


- 1 Konrad Lorenz Forschungsstelle, University of Vienna, Vienna, Austria
- 2 Department of Cognitive Biology, University of Vienna, Vienna, Austria
- 3 Institut Pluridisciplinaire Hubert CURIEN Strasbourg, Université de Strasbourg, Strasbourg, France
- 4 Centre National de la Recherche Scientifique, UMR7178, Strasbourg, France
The ability to control an immediate impulse in return for a more desirable – though delayed – outcome has long been thought to be a uniquely human feature. However, studies on non-human primates revealed that some species are capable of enduring delays in order to get food of higher quality or quantity. Recently two corvid species, common raven (Corvus corax) and carrion crow (Corvus corone corone), exchanged food for a higher quality reward though seemed less capable of enduring delays when exchanging for the same food type in a higher quantity. In the present study, we specifically investigated the ability of carrion crows to overcome an impulsive choice in a quantitative exchange task. After a short delay, individuals were asked to give back an initial reward (cheese) to the human experimenter in order to receive a higher amount of the same reward (two, four, or eight pieces). We tested six captive crows – three individuals never exchanged the initial reward for a higher quantity; the other three birds did exchange though at very low rates. We performed a preference test between one or more pieces of cheese in order to address whether crow poor performance could be due to an inability to discriminate between different quantities or not attributing a higher value to the higher quantities. All birds chose the higher quantities significantly more often, indicating that they can discriminate between quantities and that higher quantities are more desirable. Taken together, these results suggest that, although crows may possess the cognitive abilities to judge quantities and to overcome an impulsive choice, they do so only in order to optimize the qualitative but not quantitative output in the exchange paradigm.
Introduction
The ability to overcome an impulsive choice-defined as the choice of the less desirable, immediate option, over a more desirable though delayed one (Rachlin, 1974), has long been thought to be a uniquely human feature (Burns and Powers, 1975; Logue, 1988; Rachlin, 2000). Impulse control develops slowly in children (Mischel et al., 1989) and often represents serious difficulties to non-human animals (Stephens and Anderson, 2001; Stevens et al., 2005; Evans and Beran, 2007a; Anderson et al., 2010). This is particularly true for several bird species, which hardly control an impulsive choice for longer than a few seconds in experimental situations (pigeons, Columba livia: Ainslie, 1974; Mazur, 1987; Logue et al., 1988; Green et al., 2004; African gray parrots, Psittacus erithacus: Vick et al., 2010; domestic fowl, Gallus gallus: Abeyesinghe et al., 2005).
The difficulties of non-human animals in controlling impulses seem to reflect primarily a cognitive problem. From an evolutionary perspective, choices optimizing long-term gain over immediate benefits in foraging (Stephens and Krebs, 1986; Kacelnik, 2003) or mating situations (Stephens and Krebs, 1986; Stevens, 2010) should be selected for and, thus, found in a variety of species. However, several authors suggest that in some contexts, delaying food intakes might add additional risk to survival. In those cases, a fast discounting of food value over time, i.e., a preference for immediacy, may be the most adaptive strategy (Kagel et al., 1986; Stephens, 2008) and choices in several species have been described not to be consistent with rate maximization (Stevens and Mühlhoff, 2012). In term of the mechanisms involved, the capacity of an individual to overcome an impulsive choice may depend on the subjective value of the expected reward (Mitchell and Wilson, 2010) relative to the amount of the reward (Kacelnik and Bateson, 1996; Green et al., 1997; Bateson, 2002a), its quality (Bruce et al., 2010), and in relation to the delay until the reward is received (Green et al., 1999, 2004).
