Abstract
Japanese medaka (Oryzias latipes) is an acceptable small laboratory fish model for the evaluation and assessment of endocrine-disrupting chemicals (EDCs) found in the environment. In this research, we used this fish as a potential tool for the identification of EDCs that have a significant impact on human health. We conducted an electronic search in PubMed (http://www.ncbi.nlm.nih.gov/pubmed) and Google Scholar (https://scholar.google.com/) using the search terms, Japanese medaka, Oryzias latipes, and endocrine disruptions, and sorted 205 articles consisting of 128 chemicals that showed potential effects on estrogen–androgen–thyroid–steroidogenesis (EATS) pathways of Japanese medaka. From these chemicals, 14 compounds, namely, 17β-estradiol (E2), ethinylestradiol (EE2), tamoxifen (TAM), 11-ketotestosterone (11-KT), 17β-trenbolone (TRB), flutamide (FLU), vinclozolin (VIN), triiodothyronine (T3), perfluorooctanoic acid (PFOA), tetrabromobisphenol A (TBBPA), terephthalic acid (TPA), trifloxystrobin (TRF), ketoconazole (KTC), and prochloraz (PCZ), were selected as references and used for the identification of apical endpoints within the EATS modalities. Among these endpoints, during classification, priorities are given to sex reversal (masculinization of females and feminization of males), gonad histology (testis–ova or ovotestis), secondary sex characteristics (anal fin papillae of males), plasma and liver vitellogenin (VTG) contents in males, swim bladder inflation during larval development, hepatic vitellogenin (vtg) and choriogenin (chg) genes in the liver of males, and several genes, including estrogen–androgen–thyroid receptors in the hypothalamus–pituitary–gonad/thyroid axis (HPG/T). After reviewing 205 articles, we identified 108 (52.68%), 46 (22.43%), 19 (9.26%), 22 (17.18%), and 26 (12.68%) papers that represented studies on estrogen endocrine disruptors (EEDs), androgen endocrine disruptors (AEDs), thyroid endocrine disruptors (TEDs), and/or steroidogenesis modulators (MOS), respectively. Most importantly, among 128 EDCs, 32 (25%), 22 (17.18%), 15 (11.8%), and 14 (10.93%) chemicals were classified as EEDs, AEDs, TEDs, and MOS, respectively. We also identified 43 (33.59%) chemicals as high-priority candidates for tier 2 tests, and 13 chemicals (10.15%) show enough potential to be considered EDCs without any further tier-based studies. Although our literature search was unable to identify the EATS targets of 45 chemicals (35%) studied in 60 (29.26%) of the 205 articles, our approach has sufficient potential to further move the laboratory-based research data on Japanese medaka for applications in regulatory risk assessments in humans.
1 Introduction
Due to the increase in industrial and agricultural activities, endocrine-disrupting chemicals (EDCs), defined by the World Health Organization (WHO) as “Exogeneous substances that alter function(s) of the endocrine system and consequently cause adverse health effects in an intact organism or its progeny, or (sub)populations,” are accumulated in the environment. A strategic approach to identify EDCs would be utilized by the existing knowledge to prioritize and focus on the screening and environmental monitoring efforts of these chemicals. The European Commission also set criteria for the identification of EDCs that require regulatory action. Currently, endocrine disruptors (EDs) are identified on a case-by-case basis using the available guidance provided in the OECD Guidance Document 150 (2018). The OECD Conceptual Framework for Testing and Assessment of EDs provided a tiered framework for the organization of study information to assess endocrine activity. This framework provides guidance for prioritizing relevant data streams and methods according to the type and level of information needed for a regulatory assessment. In the USA, EPA’s EDSP has developed the requirements for the prioritization, screening, and testing of environmental contaminants, including pesticides, commercial chemicals, and agricultural products, for their potential to impact the endocrine system, especially in relation to estrogen, androgen, and thyroid (EAT) hormones and their nuclear receptors (NIEHS, 2018). Moreover, the perturbation of the enzymes of steroidogenesis by EDCs has potential effects on EAT pathways. Therefore, a two-tier testing approach was designed by EDSP. Tier 1 assays detect the potential effects of a chemical by various modes of action (Tier 1: screening) on EATS pathways. The results of the Tier 1 assays are evaluated by using a “weight of evidence” approach to determine whether the potential of the chemical is to interact with EATS and whether a Tier 2 assay is necessary. The purpose of Tier 2 studies is to use in vivo testing to further characterize the EATS effects and establish a dose–response relationship for adverse effects produced by the chemicals. Tier 2 tests are much longer-term studies that include exposure during critical life stages and have a broad range of more tightly spaced treatment than Tier 1. Moreover, Tier 2 tests can encompass multiple generations, covering effects on fecundity and fertility, development, growth, and sexual maturity. The successful completion of Tier 2 testing provided information to establish exposure and effect relationships, and assessed relevant endpoints across most life stages.
In aquatic environments, fish are considered one of the primary risk organisms for EDCs, especially those interacting with reproductive hormones. Sex determination in fish is very labile and can be disrupted or functionally reversed by external agents at critical developmental stages (Francis, 1992). Fish populations are directly exposed to a wide variety of EDCs, originating from industrial, agricultural, or municipal effluents (Ternes et al., 1999; Chen et al., 2007; Kim et al., 2014a). Evidence shows that EDCs can have long-term effects on reproduction and subsequent population development in natural fish populations (Kidd et al., 2007). The effects of EDCs on nuclear receptors have been studied extensively in small fish models like zebrafish, Japanese medaka, stickleback, and roach (Rutilus rutilus) (Iguchi et al., 2006; Lange et al., 2009; Tohyama et al., 2015). Since endocrine disruptions are linked to the receptor level, to predict ED effects, the identification of appropriate biomarkers at molecular levels is necessary.
Japanese medaka (Oryzias latipes) fish are small, freshwater teleost fish that inhabit gently flowing rivers and waterways. Like zebrafish (Danio rerio) and fathead minnows (Pimephales promelas), it is one of the small fish models (vertebrate) used in EDC studies (OECD, 2018). The sex determination locus has been identified in this fish species, and external sex-specific markers (chromatophores, shape of the anal and dorsal fins, anal fin papillae) can be used to easily differentiate males from females both from phenotypic and genotypic standpoints (Scholz and Mayer, 2008). Several OECD test guidelines (OECD TG 229; OECD TG 240) were used during the evaluation of EDCs in Japanese medaka, following tier-based approaches (Tier 1 and Tier 2). Moreover, the effects of endocrine active chemicals on Japanese medaka were reviewed previously (Urushitani et al., 2007; Flynn et al., 2017; Onishi et al., 2021; Kawashima et al., 2022). Based on the available publications found in public databases, we hypothesized that a literature search can identify the number and sources of EDCs that disrupted the EATS-related pathways of Japanese medaka (O. latipes) and correlate the effects with specific receptors at the molecular level.
In this review, we summarized the data on EDCs available in public databases, highlighting the links between molecular, phenotypic, and physiological endpoints using Japanese medaka as a single fish species. Although majority of the data refer to interfering with reproductive and thyroid hormone signaling pathways (EATS), limited information about the disruption of other endocrine organs, like the endocrine pancreas and interrenal gland (fish homolog of the adrenal gland), is also available (Dasmahapatra and Tchounwou, 2022a; b, 2023a; b). We evaluated the selective effects of 128 EDCs reported in 205 articles. As a result, we believed that 43 of them (EDCs) show potential to proceed to Tier 2 tests, and 13 chemicals should be considered EDCs without any further tier-based studies.
2 Materials and methods
2.1 Literature search strategy
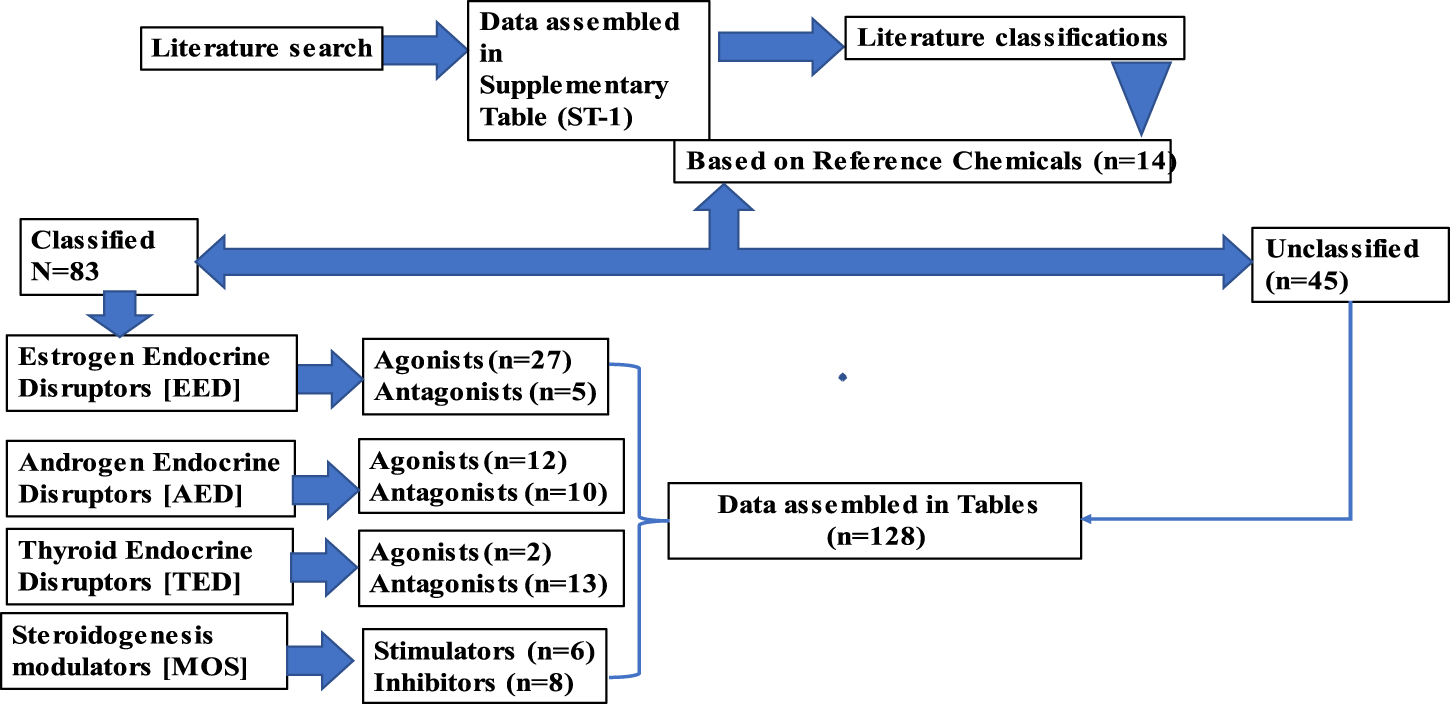
The objectives of the literature search were to identify the relevant studies published in peer-reviewed journals that focused on the endocrine disruption of Japanese medaka (O. latipes) induced by various chemicals detected in the aquatic environments. The search was performed in PubMed (http://www.ncbi.nlm.nih.gov/pubmed) and Google Scholar (https://scholar.google.com/). PubMed was considered the main and reliable source of information; however, Google Scholar was used if the full text article was not available in PubMed. We initiated our search in PubMed using the search term Japanese medaka (Oryzias latipes), which provided 3,747 results (until 30 June 2023). We narrowed down the search by adding the term “endocrine” (Japanese medaka, Oryzias latipes, and endocrine), which reduced the number to 646, and finally, the addition of the term “endocrine disruption” reduced the results to 239 (Figure 1). We finally sorted 205 articles for review that focused on EATS pathways of Japanese medaka (Figure 1). We identified 128 chemicals that have potential ED effects on this fish (Japanese medaka, O. latipes) (Figure 2). After a literature search, we assembled ED-related information in Supplementary Table S1, which was also deposited in Figshare (doi/10.6084/mg.figshare. 22598068). For classification of these compounds as selective disruptors of EATS pathways, 14 chemicals from 128 searched chemicals were selected as reference chemicals (Figure 2; Table 1). For estrogen endocrine disruptors (EEDs), E2 and EE2 were used as reference chemicals for agonists, and TAM was used for antagonists. For androgen endocrine disruptors (AEDs), 11-KT and TRB were used for agonists, and FLU and VIN were used for antagonists. For thyroid endocrine disruptors, (TEDs), T3 was used for agonists, and PFOA and TBBPA were used for antagonists. For steroidogenesis, TPA and TRF were used for stimulators, and KTC and PCZ were used for inhibitors (Table 1). After critical evaluation of the ED effects of these reference chemicals, the criteria of evaluation of endocrine disruption induced by an EDC on Japanese medaka are determined (Table 1). The chemicals which were unable to fulfill the criteria were considered unclassified.
FIGURE 1

Strategies for the selection of literature reports from peer-reviewed articles published on Japanese medaka (Oryzias latipes).
FIGURE 2
Strategies for the selection of chemicals from peer-reviewed articles published on Japanese medaka (O. latipes).
TABLE 1
| Endocrine targets | Reference chemicals | Agonists/Antagonists/Stimulator/Inhibitor | Affinity | Literature | End points | |
| Agonists | Antagonists | |||||
| EED | E2 | Agonist | In vitro reporter gene assay (affinity) (Tohyama et al., 2015)i) esr1: EC50 = 1.31 × 10−10 Mii) esr1: EC50 = 1.31 × 10−10 Miii) esr2b: EC50 = 8.16 × 10−11 M | Nimrod and Benson (1998), Patyna et al. (1999), Foran et al. (2000), Koger et al. (2000), Shioda and Wakabayashi (2000), Metcalfe et al. (2001), Tabata et al. (2001), Kang et al. (2002a), Kashiwada et al. (2002), Oshima et al. (2003), Balch et al. (2004b), Hall et al. (2005), Zeng et al. (2005), Balch and Metcalfe (2006), Hirai et al. (2006), Zhang et al. (2008d), Sun et al. (2009), Jin et al. (2011a), Kamata et al. (2011), Hirakawa et al. (2012), Lee et al. (2012), Flynn et al. (2013), Lei et al. (2013), Green et al. (2015), Tohyama et al. (2015), Inagaki et al. (2016), Flynn et al. (2017), Lee et al. (2017a), Lee Pow et al. (2017), Bertotto et al. (2019), Kang et al. (2019), Myosho et al. (2019), Ishibashi et al. (2020), Spirhanzlova et al. (2020), Onishi et al. (2021), Pandelides et al. (2021), Horie et al. (2022a). | 1. Female biased sex ratio, testis-ova (male feminization) | |
| 2. Significant decrease in fecundity | ||||||
| 3. Serum VTG level increased in males and females | ||||||
| 4. Inhibition of swim bladder inflation | ||||||
| 5. Intersex gonad | ||||||
| 6. Increased HSI in males | ||||||
| 7. Secondary sexual features reduced | ||||||
| 8. gsdf expression in XY embryos remained unaltered | ||||||
| 9. Vacuolization of hepatocytes in the liver | ||||||
| 10. Hydropic degeneration in glomerulus of the kidney | ||||||
| 11. Reproductive behavior suppressed with in both sexes | ||||||
| 12. No effect on male sexual behavior | ||||||
| 13. Histological structure of the kidney disrupted | ||||||
| 14. Gene expression | ||||||
| A: Brain: | ||||||
| (i) Male: | ||||||
| 1. Upregulation of gnrh1, cyp19b, esr1, fshβ, lhβ | ||||||
| 2. Downregulation of gnrhR1, gnrhR2, arα | ||||||
| (ii) Female: | ||||||
| 1. Upregulation of esr1 and esr2a | ||||||
| 2. cyp19b remained unaltered | ||||||
| B. Liver: | ||||||
| (i) Male: | ||||||
| 1. Upregulation of esr1, vtg 1, vtg 2, chgH, chgHm, chgL, cyp1c | ||||||
| 2. No alteration in esr1, esr2a, and arα | ||||||
| (ii) Female: | ||||||
| 1. vtg1 and vtg2 remained unaltered | ||||||
| In vitro reporter gene assay (Onishi et al., 2021): EC50 = 0.00098 µM (medaka esr1); IC50 = 2.0 µM (medaka arβ) | 2. esr2a and arα remained unaltered | |||||
| C. Gonad | ||||||
| i) Testis: | ||||||
| 1. Aromatase expression increased | ||||||
| 2. DNA methylation pattern reduced | ||||||
| 3. esr1 transcripts decreased | ||||||
| 4. Downregulation of cyp11a, cyp17 | ||||||
| 5. fshR reduced | ||||||
| (ii) Ovary: | ||||||
| 1. Downregulation of cyp19a | ||||||
| EE2 | Agonist | In vitro reporter gene assay: EC50 = 0.00088 µM (Medaka esr1 agonist assay); IC50 = 0.14 µM (medaka arβ antagonist assay (Onishi et al., 2021) | Scholz and Gutzeit (2000), Metcalfe et al. (2001), Foran et al. (2002), Islinger et al. (2002), Lee et al. (2002), Seki et al. (2002), Balch et al. (2004a), Nozaka et al. (2004), Chikae et al. (2004), Hano et al. (2005), Zeng et al. (2005), Orn et al. (2006), Zhang et al. (2008c), Hashimoto et al. (2009), Park et al. (2009), Sun et al. (2011a), Hirakawa et al. (2012), Liao et al. (2014), Schiller et al. (2014), Abdel-Moneim et al. (2015), Bhandari et al. (2015), Bhandari et al. (2020), Thayil et al. (2020), Onishi et al. (2021), Pandelides et al. (2021), Horie et al. (2022a), Myosho et al. (2022) | |||
| TAM | Antagonist | In vitro reporter gene assay: IC50 = 0.14 µM (medaka esr1 antagonist assay; (Onishi et al., 2021) | Chikae et al. (2004), Sun et al. (2007a), Sun et al. (2011a), Flynn et al. (2017), Onishi et al. (2021). | 1. Significant reduction in fecundity | ||
| 2. Reduction in hatchability, hatching delay, developmental abnormalities | ||||||
| 3. Liver VTG in male increased, reduced in females | ||||||
| 4. Liver histology of male fish disrupted | ||||||
| 5. HSI remained unaltered | ||||||
| 6. Secondary sexual features reduced | ||||||
| A: Brain: | ||||||
| i) Male: | ||||||
| 1. Upregulation of arα, esr1 and cyp19a | ||||||
| 2. Downregulation of cyp19b | ||||||
| ii) Female: | ||||||
| 1. No alteration in esr1 and esr2 | ||||||
| 2. Upregulation of arα and cyp19a | ||||||
| 3. Downregulation of cyp19b | ||||||
| B: Liver: | ||||||
| i) Male: | ||||||
| 1. Upregulation of vtg1, vtg2, esr1, arα | ||||||
| 2. No alteration in esr2a | ||||||
| ii) Female: | ||||||
| 1. Upregulation of esr2a | ||||||
| 2. Downregulation of vtg1 and vtg2 and esr1 | ||||||
| 3. arα remained unaltered | ||||||
| C; Gonad | ||||||
| i) Testis: | ||||||
| 1. Upregulation of StAR, cyp19b | ||||||
| 2. Downregulation of esr1 and esr2a, cyp17a, cyp17b, cyp19a | ||||||
| 3. cyp11a, cyp11b remained unaltered | ||||||
| ii) Ovary: | ||||||
| 1. Upregulation of StAR, cyp11a | ||||||
| 2. Downregulation of esr1, esr2a and arα | ||||||
| 3. cyp11b remained unaltered | ||||||
| AED | 11-KT | Agonist | In vitro reporter gene assay: IC50 = 0.0027 (medaka arβ agonist assay; Onishi et al., 2021) | Asahina et al. (1989), Leon et al. (2007), Leon et al. (2008), Grillitsch et al. (2010), Onishi et al. (2021), Watanabe et al. (2023). | 1. Increased anal fin papillary processes (masculinization in females) | |
| 2. Enhancement of growth is sex-specific (males are larger than females) | ||||||
| 3. Hypertrophy in thyroid follicular cells induced in both sexes | ||||||
| 4. Germ cell necrosis is induced in both sexes | ||||||
| 5. Male biased sex-ratio | ||||||
| 6. Upregulation of gsdf mRNA in XX embryos (sex reversal of the XX fish) | ||||||
| 7. Ovo-testis | ||||||
| 8. Decrease in VTG content in females | ||||||
| 9. Gene Expression | ||||||
| A: Brain: | ||||||
| TRB | Agonist | In vitro reporter gene assay: EC50 = 0.0036 µM(medaka arβ agonist assay; Onishi et al., 2021) | Orn et al. (2006), Seki et al. (2006), Zhang et al. (2008b), Park et al. (2009), Grillitsch et al. (2010), Flynn et al. (2013), Flynn et al. (2017), Abdel-Moneim et al. (2015), Mazukami-Murata et al. (2016), Kang et al. (2019), Onishi et al. (2021), Horie et al. (2022a), Myosho et al (2022). | i) Female | ||
| 1. Upregulation of gnrhR2, cyp19b | ||||||
| B: Liver: | ||||||
| 1. down regulation of vtg1, vtg2, chgH, chgHm mRNAs in the both sexes | ||||||
| 2. Down regulation of esr1 in males | ||||||
| 3. Upregulation of cyp3A and annexin max2 in females | ||||||
| C; Gonad | ||||||
| (i) Testis: | ||||||
| 1. Down regulation of StAR and cyp11b | ||||||
| (ii) Ovary: | ||||||
| 1. Upregulation of cyp19a | ||||||
| 2. Upregulation of a | ||||||
| FLU | Antagonist | In vitro reporter gene assay: IC50 = 12 µM (medaka arβ agonist assay; Onishi et al. (2021), Horie et al. (2022a) | Chikae et al. (2004), Nozaka, (2004), Kang et al. (2006), Leon et al. (2007), Leon et al. (2008), Nakamura et al. (2014b), Schiller et al. (2014), Onishi et al. (2021). | 1. Increased plasma VTG levels in females, not in males | ||
| 2. Hepatic VTG unaltered in males, decreased in females | ||||||
| 3. Fecundity and fertility were significantly decreased | ||||||
| 4. Growth (length and weight) was inhibited in males, not in females (sex-specific) | ||||||
| 5. No sex reversal | ||||||
| 6. Formation of testis-ova; disruption of spermatogenesis and ovarian cell necrosis | ||||||
| VIN | Antagonist | In vitro reporter gene assay: IC50 = 5.1 µM(medaka arβ antagonist assay; Onishi et al., 2021). | Kiparissis et al. (2003b), Nakamura et al. (2014b), Sun et al. (2016b), Flynn et al. (2017), Onishi et al. (2021) | 7. Hypertrophy of thyroid follicular cells in males | ||
| 8. Decrease in papillary process in the anal fin of male fish; females did not develop papillary process in the anal fin | ||||||
| 9. Reduced expression of gnrhr2, cyp11b and 3βhsd | ||||||
| 10. Repressed esr2a and cyp19a1b | ||||||
| 11. No significant induction of gsdf expression in XY embryos | ||||||
| 12. Gene expression: | ||||||
| A: Brain: | ||||||
| (i) Male: | ||||||
| 1. Upregulation of esr2a, arα and cyp19a and cyp19b genes | ||||||
| B: Testis: | ||||||
| 1. esr1 and cyp17b mRNAs were upregulated | ||||||
| 2. cyp19a and cyp19b downregulated | ||||||
| TED | T3 | Agonist | Godfrey et al. (2019), Horie et al. (2022d) | 1. Decrease in the surface area of swim bladder in females | ||
| 2. Upregulation of trα and trβ mRNAs | ||||||
| PFOA | Antagonists | Ji et al. (2008), Lee et al. (2017b), Kang et al. (2019), Godfrey et al. (2019) | 1. Fecundity suppressed | |||
| 2. Females displayed larger swim bladder | ||||||
| 3. Thyroid follicles showed hyperplasia, hypertrophy, and colloidal depletion | ||||||
| 4. No change in sex ratio | ||||||
| TBBPA | Antagonists | Horie et al. (2023a) | 5. In liver, vtg1 in males, and vtg1 and vtg2 in females increased; VTG protein in males reduced | |||
| 6. chgH and chgHm mRNA expression in liver of males | ||||||
| 7. Increased tshβ, trβ and vtg in females | ||||||
| 8. Upregulation of esr2a and vtg in males | ||||||
| 9. No effect on expression of dio1 and dio2 | ||||||
| MOS | TPA | Stimulator | Jang and Ji (2015). | 1. Upregulation of cyp19a, cyp19b, StAR, and cyp17 mRNAs in a concentration-dependent manner | ||
| 2. erα, vtg1, vtg2, cyp11a, hsd3b. trα, dio2, ahr remained unaltered | ||||||
| TRF | Stimulator | Zhu et al., (2015) | ||||
| KTC | Inhibitor | In vitro reporter gene assay: IC50 = 4.2 µM (medaka arβ antagonist assay) (Onishi et al., 2021) |
Zhang et al. (2008a), Onishi et al. (2021) |
1. Reduced fecundity | ||
| 2. Anal fin papillae increased in males not in females | ||||||
| 3. Liver VTG decreased in both sexes | ||||||
| 4. Gene expression | ||||||
| PCZ | Inhibitor | Zhang et al. (2008a), Schiller et al. (2014), Flynn et al. (2017), Kang et al. (2019), Onishi et al. (2021) | A: Brain: | |||
| (i) Male: | ||||||
| 1. Downregulation of cyp19b | ||||||
| (ii) Female: | ||||||
| 1. Downregulation of gnrhR2 and gnrhR3 | ||||||
| B: Liver: | ||||||
| (i) Male | ||||||
| 1. Upregulation of esr1 and arα mRNAs | ||||||
| 2. Downregulation of chgH and chgHm | ||||||
| (ii) Female | ||||||
| 1. Upregulation of arα | ||||||
| 2. Downregulation of esr1, vtgI, vtg2, cghL, chgH, chgHm | ||||||
| C: Gonad: | ||||||
| (i) Testis: | ||||||
| 1. Upregulation of esr2a, lhr, cyp19a | ||||||
| (ii) Ovary: | ||||||
| 1. Downregulation of esr2, arα, lhr | ||||||
| 2. Downregulation of StAR, cyp11a, cyp11b | ||||||
The apical endpoints of the reference chemicals related to EATS pathways in Japanese medaka.
