 Pingping Xu
Pingping Xu Qingqing Qu
Qingqing Qu Wei Shen
Wei Shen Xingshan Li
Xingshan Li- 1Key Laboratory of Behavioral Science, Institute of Psychology, Chinese Academy of Sciences, Beijing, China
- 2Department of Psychology, University of Chinese Academy of Sciences, Beijing, China
- 3Institute of Psychological Sciences, Hangzhou Normal University, Hangzhou, China
- 4Zhejiang Key Laboratory for Research in Assessment of Cognitive Impairments, Hangzhou, China
- 5Centre for Cognition and Brain Disorders, Hangzhou Normal University, Hangzhou, China
Evidence from behavior, computational linguistics, and neuroscience studies supported that semantic knowledge is represented in (at least) two semantic systems (i.e., taxonomic and thematic systems). It remains unclear whether, and to what extent taxonomic and thematic relations are co-activated. The present study investigated the co-activation of the two types of semantic representations when both types of semantic relations are simultaneously presented. In a visual-world task, participants listened to a spoken target word and looked at a visual display consisted of a taxonomic competitor, a thematic competitor and two distractors. Growth curve analyses revealed that both taxonomic and thematic competitors attracted visual attention during the processing of the target word but taxonomic competitor received more looks than thematic competitor. Moreover, although fixations on taxonomic competitor rose faster than thematic competitor, these two types of competitors started to attract more fixations than distractor in a similar time window. These findings indicate that taxonomic and thematic relations are co-activated by the spoken word, the activation of taxonomic relation is stronger and rise faster than thematic relation.
Introduction
The structure and organization of semantic knowledge are critical to nearly all aspects of human cognition such as object recognition, memory and language processing. Evidence from behavior, computational linguistics, and neuroscience studies supported that semantic knowledge is represented in (at least) two semantic systems (see Mirman et al., 2017 for a review): a taxonomic system in which semantic knowledge is organized based on categories that are defined by shared semantic features such as fruit (e.g., Collins and Loftus, 1975), and a thematic system with the organization of semantic information based on events or scenarios such as objects involved in building a house (e.g., Estes et al., 2011). For instance, mouse and dogs are taxonomically related because they share semantic features; mouse and cheese are thematically related because they often co-occur in the same scenario or event, but they do not have common features.
Behavioral studies demonstrating a dissociation between taxonomic and thematic semantics widely used semantic judgement or related semantic tasks. In a “triads” task, participants are asked to choose which of two options is most related to a target. This task has revealed qualitatively different patterns of effects for various concept domain: taxonomic relation tends to be more important for natural objects such as animals, whereas thematic relation is more important for manipulable objects such as tools (Bonthoux and Kalénine, 2007; Kalénine et al., 2009). More evidence came from the domain of spoken word production. In a picture-word interference task in which semantic relations between picture names and distractor words were manipulated, de Zubicaray et al. (2013) demonstrated that taxonomic similarity between picture names and distractor words inhibited picture naming whereas thematic relation facilitated picture naming. These findings indicate that taxonomic and thematic semantic relations are functionally distinct.
A number of studies have demonstrated the neural basis dissociation between taxonomic and thematic relations. In a large-scale study of spoken errors in picture naming produced by adults with aphasia, Schwartz et al. (2011) showed that various forms of brain lesion independently influenced semantic errors: Aphasias with lesions in the left anterior temporal lobe (ATL) appeared to produce a higher proportion of taxonomic errors (“pear” in response to apple), and those with lesions in the left temporoparietal junction (TPJ) caused more thematic errors (“worm” in response to apple). Based on these results, Schwartz et al. argued that taxonomic relation with ATL as the critical hub, and thematic relation with TPJ as the critical hub are complementary semantic systems. Although there is less consensus on the precise neural basis underlying taxonomic and thematic semantics, there is general agreement about the neural dissociation of taxonomic and thematic semantics (see Mirman et al., 2017 for a review).
More recently, eye tracking technique has opened up a new avenue for investigating semantic knowledge. In a visual world paradigm, participants listen to a spoken utterance while viewing a visual display, and their eyes are tracked (Cooper, 1974; Tanenhaus et al., 1995). Adopting this task, Mirman and Graziano (2012) demonstrated that both types of semantic knowledge were activated, with larger effect for taxonomic relation. Interestingly, individuals' relative strength of taxonomic relation vs. thematic relation in eye-tracking measurement could predict individuals' preference on taxonomic relation over thematic relation in triads task, which has been taken to indicate that individuals differ in the relative strengths of both types of semantic knowledge. In sum, taxonomic and thematic relations differentially contribute to semantic systems as demonstrated in studies above.
