 Natsu Sugama1
Natsu Sugama1 Emily T. Lew1Camilo Riquelme-Guzmán1
Emily T. Lew1Camilo Riquelme-Guzmán1 Di Sheng Lee1
Di Sheng Lee1 Xinxin Li1John S. K. Yuen2,3Taehwan Lim1Anson Kwan1Run Yi Liu4Yoshene A. Ma4
Xinxin Li1John S. K. Yuen2,3Taehwan Lim1Anson Kwan1Run Yi Liu4Yoshene A. Ma4 Scott C. Frost4
Scott C. Frost4 David L. Kaplan1*
David L. Kaplan1*- 1Tufts University School of Engineering, Medford, MA, United States
- 2Tufts University Cellular Agriculture Commercialization Laboratory, Boston, MA, United States
- 3Deco Labs, Inc., Boston, MA, United States
- 4Tufts University School of Arts and Sciences, Medford, MA, United States
Cultivated meat is emerging as a novel food source with the potential to contribute to a more sustainable and ethical food production system. However, limited research to date has explored the extent to which the nutrition and the aroma of such foods can be altered through cell culture conditions. Here, we aimed to modulate the aromatic volatile compounds in heated porcine cultivated fat cells by manipulating the media components while ensuring the preservation of robust fat differentiation. Using dynamic headspace gas chromatography–mass spectrometry (DHS-GC–MS), we demonstrated that supplementing cells with thiamine-HCl increased its intracellular concentration and promoted the production of 4-methyl-5-thiazoleethanol, contributing to milky aroma. Similarly, supplementation with L-methionine enhanced its intracellular concentration and increased the production of methional, a volatile compound with a potato-like aroma. Additionally, myoglobin significantly altered the volatile organic compound profile of cultivated fat. Notably, the concentration of γ-nonalactone, (E, E)-2,4-decadienal and 2-pentylfuran were increased, which contribute to a coconut-like, deep fat, fruity aroma, respectively, as well as elevated levels of other alcohols, aldehydes and furans. These findings highlight the potential of culture media formulations to modulate the aroma in cultivated fat production, a unique opportunity to optimize sensory features using this novel food production technology.
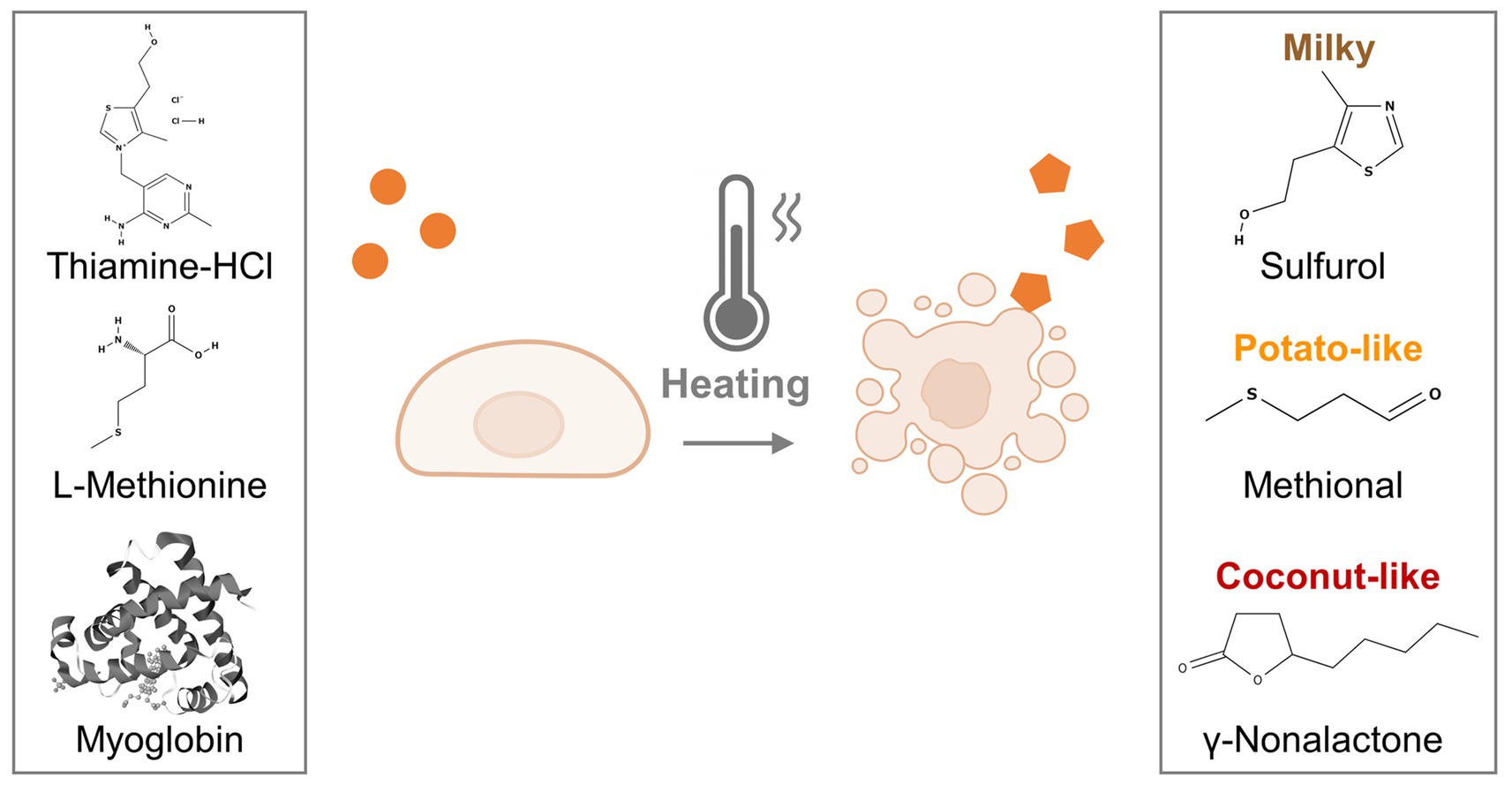
Graphical Abstract. Graphical overview of the methodology. Porcine dedifferentiated fat cells (pDFAT) were differentiated into adipocytes using adipogenesis media supplemented with aroma precursors. The cells were heated (cooked) and the resulting volatile compounds were analyzed using dynamic headspace gas chromatography–mass spectrometry (DHS-GC–MS).
Highlights
• Nutrient composition and aroma profiles of cultivated pork fat upon baking were modulated by cell culture media supplementation.
• Supplementing with thiamine-HCl, L-methionine, or myoglobin increased intracellular levels of thiamine or methionine and modulated the formation of aroma volatiles, enhancing characteristic odors such as milky, potato-like, and coconut-like notes.
1 Introduction
Cultivated meat has emerged as a promising technology to produce meat sustainably, with a significantly reduced risk of infectious diseases along with potentially improved nutrition (1, 2). A cornerstone for the success of this technology is flavor, which is a significant factor influencing consumer acceptability and purchasing decisions for meat products (3–6). Nevertheless, cultivated meat is still in its early stages of development, and many claims remain speculative rather than established facts (7). With respect to flavor, cultivated meat may differ substantially from conventional meat, particularly in its amino acid and nucleotide composition (8), however, research on its sensory characteristics remains limited (9). In terms of flavor, fat plays a crucial role in retaining aroma compounds and contributing to the persistence of scent (10, 11). Additionally, lipid oxidation products, in combination with Maillard compounds, produce a wide variety of aroma compounds in cooked meat (12–15).
Recently, several studies have reported on the aroma profiles of cultivated fat and muscle (16–18). For example, we have characterized volatile organic compounds (VOCs) released during the cooking of cultivated fat derived from porcine dedifferentiated fat cells (pDFAT) (16). Our study revealed the presence of fatty aldehydes such as pentanal, hexanal, octanal, and nonanal which contributed to fatty and buttery aromas. Sensory evaluation showed no statistical difference in response to the cooked cultivated pork fat when compared to traditional livestock-derived pork fat. Another study analyzed the VOCs of porcine adipocytes derived from porcine adipose-derived stem cells (ADSCs) cultured with scaffolds and found that both conventional and cultivated fat shared multiple common VOCs (17). A separate study revealed that porcine fibroblasts and myoblasts cultured in 10 and 15% serum-containing media exhibited significantly higher concentration of thiophenes which impart a meaty aroma than those cells maintained with 1% serum-containing media (18). These results suggested that cultivated meat and fat contain aroma volatiles similar to those found in livestock grown meat and fat, although some differences are present. Additionally, the media composition may influence the types and quantities of VOC profiles produced by baked cells.
In meat science, changing the composition of animal feed can modify VOC profiles, leading to changes in flavor (19–21). Diets with low protein and well-balanced essential amino acids significantly increased the level of 2-heptanone, which has a fruity smell, and 2,3-octanedione, which imparts the characteristic aroma of pork (21). Additionally, post-harvest treatment of cooked ham with thiamine can increase the concentration of 2-methyl-3-furanthiol and bis(2-methyl-3-furyl)-disulfide, which showed a significant difference in taste tests. These results demonstrated that the flavor of meat can be modified by adding certain nutrients to the diet of livestock. It is also possible to change the aroma during secondary processing after harvesting from the animal for processed meats like ham. However, to provoke these types of aroma changes in meat, 60 to 70 days of feeding is required (21, 22). Furthermore, the addition of aroma post-harvest requires secondary processes such as curing, and the presence of nitrites or nitrates used in processed meats can impact the changes in aroma (23). An important potential advantage of cultivated meat technology is in the ability to tailor aroma and nutritional content during cell cultivation due to the direct access of the media to the cells (24). Despite this potential, no research to date has specifically targeted the regulation of volatile aroma compounds in cultivated meat or fat through optimization of media composition.
The pathways for VOC generation in meat can be classified into the Maillard reaction, thiamine degradation, lipid oxidation, and Maillard-lipid interactions (25, 26). To address these pathways, three additives were studied: (1) thiamine plays a critical role as a coenzyme in carbohydrate metabolism and neural function, and its deficiency is associated with neurodegenerative disorders (27). In addition, during thermal degradation, thiamine generates aroma compounds such as furanthiols (28), thiophenes (29), and thiazoles (30), which contribute to meaty and nutty aromas characteristic of baked meat; (2) L-methionine is an essential amino acid involved in methylation reactions, antioxidant defense via glutathione synthesis, and hepatic function. Upon heating, it undergoes Strecker degradation to produce methional, a well-known aroma compound with a savory, potato-like odor that is commonly found in pork, beef, and chicken (31–35). (3) Myoglobin, which contributes to the generation of aroma compounds, as its iron content catalyzes lipid oxidation reactions (36). Additionally, its presence in plant-based meat increases the formation of lipid oxidation products during heating (37).
Here, we aimed to modulate the volatile compound profile of cultivated pork fat cells by manipulating media components. This approach offers a promising opportunity to leverage media formulation as a tool to enhance the sensory qualities of cultivated fat, thereby advancing applicability in food systems.
2 Results
2.1 Characterization of porcine dedifferentiated fat cells and optimization of growth and adipogenesis media
To achieve rapid cell proliferation, as well as the maintenance of adipogenic capability over multiple passages and efficient adipocyte differentiation, we evaluated proliferation and adipogenesis media using a pragmatic selection guided by previous reports (38–41), rather than through systematic optimization. Cells were cultured using three different growth media formulations which developed based on previous publications, here in after ‘20%FBS’, ‘20%FBS + ACY (A 83–01, CHIR99021 and Y-27632)’ and ‘15%FBS + bFGF (basic fibroblast growth factor)’, in (Supplementary Table S1). The results showed that cells cultured in 20%FBS experienced slower proliferation, with 72.7 h doubling time at passage 19, leading to the termination of this condition. In contrast, cells cultured with 20%FBS + ACY exhibited an average doubling time of 33.4 h, while those cultured with bFGF displayed the fastest growth, with an average doubling time of 22.8 h (Figure 1A). Cell diameters were also monitored across passages (Figure 1B). Cells cultured in medium containing 20%FBS reached an average diameter exceeding 20.0 μm (the maximum quantification limit), while those cultured in 20%FBS + ACY and 15%FBS + bFGF media maintained smaller diameters, with averages of 14.6 μm and 13.6 μm, respectively.
