 Crystal Reeck1,2
Crystal Reeck1,2 Tobias Egner1,2*
Tobias Egner1,2*- 1 Center for Cognitive Neuroscience, Duke University, Durham, NC, USA
- 2 Department of Psychology and Neuroscience, Duke University, Durham, NC, USA
Numerous theories posit that affectively salient stimuli are privileged in their capacity to capture attention and disrupt ongoing cognition. Two underlying assumptions in this theoretical position are that the potency of affective stimuli transcends task boundaries (i.e., emotional distracters do not have to belong to a current task-set to disrupt processing) and that there is an asymmetry between emotional and cognitive processing (i.e., emotional distracters disrupt cognitive processing, but not vice versa). These assumptions have remained largely untested, as common experimental probes of emotion–cognition interaction rarely manipulate task-relevance and only examine one side of the presumed asymmetry of interference. To test these propositions directly, a face–word Stroop protocol was adapted to independently manipulate (a) the congruency between target and distracter stimulus features, (b) the affective salience of distracter features, and (c) the task-relevance of emotional compared to non-emotional target features. A three-way interaction revealed interdependent effects of distracter relevance, congruence, and affective salience. Compared to task-irrelevant distracters, task-relevant congruent distracters facilitated performance and task-relevant incongruent distracters impaired performance, but the latter effect depended on the nature of the target feature and task. Specifically, task-irrelevant emotional distracters resulted in equivalent performance costs as task-relevant non-emotional distracters, whereas task-irrelevant non-emotional distracters did not produce performance costs comparable to those generated by task-relevant emotional distracters. These results document asymmetric cross-task interference effects for affectively salient stimuli, supporting the notion of affective prioritization in human information processing.
Introduction
Executive control enables organisms to act in accordance with internal goals, promoting the processing of information relevant to current objectives while mitigating distraction from irrelevant information. Several critical functions underlie this ability, including top-down attentional biasing that enhances the processing of goal- or task-relevant information relative to task-irrelevant information (Desimone and Duncan, 1995; Miller and Cohen, 2001). However, both physically salient (Yantis and Jonides, 1990) and affectively salient stimuli are adept at capturing attention in a bottom-up fashion, and may therefore disrupt ongoing goal-oriented processing (LeDoux, 2000; Ohman and Mineka, 2001; Vuilleumier and Huang, 2009). While the privileged access to processing resources exhibited by affectively salient stimuli is essential to rapid responding to stimuli that may convey a potential threat or possible reward, if affective stimuli are too adept at disrupting ongoing mental functions, harmful consequences may result. Indeed, many models of psychiatric disorders have disruptions in the interaction between executive function and affective processing at their core (Bishop, 2007; Banich et al., 2009), highlighting the necessity of characterizing the interactive influence between these two processes.
Theories positing that affectively salient items have privileged access during information processing (LeDoux, 2000; Ohman and Mineka, 2001; Vuilleumier and Huang, 2009) typically entail two key implicit assumptions regarding the properties of emotional stimuli. The first is that affective stimuli have the capacity to transcend task boundaries, disrupting ongoing processing regardless of whether they are relevant to the current task-set of the organism or not. The second is an assumption of asymmetry, whereby affective information interferes with non-affective task-sets more potently than non-affective information interferes with affective task-sets. Importantly, while these two presuppositions provide the foundation for the hypothesized privileged access of affective stimuli to attention, they are rarely ever tested, as common experimental probes of emotion–cognition interaction generally do not manipulate the task-relevance of affective stimuli or the potential for non-affective distracters to interfere with affective processing. The goal of the current experiment was to explicitly test these two assumptions.