2.2 Genes sensitive to EDCs within the EATS modalities of Japanese medaka
Within the EATS modalities, most of the EDCs function via the hormone-responsive element of a target gene by binding to the ligands of nuclear receptors (NRs), including ESRs (esr1, esr2a, and esr2b), ARs (arα and arβ), or TRs (trα and trβ). The effects of EDCs on NRs have been studied in Japanese medaka (Myosho et al., 2022; Tohyama et al., 2015). The expression of estrogen-responsive genes is known to be induced or suppressed by estrogen via ESRs with the estrogen-responsive elements (EREs) of responsive genes. Specifically, VTGs and CHGs encode complex precursor proteins in the egg yolk and eggshell, respectively, and are synthesized in the liver. EDCs with estrogenic potential induced the expression of VTG and CHG in juvenile and mature male fish, respectively, in which the expression levels of vtg and chg are typically low. Therefore, to evaluate the estrogenic potential of EDCs in Japanese medaka, vtgs (vtg1 and vtg2) and chg (chgL, chgH, and chgHm) can be used as markers.
The androgenic effects of EDCs are mediated via direct binding to ARs (arα and arβ) with distinctive binding properties or transactivation activity (Onishi et al., 2021; Kawashima et al., 2022). Molecular effects of AEDs could be identified from secondary sex characteristics (anal fin papillae of males) or indirectly by analysis of the induction/suppression of VTG (vtg1 and vtg2), LH, FSH, aromatase, ESRs, or T-hormone levels (Scholz and Mayers, 2008). The formation of papillary processes in the anal fin of Japanese medaka (males) is augmented by the bone morphogenic protein (bmp7) and lymphoid enhancer-binding factor (lef1), along with arα and arβ which can be used as markers for AEDs during evaluation (Ogino et al., 2014).
EDCs having TH-disrupting potential inhibit or accelerate TH-dependent processes, either directly or indirectly, including TH-dependent gene expression. The HPT axis is highly conserved among vertebrates, and the TH and receptors (trα and trβ) play crucial roles in the regulation of development, growth, and energy metabolism. A number of high-profile environmental pollutants adversely affect the TH system of Japanese medaka, including development, visual performance, malformation of the swim bladder, and TH-dependent gene (tshβ, trα, trβ, dio1, and dio2) expression (Godfrey et al., 2019; Dang et al., 2021; Horie et al., 2022c). Therefore, to evaluate the thyroid-disrupting potential of EDCs, in Japanese medaka, these genes (tshβ, trα, trβ, dio1, and dio2) can be used as markers for TEDs.
Moreover, within the estrogen–androgen–steroidogenesis (EAS) modalities, the steroid hormones, estrogen (E2) and androgen (A), are derived from cholesterol and secreted from the gonads (testis or ovary). The production, conversion, and breakdown of E2 and A in the endocrine glands and target tissues are carefully controlled by a range of steroidogenic enzymes (steroidogenesis), many of which belong to the cytochrome P450 family (CYP11, CYP17, and CYP19). Many EDCs have the abilities to disrupt the synthesis and function of steroidogenic enzymes, resulting in inappropriate concentrations of E2 or A, which impacts the reproduction, development, growth, and metabolism of fish (Japanese medaka). The enzyme aromatase (CYP19) converts testosterone/androgen (A) into estradiol/estrogen (E) and controls the fine balance between these two potent sex steroids. Therefore, the genes that show potential to regulate steroidogenesis in Japanese medaka are used as markers during EDC evaluation.
3 Results
Depending on the ED effects, we sorted 205 articles (Table 2) consisting of 128 chemicals (1.6 articles/chemicals or the approximate ratio is 8 articles:5 chemicals) that showed potential effects on Japanese medaka. Furthermore, based on the apical endpoints selected from 14 reference chemicals (Table 1) and after reviewing 165 articles, we identified 83 chemicals that target EATS pathways of Japanese medaka (Tables 3–10), and due to the lack of sufficient information, 45 chemicals reviewed from 60 articles remained unclassified (Table 11). Moreover, among the 83 chemicals that target EATS pathways, 43 chemicals were recommended for Tier 2 tests, and 13 chemicals show enough potential to be considered EDCs without any further tier-based studies (Tables 3–10). The rest of the EATS chemicals need further studies on Tier 1 screening. Moreover, with regard to the apical endpoints, the EATS chemicals were further classified as agonists and antagonists of EEDs, AEDs, and TEDs, and stimulators or inhibitors of steroidogenesis (Figure 2; Tables 3–10).
TABLE 2
Literature reports sorted for the evaluation of the effects of EDCs on Japanese medaka (Oryzias latipes).
Only the names of the authors are listed in the first column of the table. The cells filled in colors (red = EED; blue = AED; green = TED; yellow = MOS; black = unidentified pathways) represent the specific endocrine pathways/organs disrupted by EDCs. EDCs, endocrine-disrupting chemicals; EED, estrogen endocrine disruptor; AED, androgen endocrine disruptor; TED, thyroid endocrine disruptor; MOS, modulators of steroidogenesis.
TABLE 3
| Serial number | Name of the chemical | Nature (source) | Significant endpoints | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | AR-1260 | Polychlorinated biphenyl (persistent organic pollutant) | 1. Induced vtg, chgL, and chgHm mRNAs in the liver of males | Yum et al. (2010) | Tier 2 (thyroid-dependent mechanisms) |
| 2 | AZM | Organophosphate pesticide [agricultural] | 1. Female-biased sex ratio | Teather et al. (2005) | Tier 1 (thyroid-dependent mechanisms) |
| 3 | BZP | Antimicrobial agent [personalcare product] | 1. Serum VTG enhanced in males | Yamamoto et al. (2007) | Tier 2 (thyroid-dependent mechanisms) |
| 2. vtg1, vrg2, chgL, chgH, chgHm, esr1, and cyp1a genes upregulated in the liver of males | |||||
| 4 | BMT | Synthetic glucocorticoid (pharmaceuticals) | 1. Both vtg1 and vtg2 mRNAs were induced in the liver of male fish | Su et al. (2023) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Ova found in the testis | |||||
| 3. Serum T reduced, while E2 induced | |||||
| 5 | BPA | Raw material for polycarbonate plastic [industrial] | 1. Anal fin papillae in males disappeared | Shioda and Wakabayashi (2000) | (Thyroid-dependent mechanisms) |
| 2. Testis–ova | Yakota et al. (2000) | ||||
| 3. VTG mRNAs (vtg1 and vtg2) and protein in the liver of males increased | Metcalfe et al. (2001) | ||||
| 4. Upregulation of chgL and chgH in the liver of male fish | Tabata et al. (2001) | ||||
| 5. Expression of esr1, esr2a, and esr2b genes remained unchanged | Kang et al. (2002b) Kashiwada et al. (2002) Lee et al. (2002) Zeng et al. (2005) Kamata et al. (2011) Lee et al. (2012) Schiller et al. (2014) Bhandari et al. (2015, 2020)Tohyama et al. (2015)Inagaki et al. (2016)Li et al. (2016); Li et al. (2017)Horie et al. (2019)Ishibashi et al. (2020)Thayil et al. (2020)Kawashima et al. (2022) | ||||
| 6 | CFR | Antibiotic (pharmaceutical) | 1. Plasma E2 level increased in females | Kim et al. (2017) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Sex-specific alteration in the gene expression pattern of the HPG axis | |||||
| 7 | CLT | Organochlorine pesticide (fungicide) [agricultural] | 1. Female-biased sex ratio | Teather et al. (2005) | Tier 1 (gene analysis on EATS pathways} |
| 8 | CTC | Antimicrobial agent [pharmaceutical] | 1. Enhancement of serum E2 and liver VTG content in male fish | Kim et al. (2007) Ji et al. (2010) | Tier 2 (thyroid-dependent mechanisms) |
| 9 | p,p′-DDE | DDT metabolite [agricultural] | 1. Increased HSI | Zhang and Hu (2008) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Development of intersex | Horie et al. (2022a) | ||||
| 3. Upregulation of vtg1, vtg2, chgH, chgL, and esr1 genes in the liver of male fish | |||||
| 10 | o,p′-DDT | Organochlorine pesticide [agricultural] | 1. Female-biased sex ratio in fish | Edmunds et al. (2000) | (Thyroid-dependent mechanisms) |
| 2. Ova–testis | Cheek et al. (2001) | ||||
| 3. chgH, chgL, chgHm, vtg1, vtg2, and esr1 mRNAs induced in the liver of male | Kuhl and Brouwer (2006) Uchida et al. (2010) | ||||
| 11 | DES | Nonsteroidal estrogen [pharmaceutical] | 1. vtg1 mRNA was upregulated in males | Zeng et al. (2005) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Sex-reversed males laid eggs | Lei et al. (2016) | ||||
| 12 | EDS | Organochlorine pesticide [agricultural] | 1. Female-biased sex-ratio | Teather et al. (2005); Lee et al. (2013) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Serum VTG induced in male fish | |||||
| 13 | EQ | Metabolite of the soy isoflavone daidzein [natural product] | 1. Testis–ova formation in males | Kiparissis et al. (2003a) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Intersex | Wang et al. (2016) | ||||
| 3.11-KT in the plasma reduced | |||||
| 14 | E3 | Natural estrogen [pharmaceutical] | 1. Testis–ova | Metcalfe et al. (2001) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Induced vtg1 mRNA in males | Zeng et al. (2005) | ||||
| 15 | E1 | Natural estrogen [pharmaceutical] | 1. Female-biased sex ratio | Metcalfe et al. (2001) | (Thyroid-dependent mechanisms) |
| 2. Liver VTG induced in both sexes | Nakamura et al. (2014a) Kawashima et al. (2022) | ||||
| 16 | 4-MBC | Camphor derivative [personal care product] | 1. Enhancement in the serum VTG in both sexes | Inui et al. (2003) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Upregulation of vtg1, vtg2, chgL, and chgH, and esr1 mRNAs in the liver of males | Liang et al. (2020) | ||||
| 3. Decrease in 11-KT in the plasma of males and enhancement of E2 in females | |||||
| 4. Inhibition of spermatogenesis in the testis | |||||
| 17 | MPB | Antimicrobial agent [personal care product] | 1. Plasma VTG content increased in males | Yamomoto et al. (2011) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Upregulation of vtg2, chgL, chgH, chgHm, and esr1 in the liver of males | Kawashima et al. (2022) | ||||
| 18 | 4-NP | Alkylphenol [industrial] | 1. Males developed testis–ova with the sex ratio skewed toward female | Gray and Metcalfe, (1997); Nimrod and Benson, (1998); Shioda and Wakabayashi, (2000); Tabata et al. (2001); Yokota et al. (2001); Islinger et al. (2002); Kashiwada et al. (2002); Lee et al. (2002); Kang et al. (2003); Seki et al. (2003a); Nozaka et al. (2004); Kobayashi et al. (2005); Zeng et al. (2005), Balch and Metcalfe, (2006); Ishibashi et al. (2006); Jin et al. (2011a); Lee et al. (2012); Miyagawa et al. (2014); Tohyama et al. (2015); Watanabe et al. (2017); Ishibashi et al. (2020); Horie et al. (2021); Kawashima et al. (2022) | (Thyroid-dependent mechanisms) |
| 2. HSI in adult males increased | |||||
| 3. Serum VTG in males and hepatic VTG in both sexes increased | |||||
| 4. Female-like anal fins in some males | |||||
| 19 | OMC | Organic UV-B filter (PCP) [personal care product] | 1. Enhancement of plasma VTG in males | Inui et al. (2003) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Upregulation of vtg1, vtg2, chgL, chgH, and esr1 mRNAs in males | |||||
| 20 | 4-OP | De-ethoxylated alkylphenol [industrial] | 1. Female-biased sex ratio | Knorr and Braunbeck (2002) | Tier 1 (thyroid-dependent mechanisms) |
| 2. Some F1 males developed testis–ova | |||||
| 21 | PCPL | Insecticide [agricultural] | 1. Nonlinear enhancement in the plasma VTG levels in males and a concentration-dependent decrease in plasma VTG levels in females | Zha et al. (2006) | Tier 1 (thyroid-dependent mechanisms) |
| 2. Testis–ova formation in males and a degenerative ovary in females | |||||
| 22 | PPB | Personal care product [personal care product] | 1. Enhancement in the plasma VTG content in males | Inui et al. (2003) | Tier 2 (interruption in swim bladder inflation needs further studies in thyroid-dependent mechanisms) |
| 2. Upregulation of vtg1, vtg2, chgL, chgH, and esr1 in the liver of male fish | Gonzalez-Doncel et al. (2014a) Kawashima et al. (2022) | ||||
| 23 | 4t-OP | Alkylphenol [industrial] | 1. Sex ratio skewed toward females | Gray et al. (1999a) | Effect on swim bladder inflation needs further study on thyroid-dependent mechanisms |
| 2. Testis–ova observed in male fish | Gray et al. (1999b) | ||||
| 3. Liver VTG increased in both sexes | Gronen et al. (1999) | ||||
| 4. Inhibition of spermatogenesis | Seki et al. (2003a) | ||||
| 5. HSI in adult males increased | Nozaka et al. (2004) | ||||
| 6. Basophilia in the male liver | Flynn et al. (2017) Horie et al. (2022a) Kawashima et al. (2022) | ||||
| 24 | 4t-PP | Alkylphenol [industrial] | 1. The appearance of secondary sexual features was reduced in males | Seki et al. (2003b) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Testis–ova in the gonad of males | Yokota et al. (2005) | ||||
| 3. Hepatic VTG enhanced in both sexes | Onishi et al., 2021 | ||||
| 4. HSI increased in males | Kawashima et al. (2022) | ||||
| 25 | TBCO | Brominated flame retardant [industrial] | 1. Upregulation of chgHm in the liver of males | Saunders et al. (2015) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Upregulation of chgH, vtg2, and esr1 in the liver of females | Sun et al. (2016c) | ||||
| 3. Downregulation of esr1, esr2a, and arα in both the testis and ovary | Devoy et al. (2023) |
Potential EED agonist chemicals identified from the literature search.
TABLE 4
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | ATZ | Herbicide [agricultural] | 1. cyp19a mRNA upregulated in the brain | Zhang et al. (2008d) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Downregulation of esr1 mRNA in the testis | Richter et al. (2016) | ||||
| 3. VTG in the liver of females reduced | |||||
| 2 | MET | Drug (pharmaceutical) | 1. Intersex observed in females | Lee et al. (2019a) | Tier 2 (studies related to thyroid-related gene expression are necessary) |
| 2. vtg1 declined in males | |||||
| 3. Thyroid histology remained unchanged | |||||
| 3 | PFOA | Fluorinated organic compounds (wastewater effluent) | 1. Reduced fecundity | Lee et al. (2017a) | Tier 2 (studies related to thyroid-related gene expression are necessary) |
| 2. Increase in the serum VTG content in F2 males | |||||
| 3. Male-biased sex ratio with no change in intersex either in F1 or F2 | |||||
| 4 | TPhP | Flame retardant/plasticizer [industrial] | 4. Larval exposure reduced ovarian development in females | Li et al. (2019) | Tier 2 (thyroid-dependent mechanisms) |
| 5. Plasma T enhanced in females | Kawashima et al. (2022) | ||||
| 6. Hepatic VTG in females reduced |
Potential EED antagonist chemicals identified by the literature search.