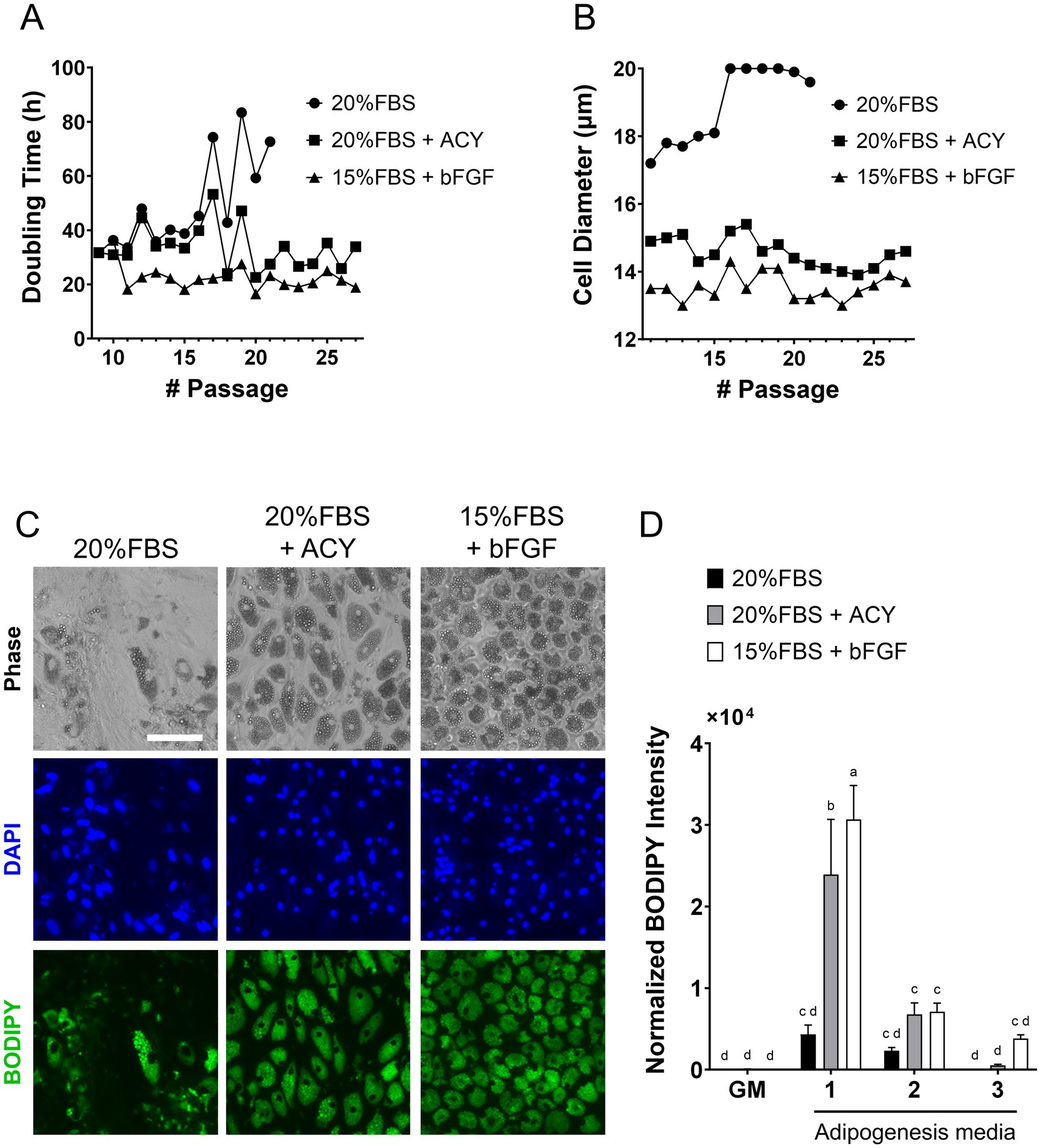
Figure 1. Characterization of pDFAT cells maintained under three different proliferation media, ‘20%FBS’, ‘20%FBS + ACY’ and ‘15%FBS + bFGF’, as well as three different adipogenesis media (1 to 3) to achieve rapid cell growth and maintaining adipogenic capability during continuous passage. (A) Hours per cell doubling. (B) Cell diameter. (C) Morphology of adipocytes maintained with three different proliferation media. Lipids were stained with BODIPY (p23). Scale bars, 100 μm. (D) Lipid quantification was performed using BODIPY staining. Average BODIPY integrated intensity was multiplied by BODIPY count and divided by the number of nuclei. GM refers to growth media. Adipogenesis was induced using three different adipogenesis media (Media 1, 2 and 3). n = 5 for each group. Statistical significance was determined using Two-way ANOVA with multiple comparisons. Different letters indicate statistically significant differences among groups (p < 0.05, Mann–Whitney test).
The optimal composition of adipogenesis media were studied. The three different adipogenesis media formulations were based on published media (42–44) (Supplementary Table S1), here in after referred to as “Media1, 2 and 3.” The morphology of the cells maintained in ‘20%FBS’, ‘20%FBS + ACY’ and ‘15%FBS + bFGF’ proliferation media and induced adipogenesis by Adipogenesis Media1 was stained with BODIPY (Figure 1C). Among the condition of proliferation media, ‘20%FBS’, ‘20%FBS + ACY’ or ‘15%FBS + bFGF’, and Adipogenesis Media1, 2 and 3 were tested, the combination of cells maintained in ‘15%FBS + bFGF’ proliferation media and induced to undergo adipogenesis using Media1 exhibited the highest lipid accumulation capacity compared to cells cultured under all other growth media conditions (Figure 1D). Therefore, the combination of proliferation media ‘15%FBS + bFGF’ and Adipogenesis Media1 was selected for subsequent experiments.
2.2 Analysis of cell proliferation and adipogenic efficiency with aroma precursor supplementation
To investigate the effects of thiamine-HCl or L-methionine on cell proliferation, relative DNA amount was quantified (Figure 2A). Additionally, the effects of thiamine-HCl, L-methionine and myoglobin on lipid accumulation were assessed (Figures 2B, C). During the adipogenesis lipid accumulation period, supplementation with thiamine-HCl and L-methionine did not result in a decrease in lipid quantities. However, during the cell proliferation period, supplementation with L-methionine at concentrations above 2.0 mM reduced cell growth. Therefore, all supplements were added only during the adipogenesis lipid accumulation period since there was no beneficial effect of the supplementation on cell proliferation. Optimal concentrations, 500 μM thiamine-HCl, 5.0 mM L-methionine, and 3.0 mg/mL myoglobin were chosen for the supplementation for the rest of this work.
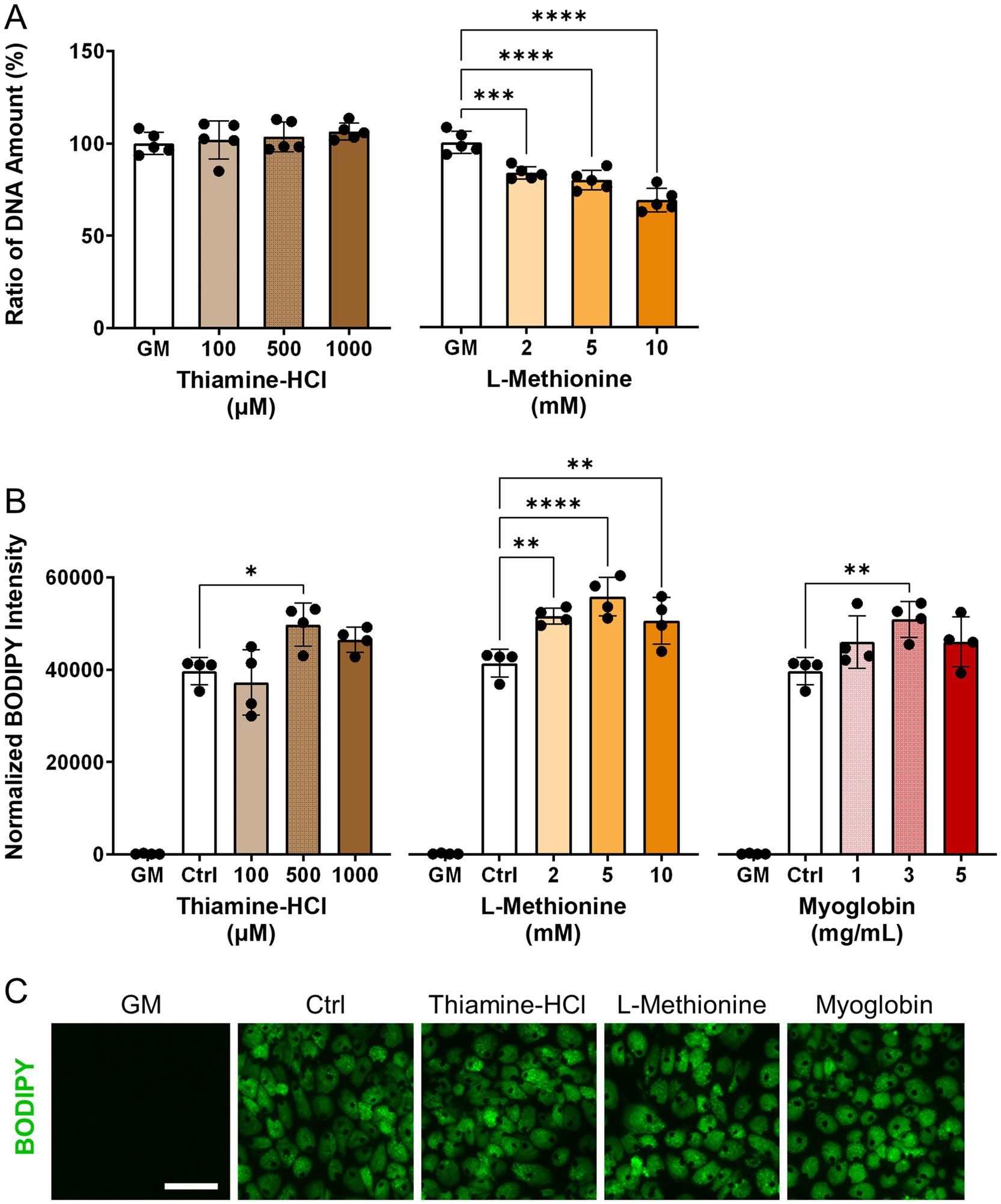
Figure 2. Effect of media supplements on proliferation and adipogenesis in pDFAT cells. (A) DNA quantification for pDFAT cells maintained with the proliferation media supplemented with different media additives (n = 5). The non-supplemented growth media condition (GM) was considered as 100%. (B) Lipid quantification of pDFAT-derived adipocytes treated with supplements during adipogenesis lipid accumulation period. Average BODIPY integrated intensity was multiplied by BODIPY count and divided by the number of nuclei. GM; cultured in proliferation media, Ctrl; cultured in adipogenesis media without supplementation. Statistical significance was determined using one-way ANOVA followed by Dunnett’s test (*, **, ***, **** denote p < 0.05, p < 0.01, p < 0.001 and p < 0.0001, respectively) compared to Ctrl. (C) Morphology of pDFAT-derived adipocytes cultured in adipogenesis media supplemented with 500 μM Thiamine-HCl, 5 mM L-Methionine or 3 mg/mL Myoglobin. Scale bars, 100 μm.
2.3 Metabolite analysis of cultivated fat before cooking
Relative levels of thiamine, thiamine pyrophosphate, methionine, methionine sulfoxide, S-adenosyl-L-homocysteine (SAH), and S-adenosyl-L-methionine (SAM) were quantified. No statistically significant differences were detected; however, a tendency toward increased thiamine pyrophosphate, a downstream metabolite of thiamine, was observed (Figure 3A). Similarly, levels of SAH, an immediate downstream metabolite, and SAM, a subsequent downstream metabolite of L-methionine, were not significantly different but both showed an increasing trend relative to the non-supplemented control (Figure 3B). Methionine sulfoxide, an oxidative product of L-methionine (45), also exhibited a tendency toward elevation. The metabolic pathways of thiamine and L-methionine in pig (Sus scrofa) were confirmed using the KEGG database (ssc00730, ssc00270).
