 Ping Zeng
Ping Zeng Yoichi Enatsu
Yoichi Enatsu Guanyu Zhou1
Guanyu Zhou1- 1Institute of Fundamental and Frontier Sciences, University of Electronic Science and Technology of China, Chengdu, China
- 2Institute of Arts and Sciences, Oshamambe Division, Tokyo University of Science, Oshamambe, Hokkaido, Japan
Competitive interactions between multiple cell populations are crucial for modeling biological processes such as tumor growth and tissue regeneration. In this study, we investigate the nonlocal advection model for two-species competition, which characterizes cell growth and dispersion phenomena in co-culture experiments. To capture realistic phenomena in biology, we introduce the time delay representing population migration and resources recovering time. The primary objectives are to investigate the impact of time delays on competitive dynamics under various parameter settings and to develop numerical methods that ensure biological feasibility of solutions. Accordingly, we design a positivity-preserving finite volume scheme based on an upwind flux approach, guaranteeing non-negative population densities and discrete conservation properties. We examine the convergence orders of the scheme through the numerical experiments and explore the effects of time delays on species competition dynamics under different parameter settings.
1 Introduction
The mathematical modeling of multi-species population dynamics provides a theoretical framework for studying species interactions in biological and ecological systems [1, 2]. In biochemical research, scientists employ co-culture systems to elucidate competitive relationships between distinct cell types, such as cancer cells and normal cells [3]. Early models included nonlinear diffusion and reaction-diffusion systems [4, 5, 6], revealing that spatial segregation can promote species coexistence by mitigating interspecific competition. A key approach in modern models involves nonlocal advection systems [7, 8], where population movement is influenced by long-range interactions instead of purely local diffusion. These models are often associated with chemotaxis, the directed movement of cells or organisms along chemical concentration gradients [9, 10, 11, 12]. The well-posedness of nonlocal advection models for two species (or a single species) has been studied in [7, 8, 13, 14]. Moreover, nonlocal advection systems are closely connected to continuum models of pedestrian dynamics, where an individual’s velocity is influenced not only by local density but also by spatially distributed crowd information [15, 16]. Extensions considering anisotropic interactions and domain boundaries further demonstrate the relevance of nonlocal frameworks to realistic crowd flow modeling [17, 18].
Time delays capture the inherent non-instantaneous nature of biological processes, yielding more realistic models that can exhibit oscillatory, bifurcation, and stability behaviors observed in real systems [19, 20]. Following the pioneering work of [21], numerous studies have introduced time delays in population dynamics models. In single species models such as the delayed logistic model [22, 23, 24], the delay represents the maturation time before individuals become reproductively active and can destabilize population dynamics by inducing oscillations or periodic behavior once it exceeds a critical threshold. The time delayed Lotka–Volterra predator-prey model [25, 23, 26] incorporates the predator’s delayed response to changes in prey population, enabling more accurate simulations of periodic oscillations and stability shifts in ecosystems. Time lags in competition models [27, 28] physically represent ecological processes such as resource recovery or interspecific interaction lags, and they can destabilize equilibrium states to induce stable periodic oscillations or even chaotic dynamics through Hopf bifurcation or period-doubling bifurcations.
This study aims to extend the existing nonlocal advection model for two competing species by incorporating time delays. Previous studies focused on population mobility and interactions, while our study introduces time delays to characterize the interaction between population migration and resource recovery. To ensure biologically realistic population densities, we develop a positivity-preserving finite volume scheme. We conduct extensive numerical experiments to: (1) examine the numerical convergence orders; (2) investigate the influence of time delays on competitive dynamics under various parameter settings.
In this work, we first introduce the mathematical model—the two species hyperbolic Keller–Segel system with time delays, and then design a finite volume scheme satisfying the positivity-preserving property, where we adopt the upwind type flux similar to [29–32, 33]. We investigate the experimental convergence orders of the proposed scheme. According to the numerical simulation, under various parameter settings, we find that small, identical delays in both species lead to a steady state, whereas larger delays may induce unsteady and potentially the extinction of one or both species. Moreover, asymmetric delay parameters between the two species tend to confer a competitive advantage to one species.
The rest of this paper is organized as follows. In Section 2, we extend the nonlocal advection Keller–Segel model by incorporating time delay and develop a positivity-preserving finite volume method that also satisfies discrete conservation laws. Numerical examples are presented in Section 3.
2 The mathematical model and the finite volume scheme
In this section, we introduce the time delay to the classical hyperbolic Keller–Segel system and design a positivity-preserving finite volume scheme.
2.1 PDE model
We consider the nonlocal advection system for two competing species with time delays in an interval
where
where
2.2 The finite volume scheme
We introduce the fully discrete finite volume scheme for problem Equations 2.1a–2.1f. We hereafter denote by
2.2.1 The finite volume scheme
For simplicity, we divide the computational domain
where
We denote by
We introduce the space of the piecewise constant function:
where
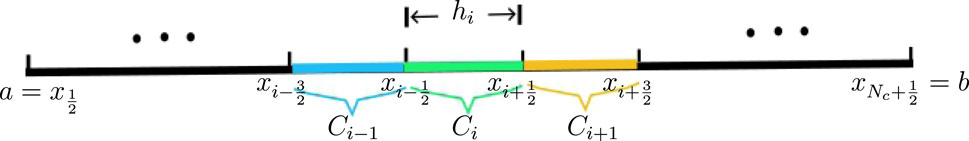
Figure 1. The mesh for the interval
We assume that there are two integers
Let
and we use the backward Euler to approximate time-differential, i.e.,
Let
By integrating Equations 2.1a–2.1c on each cell
Using
We use the approximation:
(by Equation 2.1d)
We apply the upwind discretization to treat the flux: for
where
For simplicity, we set the notations:
The time delay terms
where the integers
For
2.2.2 Positivity-preserving property
In this section, we show the positivity-preserving for the discrete solutions. We set
Theorem 2.1. Assume that
Proof. The discrete system Equations 2.4b, 2.4c can be rewritten into the matrix forms:
where
We assume that
The following statements hold for
1.
2.
3. All the column sums of
According to a convenient sufficient but not necessary condition of M-matrices (cf. [5, Appendix]), we conclude that
According to the properties of M-matrices, this implies that
2.2.3 The discrete conservation laws
The purpose of this section is to establish the mass conservation property of the (FVM) scheme (see Equations 2.4a–2.4c).
Theorem 2.2. Let
Moreover, under the assumptions that
Proof. Summing up Equations 2.4b, 2.4c with respect to
We see that
where
(by Equation 2.3).
Hence, we have
Moreover, if we assume that
Hence the proof is complete.
Remark 2.3. For hyperbolic systems, there exist other positivity-preserving strategies, such as the linear scaling limiter [35, 36], which enforces bounds in finite volume and discontinuous Galerkin methods through constraints at Legendre Gauss-Lobatto points.