Moreover, there is evidence that taxonomic and thematic relations differ in the time courses of activation. Time courses of these two semantic effects have been assessed in eye-tracking visual world task which does not involve metalinguistic decision, and thus has strengths than widely used semantic priming or judgement tasks. Kalénine et al. (2012) observed that thematic relation produces earlier and more transient effects than artificial taxonomic relation. A subsequent EEG study (Wamain et al., 2015) combining the semantic priming task demonstrated that early ERP components (N1 and P3) were only sensitive to thematic relation, and late N400 component was modulated by both semantic relations, reflecting earlier activation of thematic representations compared to taxonomic representations. However, results from recent behavioral research have demonstrated that activation of taxonomic and thematic relations may proceed in parallel. For example, Jones and Golonka (2012) compared taxonomic and thematic relations (along with integrative relation, see Estes and Jones, 2009; Jones et al., 2017 for integrative relation) in a lexical decision task across three SOAs (100, 500 or 800 ms), in which participants judged whether the target (following integrative, thematic taxonomic or unrelated prime) was a real word or not. Results showed no difference in strength or time course of priming between thematic and taxonomic relations.
In the visual-world studies reviewed above, the two types of semantic competitors were presented on separate displays, as in the classical version of the visual world paradigm and other relevant priming tasks, which caused an indirect way to assess the strength and timing of these two types of relations. It remains unclear whether and to what extent taxonomic and thematic relations are co-activated when both types of semantic relations are simultaneously presented on a trial and measured by the online method (VWP). In the present study, we presented the two critical types of semantic competitors within a single visual display, with the aim to investigate whether the two types of semantic relations are co-activated. If this is the case, what is the relative strength and timing of taxonomic vs. thematic relations? Moreover, in previous studies, target objects were often co-present in the visual scene. When target objects are co-present in the visual display, most of fixations tend to shift toward these target objects, causing less fixations toward the other objects (Huettig and Altmann, 2005). In order to maximally capture looking behavior driven by semantic relations, we presented the targets verbally only and not visually to explore the activation of semantic relations when targets are not presented in visual display. In the experiment, participants listened to a spoken target word while viewing a visual display of four objects consisting of two competitors and two distractors. Fixation proportions on different types of objects in continuous time courses reflect participants' activation of the semantic relations. Based on the studies reviewed above, we expected both of the taxonomic and thematic competitors to attract more fixations than unrelated distractors. The more interesting question was whether the relative strength and time courses of activation of taxonomic vs. thematic relations differ.
Methods
Participants
Twenty-five native Chinese speakers (9 females, age 20–30 years, mean age 23 years) participated in the experiment. They were undergraduate students from universities in Beijing, China, and were paid RMB 20 (about 3.5 US $) for their participation. All participants had normal or corrected-to-normal vision. This study was approved by the ethics committee of the Institute of Psychology, Chinese Academy of Sciences. We obtained written informed consent from the participants of this study.
Materials and Design
Forty words were selected as the spoken target words to construct 40 visual displays, but four of these sets were eliminated for the reason of counterbalance of object positions, leaving 36 sets of critical items. All spoken words were recorded in a natural tone by a female native Chinese speaker. The mean duration of spoken words was 689 ms (ranged from 539 to 836 ms, SD = 76). Each visual display consisted of four pictures of common objects, one taxonomic competitor, one thematic competitor and two unrelated distractors (see Figure 1 for an example). The competitors were selected under criteria that the taxonomic competitor was as low as possible in thematic relatedness with the target and the thematic competitor was as low as possible in taxonomic relatedness with the target. Moreover, all word pairs were phonologically or orthographically unrelated (see Appendix 1 for a complete list of stimuli). The positions of the four pictures were fully counterbalanced in the display. In addition, across experimental conditions, the names of objects in displays were matched closely on word frequency, name agreement, and naming response time (Fs < 1, ps > 0.40; see Table 1 for the properties of the experimental materials).

Figure 1. For the spoken word “耳朵” (er3duo, “ear”), the visual display consisted of a taxonomic competitor “牙齿” (ya2chi3, “tooth”), a thematic competitor object “电话” (dian4hua4, “phone”) and two unrelated distractors, “蚂蚁” (ma3yi3, “ant”), and “镜子” (jing4zi, “mirror”).

Table 1. Properties and mean rating scores across conditions (standard deviations in parentheses).
