 Sebotse Dolly Kgoale1
Sebotse Dolly Kgoale1 Abdullahi Adekilekun Jimoh1
Abdullahi Adekilekun Jimoh1 Fidele Tugizimana2,3
Fidele Tugizimana2,3 Adrian Abrahams4
Adrian Abrahams4 Oluwafemi Ayodeji Adebo1*
Oluwafemi Ayodeji Adebo1*- 1Centre for Innovative Food Research (CIFR), Department of Biotechnology and Food Technology, Faculty of Science, University of Johannesburg, Doornfontein Campus, Johannesburg, South Africa
- 2Research Centre for Plant Metabolomics, Department of Biochemistry, University of Johannesburg, Johannesburg, South Africa
- 3International Research and Development Division, Omnia Group, Ltd., Johannesburg, South Africa
- 4Department of Biotechnology and Food Technology, Faculty of Science, University of Johannesburg, Doornfontein Campus, Johannesburg, South Africa
Most existing research on amahewu has primarily examined the microbial and physicochemical properties of beverages, leaving the overall metabolite profile insufficiently characterized. Metabolomic analysis enables the monitoring of metabolite shifts throughout fermentation, revealing how microbial processes drive variations in bioactive compounds. Therefore, this study employed targeted metabolomics to investigate dynamic changes in amino acids and phenolic compounds during the production of amahewu, a traditional Southern African fermented maize beverage. White and yellow Zea mays L. varieties were fermented with malted sorghum inoculum, and samples were analyzed using liquid chromatography–tandem mass spectrometry (LC-MS/MS) with multiple reaction monitoring across four production stages. Chemometric analyses (PCA, OPLS-DA) revealed distinct stage- and maize type-dependent clustering with model validation (Q2 > 0.5; CV-ANOVA, p < 0.05). Across both maize types, seven metabolites discriminated raw from cooked maize, with valine (white maize) and alanine (yellow maize) serving as unique markers. Proline was the most abundant amino acid (>1.0 × 108 ng/g in raw maize), but declined by >80% during fermentation, consistent with microbial utilization. Conversely, threonine and valine increased >3-fold in fermented samples, reflecting proteolysis and microbial synthesis. Apigenin represented >70% of quantified phenolics at all stages, whereas caffeic acid decreased sharply post-cooking (FC = 0.07, p = 7.03 × 10−9), and avenanthramides emerged de novo in fermented samples. Correlation analysis showed strong positive feed-forward associations among amino acids (r > 0.8) and feedback-driven phenolic transformations, while pathway enrichment identified isoquinoline alkaloid biosynthesis (impact = 0.41) in white maize and alanine/aspartate/glutamate metabolism (impact = 0.13) in yellow maize as dominant routes. Collectively, this study provides insight into the quantitative changes in metabolic composition at different stages of production, which reflect changes in nutritional, biochemical, and health-promoting properties, as well as microbial communities.
1 Introduction
Maize (Zea mays) is a monoic annual crop belonging to the family Gramineae and is grown in nearly every part of the world (1). It provides up to 30% of the world's food calories to 4.5 billion people (2, 3). The most used maize types are white and yellow maize, for human consumption (4, 5). These maize types undergo different processes to be edible, including fermentation which can be performed in two ways: the direct fermentation of maize kernels or by first milling the maize to produce a maize meal. Traditional maize fermentation requires minimal ingredients and involves only basic preparation and processing. There is a multitude of diverse maize-fermented beverages, such as Busaa (Kenya), Togwa (Tanzania), Ogi (Nigeria), Amahewu (South Africa) Pozol (Mexico), and Chicha (Brazil), due to numerous food-microbe pairings (3, 6–8). Among these, amahewu stands out as one of Southern Africa's most consumed non-alcoholic fermented beverages, with an estimated annual household consumption exceeding 40 million liters across South Africa, Zimbabwe, and neighboring countries (9).
Amahewu is valued for its refreshing taste and potential health benefits, which are influenced by its nutritional composition (9, 10). It is used as a refreshing beverage by adults and children in preschool and school-age groups, and as weaning food for infants among low-income rural dwellers (11, 12). The chemistry and metabolite profile of amahewu is rich and complex, encompassing a variety of organic acids, vitamins, minerals, amino acids, and bioactive compounds (9). Despite its nutritional and cultural importance, amahewu remains largely artisanal, with unstandardized fermentation leading to marked variability in quality, safety, and sensory attributes. The absence of comprehensive biochemical profiling has limited efforts to enhance consistency and nutritional optimization. Thus, metabolomics analysis is urgently needed to elucidate the molecular transformations during fermentation, identify key nutritional and bioactive compounds, and provide a scientific basis for product standardization at both community and industrial scales.
The growth and use of foodomics have kept pace with consumers' growing interest and worry over the ingredients in their food and its safety. This is an emerging interdisciplinary field that integrates advanced omics technologies (such as genomics, transcriptomics, proteomics, metabolomics, and lipidomics) to study food (such as amahewu) and its impact on human health and nutrition (13). For this study, we employed targeted metabolomics, a hypothesis-driven approach that focuses on detecting specific metabolites associated with a particular class or metabolic pathway, rather than all components in the sample (14). This helps detect, identify, and monitor changes throughout the production of amahewu, as well as related main raw materials and the inoculum used (15). The combination of mass spectrometry and liquid chromatographic separation methods significantly enhances the chemical analysis capabilities of highly complex food samples. This technology enables the physical separation of thousands of metabolites, providing a more comprehensive view of the metabolome (16). The data from the metabolome is multidimensional, intricate, and complex, requiring the application of chemometric approaches to investigate the rich information it contains. Examples of such techniques include orthogonal partial least square discriminant analysis (OPLS-DA) and principal component analysis (PCA), which have been used to give a summary of the dataset, highlighting treatment-related trends, and indicating the metabolites that primarily contribute to differences in treatments, sample sources and other experimental variations (17–20).
As primary and secondary metabolites, amino acids and phenols have many prominent functions. They play pivotal roles in the flavor, nutritional value, and overall quality of food products including amahewu. Amino acids are fundamental to protein synthesis, scavenge free radicals and thus have antioxidant properties as well as influence the sensory attributes of the final product, while phenolics are known for their antioxidant properties and contribution to the beverage's taste and color (21). In addition to nutritional and functional interest, amino acids and phenolic acids have been recognized as important biomarkers for foodomics studies (21, 22).
