 Priyanka Mishra
Priyanka Mishra Kishore C. Panigrahi
Kishore C. Panigrahi- Plant Science Lab, School of Biological Sciences, National Institute of Science Education and Research, Bhubaneshwar, India
GIGANTEA (GI) is a plant specific nuclear protein and functions in diverse physiological processes such as flowering time regulation, light signaling, hypocotyl elongation, control of circadian rhythm, sucrose signaling, starch accumulation, chlorophyll accumulation, transpiration, herbicide tolerance, cold tolerance, drought tolerance, and miRNA processing. It has been five decades since its discovery but the biochemical function of GI and its different domains are still unclear. Although it is known that both GI transcript and GI protein are clock controlled, the regulation of its abundance and functions at the molecular level are still some of the unexplored areas of intensive research. Since GI has many important pleotropic functions as described above scattered through literature, it is worthwhile and about time to encapsulate the available information in a concise review. Therefore, in this review, we are making an attempt to summarize (i) the various interconnected roles that GI possibly plays in the fine-tuning of plant development, and (ii) the known mutations of GI that have been instrumental in understanding its role in distinct physiological processes.
Introduction
GIGANTEA (GI), the unique plant specific nuclear protein, although identified way back (Rédei, 1962) as a late flowering mutant (gi) in Arabidopsis thaliana (At), its precise biochemical roles are far from being understood (de Montaigu et al., 2010). The genomic organization of GI was evident after it was fine-mapped to chromosome 1 and subsequently, the GI cDNA was isolated (Fowler et al., 1999). The genomic locus of GI of At consists of 14 exons and encodes for a protein of 1173 amino acids (Fowler et al., 1999; Park et al., 1999). GI expression is ubiquitous and is detected throughout various stages of plant development indicative of its involvement in several functions summarized in Figure 1 (Fowler et al., 1999; Park et al., 1999). It is interesting to note the ubiquitous expression of GI that reflect upon its pleiotropic roles in multitude of responses ranging from breaking of seed dormancy, hypocotyl elongation, initiating the circadian rhythm in seeds to the setting of fruits in the adult plant. Many of the above listed responses integrate information from the light input and external temperature, making it an interesting but complicated area of plant science.
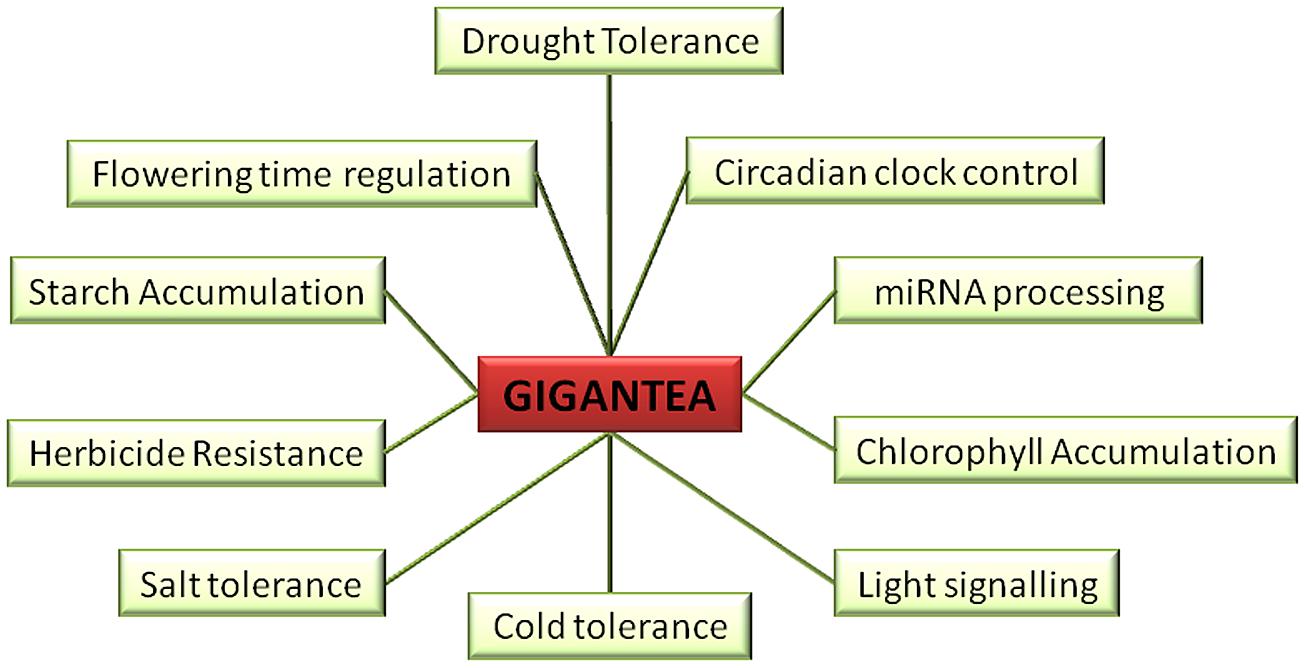
FIGURE 1. Multiple roles of GIGANTEA (GI). GI is known to play role in drought tolerance, circadian clock control, miRNA processing, chlorophyll accumulation, light signaling, cold tolerance, salt tolerance, herbicide resistance, starch accumulation, and flowering time regulation.