3 Numerical experiments
In this section, we conduct extensive numerical experiments, where we examine the numerical convergence orders, compare the results with the classical hyperbolic Keller–Segel system, and investigate the influence of time delays on competitive dynamics under various parameter settings. We take
3.1 Example 1 (non-segregated initial functions)
We set
The initial conditions are chosen as follows:
To examine the influence of time delays on competitive dynamics under varying parameter settings, the following cases are considered.
1. the classical hyperbolic Keller–Segel system
2.
3.
To obtain the experimental convergence order of the mesh size
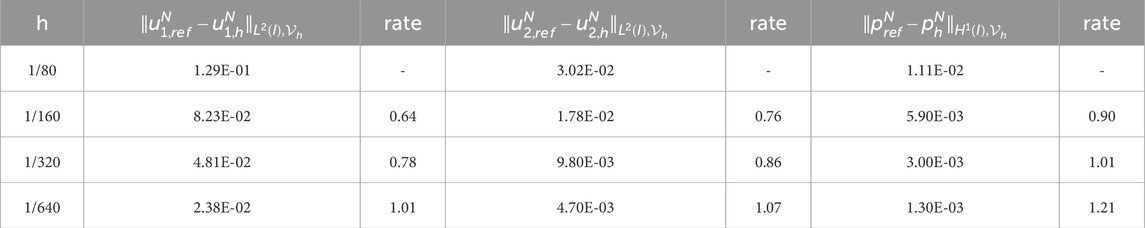
Table 1. Experimental errors and convergence order of
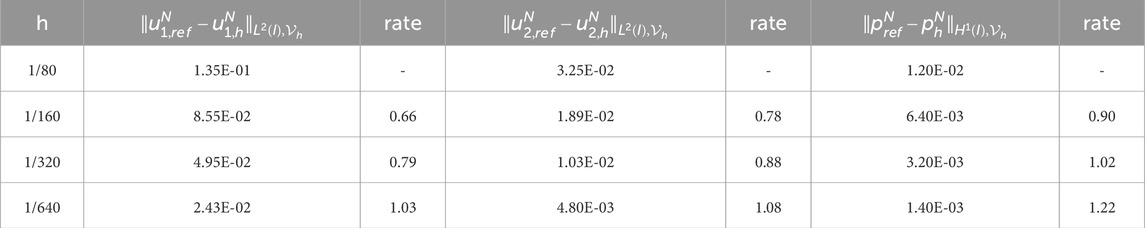
Table 2. Experimental errors and convergence order of
The evolution of the solutions for the classical hyperbolic Keller–Segel system (i.e.,
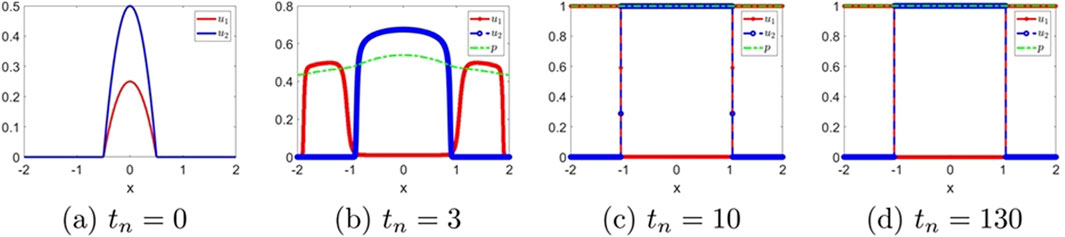
Figure 2. The numerical solution for
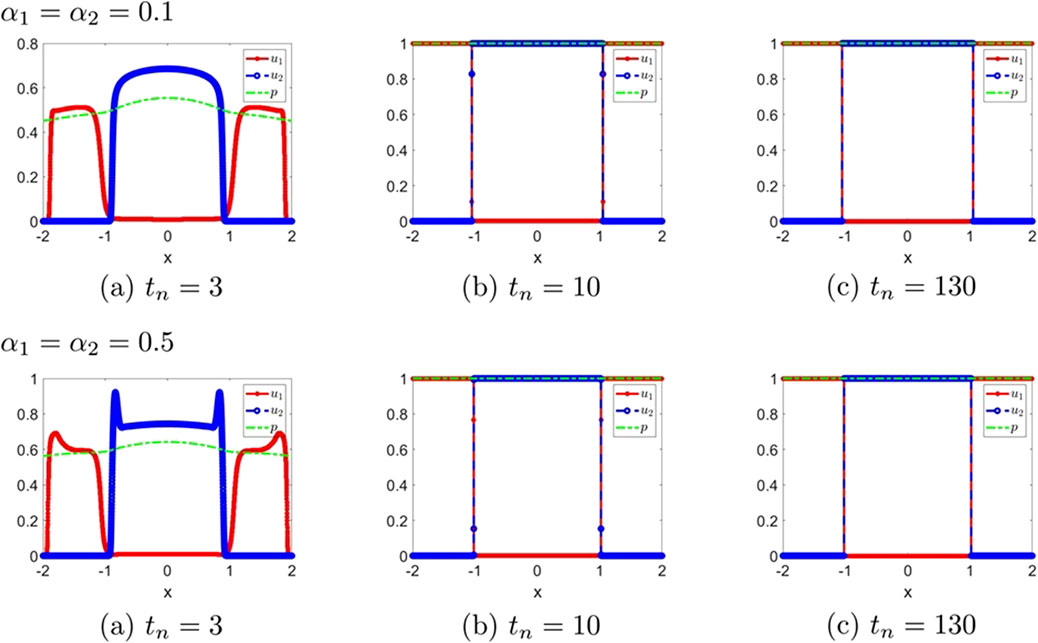
Figure 3. The numerical solution (Examples 1-2-1, 1-2-2). (a–c) show the evaluation of the numerical solution at different times.