Three subjective rating studies were conducted to evaluate the general semantic relatedness, taxonomic relatedness and thematic relatedness between the spoken target and the four objects within one set. The first rating asked participants to judge the general semantic relatedness between word pairs (either taxonomically or thematically related). The second rating asked participants to judge the taxonomic relatedness between word pairs, i.e., the degree of being in the same taxonomy. The third rating asked participants to judge the taxonomic relatedness between word pairs, i.e., the degree of connected via events or scenarios. All of the rating were based on a 7-point scale (1 = not related at all, 7 = strongly related), each collected rating scores form different sets of 17 participants. We followed the instructions (Appendix 2) used in Jones and Golonka (2012). Each rating contained all of the spoken targets, paired with the two competitors and two distractors, which were presented randomly. The means, SDs, minimums, and maximums on each of the three rating tasks are shown for each rating in Table 2. Separate One-way ANOVAs and LSD post-hoc tests (see Table 3) on the rating scores confirmed that: (1) both types of competitors were rated higher semantically related with the spoken target than the distractors, rating scores did not differ significantly between the two competitors, neither between the two distractors, (2) taxonomic competitor was highest rated in taxonomic relatedness rating, follow by thematic competitor, and two distractors were rated comparably low, and (3) thematic competitor was highest rated in thematic relatedness rating, follow by taxonomic competitor, and two distractors were rated comparably low. These rating results suggest that the selection of material was valid.
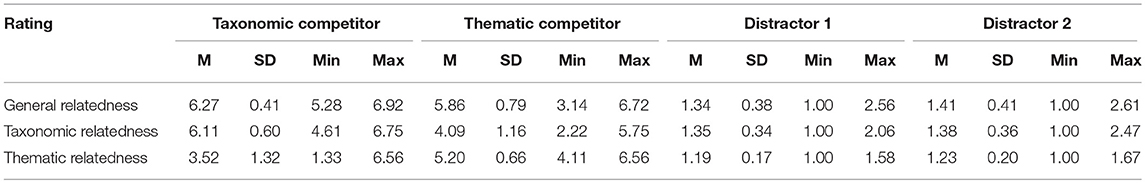
Table 2. Means, Standard Deviations, Minimums, and Maximums of ratings.
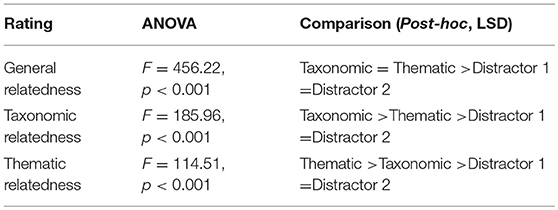
Table 3. Differences among objects for each rating.
Besides, 40 filler trials were added. Each filler set consisted of four objects with no overlap in meaning and visual form. One of the four objects was randomly selected as the target word. In total, 304 pictures were used, 218 of which were selected from the data base developed by Liu et al. (2011), and the other 86 were from Severens et al. (2005). All of the pictures were black-and-white line drawings and adjusted to the same size of 350 × 250 pixels and of approximately 13.67° × 9.77° visual angle.
Apparatus
Eye movements were recorded using an EyeLink 1000 tracker (SR Research, Mississauga, Ontario, Canada). Experimental materials were presented on a 21-in. CRT monitor (Sony Multiscan G520) with a 1,024 × 768 pixels resolution and a refresh rate of 150 Hz. The eye-tracking system was sampled at 1000 Hz. The participants placed their chins on a chin-rest and leaned their foreheads on a forehead rest to minimize head movements. Although viewing was binocular, eye movement data were collected only from the right eye. Participants were seated 58 cm from the video monitor.
Procedure
Participants were first asked to familiarize themselves with the pictures by viewing them in a booklet, with the name printed underneath each picture. Subsequently, the eye tracker was calibrated and validated, and re-calibration was conducted whenever the error was >1° during the experiment. The participants performed this procedure by means of a nine-point calibration, and the validation error was smaller than 0.5° of visual angle on average. A drift check was performed at the beginning of each trial, and then a blank screen was presented for 600 ms. The visual display was presented 2,000 ms before the onset of the spoken target word. Spoken target words were presented to participants through a headphone (Philips, SHM6110, China). Participants were asked to listen to the spoken words and view the displayed pictures without performing any explicit task. The visual display disappeared 4,000 ms after the spoken word onset. Each participant performed 36 critical and 40 filler trials which were randomly intermixed, following 8 practice trials. Trials were presented in a random order, and the entire experiment lasted approximately 20 min.
Results
Four areas of interest were defined, each overlapped with one of the pictures, corresponding to a rectangle (350 × 250 pixels). We calculated the mean fixation proportions of taxonomic competitor, thematic competitor, and distractors over successive 100 ms intervals. Figure 2 presents the distributions of the mean fixation proportions to each condition (distractor averaged from fixations on two distractors) from the onset of the target to 2,000 ms after the onset of the spoken target word.