Food processing, such as fermentation produces substantial metabolic changes in food products, highlighting the need to grasp these alterations. Identifying and understanding the metabolites involved is crucial for unraveling the functions and transformations that occur during production. This study aims to profile amino acids and phenolics during amahewu production as both white and yellow Zea mays L. varieties possess unique features that could influence fermentation dynamics and the resultant amehewu's composition. This was carried out by employing targeted metabolomics using LC-MS/MS in combination with multivariate analysis PCA and OPLS-DA to track variations and offer a full profile of amino acids and phenolic content at each stage of amahewu production.
2 Methods
2.1 Sample collection
White maize (Zea mays L.) and yellow maize (Zea mays L.) grains harvested in 2023 were obtained from the Agricultural Research Council (ARC-LNR Teliersaad breeder seed), Pretoria, South Africa. Sorghum grains (Sorghum bicolor (L.) Moench, cultivar NS 5655, 2023 harvest) were sourced from Agricol, Pretoria, South Africa. All samples were transported under dry conditions and stored at room temperature until further use.
2.2 Maize preparation
The white and yellow maize grains (4 kg) were weighed using a Mettler Toledo PL202-S scale (Greifensee, Switzerland), washed, and dried overnight at 38 °C in a drying oven (Prolab, Manama, Bahrain). The dried maize grains were milled using a laboratory mill (3600 Perten, Hudding, Sweden) with a 1.5 mm grain size sieve and then passed through a sieve shaker (Analysette 3 Spartan, Fritsch GmbH, Idar-Oberstein, Germany).
2.3 Sorghum preparation and malting
Sorghum grains (2 kg) were weighed on a weighing balance (Mettler Toledo PL202-S, Greifensee, Switzerland) and submerged in water for 1 day at room temperature. After a day, the water was drained and the sorghum grains were spread on a tray to germinate at room temperature, with water spraying in between the 48 h of germination. Two to four centimeters of sprouts germinated and were dried at 40 °C overnight in a drying oven (Prolab, Manama, Bahrain). The dried sprouted grains were milled using a laboratory mill (3600 Perten, Hudding, Sweden) with a 1.5 mm grain size and then passed through a sieve shaker (Analysette 3 Spartan, Fritsch GmbH, Idar-Oberstein, Germany).
2.4 Preparation of amahewu
Approximately 200 g of the milled maize (yellow and white maize) was mixed with 500 mL of autoclaved water in a 1 L autoclaved glass beaker (Sample 1; RWM/RYM). The mixture was cooked on a hot plate at 95 ± 2 °C for 20–30 min with continuous stirring using a sterilized glass rod until a thick porridge was formed (Sample 2; CWM/CYM). After cooking, the porridge was allowed to cool to about 30 °C and transferred into a 1 L Schott bottle. As an inoculum, 30 g of malted sorghum flour was added to the cooled porridge, thoroughly mixed in a 1 L Schott bottle (Sample 3; IWM/IYM), and then incubated at 30 °C (Labcon, Krugersdorp, South Africa) for 26 h to obtain the fermented amahewu (WMA/YMA). The malted sorghum used as inoculum was designated as Sample 5 (Sinc).
After each phase and production, samples of raw maize, cooked maize, inoculated paste, and fermented amahewu (Table 1) were collected into sterile 50 mL Falcon tubes for subsequent physicochemical and metabolomic analyses. The remaining samples were refrigerated at −80 °C and then freeze-dried using a benchtop freeze dryer (Beijer Electronics HT40; Telstar LyoQuest, Terrassa, Spain). The flours obtained from freeze-drying were stored in Ziplock bags at 4 °C for further analysis.

Table 1. Sample details, processing steps and identifications.
2.5 Determination of amino acids and phenolics using liquid chromatography-mass spectrometry
Approximately 2.5 g of each powdered sample was weighed, and 6 mL of 80% methanol was added as an extraction solvent. This corresponds to a solid-to-solvent ratio of approximately 1:2.4 (w/v). The mixture was vortex-mixed for 1 min to ensure uniform dispersion. The solution was ultrasonicated for 30 min (Scientech 704, Labotech, Johannesburg, South Africa) at a temperature of 40 °C, followed by centrifugation for 5 min at 3.000 × g (Eppendorf 5702R, Merck, Johannesburg, South Africa). The supernatant was filtered through a 0.22 μm polyethersulfone membrane filter into vials.
Samples and working solutions of standards were analyzed on an ultra-fast liquid chromatography (UFLC) system, fitted with Shim-pack GIST C18 column (2 μm; 100 × 2.1 mm l.D) (Shimadzu, Kyoto, Japan), housed in an oven with thermostat set at 40 °C. Chromatographic separation was performed using gradient elution, utilizing eluent A (MilliQ water with 0.1% formic acid) and eluent B (methanol with 0.1% formic acid) (Romil Chemistry, UK) at a constant flow rate of 0.2 mL min−1. The gradient elution conditions used were as follows: 1 min 98% A, 2% B; 1–3 min 95% A, 5% B; 2 min 90% A 10% B; 2 min 50% A, 50% B, then changed to the initial conditions. The total chromatographic run time was 10 min, and the injection volume was 2 μL for phenolics and 3 μL for amino acids. The MS conditions were as follows: nitrogen was supplied as the drying gas at a flow rate of 10 L min−1 and as the nebulizing gas at 3 L min−1; the heating gas flow was maintained at 10 L min−1. The interface temperature and voltage were set at 300 °C and 4 kV, respectively, while the DL temperature was 250 °C, and the heat block temperature was set to 400 °C. Three biological replicates were prepared for each sample, and each biological replicate was analyzed in triplicate on the LC-MS system.
2.6 Data analysis and interpretation
Data generated from LC-MRM-MS (liquid chromatography-mass spectrometry with multiple reaction monitoring) analyses were processed using LabSolution Quant BrowserTM (Shimadzu, Kyoto, Japan), where the concentrations of the unknown samples were extrapolated from the respective calibration curves. From obtaining the extrapolated concentrations expressed in parts per billion (ppb) (for amino acids) and parts per million (ppm) (for phenolics), the final concentrations were calculated and then converted to ng/g, respectively. The generated data was first imported into the SIMCA software, version 14.1 (Umetrics, Umea, Sweden) for chemometrics/multivariate statistical analysis. The imported data was scaled to unit variance prior to PCA and OPLS-DA.