The question of affective influences on cognitive processing has prompted the development of experimental paradigms seeking to examine affective modulation of executive function, particularly its influence on attention as gaged in classic conflict processing tasks, such as the Stroop paradigm (Stroop, 1935; Macleod, 1991). In the color–word Stroop task, color words (e.g., “red”) are printed in an ink color (e.g., the color red), and the participant identifies the color of the ink while ignoring the semantic meaning of the word. Thus, the ink color is the target, whereas the semantic meaning of the word is the distracter. Typically, performance is both slower and more error prone when the distracter is semantically incongruent with the target (e.g., the word “green” printed in red ink), likely due to the fact that these two features are both semantically conflicting and prime mutually exclusive responses (Kornblum et al., 1990; Macleod, 1991). A well-known adaptation of the traditional Stroop paradigm to the affective domain includes non-affective (e.g., “car”) and affectively salient (e.g., “death”) words as irrelevant stimulus features, and the potency of emotional distracters is gaged by comparing response times (RTs) between neutral and affective distracter conditions (Mathews and Macleod, 1985; McKenna, 1986; Whalen et al., 1998; Isenberg et al., 1999; Compton et al., 2003). However, this adaptation suffers from several conceptual and practical limitations. First, it does not really constitute an affective equivalent of the classic Stroop task, as the inclusion of affectively salient stimuli in the distracting feature dimension may capture attention but it does not produce semantic or response conflict with the target feature (Algom et al., 2004). For instance, the semantic meaning of a negative affective word is not directly incongruent with the ink color in the same way that the semantic meaning of a color–word would be, nor do affective words prime competing responses in this case. Second, although this adaption could in principle gauge the capacity of task-irrelevant affective stimuli to interfere with an ongoing non-affective task-set (color-naming), it does not probe whether this type of interference is asymmetrical. Finally, if this protocol tests the capacity of affective stimuli to interfere with a non-affective task-set, results obtained with this task should actually raise doubts as to whether affective information is in fact capable of transcending task boundaries, as behavioral findings of an “emotional Stroop effect” have been highly inconsistent, especially in healthy subject populations (McKenna, 1986; Williams et al., 1996; Whalen et al., 1998; Isenberg et al., 1999; Compton et al., 2003).
An alternative approach has been to design tasks where an affective distracter could conflict directly with the target of the ongoing task-set, by combining affective distracters with affective targets and task-sets. One approach is a modified face–word Stroop task, in which participants must make a judgment about a visually presented face image while ignoring an overlaid word label. In affective versions of this task, the faces are modeling affective expressions and the participant must categorize the expression (e.g., happy or fearful) while ignoring an affective word label (e.g., “happy,” “fear”; Etkin et al., 2006; Egner et al., 2008). Therefore, the distracter and the target are semantically related, and incongruent distracters would likely generate both stimulus and response conflict. Investigations using this type of paradigm reliably report significant behavioral slowing on incongruent trials (Etkin et al., 2006; Haas et al., 2006; Egner et al., 2008; Ochsner et al., 2009), indicative of a robust effect of emotional conflict. However, this type of protocol does not examine the interaction between affective salience and task-set relevance, leaving the assumption that affective stimuli asymmetrically interfere with non-affective task-sets unexamined. Thus, although tasks employing affective targets, distracters, and task-sets have expanded our knowledge about emotional conflict processing, they preclude the investigation of interactions between task-sets, target features, and affective salience.
In sum, previous paradigms for investigating the impact of affective distracters on non-affective task-sets have produced inconsistent results regarding the capacity of affective distracters to transcend task-set boundaries, whereas tasks employing affective task-sets have only shown a reliable capacity of affective distracters to disrupt performance during an affective task-set. Neither type of protocol has tested whether interference effects across task-sets are asymmetrical in nature, with affective stimuli having a more potent effect than non-affective information. The extent to which affective stimuli are prioritized in human information processing therefore remains unclear.
The current study sought to examine the effects of distracter task-set relevance and affective salience on behavioral performance. A variant of the face–word Stroop task was adapted to independently manipulate distracter congruence with the target stimulus as well as membership in the current task-set, which could involve either an affective or a non-affective judgment of the target. This manipulation enabled the examination of the relative capacity of both affective and non-affective stimuli to transcend task boundaries in altering performance.
Materials and Methods
Participants
Thirty-eight healthy college students (Mean Age 19.0 years, SD 1.0; 27 women) participated in this study for course credit. All participants were fluent in English and had normal or corrected-to-normal vision. Participants were screened via self-report to exclude those with previous or current psychiatric or neurologic conditions. All participants gave informed consent and this study was approved for use in human subjects in accordance with the Duke University Medical Center Institutional Review Board.
Materials
Stimuli consisted of black and white face images from the NimStim Set of Facial Expressions1 (Tottenham et al., 2009) overlaid with word labels. The stimulus set consisted of four female and four male individuals posing both fearful and happy expressions, resulting in a total of 16 distinct face stimuli. Images were cropped to an oval that consisted of the main facial features to standardize image size and shape across the different identities and expressions. Additionally, image cropping removed non-face information, such as hair, which could facilitate performance during gender discrimination. There were eight different versions of each of the 16 distinct face stimuli, as each face was paired with gender (“male,” “female”) and expression (“fear,” “happy”) word labels that were superimposed on the center of the face images (in Helvetica font and red ink), and which could be displayed in lower or upper case lettering. Thus, the complete stimulus set consisted of 128 unique face–word compound stimuli. Four example stimuli are displayed in Figure 1. The face images subtended approximately 12.4° of visual angle vertically and 7.2° horizontally. The labels subtended approximately 1.4° of visual angle vertically and 3.8°–8.1° horizontally. Stimulus presentation and data collection were performed using Psychophysics Toolbox Version 3 (Brainard, 1997; Pelli, 1997) on Dell Optiplex 960 computers with Dell 19″ LCD monitors. A chin rest was use to ensure that participants maintained a distance of approximately 60 cm from the monitor throughout the experiment.