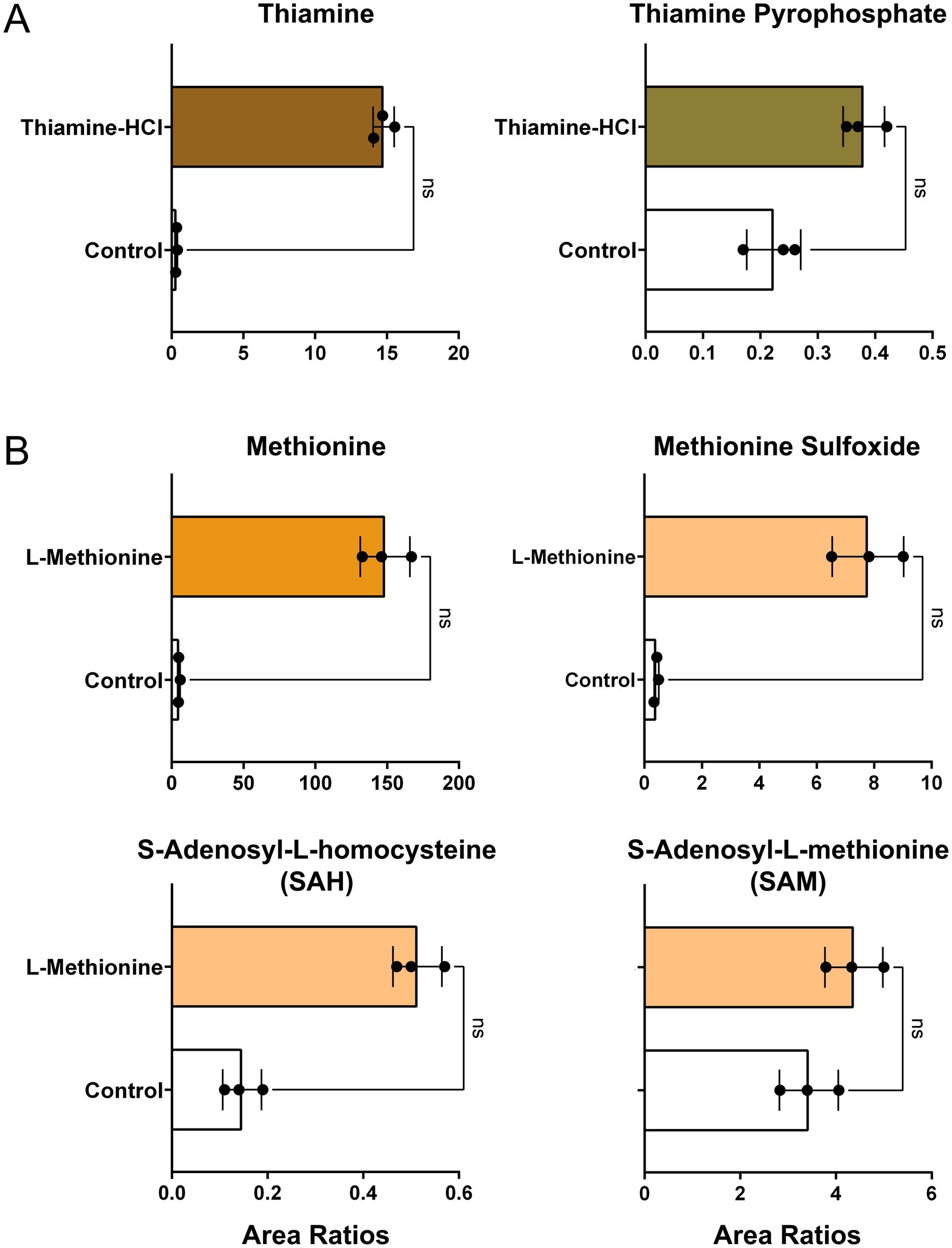
Figure 3. Relative levels of metabolites in cultivated fat. (A) Peak area ratios of thiamine and thiamine pyrophosphate in non-supplemented control and thiamine-HCl-supplemented cells prior to baking (mean±SD, n = 3). (B) Peak area ratios of methionine and its downstream intermediates in non-supplemented control and L-methionine-supplemented samples prior to baking (mean±SD, n = 3). Area ratios were calculated using the following formula: [{(Target Peak Area)/(Heavy Carbon-Labeled Phenylalanine or Methionine) divided by protein content measured by BCA assay. Statistical significance was assessed using the non-parametric Mann–Whitney test.
2.4 Fatty acid analysis of harvested fat before cooking
To determine the specific types of fatty acids accumulated as triglycerides or phospholipids in cultivated fat, fatty acid analysis was performed. Additionally, the fatty acid profiles of both non-supplemented and myoglobin-supplemented cultivated fat were analyzed prior to baking, as the iron in myoglobin could potentially influence the composition. The results showed that the fatty acid composition of the non-treated samples consisted of 45.6% C18:1 (cis-9), 17.1% C18:2 (all-cis-9,12), 14.2% C16:0, and 9.89% C18:0, as the top four fatty acids identified (Figure 4A). There was no statistically significant difference between non-supplemented and myoglobin-treated cultivated fat in either unsaturated or saturated fatty acid content (Figure 4B).
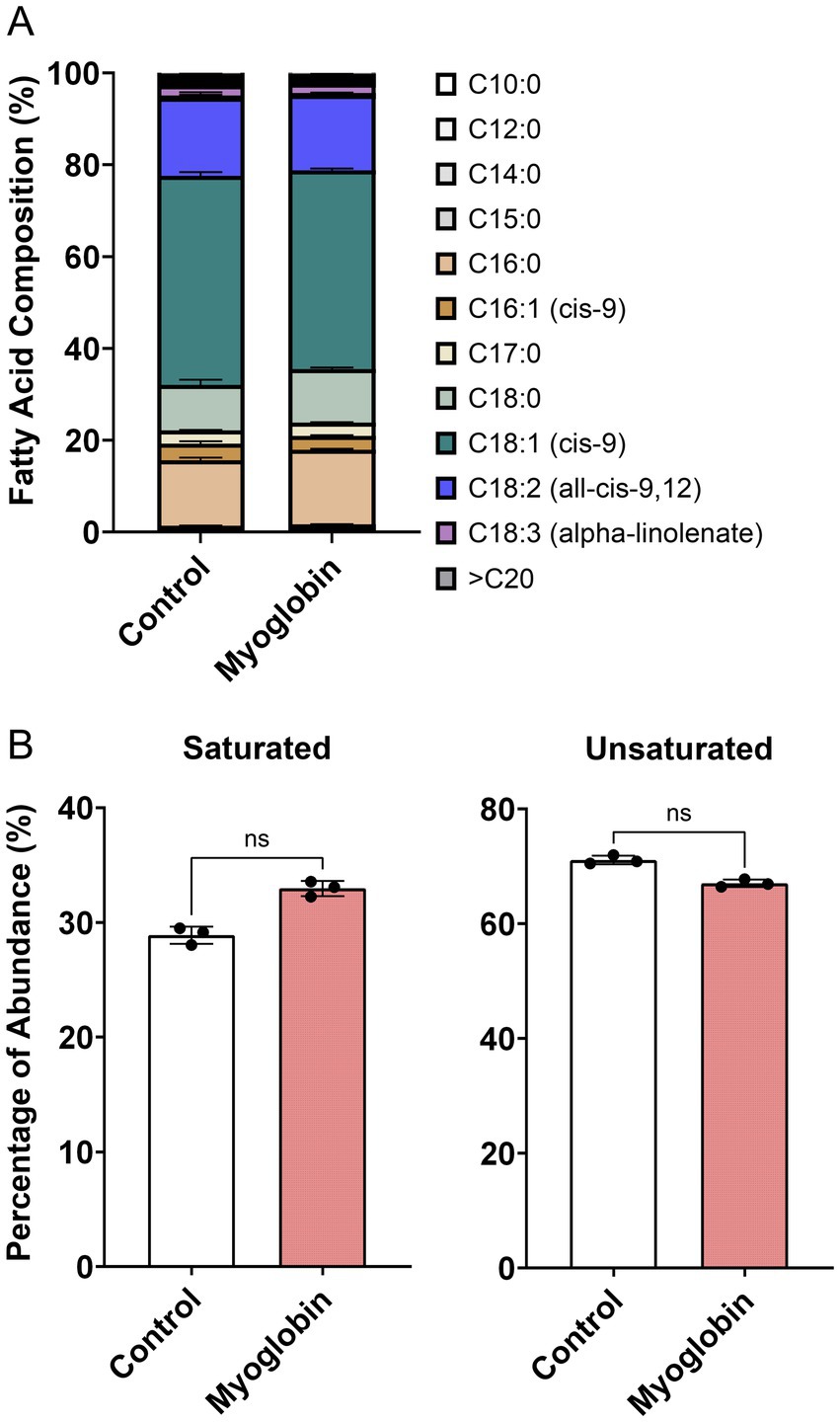
Figure 4. Fatty acid profile of cultivated fat. (A) Fatty acid composition between non-treated control and myoglobin, prior to baking (mean±SD, n = 3). (B) Percentage of total saturated and unsaturated fatty acids. Statistical significance was assessed using the non-parametric Mann–Whitney test.
2.5 Impact of aroma precursor supplementation on the concentration and profiles of volatile compounds from cells
The VOCs produced upon the addition of aroma precursors to the medium and subsequent heating/cooking were analyzed using DHS-GC–MS. The peaks of all compounds identified through deconvolution, were normalized using the internal standards and cell mass, described in ‘GC/MS data processing’ were represented in dot plots. Thiamine-HCl significantly induced the generation of 4-methyl-5-thiazoleethanol (sulfurol), milky aroma compound which derived from thiamine degradation (46, 47) (Figure 5A). L-methionine promoted the formation of methional, a well-known potato-like aroma compound (Figure 5B). The addition of myoglobin caused significant changes to the VOC profile, beginning with the formation of γ-nonalactone, (E, E)-2,4-decadienal, 2-pentylfuran, δ-decalactone, and benzeneacetaldehyde, which are responsible for the coconut-like, deep fat, fruity, peachy, and honey aroma of meat (48–51), while increasing heptanal and 1-pentanol which imparts green, fatty aroma (52, 53) (Figure 5C). Furthermore, myoglobin enhanced the production of various lipid degradation products, including aldehydes, alcohols, furans and some of ketones, fatty acids, and hydrocarbons, and phenolic derivatives. Some of the esters also showed an increase; however, the average fold change was not as significant compared to that of aldehydes, alcohols, and furans (Table 1). No significant changes were observed in the levels of pyrroles and thiazoles (Figure 6). All compounds and peaks found in the non-supplemented controls, along with those supplemented with thiamine-HCl, L-methionine and myoglobin are shown in (Tables 1–3).
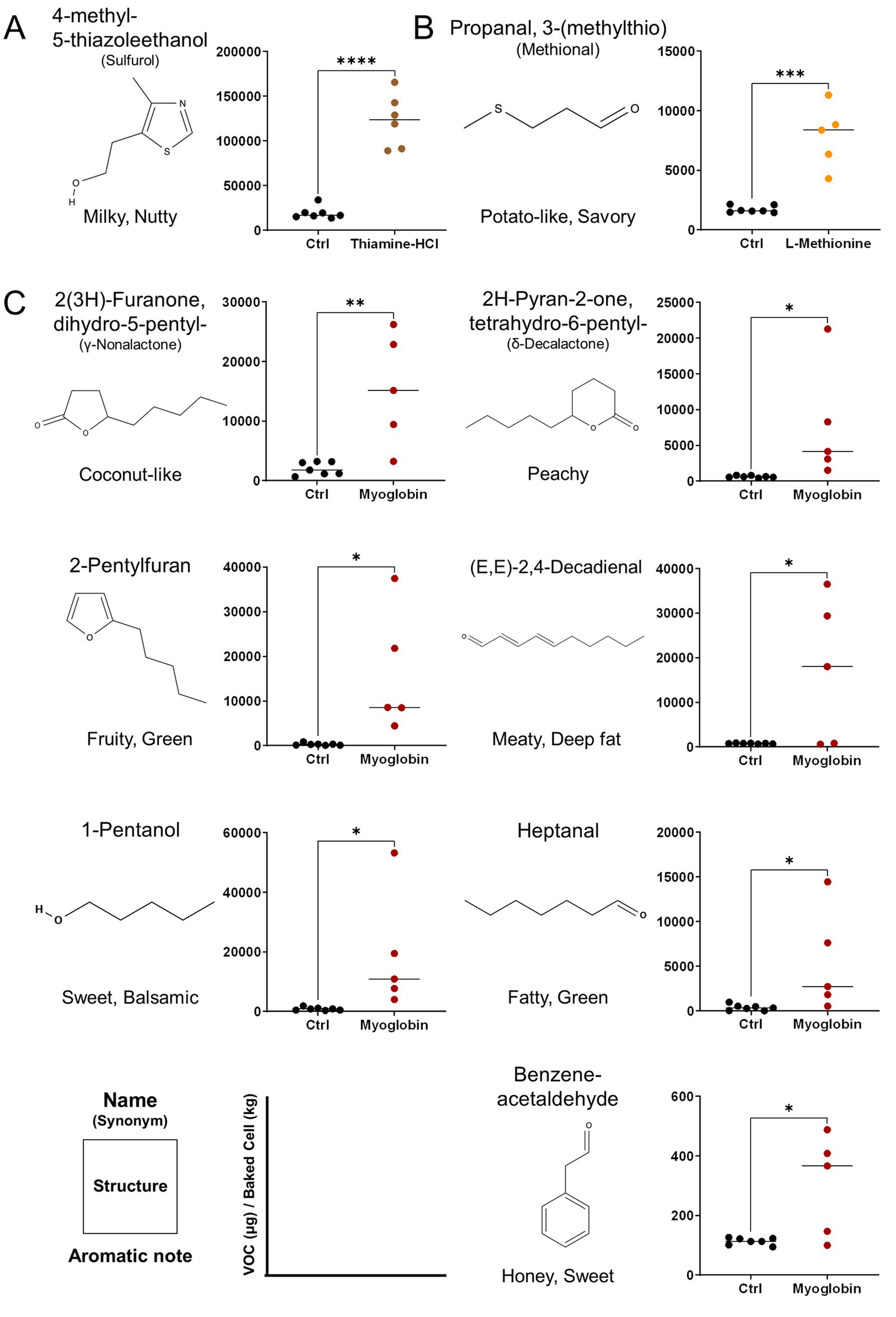
Figure 5. Major volatile organic compounds (VOCs) provoked by media supplementations and detected by DHS-GC–MS derived from pDFAT-derived adipocytes upon baking. VOCs altered under (A) 500 μM of Thiamine-HCl, (B) 5 mM of L-Methionine, (C) 3 mg/mL Myoglobin-supplemented conditions. VOCs were quantified by normalizing peak areas to two internal standards and converting the values to mass using authentic standard curves. The resulting concentrations were further normalized to cell mass after baking. ‘Ctrl’ indicates the non-supplemented cell condition. Replicates 5 to 7 include biological triplicates. Statistical significance was determined using unpaired t-test (*, **, ***, **** denote p < 0.05, p < 0.01, p < 0.001 and p < 0.0001, respectively). Compounds were identified by retention index (RI) and Mass Spec referencing NIST17 compared with authentic standards.