To perform supervised and unsupervised multivariate statistical analysis, PCA was performed to visualize the datasets firsthand and detect outliers, while OPLS-DA was used to assess the methodology's ability to discriminate between production steps, maize type and inoculum. These models were validated after being built using 7-fold cross-validation. For PCA, the cumulative R2 (explained variation) and Q2 (predicted variation) of the models used were higher than 0.5. Furthermore, for OPLS-DA models, the analysis of variance testing of cross-validated predictive residuals (CV-ANOVA), p-values were below 0.05 (23). All curves showed excellent linearity (R2 = 0.991–0.999).
The generated concentration data matrix was also uploaded into MetaboAnalyst 5.0 (http://www.metaboanalyst.ca) for metabolite-metabolite correlation analysis (MMCA) using Pearson's correlation function; and for pathways analysis using metabolomics pathway analysis (MetPA) tool, a part of MetaboAnalyst 5.0. Since many pathways are tested at the same time, both Holm-Bonferroni and false discovery rate (FDR) procedures were used to adjust for the statistical p-values from enrichment analysis.
3 Results and discussion
3.1 Multivariate data analysis of amino acids and phenolics profiles from white and yellow maize-produced amahewu inoculated with malted sorghum.
As mentioned in the experimental section, chemometrics modeling was applied to reduce data complexity, enhance interpretation, and identify key metabolic profiles that define the production of amahewu. The computed PCA model revealed sample groupings and natural trends related to maize type (yellow vs. white), treatment (fermented vs. non-fermented), and different production stages (Figure 1). Figure 1A shows a clear separation of sample groups based on their metabolite profiles, with IS, CYM, and RYM forming distinct clusters away from the central overlapping groups (CWM, FWM, FYM, IWM, IYM, and RWM). The tight clustering of IS indicates high within-group similarity, while the wide separation of CYM along principal component 1 (PC1) and RYM along principal component 2 (PC2) suggests that these groups possess unique biochemical compositions compared to the others. In contrast, the central overlap of several groups reflects shared metabolic features and less pronounced differentiation.

Figure 1. Principal component analysis (PCA) plots displaying the sample clusters that were investigated. (A) PCA score plot of both yellow and white maize (PC1; 23.8% vs. PC2; 4.7%). (B) PCA score plot of white maize (PC1; 33.4% vs. PC2; 18.6%) (C) PCA score plot of yellow maize (PC1; 33.8% vs. PC2; 18.6%). The PCA scores represent the production steps of amahewu using white and yellow maize with malted sorghum as an inoculum. CWM, cooked white maize; FWM, fermented white maize; IWM, inoculated white maize; RWM, raw white maize; CYM, cooked yellow maize; FYM, fermented yellow maize; IYM, inoculated yellow maize; RYM, raw yellow maize; IS, inoculated sorghum.
In the score plots indicating white maize production, a clear separation of IS and FWM from the other groups was observed, with IS positioned at the top and FWM at the lower end, exhibiting minimal overlaps. RWM is distinctly separated at the top, while CWM and IWM clustered closely together at the lower region (Figure 1B). In the score plots indicating yellow maize production, RYM, CYM, and IYM are closely clustered to each other (Figure 1C). The separation and clustering patterns highlight the extent of metabolic alterations occurring during the processing of amahewu, particularly the influence of the inoculum on the metabolite composition of the final products.
The PCA loading plots revealed clear metabolic differentiation across the production stages of amahewu (Figure 2). In Figure 2A, samples representing the final product and inoculum were separated along the positive axis of PC1, whereas the raw, cooked, and inoculated pastes clustered negatively, indicating progressive biochemical changes associated with fermentation. For white maize (Figure 2B), the RWM and IWM stages clustered with caffeic acid, apigenin, and ferulic acid, compounds typically found in unfermented maize. The final product (FWM) correlated with valine, threonine, and proline, suggesting increased amino acid synthesis during fermentation. Similarly, in yellow maize (Figure 2C), RYM and IYM samples were closely associated with apigenin and caffeic acid, while the FYM aligned with amino acids such as valine, threonine, and proline. The inoculum occupied a central position, linked to ferulic acid and serine, reflecting its intermediate enzymatic role in initiating the fermentation process.

Figure 2. Principal component analysis (PCA) loading plots displaying the discriminant metabolites (A) yellow and white maize (PC1; 23.8% vs. PC2; 4.7%), (B) white maize (PC1; 33.4% vs. PC2; 18.6%), (C) yellow maize (PC1; 33.8% vs. PC2; 18.6%).
The unsupervised chemometric analysis therefore underscores the transformative effect of fermentation on the metabolome of cooked maize and the baseline metabolic differences between raw maize and malted sorghum. These differences in grouping reflect underlying metabolic profiles shaped by carbohydrate availability, enzymatic activity, and secondary metabolite release. Raw maize is rich in starch, which is structurally resistant to enzymatic hydrolysis due to its compact granule structure and higher amylose-to-amylopectin ratio (24). This makes raw maize less directly fermentable, as microbial metabolism requires additional enzymatic activity to liberate fermentable sugars (25). Conversely, malted sorghum exhibits elevated endogenous enzymatic activity, particularly α-amylases and β-glucanases, which are activated and hydrolyze starches into simpler sugars, enhancing microbial accessibility and fermentation processes (26, 27). The close clustering of intermediate maize samples (CWM, IWM, CYM, IYM) suggests gradual yet coordinated biochemical transitions, where starch hydrolysis, protein breakdown, and phenolic release occur incrementally.