Figure 1. Examples of experimental stimuli for (A) the emotion expression task and (B) the gender task. The experiment varied distracter congruency and relevance by manipulating the labels superimposed on the face stimuli. The labels were printed in red ink in the experiment, but have been presented in white ink here for display purposes.
Procedure
After providing informed consent, participants were randomly assigned to either a gender task or an emotion expression task in a face–word Stroop paradigm adapted from previous work (Etkin et al., 2006; Egner et al., 2008). On each trial, participants were presented with a compound face–word stimulus and instructed to make a judgment of the face image as quickly and accurately as possible while ignoring the word label. Participants assigned to the gender task made a gender judgment (male or female) while those assigned to the emotion expression task made an expression judgment (fearful or happy). Responses were made via keyboard presses using the index and middle fingers of the right hand. Stimuli were presented for 1 s and followed by a variable inter-trial fixation interval of 2, 3, or 4 s drawn from a uniform distribution. Following a brief practice, participants completed three runs of 145 trials each, with the first trial in each run serving as a filler to mitigate any preparatory effects.
Similar to previous versions of this task (Etkin et al., 2006; Egner et al., 2008), the labels presented superimposed on the images could be either semantically congruent or incongruent with the target face image. For instance, a fearful male face could be accompanied by a congruent “male” (or “fear”) label or by an incongruent “female” (or “happy”) label. In a departure from earlier versions, however, the labels were drawn not only from the semantic category that was relevant to the current task-set (e.g., gender labels presented during the gender task) but also from the category that was irrelevant to the current task-set (e.g., emotion labels presented during the gender task; Figure 1). We refer to the former as “task-relevant distracters” and to the latter as “task-irrelevant distracters.” Importantly, task-relevant distracters could be associated with both (semantic) stimulus conflict and response conflict, because their semantic meaning could clash with the face gender/emotion (potentially inducing stimulus conflict), and in addition their meaning corresponded to a valid response option in the task-set (potentially eliciting response conflict). By contrast, task-irrelevant distracters could not generate response conflict, as they had no correspondence with valid response options in the task-set. Task-irrelevant distracters, however, could be associated with (task-irrelevant) semantic or stimulus conflict, and one of the main goals of this study was to determine whether task-irrelevant stimulus congruency could in fact affect responses to task-relevant stimulus features, and whether this type of effect would be dependent on the affective nature of the distracters. It should be emphasized that the potential semantic or stimulus conflict elicited in the current experiment would be due to completely task-irrelevant distracters and is thus different from task-relevant stimulus conflict that can be evoked, for example, by including response-ineligible colors in a regular color-naming Stroop task (Milham et al., 2001), or by employing many-to-few stimulus–response mappings in flanker or Stroop paradigms (De Houwer, 2003).
Each experimental run was balanced to include equal numbers of trials in each of the four congruence and relevance conditions (i.e., congruent–relevant, congruent–irrelevant, incongruent–relevant, incongruent–irrelevant). Face stimuli in each condition were equally likely to be male or female, fearful or happy. Additionally, the trial order was pseudo-randomized to ensure that there were an equal number of trial transitions between each of the four main conditions. Factors that could impact experimental effects of distracter congruence and relevance, such as repetition priming (Mayr et al., 2003) or cross-trial feature binding effects (Hommel, 1998), were controlled by preventing any repetitions of the same face identity on consecutive trials and alternating each trial between uppercase and lowercase distracter word labels. Thus, no exact perceptual features of either the target or the distracter ever repeated across successive trials.
Analysis
Analyses of categorization accuracy and RT were conducted using SPSS (SPSS Inc., Chicago, IL, USA). RT data exclude error trials and post-error trials. Outlier RT values that were below or above 2 SDs from the participant’s grand mean were also removed, resulting in the exclusion of 4.8% of trials on average (SD 0.9%). Two participants whose RTs were more than 2 SDs higher than the sample average in all four RT conditions were excluded from subsequent analyses. The final sample sizes were therefore 19 participants (13 female) completing the emotion expression task and 17 participants (12 female) completing the gender task. Response accuracy and RT data were submitted to separate 2 × 2 × 2 repeated-measures ANOVAs with distracter congruence (congruent versus incongruent) and task-relevance (relevant or irrelevant) as within-subjects factors and task (gender or emotion expression) as a between-subjects factor. Significant results in the main ANOVAs were examined using t-tests and ANOVAs. Means and SDs for participant RT and accuracy are reported for each condition in Tables 1 and 2 respectively.