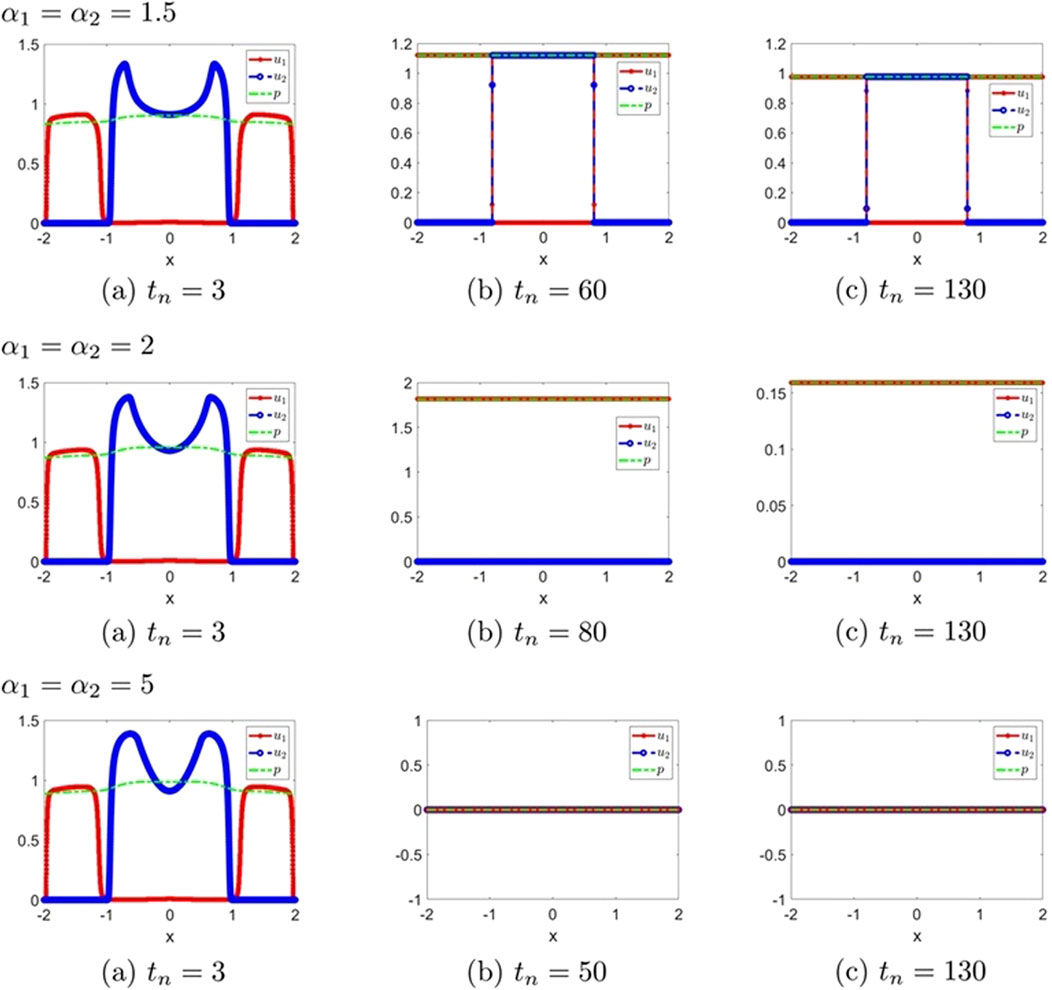
Figure 4. The numerical solution (Examples 1-2-3, 1-2-4, 1-2-5). (a–c) show the evaluation of the numerical solution at different times.
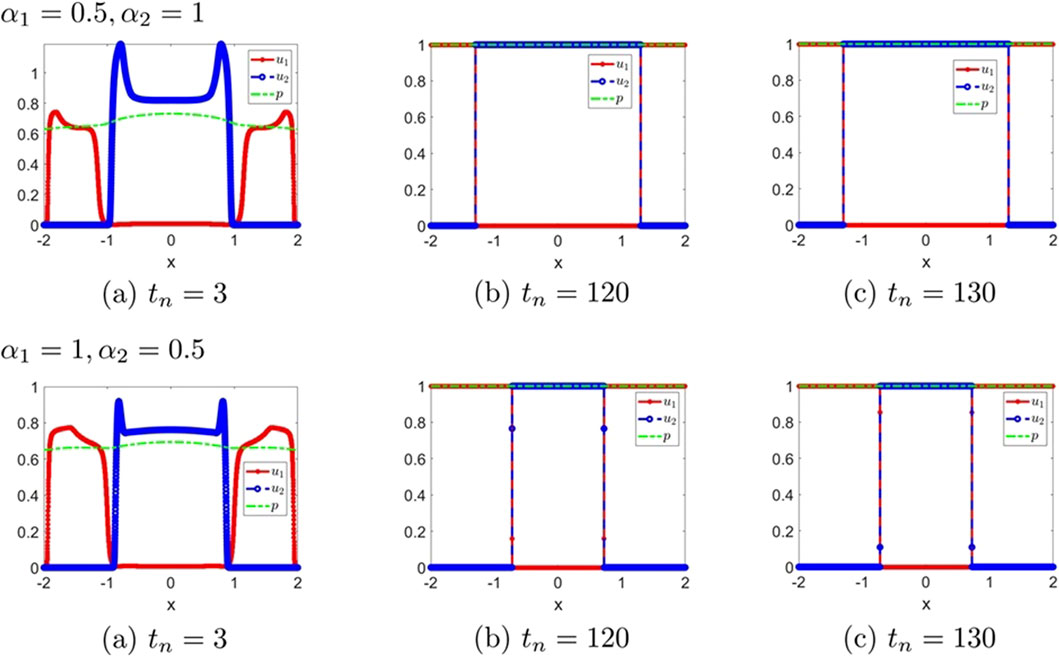
Figure 5. The numerical solution (Examples 1-2-6, 1-2-7). (a–c) show the evaluation of the numerical solution at different times.
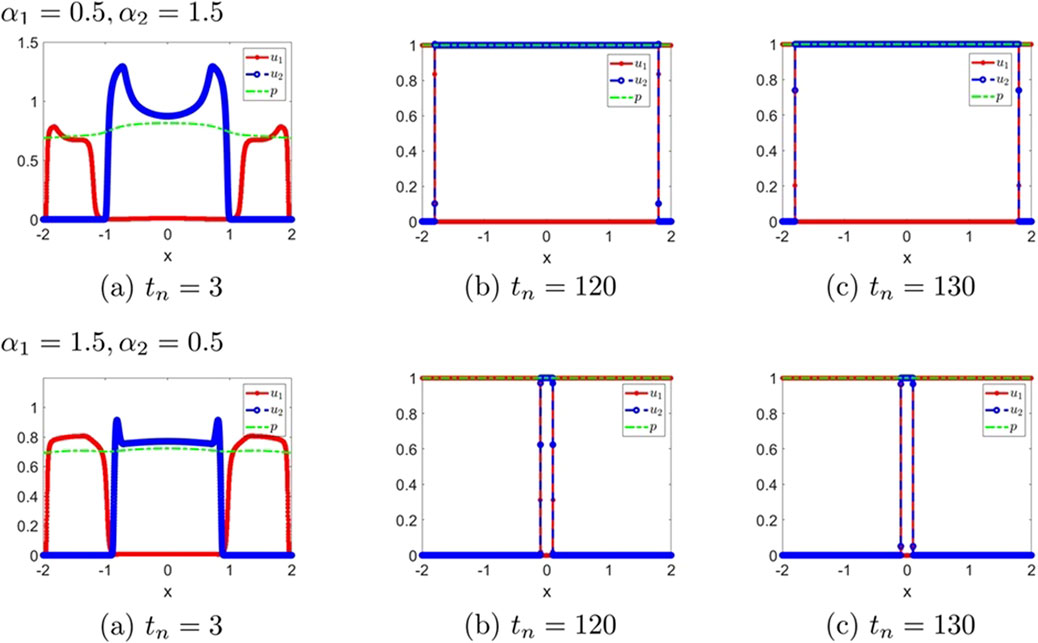
Figure 6. The numerical solution (Examples 1-2-8, 1-2-9). (a–c) show the evaluation of the numerical solution at different times.