The effects of delay in gratification on self-control are fairly well investigated in primates. In classical delay of gratification tasks, subjects are given a choice between an immediate reward of low value and a delayed option of higher value. Once the choice is made they cannot revert their decision and have to wait for the whole duration of the delay before obtaining the larger reward. In these circumstances, only great apes accept delays over a minute (great apes: Rosati et al., 2007; long-tailed macaque: Tobin et al., 1996; rhesus macaques, Macaca mulatta: Szalda-Petree et al., 2004). By contrast, other techniques allow subjects to reevaluate their choice during the delay. This is the case in delay maintenance tasks such as accumulation tasks where subject can stop an accumulation of food rewards by reaching for them, and in delayed exchange tasks where individuals endowed with an initial low value item can give up (consume the item) at any time during the delay prior to being asked to exchange it for a higher value food. In both the delayed exchange task (chimpanzees, Pan troglodytes: Dufour et al., 2007; brown capuchin monkeys, Cebus apella: Drapier et al., 2005; long-tailed macaques, Macaca fascicularis: Pelé et al., 2010) and in the accumulation task (chimpanzees: Beran and Evans, 2009; orangutan, Pongo pygmaeus: Beran, 2002) individuals’ ability to overcome the impulsive choice decreases with increasing delay time. In those studies, the type and amount of the food reward affects the subjects’ responses. Generally, they endure longer delays with high qualities and high quantities. Great apes and children have even been described to engage in self-distractive behaviors, probably in an attempt to better cope with long delays (Evans and Beran, 2007b; Steelandt et al., 2012).
The low performance of birds compared to primates in self-control tasks questions whether birds evaluate the value of a reward differently or if they are generally more impulsive. In the present study, we investigated whether carrion crows (Corvus corone corone) could overcome an impulsive choice in order to maximize the quantity of their reward. Corvids are expected to be better at impulse control than most bird species tested so far, as they are renowned for their cognitive skills during competition for hidden food (Emery and Clayton, 2004; Bugnyar and Heinrich, 2005; Clayton and Emery, 2005). Like other corvids, crows engage in scatter-hoarding. This means that in everyday life, they routinely delay immediate consumption and store food for future use (Glutz von Blotzheim, 1985; Heinrich and Pepper, 1998). In contrast to long-term cachers, crows frequently retrieve their caches fairly quickly (minutes to hours; personal observation) or up to a couple of days (Waite, 1985; Fjeld and Sonerud, 1988) after the caches have been made. In a previous study, we tested the willingness of crows and ravens to exchange a non-preferred food (bread) against a more desirable reward (grape, cheese, and sausage). In this task, birds could overcome their impulsive response to eat the immediately available food for more than just a few seconds and in some trials two birds even waited up to a couple of minutes (Dufour et al., 2012). However, along with the qualitative exchange task, we ran quantitative exchange training and in the latter found the birds failed to exchange one piece of cheese for a higher quantity (Wascher, C. A. F., and Braun, A., unpublished data). Here, we followed up on the birds apparent problem at maximizing a quantitative income. We tested if carrion crows can control their impulsivity in an exchange task in order to maximize the quantity of their reward. To control for preferences based on quantitative assessment, we also ran a binary food choice task, where individuals were asked to choose between the options one vs. two, one vs. four, and one vs. eight pieces of cheese. By doing so, we aimed at determining whether any failure to respond in the context of quantitative maximization would rely on a lack of impulse control or on a failure to discriminate quantities.
Materials and Methods
Study Subjects and Keeping
Seven captive crows (three females, four males) were involved in this study. Although they differed in origin, age, and experience with humans (Table 1), they were all tame and already used to experimental testing in physical and visual isolation (see below). Crows were held in large outdoor aviaries (15–45 m2) at the Konrad Lorenz Forschungsstelle (KLF) in Austria. At the time of the study, they were kept in three adjacent subgroups, consisting of two male–female pairs and a trio of two males and one female. One male (“Hugo”) did not learn to exchange a token with a human experimenter; therefore only six individuals participated in the exchange experiment (Task A), whereas all seven birds participated in the quantity discrimination experiment (Task B).

Table 1. List of crows participating in the present study.