TABLE 5
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | 11-OA | Glucocorticoid metabolite [pharmaceutical] | 1. Male-biased sex ratio | Grillitsch et al. (2010) | Tier 1 (gene expression analysis and thyroid-dependent mechanisms) |
| 2 | BF | Pyrethroid insecticide [agricultural] | 1. Induced masculinization in the anal fin papillae | Bertotto et al. (2019) | Tier 2 (gene expression analysis related to EATS pathways and thyroid-dependent mechanisms) |
| 2. Male-biased sex ratio | |||||
| 3 | CFD | Antibiotic (pharmaceutical) | 1. Plasma E2 was decreased in males and enhanced in females | Kim et al. (2017) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Downregulation of cyp19a in the testis and upregulation of cyp19a in the ovary | |||||
| 3. Sex-specific alteration in the gene expression of the HPG axis | |||||
| 4 | DHT | Metabolite of testosterone [pharmaceutical] | 1. Anal fin papillae increased in both sexes | Spirhanzlova et al. (2020); Onishi et al. (2021) | Tier 1 (gene expression analysis related to EATS pathways and thyroid-dependent mechanisms) |
| 2. Sex ratio skewed toward males | |||||
| 5 | GEN | Isoflavone [natural Product] | 1. Masculinization features in the secondary sex characteristics of XX females | Hishida and Kawamoto (1970); Kiparissis et al. (2003a)Schiller et al. (2013, 2014) | Tier 2 (downregulation of dio2 indicated more studies on thyroid-dependent mechanisms are necessary) |
| 6 | LNG | Second-generation progestin (pharmaceutical) | 1. Liver VTG downregulated in females | Onishi et al. (2021); Pandelides et al. (2021); Watanabe et al. (2023) | Tier 2 (effects on the swim bladder suggest more studies on thyroid-dependent mechanisms are necessary) |
| 2. Masculinization of the anal fin papillae in females | |||||
| 3. Ovotestis in females | |||||
| 7 | MT | Synthetic androgen [pharmaceutical] | 1. Sex reversal of XX females | Papoulias et al. (2000); Chikae et al. (2004); Nozaka et al. (2004); Seki et al. (2004); Kang et al. (2008); Ogino et al. (2014); Myosho et al. (2019); Onishi et al. (2021) | Thyroid-dependent mechanisms |
| 2. Serum VTG decreased in females | |||||
| 3. Upregulation of gsdf mRNA in XX fish | |||||
| 8 | P4 | Female hormone (steroid) [pharmaceutical] | 1. Females developed papillae on the anal fin rays | Onishi et al. (2021) | Tier 2 (thyroid-dependent mechanisms) |
| 9 | SPR | Synthetic aldosterone receptor agonist [pharmaceutical] | 1. Anal fin papillae increased in both sexes | LaLone et al. (2013) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Hepatic vtg reduced in female fish | |||||
| 10 | T | Male hormone (steroid) [pharmaceutical] | 1. Intersex gonad | Koger et al. (2000) | Tier 1 (thyroid-dependent mechanisms) |
Potential androgen endocrine-disrupting agonist chemicals identified from the literature search.
TABLE 6
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | CPA | Male contraceptive [pharmaceutical] | 1. Testis–ova observed in male fish | Kiparissis et al. (2003b) | Tier 2 (gene expression analysis of EATS pathways and thyroid-related mechanisms) |
| 2. No difference in the phenotypic sex ratio | |||||
| 3. Inhibition of spermatogenesis | |||||
| 2 | DZ | Organophosphate insecticide [agricultural] | 1. Number of anal fin papillae in F1 male fish reduced | Hamm and Hinton, (2000); Flynn et al. (2018); Kawashima et al. (2022) | Tier 2 (effects on the swim bladder suggests more studies on thyroid-dependent mechanisms are necessary) |
| 3 | 2-EHHB | Antimicrobial agent (personal care product) | 1. Hepatic vtg1 upregulated in F1 males and downregulated in F2 males | Matten et al. (2023) | Tier 2 (gene expression analysis of EATS pathways and thyroid-related mechanisms) |
| 2. Anal fin papillae in F2 males reduced | |||||
| 3. Delay in reproductive tract development in F1 males | |||||
| 4. Eosinophilia observed in renal ducts (kidney) of females | |||||
| 4 | FNT | Organophosphate pesticide [agricultural] | 1. Number of papillary processes decreased in XY medaka | Horie et al. (2017; 2022a) | Tier 1 (thyroid-related mechanisms) |
| 5 | KC-400 | Polychlorinated biphenyl (industrial) | 1. Downregulation of chgL, chgHm, and arα, in both males and females | Nakayama et al. (2011) | Tier 1 (thyroid-related mechanisms) |
| 2. Downregulation of vtg1 in males and upregulation in females | |||||
| 6 | LD-BP | Structural analog of bisphenol A [industrial] | 1. Liver VTG in males and females increased | Li et al. (2016, 2017) | Tier 1 (thyroid-related mechanisms) |
| 2. Aggregation and hyperplasia of interstitial cells occurred in the testis, while atretic follicles, with interstitial cell fibrosis, occurred in the ovary | |||||
| 7 | PCB 126 | Coplanar PCB (persistent organic pollutants) | 1. Downregulation of chgL, chgHm, and arα, in both males and females | Nakayama et al. (2011) | Tier 1 (thyroid-related mechanisms) |
| 2. Downregulation of vtg1 in males and upregulation of vtg1 in females | |||||
| 8 | TCrP | Organophosphate flame retardant [industrial] | 3. Suppression of 11-KT and T levels and enhanced E2 level in the plasma of male fish | Chen et al. (2022) | Tier 1 (thyroid-related mechanisms) |
| 4. Dilated the efferent duct of the testis | |||||
| 5. Intersex development |
Potential AED antagonist chemicals identified by the literature search.
TABLE 7
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | MTC | Herbicide [agricultural] | 1. Upregulation of the expression of trα, trβ, and dio2 mRNAs in females | Jin et al. (2011b) | Tier 1 (thyroid histopathology and EATS-dependent mechanisms) |
Potential TED agonist chemicals identified from the literature search.
TABLE 8
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | ATBC | Non-phthalate plasticizer [industrial] | 1. Disruption of swim bladder inflation | Horie et al. (2022b); Horie et al. (2023b) | Tier 1 (downregulation of vtg1 and vtg2 mRNAs in the liver of XX fish indicated more studies needed on EAS pathways are required) |
| 2. Downregulation of trα, trβ, and dio2 | |||||
| 2 | DEHS | Plasticizer [industrial] | 1. Downregulation of dio2 | Horie et al. (2022c) | Tier 1 (more studies needed on EATS pathways) |
| 3 | DIC | NSAID [pharmaceutical] | 1. Swim bladder inflation inhibition in larvae | Hong et al. (2007); Lee et al. (2011): Yokata et al. (2017), Yokata et al. (2018); Pandelides et al. (2021) | Tier 2 (more studies needed on EATS pathways) |
| 4 | EHMC | Organic ultraviolet UV-B filter [personal care products] | 1. T3 and T4 concentrations decreased | Lee et al. (2019b) | Tier 2 (studies other than those based on EATS pathways are necessary) |
| 2. Downregulation of dio2 | |||||
| 3. Upregulation of trh | |||||
| 5 | PFBA | Halogenated chemical [industrial] | 1. No swim bladder inflation | Godfrey et al. (2019); Horie et al. (2022d) | Tier 1 (more studies needed on EAS pathways) |
| 6 | (PFOS/PFOSA) | Halogenated compound [industrial] | 1. Hyperplasia, hypertrophy, and colloidal depletion in thyroid follicles | Ji et al. (2008) | Tier 1 (more studies needed on EATS pathways) |
| Kang et al. (2019) | |||||
| 7 | PTU | Anti-thyroid medicine [pharmaceutical] | 1. Modulation of swim bladder inflation | Horie et al. (2023a) | Tier 1 (studies related to EAS pathways) |
| 8 | SPC | Anti-thyroid chemical [industrial] | 1. Downregulation of trα and trβ genes | Lee et al. (2014) | Tier 1 (studies related to EAS pathways) |
| 2. Upregulation of dio2 | |||||
| 3. Decrease in T4 levels but T3 remained unaltered | |||||
| 4. Fecundity decreased with the increase in temperature | |||||
| 9 | TU | Anti-thyroid chemical [industrial] | 5. Decreased thyroid hormone levels in adult fish and fertilized eggs | Tagawa and Hirano (1991) | Tier 1 (although anti-thyroid effects were established, EAS-mediated pathways need to be investigated) |
| 6. No effect on the length and weight of the larvae | |||||
| 10 | RND | Herbicide (commercial formulation of glyphosate) [agricultural] | 1. Uninflated swim bladder | Smith et al. (2019) | Tier 1 (studies related to thyroid histophysiology and thyroid-dependent gene expression) |
| 11 | TDCPP | Halogen-containing organophosphorus compound [industrial] | 1. Females failed to inflate the swim bladder | Godfrey et al. (2019) | Tier 1 (upregulation of vtg1 and vrg2 mRNAs indicates further studies on EAS mechanisms are necessary) |
| Horie et al. (2022a); Horie et al. (2022d) |
Potential TED antagonist chemicals identified from the literature search.
TABLE 9
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | OCL | Organic UV filter (PCP) [personal care product] | 1. Upregulation of fshβ, lhβ, fshr, lhr, ar, esr1, esr2a, StAR, hsd3β, cyp17α, and cyp19β mRNAs in the HPG axis | Yan et al. (2020) | Tier 2 (thyroid-dependent mechanisms) |
| 2. E2 and 11-KT increased in plasma | |||||
| 3. Upregulation of vtg in the liver of males and females | |||||
| 2 | FPN | Phenylpyrazole insecticide [agricultural] | 1. Upregulation of StAR, cyp17a, and cyp19b in males | Sun et al. (2014) | Tier 2 (thyroid-dependent studies) |
| 2. Upregulation of both vtg1 and vtg2 mRNAs in both sexes | Wagner et al. (2017) | ||||
| 3. No alteration occurred in esr1, esr2a, and arα in both sexes | |||||
| 3 | RCT | β-adrenergic agonist drug [pharmaceutical] | 4. Upregulation of cyp19a and cyp19b mRNAs in females | Sun et al. (2016a) | Tier 2 (thyroid-dependent mechanisms) |
| 5. Upregulation of vtg1, vtg2, esr1, and esr2 mRNAs in females | |||||
| 4 | TRI | Pharmaceuticals [pharmaceutical] | 1. Upregulation of StAR, 3β-hsd, 20β-hsd, cyp11a, cyp11b, cyp17a, cyp17b, and cyp19a in males | Sun et al. (2014) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Upregulation of vtg1 and vtg2 in males and downregulation of vtg1 and vtg2 in females | |||||
| 3. Upregulation of esr1 and arα in males |
Potential steroidogenesis stimulating EDCs identified from the literature search.
TABLE 10
| Serial number | Name of the chemical | Nature (source) | Significant endpoint | Reference | Recommendation |
|---|---|---|---|---|---|
| 1 | BP | UV filters used in cosmetics [personal care product] | 1. Liver VTG in both male and females increased by BP2 | Coronado et al. (2008) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Enhanced T concentration in the serum of male fish by BP3 | Kim et al. (2014); Kawashima et al. (2022) | ||||
| 3. Upregulation of vtg1 and vtg2 mRNAs, and the VTG protein in the liver of male fish by BP3 | |||||
| 4. Downregulation of gonadal StAR, cyp17, hsd3b, hsd17b3, and cyp19a by BP3 | |||||
| 2 | FAD | Nonsteroidal aromatase inhibitor [pharmaceutical] | 1. Aromatase enzyme activity reduced | Suzuki et al. (2004) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Upregulation of cyp19a in the ovary | Kuhl and Brouwer, (2006) | ||||
| 3. Downregulation of esr1 and chgL in the liver of females | Thresher et al. (2011) Park et al. (2008) Zhang et al. (2008b) | ||||
| 3 | LET | Nonsteroidal triazole [pharmaceutical] | 1. Male-biased sex ratio | Sun et al. (2007b, 2009, 2011a) | Tier 2 (thyroid-dependent mechanisms) |
| 2. Downregulation of esr1, vtg1, and vtg2 in the liver of males | Liao et al. (2014) | ||||
| 3. Serum VTG levels remained unaltered in males and decreased in females | |||||
| 4. Upregulation of StAR, cyp11a, cyp11b, cyp17a, cyp17b, and esr2 and downregulation of cyp19 b and arα in the ovary | |||||
| 5. Upregulation of cyp11a and cyp11b and no alteration in cyp17a, cyp17b, cyp19a, and cyp19b mRNAs in the testis | |||||
| 4 | LNR | Herbicide [agricultural] | 1. Downregulation of 3β-hsd and cyp11b | Schiller et al. (2014) | Tier 1 (gene expression analysis related to EATS pathways and thyroid-dependent mechanisms) |
| 2. E2 or T-induced expression of chgH was downregulated | Spirhanzlova et al. (2017) | ||||
| 5 | PRN | Herbicide [agricultural] | 1. Downregulation of cyp11b, 3β-hsd, gnrhr2, and cyp19a1b | Schiller et al. (2014) | Tier 1 (gene expression analysis related to EATS pathways and thyroid-dependent mechanisms) |
| 6 | TPT-Cl | Organotin compound [industrial] | 1. Downregulation of 17β-hsd1 and cyp19a in the ovary | Zhang et al. (2008e) | Tier 1 (studies on EATS pathways and thyroid-dependent mechanisms) |
| 2. Upregulation of cyp1a and cyp2a1 | Horie et al. (2022a) | ||||
| 3. ugt2a3 and 17β-hsd1 in the liver of both sexes | |||||
| 4. No change in gsdf mRNA expression in both XX and XY embryos |
Potential steroidogenesis inhibitory EDCs identified by the literature search.
TABLE 11
| Serial number | Name of the chemical | Nature (source) | Reference | Reason |
|---|---|---|---|---|
| 1 | ACT | NSAID (pharmaceutical) | Kim et al. (2012) | Limited data (nonlinear induction of hepatic VTG in males was due to stress) |
| 2 | AMT | Herbicide [agricultural] | Horie et al. (2022a) | Insufficient data |
| 3 | BZT-UV | UV stabilizer; persistent organic pollutants (POPs) [personal care product] | Fujita et al. (2022) | Due to stress |
| 4 | BKC | Quaternary ammonium compound [personal care product] | Kim et al. (2020) | Insufficient data (enhancement of vtg1 in the whole body was probably due to stress) |
| 5 | i-BP | Antimicrobial [personal care product) | Yamamoto et al. (2007) | Insufficient data (estrogenic potential) |
| 6 | n-BP | Antimicrobial [personal care product) | Yamamoto et al. (2007) | Insufficient data (estrogenic potential) |
| 7 | Cd | Metal [inorganic] | Tilton et al. (2003), Hirako et al. (2017) | Insufficient data (anti-androgenic effects were probably mediated through stress) |
| 8 | ClxBPA | Chlorinated product of BPA | Tabata et al. (2004) | Limited data (the compound showed estrogenic potential with regard to serum VTG in male fish) |
| 9 | CMP | Biocide [personal care product] | Flynn et al. (2017); Onishi et al. (2021) | Inconsistent alteration of liver VTG in both sexes indicate the estrogenic potential of the compound |
| 10 | CYN | Herbicide [agricultural] | Kawashima et al. (2022) | Effects are not mediated through EATS pathways |
| 11 | CHDM | Plasticizer [industrial] | Jang and Ji, (2015) | Effects are not mediated through EATS pathways |
| 12 | DBP | Plasticizer [industrial] | Nozaka et al. (2004) | VTG in male fish remained unchanged |
| 13 | DEHP | Plasticizer [industrial] | Metcalfe et al. (2001) | Effects are not mediated through EATS pathways |
| 14 | DIBP | Plasticizer [industrial] | Kawashima et al. (2022) | Limited AED features (hepatic VTG reduced in females) |
| 15 | END | Organochlorine pesticide [agricultural] | Horie et al. (2022a) | Limited information (not related to EATS-mediated pathways) |
| 16 | FNC | Insecticide [agricultural] | Spirhanzlova et al. (2017) | Effects not related to EATS pathways |
| 17 | FV | Pyrethroid insecticide [agricultural] | Kawashima et al. (2022) | No effects on estrogen-dependent mechanisms |
| 18 | FLX | Antidepressant [pharmaceutical] | Foran et al. (2004) | Mostly due to toxicity and not mediated through EATS pathways |
| 19 | FLR | Herbicide (Agricultural) | Jin et al. (2020) | Effects are mediated through oxidative stress |
| 20 | GLP | Herbicide [agricultural] | Smith et al. (2019) | Effects are mediated through oxidative stress |
| 21 | GO | Nanocarbon [inorganic] | Dasmahapatra et al. (2020a, b) | Effects are not mediated through EATS pathways |
| Myla et al. (2021a); Myla et al. (2021b) | ||||
| Asala et al. (2021); Asala et al. (2022) | ||||
| Dasmahapatra and Tchounwou (2022a); Dasmahapatra and Tchounwou (2022b); Dasmahapatra and Tchounwou (2023a); Dasmahapatra and Tchounwou (2023b) | ||||
| 22 | IBP | Nonsteroidal anti-inflammatory drug [pharmaceutical] | Flippin et al. (2007); Han et al. (2010) | VTG induction in male fish serum is probably due to stress |
| 23 | LIN | Antibiotic (pharmaceutical) | Kim et al. (2012) | Insufficient data (insignificant increase in hepatic VTG in male fish) |
| 24 | MTZ | Goitrogen [pharmaceutical] | Godfrey et al. (2019) | Insufficient data (vtg gene expression upregulated in males) |
| 25 | MXC | Organochlorine pesticide [agricultural] | Nimrod and Benson (1998) | Insufficient data |
| Zeng et al. (2005) | ||||
| 26 | MCB | Fungicide [agricultural] | Lin et al. (2014) | Induced cyp3a enzyme activities |
| 27 | 1NT | Insecticide [agricultural] | Kawashima et al. (2022) | Limited data (hepatic VTG enhanced in females) |
| 28 | NPX | NSAID [pharmaceutical] | Kwak et al. (2018) | Although transcription of vtg1, erβ2, and cyp17 genes significantly increased, data are still limited for consideration as EEDs |
| 29 | NDEA | Carcinogen [industrial] | Nair et al. (2017) | Limited data (sex-specific reduction in germ cells occurred only in the ovary) |
| 30 | OYZ | Herbicide [agricultural] | Hall et al. (2005, 2007) | Insufficient data (induction of choriogenin in liver and abnormal gonad histology) |
| 31 | OXF | Herbicide [agricultural] | Powe et al. (2018) | Toxicological effects |
| 32 | OTC | Antibiotic [pharmaceutical] | Ji et al. (2010, 2012) | Insufficient data |
| 33 | PDM | Herbicide [agricultural] | Kawashima et al. (2022) | Insufficient data (hepatic VTG enhanced in males) |
| 34 | PHN | Aromatic hydrocarbon [burning of fuels] | Horng et al. (2010) | No significant EATS-mediated effects |
| 35 | PHT | Epileptic drug [pharmaceutical] | Kawashima et al. (2022) | Insufficient data |
| 36 | RLX | SERM [pharmaceutical] | Onishi et al. (2021) | Insufficient data (liver VTG enhanced in males and reduced in females) |
| 37 | SFT | Veterinary pharmaceutical [pharmaceutical] | Ji et al. (2010) | Limited data (enhancement of the serum E2 level in male fish) |
| 38 | SRF | Herbicide [agricultural] | Hall et al. (2005) | Limited data (only chg in males enhanced) |
| 39 | BDE-47 | Flame retardants [industrial] | Gonzalez-Doncel et al. (2014b); Gonzalez-Doncel et al. (2016) | Lack of ED effects related to EATS pathways |
| Gonzalez-Doncel et al. (2017) | ||||
| Beltran et al. (2022) | ||||
| 40 | TRA | Metabolite of TRB (agricultural) | Robinson et al. (2017) | Lack of adverse effects on fecundity |
| 41 | TRF | Fungicide (agricultural) | Lin et al. (2014) | Induced cyp1a and cyp3a activities in the liver |
| 42 | TRD | Fungicide [agricultural] | Lin et al. (2014); Chu et al. (2016); Liu et al. (2018) | Limited information (upregulation of vtg2 and cyp3a40 and downregulation of cyp3a38, vtg1, esr1, and cyp1a in the liver of females) |
| 43 | TBT | Biocide [agricultural] | Nozaka et al. (2004) | Limited data (inhibition of brain aromatase) |
| Kuhl and Brouwer, (2006) | ||||
| Hano et al. (2007) | ||||
| Zhang et al. (2008d) | ||||
| Horie et al. (2018, 2022a) | ||||
| 44 | TCS | Antimicrobial [industrial] | Foran et al. (2000) | Inconsistent data (hepatic VTG increased in males) |
| Ishibashi et al. (2004) | ||||
| Mihaich et al. (2019) | ||||
| Song et al. (2020) | ||||
| Kawashima et al. (2022) | ||||
| 45 | Nano zinc oxide (nZnO)/zinc sulfate (ZnSO4) | Metal [inorganic] | Paul et al. (2021) | Toxic effects (reduced follicular growth and maturation in the ovary) |
Potential EDCs with unidentified EATS pathways.