Table 1. Means and SDs of participant response times (ms).
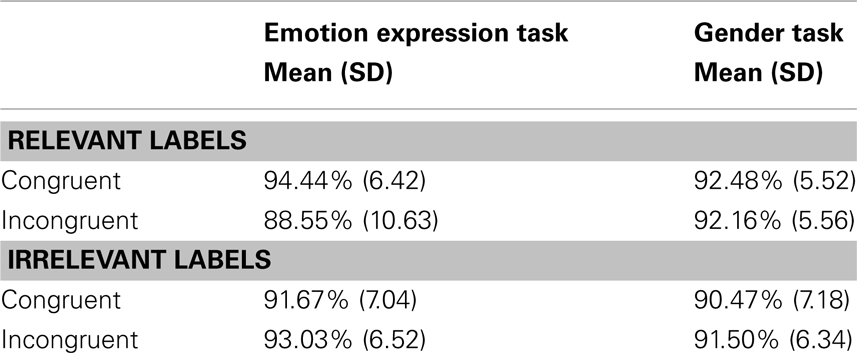
Table 2. Means and SDs of participant accuracy.
Results
Response Time Data
Analyses of RT data revealed a main effect of distracter congruence [F(1,34) = 24.4, p < 0.001], with slower responses to incongruent than congruent stimuli. There were no significant main effects of either distracter relevance or task (p’s > 0.8). However, distracter relevance interacted with distracter congruence [F(1,34) = 26.1, p < 0.001], as the congruency effect was significant only for relevant labels [t(35) = 6.9, p < 0.001] but not irrelevant labels (p > 0.8). Importantly, there was a three-way interaction between distracter congruence, distracter relevance, and task [F(1,34) = 5.6, p = 0.024, Figure 2]. To explore the three-way interaction, 2 × 2 repeated-measures ANOVAs were conducted with distracter relevance as a within-subjects factor and task as a between-subjects factor separately for congruent and incongruent trials. On congruent trials, the main effect of relevance was significant [F(1,34) = 19.0, p < 0.001], with relevant labels resulting in faster responses than irrelevant labels. Thus, for both tasks, congruent distracters facilitated performance to a greater extent when the distracter was relevant to the current task-set. Neither the main effect of task nor the interaction between distracter relevance and task was significant (p’s > 0.1). On incongruent trials, the main effect of relevance was also significant [F(1,34) = 13.0, p = 0.001], as relevant labels resulted in slower responses than irrelevant labels. Critically, the interaction between distracter relevance and task was significant [F(1,34) = 4.4, p = 0.043]. Participants completing the emotion expression task exhibited slower RTs on incongruent trials with task-set relevant labels (i.e., affective labels) than those with task-set irrelevant labels (i.e., gender labels) [t(18) = 3.5, p = 0.002]. However, participants completing the gender task exhibited equally slow RTs on incongruent trials with task-set relevant and irrelevant labels (p > 0.15), indicating that the incongruent affective labels interfered with their performance despite their irrelevance to the task-set. Furthermore, RTs did not differ across the congruent and incongruent–irrelevant trials (p > 0.4), indicating that affective labels in general interfered with task performance, regardless of congruency. Thus, whereas gender labels did not affect performance during the emotion expression task, affective labels produced marked impairments of performance during the gender task, consistent with the capacity of affective stimuli to capture attention irrespective of their relevance to an ongoing task-set.
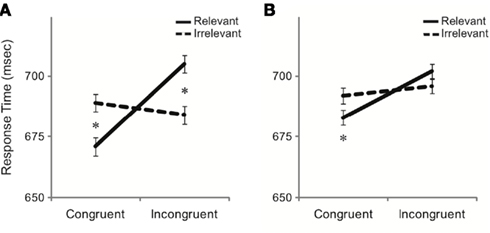
Figure 2. Mean response times by distracter relevance and congruence presented separately for (A) the emotion expression task and (B) the gender task. Error bars represent SE of the mean.