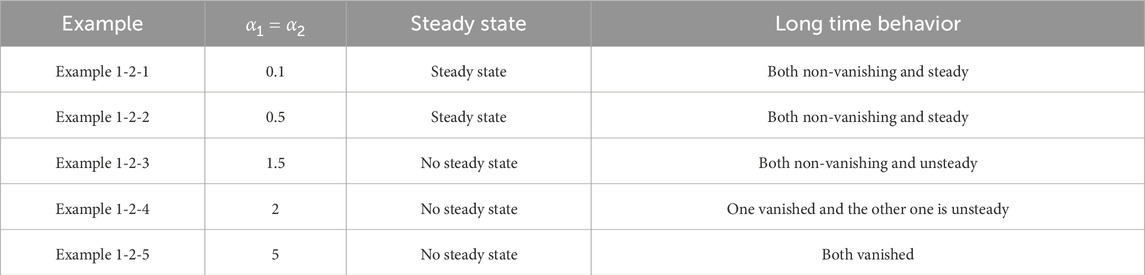
Table 3. Classification of asymptotic behavior
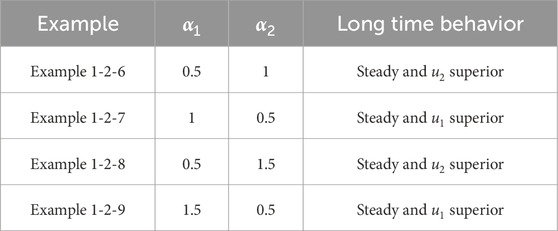
Table 4. Classification of asymptotic behavior
3.2 Example 2 (segregated initial functions)
Set
The initial conditions are set to
We consider the following settings.
1. the classical hyperbolic Keller–Segel system
2.
3.
We fix
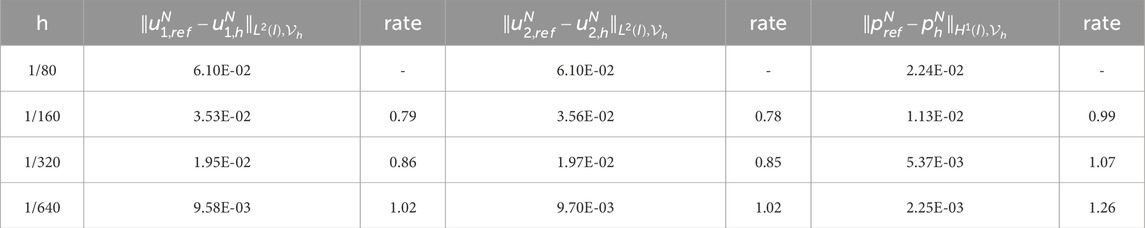
Table 5. Experimental errors and convergence order of
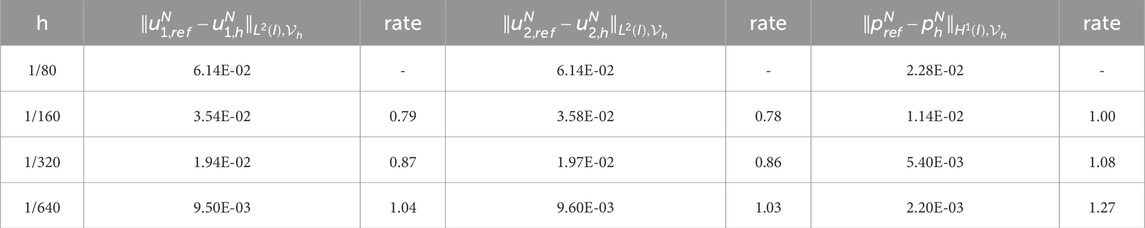
Table 6. Experimental errors and convergence order of
Figure 7 displays the evolution of solutions for the classical Keller–Segel system with