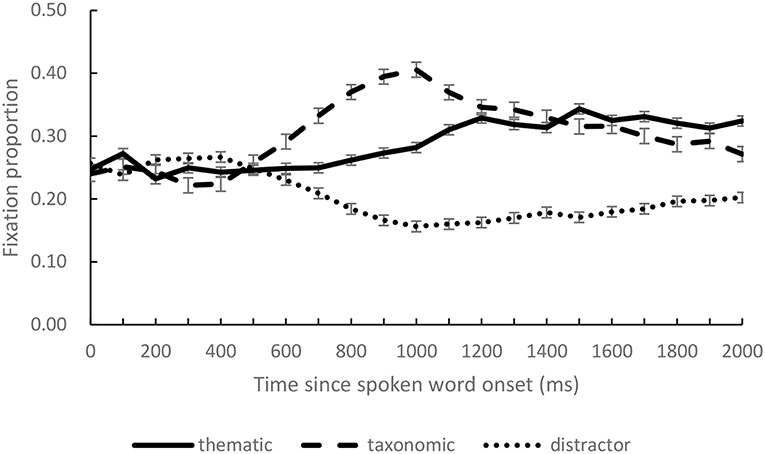
Figure 2. The fixation proportions on thematic competitor, taxonomic competitor, and distractor objects in the critical time window (0 on the x-axis indicates the onset of the spoken target words).
We adopted the growth curve analysis (GCA), a multilevel regression modeling technique using fourth-order orthogonal polynomials (Mirman et al., 2008), to quantify the effects of semantic relations. Four terms are included in such analyses, the intercept term represents the mean proportion of fixations over the entire window, the linear term captures variation in how rapidly looks to an object rise over time, the quadratic term captures variation in the curvature of the line representing looks to each object, and the cubic and quartic terms reflect the inflections at the extremities of the curve (Kalénine et al., 2012; Gambi et al., 2016). All analyses were carried out in R version 3.3.2 using the lme4 package (version 1.1-17). The first model compared fixed effects of object type (taxonomic competitor, thematic competitor and unrelated distractor) on all time terms, with unrelated distractors as the baseline, which compared the two types of competitors with the distractors respectively. The model also included random effects of participants on all time terms, which were captured with the crossed random effects structure, which estimates the random effects of participant and participant: object type (Mirman, 2014, p 70)1. We analyzed fixation proportions during the time window of 500–1,000 ms, which started from the earliest onset (539 ms) and ended at the latest offset of spoke target word (836 ms) plus about 200 ms (needed to programme and launch an eye movement, e.g., Hallett, 1986). This time window reflected the online processing of the spoken word. When a model includes three levels of a variable, the model first assessed baseline (Mirman, 2014, p. 95) and showed significant intercept (estimate = 0.198, p < 0.001) and linear (estimate = −0.078, p < 0.001) terms. Importantly, paired comparisons showed that both types of semantic competitors received more fixations than distractors (for intercept term, taxonomic vs. distractor: estimate = 0.144, p < 0.001; thematic vs. distractor: estimate = 0.062, p < 0.001). Effects on linear term were also significant, suggesting that fixation rose faster on both of the competitors than distractors (taxonomic vs. distractor: estimate = 0.207, p < 0.001; thematic vs. distractor: estimate = 0.109, p < 0.001) (see Table 4 for details).
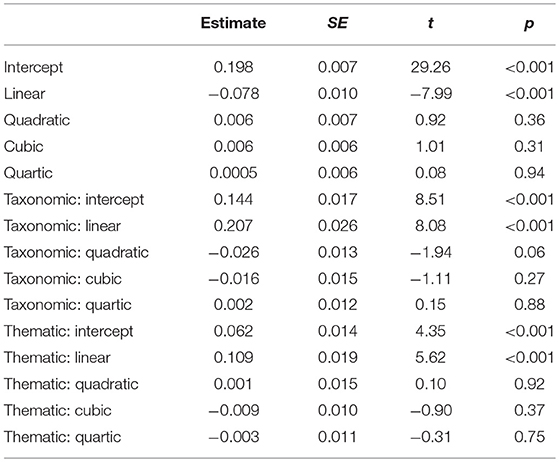
Table 4. Parameter Estimated for Analysis of Effect of Object Type (first model).
In the second model, we compared the fixation proportions on taxonomic and thematic competitors, with taxonomic competitors as the baseline. This model revealed a significant difference (intercept term, estimate = 0.342, p < 0.001; linear term, estimate = 0.130, p < 0.001). Paired comparison between the two competitors showed significant intercept term, estimate = −0.082, p < 0.001, indicating more fixations on the taxonomic than the thematic competitor. The linear term suggested that fixations on taxonomic competitor rose faster than that on the thematic competitor (estimate = −0.098, p < 0.001). Moreover, other inflections on the time courses of taxonomic and thematic effects did not differ significantly, as suggested by the quadratic, cubic and quartic terms (see Table 5 for details).