Accuracy
Analyses of response accuracy revealed no significant main effects of distracter relevance, distracter congruence, or task (p’s > 0.05). There were significant interactions between distracter relevance and task [F(1,34) = 4.2, p = 0.049], distracter congruence and task [F(1,34) = 7.0, p = 0.012], and distracter relevance and congruence [F(1,34) = 13.2, p = 0.001]. However, these interactions were qualified by a significant three-way interaction of distracter relevance, distracter congruence, and task [F(1,34) = 6.2, p = 0.018, Figure 3]. To investigate this three-way interaction, 2 × 2 repeated-measures ANOVAs were conducted with distracter relevance as a within-subjects factor and task as a between-subjects factor separately for congruent and incongruent trials. On congruent trials, the main effect of relevance was significant [F(1,34) = 14.0, p = 0.001], with higher accuracy on relevant compared to irrelevant congruent trials. Relevant congruent distracters facilitated performance relative to irrelevant congruent distracters on both tasks. There was no main effect of task nor was there a significant interaction between distracter relevance and task (p’s > 0.5). On incongruent trials, there was a main effect of relevance [F(1,34) = 4.2, p = 0.047], with lower accuracy in response to relevant than irrelevant incongruent distracters. Importantly, there was an interaction between distracter relevance and task [F(1,34) = 7.6, p = 0.009]. Paired t-tests comparing accuracy on irrelevant and relevant incongruent trials showed a significant difference for participants performing the emotion expression task [t(18) = 2.9, p = 0.009], such that accuracy was lower on relevant compared to irrelevant incongruent trials. No such difference was found for participants performing the gender task (p > 0.5). Accuracy on irrelevant congruent and incongruent stimuli did not differ (p > 0.2), indicating that affective labels interfered with performance, irrespective of their congruency, and despite being irrelevant to the current task-set. Consistent with the results from analyses of RT, these findings indicate that the irrelevant emotion labels in the gender task asymmetrically disrupt task performance compared to the irrelevant gender labels in the emotion expression task.
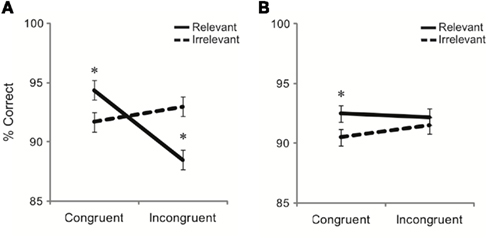
Figure 3. Mean judgment accuracy by distracter relevance and congruence presented separately for (A) the emotion expression task and (B) the gender task. Error bars represent SE of the mean.
Effects of Target Valence
The observed interaction between task-set, distracter relevance, and affective salience could additionally be modulated by the affective content of the target stimulus. Although all the target face stimuli modeled an affective expression (either happiness or fear) and were counterbalanced across the cells of the factorial design, the specific affective valence of the target stimuli could nevertheless interact with observed effects of distracter relevance and affective salience. To interrogate this possibility, additional analyses were conducted in which trials were classified based on target valence (happy or fear expression) as well as distracter relevance and congruence. These analyses revealed no interactions between target valence and the distracter relevance by congruence by task-set interaction described above for either RT or accuracy (p’s > 0.25). The present findings, therefore, appear to be driven by the affective nature of the distracters and their relevance to the current task-set as opposed to differences in affective valence of the targets in this task. The absence of an effect of target valence is consistent with the findings from a number of previous studies of emotion–cognition interactions (Compton et al., 2003; Etkin et al., 2006; Haas et al., 2006; Egner et al., 2008; Ochsner et al., 2009).
Discussion
The present study adapted a face–word Stroop paradigm to investigate two underlying assumptions of theories postulating prioritized processing of affective stimuli: that affective stimuli can disrupt processing across task boundaries and that this capacity is not shared by non-affective stimuli. Analyses of the interaction between distracter congruence, distracter affective salience, and task-set relevance revealed that each of these factors modulates task performance. While the basic pattern of responses in the two task-sets was quite similar, the analyses revealed some significant distinctions: for both affective and non-affective task-sets, incongruent distracters relevant to the current task-set resulted in slower RTs and lower judgment accuracy compared to congruent distracters, due to the generation of semantic and response conflict. However, the influence of distracters outside the task-set on performance was driven by an interaction between affective salience and task-set. In the presence of a non-affective task-set, gender categorization, affective labels markedly slowed RTs and reduced judgment accuracy despite their irrelevance to the present task-set, demonstrating the capacity of affectively salient stimuli to disrupt ongoing processing and transcend task boundaries. This effect was specific to the task-irrelevant affective distracters, as non-affective, task-irrelevant distracters did not similarly impair performance in the presence of an affective task-set, the emotion expression categorization. Furthermore, this effect was not dependent on the congruency of the affective word distracter with respect to the equally task-irrelevant affective facial expression, indicating that task-irrelevant affective salience interfered with task performance regardless of congruency. In sum, affectively salient distracter stimuli, but not non-emotional distracter stimuli, interfered with ongoing performance despite their irrelevance to current task-set, indicating that affective stimuli have the capacity to transcend task boundaries and disrupt ongoing executive processes.