Experiments
Experiments were conducted from February until April 2010. We performed two daily sessions – one of task (A) and one of task (B) – in counterbalanced order between 0800–1000 and 1600–1800. All birds voluntarily participated in the experiments, entering the test compartment after being called by name. They were fed twice a day (morning and late afternoon, after the experiments) with a mixture of meat, milk products, vegetables, and fruits and had ad libitum access to water. All procedures were in accordance with the laws of Austria and the federal State of Upper Austria.
Task (A): Exchange task
From spring 2007 to fall 2009, we successfully trained six carrion crows to exchange a token (stone or piece of wood) for food with a human experimenter via progressive behavioral shaping. It took the birds approximately 25–30 sessions of eight trials each to learn how to exchange. Once a bird exchanged in 80% of the trials over two consecutive sessions, the training procedure was considered to be complete.
Crows were tested individually, separated from the experimenter (Claudia A. F. Wascher) by a wire mesh. Individuals were presented the initial item (one piece of cheese; 5 mm × 3 mm × 1 mm) in one hand (left and right in counterbalanced order) and a higher number of cheese pieces (two, four, or eight) in the other hand. The initial item was given to the subject and after a short delay (approximately 2 s) individuals were given the chance to exchange the initial item in order to receive the larger amount of cheese. In order to succeed, birds had to put the initial item into the open palm of the experimenter. During this sequence the reward was visible for the crow. We conducted 12 sessions, consisting of nine trials each, offering each quantity (two, four, and eight) three times per session in randomized order.
Task (B): Preference task
Individuals were familiar with different forms of two-choice studies (Mikolasch et al., 2012; Braun, A., and Bugnyar, T., unpublished data) and had already been involved in food preference tasks where they had to choose between two food items differing in quality by pointing with their beak. For the current study, they were given the opportunity to choose between two different amounts of the same highly desirable food reward. Each amount was presented in the hands of the human experimenter (Claudia A. F. Wascher) until birds indicated their chosen option. During each trial, the experimenter gazed at the ground in order to avoid giving cues to the focal individual. Note that we explicitly tested the same crows for using human-given cues in a choice experiment, which they failed to do (Cibulski, L., unpublished data). The different amounts were presented as followed: one vs. two, one vs. four, one vs. eight pieces of cheese reward. We ran 12 sessions consisting of nine trials, with each quantity (two, four, and eight pieces) being offered three times in randomized order. The hand (left/right), which presented the higher quantity, was counterbalanced across session.
Statistical Analysis
To test whether the birds’ choice for the higher quantity deviated from chance level in task (B) we applied binomial tests. A Friedman test and post hoc Wilcoxon signed rank tests were conducted to investigate differences in performance between different conditions (one vs. two, one vs. four, one vs. eight). Tests were conducted two-tailed and considered significant when p < 0.05. Statistical analysis was conducted using SPSS 19.0 statistical package.
Results
Experiment (A): Exchange Task
Crows performed at very low levels in all three conditions of this experiment (Figure 1A). From six birds tested, three individuals never successfully exchanged one piece of cheese for a larger amount of cheese. The remaining three individuals exchanged at low frequencies (Table 2). Out of a total of 108 trials, individual 1 (“Gabi”) had one successful trial in the condition ×8; individual 2 (“Toeffel”) had five successful trials in the conditions ×4 and ×8; individual three (“Franz”) had 13 successful trials spread between all three conditions (Figure 1A). Table 2 also lists the performance of the crows in the qualitative exchange task of Dufour et al. (2012). It is notable that the two individuals that exchanged more than once in the current task were not among the best in the qualitative exchange task.