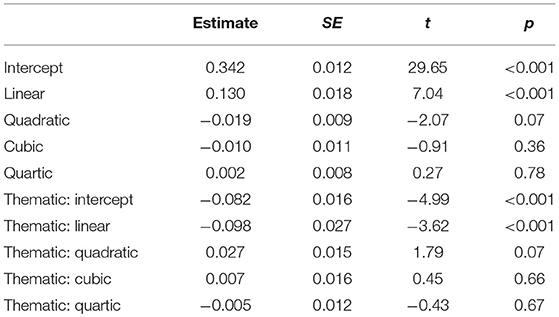
Table 5. Parameter Estimated for Analysis of Effect of Object Type (second model).
Besides the critical time window, we also calculated fixation proportions over successive 100 ms intervals during the preview period to test if there was any preference on the pictures caused by factors other than the spoken activation. We compared fixation proportions on the four objects in the same way as the first model in analysis of critical time window. The results suggested that fixation proportions on both of the competitors did not differ significantly from that on the distractors (taxonomic competitor: estimate = 0.011, p = 0.35; thematic competitor: estimate = −0.008, p = 0.36). Actually, during the preview period, fixation proportions on the four objects approximated the chance level 0.25 (see Figure 3).
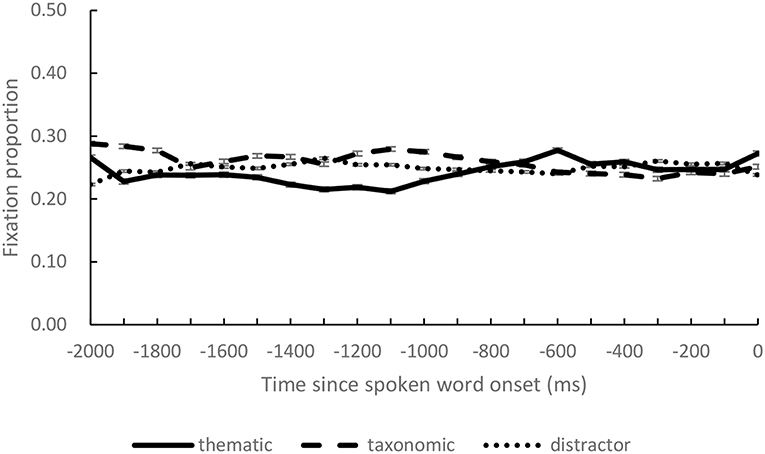
Figure 3. The fixation proportions on thematic competitor, taxonomic competitor, and distractor objects during preview (0 on the x-axis indicates the onset of the spoken target words).
Discussion
The present study examined the semantic representations invoked by a spoken word when the referent object was absent in the visual scene. Two types of semantic relations were compared, which are proposed as complementary systems and investigated widely (Mirman et al., 2017). We found that both the taxonomic and thematic competitors were fixated more than the unrelated distractors, indicating that the spoken input induced activation of these semantic representations. Rise of fixation proportion on taxonomic and thematic competitors started at similar time points, about 500 ms after the spoken word onset.
The finding that the magnitude of taxonomic effect and the rise speed of fixations on the taxonomic competitor was greater than the thematic effect was consistent with findings with the previous studies in which target objects were presented in visual displays (e.g., Mirman and Graziano, 2012). Mirman and Graziano (2012) proposed three possibilities for the stronger taxonomic effects relative to thematic effects: first, in their studies, the taxonomic competitors were also thematically related eliciting stronger semantic relation in taxonomic pairs; Second, since taxonomic relation is defined by shared semantic features, taxonomic competitors highly possibly shared visual features with the targets, which would cause more visual attention, as suggested by the finding that competition effects for shape similarity are earlier than for non-perceptual similarity (Yee et al., 2011); and Third, recognition of concrete objects used in their experiments was dominated by taxonomic knowledge (see also Duñabeitia et al., 2009), which resulted in stronger activation of taxonomic relation. In the present study, ratings on semantic relatedness suggested that taxonomic competitor and thematic competitor did not differ in general semantic relatedness, which rules out the possibility that taxonomic competitors are stronger in semantic relatedness. We speculate that visual similarity between taxonomic competitor and the target could have caused the larger taxonomic effect in the present study. Visual similarity naturally arises in taxonomically related items. As Mirman and Graziano (2012) emphasized, it is not practical to choose taxonomic competitor not visually similar with the target, Moreover, all of the concepts we used are concrete, and such concreteness of the stimuli used in the present study could also be responsible for the larger magnitude. This explanation is in agreement with the argument that concrete words are organized based on semantic features rather than association (Duñabeitia et al., 2009).