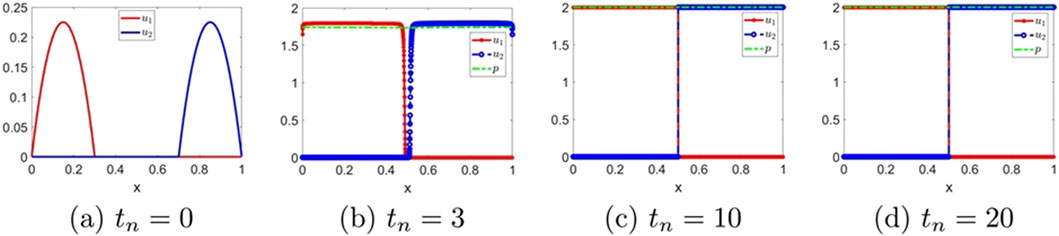
Figure 7. The numerical solution for
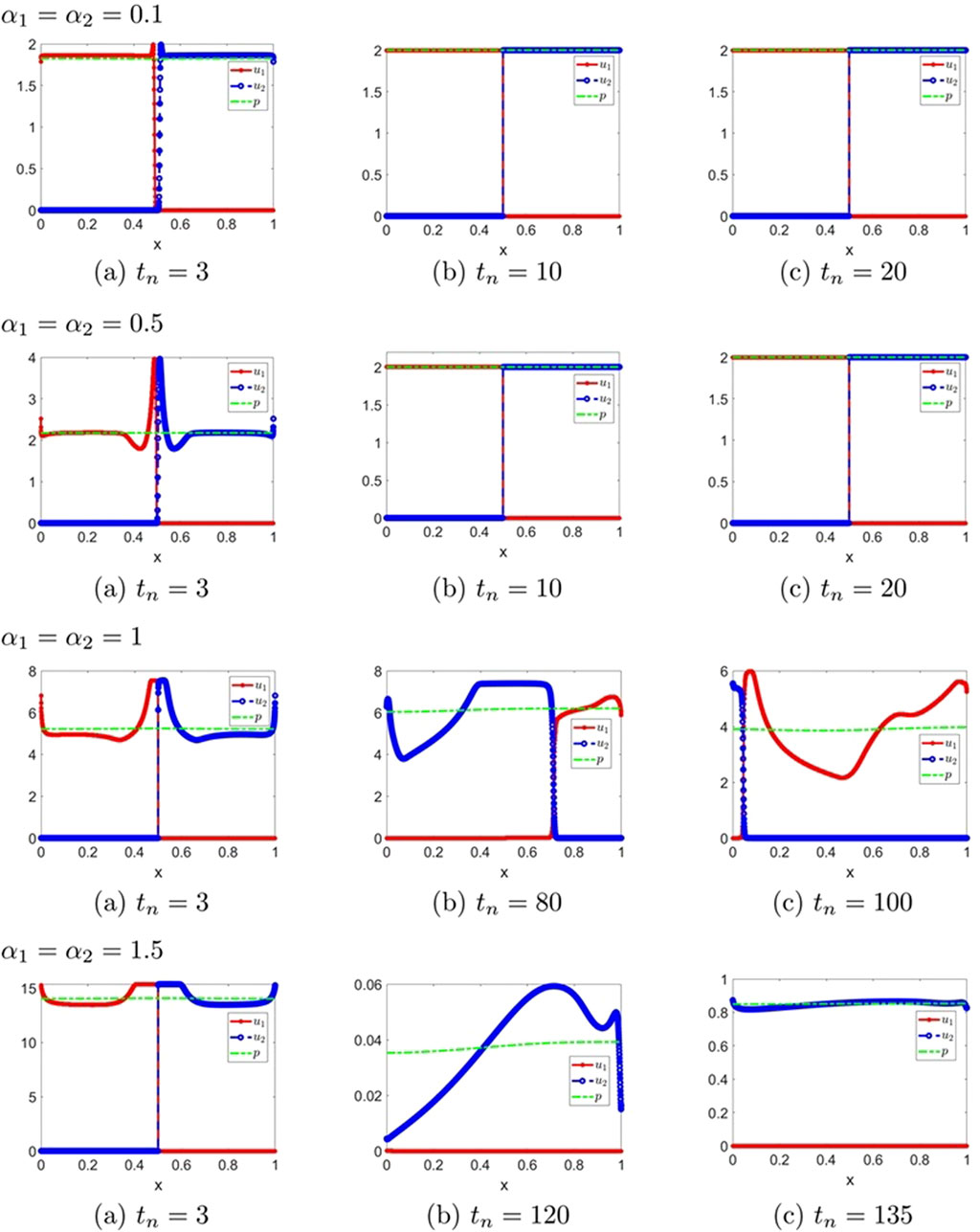
Figure 8. The numerical solution (Examples 2-2-1 through 2-2-4). (a–c) show the evaluation of the numerical solution at different times.
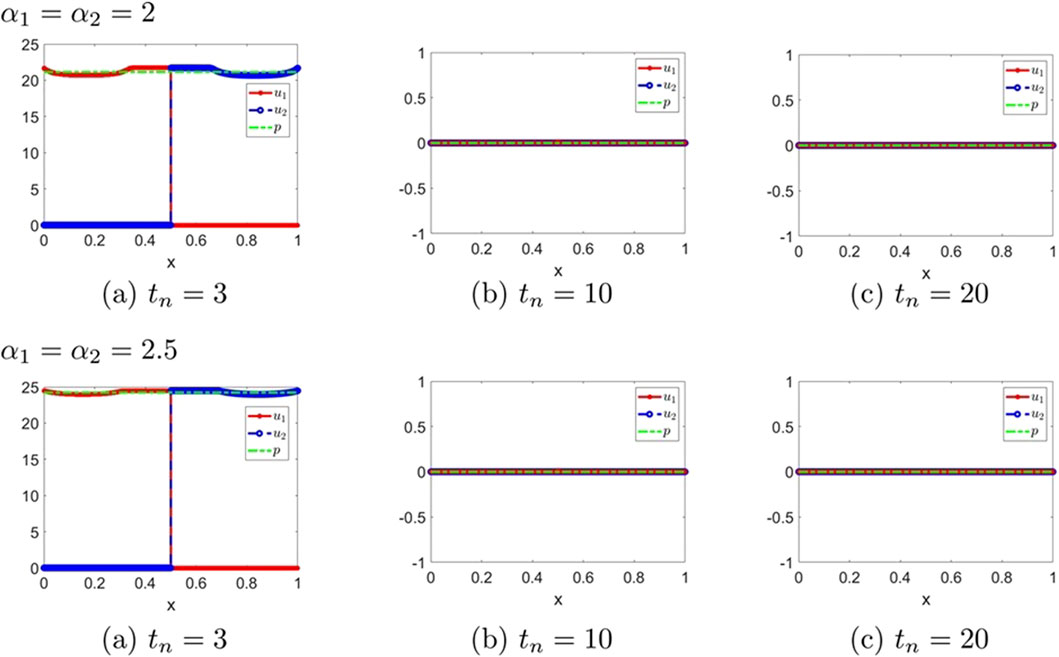
Figure 9. The numerical solution (Examples 2-2-5, 2-2-6). (a–c) show the evaluation of the numerical solution at different times.
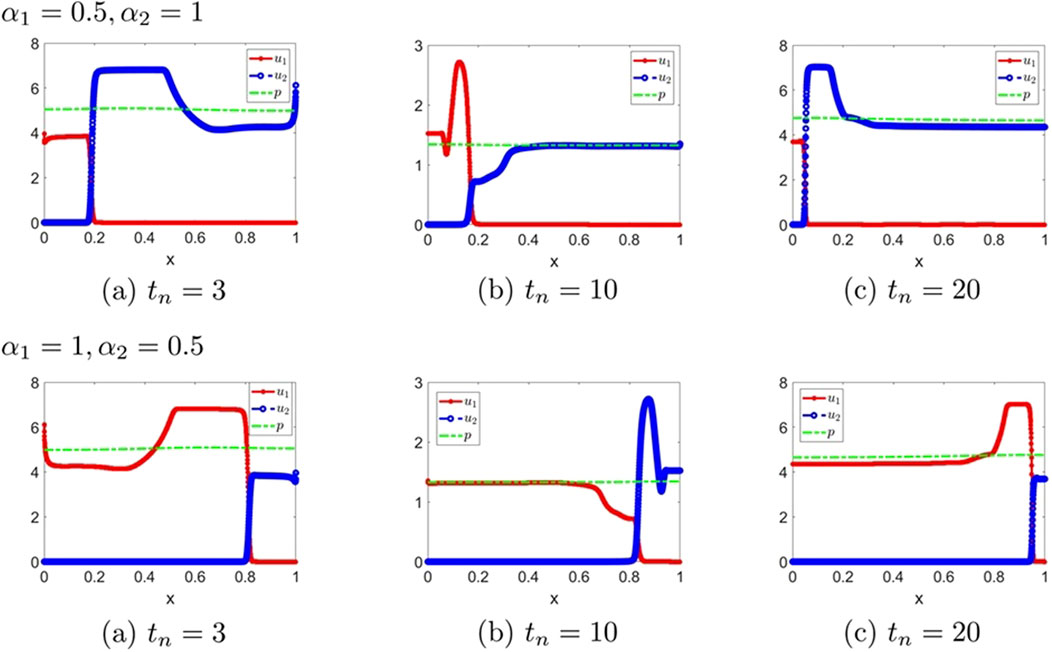
Figure 10. The numerical solution (Examples 2-2-7, 2-2-8). (a–c) show the evaluation of the numerical solution at different times.