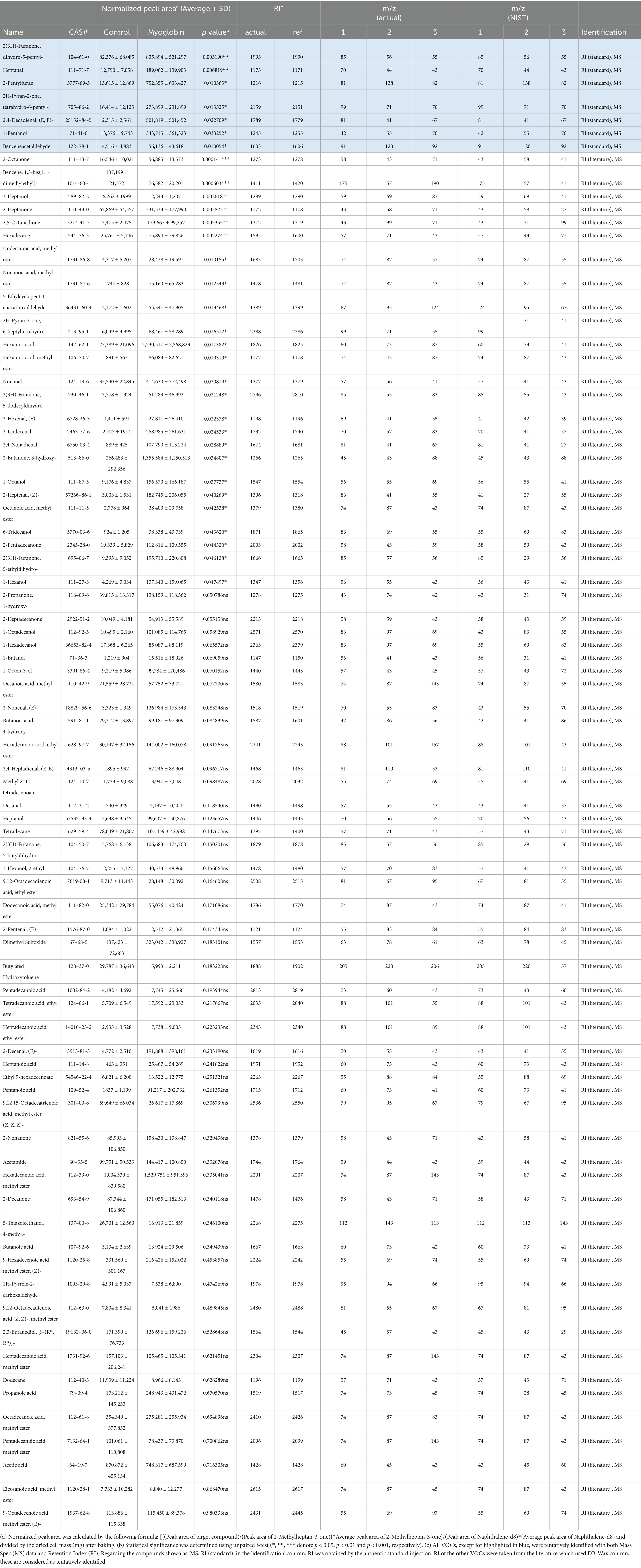
Table 1. All the volatile organic compounds (VOCs) found in 120 °C heated cultivated fat maintained with non-supplemented media (Control) and supplemented with 3 mg/mL myoglobin.
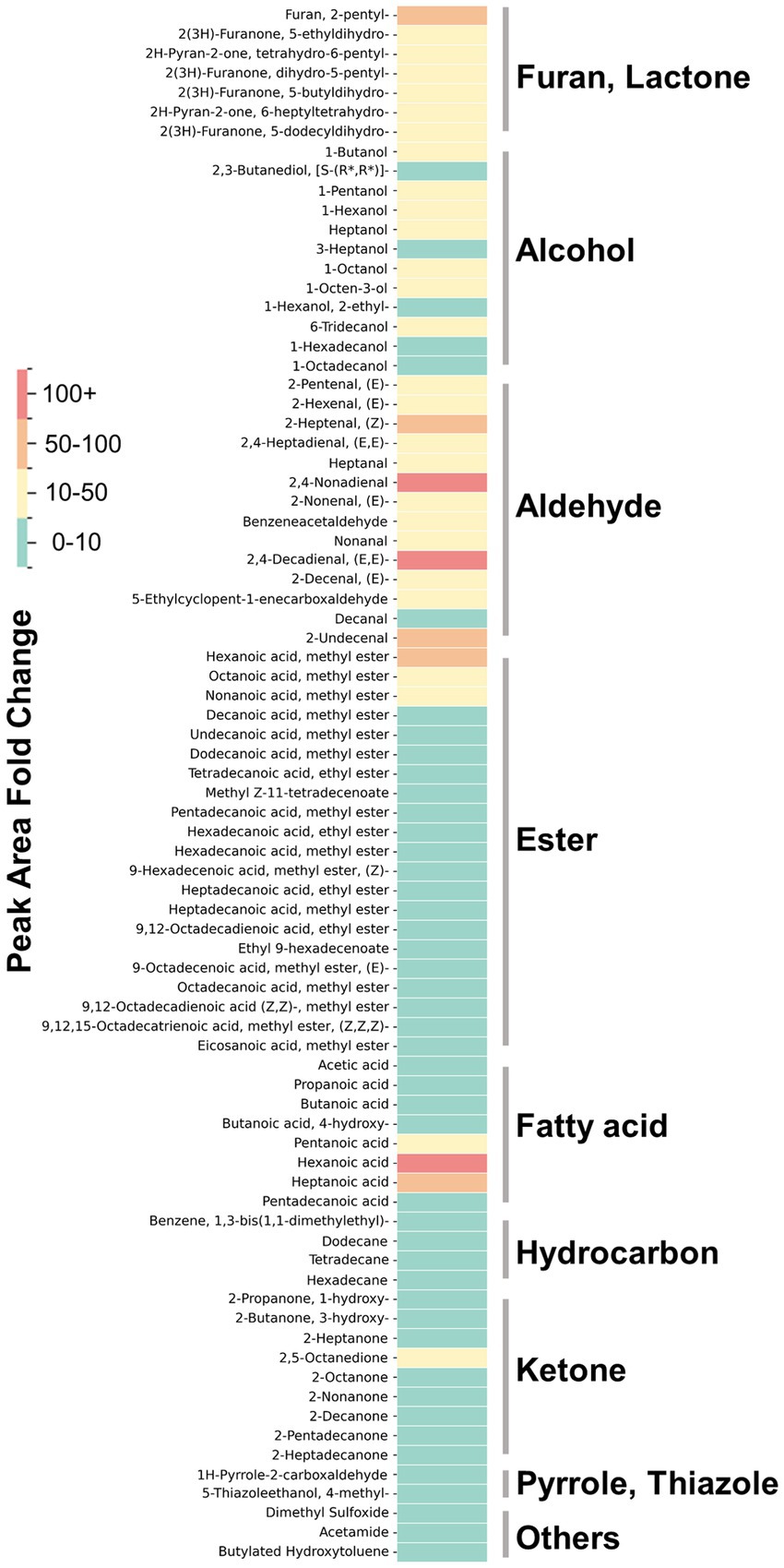
Figure 6. The fold change in the normalized peak area of VOCs found in myoglobin-supplemented cultivated fat, compared to non-supplemented control cells. The color intensity represents the value of the fold changes. All VOCs are tentatively identified except for VOCs shown in Figure 3.
3 Discussion
In this study, we demonstrated that aroma volatiles in cultivated fat cells can be altered by media supplementation, resulting in different flavors in fat, while maintaining rapid proliferation and robust adipogenesis capability. Prior to the investigation of flavor precursor supplementation, we aimed to select comparatively better media conditions to achieve rapid cell proliferation between the three proliferation media previously reported (40, 41, 54). While bFGF has a positive effect on both proliferation and differentiation for the cells (55, 56), the cost has been highlighted as an issue (57). Therefore, as an alternative we considered the addition of small molecule compounds, as previously reported in studies with MSCs (40). An improvement in cell proliferation rate was observed with the addition of bFGF or ACY, which aligns with previous reports on pDFAT or human MSCs (Figure 1A) (40, 41). Additionally, cells maintained in ‘20%FBS + ACY’ or ‘15%FBS + bFGF’ exhibited smaller cell sizes compared to those maintained in ‘20%FBS’ (Figure 1B). Several prior studies have reported that aged cells tend to have larger sizes, whereas stem cells or those undergoing rapid self-renewal cycles are typically smaller (58, 59), consistent with the results reported here, where the cells maintained with ‘20%FBS + ACY’ or ‘15%FBS + bFGF’ showed faster proliferation and higher adipogenic capability (Figure 1D). These results showed that bFGF remains a crucial supplement for maintaining rapid cell proliferation and robust differentiation capability. Small molecule cocktails such as ACY are desirable as a substitute for bFGF, however, there was a subsequent unexpected challenge in that cells maintained with ‘20%FBS + ACY’ exhibited stronger cell adhesion, requiring over 20 min of cell dissociation treatment after the 20th passage (data not shown). The exploration of appropriate combinations and concentrations of each inhibitor could resolve this cell dissociation problem. Further, if these compounds can be replaced with food-grade materials, they could be utilized as effective growth promoters and factors for maintaining adipogenic capability, yet also keep costs lower and thus potentially improve regulatory acceptability.
In terms of adipogenic differentiation, three different adipogenic media which were previously reported were tested in the present study (42–44). Media2 contains commonly used components for inducing adipogenesis, including insulin, IBMX (isobutylmethylxanthine), rosiglitazone, and dexamethasone (Dex) (43). Media3 includes only two inducers, insulin and rosiglitazone additions to the essential minimum required for adipogenesis (44). Media1 is based on a previously reported medium (42), to which Intralipid, has been added. Media1, which contained Intralipid, demonstrated the most efficient lipid accumulation in our isolated pDFAT cells (Figure 1D). It is hypothesized that Intralipid, which contain lecithin and fatty acids, significantly enhance adipogenic differentiation (60–62). Therefore, it can be inferred that these components contribute substantially to the highest lipid accumulation among three different adipogenesis media studied in the present research. Future research should focus on the consideration of lipid-based additives that can balance the regulation of fatty acid composition with the promotion of efficient fat accumulation. Although supplementation with aroma precursors such as L-methionine inhibited cell proliferation at concentrations above 2.0 mM, it was well tolerated during the adipogenic phase and did not impair lipid accumulation (Figures 2A,B,C). This observation implies the possibility that the altered metabolic state of differentiating adipocytes confers greater tolerance to L-methionine, thereby enabling the use of higher concentrations required for efficient volatile compound production.
A tendency toward intracellular accumulation of thiamine and L-methionine upon media supplementation was observed, although the differences did not reach statistical significance (Figures 3A,B). High doses of thiamine-HCl or L-methionine raise concerns regarding the potential formation of undesirable thermal by-products—such as certain heterocyclic amines (63) or compounds generated through oxidative stress (64) —during the cooking process, rendering further safety assessment essential. Additionally, further testing of various media supplements would be desirable to achieve enhanced nutritional fortification.
Thiamine-HCl supplementation led to the statistically significant increase of 4-methyl-5-thiazoleethanol (Figure 5A), and there is a tendency for thiazole, tetrahydro-, which is predictably identified, to be induced; however, no statistically significant difference was observed (Table 2). On the other hand, thiols and thiophenes which were supposed to be derived from thiamine, were not detected in this experimental setup. Higher sensitivity detection methods, such as GC–MS/MS, could potentially confirm additional thiamine degradation products in the cooked cultivated fat supplemented with thiamine-HCl. Furthermore, it’s possible that the amount of cell sample influences the detection of sulfide-containing VOCs. Therefore, if we prepare the cell samples through suspension culture and provide a larger amount cells, such as on a gram scale, we may be able to detect such VOCs (18).