What could be the reason for the similar start points of two effects, in contrast to an earlier thematic effect relative to a taxonomic effect previously found by Kalénine et al. (2012)? Differences in experimental stimuli could be one potential reason. The stimuli used in the current study were mixed with natural and man-made objects, whereas all stimuli in Kalénine et al. were man-made objects. It is well-established that the role of taxonomic relation and thematic relation differ by concept domain, with thematic relation more important for artifacts/man-made/manipulable objects and taxonomic relation more important for natural objects (Bonthoux and Kalénine, 2007; Kalénine et al., 2009). The stimuli of artifacts such as tools used in Kalénine et al. could elicit earlier thematic effects, whereas the combined usage of natural and man-made objects could elicit the synchronization of taxonomic and thematic effects in the present study.
One of novelty in the current study was to adopt target-absent visual displays (Huettig et al., 2011 for a review). We provide evidence for activation of both types of semantic relations when the target was absent, as is the case in studies with target objects presented in the visual display (e.g., Kalénine et al., 2012; Mirman and Graziano, 2012), hence arguing for the idea that the semantic activation of a word does not depend on the its visual referent, but on the representations in mental lexicon. The activation invoked by spoken input without visual referent was also observed in children. Swingley and Fernald (2002) found that 2-year-old children maintained their fixation if the fixated object was the exact referent of what they heard, but shifted their gaze and searched other objects if the spoken word could not apply to the fixated object. These findings were interpreted as evidence that by 24 months, rapid activation in word recognition does not depend on the presence of the words' referents. Rather, very young children are capable of quickly and efficiently interpreting words in the absence of visual supporting context. Visual attention shift occurred because the visible objects did not match the representation activated by the spoken word. A recent auditory priming study (Willits et al., 2013) found semantic priming effect with bare spoken word stimuli (e.g., shorter looking time for “kitty-kitty-kitty” followed semantic prime “dog-dog-dog” than unrelated word “juice-juice-juice”), suggesting that young children activate lexical semantic knowledge in the absence of visual referents or sentence contexts. The present study, with adult participants, further suggests that spoken word invoked multiple semantic representations, reflected by more attention paid on the taxonomic and thematic related objects, when the visual display did not contain the target object.
In the present study, taxonomic and thematic competitors were depicted within the same visual display and both attracted more fixations than the unrelated distractors. As we mentioned in the Introduction section, taxonomic and thematic relations are found to be dissociated both in behavioral and neural studies (e.g., Bonthoux and Kalénine, 2007; Kalénine et al., 2009; de Zubicaray et al., 2013). In the perspective of language function, taxonomic relation contributes to organize classes of objects, concepts, and even people appropriately based on features, while thematic relation conveys knowledge about events and scenarios, which help us to establish expectation about upcoming events and complement one's knowledge about features and taxonomic relations (Estes et al., 2011). The present finding implies that although located in different hubs, taxonomic and thematic relations are activated by the same linguistic input, which might help us to establish a more complete representation of the intended concept.
In conclusion, the present study suggested that taxonomic and thematic relations are co-activated by the spoken word, even when the referent object is absent from the visual context. These two relations are activated at similar time points, but the taxonomic effect is stronger and develops faster than the thematic relation in the processing of concrete concepts.
Ethics Statement
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with APA regulations.
Author Contributions
PX, QQ, and XL inspired the idea of the study, designed the experiment, collected the data, conducted the data analysis, interpreted the results, and wrote the manuscript. WS provided the instruction to the execution of the experiment and data analysis. All authors agreed on the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (NSFC), No. 31771212, and the German Research Foundation (DFG) and NSFC in joint project Cross-modal Learning, DFG TRR-169/NSFC No. 61621136008 to the corresponding author and the last author.
Footnotes
1. ^R syntax for the models (model 1 and 2 used the same syntax, differed in the content of data):
t <-poly(unique(fix$timeBin),4)
fix[,paste(“ot”,1:4,sep = "“)] <-
t[fix$timeBin,1:4]
m.full1 <-lmer(meanFix~(ot1+ot2+ot3+ot4)*Object +
((ot1+ot2+ot3+ot4)*Object | Subject),
control = lmerControl(optimizer = ”bobyqa"),
data = fix, REML = FALSE)”
References
Bonthoux, F., and Kalénine, S. (2007). Preschoolers' superordinate taxonomic categorization as a function of individual processing of visual vs. contextual/functional information and object domain. Cogn. Brain Behav. 11, 713–722.
Collins, A. M., and Loftus, E. F. (1975). A spreading-activation theory of semantic processing. Psychol. Rev. 82, 407–428.
Cooper, R. M. (1974). The control of eye fixation by the meaning of spoken language: a new methodology for the real-time investigation of speech perception, memory, and language processing. Cogn. Psychol. 6, 84–107.
de Zubicaray, G. I., Hansen, S., and McMahon, K. L. (2013). Differential processing of thematic and categorical conceptual relations in spoken word production. J. Exp. Psychol. 142, 131–142. doi: 10.1037/a0028717
Duñabeitia, J. A., Avilés, A., Afonso, O., Scheepers, C., and Carreiras, M. (2009). Qualitative differences in the representation of abstract versus concrete words: evidence from the visual-world paradigm. Cognition 110, 284–292. doi: 10.1016/j.cognition.2008.11.012
Estes, Z., Golonka, S., and Jones, L. L. (2011). “Thematic thinking: The apprehension and consequences of thematic relations,” in Psychology of Learning and Motivation, eds B. Ross, Vol. 54 (Burlington, MA: Academic Press), 249–294.