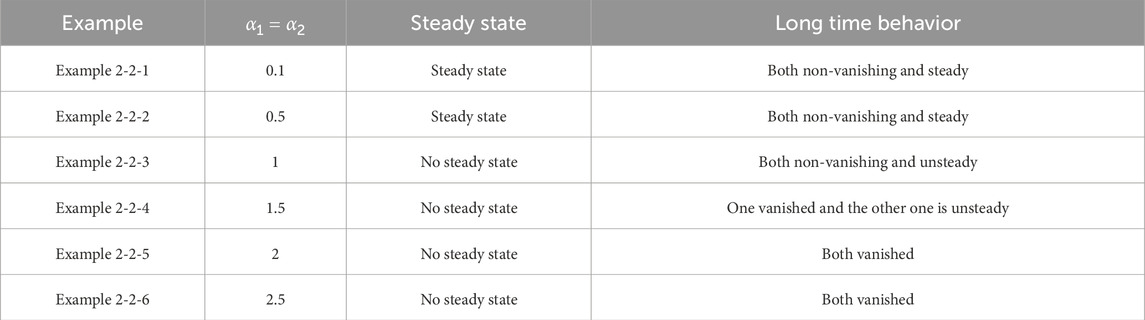
Table 7. Classification of asymptotic behavior
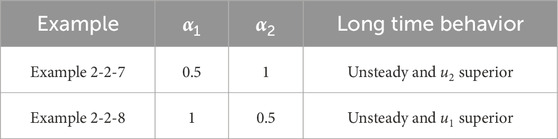
Table 8. Classification of asymptotic behavior
3.3 Example 3 (segregated initial functions)
The settings are the same as those in Example 2, except that we choose
The following parameters are considered.
1. the classical hyperbolic Keller–Segel system
2.
3.
We fix
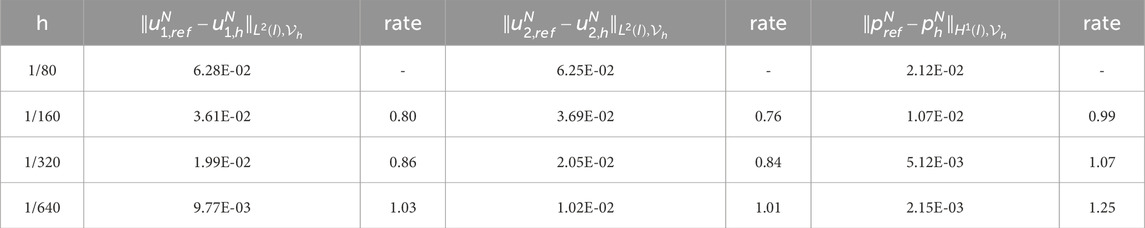
Table 9. Experimental errors and convergence order of
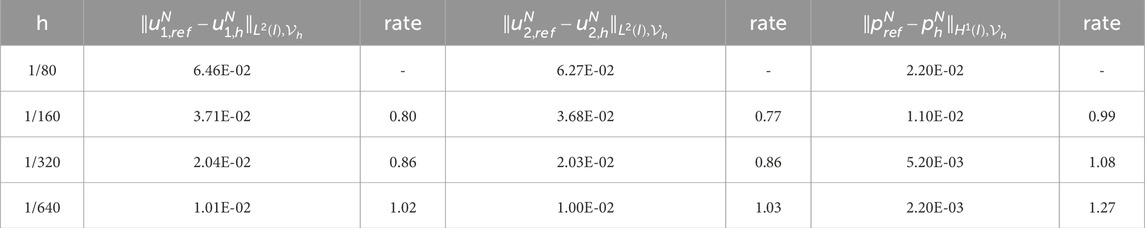
Table 10. Experimental errors and convergence order of
The evolutions of the solution for the classical Keller–Segel system
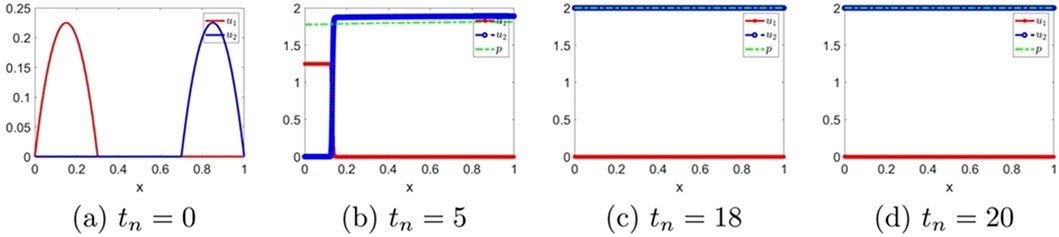
Figure 11. The numerical solution for
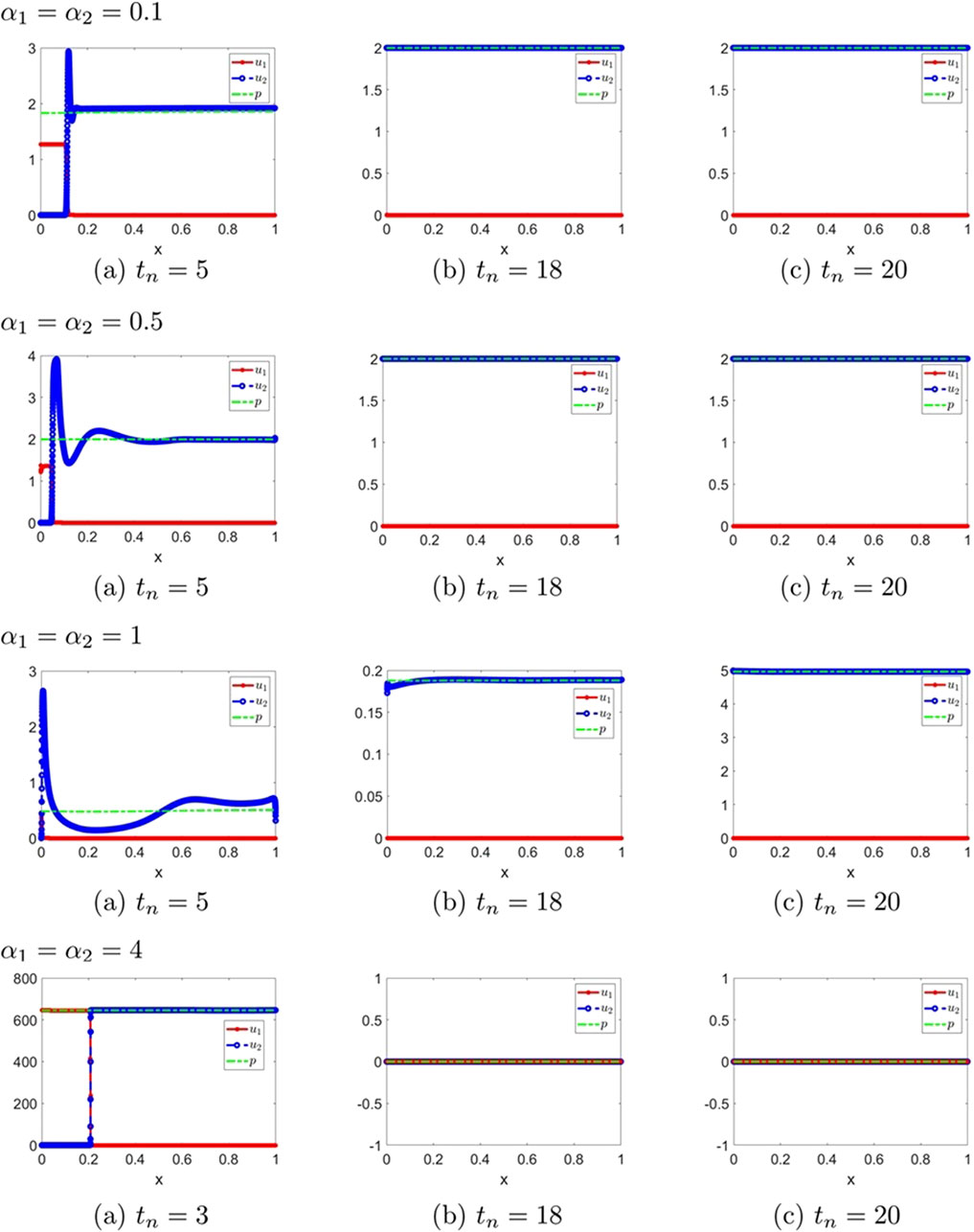
Figure 12. The numerical solution (Examples 3-2-1 through 3-2-4). (a–c) show the evaluation of the numerical solution at different times.
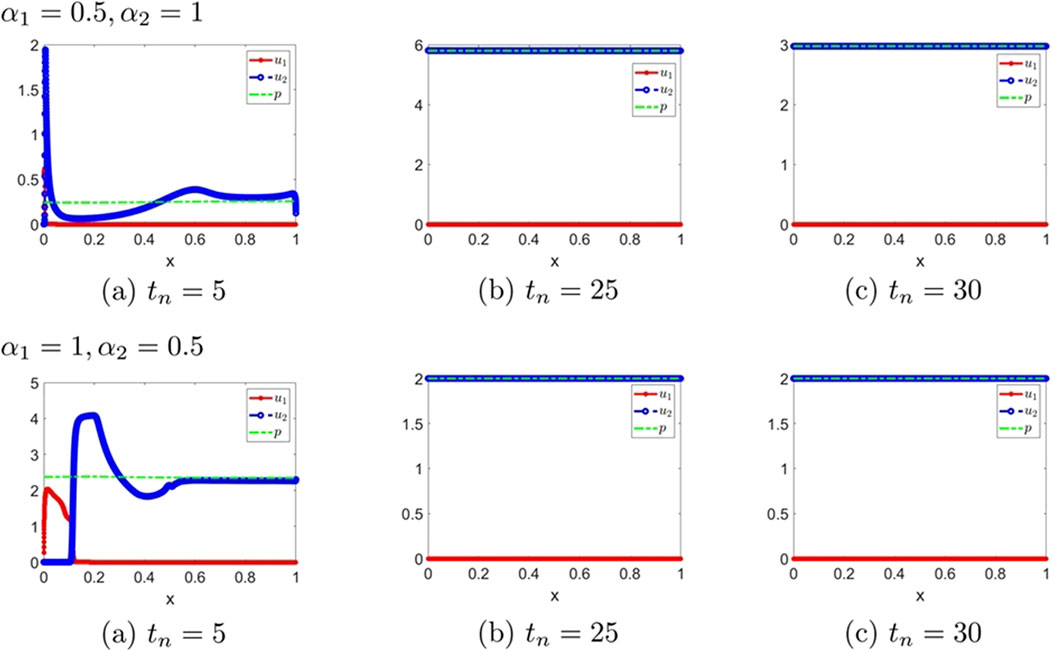
Figure 13. The numerical solution (Examples 3-2-5, 3-2-6). (a–c) show the evaluation of the numerical solution at different times.
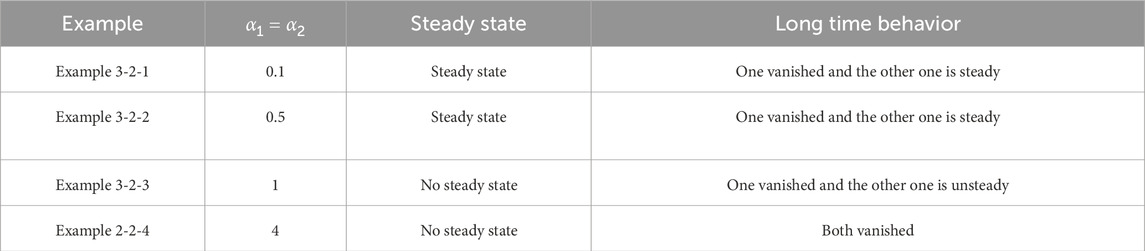
Table 11. Classification of asymptotic behavior
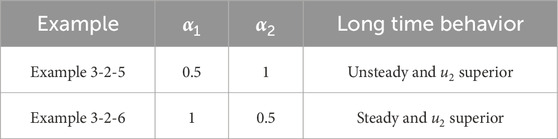
Table 12. Classification of asymptotic behavior
Remark 3.1. We observe that small symmetric delays
4 Conclusion
This study proposes a nonlocal advection model with time delays to investigate the population dynamics of two competing species, extending previous frameworks by incorporating delayed interactions between migration and resource recovery. A positivity-preserving finite volume scheme is developed to ensure biologically realistic solutions. Numerical experiments demonstrate that small symmetric delays
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
PZ: Writing – original draft, Data curation, Writing – review and editing, Methodology. YE: Writing – review and editing, Funding acquisition, Supervision, Methodology. GZ: Funding acquisition, Methodology, Supervision, Investigation, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by JSPS KAKENHI Grant Number 25K07137, NSFC General Project No. 12171071, and the Natural Science Foundation of Sichuan Province No. 2023NSFSC0055.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
3. Pasquier J, Galas L, Boulangé-Lecomte C, Rioult D, Bultelle F, Magal P. Different modalities of intercellular membrane exchanges mediate cell-to-cell p-glycoprotein transfers in MCF-7 breast cancer cells. J Biol Chem (2012) 287(10):7374–87. doi:10.1074/jbc.M111.312157
4. Lou Y, Ni W. Diffusion, self-diffusion and cross-diffusion. J Differ Equ (1996) 131(1):79–131. doi:10.1006/jdeq.1996.0157
5. Lou Y, Ni W. Diffusion vs cross-diffusion: an elliptic approach. J Differ Equ (1999) 154(1):157–90. doi:10.1006/jdeq.1998.3559