3.1 EEDs
For the identification and classification of EEDs from the searched chemicals, we considered three chemicals as references, E2 and EE2 as agonists, and TAM as antagonists (Table 1). Based on these reference chemicals, several endpoints, such as the female-biased sex ratio, induction of serum VTG (protein) in male fish, alteration of the secondary sex characteristics (anal fin papillae in the male fish), and up- or downregulation of vtg and chg genes/mRNAs in the liver of male fish, as well as the estrogen receptors (ERs) of the HPG axis in both sexes, were considered (Table 1). Using these strategies, we reviewed 108 articles, which is 52.68% of the searched articles, consisting of 25 chemicals as agonists and 4 chemicals as antagonists (Tables 3, 4). Adding three reference chemicals to the list, the number of EED agonists increased to 27 (21.09% of 128 chemicals) and antagonists to 5 (3.9% of 128 chemicals), altogether 32, which is 25% of the total (128 chemicals) chemicals searched by the literature survey. Alternatively, it appears that for every 100 EDCs, ∼21 of them are identified as EED agonists and ∼4 of them are identified as EED antagonists. Moreover, considering the 108 articles that studied EEDs, every EED chemical was studied in 3.375 articles (27 articles: 8 EEDs). Moreover, among EED agonists, other than two reference chemicals (E2 was reviewed in 37 articles and EE2 in 27 articles), 4-nonylphenol (4-NP; 23 articles), bisphenol A (BPA; 21 articles), and 4-tert-octylphenol (4-t-OP; 8 articles) are the most studied EED agonist chemicals in Japanese medaka (Table 3). Among others, o,p′-DDT (4 articles), 4t-PP (4 articles), E1 (3 articles), PPB (3 articles), and TBCO (3 articles) have drawn significant interest among investigators. The remaining 17 estrogen agonists were studied either twice (8 chemicals) or once (9 chemicals). For EED antagonists, the reference chemical TAM was studied in five articles, whereas ATZ was studied twice (Ritcher et al., 2016), MET once (Lee et al., 2019), and TPhP in two articles (Li et al., 2019; Kawashima et al., 2022). Moreover, 16 of the EEDs as agonists and 4 as antagonists were recommended for Tier 2 tests. Therefore, based on the literature search, we recommend that eight chemicals (E1, E2, EE2, BPA, o,p′-DDT, 4-NP, 4-t-OP, and TAM) showed enough potential to be considered EEDs in Japanese medaka and did not require any further Tier 2 tests for estrogen signaling mechanisms. Furthermore, except PPB (Gonzalez-Doncel et al., 2014a), 4t-OP (Gray et al., 1999b), and MET (Lee et al., 2019), in most of the EED chemicals, whether agonists or antagonists, the thyroid-related endpoints remained uninvestigated, even though the reference agonists (E2 and EE2) have the potential to inhibit swim bladder inflation (a thyroid-related endpoint) in a concentration-dependent manner in larvae if the embryos were exposed either to E2 or EE2 during development (Pandelides et al., 2021).
3.2 AEDs
For AEDs, four chemicals, 11-KT and TRB as agonists and FLU and TRB as antagonists, were considered references (Table 1). Based on these reference chemicals, the apical endpoints, such as masculinization of females (development of anal fin papillae), male-biased sex ratio, upregulation of gsdf mRNA in XX embryos, ovotestis, and downregulation of vtg1, vtg2, chgH, and chgHm gene transcripts in the liver of both male and female fish (Table 1), were mostly considered during the evaluation of AEDs. With these efforts, from 46 articles, which is 22.43% of the sorted articles (Table 2), we identified 10 chemicals as agonists (Table 5) and 8 chemicals as antagonists (Table 6). With the addition of four reference chemicals, the number of AEDs increased to 22 (∼9% agonists and ∼8% antagonists), which is 17.18% of the 128 chemicals screened through the literature search. Alternatively, for every 100 EDCs, ∼9 chemicals are identified as AED agonists and ∼8 chemicals are identified as AED antagonists. Moreover, with regard to 46 articles that studied 22 AEDs, it appears that one AED chemical was studied in 2.09 articles (approximately 2 articles:1 AED). Moreover, among the reference chemicals, effects of TRB were observed in 14 articles, FLU was in 8 articles, and 11-KT and VIN were included in 5 articles (Table 1). Other than the references, the ED effects of three compounds, DHT, LNG, and P4, were evaluated together (Onishi et al., 2021). Furthermore, among the androgen agonists, the AED effects of MT were peer-reviewed in eight articles, followed by GEN (four articles) (Table 4). Among the other agonists, LNG was reviewed in three articles, and the remaining seven chemicals were studied only once (Table 5). Among the apical endpoints, masculinization was induced by BF, GEN, LNG, P4, and SPR, while downregulation of hepatic vtg in females was observed in MT and SPR (Table 5). Among the eight chemicals identified as potential antagonists, the most studied chemical was DZ, which was studied in three articles (Hamm and Hinton, 2000; Flynn et al., 2018; Kawashima et al., 2022), followed by LD-BP and FNT, which were studied in two articles each (Li et al., 2016; 2017; Horie et al., 2017; 2022a). Other than these chemicals, the remaining five chemicals were studied once (one article/chemical). Moreover, based on the targeted apical endpoints related to AED and the literature review, we recommend that nine chemicals showed enough potential to proceed to Tier 2 tests, and five chemicals (FLU, 11-KT, MT, TRB, and VIN) did not require Tier 2 tests for the evaluation of androgen signaling mechanisms. In addition, similar to EEDs, the thyroid-related apical endpoints, such as hypertrophy of thyroid follicular cells, were induced by 11-KT (reference agonist) and FLU (reference antagonist) in Japanese medaka (Leon et al., 2007). Other than the references, LNG (agonist) and DZ (antagonist) showed the potential to modulate swim bladder inflation in Japanese medaka larvae during development (Hamm and Hinton, 2000; Pandelides et al., 2021). Furthermore, GEN (agonist) shows potential to regulate the expression of dio2 mRNAs in larvae if the embryos were exposed to GEN during development (Schiller et al., 2013; 2014). Therefore, during the classification of EDCs as AED, the thyroid-related apical endpoints should not be ignored.
3.3 TEDs
For TEDs, three chemicals, T3 as the agonist and PFOA and TBBPA as antagonists, were considered references (Table 1). The apical endpoints, such as swim bladder inflation in larvae, disruption of thyroid histopathology, and up- or downregulation of TH receptor genes (trα and trβ) and deiodinases (dio1 and dio2), were considered during TED evaluations. Our literature search found only 19 articles, which is 9.26% of the total articles (205 articles) sorted are focused on TED. From these articles, 12 chemicals, one as agonist (Table 7), and 11 chemicals as antagonists, were identified as TEDs (Table 8). Considering three references, 15 chemicals, 2 as agonists (1.56% of 128 EDCs) and 13 as antagonists (10.16% of 128 EDCs), which is only 11.72% of the screened chemicals (128 chemicals), showed TED effects on Japanese medaka. Alternatively, for 100 EDCs, 1.56 chemicals are identified as TED agonists, and ∼10 chemicals are identified as TED antagonists. Moreover, 19 articles identified 15 chemicals, which indicated that one TED was reviewed in 1.266 articles (approximately 5 articles:4 chemicals). Moreover, 5 chemicals, including three references and two antagonists (DIC and EHMC), were recommended to proceed to Tier 2 tests. The reference agonist T3 was studied in two articles, and the reference antagonists PFOA and TBBPA were included in four articles and 1 article, respectively (Table 1). Other than the references, the most studied chemical as a TED antagonist in Japanese medaka was DIC, which was peer-reviewed in five articles (Hong et al., 2007; Lee et al., 2011; Yokota et al., 2017; 2018; Pandelides et al., 2021). Other chemicals, such as ATBC, PFBA, PFOS/PFOSA, and TDCPP, were studied in two articles each. The remaining four antagonists were studied only once (Table 8). Although ATBC and TDCPP were evaluated as TED antagonists, the downregulation of liver vtg1 and vtg2 genes in XX fish by ATBC (Horie et al., 2023b) and upregulation of vtg mRNA in both male and female larvae by TDCPP (Godfrey et al., 2019) indicated that TED chemicals have the potential to regulate EAS pathways, which need further verifications.
3.4 MOS
For identification of the MOS chemicals in Japanese medaka, four chemicals, TPA and TRF as stimulators and KTC and PCZ as inhibitors, were used as reference chemicals (Table 1). The apical endpoints selected for steroidogenesis are either the up- or downregulation of cyp19 genes that show potential to regulate the aromatase enzyme activity and lead to an increase or decrease in the circulating estrogen level in Japanese medaka. Our literature search selected 26 articles, which is 12.68% of the sorted articles, for the evaluation of steroidogenesis in Japanese medaka (Table 2). After reviewing these literature reports, four chemicals were considered stimulators of steroidogenesis and six chemicals were considered inhibitors (Table 9). Including the references, the total number of chemicals that interrupt steroidogenesis is 14, 6 stimulators (∼5%), and 8 inhibitors (∼6%), which is 10.93% of the identified chemicals that showed potential ED activities in Japanese medaka. Alternatively, for every 100 EDCs, 5 chemicals show potential to stimulate steroidogenesis and 8 chemicals inhibit steroidogenesis. Moreover, 14 MOS were identified after reviewing 26 articles, which indicated that for the identification of a chemical as MOS, 1.857 articles/MOS are reviewed (approximately 9 articles: 5 chemicals). Moreover, although the thyroid-related endpoints were not considered in these chemicals, including two references (TPA and TRF as agonists), nine chemicals (six as agonists and three as antagonists) were recommended for Tier 2 tests (Tables 9, 10). Among the stimulators, the ED activities of FPN, an insecticide, and TRI, a pharmaceutical product, were studied together (Sun et al., 2014). However, FPN was included separately in two articles (Sun et al., 2014; Wagner et al., 2017); the remaining three chemicals, OCL, RCT, and TRI, were investigated once (Sun et al., 2014; 2016a; Yan et al., 2020) (Table 9). Among inhibitors, the most studied chemical is FAD, a nonsteroidal aromatase inhibitor, which was studied in five articles (Suzuki et al., 2004; Kuhl and Brower, 2006; Zhang et al., 2008b; Park et al., 2008; Thresher et al., 2011). Moreover, LET, a nonsteroidal triazole, was included in four articles (Sun et al., 2007b; 2009; 2011a; Liao et al., 2014). BP, a UV filter used in cosmetics, was evaluated in three articles (Coronado et al., 2008; Kim et al., 2014; Kawashima et al., 2022), while the herbicide LNR and the organotin compound TPT-Cl were studied in two articles each (Table 10), and PRN was studied only once (Schiller et al., 2014). Although the apical endpoints of MOS are mainly concentrated on aromatase enzyme genes and enzyme activities, the ED effects of these compounds on Japanese medaka either as an EED or AED can also be observed in TRI (Yan et al., 2020), RCT (Sun et al., 2016a), BP (Coronado et al., 2008; Kawashima et al., 2022), FAD (Zhang et al., 2008b), and LET (Sun et al., 2007b).
3.5 Unclassified
Due to limitations in the selection of apical endpoints, we were unable to identify the targeted EATS pathways of 45 chemicals (35.15% of the EDCs) identified from 60 (29.26% of the articles sorted) articles (Table 11). Alternatively, among 100 EDCs, 35 chemicals remained unclassified within the EATS modalities due to the lack of sufficient information (Table 11). Moreover, 45 unidentified EDCs in 60 sorted articles indicated that one chemical remained unidentified in 1.33 articles reviewed (4 articles:3 chemicals). Among these chemicals, the ED potential of GO was described in the maximum number of articles (10 articles) targeting the gonads, thyroid, interrenal glands, and endocrine pancreas of Japanese medaka (Dasmahapatra et al., 2020a; Dasmahapatra et al., 2020b; Myla et al., 2021; Asala et al., 2021; Myla et al., 2021; Dasmahapatra and Tchounwou, 2022a; Asala et al., 2022; Dasmahapatra and Tchounwou, 2022b; Dasmahapatra and Tchounwou, 2023a; Dasmahapatra and Tchounwou, 2023b). Moreover, TBT, a biocide used in agriculture, has been studied in six articles and showed the potential to inhibit brain aromatase in Japanese medaka (Nozaka et al., 2004; Khul and Brouwer, 2006; Hano et al., 2007; Zhang et al., 2008; Horie et al., 2018; Horie et al., 2022a). Furthermore, TCS, an antimicrobial product, was peer-reviewed in five articles that showed potential to enhance hepatic VTG in male fish (Foran et al., 2000; Ishibashi et al., 2004; Mihaich et al., 2019; Song et al., 2020; Kawashima et al., 2022). In addition, the flame retardant 2,2′,4,4′-BDE47 was peer-reviewed in four articles, although it was unable to target any of the EATS-related pathways in Japanese medaka (Gonzalez-Doncel et al., 2014b; 2016; 2017; Beltran et al., 2022). Among others, ACT, BKC, ClxBPA, CMP, IBP, LIN, MET, PDM, RLX, SFT, and TCS, although studied in a limited number of articles (except TCS, in most cases one or two articles), showed estrogenic potential by inducing the serum or liver VTG content in male fish (Foran et al., 2000; Ishibashi et al., 2004; Kim et al., 2012; Flynn et al., 2017; Godfrey et al., 2019; Mihaich et al., 2019; Kim et al., 2020; Song et al., 2020; Onishi et al., 2021; Kawashima et al., 2022). Furthermore, NPX, a NSAID, showed estrogenic potential by upregulating the expression of vtg1, erβ, and cyp17 genes in Japanese medaka (Kwak et al., 2018). Moreover, the potential ED effects produced by the rest of the chemicals (Table 11) are either due to induction of stress or mediated through pathways other than EATS.
4 Discussion
Japanese medaka (Oryzias latipes) is one of the small laboratory fish models used for the evaluation of EDCs found in the environment (OECD, 2018). Like all other vertebrates, EATS pathways and their associated hypothalamus pituitary-releasing and -stimulating hormones are targeted by EDCs and disrupt the normal development and reproductive processes of this fish. For the identification of EDCs that specifically affect the endocrine systems of Japanese medaka (O. latipes), we searched the research articles in PubMed (http://www.ncbi.nlm.nih.gov/pubmed) and Google Scholar (https://scholar.google.com/) databases with the search terms, Japanese medaka, O. latipes, and endocrine disruptions. We hypothesized that literature search and evaluation can identify the number and sources of EDCs that disrupted the EATS-related pathways of Japanese medaka (Oryzias latipes) and provide additional evidence for the selection of a chemical as to whether to proceed to Tier 2 tests or not.
We sorted 205 articles that involved 128 chemicals for review (Figures 1, 2; Tables 1–11). Due to wide variations in experimental protocols and methodologies described in the research articles (n = 205), especially in non-TG studies, interpretation of the data from the literature survey became more complex. Moreover, the use of different life stages (embryos/larvae/adults), diversity in the modes of exposure (injection, immersion, and feeding), or in the duration of exposure (restricted either only in one generation or continued through multiple generations) made the problem even more complex. Therefore, to maintain consistency in the apical endpoints associated with ED effects, among the 128 identified chemicals, we selected 14 chemicals as the reference (Table 1). These chemicals (references) are either evaluated in this model (Japanese medaka) as reference chemicals by other investigators or screened through Tier 2 tests, following OECD guidelines (Flynn et al., 2017; Onishi et al., 2021; Kawashima et al., 2022; Myosho et al., 2022). Among these chemicals, E2 and EE2 (estrogen agonists), TAM (estrogen antagonist), 11-KT and TRB (androgen agonists), FLU and VIN (androgen antagonists), and KTC and PCZ (steroidogenesis inhibitors) were verified as agonists or antagonists for esr1 (for estrogen) and arβ (androgen) genes of Japanese medaka in vitro by RGA (Onishi et al., 2021; Kawashima et al., 2022). Additionally, the potential of E2, TAM, TRB, VIN, KTC, and PCZ as an EDC was evaluated in medaka through Tier 2 tests, following the MEOGRT protocol (Flynn et al., 2017). For stimulators of steroidogenesis, we considered TPA and TRF as reference chemicals (Jang and Ji, 2015; Zhu et al., 2015). For the thyroid, T3 as the agonist and PFOA and TBBPA as antagonists were considered, which were recently referenced by Godfrey et al. (2019) and Horie et al. (2023a) in Japanese medaka. Therefore, we think that the selection of reference chemicals for the identification of EATS-related apical endpoints and to set up guidelines is very reasonable and acceptable. Our approach identified 69 chemicals that show potential to target the EATS pathways of Japanese medaka, and 45 chemicals remained unclassified due to limited information, even though these unclassified chemicals induced ED-like effects in Japanese medaka (Table 11). Taken together, considering 14 references, 83 (69 identified +14 references = 83) chemicals are identified as EDCs (∼65%) that disrupt EATS pathways of Japanese medaka (O. latipes), and 45 EDCs (∼35%) remain unclassified due to the lack of sufficient information.
We further classified the EATS chemicals as agonists/stimulators and antagonists/inhibitors of EEDs, AEDs, and TEDs, and MOS. The apical endpoints selected for agonists should be in contrast with antagonists, and in many cases, these borderlines cannot be maintained. For example, one of the significant apical endpoints of an EED as an agonist is the upregulation of VTG in the liver of male (XY) medaka (Flynn et al., 2017); however, TAM, which was used as a reference chemical of the EED antagonist, increased the liver VTG content in male fish (Flynn et al., 2017). To avoid complicacy, during analysis, we ignored the classification of EATS chemicals as agonists and antagonists, and simply included all the agonists and antagonists together and expressed them as EEDs, AEDs, TEDs, and MOS where applicable (Table 2).
As mentioned previously, 128 EDCs were identified after reviewing 205 individual articles, which indicates that for the identification of a chemical as an EDC in Japanese medaka, more than one article was reviewed (1.60 articles/chemical, or the approximate ratio is 8 chemicals: 13 articles). Our studies also showed that after reviewing 165 articles, 83 EDCs were identified that targeted EATS pathways (Tables 3–10), and 45 chemicals remained unidentified after reviewing 60 articles (Table 11). Accordingly, approximately 65% of the EDCs were identified with their specific EATS targets after reviewing 80% of the searched articles and 35% of the EDCs remained unclassified after reviewing 20% of the searched articles (Table 2). Therefore, it appears that the databases consist of more articles as classified EDCs (related to EATS) than unclassified EDCs (Table 2). Moreover, as the EATS pathways are interdependent on each other through the common hypothalamus–pituitary axis (HP axis), it is very difficult to classify the EDCs on the basis of apical endpoints specific to the EATS pathways. However, our studies showed that more than 65% of the articles identified EDCs as EED, 28% of the articles identified EDCs as AED, 12% of the articles identified EDCs as TED, and 16% of the articles identified EDCs as MOS (Table 2), which can be arranged in the order of TED < MOS < AED < EED. Furthermore, among 83 EDCs that targeted EATS pathways, 39% of them are identified as EEDs, 27% are AEDs, 18% are TEDs, and 17% are MOS (Tables 3–10), and the order of arrangement appears to be MOS < TED < AED < EED. Therefore, the potential of literature searching to identify EATS-targeted chemicals in Japanese medaka partially supports the concept that the more the number of articles in the databases, the more the number of EDCs should be identified.
As recommended by USEPA, the effects of an EDC should be evaluated using a tier-based approach. In Tier-1 studies, the endpoints are focused mainly on lethal concentrations (LC/LD/IC50, NOEC, and LOEC), reproductive activity (fecundity, fertility, breeding behavior, and hatching of the embryos), sex reversal, secondary sexual features (the number of papillae in the anal fin rays which are present in juvenile/adult males and absent in female Japanese medaka), VTG (the egg yolk precursor protein), and choriogenins (the eggshell protein), which are absent in the liver of male fish, and histopathology of the gonad, liver, and kidney. The Tier 2 approach is multigenerational, consisting mostly of the same features evaluated in Tier 1 (fecundity, fertility, hatching, VTG content of the male fish liver, secondary sexual features, sex reversal, survivability of embryos, larvae, and adults, and histopathology of the gonad, liver, and kidney). Even though the Tier 2 tests are time-consuming, expensive, and need proper validation of the chemicals as an EDC through Tier 1 screening, for proper classification of the EDCs and their respective target endocrine organs or hormones in fish (Japanese medaka), multigenerational studies (Tier 2) are necessary (Kawashima et al., 2022). Accordingly, among 83 EATS (69 classified and 14 references), we recommend that six of the references (11-KT, T3, PFOA, TBBPA, TPA, and TRF), due to the limited number of articles (studies in Japanese medaka), should be considered high-priority candidate substances for Tier 2 testing. The eight other references (E2, EE2, TAM, TRB, FLU, VIN, KTC, and PCZ) were already verified either as reference chemicals during the evaluation of other EDCs or through multigenerational MEOGRT tests (OECD TG 240) (Flynn et al., 2017; Onishi et al., 2021; Kawashima et al., 2022). Therefore, these eight reference chemicals did not need any further Tier 2 tests for potential EAS-related effects; however, evaluation of thyroid-dependent mechanisms of these chemicals may require investigation (Myosho et al., 2019, 2021; Pandelides et al., 2021).