Estes, Z., and Jones, L. L. (2009). Integrative priming occurs rapidly and uncontrollably during lexical processing. J. Exp. Psychol. 138, 112–130. doi: 10.1037/a0014677
Gambi, C., Pickering, M. J., and Rabagliati, H. (2016). Beyond associations: Sensitivity to structure in pre-schoolers' linguistic predictions. Cognition 157, 340–351. doi: 10.1016/j.cognition.2016.10.003
Hallett, P. E. (1986). “Eye movements,” in Handbook of Perception and Human Performance, eds K. R. Boff, L. Kaufman, and J. P. Thomas (New York, NY: Wiley), 10.1–10.112.
Huettig, F., and Altmann, G. T. (2005). Word meaning and the control of eye fixation: semantic competitor effects and the visual world paradigm. Cognition 96, B23–B32. doi: 10.1016/j.cognition.2004.10.003
Huettig, F., Rommers, J., and Meyer, A. S. (2011). Using the visual world paradigm to study language processing: a review and critical evaluation. Acta Psychol. 137, 151–171. doi: 10.1016/j.actpsy.2010.11.003
Jones, L. L., and Golonka, S. (2012). Different influences on lexical priming for integrative, thematic, and taxonomic relations. Front. Hum. Neurosci. 6:205. doi: 10.3389/fnhum.2012.00205
Jones, L. L., Wurm, L. H., Calcaterra, R. D., and Ofen, N. (2017). Integrative priming of compositional and locative relations. Front. Psychol. 8:359. doi: 10.3389/fpsyg.2017.00359
Kalénine, S., Mirman, D., Middleton, E. L., and Buxbaum, L. J. (2012). Temporal dynamics of activation of thematic and functional knowledge during conceptual processing of manipulable artifacts. J. Exp. Psychol. Learn. Mem. Cogn. 38, 1274–1295. doi: 10.1037/a0027626
Kalénine, S., Peyrin, C., Pichat, C., Segebarth, C., Bonthoux, F., and Baciu, M. (2009). The sensory-motor specificity of taxonomic and thematic conceptual relations: a behavioral and fMRI study. Neuroimage 44, 1152–1162. doi: 10.1016/j.neuroimage.2008.09.043
Liu, Y., Hao, M., Li, P., and Shu, H. (2011). Timed picture naming norms for Mandarin Chinese. PLoS ONE 6:e16505. doi: 10.1371/journal.pone.0016505
Mirman, D. (2014). Growth Curve Analysis and Visualization Using R. Boca Raton, FL: Chapman and Hall/CRC.
Mirman, D., Dixon, J. A., and Magnuson, J. S. (2008). Statistical and computational models of the visual world paradigm: growth curves and individual differences. J. Mem. Lang. 59, 475–494. doi: 10.1016/j.jml.2007.11.006
Mirman, D., and Graziano, K. M. (2012). Individual differences in the strength of taxonomic versus thematic relations. J. Exp. Psychol. 141, 601–609. doi: 10.1037/a0026451
Mirman, D., Landrigan, J. F., and Britt, A. E. (2017). Taxonomic and thematic semantic systems. Psychol. Bull. 143:499. doi: 10.1037/bul0000092
Schwartz, M. F., Kimberg, D. Y., Walker, G. M., Brecher, A., Faseyitan, O. K., Dell, G. S., et al. (2011). Neuroanatomical dissociation for taxonomic and thematic knowledge in the human brain. Proc. Natl. Acad. Sci.U.S.A. 108, 8520–8524. doi: 10.1073/pnas.1014935108
Severens, E., Van Lommel, S., Ratinckx, E., and Hartsuiker, R. J. (2005). Timed picture naming norms for 590 pictures in Dutch. Acta Psychol. 119, 159–187. doi: 10.1016/j.actpsy.2005.01.002
Swingley, D., and Fernald, A. (2002). Recognition of words referring to present and absent objects by 24-month-olds. J. Mem. Lang. 46, 39–56. doi: 10.1006/jmla.2001.2799
Tanenhaus, M. K., Spivey-Knowlton, M. J., Eberhard, K. M., and Sedivy, J. C. (1995). Integration of visual and linguistic information in spoken language comprehension. Science 268:1632.