6. Shigesada N, Kawasaki K, Teramoto E. Spatial segregation of interacting species. J Theor Biol (1979) 79(1):83–99. doi:10.1016/0022-5193(79)90258-3
7. Fu X, Griette Q, Magal P. A cell-cell repulsion model on a hyperbolic Keller–Segel equation. J Math Biol (2020) 80(7):2257–300. doi:10.1007/s00285-020-01495-w
8. Fu X, Magal P. Asymptotic behavior of a nonlocal advection system with two populations. J Dyn Differ Equ (2022) 34(3):2035–77. doi:10.1007/s10884-021-09956-6
9. Horstmann D. From 1970 until present: the Keller–Segel model in chemotaxis and its consequences. Jahresber Dtsch Math.-Ver (2003) 195(1):103–65. Available online at: https://cir.nii.ac.jp/crid/1571698600200464128
10. Horstmann D. From 1970 until present: the Keller–Segel model in chemotaxis and its consequences. Jahresber Dtsch Math.-Ver (2004) 106:51–69. Available online at: https://cir.nii.ac.jp/crid/1570291226041248512
11. Keller EF, Segel LA. Model for chemotaxis. J Theor Biol (1971) 30(2):225–34. doi:10.1016/0022-5193(71)90050-6
12. Patlak CS. Random walk with persistence and external bias. Bull Math Biophys (1953) 15:311–38. doi:10.1007/bf02476407
13. Fu X, Griette Q, Magal P. Existence and uniqueness of solutions for a hyperbolic Keller–Segel equation, Discrete Contin. Dyn Syst Ser B (2021) 26(4):1931–66. doi:10.3934/dcdsb.2020326
14. Perthame B, Dalibard A. Existence of solutions of the hyperbolic Keller–Segel model. Trans Am Math Soc (2009) 361(5):2319–35. doi:10.1090/s0002-9947-08-04656-4
15. Bruno L, Tosin A, Tricerri P, Venuti F. Non-local first-order modelling of crowd dynamics: a multidimensional framework with applications. Appl Math Model (2011) 35(1):426–45. doi:10.1016/j.apm.2010.07.007
16. Colombo RM, Garavello M, Lécureux-Mercier M. A class of nonlocal models for pedestrian traffic. Math Models Methods Appl Sci (2012) 22(4):1150023. doi:10.1142/s0218202511500230
17. Bürger R, Goatin P, Inzunza D, Villada LM. A non-local pedestrian flow model accounting for anisotropic interactions and domain boundaries. Math Biosci Eng (2020) 17(5):5883–906. doi:10.3934/mbe.2020314
18. Goatin P, Inzunza D, Villada LM. Numerical comparison of nonlocal macroscopic models of multi-population pedestrian flows with anisotropic kernel. Springer (2022). p. 371–81.
20. Smith HL. An introduction to delay differential equations with applications to the life sciences. New York: Springer (2011).
21. Volterra V. Leçons sur la Théorie Mathématique de la Lutte pour la Vie. Paris: Gauthier-Villars (1931).
22. Cushing JM. Time delays in single species growth models. J Math Biol (1977) 4(3):257–64. doi:10.1007/bf00280975