During screening of EEDs, among the identified chemicals, we recommend 16 (AR-1260, BZP, BMT, CFR, CTC, p,p′-DDE, DES, EDS, EQ, E3, 4-MBC, MPB, OMC, PPB, 4t-PP, and TBCO) as agonists, and 4 chemicals (ATZ, MET, PFAA, and TPhP) as antagonists were high-priority chemicals for Tier 2 tests. Among the rest, EED potentials of o,p′-DDT and 4-t-OP were evaluated by multigenerational MEOGRT tests (Flynn et al., 2017) and probably did not require any further Tier 2 tests as well (Flynn et al., 2017). In addition, BPA, E1, 4-NP, and 4-t-OP were recommended for Tier 2 tests after successful evaluation through the OECD TG 229 protocol (Kawashima et al., 2022). Moreover, our literature search found that BPA was reviewed in 21 articles, 4-NP in 23 articles, 4-t-OP in 8 articles, and E1 in 3 articles (Table 3). Therefore, we believe that these EEDs (E1, BPA, o,p′-DDT, 4-NP, and 4-t-OP) showed enough potential to be considered EED agonists without performing any further Tier 2 tests. In AEDs, six chemicals (BF, CFD, GEN, LNG, P4, and SPR) as agonists and three chemicals (CPA, DZ, and 2-EHHB) as antagonists were recommended for Tier 2 tests (Tables 5, 6). Moreover, our literature search showed that MT was studied in eight articles and probably did not require Tier 2 tests anymore. However, P4 and LNG, as progestins, induced secondary sexual features in female Japanese medaka (XX) (Onishi et al., 2021), and further evaluation by Tier 2 tests is necessary. Among TEDs, two antagonists (DIC and EHMC) were recommended for Tier 2 tests and for MOS, four chemicals (OCL, FPN, RCT, and TRI), as stimulators, and three chemicals (BP, FAD and LET), as inhibitors, were recommended for Tier 2 tests. Taken together, among the 83 EDCs that targeted EATS pathways, 43 chemicals were recommended for Tier 2 tests, and 13 chemicals can be considered potential EDCs without any further Tier 2 tests in Japanese medaka.
Our literature search did not classify the EATS pathways of 45 chemicals (35%), even though several of them induced specific EATS-related apical endpoints (Table 11). Generally, in in vivo studies, probably due to the HPG and HPT axes, the overlapping effects of the chemicals within the EATS pathways cannot be ruled out; therefore, many of these unclassified chemicals demonstrated effects on endocrine-related apical endpoints, such as alteration in the liver VTG content (upregulated by CMP, CHDM, IBP, MTZ, NPX, 1NT, PDM, and RLX, and downregulated by DIBP), upregulation of chg in the liver of male fish (OXY and SRF), impaired reproductive activity and gonad histology (Cd, LD-BP, and nZnO), histopathological changes in the thyroid (BDE-47), inhibition of aromatase (TBT), and regulation of the E2 concentration in the blood of fish (Cd and SFT) (Table 11). In addition, several of the unclassified EDCs have potential as ESR agonists (CMP, DIBP, and FU) or antagonists (CYN, PHT, and RLX), and the ESR agonist and ARβ agonist (INT) and ESR agonist and ARβ antagonist (CMP) were observed in in vitro RGA with medaka esr1 and arβ genes (Onishi et al., 2021; Kawashima et al., 2022). Moreover, the nanocarbon, GO, was evaluated in 10 articles targeting the gonads, thyroid, interrenal glands, and pancreas in adults; and gonads, thyroids, and interrenal glands in larvae (Dasmahapatra et al., 2020a; Dasmahapatra et al., 2020b; Dasmahapatra and Tchounwou, 2022a; Dasmahapatra and Tchounwou, 2022b; Dasmahapatra and Tchounwou, 2023a; Dasmahapatra and Tchounwou, 2023b; Asala et al., 2021; 2022; Myla et al., 2021a; b). Despite the histopathological alterations and cellular disruptions induced in the gonads, liver, kidneys, thyroid, interrenal glands, and pancreas of the adults and larvae of Japanese medaka by GO, due to the lack of specific Tier 1 and Tier 2 tests, GO remained unclassified without identifying any EATS-specific pathways (Dasmahapatra et al., 2020a; Dasmahapatra et al., 2020b; Dasmahapatra and Tchounwou, 2022a; Dasmahapatra and Tchounwou, 2022b; Dasmahapatra and Tchounwou, 2023a; Dasmahapatra and Tchounwou, 2023b; Asala et al., 2021; 2022; Myla et al., 2021a; b). Therefore, we think that, before excluding the potential of these unclassified chemicals as an ED, further validations using tier-based approaches are necessary. Alternatively, the effects should be considered nonspecific, mediated through oxidative stress, or not related to EATS-specific mechanisms.
Although the effects of 128 EDCs in Japanese medaka are classified based on EATS modalities, the disruptions of non-EATS pathways by these chemicals need to be investigated carefully (Martyniuk et al., 2022). Moreover, compared to EATS, less attention has been given to other endocrine organs, including the endocrine pancreas and the interrenal gland (adrenal gland), which should belong to non-EATS pathways of Japanese medaka. Due to the lack of validated in vivo or in vitro methods and the availability of the appropriate literature in the public databases, the evaluation of EDCs targeting non-EATS modalities of Japanese medaka is not properly focused on this review. Our literature search on the effects of EDCs on the endocrine pancreas and interrenal glands of Japanese medaka found only four articles, two for pancreas (Dasmahapatra and Tchounwou, 2023a; Dasmahapatra and Tchounwou, 2023b), and two for interrenal glands (Dasmahapatra and Tchounwou, 2022a; Dasmahapatra and Tchounwou, 2022b) (Tables 2, 11) in PubMed (www.ncbi.gov). Therefore, despite the significant importance of non-EATS modalities in Japanese medaka, due to the lack of sufficient literature and standard methods, the evaluation of EDCs mediated through non-EATS pathways is not appropriately described in this review article.
In conclusion, our strategies on the literature survey sorted 205 articles on Japanese medaka (O. latipes) that focused on 128 chemicals as EDCs. We found that 83 chemicals (∼65%) show potential as EDs targeting the EATS pathways. Although the overlapping of the endocrine-related apical endpoints cannot be ruled out, from the literature search, we classified 32 chemicals from 108 articles as EEDs, 22 chemicals from 46 articles as AEDs, 15 chemicals from 19 articles as TEDs, and 14 chemicals from 26 articles as MOS, and 45 EDCs from 60 articles remained unclassified. The number of EATS chemicals arranged in order (MOS < TED < AED < EED) fits well with the numbers identified by the literature search (TED < MOS < AED < EED). Moreover, 43 EDCs belonging to EATS are recommended for Tier 2 tests (∼34%), and 13 chemicals showed enough potential to be considered EDCs without any further tier-based studies (∼10%). Our evaluation of EDCs in Japanese medaka shows significant potential to further apply the laboratory-based research data for applications in regulatory risk assessments in humans.
Statements
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
AD: conceptualization, resources, data curation, formal analysis, investigation, methodology, and writing–original draft and review and editing. CW: formal analysis, investigation, resources, writing–original draft, and review and editing, validation, and visualization. AM: formal analysis, validation, and writing–original draft. ST: validation and writing–original draft. PT: conceptualization, funding acquisition, resources, supervision, and writing–review and editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The research was supported by the NIH/NIMHD grant #G12MD07581 (RCMI Center for Environmental Health), NIH/NIMHD grant #1U54MD015929 (RCMI Center for Health Disparities Research) and NSF grant #HRD 1547754 (CREST Center for Nanotoxicity Studies) at Jackson State University, Jackson, Mississippi, United States, and NIH/NIMHD grant #U54MD013376 (RCMI Center for Urban Health Disparities Research and Innovation) at Morgan State University, Baltimore, Maryland, United States. The content is solely the responsibility of the authors and does not necessarily represent the official view of the NIH or NSF.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/ftox.2023.1272368/full#supplementary-material
Glossary
| ACT | Acetaminophen |
| ATBC | Acetyl tributyl citrate |
| APEO | Alkylphenol polyethoxylate surfactants |
| AMT | Amitrole |
| AChE | Acetylcholinesterase enzyme |
| 11-OA | 5α-Androstan-3,11,17-trione |
| AED | Androgen endocrine disruptors |
| AR | Androgen receptor |
| AR-1260 | Aroclor 1260 |
| ATZ | Atrazine |
| AZM | Azinphos-methyl |
| BP2 | Benzophenone 2 |
| BP3 | Benzophenone 3 |
| BF | Bifenthrin |
| DEHS | Bis(2-ethylhexyl) sebacate |
| BPA | Bisphenol A |
| BZT-UV | Benzotriazole ultraviolet stabilizer |
| BKC | Benzalkonium chloride |
| i-BP | i-Butylparaben |
| n-BP | n-Butylparaben |
| Cd | Cadmium |
| CFD | Cefadroxil |
| CFR | Cefradine |
| ClxBPA | Chlorinated BPA |
| CMP | 4-Chloro-3-methylphenol |
| CLT | Chlorothalonil |
| CTC | Chlortetracycline |
| CHG | Choriogenin |
| CYN | Cyanazine |
| CHDM | 1,4-Cyclohexanedimethanol |
| CPA | Cyproterone acetate |
| dpf | Day post-fertilization |
| dph | Day post-hatch |
| DZ | Diazinon |
| DBP | Dibutyl phthalate |
| DIC | Diclofenac |
| p,p′-DDE | Dichlorodiphenyldichloroethylene |
| o,p′-DDT | Dichlorodiphenyltrichloroethane |
| DEHP | Diethylhexyl phthalate |
| DES | Diethylstilbestrol |
| DHT | 5α-Dihydrotestosterone |
| DIBP | Diisobutyl phthalate |
| EDs | Endocrine disruptors |
| EDSP | Endocrine Disruptor Screening Program |
| EDS | Endosulfan |
| END | Endrin |
| E2 | 17-β-Estradiol |
| E3 | Estriol |
| ESR | Estrogen receptor |
| E1 | Estrone |
| EQ | Equol |
| EAS | Estrogen–androgen–steroidogenesis |
| EATS | Estrogen–androgen–thyroid–steroidogenesis |
| EHMC | 2-Ethylhexyl-4-methoxycinnamate |
| EFSA | European Food Safety Authority |
| EMA | European Medicine Agency |
| EE2 | 17α-Ethinylestradiol |
| EU | European Union |
| FAD | Fadrozole |
| FNT | Fenitrothion |
| FNC | Fenoxycarb |
| FV | Fenvalerate |
| FLFII | Female leukophore-free strain |
| FAT | Fish acute toxicity test |
| FELS | Fish, early life stage toxicity test |
| FET | Fish embryo toxicity test |
| FSTRA | Fish short-term reproduction assay |
| FPN | Fipronil |
| FLX | Fluoxetine |
| FLR | Fluridone |
| FLU | Flutamide |
| GEN | Genistein |
| GLP | Glyphosate |
| gsdf | Gonadal soma-derived factor |
| GSI | Gonadosomatic index |
| GO | Graphene oxide |
| HSI | Hepatosomatic index |
| HBCD | Hexabromocyclododecane |
| hpf | Hour post fertilization |
| HPA | Hypothalamus–pituitary–adrenal axis |
| HPG | Hypothalamus–pituitary–gonadal axis |
| HPT | Hypothalamus–pituitary–thyroid axis |
| IBP | Ibuprofen |
| KC-400 | Kanecholor 400 |
| KTC | Ketoconazole |
| 11-KT | 11-Ketotestosterone |
| LC | Lethal concentration |
| LET | Letrozole |
| LNG | Levonorgestrel |
| LD-BP | Lignin-derived bisphenol |
| LIN | Lincomycin |
| LNR | Linuron |
| LOEC | Lowest observed effect concentration |
| LOEL | Lowest observed effect level |
| MEOGRT | Medaka extended one-generation reproduction test |
| MELA | Medaka embryo-larval development assay |
| MDA | Malondialdehyde |
| MET | Metformin |
| MTZ | Methimazole |
| MXC | Methoxychlor |
| 4-MBC | 3-(4-Methylbenzylidene) camphor |
| MPB | Methylparaben |
| MT | Methyltestosterone |
| MTC | Metolachlor |
| MOE | Ministry of Environment |
| mph | Month post-hatch |
| MOS | Modulator of steroidogenesis |
| MCB | Myclobutanil |
| NPX | Naproxen |
| 1NT | 1-Naphthol |
| NDEA | N-Nitrosodiethylamine |
| NSAID | Nonsteroidal anti-inflammatory drug |
| 4-NP | 4-Nonylphenol |
| NP1EC; NP2EC | Nonylphenol ethoxycarboxylate |
| NP1EO; NP2EO; NP9EO | Nonylphenol ethoxylate |
| NPEO | Nonylphenol polyethoxylates |
| NOEC | No-observed-effect concentration |
| OCL | Octocrylene |
| OMC | Octyl methoxycinnamate |
| 4-OP | 4-Octylphenol |
| OECD | Organization of Economic Cooperation and Development |
| OYZ | Oryzalin |
| OXF | Oxyfluorfen |
| OTC | Oxytetracycline |
| PDM | Pendimethalin |
| PCPL | Pentachlorophenol |
| 4t-PP | 4-Tert-pentylphenol |
| PFAA | Perfluoroalkyl acid |
| PFBA | Perfluorobutyric acid |
| PFOA | Perfluorooctanoic acid |
| PFOS | Perfluorooctane sulfonate |
| PFOSA | Perfluorooctane sulfonic acid |
| PFAA | Perfluoroalkyl acid |
| PFBS | Perfluorobutane sulfonate |
| PFNA | Perfluorononanoic acid |
| PPAR | Peroxisome proliferator-activated receptor |
| PHT | Phenytoin |
| PCB 126 | Polychlorinated biphenyl 126 |
| PCZ | Procloraz |
| P4 | Progesterone |
| PRN | Propanil |
| PPB | Propylparaben |
| PTU | 6-Propyl-2-thiouracil |
| RCT | Ractopamine |
| RLX | Raloxifene |
| REP | Relative estrogenic potency |
| RGA | Reporter gene assay |
| RND | Roundup |
| SERM | Selective estrogen receptor modulator |
| SDS | Sodium dodecyl sulfate |
| SPC | Sodium perchlorate |
| SPR | Spironolactone |
| SFT | Sulfathiazole |
| SRF | Surflan |
| TAM | Tamoxifen |
| TPA | Terephthalic acid |
| TG | Test guidelines |
| 4t-OP | 4-Tert-octylphenol |
| T | Testosterone |
| TBBPA | Tetrabromobisphenol A |
| TBCO | 1.2,5,6-Tetrabromocyclooctane |
| BDE-47 | 2,2′,4,4′Tetrabromodiphenyl ether |
| TU | Thiourea |
| TED | Thyroid endocrine disruptors |
| TH | Thyroid hormone |
| TR | Thyroid hormone receptor |
| TSH | Thyroid-stimulating hormone |
| T4 | Thyroxine |
| TRA | 17α-Trenbolone |
| TRB | 17β-Trenbolone |
| TRD | Triadimenol |
| TRF | Triadimefon |
| TBT | Tributyltin |
| TCS | Triclosan |
| TCrP | Tricresyl phosphate |
| TRF | Trifloxystrobin |
| T3 | Triiodothyronine |
| TRI | Trilostane |
| TPhP | Triphenyl phosphate |
| TPT-Cl | Triphenyltin chloride |
| TDCPP | Tris (1,3-dichloro-2-propyl) phosphate |
| USEPA; EPA | United States Environmental Protection Agency |
| USFDA | United States Food and Drug Administration |
| VIN | Vinclozolin |
| VTG | Vitellogenin |
| wph | Week post-hatch |
| nZnO | Nano zinc oxide |
| ZnSO4 | Zinc sulfate |
References
1
Abdel-Moneim A. Mahapatra C. T. Hatef A. Sepulveda M. S. (2015). Ovarian structure protein 1: a sensitive molecular biomarker of gonadal intersex in female Japanese medaka after androgen exposure. Environ. Toxicol. Chem.34, 2087–2094. 10.1002/etc.3032
2
Asahina K. Urabe A. Sakai T. Hirose H. Hibiya T. (1989). Effects of various androgens on the formation of papillary processes on the anal fin rays in the female medaka <i>Oryzias latipes</i>. Oryzias latipes Nippon. Gakkaishi55, 1871. 10.2331/suisan.55.1871
3
Asala T. E. Dasmahapatra A. K. Myla A. Tchounwou P. B. (2021). Exerimental data sets on the evaluation of graphene oxide as a thyroid endocrine disruptor and modulator of gas gland cells in Japanese medaka medaka (Oryzias latipes) larvae at the onset of maturity. Data Brief.39, 107625. 10.1016/j.dib.2021.107625
4
Asala T. E. Dasmahapatra A. K. Myla A. Tchounwou P. B. (2022). Histological and histochemical evaluation of graphene oxide on thyroid follicles and gas gland of medaka larvae (Oryzias latipes). Chemosphere286, 131719. 10.1016/j.chemosphere.2021.131719
5
Balch G. Metcalfe C. (2006). Developmental effects in Japanese medaka (Oryzias latipes) exposed to nonylphenol ethoxylates and their degradation products. Chemosphere62, 1214–1223. 10.1016/j.chemosphere.2005.02.100
6
Balch G. C. Mackenzie C. Metcalfe C. D. (2004a). Alterations of gonadal development and reproductive success in Japanese medaka (Oryzias latipes) exposed to 17α-ethinylestradiol. Environ. Toxicol. Chem.23, 782–791. 10.1897/02-539
7
Balch G. C. Shami K. Wilson P. J. Wakamatsu Y. Metcalfe C. D. (2004b). Feminization of female leukophore-free strain of Japanese medaka (Oryzias latipes) exposed to 17β-estradiol. Environ. Toxicol. Chem.23, 2763–2768. 10.1897/03-633
8
Beltran E. M. Gonzalez-Doncel M. Garcia-Maurino J. E. Hortigiiela P. G. Pablos M. V. (2022). Effects of lifecycle exposure to dietary 2, 2′, 4,4′-tetrabromodiphenyl ether (BDE-47) on medaka fish (Oryzias latipes). Aqua Toxicol.245, 106133. 10.1016/j.aquatox.2022.106133
9
Bertotto L. B. Bruce R. Li S. Richards J. Sikder R. Baljkas L. et al (2019). Effects of bifenthrin on sex determination in Japanese medaka (Oryzias latipes). Environ. Res.177, 108564. 10.1016/j.envres.2019.108564
10
Bhandari R. K. vom Saal F. S. Tillitt D. E. (2015). Transgenerational effects from early developmental exposure to bisphenol A or 17 alpha-ethinylestradiol in medaka, Oryzias latipes. Sci. Rep55, 9303. 10.1038/srep09303
11
Bhandari R. K. Wang X. vom Saal F. S. Tillitt D. E. (2020). Transcriptome analysis of testis reveals the effects of developmental exposure to bisphenol A or 17α-ethinylestradiol in medaka (Oryzias latipes). Aquat. Toxicol.225, 105553. 10.1016/j.aquatox.2020.105553
12
Cheek A. O. Brouwer T. H. Carroll S. Manning S. McLachlan J. A. Brouwer M. (2001). Experimental evaluation of vitellogenin as a predictive biomarker for reproductive disruption. Environ. Health Perspt.109, 681–690. 10.1289/ehp.01109681
13
Chen P.-J. Rosenfeldt E. J. Kullman S. W. Hinton D. E. Linden K. G. (2007). Biological assessments of a mixture of endocrine disruptors at environmentally relevant concentrations in water following UV/H2O2 oxidation. Sci. Total Environ.376, 18–26. 10.1016/j.scitotenv.2006.12.051
14
Chen R. He J. Li Y. An L. Hu J. (2022). Tricresyl phosphate inhibits fertilization in Japanese medaka (Oryzias latipes): emphasizing metabolic toxicity. Environ. Pollut.297, 118809. 10.1016/j.envpol.2022.118809
15
Chikae M. Ikeda R. Hasan Q. Morita Y. Tamiya E. (2004). Effects of Tamoxifen, 17α-ethynylestradiol, flutamide, and methyltestosterone on plasma vitellogenin levels of male and female Japanese medaka (Oryzias latipes). Environ. Toxicol. Pharmacol.17, 29–33. 10.1016/j.etap.2004.02.002
16
Chu S. H. Liao P. H. Chen P. J. (2016). Developmental exposures to an azole fungicide triadimenol at environmentally relevant concentrations cause reproductive dysfunction in females of medaka fish. Chemosphere152, 181–189. 10.1016/j.chemosphere.2016.02.078
17
Coronado M. De Haro H. Deng X. Rempel M. A. Lavado R. Schlenk D. (2008). Estrogenic activity and reproductive effects of the UV-filter oxybenzone(2-hydroxy-4-methoxyphenyl-methanone) in fish. Aquat. Tox90, 182–187. 10.1016/j.aquatox.2008.08.018
18
Dang Z. Arena M. Kienzler A. (2021). Fish toxicity testing for identification of thyroid disrupting chemicals. Environ. Pollut.284, 11734. 10.1016/j.envpol.2021.117374
19
Dasmahapatra A. K. Powe D. K. Dasari T. P. S. Tchounwou P. B. (2020a). Assessment of reproductive and developmental effects of graphene oxide on Japanese medaka (Oryzias latipes). Chemosphere259, 127221. 10.1016/j.chemosphere.2020.127221
20
Dasmahapatra A. K. Powe D. K. Dasari T. P. S. Tchounwou P. B. (2020b). Experimental data sets on the characterization of graphene oxide and its reproductive and developmental effects of graphene oxide on Japanese medaka (Oryzias latipes). Data. Brief.32, 106218. 10.1016/j.dib.2020.106218(2020b)
21
Dasmahapatra A. K. Tchounwou P. B. (2022a). Histological evaluation of the interrenal gland (adrenal homolog) of Japanese medaka (Oryzias latipes) exposed to graphene oxide. Environ. Toxicol.37, 2460–2482. 10.1002/tox.23610
22
Dasmahapatra A. K. Tchounwou P. B. (2022b). Experimental datasets on the histopathological and immunohistological assessment of the interrenal gland (adrenal homolog) of Japanese medaka (Oryzias latipes) fish exposed to graphene oxide. Data Brief.45, 108693. 10.1016/j.dib.2022.108693
23
Dasmahapatra A. K. Tchounwou P. B. (2023a). Evaluation of pancreatic δ-cells as a potential target site of graphene oxide toxicity in Japanese medaka (Oryzias latipes) fish. Ecotoxicol. Environ. Saf.253, 114649. 10.1016/j.ecoenv.2023.114649
24
Dasmahapatra A. K. Tchounwou P. B. (2023b). Experimental datasets on the immunohistological assessment of δ-cells in the islet organ of the endocrine pancreas of Japanese medaka (Oryzias latipes) fish exposed to graphene oxide. Data Brief.48, 109213. (in press). 10.1016/j.dib.2023.109213
25
Devoy C. Raza Y. Kleiner M. Jones P. D. Doering J. A. Wiseman S. (2023). The brominated flame retardant, 1,2,5,6, -tetrabromocyclooctane(TBCO) causes multigenerational effects on reproductive capacity of Japanese medaka (Oryzias latipes). Chemosphere313, 137561. 10.1016/j.chemosphere.2022.137561
26
Edmunds J. S. McCarthy R. A. Ramsdell J. S. (2000). Permanent and functional male to female sex reversal in d-rR strain medaka (Oryzias latipes) following egg microinjection of o,p’-DDT. Environ. Health Perspect.108, 219–224. 10.1289/ehp.00108219
27
Flippin J. L. Huggett D. Foran C. M. (2007). Changes in the timing of reproduction following chronic exposure to ibuprofen in Japanese medaka, Oryzias latipes. Aquat. Toxicol.81, 73–78. 10.1016/j.aquatox.2006.11.002
28
Flynn K. Lothenbach D. Whiteman F. Hammermeister D. Swintek J. Etterson M. et al (2018). The effects of continuous diazinon exposure on growth and reproduction in Japanese medaka using a modified Medaka Extended One Generation Reproduction Test (MEOGRT). Ecotoxicol. Environ. Saf.162, 438–445. 10.1016/j.ecoenv.2018.06.088
29
Flynn K. Lothenbach D. Whiteman F. Hammermeister D. Touart L. W. Swintek J. et al (2017). Summary of the development the US environmental protection agency's medaka extended one generation reproduction test (MEOGRT) using data from 9 multigenerational medaka tests. Environ. Toxicol. Chem.36, 3387–3403. 10.1002/etc.3923
30
Flynn K. Swintek J. Johnson R. (2013). Use of gene expression data to determine effects on gonad phenotype in Japanese medaka after exposure to trenbolone or estradiol. Environ.Toxicolo. Chem.32, 1344–1353. 10.1002/etc.2186
31
Foran C. M. Bennett E. R. Benson W. H. (2000). Developmental evaluation of a potential non-steroidal estrogen: triclosan. Mar. Environ. Res.50, 153–156. 10.1016/s0141-1136(00)00080-5
32
Foran C. M. Peterson B. N. Benson W. H. (2002). Transgenerational and developmental exposure of Japanese medaka (Oryzias latipes) to ethinylestradiol as adults. Toxicol. Sci.68, 389–402.