The present findings extend those from previous investigations of the interaction between affective processing and executive function. Previous adaptations of the Stroop paradigm utilizing task-irrelevant affective distracters and non-affective task-sets (such as adaptations of the color–word Stroop) examined whether affective stimuli could disrupt ongoing non-affective top-down processing but have met with mixed results (Mathews and Macleod, 1985; McKenna, 1986; Whalen et al., 1998; Isenberg et al., 1999; Compton et al., 2003). Other adaptations utilizing task-relevant affective distracters in the presences of affective task-sets exhibited robust interference effects but did not speak to the capacity of affective stimuli to disrupt non-affective processing (Etkin et al., 2006; Egner et al., 2008; Ochsner et al., 2009). Finally, neither of the sets of studies above addressed the question as to whether affective distracters have a greater potency for cross-task interference than non-affective distracters. By contrast, the present study manipulated the affective nature and task-relevance of target and distracter features independently, and demonstrated the asymmetry of these interference effects. Affective stimuli appear to disrupt ongoing processing regardless of the task-set, while non-affective stimuli seem to impair performance only in the presence of relevant task-sets and response contingencies.
The capacity of affectively salient stimuli to override top-down biasing may be adaptive under certain circumstances. Affective stimuli may help direct attention to, and thus improve the detection of, stimuli that convey biologically relevant information, such as threat (Ohman and Mineka, 2001; Vuilleumier and Huang, 2009; Notebaert et al., 2011) or reward (Maunsell, 2004; Field et al., 2009; Krajbich et al., 2010). The prioritized processing of affective stimuli may also improve the speed and sensitivity of processing of related information (Anderson and Phelps, 2001; Vuilleumier et al., 2001; Phelps et al., 2006; Pourtois et al., 2006; Lim et al., 2009), therein enabling rapid, appropriate responding to salient information. Moreover, affectively salient stimuli can help mitigate the influence of factors that might otherwise impair executive function, such as by eliminating Stroop dilution effects (Chajut et al., 2010) and overcoming the attentional blink (Anderson and Phelps, 2001; Lim et al., 2009). Thus, the ability of affective stimuli to alter top-down processing can help promote adaptive responding.
The interaction between affective processing and executive function may, however, be disrupted in certain clinical disorders (Bishop, 2007; Banich et al., 2009). Heightened levels of anxiety, for instance, may result in hyper-vigilance for threat, and afflicted individuals may have difficulty disengaging from affective stimuli which disrupt ongoing top-down processing (Fox et al., 2001; Koster et al., 2004; Salemink et al., 2007), diminishing their ability to respond adaptively to subsequent stimuli and function normally. In the present study, all participants were screened for psychological and neurological disorders and subclinical variability in anxiety and depression were not assessed; however, the present findings anticipate that high-anxious individuals are likely to display greater cross-task interference from affective distracters than low-anxious individuals. Future studies should investigate the interactions between affective processing and executive function in relation to individual differences and clinical populations, ideally in conjunction with neuroimaging techniques, to elucidate the mechanisms underlying clinical disorders.
The present study documented the capacity of affectively salient distracters to interfere with ongoing processing despite irrelevance to the current task-set. This interference could be produced by rapid attentional orienting to the affective distracters, difficulty disengaging attention from the affective distracters, or some combination of both orienting and disengagement processes. Clarifying the differential roles of orienting and disengagement processes in selective attention has proved critical to elucidating the processes underlying selective attention to threat, particularly amongst anxious individuals (Fox et al., 2001; Koster et al., 2004; Salemink et al., 2007). However, in the present paradigm, the influences of these two processes cannot be distinguished. Future research should endeavor to characterize the unique contributions of orienting and disengagement to the type of affective interference documented in the current experiment. The present findings also indicate that the capacity of distracters to transcend task boundaries in generating interference is restricted to affectively salient distracters and not shared by non-affective distracters. Note though that non-affective distracters may be capable of crossing task-set boundaries in generating interference when the relevant task is non-affective in nature. The affective nature of a task-set may help buffer it from task-irrelevant interference generally, producing the lack of observed interference by non-affective task-irrelevant distracters. Either increased cross-task interference produced by affective distracters or enhanced task-set shielding for affective task-sets would support the asymmetrical effects of interference reported here.
In conclusion, through the adaptation of a face–word Stroop paradigm, the present study tested two key assumptions of theories of prioritized processing of affective stimuli, namely, that such stimuli can transcend task-set boundaries and that this capacity is not shared by non-affective stimuli. The current findings support both of these assumptions, demonstrating that affective stimuli disrupt top-down processing regardless of task-set, but that non-affective stimuli only produce interference if relevant for the task-set at hand. This asymmetric modulation of executive function confirms the prioritized processing of affective stimuli and highlights the importance of understanding the interactive influence of affect and cognition.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was supported by NIMH R01 MH087610-01 (Tobias Egner) and an NSF Graduate Research Fellowship (Crystal Reeck). The authors thank Sora Ely and Amanda Arulpragasam for assistance with data collection.
