fermentation improves production of bioactive compounds and antioxidant activity of Withania somnifera extract and its metabolic signatures as revealed by LC-MS/MS. J Microbiol Biotechnol. (2022) 32:473–83. doi: 10.4014/jmb.2111.11018
6. Nkhata, SG, Ayua, E, Kamau, EH, and Shingiro, JB. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci Nutr. (2018) 6:2446–58. doi: 10.1002/fsn3.846
7. Pranoto, Y, Anggrahini, S, and Efendi, Z. Effect of natural and Lactobacillus plantarum fermentation on in-vitro protein and starch digestibilities of sorghum flour. Food Biosci. (2013) 2:46–52. doi: 10.1016/j.fbio.2013.04.001
8. Jin, Y, Wu, J, Hu, D, Li, J, Zhu, W, Yuan, L, et al. Gamma-aminobutyric acid-producing Levilactobacillus brevis strains as probiotics in litchi juice fermentation. Foods. (2023) 12:302. doi: 10.3390/foods12020302
9. Ngo, D-H, and Vo, TS. An updated review on pharmaceutical properties of gamma-aminobutyric acid. Molecules. (2019) 24:2678. doi: 10.3390/molecules24152678
10. Bruni, O, Ferini-Strambi, L, Giacomoni, E, and Pellegrino, P. Herbal remedies and their possible effect on the GABAergic system and sleep. Nutrients. (2021) 13:530. doi: 10.3390/nu13020530
11. Hepsomali, P, Groeger, JA, Nishihira, J, and Scholey, A. Effects of oral gamma-aminobutyric acid (GABA) administration on stress and sleep in humans: a systematic review. Front Neurosci. (2020) 14:923. doi: 10.3389/fnins.2020.00923
12. Jeong, A-H, Hwang, J, Jo, K, Kim, S, Ahn, Y, Suh, HJ, et al. Fermented gamma-aminobutyric acid improves sleep behaviors in fruit flies and rodent models. Int J Mol Sci. (2021) 22:3537. doi: 10.3390/ijms22073537
13. Wu, S-J, Chang, C-Y, Lai, Y-T, and Shyu, Y-T. Increasing γ-aminobutyric acid content in vegetable soybeans via high-pressure processing and efficacy of their antidepressant-like activity in mice. Foods. (2020) 9:1673. doi: 10.3390/foods9111673
14. Quillin, SJ, Tran, P, and Prindle, A. Potential roles for gamma-aminobutyric acid signaling in bacterial communities. Bioelectricity. (2021) 3:120–5. doi: 10.1089/bioe.2021.0012
15. Dhakal, R, Bajpai, VK, and Baek, KH. Production of gaba (γ-aminobutyric acid) by microorganisms: a review. Braz J Microbiol. (2012) 43:1230–41. doi: 10.1590/s1517-83822012000400001
16. Zhang, Y, Zhu, M, Lu, W, Zhang, C, Chen, D, Shah, NP, et al. Optimizing Levilactobacillus brevis NPS-QW 145 fermentation for gamma-aminobutyric acid (GABA) production in soybean sprout yogurt-like product. Foods. (2023) 12:977. doi: 10.3390/foods12050977
17. Cataldo, PG, Urquiza Martínez, MP, Villena, J, Kitazawa, H, Saavedra, L, and Hebert, EM. Comprehensive characterization of γ-aminobutyric acid (GABA) production by Levilactobacillus brevis CRL 2013: insights from physiology, genomics, and proteomics. Front Microbiol. (2024) 15:1408624. doi: 10.3389/fmicb.2024.1408624
18. Luo, X, Wang, Y, Li, Q, Wang, D, Xing, C, Zhang, L, et al. Accumulating mechanism of γ-aminobutyric acid in soybean (Glycine max L.) during germination. Int J Food Sci Technol. (2018) 53:106–11. doi: 10.1111/ijfs.13563
19. Gomez Cahuata, JF, Rosas-Quina, YE, and Pachari Vera, E. Cañihua (Chenopodium pallidicaule Aellen) a promising superfood in food industry: a review. Nutr Food Sci. (2022) 52:917–28. doi: 10.1108/NFS-09-2021-0277
20. Suomela, J-P, Repo-Carrasco-Valencia, R, and Lutz, M. Andean native grains, quinoa, and lupin as sources of bioactive components In: Native crops in Latin America. Boca Raton, Florida, USA: CRC Press (2022).