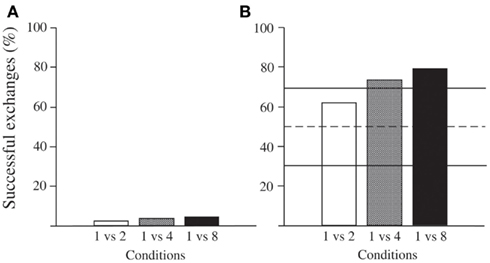
Figure 1. Performance of carrions crows in the quantitative exchange task (A) and in the preference test (B). In task (B), individuals chose higher quantities significantly more often than expected by chance in condition one vs. four and one vs. eight. Dashed line represents 50% chance; full lines represent significance levels (p < 0.05).

Table 2. Exchange performance (percentage of successful exchanges at 2 s delay) of crows tested in the quantitative exchange Task (A) of the current study and the qualitative exchange task of Dufour et al. (2012).
Experiment (B): Preference Task
In contrast to experiment (A), individuals behaved consistently in task (B). Crows chose the higher quantities significantly above chance in the conditions one vs. four (Binomial test: p = 0.01) and one vs. eight (Binomial test: p = 0.001). In condition one vs. two individuals chose the higher quantity more than 50% of the time though not significantly different from chance (mean: 22.4 correct choices out of 36 trials; Binomial test: p = 0.243, Figure 1B).
The individuals choice of bigger quantities differed significantly across conditions (Friedman: N = 7, χ2 = 8.857, df = 2, p = 0.008). Specifically, crows chose the higher quantities more often in the one vs. eight and one vs. four compared to the one vs. two condition (Wilcoxon signed rank test: N = 7, one vs. two – one vs. four: T+ = 27, p = 0.028; one vs. two – one vs. eight: T+ = 28, p = 0.018; only the latter remains significant after Bonferroni correction). Performance did not differ between one vs. four and one vs. eight condition (Wilcoxon signed rank test: N = 7, T+ = 20, p = 0.307).
Discussion
The results of the present study show that carrion crows are able to discriminate quantities in a two-choice preference task and, as expected, prefer larger quantities. Nevertheless, they do not perform above chance when asked to exchange one piece of food for a higher quantity of the same reward. These findings indicate that crows have problems in overcoming impulsive choice in the context of quantity to maximize the number of rewards gained. Surprisingly, the same birds performed well in a qualitative exchange task, not only overcoming an initial impulsiveness in order to exchange, but also coping with extended delays in order to optimize the quality of the gained food reward (Dufour et al., 2012). This suggests that, although carrion crows generally have the cognitive capacities to overcome an impulsive choice, their motivation to do so is strongly dependent on context. In the current task, they seemingly do not attribute quantity a high enough value to be willing to exchange for a higher number of rewards.
The question arises if the two chosen tasks of the present study deliver reliable results. The findings of our two-choice task are in accordance with other studies on quantity judgment and number competence in corvids (Koehler, 1941; Ujfalussy, D. J., unpublished data). Moreover, the very same crows performed poorly in a parallel experiment testing their use of human-given cues (Cibulski, L., unpublished data). Together, this suggests that crows’ consistent choice of higher quantities in the current experiment is due to their discrimination ability of quantities rather than any form of cueing. Focusing on our main result, the failure of the crows in exchanging food for higher quantities, we are confident that this cannot be attributed to problems with the exchange paradigm per se. Birds had been trained to exchange 2 years prior the present study and successfully participated in a number of studies applying this specific paradigm (Dufour et al., 2012; Wascher, C. A. F., and Bugnyar, T., unpublished data). The food exchange paradigm was therefore not new to them, and similarly lack of experience with this task or with the use of food rather than tokens cannot explain their performances. It is particularly interesting that individual performance in the qualitative exchange experiment of Dufour et al. (2012) did not correlate with the birds’ performance in the quantitative exchange. Of the two crows performing best in the qualitative exchange, one never exchanged in the quantitative exchange and one individual exchanged in a single out of 108 trials. The most successful individual in the quantitative exchange (13 successful out of 108 trials) was among the worst performing individuals in the qualitative exchange (Table 2).