33
Foran C. M. Weston J. Slattery M. Brooks B. W. Huggett D. B. (2004). Reproductive assessment of Japanese medaka (Oryzias latipes) following a four-week fluoxetine (SSRI) exposure. Arch. Environ. Contam. Toxicol.46, 511–517. 10.1007/s00244-003-3042-5
34
Francis R. C. (1992). Sexual lability in teleosts: developmental factors. Q. Rev. Biol.67, 1–18. 10.1086/417445
35
Fujita K. K. Doering J. Stock E. Lu Z. Montina T. Wiseman S. (2022). Effects of dietary 2(2H-benzotriazol-2yl)-4methyl-phenol (UV-P) exposure on Japanese medaka (Oryzias latipes) in a short-term reproduction assay. Aquat. Tocicol.248, 106206–206. 10.1016/j.aquatox.2022.106206
36
Godfrey A. Hooser B. Abdelmoneim A. Sepulveda M. S. (2019). Sex-specific endocrine disrupting effects of three halogenated chemicals in Japanese medaka. J. Appl. Toxicol.39, 1215–1223. 10.1002/jat.3807
37
Gonzalez-Doncel M. Carbonell G. Garcia-Maurino J. E. Sastre S. Beltran E. M. Fernandez Toriza C. (2016). Effects of diatery2, 2,′4,4′-tetrabromodiphenyl ether (BDE-47) exposure in growing medaka fish (Oryzias latipes). Aquat. Toxicol.178, 141–152. 10.1016/j.aquatox.2016.07.017
38
Gonzalez-Doncel M. Garcia-Maurino J. E. Segundo L. S. Beltran E. M. Sastre S. Torija C. F. (2014a). Embryonic exposure of medaka (Oryzias latipes) to propylparaben: effects on early development and post-hatching growth. Environ. Pollut.184, 360–369. 10.1016/j.envpol.2013.09.022
39
Gonzalez-Doncel M. Sastre S. Carbonell G. Beltran E. M. Anaya C. G. Garcia-Maurino J. E. et al (2017). Bioaccumulation, maternal transfer and effects of dietary 2,2’,4,4’-tetrabromodiphenyl ether (BDE-47) exposure on medaka fish (Oryzias latipes) offspring. Aquat. Tox192, 241–250. 10.1016/j.aquatox.2017.09.024
40
Gonzalez-Doncel M. Torija C. F. Beltran E. M. Garcia-Maurino J. E. Sastre S. Carbonell G. (2014b). Limitations of waterborne exposure of fish early life stages to BDE-47. Aqua Tox48, 184–194. 10.1016/j.aquatox.2014.01.015
41
Gray M. A. Metcalfe C. D. (1997). Induction of testis-ova in Japanese medaka (Oryzias latipes) exposed to p-nonylphenol. Environ. Toxicol. Chem.22, 1082–2445. 10.1897/1551-5028(1997)016<1082:iotoij>2.3.co;2
42
Gray M. A. Niimi A. J. Metcalfe C. D. (1999a). Factors affecting the development of testis-ova in medaka, Oryzias latipes, exposed to octylphenol. Environ. Toxicol. Chem.18, 1835–1842. 10.1897/1551-5028(1999)018<1835:fatdot>2.3.co;2
43
Gray M. A. Teather K. L. Metcalfe C. D. (1999b). Reproductive success and behavior of Japanese medaka (Oryzias latipes) exposed to 4-tert-octylphenol. Environ. Toxicol. Chem.18, 2587–2594. 10.1897/1551-5028(1999)018<2587:rsaboj>2.3.co;2
44
Green C. Brian J. Kanda R. Scholze M. Williams R. Jobling S. (2015). Environmental concentrations of antiandrogenic pharmaceuticals do not impact sexual disruption in fish alone or in combination with steroid oestrogens. Aquat. Toxicol.100, 117–127. 10.1016/j.aquatox.2014.12.022
45
Grillitsch B. Altmann D. Schabuss M. Zornig H. Sommerfeld-Stur I. Mostl E. (2010). Mammalian glucocorticoid metabolites act as androgenic endocrine disruptors in the medaka (Oryzias latipes). Environ. Toxicol. Chem.29, 1613–1620. 10.1002/etc.176
46
Gronen S. Denslow N. Manning S. Barnes S. Barnes D. Brouwer M. (1999). Serum vitellogenin levels and reproductive impairment of male Japanese medaka (Oryzias latipes) exposed to 4-tert-octylphenol. Environ. Health Perspect.107, 385–390. 10.1289/ehp.99107385
47
Hall L. C. Okihiro M. Johnson M. L. The S. J. (2007). Surflan TM and Oryzalin impair reproduction in the teleost medaka (Oryzias latipes). Mar. Environ. Res.63, 15–131. 10.1096/j.marenvres.2006.07.003
48
Hall L. C. Rogers J. M. Denison M. S. Johnson M. L. (2005). Identification of the herbicide surflan and its active ingredient oryzalin, a dinitrosulfonamide, as xenoestrogens. Arch. Environ. Contam. Toxicol.48, 201–208. 10.1007/s00244-003-0164-8
49
Hamm J. T. Hinton D. E. (2000). The role of development and duration of exposure to the embryotoxicity of diazinon. Aquat. tox48, 403–418. 10.1016/s0166-445x(99)00065-x
50
Han S. Choi K. Kim J. Ji K. Kim S. Ahn B. et al (2010). Endocrine disruption and consequences of chronic exposure to ibuprofen in Japanese medaka (Oryzias latipes) and freshwater cladocerans Daphnia magna and Moina macrocopa. Aqaut Toxicol.98, 256–264. 10.1016/j.aquatox.2010.02.013
51
Hano T. Oshima Y. Oe T. Kinoshita M. Tanaka M. Wakamatsu Y. et al (2005). Quantitative bioimaging analysis for evaluation of sexual differentiation in germ cells of olvas-GFP/STIIYI medaka (Oryzias latipes) nanoinjected in ovo with ethinylestradiol. Environ.Toxicol. Chem.24, 70–77.
52
Hano T. Oshima Y. Kim S. G. Satone H. Oba Y. Kitano T. et al (2007). Tributylin causes abnormal development in embryos of medaka, Oryzias latipes. Chemosphere69, 927–933. 10.1016/j.chemosphere.2007.05.093
53
Hashimoto S. Watanabe E. Ikeda M. Terao Y. Strussmann C. A. Inoue M. et al (2009). Effects of ethinylestradiol on medaka (Oryzias latipes) as measured by sperm mortality and fertilization success. Arch. Environ. Contam. Toxicol.56, 253–259. 10.1007/s00244-008-9183-9
54
Hirai N. Nanba A. Koshio M. Kondo T. Morita M. Tatarazako N. (2006). Feminization of Japanese medaka (Oryzias latipes) exposed to 17β-estradiol: effect of exposure period on spawning performance in sex-transformed females. Aquat. Toxicol.79, 288–295. 10.1016/j.aquatox.2006.06.018
55
Hirakawa I. Miyagawa S. Katsu Y. Kagami Y. Tatarazako N. Kobayashi T. et al (2012). Gene expression profiles in the testis associated with testis-ova in adult Japanese medaka (Oryzias latipes) exposed to 17 alpha-ethinylestradiol. Chemosphere87, 668–674. 10.1016/j.chemosphere.2011.12.047
56
Hirako A. Takeoka Y. Furukawa S. Sugiyama A. (2017). Effects of cadmium exposure on medaka (Oryzias latipes) testes. J. Toxicol. Pathol.30, 255–260. 10.1293/tox.2017-0015
57
Hishida T.-O. Kawamoto N. (1970). Androgenic and male-induced effects of 11-ketotestosterone on a teleost, the medaka (Oryzias latipes). J. Exp. Zool.173, 279–283. 10.1002/jez.1401730306
58
Hong H. N. Kim H. N. Park K. S. Lee S.-K. Gu M. B. (2007). Analysis of the effects diclofenac has on Japanese medaka (Oryzias latipes) using real-time PCR. Chemosphere67, 2115–2121. 10.1016/j.chemosphere.2006.12.090
59
Horie Y. Kanazawa N. Takahashi C. Tatarazako N. Iguchi T. (2019). Bisphenol A induces a shift in sex differentiation gene expression with testis-ova or sex reversal in Japanese medaka (Oryzias latipes). J. Appl. Toxicol.40, 804–814. 10.1002/jat.3945
60
Horie Y. Kanazawa N. Takahashi C. Tatarazako N. Iguchi T. (2021). Exposure to 4-nonylphenol induces a shift in the gene expression of gsdf and testis-ova formation and sex reversal in Japanese medaka (Oryzias latipes). Appl. Toxicol.41, 399–409. 10.1002/jat.4051
61
Horie Y. Kanazawa N. Takahashi C. Tatarazako N. Iguchi T. (2022a). Gonadal soma-derived factor expression is a potential biomarker for predicting effects of endocrine disrupting chemicals on gonadal differentiation in Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem.41, 1875–1884. 10.1002/etc.5353
62
Horie Y. Nomura M. Ernesto U. D. L. Naija A. Akkajit P. Okamura H. (2023b). Impact of acetyl tributyl citrate on gonadal sex differentiation and expression of biomarker genes for endocrine disruption in Japanese medaka. Aqut Tox106553, 106553. 10.1016/j-aquatox.2023.106553
63
Horie Y. Nomura M. Okamoto K. Takahashi C. Sato T. Miyagawa S. et al (2022d). Effect of thyroid hormone-disrupting chemicals on swim bladder inflation and thyroid hormone-related gene expression in Japanese medaka and zebrafish. J. Appl. Toxicol.42, 1385–1395. 10.1002/jat.4302
64
Horie Y. Nomura M. Ramaswami B. R. Harino H. Yap C. K. Okamura H. (2022c). Effects of nonphthalate plasticizer bis(2-ethylhexyl) sebacate (DEHS) on the endocrine system in Japanese medaka (Oryzias latipes). Comp. Biochem. Physiol. C Toxicol. Pharmacol.264, 109531. 10.1016/j.cbpc.2022.109531
65
Horie Y. Watanabe H. Takanobu H. Yagi A. Yamagishi T. Iguchi T. et al (2017). Development of an invivo anti-androgenic activity detection assay using fenitrothion in Japanese medaka (Oryzias latipes). J. Appl. Toxicol.37, 339–346. 10.1002/jat.3365
66
Horie Y. Yamagihsi T. Yamamoto J. Suzuki M. Onishi Y. Chiba T. et al (2023a). Adverse effects of thyroid-hormone-disrupting chemicals 6-propyl-2-thiouracil and tetrabromobisphenol A on Japanese medaka (Oryzias latipes). Comp. Biochem. Physiol. C Toxicol. Pharmacol.263, 109502. 10.1016/j.cbpc.2022.109502
67
Horie Y. Yamagishi T. Shintaku Y. Iguchi T. Tatarazako N. (2018). Effects of tributyltin on early life-stage, reproduction, and gonadal sex differentiation in Japanese medaka (Oryzias latipes). Chemosphere203, 418–425. 10.1016/j.chemosphere.2018.03.135
68
Horie Y. Yap C. K. Okamura H. (2022b). Developmental toxicity and thyroid hormone-disrupting effects of acetyl tributyl citrate in zebrafish and Japanese medaka. J. Hazard. Mat. Adv.8, 100199. 10.1016/j.hazadv.2022.100199
69
Horng C.-Y. Lin H.-C. Lee W. (2010). Reproductive toxicology study of phenanthrene in medaka (Oryzias latipes). Arch. Environ. Contam. Toxicol.58, 131–139. 10.1007/s00244-009-9335-6
70
Iguchi T. Watanabe H. Katsu H. (2006). Application of ecotoxicogenomics for studying endocrine disruption in vertebrates and invertebrates. Environ. Health Perspect.114, 101–105. 10.1289/ehp.8061
71
Inagaki T. Smith N. Lee E. K. Ramakrishnan S. (2016). Low dose exposure to Bisphenol A alters development of gonadotropin-releasing hormone 3 neurons and larval locomotor behavior in Japanese Medaka. NeuroToxicology52, 188–197. 10.1016/j.neuro.2015.12.003
72
Inui M. Adachi T. Takenaka S. Inui H. Nakazawa M. Ueda M. et al (2003). Effect of UV screens and preservatives on vitellogenin and choriogenin production in male medaka (Oryzias latipes). Toxicology194, 43–50. 10.1016/s0300-483x(03)00340-8
73
Ishibashi H. Hirano M. Matsumura N. Watanabe N. Takao Y. Arizono K. (2006). Reproductive effects and bioconcentration of 4-nonylphenol in medaka fish (Oryzias latipes). Chemosphere65, 1019–1026. 10.1016/j.chemosphere.2006.03.034
74
Ishibashi H. Matsumura M. Hirano M. Matsuoka M. Shiratsuchi H. Ishibashi Y. et al (2004). Effects of triclosan on the early life stages and reproduction of medaka Oryzias latipes and induction of hepatic vitellogenin. Aquat. Tox67, 167–179. 10.1016/j.aquatox.2003.12.005
75
Ishibashi H. Uchida M. Temma Y. Hirano M. Tominaga N. Arizono K. (2020). Choriogenin transcription in medaka embryos and larvae as an alternative model for screening estrogenic endocrine-disrupting chemicals. Eco Toxicol. Environ. Saf.193, 110324. 10.1016/j.ecoenv.2020.110324
76
Islinger M. Yuan H. Voelkl A. Braunbeck T. (2002). Measurement of vitellogenin gene expression by RT-PCR as a tool to identify endocrine disruption in Japanese medaka (Oryzias latipes). Biomarkers7, 80–93. 10.1080/13547500110086919
77
Jang S. Ji K. (2015). Effect of chronic exposure to two components of tritan copolyester on Daphnia magna, Moina macrocopa, and Oryzias latipes, and potential mechanisms of endocrine disruption using H295R cells. Ecotoxicol24, 1906–1914. 10.1007/s10646-015-1526-5
78
Ji K. Choi K. Lee S. Park S. Khim J. S. Jo E.-H. et al (2010). Effects of sulfathiazole, oxytetracycline and Chlortetracycline on steroidogenesis in the human adrenocarcinoma (H295R) cell line and freshwater fish Oryzias latipes. J. Hazard. Mat.182, 494–502. 10.1016/j.jhazmat.2010.06.059
79
Ji K. Kim S. Han S. Seo J. Lee S. Park Y. et al (2012). Risk assessment of chlortetracycline, oxytetracycline, sulfamethazine, sulfathiazole, and erythromycin in aquatic environment.: are the current environmental concentrations safe?Ecotoxicol21, 2031–2050. 10.1007/s10646-012-0956-6
80
Ji K. Kim Y. Oh S. Ahn B. Jo H. Choi K. (2008). Toxicity of perfluorooctane sulfonic acid and perfluorooctanoic acid on freshwater macroinvertebrates (Daphnia magna and Monia Macrocopa) and fish (Oryzias latipes). Environ. Toxicol. Chem.27, 2159–2168. 10.1897/07-523.1
81
Jin Y. Chen R. Wang L. Liu J. Yang Y. Zhou C. et al (2011a). Environmental cues influence EDC-induced endocrine disruption effects in different developmental stages of Japanese medaka (Oryzias latipes). Aqut. Toxicol.101, 254–260. 10.1016/j.aquatox.2010.10.005
82
Jin Y. Shu L. Huang F. Cao L. Sun L. Fu Z. et al (2011b). Effects of metolachlor on transcription of thyroid system-related genes in juvenile and adult Japanese medaka (Oryzias latipes). Gen. Comp. Endocrinol.170, 487–493. 10.1016/j.ygcen.2010.11.001
83
Kamata R. Shiraishi F. Nakajima D. Kageyama S. (2011). Estrogenic effects of leachates from industrial waste landfills measured by a recombinant yeast assay and transcriptional analysis in Japanese medaka. Quat. Tox101, 430–437. 10.1016/j.aquatox.2010.11.018
84
Kang I. J. Hano T. Oshima Y. Yokota H. Tsuruda Y. Shimasaki Y. et al (2006). Anti-androgen flutamide affects gonadal development and reproduction in Japanese medaka (Oryzias latipes). Mar. Environ. Res.62, S253–S257. 10.1016/j.marenvres.2006.04.065
85
Kang I. J. Yokota H. Oshima Y. Tsuruda Y. Hano T. Maeda M. et al (2003). Effects of 4-nonylphenol on reproduction of Japanese medaka, Oryzias latipes. Oryzias latipes Environ. Toxicol. Chem.22, 2438–2445. 10.1897/02-225
86
Kang I. J. Yokota H. Oshima Y. Tsuruda Y. Oe T. Imada N. et al (2002b). Effects of bisphenol A on the reproduction of Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem.21, 2394–2400. 10.1897/1551-5028(2002)021<2394:eobaot>2.0.co;2
87
Kang I. J. Yokota H. Oshima Y. Tsuruda Y. Shimasaki Y. Honjo T. (2008). The effects of methyl testosterone on the sexual development and reproduction of adult medaka (Oryzias latipes). Aquat. Toxicol.87, 37–46. 10.1016/j.aquatox.2008.01.010
88
Kang I. J. Yokota H. Oshima Y. Tsuruda Y. Yamaguchi T. Maeda M. et al (2002a). Effect of 17β-estradiol on the reproduction of Japanese medaka (Oryzias latipes). Chemosphere47, 71–80. 10.1016/s0045-6535(01)00205-3
89
Kang J. S. Ahn T.-G. Park J.-W. (2019). Perfluorooctanoic acid (PFOA) and perfluorooctane sulfonate (PFOS) induced different modes of action in reproduction of Japanese medaka (Oryzias latipes). J. Hazard Mat.368, 97–103. 10.1016/j.jhazmat.2019.01.034
90
Kashiwada S. Ishikawa H. Miyamoto N. Ohnishi Y. Magara Y. (2002). Fish test for endocrine-disruption and estimation of water quality of Japanese rivers. Wat Res.36, 2161–2166. 10.1016/s0043-1354(01)00406-7
91
Kawashima Y. Onishi Y. Tatarazako N. Yamamoto H. Koshio M. Oka T. et al (2022). Summary of 17 chemicals evaluated by OECD TG229 using Japanese medaka, Oryzias latipes in Extend 2016. J. Appl. Toxicol.42, 750–777. 10.1002/jat.4255
92
Kidd K. A. Blanchfield P. J. Mills K. H. Palace V. P. Evans R. E. Lazorchak J. M. et al (2007). Collapse of a fish population after exposure to a synthetic estrogen. Proc. Natl. Acad. Sci. U. S. A.104, 8897–8901. 10.1073/pnas.0609568104
93
Kim B. Ji K. Kho Y. Kim P.-G. Park K. Kim K. et al (2017). Effects of chronic exposure to cefadroxil and cefradine on Daphnia magna and Oryzias latipes. Chemosphere185, 844–851. 10.1016/j.chemosphere.2017.07.085
94
Kim P. Park Y. Ji K. Seo J. Lee S. Choi K. et al (2012). Effect of chronic exposure to acetaminophen and lincomycin on Japanese medaka (Oryzias latipes) and freshwater cladocerans Daphnia magna and Moina macrocopa, and potential mechanisms of endocrine disruption. Chemosphere89, 10–18. 10.1016/j.chemosphere.2012.04.006
95
Kim S. Ji K. Shin H. Park S. Kho Y. Park K. et al (2020). Occurrences of benzalkonium chloride in streams near a pharmaceutical manufacturing complex in Korea and associated ecological risk. Chemosphere256, 127084. 10.1016/j.chemosphere.2020.127084
96
Kim S. Jung D. Kho Y. Choi K. (2014). Effects of benzophenone-3 exposure on endocrine disruption and reproduction of Japanese medaka (Oryzias latipes), a two generations exposure study. Aquat. Toxicol.155, 244–252. 10.1016/j.aquatox.2014.07.004
97
Kim S. Lee S. Kim C. Liu X. Seo J. Jung H. et al (2014a). In vitro and in vivo toxicities of sediment and surface water in an area near a major steel industry of Korea: endocrine disruption, reproduction, or survival effects combined with instrumental analysis. Sci. Total Environ.470-471, 1509–1516. 10.1016/j.scitotenv.2013.08.010
98
Kim S. D. Cho I. Kim I. S. Vanderford B. J. Synder S. A. (2007). Occurrence and removal of pharmaceuticals and endocrine disruptors in South Korean surface, drinking, and waste waters. Water Res.41, 1013–1021. 10.1016/j.watres.2006.06.034
99
Kiparissis Y. Balch G. G. Metcalfe T. I. Metcalfe G. D. (2003a). Effects of the isoflavone genistein and equol on the gonadal development of Japanese medaka, (Oryzias latipes). Environ. Health Perspect.111, 1158–1163. 10.1289/ehp.5928
100
Kiparissis Y. Metcalfe T. L. Balch G. C. Metcalfe G. D. (2003b). Effects of the antiandrogen, vinclozolin and cyproterone acetate on gonadal development in the Japanese medaka (Oryzias latipes). Aquat. Toxicol.63, 391–403. 10.1016/s0166-445x(02)00189-3
101
Knorr S. Braunbeck T. (2002). Decline in reproductive success, sex reversal, and developmental alterations in Japanese medaka (Oryzias latipes) after continuous exposure to octylphenol. Ecotoxicol. Environ. Saf.51, 187–196. 10.1006/eesa.2001.2123
102
Kobayashi K. Tamotsu S. Yasuda K. Oishi T. (2005). Vitellogenin immunohistochemistry in the liver and the testis of the medaka Oryzias latipes, exposed to 17 beta-estradiol and p-nonylphenol. Zool. Sci.22, 453–461.