21. Zhang, Y, Zhang, M, Li, T, Zhang, X, and Wang, L. Enhance production of γ-aminobutyric acid (GABA) and improve the function of fermented quinoa by cold stress. Foods. (2022) 11:3908. doi: 10.3390/foods11233908
22. De-La-Cruz-Yoshiura, S, Vidaurre-Ruiz, J, Alcázar-Alay, S, Encina-Zelada, CR, Cabezas, DM, Correa, MJ, et al. Sprouted Andean grains: an alternative for the development of nutritious and functional products. Food Rev Int. (2023) 39:5583–611. doi: 10.1080/87559129.2022.2083158
23. Verduzco-Oliva, R, and Gutierrez-Uribe, JA. Beyond enzyme production: solid state fermentation (SSF) as an alternative approach to produce antioxidant polysaccharides. Sustainability. (2020) 12:495. doi: 10.3390/su12020495
24. Ghosh, JS. Solid state fermentation and food processing: a short review. J Nutr Food Sci. (2016) 6:1–7. doi: 10.4172/2155-9600.1000453
25. Karabulut, G, Nemzer, BV, and Feng, H. γ-Aminobutyric acid (GABA)-enriched hemp milk by solid-state co-fermentation and germination bioprocesses. Plant Foods Hum Nutr. (2024) 79:322–9. doi: 10.1007/s11130-024-01187-6
26. Ibieta, G, Ortiz-Sempértegui, J, Peñarrieta, JM, and Linares-Pastén, JA. Enhancing the functional value of Andean food plants: enzymatic production of γ-aminobutyric acid from tarwi, cañihua and quinoa real seeds’ proteins. LWT Food Sci Technol. (2025) 220:117564. doi: 10.1016/j.lwt.2025.117564
27. Villegas, JM, Brown, L, Savoy de Giori, G, and Hebert, EM. Optimization of batch culture conditions for GABA production by Lactobacillus brevis CRL 1942, isolated from quinoa sourdough. LWT Food Sci Technol. (2016) 67:22–6. doi: 10.1016/j.lwt.2015.11.027
28. Allahgholi, L, Jönsson, M, Christensen, MD, Jasilionis, A, Nouri, M, Lavasani, S, et al. Fermentation of the brown seaweed Alaria esculenta by a lactic acid bacteria consortium able to utilize mannitol and laminari-oligosaccharides. Fermentation. (2023) 9:499. doi: 10.3390/fermentation9060499
29. Wang, Q, Liu, X, Fu, J, Wang, S, Chen, Y, Chang, K, et al. Substrate sustained release-based high efficacy biosynthesis of GABA by Lactobacillus brevis NCL912. Microb Cell Factories. (2018) 17:1–8. doi: 10.1186/s12934-018-0919-6
30. Icer, MA, Sarikaya, B, Kocyigit, E, Atabilen, B, Çelik, MN, Capasso, R, et al. Contributions of gamma-aminobutyric acid (GABA) produced by lactic acid bacteria on food quality and human health: current applications and future prospects. Foods. (2024) 13:2437. doi: 10.3390/foods13152437
31. Ko, CY, Lin, H-TV, and Tsai, GJ. Gamma-aminobutyric acid production in black soybean milk by Lactobacillus brevis FPA 3709 and the antidepressant effect of the fermented product on a forced swimming rat model. Process Biochem. (2013) 48:559–68. doi: 10.1016/j.procbio.2013.02.021
32. Kim, JY, Lee, MY, Ji, GE, Lee, YS, and Hwang, KT. Production of γ-aminobutyric acid in black raspberry juice during fermentation by Lactobacillus brevis GABA100. Int J Food Microbiol. (2009) 130:12–6. doi: 10.1016/j.ijfoodmicro.2008.12.028
33. Lee, XY, Tan, JS, and Cheng, LH. Gamma-aminobutyric acid (GABA) enrichment in plant-based food – a mini review. Food Rev Int. (2023) 39:5864–85. doi: 10.1080/87559129.2022.2097257
34. Takahashi, Y, Sasanuma, T, and Abe, T. Accumulation of gamma-aminobutyrate (GABA) caused by heat-drying and expression of related genes in immature vegetable soybean (edamame). Breed Sci. (2013) 63:205–10. doi: 10.1270/jsbbs.63.205