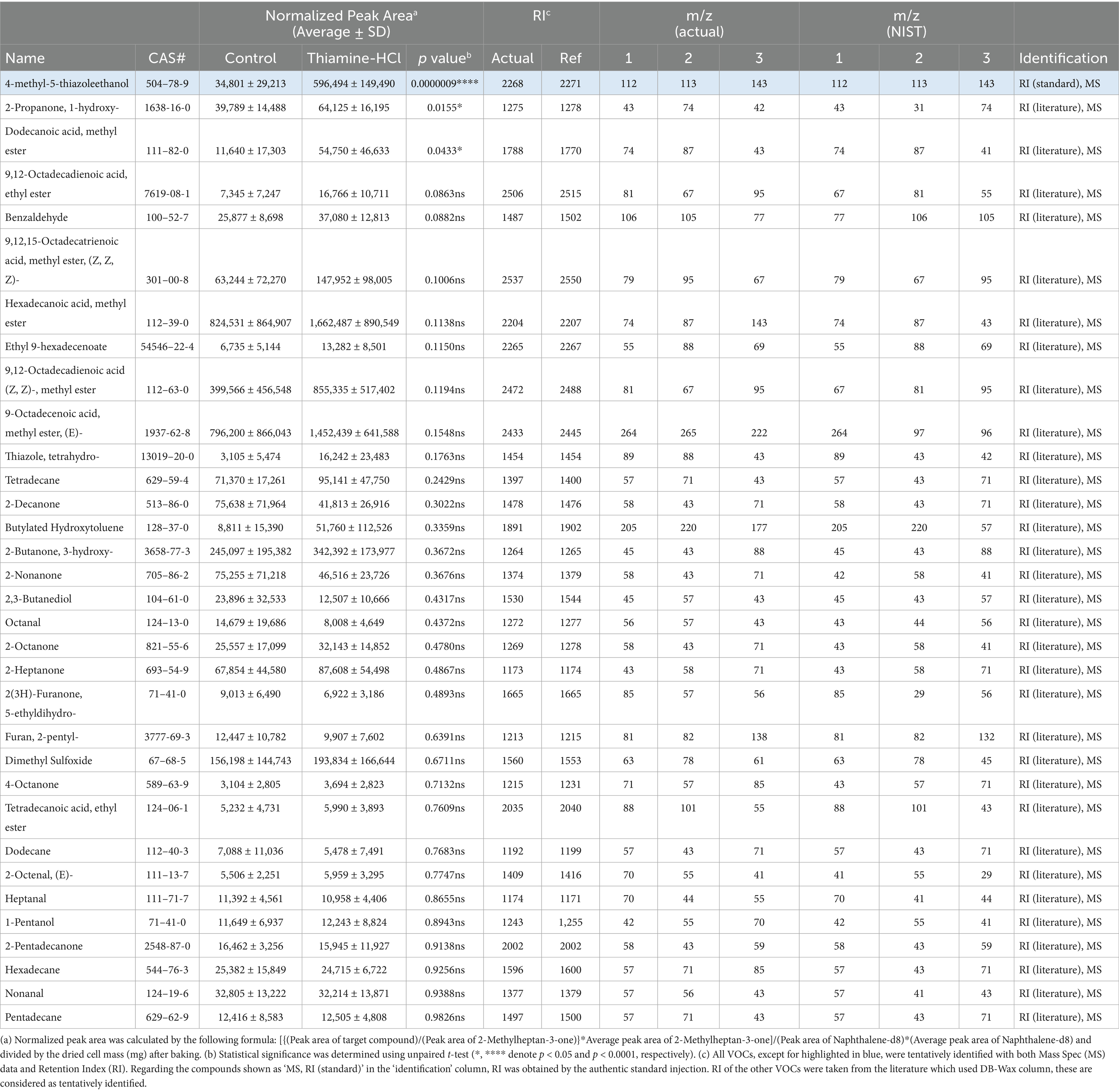
Table 2. All the volatile organic compounds (VOCs) found in 120 °C heated cultivated fat maintained with non-supplemented media (Control) and supplemented with 500 μM thiamine-HCl.
L-methionine supplementation enhanced the production of methional, which emits a potato-like aroma (31, 32) during heating/cooking. However, it was implied that L-methionine supplementation has the possibility to promote the generation of methanethiol, which is predictably identified, a degradation product of methional (65) (Table 3). Methanethiol is described as having an onion-like odor, and at higher concentrations, it can be perceived as an unpleasant smell. To fully assess the impact of L-methionine on the aroma profile of cultivated pork fat, further studies incorporating descriptive sensory panels are required.
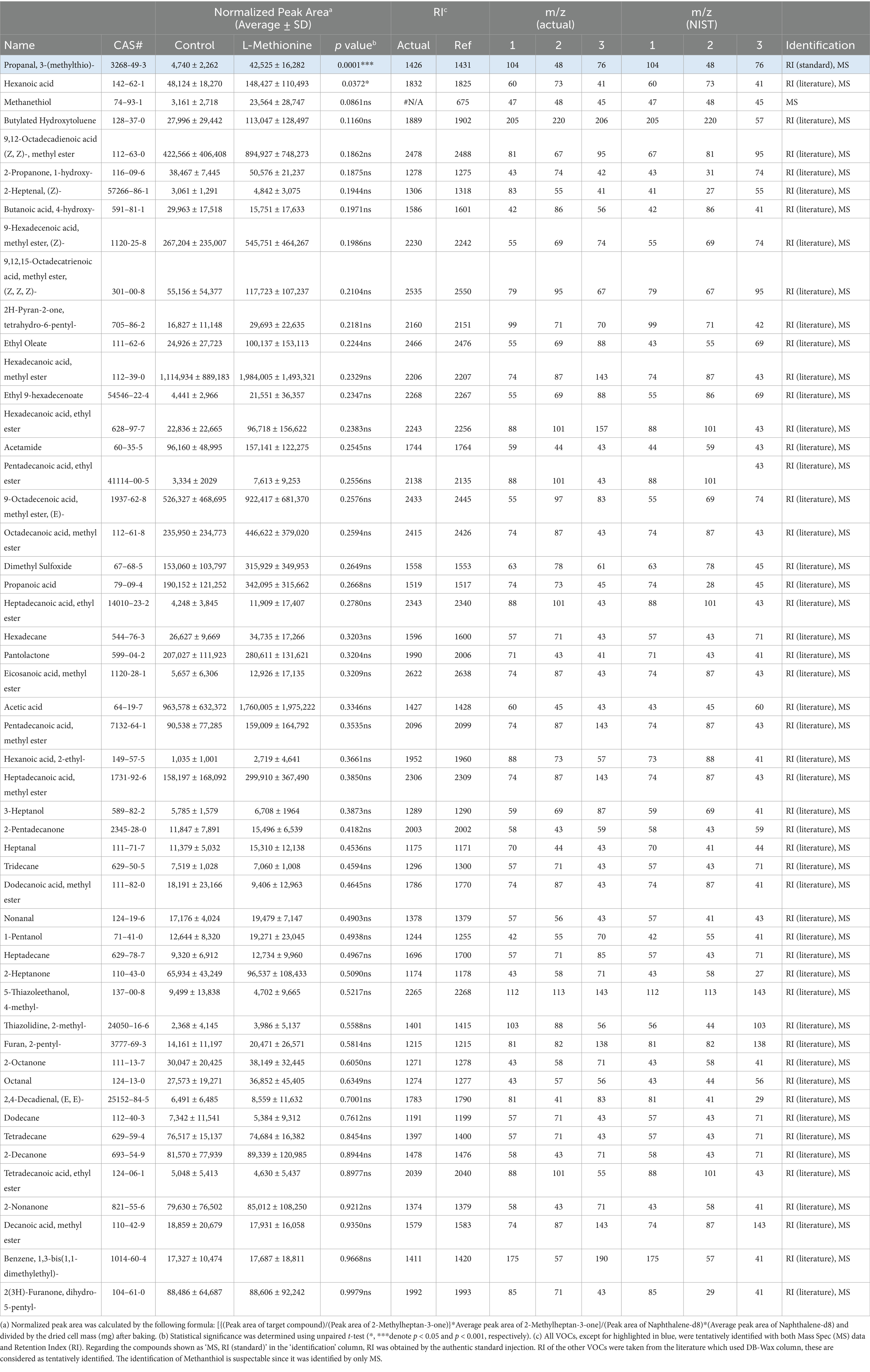
Table 3. All the volatile organic compounds (VOCs) found in 120 °C heated cultivated fat maintained with non-supplemented media (Control) and supplemented with 5 mM L-methionine.
Myoglobin in plant-based meat increases the formation of lipid oxidation products during heating, which increases the flavor complexity and is linked to characteristics such as a serum-like taste and a metallic mouthfeel (37, 66). We examined whether supplementation with myoglobin would enhance the formation of the lipid-derived aroma compounds in cultivated fat. Myoglobin significantly enhanced the formation of aldehydes, alcohols, furans, lactones and some of ketones and fatty acids (Figure 6). Notably, (E, E)-2,4-decadienal, a characteristic aroma compound known for its association with the deep-fat, meaty aroma of cooked meat, was reported to be 22 times greater than the non-supplemented condition (67) (Figure 5C). Additionally, lactones, such as γ-nonalactone and δ-decalactone, which imparts coconut-like, peachy aroma (68, 69), were each statistically significantly enhanced by 7.6 times and 12 times greater than the myoglobin-supplemented conditions. Heptanal, 1-pentanol and predictably identified aldehydes such as hexanal which exhibits fatty aroma (70, 71) also were enhanced. Hexanal has been associated with off-flavors that may be perceived as unpleasant at certain concentrations (70, 72) (Table 1). Therefore, it will be important in future studies to determine, through sensory analysis, whether its concentration reaches levels perceived as unpleasant by humans.
The results of the fatty acid analysis of the cultivated fat prior to cooking showed that the addition of myoglobin did not affect the fatty acid composition. Given the significant impact of myoglobin observed in this study, the combination of myoglobin and cultivated fat holds potential for altering aroma profiles (37, 73). Additionally, in muscle satellite cells, it has been reported that myoglobin promotes cell proliferation (74). However, considering the cytotoxicity effects of iron in cell culture (75), the post-harvest addition of myoglobin might be a more effective depending on the cell type. Fatty acid analysis revealed that cultivated fat contained 17.1% linoleic acid (Figure 4A). The decomposition of linoleic acid is known to generate aroma compounds such as (E, E)-2,4-decadienal, 2-pentylfuran and 1-pentanol, aligning with the results observed in the present study (76–79).
As part of future work, exploring the addition of supplements, such as L-glutamic acid or inosine monophosphate to enhance the umami taste, and linoleic acid rich edible oils provide further strategies to modify the flavor and nutritional profile of cultivated foods. Furthermore, investigation of masking supplements to prevent the formation of off-flavors, such as hexanal which was enhanced by myoglobin in this study is considered crucial for further enhancing desirable odor changes. Additionally, testing various combinations and concentrations of supplements could yield other insights to provoke desirable aromas. There is a possibility that both the individual effect of ribose supplementation and its combination with other flavor precursors could synergistically enhance the formation of aroma volatiles (Supplementary Figures S2, S3). Such approaches may have potential not only with porcine cells but also in cells from other tissues and other species, such as bovine (Supplementary Figure S4), as well as in muscle cells or undifferentiated (e.g., stem) cells. The present study provides evidence that the amino acid and vitamin contents, as well as the aroma volatile profiles, of cultivated fat can be systematically altered by modulating media components, thereby enhancing the potential of cultivated fat as a food ingredient.
4 Materials and methods
4.1 Cell isolation
Dedifferentiated porcine (Sus domesticus) cells (pDFAT) and bovine (Bos taurus) DFAT cells were isolated as previously described (39) from the belly (subcutaneous fat) of a 93-day-old female Yorkshire pig (DOB: 10/18/2021) and from the tailhead fat tissue (subcutaneous fat) of a 604-day-old male Angus/Holstein cross steer (DOB: 09/25/2022) were isolated. Briefly, only the modifications specific to cattle are outlined here. Adipose tissue was minced and digested in 0.2% collagenase (LS004176; Worthington Biochemical, Lakewood, NJ) dissolved in DMEM/F12 (11320033; Thermo Fisher, San Jose, CA), supplemented with 10% of antibiotic-antimycotic (100X) (15240062; Thermo Fisher) and 0.75% of 10% Pluronic F-68 (24040032; Thermo Fisher, San Jose, CA), for 1.5 h at 37 °C with shaking. The digest was filtered through 750 μm and then 300 μm cell strainers and centrifuged at 400 g for 5 min to collect mature adipocytes from the top layer of the supernatant. The lipid-rich layer was transferred to a tissue culture flask and incubated to allow stromal vascular cells to adhere, thereby separating them from the mature adipocytes. After 2 days, the floating lipids were transferred to a new tissue culture flask containing fresh media to initiate ceiling culture. Once dense colonies of lipid-laden cells were observed, the flask was flipped back to its normal position for routine maintenance.