3.2 Discriminant analysis of metabolites in white maize vs. yellow maize amahewu samples
Orthogonal partial least square discriminant analysis was employed to further interpret differential metabolite profiles, separating multivariate relationships into predictive and orthogonal variation and identifying the metabolic features driving groups discrimination. The computed and validated models showed strong classification with no overfitting, as confirmed by cross-validation and permutation testing (n = 50). Discriminant biomarkers (unique Rt and m/z values) were identified from S-plots and confirmed with VIP scores, which rank variables by contribution and stability (Supplementary Figures S1–18). Thus, statistically significant features contributing to class separation are present in Table 2. The resultant OPLS-DA model comparisons across white and yellow maize, as well as production steps (RWM vs. RYM, CWM vs. CYM, IWM vs. IYM, and FWM vs. FYM), showed good sample classification, suggesting that the two groups have differential metabolite profiles.

Table 2. Significant metabolites contributing to the differences at different stages of production.
Across both white and yellow maize productions, seven metabolites discriminate raw maize from cooked maize paste, with five AAs and two PCs. However, the specific discriminants differed: valine characterized white maize, whereas alanine was unique to yellow maize. Both are alkyl nonpolar amino acids compared to the other three AAs (cysteine, serine, and tyrosine), which are neutral polar. Between cooked and inoculated maize, eight metabolites discriminate in white maize compared to only five in yellow maize, with overlap restricted to three shared metabolites. Distinct differences included cysteine, aspartic acid, threonine, valine, and apigenin in white maize, and serine and ferulic acid in yellow maize. At the final stage, discriminants between inoculated maize and the final product comprised eight metabolites in white maize (five AAs and three PCs) vs. five in yellow maize (three AAs and two PCs). Here, alanine, cysteine, and avenonthiamide A characterized white maize, while apigenin, a phenolic compound, uniquely defined yellow maize.
The significant metabolites in white maize production increased from seven in RWM vs. CWM and CWM vs. IWM to eight in IWM vs. FWM (Table 2). Serine is only present in the first discriminant and the change addition of aspartic acid and avenonthiamide B in the second discriminant. The final stage added alanine, avenonthiamide A, and ferulic acid, whereas cysteine, threonine, and valine were consistently present throughout. In yellow maize, there was a decrease in fluctuations; seven metabolites were present as significant discriminants between RYM vs. CYM, five in CYM vs. IYM and six in IYM vs. FYM (Table 2). Avenonthiamide B and ferulic acid were additional changes in the second discrimination, complementing tyrosine, serine and cysteine that were present in the first discrimination, and the additional changes of aspartic acid, threonine, valine and apigenin in the final discrimination.
The structural characterization of metabolites, supported by OPLS-DA analysis, demonstrated that the metabolic profiles of the groups were considerably different across production stages. Complementary microbial analysis further revealed dynamic shifts in microbial populations during fermentation, indicating that microbial succession plays a central role in shaping metabolite profiles. This is consistent with the bacterial community structure previously reported by Daji, Green (28), which identified a diverse assemblage of genera, including Paenibacillus, Weissella, Lactococcus, Enterococcus, Bacillus, Leuconostoc, Pseudomonas, and Pediococcus, in optimized white- and yellow-maize mahewu. Their 16S rRNA profiling demonstrated that microbial diversity and fermentation conditions drive the acidification and flavor development of the beverage (28). Besides lactic acid bacteria (LAB), amahewu fermentation involves other microbial consortia comprising yeasts, Bacillus, and Enterobacter species. These organisms contribute to amino-acid metabolism and phenolic biotransformation, collectively driving the biochemical remodeling observed in the metabolomic profiles. The amino acid profile is particularly critical, as it directly reflects the nutritional quality of proteins and follows similar patterns of protein turnover and degradation (29). Notably, white maize production stages exhibited a greater increase in amino acid metabolites compared to yellow maize, suggesting more extensive proteolytic activity and microbial remodeling in white maize.
3.3 Quantification analysis of the discriminating metabolites
To gain a deeper understanding of the biochemical differences underlying the observed group separations, the discriminating metabolites identified through multivariate analyses were quantitatively assessed (Figure 3). Apigenin was the most abundant compound throughout all stages, consistently representing more than 70% of the total quantified phenolics (Figure 3A). This is expected, as maize naturally contains significant levels of apigenin-C-glycosides such as vitexin and orientin, which can undergo enzymatic hydrolysis during fermentation to release free apigenin (30, 31). The persistence of apigenin across all stages, including after heat treatment and extended fermentation, highlights its stability under thermal and acidic conditions, as well as its resistance to microbial degradation. This stability, coupled with its continuous liberation during fermentation, underscores its importance as a major bioactive contributor in amahewu, associated with biological properties (32).

Figure 3. Targeted differential profile during different stages of amahewu production (A) Phenolics, (B) Amino acids.
Avenonthiamides commonly referred to as Avenanthramides (Figure 3A) are phenolic amides primarily associated with oats but can also appear in maize through contamination or in sorghum via microbial activity (33). The malting sorghum introduces enzymes and LAB that can hydrolyze bound phenolics from cell walls (34) and conjugate them with amino acids. Consequently, the presence of anthranilic acid or its precursors from protein degradation in maize or sorghum, together with abundant phenolics such as p-coumaric acid and ferulic acid, can promote condensation reactions, ultimately leading to the formation of avenanthramide-like compounds.
Ferulic and caffeic acids, common hydroxycinnamic acids in maize and sorghum, were detected in moderate amounts. Both compounds are typically present in bound forms within the cell wall matrix, and their release depends on thermal softening and enzymatic hydrolysis during fermentation. For instance caffeic acid, a naturally occurring hydroxycinnamic acid widely distributed in cereals, fruits, and vegetables, contributed significantly to the discrimination between raw maize and cooked maize but not between the malted sorghum inoculum and the final products. Its presence at the early stages of processing aligns with reports that cooking enhances the release of bound phenolic acids from cell wall matrices (35). However, its absence in the final fermented products can be attributed to microbial metabolism. LAB and other fermentative microorganisms are known to decarboxylate, reduce, or conjugate caffeic acid into derivatives such as ferulic acid, dihydrocaffeic acid, and their glucosides, effectively reducing its detectable concentration in the end product (14, 36). The contrasting behavior of amino acids and phenolic acids in the discriminant profiles highlights the interplay between proteolysis and phenolic metabolism as central metabolic processes shaping the biochemical fingerprint of fermented maize beverages.