In studies investigating quantity discrimination in non-human animals, it has been suggested that the preference for higher quantities is caused by the need for optimization of the outcome (Boysen and Berntson, 1989; Boysen, 1997; Agrillo et al., 2007). Although, when it comes to foraging decisions, the choice made by individuals has been shown to be more related to context rather than absolute regarding of the reward’s value (Bateson, 2002b; Bateson et al., 2002, 2003; Shafir et al., 2002). Our results support these findings in so far as crows do not generally overcome the impulsive choice in order to optimize the size of the reward though do so in the context of quality.
It is possible that our feeding regime has removed the birds from experiencing hunger so they could afford to become more selective in selection of food types. Still, it is surprising that crows, as a food-caching species, appear to be insensitive to maximizing quantity of highly preferred food. In the present study, individuals did engage in food-caching approximately as frequently as in other experiments using the exchange paradigm (Wascher, C. A. F., unpublished data) with the consequence that they were usually caching one rather than many pieces per event. Possibly, as short-term cachers, crows are tuned to pay attention to food of different qualities (Raby et al., 2007) rather than quantities. For carrions crows and maybe birds in general it might be critical for survival to be able to fly away quickly without being handicapped by large loads. Therefore individuals attending less to quantity than to quality of the prospected income might have been favored by natural selection. If this is true, similar cognitive constraints could be expected in other food-caching species.
It is known that the incentive to cache is affected by the quality of food (Clayton and Dickinson, 1999). This could affect the results of our study in two ways: the food we used (cheese) was not preferred enough by the crows to optimize quantity. However, we consider this option as highly unlikely because cheese turned out to be one of the most preferred food items in several studies (Dufour et al., 2012) and in order to maintain its high value, it is given to the birds only during experiments. Alternatively, cheese was too preferred and crows could not inhibit the impulse to eat a single piece of it, once having it already in their beak. This could certainly be the case. However, we chose highly preferred food to ensure a motivation to participate. Future studies are needed to disentangle effects of food quality.
The endowment effect, in which individuals favor an item more when they possess it, could offer a potential explanation for the behavior of the crows in the present study. This effect has previously been described in humans (Kahneman et al., 1991), chimpanzees (Brosnan et al., 2007), and capuchin monkeys (Lakshminarayanan et al., 2008). In the present study this would mean that the birds did not exchange one piece of cheese for higher quantities because they may have valued the one item they already possessed more than the expected reward. Further studies are necessary to test this assumption, for example by applying another experimental paradigm like the accumulation task, which has been used in various primate species (Beran, 2002; Beran and Evans, 2006, 2009; Evans and Beran, 2007a; Pelé et al., 2011) and recently also in parrots (Vick et al., 2010). Similar to the results of our study, parrots performed badly in overcoming an impulsive choice in order to optimize the quantitative output. This hints toward similarities between parrots and crows regarding the evaluation of food, anyhow parrots ability to delay gratification in the context of quality has yet to be tested.
In conclusion, carrion crows are able to discriminate quantities (experiment B) and they are able to control their immediate impulses when having the chance to exchange food for better food (Dufour et al., 2012). However, crows rarely exchange food for a higher amount of the same food type (experiment A). This context dependence in exchange performance stands in contrast to recent findings in primates (Ramseyer et al., 2006; Dufour et al., 2007; Pelé et al., 2010) and leaves us with two mutually not exclusive interpretations: (i) motivational differences due to daily feeding and/or this particular testing regime and (ii) species differences regarding evaluation of food.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Kurt Kotrschal and Georgine Szipl for support and general discussion. We like to thank Rachael Miller, Michael Beran, and three anonymous referees for valuable comments on the manuscript. The project was funded by a L’Oreal Women in Science scholarship to Claudia A. F. Wascher, the ESF EUROCORES program TECT: COCOR (I105-G11), and the START program (Y366-B17) of the Austrian Science Fund (FWF) to Thomas Bugnyar. Permanent support was provided by the “Verein der Förderer” and the Herzog von Cumberland Stiftung.