4.2 Cell culture
Passage 9 pDFAT cells were thawed and cultured using three different growth media formulations which developed based on previous publications (38–41), here in after ‘20%FBS’, ‘20%FBS + ACY’ and ‘15%FBS + bFGF’ with of 0.25 μg/cm2 laminin 511-E8 (N-892021; Iwai North America Inc., San Carlos, CA) which added to the media during cell seeding, shown in (Supplementary Table S1). Cells were repeatedly passaged, and their doubling time compared. In ‘20%FBS + ACY’, the appropriate concentration of each inhibitor was determined by cell proliferation assay, described below (Supplementary Figure S1). Cells were maintained by passaging or stored by freezing as previously described (39). Passage 23 or 24 cells were used for GC–MS analysis, samples were seeded into 150 mm dishes (430,499; Corning, Tewksbury, MA). Passage 3 of bDFAT cells were maintained in ‘15%FBS + bFGF’ on laminin coated surface.
4.3 Cell proliferation assay
To determine the effect of supplementation of thiamine-HCl and L-methionine supplementation during cell proliferation, DNA amount was quantified using the CyQUANT™ Cell Proliferation Assay kit (C7026; Thermo Fisher) according to the manufacturer’s instructions. Cells were seeded in 96-well plates at 8,000 cells/cm2. Thiamine-HCl and L-methionine treatments were administered after the cells had adhered following seeding and cultured until the cells reach to 80% confluent. For the evaluation of A 83–01, CHIR99021 and Y-27632, cells were seeded in multiple 96-well plates at 6,000 cells/cm2 and cultured for 24 to 120 h. The culture medium was replaced every other day. Although cells were seeded at the same density and at the same time, each time point was measured from a separate well.
4.4 Adipogenic differentiation
After pDFAT cells reached 100% confluency in growth media and remained confluent for at least 24 h, their medium was replaced with adipogenic induction medium. The three different adipogenesis media formulations were developed based on (42–44) with replacing Chemically-defined FBS replacement changed to FBS, and 1% Penicillin/Streptomycin/Amphotericin (PSA) to 100 μg/mL Primocin (Supplementary Table S1). For all media compositions, the cells were fed every two days until Day 8. To assess the effect of thiamine-HCl and L-methionine, using the adipogenesis Media1, supplements were applied during the adipogenesis lipid accumulation phase, days 2 to 8, with day 0 defined as the day of media transition to adipogenesis induction media. Myoglobin treatment was limited to 24 h prior to cell harvest, due to its potential cytotoxicity and inhibition of cell proliferation (81). For each added compound, the concentration that yielded the most effective lipid accumulation was selected for use in cell culture for DHS-GC–MS analysis.
4.5 Lipid staining
Cultured adipocytes were stained to confirm intracellular lipid accumulation. Cells were washed twice with DPBS(−) (14190144; Thermo Fisher) to avoid cell detachment and fixed with 4% paraformaldehyde (PFA) for 20 min at room temperature (RT). After fixation, cells were first rinsed with DPBS(−), then incubated at RT for 1 h with 2 μM 4,4-difluoro-1,3,5,7,8-pentamethyl-4-bora-3a,4a-diaza-s-indacene (BODIPY 493/503, D3922; Invitrogen) diluted in DPBS(−). After BODIPY incubation, cells were rinsed three times with DPBS(−) and the cell nuclei were stained with 2 μg/mL 4′,6-diamidino-2-phenylindole (DAPI, 62247; Thermo Fisher) in DPBS(−) for 15 min at room temperature. After DAPI staining, cells were rinsed twice with DPBS(−) and stored in DPBS(−). Imaging was performed with a fluorescent widefield microscope (KEYENCE, BZ-X700, Osaka, Japan). Using this stained plate, adipogenesis efficiency was quantified by measuring normalized BODIPY intensity by Celigo Image Cytometer (200-BFFL-5C; Nexcelom Bioscience LLC, Lawrence, MA). Specifically, the average integrated intensity of BODIPY was multiplied by the area of BODIPY, and the resulting value was normalized by dividing it by the number of nuclei stained with DAPI.
4.6 Fat harvest
For GC–MS analysis, cells were prepared with Adipogenesis Accumulation Media 1 supplemented with supplementation of 500 μM of thiamine-HCl (T1270; Millipore Sigma, Burlington, MA) or 5.0 mM L-methionine (M5308; Millipore Sigma) or 25 mM ribose (R7500; Millipore Sigma) for the lipid accumulation phase, Day 2 to 8. 3 mg/mL of myoglobin (M0630; Millipore Sigma) was added only during the last 24 h before sample collection. The spent culture media was aspirated at Day 8, then rinsed with DPBS(−) (Thermo Fisher) 3 times. Dishes were then kept vertical for 3 min to thoroughly drain DPBS(−) and any remaining media. Once excess DPBS(−) was aspirated, the adipocytes were harvested using a cell lifter (08–100-240; Fisher Scientific), then transferred into a pre-weighed 2.0 mL tube. Samples were stored at −80 °C.
4.7 Metabolite analysis
Polar metabolites were extracted from frozen cell pellets using 80% methanol as previously described (80). Metabolites were separated on an Atlantis Premier BEH Z-HILIC VanGuard FIT Column: 1.7 μm, 2.1 mm × 150 mm (Waters Corporation, Milford, MA) at a flow rate of 0.175 mL/min using the following gradient: 0–20 min: linear gradient from 80–20% B; 20–20.5 min: linear gradient form 20–80% B; 20.5–28 min: hold at 80% B. Mobile Phase A was a 10 mM ammonium carbonate; Mobile Phase B was acetonitrile. To quantify thiamine and thiamine pyrophosphate, metabolites were separated on a Luna PFP(2) LC column, 3 μm, 2 mm x 100 mm (Phenomenex). Mobile Phase A was water with 0.1% formic acid; Mobile Phase B was acetonitrile, with a flow rate of 0.25 mL/min and the following gradient: 0–2 min: hold at 2% B; 2–7.5 min: linear gradient from 2–60% B; 7.5–8.5 min: linear gradient from 60–100% B; 8.5–10.5 min: hold at 100% B; 10.5–15 min: hold at 2% B. The UHPLC (Vanquish Duo; Thermo Fisher Scientific) was coupled to an Orbitrap Exploris 240 (Thermo Fisher Scientific) and full scan data were acquired in polarity switching mode at a resolution of 120,000 (m/z = 200). Relative quantitation of metabolites was performed with Skyline using a mass tolerance of 5 ppm. Compound ID was confirmed by referencing an in-house spectral library containing retention times built with authentic chemical standards.
4.8 Fatty acid analysis
Lipid extractions were performed using a scaled-down methyl tert-butyl ether (MTBE)-based method as described in a previous study (39). As an internal standard, 20 μL of nonadecanoic acid (N5252; Millipore Sigma) at 6 mg/mL in hexane (139386; Millipore Sigma, Burlington, MA) was added with a glass syringe. The doubly extracted MTBE phase was dried under nitrogen, saponified with 3 mL of 0.5 M sodium methoxide in methanol (92446; Millipore Sigma) at 55 °C for 30 min and then methylated with 3 mL of 14% boron trifluoride/methanol (15716; Millipore Sigma) under the same conditions. After cooling, the solution was transferred to a 15 mL polypropylene centrifuge tube, mixed with 2 mL saturated NaCl solution and 2 mL hexane (139386; Millipore Sigma), vortexed, and centrifuged at 3,500 rpm for 5 min. The upper organic phase was collected into sample vials (26590, RESTEK) for GC-FID analysis. Fatty acid composition was analyzed using an Agilent 6,890 N GC (Agilent Technologies, Santa Clara, CA) equipped with a flame ionization detector and a Select FAME capillary column (CP7430; Agilent Technologies, Santa Clara, CA; 100 m × 0.25 mm × 0.25 μm). Injection volume was 1 μL (split 1:20) at 250 °C, with helium as the carrier gas. The oven was held at 100 °C for 5 min, ramped at 10 °C/min to 220 °C (28 min), then to 250 °C (10 min). Fatty acids were identified by comparing retention times with a standard mixture (Food Industry FAME Mix, 35077; RESTEK, Bellefonte, PA). Concentrations were quantified relative to the internal standard using calibration curves of methyl nonadecanoate (74208; Sigma-Aldrich, St. Louis, MO), which showed R2 = 0.996 and LOD = 0.732 μg/mL (S/N > 3). Peaks with S/N < 3 were considered not detected.
5 Volatile compound analysis
5.1 Dynamic headspace GC–MS
Samples were prepared by weighing a 60 mg cell pellet into a 20 mL headspace vial (23087; RESTEK, Bellefonte, PA). As an internal standard, 1 μL of 2-Methylheptan-3-one (A284658; AmBeed, Arlington Heights, IL), prepared as 5 mg/mL in methanol, was injected into the 20 mL headspace. Additionally, prior to DHS baking, TDU tubes were loaded with Tenax® resin (11982; Millipore Sigma) and conditioned at 300 °C for 120 min under a constant flow of ultra-pure nitrogen. After conditioning, 1 μL of Naphthalene-d8 (31043; RESTEK), prepared as 10 μg/mL in dichloromethane, was injected into the Tenax® resin bed. Dichloromethane was removed by reconditioning the DHS tube at 75 °C for 5 min under a constant flow of ultra-pure nitrogen. The Naphthalene-d8 was utilized to normalize the sample injection efficiency of GC/MS, and the 2-Methylheptan-3-one was utilized to normalize the DHS extraction efficiency. The 20 mL headspace (HS) vial was transferred to the DHS module at 120 °C and incubated for 15 min. The HS vial was then purged with 200 mL ultra-pure nitrogen at 100 mL/min. The preconditioned TDU tube, packed with Tenax® resin, was then transferred to the DHS trap module at 40 °C. The HS vial was then incubated at 120 °C and purged with 1,500 mL of ultra-pure nitrogen at 50 mL/min trapping volatiles on the trap at 40 °C. The trap was then moved to the dry purge position and purged with 750 mL ultra-pure nitrogen at 100 mL/min, with the trap at 40 °C. The TDU tube was then transferred to the TDU for thermal desorption. Prior to desorption, the CIS with glass bead liner, was cooled to −120 °C with liquid nitrogen. The PTV inlet was set to a split ratio of 1:10 by selecting the solvent vent mode and setting the solvent purge to initiate at 0.01 min at 12 mL/min flow. Desorption was initiated by ramping the TDU from 40 °C to 250 °C at 720 °C/min with a 3 min hold at 250 °C. After desorption the TDU tube was removed from the TDU. The chromatographic analysis was initiated with the ramping of the CIS from −120 °C to 250 °C followed by a 3 min hold. The 7890A was outfitted with a DB-WAX UI capillary column (30 m, 0.25 mm i.d., 0.25 mm film thickness (Agilent Technologies)). The 7890A oven temperature was programmed to hold at 40 °C for 2 min; then ramp at 5 °C/min to 250 °C and then hold 250 °C for 15 min. The helium carrier gas was set to a constant flow 1.2 mL/min. The MSD was set to a solvent delay for the initial 1.25 min, with an electron energy of −70 eV, a source temperature of 250 °C, and quadrupole temperature of 150 °C. Data was acquired in scan mode ranging from 35 m/z to 450 m/z. At least three biological replicates, each with 5 to 7 injection replicates per sample, were completed.
5.2 GC–MS data processing
Chromatographic deconvolution was performed using PARADISe software version 6.1.7, which enabled batch processing of the full set of chromatograms. The software applies PARAFAC2 modeling within user-defined time intervals to resolve coeluting compounds. Intervals were defined to include the baseline on both sides of each peak. In cases where peaks appeared to overlap, a composite interval was created to encompass the full region as well as intervals for each visually distinct peak. Following deconvolution, the resulting mass spectra were matched against the NIST17 mass spectral libraries for “probable identification.” Only compounds with Match Quality rated as ‘Excellent’ or ‘Good’ were included. “Tentative identification” was further supported by comparison with published retention indices (RI) obtained using an identical column (DB-Wax) and chromatographic conditions. Finally, compound identities were confirmed using authentic reference standards. Deconvoluted peaks were normalized using the following formula. Deconvoluted peaks were normalized using the following formula:
PTG: Peak area of target compound, P2m3h: Peak area of 2-Methylheptan-3-one, PAve.2m3h: Average peak area of 2-Methylheptan-3-one between samples, PAve.d8N: Average peak area of Naphthalene-d8 between samples, Pd8N: Peak area of Naphthalene-d8, and Cell mass represents the baked cell mass (mg) after DHS baking.
5.3 Volatile compounds quantification
The quantification range and the detection limit of each compound were determined by the serial dilution of the following authentic standard compounds. Each compound’s quantification range, coefficient of determination of the calibration curve, limit of detection (LOD), reagent purity of authentic standard, manufacturer, and catalog number are provided in Supplementary Table S2. In the non-supplemented control cell sample, when a peak area larger than the LOD but smaller than the limit of quantification (LOQ) was detected, the peak area was treated as the value of the LOQ, and absolute quantification was performed using the calibration curve.