For quantification of the discriminant amino acids, proline exhibited the highest concentrations across all stages, particularly in RWM and CWM, where its levels exceeded 100 million units, making it the predominant amino acid in the system (Figure 3B). This finding aligns with literature that identifies proline as a major storage amino acid in cereals and as a crucial osmoprotectant during stress conditions (37). The sharp decline in proline content through successive fermentation stages suggests its utilization by fermentative microorganisms that often metabolize proline as an energy source to maintain redox balance driven by acidification and fermentation. This contributes to the development of characteristic sourdough-like flavors (38). During fermentation, proline also plays a protective function by stabilizing protein structures under stress conditions such as changes in pH and temperature (39). This explains its consistent appearance as a key discriminant metabolite differentiating malted sorghum inoculum from the final fermented maize product. Amino acids such as Aspartic Acid, Valine, Threonine, and Tyrosine demonstrated a noticeable increase in fermented samples, especially in FWM and FYM. Aspartic acid showed a significant elevation in fermented stages, which may reflect its role in microbial energy metabolism and as a precursor for other essential amino acids (40). Similarly, Valine and Threonine, both essential amino acids, were present in low concentrations initially but increased during fermentation, as available microorganisms consume sugars and convert them to other substrates like lactic acid, which in turn supported valine synthesis (41).
The IS displayed high levels of Alanine, Threonine, and Valine, in addition to considerable amounts of Proline, confirming its role as a source of free amino acids and enzymatic activity. The inclusion of malted sorghum not only contributes to starch hydrolysis but also enriches the amino acid pool, thereby supporting microbial activity and improving the sensory and nutritional profile of the beverage. The reduction in proline and the increase in essential amino acids such as Valine and Threonine improve the amino acid score of the beverage, enhancing its dietary protein quality. Furthermore, the liberation of amino acids during fermentation provides precursors for flavor compounds through Maillard and Strecker reactions, contributing to the characteristic taste of amahewu. This process may also facilitate the generation of bioactive peptides with antioxidant and antihypertensive properties, further positioning amahewu as a functional fermented cereal beverage.
3.4 Metabolite-metabolite correlation and pathway analysis.
Through correlation and pathway analyses, the identifiable metabolite markers can be linked to specific metabolic interactions and pathways, thereby offering mechanistic insights into the processes underlying the observed differences across production stages. Notably, metabolite–metabolite correlation analysis reveals the strength and direction of interactions, with positive correlations indicating coordinated increases or decreases, and negative correlations reflecting inverse concentration trends (42).
Strong positive correlations were observed between amino acids, namely serine, alanine, cysteine, proline, aspartic acid, threonine and valine and one phenolic compound as caffeic acid in white maize production (Figure 4A), whereas in yellow maize production, only amino acids had a strong positive correlation, namely proline, alanine, valine, threonine, aspartic acid and serine (Figure 4B). Correlation analysis proposes feed-forward and feed-back interaction of metabolites involved in the metabolic pathway. Amino acids in both productions exhibit a feed-forward interaction, where the presence of one amino acid promotes the production or accumulation of others, while phenolic compounds display a feedback interaction, resulting from the degradation of phenolic classes into derivatives that are often more bioactive than the parent compound (43).

Figure 4. Metabolite-metabolite correlation analysis heatmaps between (A) white and (B) yellow maize production.
To map these feedback interactions of metabolites in the biological pathways, the functional analysis revealed impacted pathways (Figure 5, Table 3). The leading pathway in white maize production is isoquinoline alkaloid biosynthesis, a pathway that begins with the conversion of tyrosine and phenylalanine; isoquinoline alkaloids form a family of phytochemicals or secondary metabolites that are found in several plants' precursors (44). While isoquinoline alkaloids are not typically considered primary nutrients, their presence or altered production during fermentation can lead to modification of bioactive compounds in the final product. These compounds are known for their pharmacological properties, including anti-inflammatory and antioxidant activities (45). Therefore, the presence of isoquinoline alkaloids might interact synergistically with other nutritional elements in amahewu, such as vitamins, minerals, or polyphenols, potentially enhancing the overall properties of the food product. This is evident when the addition of bambara flour protein source enhances the nutritional value of amahewu by improving important AA lysine content and minerals, as well as an improvement in the amino profile (46). Tyrosine metabolism is the second leading metabolic pathway, which is significant as an upstream precursor for synthesizing a series of valuable natural products, such as phenolic acids (47, 48). The fermentation process may lead to tyrosine breakdown into bioactive metabolites like p-hydroxy phenylacetate and homovanillic acid, which could increase the functional value of amahewu. Also, changes in tyrosine metabolism during the fermentation process could alter the concentration of free tyrosine in amahewu, affecting its amino acid profile (47).
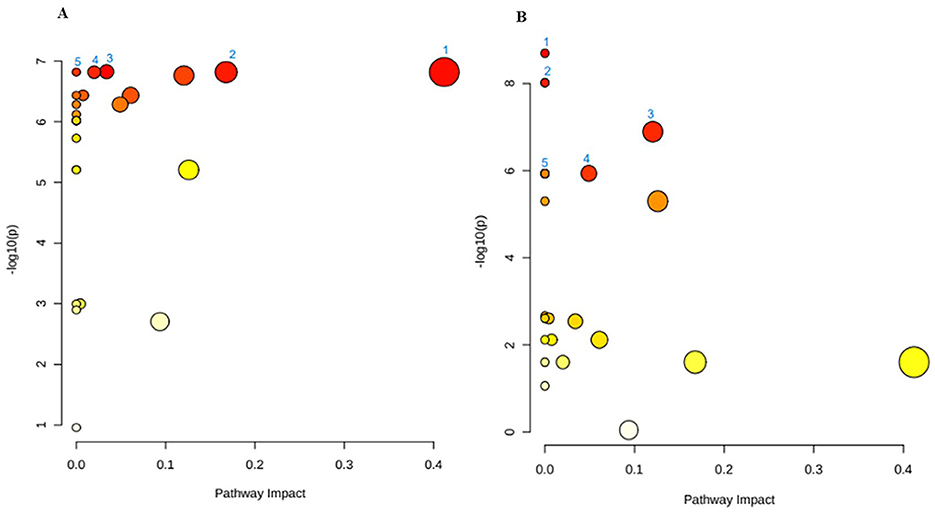
Figure 5. Metabolic pathway analysis of (A) white maize and (B) yellow maize production, with numbers (1–50) representing the main pathways and differential-difference of red = high and yellow = low. The graphs contain all the matched pathways (the metabolome) arranged by p-values (pathway enrichment analysis) on the y-axis, and the pathway impact values on the x-axis, further highlighted in Table 3 with the statistical p-values adjusted for multiple testing to account for numerous metabolic pathways that were all tested at the same time.