Footnote
- ^Development of the MacBrain Face Stimulus Set was overseen by Nim Tottenham and supported by the John D. and Catherine T. MacArthur Foundation Research Network on Early Experience and Brain Development. Please contact Nim Tottenham atdG90dDAwMDZAdGMudW1uLmVkdQ==for more information concerning the stimulus set.
References
Algom, D., Chajut, E., and Lev, S. (2004). A rational look at the emotional Stroop phenomenon: a generic slowdown, not a Stroop effect. J. Exp. Psychol. Gen. 133, 323–338.
Anderson, A. K., and Phelps, E. A. (2001). Lesions of the human amygdala impair enhanced perception of emotionally salient events. Nature 411, 305–309.
Banich, M. T., Mackiewicz, K. L., Depue, B. E., Whitmer, A. J., Miller, G. A., and Heller, W. (2009). Cognitive control mechanisms, emotion and memory: a neural perspective with implications for psychopathology. Neurosci. Biobehav. Rev. 33, 613–630.
Bishop, S. J. (2007). Neurocognitive mechanisms of anxiety: an integrative account. Trends Cogn. Sci. (Regul. Ed.) 11, 307–316.
Chajut, E., Schupak, A., and Algom, D. (2010). Emotional dilution of the Stroop effect: a new tool for assessing attention under emotion. Emotion 10, 944–948.
Compton, R. J., Banich, M. T., Mohanty, A., Milham, M. P., Herrington, J., Miller, G. A., Scalf, P. E., Webb, A., and Heller, W. (2003). Paying attention to emotion: an fMRI investigation of cognitive and emotional Stroop tasks. Cogn. Affect. Behav. Neurosci. 3, 81–96.
De Houwer, J. (2003). On the role of stimulus-response and stimulus-stimulus compatibility in the Stroop effect. Mem. Cognit. 31, 353–359.
Desimone, R., and Duncan, J. (1995). Neural mechanisms of selective visual attention. Annu. Rev. Neurosci. 18, 193–222.
Egner, T., Etkin, A., Gale, S., and Hirsch, J. (2008). Dissociable neural systems resolve conflict from emotional versus nonemotional distracters. Cereb. Cortex 18, 1475–1484.
Etkin, A., Egner, T., Peraza, D. M., Kandel, E. R., and Hirsch, J. (2006). Resolving emotional conflict: a role for the rostral anterior cingulate cortex in modulating activity in the amygdala. Neuron 51, 871.
Field, M., Munafo, M. R., and Franken, I. H. A. (2009). A meta-analytic investigation of the relationship between attentional bias and subjective craving in substance abuse. Psychol. Bull. 135, 589–607.
Fox, E., Russo, R., Bowles, R., and Dutton, K. (2001). Do threatening stimuli draw or hold visual attention in subclinical anxiety? J. Exp. Psychol. Gen. 130, 681–700.
Haas, B. W., Omura, K., Constable, R. T., and Canli, T. (2006). Intederence produced by emotional conflict associated with anterior cingulate activation. Cogn. Affect. Behav. Neurosci. 6, 152–156.
Hommel, B. (1998). Event files: evidence for automatic integration of stimulus-response episodes. Vis. cogn. 5, 183–216.
Isenberg, N., Silbersweig, D., Engelien, A., Emmerich, S., Malavade, K., Beattie, B., Leon, A. C., and Stern, E. (1999). Linguistic threat activates the human amygdala. Proc. Natl. Acad. Sci. U.S.A. 96, 10456–10459.
Kornblum, S., Hasbroucq, T., and Osman, A. (1990). Dimensional overlap – cognitive basis for stimulus-response compatibility – a model and taxonomy. Psychol. Rev. 97, 253–270.
Koster, E. H. W., Crombez, G., Verschuere, B., and De Houwer, J. (2004). Selective attention to threat in the dot probe paradigm: differentiating vigilance and difficulty to disengage. Behav. Res. Ther. 42, 1183–1192.
Krajbich, I., Armel, C., and Rangel, A. (2010). Visual fixations and the computation and comparison of value in simple choice. Nat. Neurosci. 13, 1292–1298.
Lim, S.-L., Padmala, S., and Pessoa, L. (2009). Segregating the significant from the mundane on a moment-to-moment basis via direct and indirect amygdala contributions. Proc. Natl. Acad. Sci. U.S.A. 106, 16841–16846.
Macleod, C. M. (1991). Half a century of research on the Stroop effect – an integrative review. Psychol. Bull. 109, 163–203.