6 Statistical analysis
Statistical analyses were performed using GraphPad Prism 10.4.1 (GraphPad Software, San Diego, CA, USA). Two-group comparisons were analyzed with unpaired t-tests, or with the Mann–Whitney test when sample size was limited (n = 3). For three or more groups, one-way ANOVA was used, followed by Tukey’s test for pairwise comparisons or Dunnett’s test when comparing to a non-supplemented control. Two-way ANOVA with multiple comparisons was applied for analyses across groups and time points. Data are shown as means ± SD, with significance set at p < 0.05. All experiments included at least triplicate samples (n ≥ 3).
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Adipocyte progenitor cells were isolated from pigs and cattle in accordance with approved protocols. Porcine cells were obtained from Tufts Comparative Medicine Services (CMS), while bovine cells were sourced from Tufts Cummings School of Veterinary Medicine, with approvals under IACUC protocols #B2021-32 and #G2023-65, respectively. All procedures followed the guidelines of the United States Department of Agriculture (USDA), Office of Laboratory Animal Welfare (OLAW), Massachusetts Department of Public Health (MDPH), and the Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC). Additionally, all methods were conducted in accordance with ARRIVE 2.0 guidelines.
Author contributions
NS: Conceptualization, Writing – original draft, Methodology, Data curation, Investigation, Formal analysis, Writing – review & editing. EL: Writing – review & editing, Methodology, Formal analysis. CR-G: Writing – review & editing. DL: Writing – review & editing, Data curation. XL: Data curation, Writing – review & editing. JY: Writing – review & editing, Methodology. TL: Writing – review & editing, Methodology. AK: Writing – review & editing, Methodology, Formal analysis, Data curation. RL: Writing – review & editing, Investigation. YM: Investigation, Writing – review & editing, Methodology. SF: Writing – original draft, Investigation, Writing – review & editing, Methodology. DK: Conceptualization, Writing – original draft, Supervision, Resources, Funding acquisition, Project administration, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. We thank the United States Department of Agriculture (2021–69012–35978).
Acknowledgments
We gratefully acknowledge the current and past members of the Kaplan Lab and the Tufts University Center for Cellular Agriculture. We thank the Metabolomics Core at UMass Chan Medical School (RRID: SCR_027036) for assistance with experimental design, metabolite extraction, LC–MS analysis, data processing and interpretation. We thank Courtney Bogins from Tufts Comparative Medicine Services (CMS) and Eugene C. White from Cummings School of Veterinary Medicine for their valuable contributions in providing porcine and bovine adipose tissues, respectively, which were essential for cell isolation in this study. We thank Anupam Abraham for his advice on aroma optimization. We are grateful to Ajinomoto Co., Inc. for support for NS. The graphical abstract was created using BioRender. Chemical structure was designed by MolView.
Conflict of interest
NS has been employed by Ajinomoto Co., Inc. JY has been employed by Deco Labs, Inc.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2025.1674183/full#supplementary-material
References
1. Treich, N. Cultured meat: promises and challenges. Environ Resour Econ (Dordr). (2021) 79:33–61. doi: 10.1007/s10640-021-00551-3
2. Zidarič, T, Milojević, M, Vajda, J, Vihar, B, and Maver, U. Cultured meat: meat industry hand in hand with biomedical production methods. Food Eng Rev. (2020) 12:498–519. doi: 10.1007/s12393-020-09253-w
3. Behrends, JM, Goodson, KJ, Koohmaraie, M, Shackelford, SD, Wheeler, TL, Morgan, WW, et al. Beef customer satisfaction: USDA quality grade and marination effects on consumer evaluations of top round steaks1. J Anim Sci. (2005) 83:662–70. doi: 10.2527/2005.833662x
4. Reicks, AL, Brooks, JC, Garmyn, AJ, Thompson, LD, Lyford, CL, and Miller, MF. Demographics and beef preferences affect consumer motivation for purchasing fresh beef steaks and roasts. Meat Sci. (2011) 87:403–11. doi: 10.1016/j.meatsci.2010.11.018
5. Legako, JF, Brooks, JC, O’Quinn, TG, Hagan, TDJ, Polkinghorne, R, Farmer, LJ, et al. Consumer palatability scores and volatile beef flavor compounds of five USDA quality grades and four muscles. Meat Sci. (2015) 100:291–300. doi: 10.1016/j.meatsci.2014.10.026
6. Arshad, MS, Sohaib, M, Ahmad, RS, Nadeem, MT, Imran, A, Arshad, MU, et al. Ruminant meat flavor influenced by different factors with special reference to fatty acids. Lipids Health Dis. (2018) 17:223. doi: 10.1186/s12944-018-0860-z
7. Olenic, M, and Thorrez, L. Cultured meat production: what we know, what we don’t know and what we should know. Ital J Anim Sci. (2023) 22:749–53. doi: 10.1080/1828051X.2023.2242702
8. Joo, S-T, Choi, J-S, Hur, S-J, Kim, G-D, Kim, C-J, Lee, E-Y, et al. A comparative study on the taste characteristics of satellite cell cultured meat derived from chicken and cattle muscles. Food Sci Anim Resour. (2022) 42:175–85. doi: 10.5851/kosfa.2021.e72
9. To, KV, Comer, CC, O’Keefe, SF, and Lahne, J. A taste of cell-cultured meat: a scoping review. Front Nutr. (2024) 11:1332765. doi: 10.3389/fnut.2024.1332765
10. Nishimura, T. Effect of lipids on food palatability: holding effect for aroma compounds and enhancement effect of koku attributes. J Lipid Nutr. (2021) 30:14–25. doi: 10.4010/jln.30.14
11. Drewnowski, A. Sensory properties of fats and fat replacements. Nutr Rev. (1992) 50:17–20. doi: 10.1111/j.1753-4887.1992.tb01285.x
12. Wojtasik-Kalinowska, I, Szpicer, A, Binkowska, W, Hanula, M, Marcinkowska-Lesiak, M, and Poltorak, A. Effect of processing on volatile organic compounds formation of meat—review. Appl Sci. (2023) 13:705. doi: 10.3390/app13020705
13. Mastrocola, D, Munari, M, Cioroi, M, and Lerici, CR. Interaction between Maillard reaction products and lipid oxidation in starch-based model systems. J Sci Food Agric. (2000) 80:684–90. doi: 10.1002/(SICI)1097-0010(20000501)80:6<684::AID-JSFA589>3.0.CO;2-3
14. Elmore, JS, Campo, MM, Enser, M, and Mottram, DS. Effect of lipid composition on meat-like model systems containing cysteine, ribose, and polyunsaturated fatty acids. J Agric Food Chem. (2002) 50:1126–32. doi: 10.1021/jf0108718
15. Yeo, H. Elucidation of the role of different sources of phospholipids in meat aroma formation. University of Reading (2022). Available online at: https://centaur.reading.ac.uk/111736/.
16. Lew, ET, Yuen, JSK, Zhang, KL, Fuller, K, Frost, SC, and Kaplan, DL. Chemical and sensory analyses of cultivated pork fat tissue as a flavor enhancer for meat alternatives. Sci Rep. (2024) 14:17643. doi: 10.1038/s41598-024-68247-4
17. Liu, P, Song, W, Bassey, AP, Tang, C, Li, H, Ding, S, et al. Preparation and quality evaluation of cultured fat. J Agric Food Chem. (2023) 71:4113–22. doi: 10.1021/acs.jafc.2c08004
18. Zhou, H, Loo, LSW, Ong, FYT, Lou, X, Wang, J, Myint, MK, et al. Cost-effective production of meaty aroma from porcine cells for hybrid cultivated meat. Food Chem. (2025) 473:142946. doi: 10.1016/j.foodchem.2025.142946
19. Yu, M, Li, Z, Rong, T, Wang, G, Liu, Z, Chen, W, et al. Different dietary starch sources alter the carcass traits, meat quality, and the profile of muscle amino acid and fatty acid in finishing pigs. J Anim Sci Biotechnol. (2020) 11:78. doi: 10.1186/s40104-020-00484-9
20. Nuernberg, K, Fischer, K, Nuernberg, G, Kuechenmeister, U, Klosowska, D, Eliminowska-Wenda, G, et al. Effects of dietary olive and linseed oil on lipid composition, meat quality, sensory characteristics and muscle structure in pigs. Meat Sci. (2005) 70:63–74. doi: 10.1016/j.meatsci.2004.12.001
21. Wang, D, Chen, G, Chai, M, Shi, C, Geng, Y, Che, Y, et al. Effects of dietary protein levels on production performance, meat quality and flavor of fattening pigs. Front Nutr. (2022) 9:910519. doi: 10.3389/fnut.2022.910519
22. Wang, D, Hou, K, Kong, M, Zhang, W, Li, W, Geng, Y, et al. Low-protein diet supplemented with amino acids can regulate the growth performance, meat quality, and flavor of the Bamei pigs. Foods. (2025) 14:946. doi: 10.3390/foods14060946
23. Thomas, C, Mercier, F, Tournayre, P, Martin, J-L, and Berdagué, J-L. Effect of added thiamine on the key odorant compounds and aroma of cooked ham. Food Chem. (2015) 173:790–5. doi: 10.1016/j.foodchem.2014.10.078
24. Broucke, K, Van Pamel, E, Van Coillie, E, Herman, L, and Van Royen, G. Cultured meat and challenges ahead: a review on nutritional, technofunctional and sensorial properties, safety and legislation. Meat Sci. (2023) 195:109006. doi: 10.1016/j.meatsci.2022.109006
25. Resconi, VC, Escudero, A, and Campo, MM. The development of aromas in ruminant meat. Molecules. (2013) 18:6748–81. doi: 10.3390/molecules18066748
26. Sun, A, Wu, W, Soladoye, OP, Aluko, RE, Bak, KH, Fu, Y, et al. Maillard reaction of food-derived peptides as a potential route to generate meat flavor compounds: a review. Food Res Int. (2022) 151:110823. doi: 10.1016/j.foodres.2021.110823
27. Butterworth, RF. Thiamin deficiency and brain disorders. Nutr Res Rev. (2003) 16:277–84. doi: 10.1079/NRR200367
28. Tang, W, Jiang, D, Yuan, P, and Ho, C-T. Flavor chemistry of 2-methyl-3-furanthiol, an intense meaty aroma compound. J Sulfur Chem. (2013) 34:38–47. doi: 10.1080/17415993.2012.708933
29. de Sousa Fontes, VM, de Sousa, GM, Moreira de Carvalho, L, do Nascimento Guedes, FL, Dos Santos Lima, M, Alencar Bezerra, TK, et al. Thiamine, cysteine and xylose added to the Maillard reaction of goat protein hydrolysate potentiates the formation of meat flavoring compounds. Food Chem. (2024) 445:138398. doi: 10.1016/j.foodchem.2024.138398
30. Li, L, Perea-Sanz, L, Salvador, A, Belloch, C, and Flores, M. Understanding the impact of nitrogen and sulfur precursors on the aroma of dry fermented sausages. Meat Sci. (2022) 192:108896. doi: 10.1016/j.meatsci.2022.108896
31. Albrecht, E, Teuscher, F, Ender, K, and Wegner, J. Growth- and breed-related changes of marbling characteristics in cattle. J Anim Sci. (2006) 84:1067–75. doi: 10.2527/2006.8451067x
32. Specht, K, and Baltes, W. Identification of volatile flavor compounds with high aroma values from shallow-fried beef. J Agric Food Chem. (1994) 42:2246–53. doi: 10.1021/jf00046a031
33. Lee, M, and Kim, Y-S. Analysis of volatile and odor-active compounds in charcoal-grilled marinated beef using gas chromatography-mass spectrometry and gas chromatography-olfactometry. Food Sci Biotechnol. (2025) 34:1339–49. doi: 10.1007/s10068-024-01783-x
34. Shakoor, A, Al-Dalali, S, Xie, J, Zhang, C, and Hossen, I. Insight into the effect of GSH curing treatment on the flavor formation of chicken meat. Food Chem. (2025) 468:142488. doi: 10.1016/j.foodchem.2024.142488
35. Wang, X, Huang, M, Yao, Y, Yu, J, Cui, H, Hayat, K, et al. Difference comparison of characteristic aroma compounds between braised pork cooked by traditional open-fire and induction cooker and the potential formation cause under electromagnetic cooking. Food Res Int. (2024) 188:114506. doi: 10.1016/j.foodres.2024.114506