References
Abeyesinghe, S. M., Nicol, C. J., Hartnell, S. J., and Wathes, C. M. (2005). Can domestic fowl, Gallus gallus domesticus, show self-control. Anim. Behav. 70, 1–11.
Agrillo, C., Dadda, M., and Bisazza, A. (2007). Quantity discrimination in female mosquitofish. Anim. Cogn. 10, 63–70.
Anderson, J. R., Kuroshima, H., and Fujita, K. (2010). Delay of gratification in capuchin monkeys (Cebus apella) and squirrel monkeys (Saimiri sciureus). J. Comp. Psychol. 124, 205–210.
Bateson, M. (2002a). Recent advances in our understanding of risk-sensitive foraging preferences. Proc. Nutr. Soc. 61, 1–8.
Bateson, M. (2002b). Context-dependent foraging choices in risk-sensitive starlings. Anim. Behav. 64, 251–260.
Bateson, M., Healy, S. D., and Hurly, T. A. (2002). Irrational choices in hummingbird foraging behaviour. Anim. Behav. 63, 587–596.
Bateson, M., Healy, S. D., and Hurly, T. A. (2003). Context-dependent foraging decisions in rufous hummingbirds. Proc. Biol. Sci. 270, 1271–1276.
Beran, M., and Evans, T. (2009). Delay of gratification by chimpanzees (Pan troglodytes) in working and waiting situations. Behav. Processes 80, 177–181.
Beran, M. J. (2002). Maintenance of self-imposed delay of gratification by four chimpanzees (Pan troglodytes) and an orangutan (Pongo pygmaeus). J. Gen. Psychol. 129, 49–66.
Beran, M. J., and Evans, T. A. (2006). Maintenance of delay of gratification by four chimpanzees (Pan troglodytes): the effects of delayed reward visibility, experimenter presence, and extended delay intervals. Behav. Processes 73, 315–324.
Boysen, S. T., and Berntson, G. G. (1989). Numerical competence in a chimpanzee. J. Comp. Psychol. 103, 23–31.
Brosnan, S. F., Jones, O. D., Lambeth, S. P., Mareno, M. C., Richardson, A. S., and Schapiro, S. J. (2007). Endowment effects in chimpanzees. Curr. Biol. 17, 1704–1707.
Bruce, T. J. A., Midega, C. A. O., Birkett, M. A., Pickkett, J. A., and Zeyaur, K. (2010). Is quality more important than quantity? Insect behavioural responses to changes in a volatile blend after stemborer oviposition on an African grass. Biol. Lett. 6, 314–317.
Bugnyar, T., and Heinrich, B. (2005). Food-storing ravens differentiate between knowledgeable and ignorant competitors. Proc. Biol. Sci. 272, 1641–1646.
Burns, D. J., and Powers, R. B. (1975). Choice and self-control in children: a test of Rachlin’s model. Bull. Psychon. Soc. 5, 156–158.
Clayton, N. S., and Dickinson, A. (1999). Motivational control of caching behaviour in the scrub jay, Aphelocoma coerulescens. Anim. Behav. 57, 435–444.
Drapier, M., Chauvin, C., Dufour, V., Uhlrich, P., and Thierry, B. (2005). Food-exchange with humans in brown capuchin monkeys. Primates 46, 241–248.
Dufour, V., Pelé, M., Sterck, E. H. M., and Thierry, B. (2007). Chimpanzee (Pan troglodytes) anticipation of food return: coping with waiting time in an exchange task. J. Comp. Psychol. 121, 145–155.
Dufour, V., Wascher, C. A. F., Braun, A., Miller, R., and Bugnyar, T. (2012). Corvids can decide if a future exchange is worth waiting for. Biol. Lett. 8, 201–204.