Mathews, A., and Macleod, C. (1985). Selective processing of threat cues in anxiety-states. Behav. Res. Ther. 23, 563–569.
Maunsell, J. H. R. (2004). Neuronal representations of cognitive state: reward or attention? Trends Cogn. Sci. 8, 261–265.
Mayr, U., Awh, E., and Laurey, P. (2003). Conflict adaptation effects in the absence of executive control. Nat. Neurosci. 6, 450–452.
McKenna, F. P. (1986). Effects of unattended emotional stimuli on color-naming performance. Curr. Psychol. Res. Rev. 5, 3–9.
Milham, M. P., Banich, M. T., Webb, A., Barad, V., Cohen, N. J., Wszalek, T., and Kramer, A. F. (2001). The relative involvement of anterior cingulate and prefrontal cortex in attentional control depends on nature of conflict. Cogn. Brain Res. 12, 467–473.
Miller, E. K., and Cohen, J. D. (2001). An integrative theory of prefrontal cortex function. Annu. Rev. Neurosci. 24, 167–202.
Notebaert, L., Crombez, G., Van Damme, S., De Houwer, J., and Theeuwes, J. (2011). Signals of threat do not capture, but prioritize, attention: a conditioning approach. Emotion 11, 81–89.
Ochsner, K. N., Hughes, B., Robertson, E. R., Cooper, J. C., and Gabrieli, J. D. E. (2009). Neural systems supporting the control of affective and cognitive conflicts. J. Cogn. Neurosci. 21, 1841–1854.
Ohman, A., and Mineka, S. (2001). Fears, phobias, and preparedness: toward an evolved module of fear and fear learning. Psychol. Rev. 108, 483–522.
Pelli, D. G. (1997). The VideoToolbox software for visual psychophysics: transforming numbers into movies. Spat. Vis. 10, 437–442.
Phelps, E. A., Ling, S., and Carrasco, M. (2006). Emotion facilitates perception and potentiates the perceptual benefits of attention. Psychol. Sci. 17, 292–299.
Pourtois, G., Schwartz, S., Seghier, M. L., Lazeyras, F., and Vuilleumier, P. (2006). Neural systems for orienting attention to the location of threat signals: an event-related fMRI study. Neuroimage 31, 920–933.
Salemink, E., Van Den Hout, M. A., and Kindt, M. (2007). Selective attention and threat: quick orienting versus slow disengagement and two versions of the dot probe task. Behav. Res. Ther. 45, 607–615.
Stroop, J. R. (1935). Studies of interference in serial verbal reactions. J. Exp. Psychol. 18, 643–662.
Tottenham, N., Tanaka, J. W., Leon, A. C., Mccarry, T., Nurse, M., Hare, T. A., Marcus, D. J., Westerlund, A., Casey, B. J., and Nelson, C. (2009). The NimStim set of facial expressions: judgments from untrained research participants. Psychiatry Res. 168, 242–249.
Vuilleumier, P., Armony, J. L., Driver, J., and Dolan, R. J. (2001). Effects of attention and emotion on face processing in the human brain: an event-related fMRI study. Neuron 30, 829–841.
Vuilleumier, P., and Huang, Y. M. (2009). Emotional attention: uncovering the mechanisms of affective biases in perception. Curr. Dir. Psychol. Sci. 18, 148–152.
Whalen, P. J., Bush, G., Mcnally, R. J., Wilhelm, S., Mcinerney, S. C., Jenike, M. A., and Rauch, S. L. (1998). The emotional counting Stroop paradigm: a functional magnetic resonance imaging probe of the anterior cingulate affective division. Biol. Psychiatry 44, 1219–1228.
Williams, J. M. G., Mathews, A., and Macleod, C. (1996). The emotional Stroop task and psychopathology. Psychol. Bull. 120, 3–24.
Keywords: attention, affect, interference resolution, emotional conflict, executive function, conflict, cognitive control, Stroop
Citation: Reeck C and Egner T (2011) Affective privilege: asymmetric interference by emotional distracters. Front. Psychology 2:232. doi: 10.3389/fpsyg.2011.00232
Received: 16 May 2011;
Accepted: 29 August 2011;
Published online: 16 September 2011.
Edited by:
Wim Notebaert, Ghent University, BelgiumReviewed by:
Maurits W. Van Der Molen, University of Amsterdam, NetherlandsAnnekathrin Schacht, University of Goettingen, Germany
Copyright: © 2011 Reeck and Egner. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Tobias Egner, Center for Cognitive Neuroscience, Department of Psychology and Neuroscience, Duke University, LSRC Building, Research Drive, Box 90999, Durham, NC 27708, USA. e-mail:dG9iaWFzLmVnbmVyQGR1a2UuZWR1