Table 3. Significant metabolic pathway altered in white and yellow maize production of amahewu from Metabolic Pathway Analysis (MetPA).
The findings from the production of amahewu using yellow maize and malted sorghum as inoculums indicate that amino acid metabolism was dominant. The metabolism of alanine, aspartate and glutamate, as well as glycine, serine and threonine (Gly-Ser-Thr), are the two leading impacted metabolic pathways. First, the amino acids were generated through enzymatic reactions with the help of present microorganisms, and then they were metabolized by the presumption of fermentation time, temperature and the application of inoculum. Alanine and aspartate can serve as substrates and precursors which are ultimately responsible for energy production and removing excess nitrogen from the body, while glutamate is widely known for its role as a flavor enhancer, contributing to the umami taste in foods (49). Similarly during the fermentation of amahewu from yellow maize, glutamate levels can increase, enhancing the sensory characteristics of the final product and leading to a more savory and appealing taste, making amahewu more enjoyable to consume. Additionally, it is speculated that antioxidants are produced through the Gly-Ser-Thr metabolic axis (50). Both Gly and Thr are critical components of glutathione, and since the considered factors in this study lead to their increase, amahewu could potentially have enhanced antioxidant properties, helping to combat oxidative stress and cellular damage.
4 Conclusion
This study revealed distinct metabolomic variations across processing stages and maize types during amahewu production, demonstrating that cooking, inoculation, and fermentation markedly reshape the beverage's biochemical composition. An increase in amino acids and phenolic content contributes enhanced the nutritional value of the beverage. The trend was consistent, throughout the study, as white maize showed steady metabolite increases, whereas yellow maize exhibited fluctuations of decreasing metabolites and lower fermentation efficiency. Thus, white maize–based amahewu showed more consistent and extensive metabolic remodeling than the yellow maize variant, reflecting higher fermentation efficiency, nutrient enrichment, and biochemical stability. Key metabolites such as valine, alanine, and apigenin emerged as reliable markers of fermentation progress. The chemometric analyses (PCA and OPLS-DA) confirmed clear separations among production stages, emphasizing the effect of fermentation on maize metabolism. Metabolite–metabolite correlation analysis revealed strong positive associations between amino acids and feedback-driven phenolic conversions. Pathway enrichment analysis identified isoquinoline alkaloid biosynthesis (in white maize) and alanine/aspartate/glutamate metabolism (in yellow maize) as central pathways shaping the metabolic outcomes. With further research, this approach could lead to improvements in the amahewu production process, ultimately resulting in a higher quality and more consistent final product.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
SK: Investigation, Formal analysis, Methodology, Writing – original draft, Data curation. AJ: Writing – original draft, Data curation, Writing – review & editing. FT: Methodology, Formal analysis, Data curation, Validation, Writing – review & editing, Resources, Supervision. AA: Resources, Validation, Data curation, Writing – review & editing, Methodology, Supervision. OA: Resources, Funding acquisition, Writing – review & editing, Supervision, Methodology, Conceptualization, Data curation, Validation.
Funding
The author(s) declared that financial support was received for this work and/or its publication. This work was financially supported by the Food and Beverage Manufacturing Sector Education and Training Authority (FoodBev SETA) Research and Innovation Masters' Bursary, South Africa, which was awarded to the main author (Kgoale SDP). Funding from the National Research Foundation of South Africa Thuthuka Grant (Grant number: 121826), the University of Johannesburg (UJ) Faculty of Science, and the UJ Research Committee (URC) Grant awarded to Adebo OA are duly acknowledged.
Conflict of interest
Author Fidele Tugizimana was employed by Omnia Group, Ltd.
The remaining author(s) declared that this work was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author FT declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declared that generative AI was not used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fnut.2025.1697672/full#supplementary-material
References
1. Kaushal M, Sharma R, Vaidya D, Gupta A, Saini HK, Anand A, et al. Maize: an underexploited golden cereal crop. Cereal Res Commun. (2022) 51:3–14. doi: 10.1007/s42976-022-00280-3
2. Chaudhary DP, Kumar S, Langyan S. Maize: nutrition dynamics and novel uses. Berlin: Springer (2014). doi: 10.1007/978-81-322-1623-0
3. Chaves-López C, Rossi C, Maggio F, Paparella A, Serio A. Changes occurring in spontaneous maize fermentation: an overview. Fermentation. (2020) 6:36. doi: 10.3390/fermentation6010036
4. Kaul J, Jain K, Olakh D. An overview on role of yellow maize in food, feed and nutrition security. Int J Curr Microbiol Appl Sci. (2019) 8:3037–48. doi: 10.20546/ijcmas.2019.802.356