23. Cushing JM. Integrodifferential equations and delay models in population dynamics, 20. Berlin: Springer (2013).
24. Miller RK. On Volterra’s population equation. SIAM J Appl Math (1966) 14(3):446–52. doi:10.1137/0114039
25. Bownds JM, Cushing JM. On the behavior of solutions of predator-prey equations with hereditary terms. Math Biosci (1975) 26(1-2):41–54. doi:10.1016/0025-5564(75)90093-0
26. Macdonald N. Time delay in prey-predator models. Math Biosci (1976) 28(3-4):321–30. doi:10.1016/0025-5564(76)90130-9
27. Gopalsamy K, Aggarwala BD. Limit cycles in two spacies competition with time delays. J Austral Math Soc (1980) 22(2):148–60. doi:10.1017/s033427000000223x
28. Shibata A, Saitô N. Time delays and chaos in two competing species. Math Biosci (1980) 51(3-4):199–211. doi:10.1016/0025-5564(80)90099-1
29. Saito N. Conservative upwind finite-element method for a simplified Keller–Segel system modelling chemotaxis. IMA J Numer Anal (2007) 27(2):332–65. doi:10.1093/imanum/drl018
30. Saito N. Error analysis of a conservative finite-element approximation for the Keller–Segel system of chemotaxis. Commun Pure Appl Anal (2011) 11(1):339–64. doi:10.3934/cpaa.2012.11.339
31. Saito N, Suzuki T. A finite difference scheme to the system of self-interacting particles. RIMS Kôkyûroku (2003) 1320:18–28. Available online at: http://hdl.handle.net/2433/43075
32. Saito N, Suzuki T. Notes on finite difference schemes to a parabolic-elliptic system modelling chemotaxis. Appl Math Comput (2005) 171(1):72–90. doi:10.1016/j.amc.2005.01.037
33. Zhou G, Saito N. Finite volume methods for a Keller–Segel system: discrete energy, error estimates and numerical blow-up analysis. Numer Math (2017) 135(1):265–311. doi:10.1007/s00211-016-0793-2
34. Cross L, Zhang X. On the monotonicity of Q2spectral element method for Laplacian on quasi-uniform rectangular meshes. Commun Comput Phys (2024) 35(1):60–180. doi:10.4208/cicp.oa-2023-0206
35. Zhang X, Shu C. On maximum-principle-satisfying high order schemes for scalar conservation laws. J Comput Phys (2010) 229(9):3091–120. doi:10.1016/j.jcp.2009.12.030
Keywords: two species model, time delay, hyperbolic PDE, Keller–Segel model, advection, nonlocal
Citation: Zeng P, Enatsu Y and Zhou G (2025) A nonlocal advection system for two competing species with resources recovering time. Front. Phys. 13:1657967. doi: 10.3389/fphy.2025.1657967
Received: 02 July 2025; Accepted: 19 September 2025;
Published: 03 November 2025.
Edited by:
Ryosuke Yano, Tokio Marine dR Co., Ltd., JapanReviewed by:
Abdellatif Ben Makhlouf, University of Sfax, TunisiaNorikazu Saito, The University of Tokyo, Komaba, Japan
Copyright © 2025 Zeng, Enatsu and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yoichi Enatsu, eWVuYXRzdUBycy50dXMuYWMuanA=