35. Hwang, HJ, Lee, SY, Kim, SM, and Lee, SB. Fermentation of seaweed sugars by Lactobacillus species and the potential of seaweed as a biomass feedstock. Biotechnol Bioprocess Eng. (2011) 16:1231–9. doi: 10.1007/s12257-011-0278-1
36. Vieira, AT, Galvão, I, Macia, LM, Sernaglia, EM, Vinolo, MAR, Garcia, CC, et al. Dietary fiber and the short-chain fatty acid acetate promote resolution of neutrophilic inflammation in a model of gout in mice. J Leukoc Biol. (2017) 101:275–84. doi: 10.1189/jlb.3A1015-453RRR
37. Watanabe, T, Nishio, H, Tanigawa, T, Yamagami, H, Okazaki, H, Watanabe, K, et al. Probiotic Lactobacillus casei strain Shirota prevents indomethacin-induced small intestinal injury: involvement of lactic acid. American journal of physiology-gastrointestinal and liver. Physiology. (2009) 297:G506–13. doi: 10.1152/ajpgi.90553.2008
38. Garrote, GL, Abraham, AG, and Rumbo, M. Is lactate an undervalued functional component of fermented food products? Front Microbiol. (2015) 6:629. doi: 10.3389/fmicb.2015.00629
39. del Cueto, AG, Martinez, W, and Frampton, V. Heat effects on peas, effect of autoclaving on the basic amino acids and proteins of the chick pea. J Agric Food Chem. (1960) 8:331–2. doi: 10.1021/jf60110a022
40. Srihara, P, and Alexander, J. Protein quality of raw and autoclaved plant protein blends. Can Inst Food Sci Technol J. (1983) 16:63–7. doi: 10.1016/S0315-5463(83)72021-3
41. Donzella, S, Fumagalli, A, Arioli, S, Pellegrino, L, D’Incecco, P, Molinari, F, et al. Recycling food waste and saving water: optimization of the fermentation processes from cheese whey permeate to yeast oil. Fermentation. (2022) 8:341. doi: 10.3390/fermentation8070341
42. Tang, Q, Tan, P, Ma, N, and Ma, X. Physiological functions of threonine in animals: beyond nutrition metabolism. Nutrients. (2021) 13:2592. doi: 10.3390/nu13082592
43. Alves, A, Bassot, A, Bulteau, A-L, Pirola, L, and Morio, B. Glycine metabolism and its alterations in obesity and metabolic diseases. Nutrients. (2019) 11:1356. doi: 10.3390/nu11061356
44. Pérez-Torres, I, María Zuniga-Munoz, A, and Guarner-Lans, V. Beneficial effects of the amino acid glycine. Mini Rev Med Chem. (2017) 17:15–32. doi: 10.2174/1389557516666160609081602
45. Gorissen, SHM, and Phillips, SM. Chapter 17 - branched-chain amino acids (leucine, isoleucine, and valine) and skeletal muscle In: S Walrand, editor. Nutrition and skeletal muscle. San Diego, California: Academic Press (2019).
Keywords: solid state fermentation (SSF), Levilactobacillus brevis, γ-aminobutyric acid (GABA), amino acid profile, Andean seeds, short-chain fatty acids (SCFAs)
Citation: Ibieta G, Ortiz-Sempértegui J, Grey C, Peñarrieta JM and Linares-Pastén JA (2025) γ-Aminobutyric acid (GABA) production and soluble free amino acid profile change in Andean seeds by Levilactobacillus brevis fermentation. Front. Nutr. 12:1693053. doi: 10.3389/fnut.2025.1693053
Edited by:
Bin Wang, Shaoguan University, ChinaReviewed by:
Guillermo H. Peralta, CONICET Instituto de Lactología Industrial (INLAIN), ArgentinaBruntha Devi, Pondicherry University, India
Copyright © 2025 Ibieta, Ortiz-Sempértegui, Grey, Peñarrieta and Linares-Pastén. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gabriela Ibieta, Z2FicmllbGEuaWJpZXRhX2ppbWVuZXpAcGxlLmx0aC5zZQ==; Javier A. Linares-Pastén, amF2aWVyLmxpbmFyZXNfcGFzdGVuQHBsZS5sdGguc2U=