5. Ranum P, Pena-Rosas JP, Garcia-Casal MN. Global maize production, utilization, and consumption. Ann N Y Acad Sci. (2014) 1312:105–12. doi: 10.1111/nyas.12396
6. Chelule PK, Mbongwa HP, Carries S, Gqaleni N. Lactic acid fermentation improves the quality of amahewu, a traditional South African maize-based porridge. Food Chem. (2010) 122:656–61. doi: 10.1016/j.foodchem.2010.03.026
7. Blandino A, Al-Aseeri ME, Pandiella SS, Cantero D, Webb C. Cereal-based fermented foods and beverages. Food Res Int. (2003) 36:527–43. doi: 10.1016/S0963-9969(03)00009-7
8. Puerari C, Magalhaes-Guedes KT, Schwan RF. Physicochemical and microbiological characterization of chicha, a rice-based fermented beverage produced by Umutina Brazilian Amerindians. Food Microbiol. (2015) 46:210–7. doi: 10.1016/j.fm.2014.08.009
9. Daji GA, Green E, Adebo OA. Nutritional and phytochemical composition of Mahewu (a Southern African Fermented Food Product) derived from white and yellow maize (Zea mays) with different inocula. Fermentation. (2023) 9:58. doi: 10.3390/fermentation9010058
10. Maakelo PK, Bultosa G, Kobue-Lekalake RI, Gwamba J, Sonno K. Effects of watermelon pulp fortification on maize mageu physicochemical and sensory acceptability. Heliyon. (2021) 7:e07128. doi: 10.1016/j.heliyon.2021.e07128
11. Awobusuyi TD, Siwela M, Kolanisi U, Amonsou EO. Provitamin A retention and sensory acceptability of amahewu, a non-alcoholic cereal-based beverage made with provitamin A-biofortified maize. J Sci Food Agric. (2016) 96:1356–61. doi: 10.1002/jsfa.7230
12. Oyeyinka AT, Siwela M, Pillay K. A mini review of the physicochemical properties of amahewu, a Southern African traditional fermented cereal grain beverage. LWT. (2021) 151:112159. doi: 10.1016/j.lwt.2021.112159
13. Singh N, Barthwal R, Negi A, Aggarwal S, Kathuria D, Kumar V, et al. Foodomics: futuristic omic strategies to assess the impact of food nutrients on human health and gut microbiome. Int J Food Sci Technol. (2024) 59:4194–212. doi: 10.1111/ijfs.17041
14. Adebo OA. African sorghum-based fermented foods: past, current and future prospects. Nutrients. (2020) 12:1111. doi: 10.3390/nu12041111
15. Daji GA, Green E, Akanni GB, Adebo OA. GC–MS-based metabolomics elucidates the impact of cereal-based microbial starters on the metabolite profile of mahewu prepared from yellow maize. Int J Food Sci Technol. (2023) 59:4268–80. doi: 10.1111/ijfs.16667
16. Pascale R, Onzo A, Ciriello R, Scrano L, Bufo SA, Bianco G. LC/MS based food metabolomics. In: Comprehensive Foodomics. Amsterdam: Elsevier (2021). p. 39–53. doi: 10.1016/B978-0-08-100596-5.22774-1
17. Cui Y, Du K, Hou S, Yang R, Qi L, Li J, et al. A comprehensive strategy integrating metabolomics with multiple chemometric for discovery of function related active markers for assessment of foodstuffs: a case of hawthorn (Crataegus cuneata) fruits. Food Chem. (2022) 383:132464. doi: 10.1016/j.foodchem.2022.132464
18. Cocuron JC, Koubaa M, Kimmelfield R, Ross Z, Alonso AP. A Combined Metabolomics and fluxomics analysis identifies steps limiting oil synthesis in maize embryos. Plant Physiol. (2019) 181:961–75. doi: 10.1104/pp.19.00920
19. Carballo-Meilan A, McDonald L, Pragot W, Starnawski LM, Saleemi AN, Afzal W. Development of a data-driven scientific methodology: from articles to chemometric data products. Chemom Intell Lab Syst. (2022) 225:104555. doi: 10.1016/j.chemolab.2022.104555
20. Li D, Peng J, Kwok L-y, Zhang W, Sun T. Metabolomic analysis of Streptococcus thermophilus S10-fermented milk. LWT. (2022) 161:113368. doi: 10.1016/j.lwt.2022.113368
21. Zhao L, Zhao X, Xu Y, Liu X, Zhang J, He Z. Simultaneous determination of 49 amino acids, B vitamins, flavonoids, and phenolic acids in commonly consumed vegetables by ultra-performance liquid chromatography-tandem mass spectrometry. Food Chem. (2021) 344:128712. doi: 10.1016/j.foodchem.2020.128712
22. Sarker U, Oba S. Salinity stress enhances color parameters, bioactive leaf pigments, vitamins, polyphenols, flavonoids and antioxidant activity in selected Amaranthus leafy vegetables. J Sci Food Agric. (2019) 99:2275–84. doi: 10.1002/jsfa.9423
23. Tugizimana F, Djami-Tchatchou AT, Steenkamp PA, Piater LA, Dubery IA. Metabolomic analysis of defense-related reprogramming in sorghum bicolor in response to colletotrichum sublineolum infection reveals a functional metabolic web of phenylpropanoid and flavonoid pathways. Front Plant Sci. (2018) 9:1840. doi: 10.3389/fpls.2018.01840
24. Bello-Pérez LA, Flores-Silva PC, Sifuentes-Nieves I, Agama-Acevedo E. Controlling starch digestibility and glycaemic response in maize-based foods. J Cereal Sci. (2021) 99:103222. doi: 10.1016/j.jcs.2021.103222
25. Dabija A, Ciocan ME, Chetrariu A, Codină GG. Maize and sorghum as raw materials for brewing, a review. Appl Sci. (2021) 11:3139. doi: 10.3390/app11073139
26. Nkhata SG, Ayua E, Kamau EH, Shingiro JB. Fermentation and germination improve nutritional value of cereals and legumes through activation of endogenous enzymes. Food Sci Nutr. (2018) 6:2446–58. doi: 10.1002/fsn3.846
27. Thaoge M, Adams M, Sibara M, Watson T, Taylor J, Goyvaerts E. Production of improved infant porridges from pearl millet using a lactic acid fermentation step and addition of sorghum malt to reduce viscosity of porridges with high protein, energy and solids (30%) content. World J Microbiol Biotechnol. (2003) 19:305–10.