Wamain, Y., Pluciennicka, E., and Kalénine, S. (2015). A saw is first identified as an object used on wood: ERP evidence for temporal differences between thematic and functional similarity relations. Neuropsychologia 71, 28–37. doi: 10.1016/j.neuropsychologia.2015.02.034
Willits, J. A., Wojcik, E. H., Seidenberg, M. S., and Saffran, J. R. (2013). Toddlers activate lexical semantic knowledge in the absence of visual referents: evidence from auditory priming. Infancy 18, 1053–1075. doi: 10.1111/infa.12026
Yee, E., Huffstetler, S., and Thompson-Schill, S. L. (2011). Function follows form: activation of shape and function features during object identification. J. Exp. Psychol. 140, 348–363. doi: 10.1037/a0022840
Appendix 1: Stimuli Used in the Critical Trials

Appendix 2: Instructions for Rating Studies
General Relatedness
请判断下列词对之间的语义相关性, 1表示无关, 7表示非常相关。请用1-7((数字1, 2, 3, 4, 5, 6, 7))进行评定。语义相关性包括类别相关(属于同一个类别的词对, 如蝴蝶-蜜蜂都是昆虫, 应该评为高相关, 蝴蝶-筷子分别属于不同类别, 应该评为低相关)和主题相关性(两个词虽然不是同一类事物, 但是可以通过事件、功能或场景产生联系, 如螃蟹-大海, 伤口-绷带都应该评定为高相关, 螃蟹-考试, 伤口-水杯为低相关)。
Please rate the semantic relatedness between each of the following word pairs on a scale from 1 (not thematically connected) to 7 (highly thematically connected). Please use the full range of the scale (1, 2, 3, 4, 5, 6, or 7) in indicating your responses. Semantic relatedness includes taxomomical relatedness (two words belong to the same specific category, e.g., BUTTERFLY and BEE are both types of insect, thus are taxonomically connected, whereas BUTTERFLY and CHOPSTIC belong to different categories) and thematic relatedness (the concepts are linked together in a common scenario, event, or function, e.g., CRAB-SEA and WOUND-BANDAGE are both highly related, whereas CRAB-EXAM and WOUND-CUP are lowly related).
Taxonomic Relatedness
请判断下列词对之间的类别相关性, 1表示无关, 7表示非常相关。请用1-7((数字1, 2, 3, 4, 5, 6, 7))进行评定。类别相关性是指两个词语所代表的事物属于同一类别的程度, 如蝴蝶-蜜蜂都是昆虫, 应该评为高相关。蝴蝶-筷子分别属于不同类别, 应该评为低相关。
Please rate the taxonomic relatedness between each of the following word pairs on a scale from 1 (not thematically connected) to 7 (highly thematically connected). Please use the full range of the scale (1, 2, 3, 4, 5, 6, or 7) in indicating your responses. Taxonomic relatedness refers to the extent to which the two words belong to the same specific category. For example, BUTTERFLY and BEE are both types of insect, thus these items are taxonomically connected. Whereas BUTTERFLY and CHOPSTIC belong to different categories.
Thematic Relatedness
请判断下列词对之间的类别相关性, 1表示无关, 7表示非常相关。请用1-7((数字1, 2, 3, 4, 5, 6, 7))进行评定。类别相关性是指两个词语所代表的事物属于同一类别的程度, 如蝴蝶-蜜蜂都是昆虫, 应该评为高相关。蝴蝶-筷子分别属于不同类别, 应该评为低相关。
Please rate the thematic relatedness between each of the following word pairs on a scale from 1 (not thematically connected) to 7 (highly thematically connected). Please use the full range of the scale (1, 2, 3, 4, 5, 6, or 7) in indicating your responses. Thematic relatedness refers to the extent to which the concepts are linked together in a common scenario, event, or function. For example, CREAM is often added to COFFEE, and CHALK is used to write on a BLACKBOARD. Thus, these items are thematically connected. Please note that thematically connected items may not necessarily share similar features. For example, BLACKBOARD and CHALK are different shapes and sizes and are made out of different materials.
Keywords: semantic system, taxonomic relation, thematic relation, co-activation, visual-world paradigm
Citation: Xu P, Qu Q, Shen W and Li X (2019) Co-activation of Taxonomic and Thematic Relations in Spoken Word Comprehension: Evidence From Eye Movements. Front. Psychol. 10:964. doi: 10.3389/fpsyg.2019.00964
Received: 16 January 2019; Accepted: 11 April 2019;
Published: 03 May 2019.
Edited by:
Ludovic Ferrand, Centre National de la Recherche Scientifique (CNRS), FranceReviewed by:
Solene Kalenine, UMR9193 Laboratoires Sciences Cognitives et Sciences Affectives (SCALab), FranceLara L. Jones, Wayne State University, United States
Copyright © 2019 Xu, Qu, Shen and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Qingqing Qu, cXVxcUBwc3ljaC5hYy5jbg==