Emery, N. J., and Clayton, N. (2004). The mentality of crows: convergent evolution of intelligence in corvids and apes. Science 306, 1903–1907.
Evans, T. A., and Beran, M. J. (2007a). Delay of gratification and delay maintenance by rhesus macaques (Macaca mulatta). J. Gen. Psychol. 134, 199–216.
Evans, T. A., and Beran, M. J. (2007b). Chimpanzees use self-distraction to cope with impulsivity. Biol. Lett. 3, 599–602.
Fjeld, P. E., and Sonerud, G. A. (1988). Food caching, cache recovery, and the use of an egg shell dump in hooded crows (Corvus corone cornix). J. Avian Biol. 19, 268–274.
Green, L., Myerson, J., Holt, D. D., Slevin, J. R., and Estle, S. J. (2004). Discounting of delayed food rewards in pigeons and rats: is there a magnitude effect? J. Exp. Anal. Behav. 81, 39–50.
Green, L., Myerson, J., and Mcfadden, E. (1997). Rate of temporal discounting decreases with amount of reward. Mem. Cognit. 25, 715–723.
Green, L., Myerson, J., and Ostaszewski, P. (1999). Discounting of delayed rewards across the life span: age differences in individual discounting functions. Behav. Processes 1, 89–96.
Heinrich, B., and Pepper, J. (1998). Influences of competitors on caching behaviour in common ravens, Corvus corax. Anim. Behav. 56, 1083–1090.
Kacelnik, A. (2003). “The evolution of patience,” in Time and decision. Economic and psychological perspectives on intertemporal choice, eds. G. Loewenstein, D. Read, and R. F. Baumeister (New York: Russell Sage Foundation), 115–138.
Kacelnik, A., and Bateson, M. (1996). Risky theories- the effects of variance on foraging decisions. Am. Zool. 36, 402–434.
Kagel, J. H., Green, L., and Caraco, T. (1986). When foragers discount the future: constraint or adaptation? Anim. Behav. 34, 271–283.
Kahneman, D., Knetsch, J. L., and Thaler, R. (1991). The endownment effect, loss aversion, and the status quo bias. J. Econ. Perspect. 5, 193–206.
Koehler, O. (1941). Vom Erlernen unbenannter Anzahlen bei Vögeln. Naturwissenschaften 14/15, 201–218.
Lakshminarayanan, V., Chen, K. M., and Santos, L. R. (2008). Endowment effect in capuchin monkeys. Philos. Trans. R. Soc. Lond. B Biol. Sci. 363, 3837–3844.
Logue, A. W. (1988). Research on self-control: an integrating framework. Behav. Brain Sci. 11, 665–679.
Logue, A. W., Chavarro, A., Rachlin, H., and Reeder, R. W. (1988). Impulsiveness in pigeons living in the experimental chamber. Learn. Behav. 16, 31–39.
Mazur, J. E. (1987). “An adjusting procedure for studying delayed rein-forcement: the effects of delay and intervening events on reinforcement value,” in Quantitative Analyses of Behavior, eds. M. L. Commons, J. E. Mazur, J. A. Nevin, and H. Rachlin (Hillsdale: Erlbaum), 55–73.
Mikolasch, S., Kotrschal, K., and Schloegl, C. (2012). Is caching the key to exclusion in corvids? The case of carrion crows (Corvus corone corone). Anim. Cogn. 15, 73–82.
Mischel, W., Shoda, Y., and Rodriguez, M. L. (1989). Delay of gratification in children. Science 244, 933–938.
Mitchell, S. H., and Wilson, V. B. (2010). The subjective value of delayed and probabilistic outcomes: outcome size matters for gains but not for losses. Behav. Processes 83, 36–40.
Pelé, M., Dufour, V., Micheletta, J., and Thierry, B. (2010). Long-tailed macaques display unexpected waiting abilities in exchange tasks. Anim. Cogn. 13, 263–271.