28. Daji GA, Green E, Abrahams A, Oyedeji AB, Masenya K, Kondiah K, et al. Physicochemical properties and bacterial community profiling of optimal mahewu (a fermented food product) prepared using white and yellow maize with different inocula. Foods. (2022) 11:3171. doi: 10.3390/foods11203171
29. Cui L, Li Dj, Liu CQ. Effect of fermentation on the nutritive value of maize. Int J Food Sci Technol. (2012) 47:755–60. doi: 10.1111/j.1365-2621.2011.02904.x
30. Adebo OA, Oyedeji AB, Adebiyi JA, Chinma CE, Oyeyinka SA, Olatunde OO, et al. Kinetics of phenolic compounds modification during maize flour fermentation. Molecules. (2021) 26:6702. doi: 10.3390/molecules26216702
31. Paucar-Menacho LM, Castillo-Martinez WE, Simpalo-Lopez WD, Verona-Ruiz A, Lavado-Cruz A, Martinez-Villaluenga C, et al. Performance of thermoplastic extrusion, germination, fermentation, and hydrolysis techniques on phenolic compounds in cereals and pseudocereals. Foods. (2022) 11:1957. doi: 10.3390/foods11131957
32. Hostetler GL, Ralston RA, Schwartz SJ. Flavones: food sources, bioavailability, metabolism, and bioactivity. Adv Nutr. (2017) 8:423–35. doi: 10.3945/an.116.012948
33. Xie X, Lin M, Xiao G, Liu H, Wang F, Liu D, et al. Phenolic amides (avenanthramides) in oats–an update review. Bioengineered. (2024) 15:2305029. doi: 10.1080/21655979.2024.2305029
34. Mbock E, Emmambux M, Duodu KG. Effect of malting and fermentation on the functional and health-promoting properties of red and white sorghum cultivars. Pretoria: University of Pretoria (2025).
35. Toydemir G, Gultekin Subasi B, Hall RD, Beekwilder J, Boyacioglu D, Capanoglu E. Effect of food processing on antioxidants, their bioavailability and potential relevance to human health. Food Chem X. (2022) 14:100334. doi: 10.1016/j.fochx.2022.100334
36. Svensson L, Sekwati-Monang B, Lutz DL, Schieber A, Ganzle MG. Phenolic acids and flavonoids in nonfermented and fermented red sorghum (Sorghum bicolor (L.) Moench). J Agric Food Chem. (2010) 58:9214–20. doi: 10.1021/jf101504v
37. Khan P, Abdelbacki AMM, Albaqami M, Jan R, Kim KM. Proline promotes drought tolerance in maize. Biology. (2025) 14:41. doi: 10.3390/biology14010041
38. Gobbetti M, De Angelis M, Di Cagno R, Calasso M, Archetti G, Rizzello CG. Novel insights on the functional/nutritional features of the sourdough fermentation. Int J Food Microbiol. (2019) 302:103–13. doi: 10.1016/j.ijfoodmicro.2018.05.018
39. Takagi H, Taguchi J, Kaino T. Proline accumulation protects Saccharomyces cerevisiae cells in stationary phase from ethanol stress by reducing reactive oxygen species levels. Yeast. (2016) 33:355–63. doi: 10.1002/yea.3154
40. Azevedo Rd, Lancien M, Lea P. The aspartic acid metabolic pathway, an exciting and essential pathway in plants. Amino acids. (2006) 30:143-62. doi: 10.1007/s00726-005-0245-2
41. Blombach B, Arndt A, Auchter M, Eikmanns BJ. L-Valine production during growth of pyruvate dehydrogenase complex-deficient Corynebacterium glutamicum in the presence of ethanol or by inactivation of the transcriptional regulator SugR. Appl Environ Microbiol. (2009) 75:1197–200. doi: 10.1128/AEM.02351-08
42. Rao G, Sui J, Zhang J. Metabolomics reveals significant variations in metabolites and correlations regarding the maturation of walnuts (Juglans regia L.). Biol Open. (2016) 5:829–36. doi: 10.1242/bio.017863
43. Leonard W, Zhang P, Ying D, Adhikari B, Fang Z. Fermentation transforms the phenolic profiles and bioactivities of plant-based foods. Biotechnol Adv. (2021) 49:107763. doi: 10.1016/j.biotechadv.2021.107763
44. Khan AY, Suresh Kumar G. Natural isoquinoline alkaloids: binding aspects to functional proteins, serum albumins, hemoglobin, and lysozyme. Biophys Rev. (2015) 7:407–20. doi: 10.1007/s12551-015-0183-5
45. Aryal B, Raut BK, Bhattarai S, Bhandari S, Tandan P, Gyawali K, et al. Potential therapeutic applications of plant-derived alkaloids against inflammatory and neurodegenerative diseases. Evid Based Complement Alternat Med. (2022) 2022:7299778. doi: 10.1155/2022/7299778
46. Awobusuyi TD, Siwela M. Nutritional properties and consumer's acceptance of provitamin A-biofortified amahewu combined with bambara (Vigna Subterranea) flour. Nutrients. (2019) 11:1476. doi: 10.3390/nu11071476
47. Li G, Chen Z, Chen N, Xu Q. Enhancing the efficiency of L-tyrosine by repeated batch fermentation. Bioengineered. (2020) 11:852–61. doi: 10.1080/21655979.2020.1804177
48. Li Y, Mao J, Song X, Wu Y, Cai M, Wang H, et al. Optimization of the l-tyrosine metabolic pathway in saccharomyces cerevisiae by analyzing p-coumaric acid production. 3 Biotech. (2020) 10:258. doi: 10.1007/s13205-020-02223-3
49. Yamamoto T, Inui-Yamamoto C. The flavor-enhancing action of glutamate and its mechanism involving the notion of kokumi. NPJ Sci Food. (2023) 7:3. doi: 10.1038/s41538-023-00178-2
Keywords: metabolomics, metabolites, targeted, phenolics, amino acids
Citation: Kgoale SD, Jimoh AA, Tugizimana F, Abrahams A and Adebo OA (2026) Targeted metabolomics to quantitatively profile changes in amino acids and phenolics at every step of amahewu production from two Zea Mays L. maize types (white and yellow). Front. Nutr. 12:1697672. doi: 10.3389/fnut.2025.1697672
Received: 02 September 2025; Revised: 21 November 2025;
Accepted: 08 December 2025; Published: 12 January 2026.
Edited by:
Gabriel Cardoso-Ugarte, Popular Autonomous University of the State of Puebla, MexicoReviewed by:
Myriam Lamine, Center of Biotechnology of Borj Cedria (CBBC), TunisiaRifqi Ahmad Riyanto, Sultan Ageng Tirtayasa University, Indonesia
Remziye Yilmaz, Hacettepe University, Türkiye
Copyright © 2026 Kgoale, Jimoh, Tugizimana, Abrahams and Adebo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Oluwafemi Ayodeji Adebo, b2FkZWJvQHVqLmFjLnph; b2FhZGVib0BnbWFpbC5jb20=