103
Koger C. S. Teh S. J. Hinton D. E. (2000). Determining the sensitive developmental stages of intersex induction in medaka (Oryzias latipes) exposed to 17β-estradiol or testosterone. Mar. Environ. Res.50, 201–206. 10.1016/s0141-1136(00)00068-4
104
Kuhl A. J. Brouwer M. (2006). Antiestrogens inhibit xenoestrogen-induced brain aromatase activity but do not prevent xenoestrogen-induced feminization in Japanese medaka (Oryzias latipes). Environ. Health Persp.114, 500–506. 10.1289/ehp.8211
105
Kwak K. Ji K. Kho Y. Kim P. Lee J. Ryu J. et al (2018). Chronic toxicity and endocrine disruption of naproxen in freshwater waterfleas and fish, and steroidogenic alteration using H295R cell assay. Chemosphere204, 156–162. 10.1016/j.chemosphere.2018.04.035
106
LaLone C. A. Villeneuve D. L. Cavallin J. E. Kahl M. D. Durhan E. J. Makynen E. A. et al (2013). Cross species sensitivity to a novel androgen receptor agonist of potential environmental concern, spironolactone. Environ. Toxicol. Chem.32, 2528–2541. 10.1002/etc.2330
107
Lange A. Paull G. C. Coe T. S. Katsu Y. Urushitani H. Iguchi T. et al (2009). Sexual reprogramming and estrogenic sensitization in wild fish exposed to ethinylestradiol. Environ. Sci. Technol.43, 1219–1225. 10.1021/es802661p
108
Lee C. Na J. G. Lee K.-C. Park K. (2002). Choriogenin mRNA induction in male medaka, Oryzias latipes is a biomarker of endocrine disruption. Aqua. Tox.61, 233–241. 10.1016/s0166-445x(02)00060-7
109
Lee I. Lee J. Jung D. Kim S. Choi K. (2019b). Two-generation exposure to 2-ethylhexyl 4-methoxycinnamate (EHMC) in Japanese medaka (Oryzias latipes) and its reproduction and endocrine-related effects. Chemosphere228, 478–484. 10.1016/j.chemosphere.2019.04.123
110
Lee J. Ji K. Kho Y. L. Kim P. Choi K. (2011). Chronic exposure to diclofenac on two freshwater cladocerans and Japanese medaka. Ecotoxicol. Environ. Saf.74, 1216–1225. 10.1016/j.ecoenv.2011.03.014
111
Lee J. W. Lee J.-W. Kim K. Shin Y.-J. Kim J. Kim S. et al (2017b). PFOA-induced metabolism disturbance and multi-generational reproductive toxicity in Oryzias latipes. J. Hazard Mat.340, 231–240. 10.1016/j.jhazmat.2017.06.058
112
Lee J. W. Lee J. W. Shin Y. J. Kim J. E. Ryu T. K. Ryu J. et al (2017a). Multigenerational xenoestrogenic effects of perfluoroalkyl acids (PFAAs) mixture on Oryzias latipes using a flow through exposure system. Chemosphere169, 212–223. 10.1016/j.chemosphere.2016.11.035
113
Lee J. W. Shin Y.-J. Kim H. Kim H. Kim J. Min S.-A. et al (2019a). Metformin-induced endocrine disruption and oxidative stress of Oryzias latipes on two-generational condition. J. Haz. Mat.367, 171–181. 10.1016/j.jhazmat.2018.12.084
114
Lee S.-E. Young-Woong C. Mo H. Son J. Park K. Cho K. (2013). Endosulfan-induced biomarkers in Japanese rice fish (Oryzias latipes) analyzed by SELDI-TOF-MS. Int. J. Biol. Sci.9, 343–349. 10.7150/ijbs.5501
115
Lee W. Kang C.-W. Su C.-K. Okubo K. Nagahama Y. (2012). Screening estrogenic activity of environmental contaminants and water samples using a transgenic medaka embryo bioassay. Chemosphere88, 945–952. 10.1016/j.chemosphere.2012.03.024
116
Lee W. Kang C.-W. Su C.-K. Okubo K. Nagahama Y. (2014). Effects of water temperature on perchlorate toxicity to the thyroid and reproductive system of Oryzias latipes. Ecotoxicol. Environ. Safe108, 311–317. 10.1016/j.ecoenv.2014.07.016
117
Lee Pow C. S. D. Tilahun K. Creech K. Law J. M. Cope W. G. Kwak T. J. et al (2017). Windows of susceptibility and consequences of early life exposures to 17β-estradiol on medaka (Oryzias latipes) reproductive success. Environ. Sci. Tech.51, 5296–5305. 10.1021/acs.est.7b01568
118
Lei B. Kang J. Yu Y. Zha J. Li W. Wang Z. (2013). Β-Estradiol 17 valerate affects embryonic development and sexual differentiation in Japanese medaka (Oryzias latipes). Aquat. Toxicol.160, 128–134. 10.1016/j.aquatox.2013.03.011
119
Lei B. Peng W. Li W. Yu Y. Xu J. Wang Y. (2016). Diethylstilbestrol at environmental levels affects the development of early life stage and target gene expression in Japanese medaka (Oryzias latipes). Ecotoxicol25, 563–573. 10.1007/s10646-016-1615-0
120
Leon A. The S. J. Hall L. C. The F. C. (2007). Androgen disruption of early development in quart strain medaka (Oryzias latipes). Aquat. Toxicol.82, 195–203. 10.1016/j.aquatox.2007.02.012
121
Leon A. Wu P.-S. Hall L. C. Johnson M. L. Teh S. J. (2008). Global gene expression profiling of androgen disruption in Qurt strain medaka. Environ. Sci. Technol.42, 962–969. 10.1021/es071785c
122
Li D. Chen Q. Cao J. Chen H. Li L. Cedergreen N. et al (2016). The chronic effects of lignin-derived bisphenol and bisphenol-A in Japanese medaka Oryzias latipes. Aquat. Toxicol.170, 199–207. 10.1016/j.aquatox.2015.11.024
123
Li D. Ren B. Chen H. Mu L. Zhang L. Chen Q. et al (2017). The acute toxicity of bisphenol A and lignin-derived bisphenol in algae, daphnids, and Japanese medaka. Environ. Sci. Pollut. Res.24 (30), 23872–23879. 10.1007/s11356-017-0018-y
124
Li Y. Chen R. He J. Ma H. Zhao F. Tao S. et al (2019). Triphenyl phosphate at environmental levels retarded ovary development and reduced egg production in Japanese medaka (Oryzias latipes). Environ. Sci. Technol.53, 14709–14715. 10.1021/acs.est.9b05669
125
Liang M. Yan S. Chen R. Hong X. Zha J. (2020). 3-(4-Methylbenzylidene) camphor induced reproduction toxicity and antiandrogenicity in Japanese medaka (Oryzias latipes). Chemosphere249, 126224. 10.1016/j.chemosphere.2020.126224
126
Liao P. H. Chu S. H. Tu T. Y. Wang X. H. Lin A. Y. Chen P. J. (2014). Persistent endocrine disruption effects in medaka fish with early life-stage exposure to a triazole containing aromatase inhibitor (letrozole). J. Hazard Mat.277, 141–149. 10.1016/j.jhazmat.2014.02.013
127
Lin C.-H. Chou P.-H. Chen P.-J. (2014). Two azole fungicides (carcinogenic triadimefon and non-carcinogenic myclobutanil) exhibit different hepatic cytochrome P450 activities in medaka fish. J. Hazard Mat.277, 150–158. 10.1016/j.jhazmat.2014.05.083
128
Liu N. Jin X. Zhou J. Wang Y. Yang Q. Wu F. et al (2018). Predicted no-effect concentration (PNEC) and assessment of risk for the fungicide, triadimefon based on reproductive fitness of aquatic organisms. Chemosphere207, 682–689. 10.1016/j.chemosphere.2018.05.093
129
Martyniuk C. J. Martinez R. Navarro-Martin L. Kamstra J. H. Schwendt A. Reynaud S. et al (2022). Emerging concepts and opportunities for endocrine disruptor screening of the non-EATS modalities. Environ. Res.204, 111904. 10.1016/j.envres.2021.111904
130
Matten S. Fallacara D. Kamel A. Lynn S. G. Fort D. J. Wolf J. C. et al (2023). Evaluation of multigenerational effects of 2-ethylhexyl 4-hydroxybenzoate in Japanese medaka. J. Appl. Toxicol.43, 1645–1666. 10.1002/jat.4502
131
Mazukami-Murata S. Kishi-Kadota K. Nishida T. (2016). 17β-trenbolone exposure programs metabolic dysfunction in larval medaka. Environ. Toxicol.31, 1539–1551. 10.1002/tox.22158
132
Metcalfe C. D. Metcalfe T. L. Kiparessis Y. Koenig B. G. Khan C. Hughes R. J. et al (2001). Estrogenic potency of chemicals detected in sewage treatment plant effluents as determined by in vivo assays with Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem.20, 297–308. 10.1897/1551-5028(2001)020<0297:epocdi>2.0.co;2
133
Mihaich E. Capdevielle M. Urbach-Ross D. Gallagher S. Wolf J. (2019). Medaka (Oryzias latipes) multigeneration test with triclosan. Environ. Toxicol. Chem.38, 1770–1783. 10.1002/etc.4451
134
Miyagawa S. Lange A. Hirakawa I. Tohyama S. Ogino Y. Mizutani T. et al (2014). Differing species responsiveness of estrogenic contaminants in fish is conferred by the ligand binding domain of the estrogen receptor. Environ. Sci. Technol.48, 5254–5263. 10.1021/es5002659
135
Myla A. Dasmahapatra A. K. Tchounwou P. B. (2021a). Sex reversal and Histopathological assessment of potential endocrine disrupting effects of graphene oxide on Japanese medaka (Oryzias latipes) larvae. Chemosphere279, 130768. 10.1016/j.chemosphere.2021.130768
136
Myla M. Dasmahapatra A. K. Tchounwou P. B. (2021b). Experimental data sets on Sex-Reversal and histopathological assessment of potential endocrine-disrupting effects of graphene oxide on Japanese medaka (Oryzias latipes) larvae at the onset of maturity. Data. Brief.39, 107330. 10.1016/j.dib.2021.107330
137
Myosho T. Ishibashi A. Fujimoto S. Miyagawa S. Iguchi T. Kobayashi T. (2022). Preself-feeding medaka fry provides a suitable screening system for in vivo assessment of thyroid hormone-disrupting potential. Environ. Sci. Technol.56, 6479–6490. 10.1021/acs.est.1c06729
138
Myosho T. Sato T. Nishiyama H. Watanabe A. Yamamoto J. Okamura T. et al (2019). Inter- and intraspecific variation in sex hormone-induced sex-reversal in medaka, Oryzias latipes and Oryzias sakaizumii. Zoll Sci.36, 425–431. 10.2108/zs180194
139
Nair R. U. Victor A. C. Paul V. Paul-Prasanth (2017). Effects of N-Nitrosodiethylamine, a potent carcinogen, on sexual development, gametogenesis, and oocyte maturation. Sex. Dev.11, 161–167. 10.1159/000477106
140
Nakamura A. Takanobu H. Tamura I. Yamamuro M. Iguchi T. Tatarazako N. (2014b). Verification of responses of Japanese medaka (Oryzias latipes) to anti-androgens, vinclozolin and flutamide, in short-term assays. J. Appl. Toxicol.34, 545–553. 10.1002/jat.2934
141
Nakamura A. Tamura I. Takanobu H. Yamamuro M. Iguchi T. Tatarazako N. (2014a). Fish multigeneration test with preliminary short-term reproduction assay for estrone using Japanese medaka (Oryzias latipes). J. Appl. Toxicol.35, 11–13. 10.1002/jat.2981
142
Nakayama K. Sei N. Hqandoh I. C. Shimasaki Y. Honjo T. Oshima Y. (2011). Effects of polychlorinated biphenyls on liver function and sexual characteristics in Japanese medaka (Oryzias latipes). Mar. Poll. Bull.63, 366–369. 10.1016/j.marpolbul.2011.01.015
143
NIEHS (2018). Endocrine disruptors. Available at: http://niehs.nih.gov/health/topics/agents/endocrine/index.cfm (Accessed December 28, 2018).
144
Nimrod A. C. Benson W. H. (1998). Reproduction and development of Japanese medaka following an early life stage exposure to xenoestrogens. Aqat. Toxicol.44, 141–156. 10.1016/s0166-445x(98)00062-9
145
Nozaka T. Abe T. Matsura T. Sakamoto T. Nakano N. Maeda M. et al (2004). Development of vitellogenin assay for endocrine disruption using medaka (Oryzias latipes). Environ. Sci.11 (2), 99–121.