36. Love, JD. The role of heme iron in the oxidation of lipids in red meats. Food Technol. (1983) 12:117–20.
37. Devaere, J, De Winne, A, Dewulf, L, Fraeye, I, Šoljić, I, Lauwers, E, et al. Improving the aromatic profile of plant-based meat alternatives: effect of myoglobin addition on volatiles. Foods. (2022) 11:1985. doi: 10.3390/foods11131985
38. Oki, Y, Hagiwara, R, Matsumaru, T, and Kano, K. Effect of volatile fatty acids on adipocyte differentiation in bovine dedifferentiated fat (DFAT) cells in vitro. Genes Cells. (2022) 27:5–13. doi: 10.1111/gtc.12903
39. Yuen, JSK, Saad, MK, Xiang, N, Barrick, BM, DiCindio, H, Li, C, et al. Aggregating in vitro-grown adipocytes to produce macroscale cell-cultured fat tissue with tunable lipid compositions for food applications. eLife. (2023) 12:2120. doi: 10.7554/eLife.82120
40. Feng, Y, Luo, J, Cheng, J, Xu, A, Qiu, D, He, S, et al. A small-molecule cocktails-based strategy in culture of mesenchymal stem cells. Front Bioeng Biotechnol. (2022) 10:819148. doi: 10.3389/fbioe.2022.819148
41. Peng, X, Song, T, Hu, X, Zhou, Y, Wei, H, Peng, J, et al. Phenotypic and functional properties of porcine dedifferentiated fat cells during the long-term culture in vitro. Biomed Res Int. (2015) 2015:673651. doi: 10.1155/2015/673651
42. Jurek, S, Sandhu, MA, Trappe, S, Bermúdez-Peña, MC, Kolisek, M, Sponder, G, et al. Optimizing adipogenic transdifferentiation of bovine mesenchymal stem cells: a prominent role of ascorbic acid in FABP4 induction. Adipocytes. (2020) 9:35–50. doi: 10.1080/21623945.2020.1720480
43. Dohmen, RGJ, Hubalek, S, Melke, J, Messmer, T, Cantoni, F, Mei, A, et al. Muscle-derived fibro-adipogenic progenitor cells for production of cultured bovine adipose tissue. NPJ Sci Food. (2022) 6:6. doi: 10.1038/s41538-021-00122-2
44. Mitić, R, Cantoni, F, Börlin, CS, Post, MJ, and Jackisch, L. A simplified and defined serum-free medium for cultivating fat across species. iScience. (2023) 26:105822. doi: 10.1016/j.isci.2022.105822
45. Lee, BC, and Gladyshev, VN. The biological significance of methionine sulfoxide stereochemistry. Free Radic Biol Med. (2011) 50:221–7. doi: 10.1016/j.freeradbiomed.2010.11.008
46. Liu, L, Liu, B, Cui, Y, Wu, Y, and Ouyang, J. Maillard products formed from chicken fat oxidants and amino acids act as a flavor enhancer in comminuted chicken products. J Food Sci Eng. (2013) 3:11.
47. Api, AM, Belsito, D, Botelho, D, Bruze, M, Burton, GA, Buschmann, J, et al. RIFM fragrance ingredient safety assessment, 4-methyl-5-thiazoleethanol, CAS registry number 137-00-8. Food Chem Toxicol. (2020) 144:111530. doi: 10.1016/j.fct.2020.111530
48. Tamura, Y, Iwatoh, S, Miyaura, K, Asikin, Y, and Kusano, M. Metabolomic profiling reveals the relationship between taste-related metabolites and roasted aroma in aged pork. Lebenson Wiss Technol. (2022) 155:112928. doi: 10.1016/j.lwt.2021.112928
49. Fang, Y, Zhang, J, Ma, C, Xing, L, Wang, W, and Zhang, W. Ultrasound-induced modifications of beef flavor characteristics during postmortem aging. Ultrason Sonochem. (2024) 108:106979. doi: 10.1016/j.ultsonch.2024.106979
50. Park, MK, Kim, B-G, Kang, M-C, Kim, T-K, and Choi, Y-S. Distinctive volatile compound profile of different raw meats, including beef, pork, chicken, and duck, based on flavor map. Appl Food Res. (2025) 5:100655. doi: 10.1016/j.afres.2024.100655
51. Song, S, Tang, Q, Hayat, K, Karangwa, E, Zhang, X, and Xiao, Z. Effect of enzymatic hydrolysis with subsequent mild thermal oxidation of tallow on precursor formation and sensory profiles of beef flavours assessed by partial least squares regression. Meat Sci. (2014) 96:1191–200. doi: 10.1016/j.meatsci.2013.11.008
52. Goodridge, CF, Beaudry, RM, Pestka, JJ, and Smith, DM. Solid phase microextraction-gas chromatography for quantifying headspace hexanal above freeze-dried chicken myofibrils. J Agric Food Chem. (2003) 51:4185–90. doi: 10.1021/jf0260646
53. Zhang, M, Chen, X, Hayat, K, Duhoranimana, E, Zhang, X, Xia, S, et al. Characterization of odor-active compounds of chicken broth and improved flavor by thermal modulation in electrical stewpots. Food Res Int. (2018) 109:72–81. doi: 10.1016/j.foodres.2018.04.036
54. Nobusue, H, and Kano, K. Establishment and characteristics of porcine preadipocyte cell lines derived from mature adipocytes. J Cell Biochem. (2010) 109:542–52. doi: 10.1002/jcb.22431
55. Song, X, Li, Y, Chen, X, Yin, G, Huang, Q, Chen, Y, et al. bFGF promotes adipocyte differentiation in human mesenchymal stem cells derived from embryonic stem cells. Genet Mol Biol. (2014) 37:127–34. doi: 10.1590/S1415-47572014000100019
56. Zaragosi, L-E, Ailhaud, G, and Dani, C. Autocrine fibroblast growth factor 2 signaling is critical for self-renewal of human multipotent adipose-derived stem cells. Stem Cells. (2006) 24:2412–9. doi: 10.1634/stemcells.2006-0006
57. Quek, JP, Gaffoor, AA, Tan, YX, Tan, TRM, Chua, YF, Leong, DSZ, et al. Exploring cost reduction strategies for serum free media development. Npj Sci Food. (2024) 8:107. doi: 10.1038/s41538-024-00352-0
58. Lengefeld, J, Cheng, C-W, Maretich, P, Blair, M, Hagen, H, McReynolds, MR, et al. Cell size is a determinant of stem cell potential during aging. Sci Adv. (2021) 7:eabk0271. doi: 10.1126/sciadv.abk0271
59. Mitsui, Y, and Schneider, EL. Relationship between cell replication and volume in senescent human diploid fibroblasts. Mech Ageing Dev. (1976) 5:45–56. doi: 10.1016/0047-6374(76)90007-5
60. Zhang, Y, Huang, C, Sheng, X, Gong, Z, and Zang, YQ. Lecithin promotes adipocyte differentiation and hepatic lipid accumulation. Int J Mol Med. (2009) 23:449–54.
61. Pasitka, L, Cohen, M, Ehrlich, A, Gildor, B, Reuveni, E, Ayyash, M, et al. Spontaneous immortalization of chicken fibroblasts generates stable, high-yield cell lines for serum-free production of cultured meat. Nat Food. (2023) 4:35–50. doi: 10.1038/s43016-022-00658-w
62. Intralipid. (2022). Available online at: https://www.accessdata.fda.gov/drugsatfda_docs/label/2022/020248s026lbl.pdf.
63. Zamora, R, Alcón, E, and Hidalgo, FJ. Effect of amino acids on the formation of 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) in creatinine/phenylalanine and creatinine/phenylalanine/4-oxo-2-nonenal reaction mixtures. Food Chem. (2013) 141:4240–5. doi: 10.1016/j.foodchem.2013.06.036
64. Bin, P, Huang, R, and Zhou, X. Oxidation resistance of the sulfur amino acids: methionine and cysteine. Biomed Res Int. (2017) 2017:9584932. doi: 10.1155/2017/9584932
65. Brewer, SM. The chemistry of beef flavor. (2006). Available online at: https://www.beefresearch.org/Media/BeefResearch/Docs/the_chemistry_of_beef_flavor_08-20-2020-98.pdf.
66. Post, MJ. Proteins in cultured beef In: MJ Post, editor. Proteins in food processing. Amsterdam, Netherlands: Elsevier (2018). 289–98.
67. Song, S, Zhang, X, Hayat, K, Liu, P, Jia, C, Xia, S, et al. Formation of the beef flavour precursors and their correlation with chemical parameters during the controlled thermal oxidation of tallow. Food Chem. (2011) 124:203–9. doi: 10.1016/j.foodchem.2010.06.010
68. Yao, L, Mo, Y, Chen, D, Feng, T, Song, S, Wang, H, et al. Characterization of key aroma compounds in Xinjiang dried figs (Ficus carica L.) by GC–MS, GC–olfactometry, odor activity values, and sensory analyses. Lebenson Wiss Technol. (2021) 150:111982. doi: 10.1016/j.lwt.2021.111982
69. Mattick, LR, Patton, S, and Keeney, PG. The coconut-like flavor defect from milk fat. III. Observations on the origin of δ-decalactone in fat-containing dairy products. J Dairy Sci. (1959) 42:791–8. doi: 10.3168/jds.S0022-0302(59)90655-1
70. Shahidi, F, and Pegg, RB. Hexanal as an indicator of meat flavor deterioration. J Food Lipids. (1994) 1:177–86. doi: 10.1111/j.1745-4522.1994.tb00245.x
71. Dong, T, Tian, Z, Wang, S, Sun, J, Chen, H, Wang, S, et al. Identification of key off-flavor compounds during storage of fried pepper (Zanthoxylum bungeanum maxim.) oils by sensory-directed flavor analysis and partial least squares regression (PLSR). J Food Compost Anal. (2024) 131:106268. doi: 10.1016/j.jfca.2024.106268
72. Brewer, MS, and Vega, JD. Detectable odor thresholds of selected lipid oxidation compounds in a meat model system. J Food Sci. (1995) 60:592–5. doi: 10.1111/j.1365-2621.1995.tb09834.x
73. U.S. Food and Drug Administration. GRAS notice no. GRN 000737. (2017). Available online at: https://www.fda.gov/media/116243/download.
74. Simsa, R, Yuen, J, Stout, A, Rubio, N, Fogelstrand, P, and Kaplan, DL. Extracellular heme proteins influence bovine myosatellite cell proliferation and the color of cell-based meat. Foods. (2019) 8:521. doi: 10.3390/foods8100521
75. Jiang, S, Xue, D, Li, Q, Shan, K, Zhang, M, Zhou, G, et al. Exploring the impact of myoglobin from red meat on intestinal function: insights from mouse and cell models. Food Front. (2024) 5:2672–90. doi: 10.1002/fft2.472
76. Liu, Y, Fang, Y, Wang, Z, Guo, L, Xiao, J, and Li, X. Formation and distribution of (E,E)-2,4-decadienal in deep-fried dough sticks under different conditions. Eur J Lipid Sci Technol. (2024) 126:2300194. doi: 10.1002/ejlt.202300194
77. Zhang, Q, Ke, J, Long, P, Wen, M, Han, Z, Zhang, L, et al. Formation of furan from linoleic acid thermal oxidation: (E,E)-2,4-decadienal as a critical intermediate product. J Agric Food Chem. (2024) 72:4384–92. doi: 10.1021/acs.jafc.3c08604
78. Elmore, JS, Warren, HE, Mottram, DS, Scollan, ND, Enser, M, Richardson, RI, et al. A comparison of the aroma volatiles and fatty acid compositions of grilled beef muscle from Aberdeen Angus and Holstein-Friesian steers fed diets based on silage or concentrates. Meat Sci. (2004) 68:27–33. doi: 10.1016/j.meatsci.2004.01.010
79. Song, S, Tang, Q, Fan, L, Xu, X, Song, Z, Hayat, K, et al. Identification of pork flavour precursors from enzyme-treated lard using Maillard model system assessed by GC-MS and partial least squares regression. Meat Sci. (2017) 124:15–24. doi: 10.1016/j.meatsci.2016.10.009
80. Cantor, JR, Abu-Remaileh, M, Kanarek, N, Freinkman, E, Gao, X, Louissaint, A, et al. Physiologic medium rewires cellular metabolism and reveals uric acid as an endogenous inhibitor of UMP synthase. Cell. (2017) 169:258–272.e17. doi: 10.1016/j.cell.2017.03.023
Keywords: cellular agriculture, cultivated meat, fat, adipogenesis, flavor, aroma, sensory
Citation: Sugama N, Lew ET, Riquelme-Guzmán C, Lee DS, Li X, Yuen JSK Jr, Lim T, Kwan A, Liu RY, Ma YA, Frost SC and Kaplan DL (2025) Modulation of nutritional composition and aroma volatiles in cultivated pork fat by culture media supplementation. Front. Nutr. 12:1674183. doi: 10.3389/fnut.2025.1674183
Edited by:
Jean-Francois Hocquette, INRA UMR1213 Unité Mixte de Recherche sur les Herbivores (UMRH), FranceReviewed by:
Yin Haowen, Guangdong Ocean University, ChinaXingchen Li, The Ohio State University, United States
Copyright © 2025 Sugama, Lew, Riquelme-Guzmán, Lee, Li, Yuen, Lim, Kwan, Liu, Ma, Frost and Kaplan. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: David L. Kaplan, ZGF2aWQua2FwbGFuQHR1ZnRzLmVkdQ==