Pelé, M., Micheletta, J., Uhlrich, P., Thierry, B., and Dufour, V. (2011). Delay maintenance in tonkean macaques (Macaca tonkeana) and brown capuchin monkeys (Cebus apella). Int. J. Primatol. 32, 149–166.
Raby, C. R., Alexis, D. M., Dickinson, A., and Clayton, N. (2007). Planning for the future by western scrub-jays. Nature 445, 919–921.
Ramseyer, A., Pelé, M., Dufour, V., Chauvin, C., and Thierry, B. (2006). Accepting loss: the temporal limits of reciprocity in brown capuchin monkeys. Proc. Biol. Sci. 273, 179–184.
Rosati, A. G., Stevens, J. R., Hare, B., and Hauser, M. D. (2007). The evolutionary origins of human patience: temporal preferences in chimpanzees, bonobos, and human adults. Curr. Biol. 17, 1663–1668.
Shafir, S., Waite, T. A., and Smith, B. H. (2002). Context dependent violations of rational choice in honeybees (Apis mellifera) and gray jays (Perisoreus canadensis). Behav. Ecol. Sociobiol. (Print) 51, 180–187.
Steelandt, S., Thierry, B., Broihanne, M.-H., and Dufour, V. (2012). The ability of children to delay gratification in an exchange task. Cognition 122, 416–425.
Stephens, D. W. (2008). Decision ecology: foraging and the ecology of animal decision making. Cogn. Affect. Behav. Neurosci. 8, 475–484.
Stephens, D. W., and Anderson, D. (2001). The adaptive value of preference for immediacy: when shortsighted rules have farsighted consequences. Behav. Ecol. 12, 330–339.
Stevens, J. R. (2010). “Intertemporal choice,” in Encyclopedia of Animal Behavior, eds. M. D. Breed and J. Moore (Oxford: Academic Press).
Stevens, J. R., Hallinan, E. V., and Hauser, M. D. (2005). The ecology and evolution of patience in two New World monkeys. Biol. Lett. 1, 223–226.
Stevens, J. R., and Mühlhoff, N. (2012). Intertemporal choice in lemurs. Behav. Processes 89, 121–127.
Szalda-Petree, A. D., Craft, B. B., Martin, L. M., and Deditius-Island, H. K. (2004). Self-control in rhesus macaques (Macaca mulatta): controlling for differential stimulus exposure. Percept. Mot. Skills 98, 141–146.
Tobin, H., Logue, A. W., Chelonis, J. J., Ackerman, K. T., and May, J. G. (1996). Self-control in the monkey Macaca fascicularis. Anim. Learn. Behav. 24, 168–174.
Vick, S.-J., Bovet, D., and Anderson, J. R. (2010). How do African grey parrots (Psittacus erithacus) perform on a delay of gratification task? Anim. Cogn. 13, 351–358.
Keywords: impulse control, quantity, exchange task, carrion crows, Corvus corone corone
Citation: Wascher CAF, Dufour V and Bugnyar T (2012) Carrion crows cannot overcome impulsive choice in a quantitative exchange task. Front. Psychology 3:118. doi: 10.3389/fpsyg.2012.00118
Received: 23 December 2011; Accepted: 31 March 2012;
Published online: 17 April 2012.
Edited by:
Michael Beran, Georgia State University, USAReviewed by:
Jennifer Vonk, Oakland University, USAElsa Addessi, National Research Council of Italy, Italy
Dalila Bovet, Université Paris Ouest Nanterre La Défense, France
Copyright: © 2012 Wascher, Dufour and Bugnyar. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Claudia A. F. Wascher, Konrad Lorenz Forschungsstelle für Ethologie, University of Vienna, Fischerau 11, A-4645 Grünau, Vienna, Austria. e-mail:Y2xhdWRpYS53YXNjaGVyQGtsZi5hYy5hdA==