146
OECD (2018). Revised guidance document 150 on standardized test guidelines for evaluating chemicals for endocrine disruption. Paris: OECD publishing. 10.1787/9789264304741.8.en
147
Ogino Y. Hirakawa I. Inohaya K. Sumiya E. Miyagawa S. Denslow N. et al (2014). Bmp7 and lef1 are the downstream effectors of androgen signaling in androgen-induced sex characteristics development in medaka. Endocrinology155, 449–462. 10.1210/en.2013-1507
148
Onishi Y. Tatarazako N. Koshio M. Okamura T. Watanabe H. Sawai A. et al (2021). Summary of reference chemicals evaluated by the fish short-term reproduction assay, OECD TG229, Using Japanese medaka, Oryzias latipes. J. Appl. Toxicol.41, 1200–1221. 10.1002/jat.4104
149
Orn S. Yamani S. Norrgren L. (2006). Comparison of vitellogenin induction, sex ratio, and gonad morphology between zebrafish and Japanese medaka after exposure to 17alpha-ethinylestradiol and 17beta-trenbolone. Arch. Environ. Contam. Toxicol.51, 237–243. 10.1007/s00244-005-0103-y
150
Oshima Y. Kang I. K. Kobayashi M. Nakayama K. Imada N. Honjo T. (2003). Suppression of sexual behavior in male Japanese medaka (Oryzias latipes) exposed to 17β-estradiol. Cemosphere50, 429–436. 10.1016/s0045-6535(02)00494-0
151
Pandelides Z. Ussery E. J. Overturf M. D. Guchardi J. Holdway D. A. (2021). Inhibition of swim bladder inflation in Japanese medaka (Oryzias latipes) embryos following exposure to select pharmaceuticals alone and in combination. Aqua Tox234, 105796. 10.1016/j.aquatox.2021.105796
152
Papoulias D. M. Noltie D. B. Tillit D. E. (2000). Effects of methyl testosterone exposure on sexual differentiation in medaka, Oryzias latipes. Mar. Environ. Res.51, 181–184. 10.1016/s0141-1136(00)00076-3
153
Park J. W. Tompsett A. R. Zhang X. Newsted J. L. Jones P. D. Au D. W. T. et al (2008). Flourescence in situ hybridization techniques (FISH) to detect changes in CYP19a gene expression of Japanese medaka (Oryzias latipes). Toxicol. Appl. Pharmacol.232, 226–235. 10.1016/j.taap.2008.06.012
154
Park J. W. Tompsett A. R. Zhang X. Newsted J. L. Jones P. D. Au D. W. T. et al (2009). Advanced fluorescence in situ hybridization to localize and quantify gene expression in Japanese medaka (Oryzias latipes) exposed to endocrine-disrupting compounds. Environ. Toxicol. Chem.28, 1951–1962. 10.1897/08-574.1
155
Patyna P. J. Davi R. A. Parkerton T. F. Brown R. P. Cooper K. R. (1999). A proposed multigeneration protocol for Japanese medaka (Oryzias latipes) to evaluate effects of endocrine disruptors. Sci. Total Environ.233, 211–220. 10.1016/s0048-9697(99)00227-2
156
Paul V. Krishnakumar S. Gowd G. S. Nair S. V. Koyakutty M. Paul-Prasanth B. (2021). Sex-dependent bioaccumulation of nano zinc oxide and its adverse effects on sexual behavior and reproduction in Japanese medaka. ACS Ppl. Bio. Mat.4, 7408–7421. 10.1021/acsabm.1c00575
157
Powe D. K. Dasmahapatra A. K. Russel J. L. Tchounwou P. B. (2018). Toxicity implications for early life stage Japanese medaka (Oryzias latipes) exposed to oxyfluorfen. Environ. Toxicol.33, 555–568. 10.1002/tox.22541
158
Richter C. A. Papoulias D. M. Whyte J. J. Tillitt D. E. (2016). Evaluation of potential mechanisms of atrazine-induced reproductive impairment in fathead minnow (Pimephales promelas) and Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem.35, 2230–2238. 10.1002/etc.3376
159
Robinson J. A. Staveley J. P. Constantine L. (2017). Reproductive effects on freshwater fish exposed to 17α-trenbolone and 17α-estradiol. Environ. Toxicol. Chem.36, 636–644. 10.1002/etc.3526
160
Saunders D. M. V. Podaima M. Wiseman S. Giesy J. P. (2015). Effects of the brominated flame retardant TBCO on fecundity and profiles of transcripts of the HPGL-axis in Japanese medaka. Toxicol160, 180–187. 10.1016/j.aquatox.2015.01.018
161
Schiller V. Wichmann A. Kriehuber R. Muth-Kohne E. Giesy J. P. Hecker M. et al (2013). Studying the effects of genistein on gene expression of fish embryos as an alternative testing approach for endocrine disruption. Comp. Biochem. Physiol. Part C157, 41–53. 10.1016/j.cbpc.2012.09.005
162
Schiller V. Zhang X. Hecker M. Schafers C. Fischer R. Fenske M. (2014). Species-specific considerations in using the fish embryo test as an alternative to identify endocrine disruption. Aqut. Toxicol.155, 62–72. 10.1016/j.aquatox.2014.06.005
163
Scholz S. Gutzeit H. O. (2000). 17-alpha-ethinylestradiol affects reproduction, sexual differentiation and aromatase gene expression of the medaka (Oryzias latipes). Aquat. Toxicol.50, 363–373. 10.1016/s0166-445x(00)00090-4
164
Scholz S. Mayer I. (2008). Molecular biomarkers of endocrine disruption in small moel fish. Mol. Cell Endocrinol.293, 52–70. 10.1016/j.mce.200.06.008
165
Scholz S. Mayers I. (2008). Molecular biomarkers of endocrine disruption in small model fish. Mol. Cell Endocrinol.293, 57–70. 10.1016/j.mce.2008.06.008
166
Seki M. Fujishima S. Nozaka T. Maeda M. Kobayashi K. (2006). Comparison of response to 17β-estradiol and 17β-trenbolone among three small fish species. Environ. Toxicol. Chem.22, 2742–2752. 10.1897/05-647r.1
167
Seki M. Yokota H. Maeda M. Tadokoro H. Kobayashi K. (2003a). Effects of 4-nonylphenol and 4-tert-octylphenol on sex differentiation and vitellogenin induction in medaka (Oryzias latipes). Environ. Toxicol. Chem.22, 1507–1516. 10.1897/1551-5028(2003)22<1507:eonato>2.0.co;2
168
Seki M. Yokota H. Matsubara H. Maeda M. Tadokoro H. Kobayashi K. (2003b). Fish full life cycle testing for the weak estrogen 4-tert-pentylphenol on medaka (Oryzias latipes). Environ. Toxicol. Chem.22, 1487–1496. 10.1897/1551-5028(2003)22<1487:ffltft>2.0.co;2
169
Seki M. Yokota H. Matsubara H. Maeda M. Tadokoro H. Kobayashi K. (2004). Fish full life cycle testing for androgen methyltestosterone on medaka (Oryzias latipes). Environ. Toxicol. Chem.23, 774–781. 10.1897/03-26
170
Seki M. Yokota H. Matsubara H. Tsuruda Y. Maeda M. Tadokoro H. et al (2002). Effects of ethinylestradiol on the reproduction and induction of vitellogenin and testis-ova in medaka (Oryzias latipes). Environ. Toxicol. Chem.21, 1692–1698. 10.1897/1551-5028(2002)021<1692:eoeotr>2.0.co;2
171
Shioda T. Wakabayashi M. (2000). Effect of certain chemicals on the reproduction of medaka (Oryzias latipes). Chemosphere40, 239–243. 10.1016/s0045-6535(99)00235-0
172
Smith C. M. vera M. K. M. Bhandari R. K. (2019). Developmental and epigenetic effects of roundup and glyphosate exposure on Japanese medaka (Oryzias latipes). Aquat. Toxicol.210, 215–226. 10.1016/j.aquatox.2019.03.005
173
Song X. Wang X. Bhandari R. K. (2020). Developmental abnormalities and epigenetic alterations in medaka (Oryzias latipes) embryos induced by triclosan exposure. Chemosphere261, 127613. 10.1016/j.chemosphere.2020.127613
174
Spirhanzlova P. Groef B. D. Nicholson F. E. Grommen S. V. H. Marras G. Sebillot A. et al (2017). Using short-term bioassays to evaluate the endocrine disrupting capacity of the pesticides linuron and fenoxycarb. Comp. Biochem. Physiol. part C200, 52–58. 10.1016/j.cbpc.2017.06.006
175
Spirhanzlova P. Trebulle P. Lallement J. Sebillot A. Kanamori A. Lemkine G. F. et al (2020). Transgenic medaka identify embryonic periods sensitive to disruption of sex determination. Environ. Toxicol. Chem.39, 842–851. 10.1002/etc.4674
176
Su M. Zhong Y. Xiang J. Chen Y. Liu N. Zhang J. (2023). Reproductive endocrine disruption and gonadal intersex induction in male Japanese medaka chronically exposed to betamethasone at environmentally relevant levels. J. Hazard Mat.455, 131493. 10.1016/j.jhazmat.2023.131493
177
Sun I. Zha J. Spear P. A. Wang Z. (2007a). Tamoxifen effects on the early life stagesand reproduction of Japanese medaka (Oryzias latipes). Environ. Toxicol. Pharmacol.24, 23–29. 10.1016/j.etap.2007.01.003
178
Sun I. Zha J. Spear P. A. Wang Z. (2007b). Toxicity of the aromatase inhibitor letrozole to Japanese medaka (Oryzias latipes) eggs, larvae, and breeding adults. Comp. Biochem. Physiol. C Toxicol. Pharmacol.145, 533–541. 10.1016/j.cbpc.2007.01.017
179
Sun J. Tang S. Peng H. Saunders D. M. V. Doering J. A. Hecker M. et al (2016c). Combined transcriptomic approach to identify toxicity pathways in early life stages of Japanese medaka (Oryzias latipes) exposed to 1,2,5,6-tetrabromocyclooctane (TBCO). Environ. Sci. Technol.50, 7781–7790. 10.1021/acs.est.6b01249
180
Sun L. Jin R. Peng Z. Zhou Q. Qian H. Fu Z. (2014). Effects of trilostane and fipronil on the reproductive axis in an early life stage of the Japanese medaka (Oryzias latipes). Ecotoxicology23, 1044–1054. 10.1007/s10646-014-1248-0
181
Sun L. Peng T. Liu F. Ren L. Peng Z. Ji G. et al (2016b). Transcriptional responses in male Japanese medaka exposed to antiandrogens and antiandrogen/androgen mixtures. Environ. Toxicol.31, 1591–1599. 10.1002/tox.22163
182
Sun L. Shao X. Chi J. Hu X. Jin Y. Fu Z. (2011a). Transcriptional responses in the brain, liver, and gonad of Japanese ricefish (Oryzias latipes) exposed to two anti-estrogens. Comp. Biochem. Physiol. part C153, 392–401. 10.1016/j.cbpc.2011.01.003
183
Sun L. Shao X. Hu X. Chi J. Jin Y. Ye W. et al (2011b). Transcriptional responses in Japanese medaka (Oryzias latipes) exposed to binary mixtures of an estrogen and antiestrogen. Aqut. Toxicol.105, 629–639. 10.1016/j.aquatox.2011.08.024
184
Sun L. Wang S. Lin X. Tan H. Fu Z. (2016a). Early life exposure to ractopamine causes endocrine disrupting effects in Japanese medaka (Oryzias latipes). Bull. Environ. Contam. Toxicol.96, 150–155. 10.1007/s00128-015-1659-5
185
Sun L. Zha J. Wang Z. (2009). Effects of binary mixtures of estrogen and antiestrogens on Japanese medaka (Oryzias latipes). Qut. Toxicol.93, 83–89. 10.1016/j.aquatox.2009.03.010
186
Suzuki A. Tanaka M. Shibata N. Nagahama Y. (2004). Expression of aromatase mRNA and effects of aromatase inhibitor during ovarian development in the medaka, Oryzias latipes. J. Exp. Zool.301A, 266–273. 10.1002/jez.a.20027
187
Tabata A. Kashiwada S. Ohnishi Y. Ishikawa H. Miyamoto N. Itoh M. et al (2001). Estrogenic influences of estradiol-17β, p-nonylphenol and bis-phenol-A on Japanese medaka (Oryzias latipes) at detected environmental concentrations. Water Sci. Tech.43, 109–116. 10.2166/wst.2001.0079
188
Tabata A. Watanabe N. Yamamoto I. Ohnishi Y. Itoh M. Kamel T. et al (2004). The effect of Bisphenol A and chlorinated derivatives of bisphenol A on the level of serum vitellogenin in Japanese medaka (Oryzias latipes). Water Sci. Technol.50, 125–132. 10.2166/wst.2004.0319
189
Tagawa M. Hirano T. (1991). Effects of thyroid hormone deficiency in eggs on early development of the medaka, Oryzias latipes. J. Exp. Zool.257, 360–366. 10.1002/jez.1402570309
190
Teather K. Jardine C. Gormley K. (2005). Behavioral and sex ratio modification of Japanese medaka (Oryzias latipes) in response to environmentally relevant mixtures of three pesticides. Environ. Toxicol.20, 110–117. 10.1002/tox.20084
191
Ternes T. Stumpf M. Mueller J. Haberer K. Wilken R.-D. Servos M. (1999). Behavior and occurrence of estrogens in municipal sewage treatment plants-I. Investigations in Germany, Canada, and Brazil. Sci. Total Environ.225, 81–90. 10.1016/s0048-9697(98)00334-9
192
Thayil A. J. Wang X. Bhandari P. vom Saal F. S. Tillitt D. E. Bhandari R. J. (2020). Bisphenol A and 17α-ethinylestradiol-induced transgenerational gene expression differences in the brain-pituitary-testis axis of medaka, Oryzias latipes. Biol. Reprod.103, 1324–1335. 10.1093/biolre/ioaa169
193
Thresher R. Gurney R. Canning M. (2011). Effects of lifetime chemical inhibition of aromatase on the sexual differentiation, sperm characteristics, and fertility of medaka (Oryzias latipes) and zebrafish (Danio rerio). Aquat. Toxicol.105, 355–360. 10.1016/j.aquatox.2011.07.008
194
Tilton S. C. Foran C. M. Benson W. H. (2003). Effects of cadmium on the reproductive axis of Japanese medaka (Oryzias latipes). Comp. Biochem. Physiol. Part C136, 265–276. 10.1016/j.cca.2003.09.009
195
Tohyama S. Miyagawa S. Lange A. Ogino Y. Mizutani T. Tatarazako N. et al (2015). Understanding the molecular basis for differences in responses of fish estrogen receptor subtypes to environmental estrogens. Environ. Sci. Tech.49, 7439–7447. 10.1021/acs.est.5b00704
196
Uchida M. Nakamura H. Kagami Y. Kusano T. Arizono K. (2010). Estrogenic effects of o,p’-DDT exposure in Japanese medaka (Oryzias latipes). J. Toxicol. Sci.35, 605–608. 10.2131/jts.35.605
197
Urushitani H. Katsu Y. Kato Y. Tooi O. Santo N. Kawashima Y. et al (2007). Medaka (Oryzias latipes) for use in evaluating developmental effects of endocrine active chemicals with special reference to gonadal development. Envirn. Sci.14, 211–233.
198
Wagner S. D. Kurobe T. Hammock B. G. Lam C. H. Wu G. Vasylieva N. et al (2017). Developmental effects of fipronil on Japanese medaka (Oryzias latipes) embryos. Chemosphere166, 511–520. 10.1016/j.chemosphere.2016.09.069
199
Wang C. Zhang S. Zhou Y. Huang C. Mu D. Giesy J. P. et al (2016). Equal induces gonadal intersex in Japanese medaka (Oryzias latipes) at environmentally relevant concentrations: comparison with 17β-estradiol. Environ. Sci. Technol.50, 7852–7860. 10.1021/acs.est.6b02211
200
Watanabe A. Myosho T. Ishibashi A. Yamamoto J. Toda M. Onishi Y. et al (2023). Levonorgestrel causes feminization and dose-dependent masculinization in medaka fish (Oryzias latipes): endocrine-disruption activity and its correlation with sex reversal. Sci. Total Environ.876, 162740. 10.1016/j.scitotenv.2023.162740
201
Watanabe H. Horie Y. Takanobu H. Koshio M. Flynn K. Iguchi T. et al (2017). Medaka extended one-generation reproduction test evaluating 4-nonylphenol. Environ. Toxicol. Chem.36, 3254–3266. 10.1002/etc.3895
202
Yamamoto H. Tamura I. Hirata Y. Kato J. Kagota K. Katsuki S. et al (2011). Aquatic toxicity and ecological risk assessment of seven parabens: individual and additive approach. Sci. Total Environ.410-411, 102–111. 10.1016/j.scitotenv.2011.09.040
203
Yamamoto H. Watanabe M. Hirata Y. Nakamura Y. Nakamura Y. Kitani C. et al (2007). Preliminary ecological risk assessment of Butylparaben and benzylparaben-1. Removal efficiency in wastewater treatment, acute/chronic toxicity for aquatic organisms, and effects on medaka gene expression. Environ. Sci.14, 73–87.
204
Yan S. Liang M. Chen R. Hong X. Zha J. (2020). Reproductive toxicity and estrogen activity in Japanese medaka (Oryzias latipes) exposed to environmentally relevant concentrations of octocrylene. Environ. Pol.261, 114104. 10.1016/j.envpol.2020.114104
205
Yokota H. Abe T. Nakai M. Murakami H. Eto C. Yakabe Y. (2005). Effects of 4-tert-pentylphenol on the gene expression of p450 11β-hydroxylase in the gonad of medaka (Oryzias latipes). Auat Toxicol.71, 121–132. 10.1016/j.aquatox.2004.10.017
206
Yokota H. Higashi K. Hanada E. Matsuzaki E. Tsuruda Y. Suzuki T. et al (2017). Recovery from reproductive and morphological abnormalities in medaka (Oryzias latipes) following a 14-day exposure to diclofenac. Environ. Toxicol. Chem.36, 3277–3283. 10.1002/etc.3899
207
Yokota H. Seki M. Maeda M. Oshima Y. Tadokoro H. Honjo T. et al (2001). Life cycle toxicity of 4-nonylphenol to medaka (Oryzias latipes). Environ. Toxicol. Chem.20, 2552–2560. 10.1897/1551-5028(2001)020<2552:lctont>2.0.co;2
208
Yokota H. Taguchi Y. Tanaka Y. Uchiyama M. Kondo M. Tsuruda Y. et al (2018). Chronic exposure to diclofenac induced delayed mandibular defects in medaka (Oryzias latipes) in a sex-dependent manner. Chemosphere210, 139–146. 10.1016/j.chemosphere.2018.07.016
209
Yokota H. Tsuruda Y. Maeda M. Oshima Y. Tadokoro H. Nakazono A. et al (2000). Effect of bisphenol A on the early life stage in Japanese medaka (Oryzias latipes). Environ. Toxicol. Chem.19, 1925–1930. 10.1897/1551-5028(2000)019<1925:eobaot>2.3.co;2
210
Yum S. Woo S. Kagami Y. Park H.-S. Ryu J.-C. (2010). Changes in gene expression profile of medaka with acute toxicity of Arochlor 1260, a polychlorinated biphenyl mixture. Comp. Biochem. Physiol.151C, 51–56. 10.1016/j.cbpc.2009.08.007
211
Zeng Z. Shan T. Tong Y. Lam S. H. Gong Z. (2005). Development of estrogen-responsive transgenic medaka for environmental monitoring of endocrine disrupters. Environ. Sci. Technol.39, 9001–9008. 10.1021/es050728l
212
Zha J. Wang Z. Schlenk D. (2006). Effects of pentachlorophenol on the reproduction of Japanese medaka (Oryzias latipes). Chem. Biol. Int.161, 26–36. 10.1016/j.cbi.2006.02.010
213
Zhang X. Hecker M. Jones P. D. Newsted J. Au D. Kong R. et al (2008a). Responses of the medaka HPG axis PCR array and reproduction to prochloraz and ketoconazole. Environ. Sci. Technol.42, 6762–6769. 10.1021/es800591t
214
Zhang X. Hecker M. Park J.-W. Tompsett A. R. Jones P. D. Newsted J. et al (2008b). Time-dependent transcriptional profiles of genes of the hypothalamic-pituitary-gonadal axis in medaka (Oryzias latipes) exposed to Fadrozole and 17β-trenbolone. Environ. Toxicol. Chem.27, 2504–2511. 10.1897/08-082.1
215
Zhang X. Hecker M. Park J.-W. Tompsett A. R. Newsted J. Nakayama K. et al (2008c). Real-time PCR array to study effects of chemicals on the hypothalamic-Pituitary-Gonadal axis of the Japanese medaka. Aqua. Toxicol.88, 173–182. 10.1016/j.aquatox.2008.04.009
216
Zhang Z. Hu J. Zhen H. Wu X. Huang C. (2008e). Reproductive inhibition and transgenerational toxicity of triphenyltin on medaka (Oryzias latipes) at environmentally relevant levels. Environ. Sci. Tech.42, 8133–8139. 10.1021/es801573x
217
Zhang Z. Hu Y. (2008). Effects of p,p′-DDE exposure in gonadal development and gene expression in Japanese medaka (Oryzias latipes). J. Environ. Sci.20, 347–352. 10.1016/s1001-0742(08)60054-6
218
Zhang Z.-B. Hu J.-Y. Sai S.-X. Zhao Y.-B. Huang C. Tian X.-J. (2008d). Gene cloning, sequence analysis and tissue expression of estrogen-related receptor alpha (Erralpha) in Japanese medaka and its transcriptional responses after differential EDCs exposure. Huan Jing Ke Xue29, 3153–3158. (article in Chinese].
219
Zhu L. Wang H. Liu H. Li W. (2015). Effects of trifloxystrobin on hatching, survival, and gene expression of endocrine biomarkers in early life stages of medaka (Oryzias latipes). Environ. Toxicol.30, 648–655. 10.1002/tox.21942
Summary
Keywords
Japanese medaka, endocrine disruptors, EATS pathways, systematic review, risk assessment
Citation
Dasmahapatra AK, Williams CB, Myla A, Tiwary SK and Tchounwou PB (2023) A systematic review of the evaluation of endocrine-disrupting chemicals in the Japanese medaka (Oryzias latipes) fish. Front. Toxicol. 5:1272368. doi: 10.3389/ftox.2023.1272368
Received
04 August 2023
Accepted
10 October 2023
Published
27 November 2023
Volume
5 - 2023
Edited by
Rosaria Meccariello, University of Naples Parthenope, Italy
Reviewed by
Massimo Venditti, Second University of Naples, Italy
Azza Naija, Qatar University, Qatar
Updates
Copyright
© 2023 Dasmahapatra, Williams, Myla, Tiwary and Tchounwou.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paul. B. Tchounwou, paul.tchounwou@morgan.edu
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.