Experiments aimed at understanding the abundance of the transcript and the protein are typically carried out in controlled cabinets, where the subjective time of the diurnal cycle are referred as the Zeitgeber time (ZT). Both the GI transcript and GI protein are under the control of diurnal regulation. Under long day (LD) growth cycle of 16 h light and 8 h dark (16 hL/8 hD), the GI mRNA peaks at ZT 10 and shows a trough at ZT 0, while under short day (SD) cycle of 8 hL/16 hD, GI transcript level peaks at ZT 8 (Fowler et al., 1999). The GI protein abundance also follows a similar pattern to its transcript accumulation (David et al., 2006). The regulation of GI is important for the control of circadian clock and several genes such as FLAVIN-BINDING, KELCH REPEAT, F BOX 1 (FKF1), a blue light photoreceptor, and CYCLING DOF FACTORs (CDFs), which are involved in the transcription of a flowering time regulator CONSTANS (CO; Fornara et al., 2009). In addition, the diurnal regulation of the protein might also play an important role in the diurnal control of stomatal opening (Ando et al., 2013).
In order to assign a function to GI, it was of interest to enumerate its precise sub-cellular localization. Therefore, N-terminal GFP fusion of GI was constructed and transiently transfected in onion epidermal cells. The fluorescence microscopy of the fusion protein for the first time demonstrated that GI is predominantly localized to the nuclei and forms nuclear bodies (Huq et al., 2000). Later, the GI protein was also found to be localized in the nucleus of different cell types of transgenic At plants over-expressing GI:GFP (Mizoguchi et al., 2005). Four clusters of basic amino acids resembling the nuclear localization signal (NLS) in the GI sequence explained its nuclear abundance (Huq et al., 2000). GI has been shown to form nuclear bodies of diverse numbers, size, and shape (Kim et al., 2013c). To understand the molecular composition of GI nuclear bodies, attempts were made to evaluate the co-localization of GI with marker proteins of known sub-nuclear compartments such as heterochromatin bundles, nucleoli, spliceosome, and Cajal bodies. This piece of work demonstrated that GI did not localize to any of the above known nuclear compartments (Kim et al., 2013c). This suggested that GI might not have role in processes such as biogenesis of rRNA and snRNP, pre-mRNA splicing, and protein degradation. Since these co-localization studies were carried out in Arabidopsis mesophyll protoplasts using a transient over-expression method, it does not mimic the exact physiological environment. Furthermore, the association and dissociation rate of proteins to nuclear bodies has been shown to be affected by specific post-translational modifications. The spatio-temporal mis-localization of proteins can also affect its post-translational modifications. With so many complexities involved, stable transgenic lines expressing fluorescent tagged marker proteins and GI under their native promoters would be an impressive feat to achieve in order to understand the molecular composition of the GI complexes. Understanding the molecular composition of GI nuclear bodies (NBs) at different diurnal time-points would be a valuable asset.
The formation of GI nuclear bodies is light dependent since, the sequestration of GI into NBs is facilitated by EARLY FLOWERING 4 (ELF4) during the day, thus inhibiting the CO transcription. Likewise, EARLY FLOWERING 3 (ELF3) promotes the interaction of GI and CONSTITUTIVE PHOTOMORPHOGENESIS 1 (COP1) to form NBs which degrade GI in planta (Yu et al., 2008). The dynamic association of GI with heterogenous nuclear bodies during the light to dark transition needs to be evaluated. In other words, the question still remains, if GI associates and dissociates in a light dependent manner on a core complex within the nuclei based on its differential post-translational modification status.
Although studies showed the presence of GI predominantly in the nuclei, in silico analysis predicted the presence of 11 transmembrane domains in AtGI which argues in favor of a possible membrane localization (Fowler et al., 1999). Furthermore, membrane localized GI possibly has a role in the regulation of ion channels during salt stress and stomatal opening as seen in phototropins (Stoelzle et al., 2003). Purified recombinant GI from Escherichia coli when subjected to electron microscopic study, revealed a tetrameric arrangement in vitro. However, its quaternary structure in vivo is still unclear (Black et al., 2011). This multimeric organization of a protein would not only offer more epitopes for interactions with diverse regulators but also would offer additional layers of control on its stability.
Alleles of GIGANTEA with Distinct Phenotypes
The gi mutants were described as late flowering mutants for the first time (Rédei, 1962). There are several gi mutants described in literature such as gi-1, gi-2, gi-3, gi-4, gi-5, gi-6, gi-11, gi-12, gi-100, gi-200, gi-201, gi-596, and gi-611 (summarized in Table 1; Figure 2). Some of the gi mutants were shown to influence the activity of the circadian clock, while others alter diverse responses (Park et al., 1999). The gi-1 allele, lacking the C-terminal part of GI, was responsible for shortening the period of the clock, while the gi-2 allele, lacking both the C-terminal and the central region of GI, lengthened the period. While the gi-1 mutation shortened the period of CAB2 expression, the gi-2 mutation lengthened the period of CAB2 expression (Park et al., 1999). This suggests that the central region of the protein or the terminal half of the protein most probably fine-tunes the period length of the circadian clock.
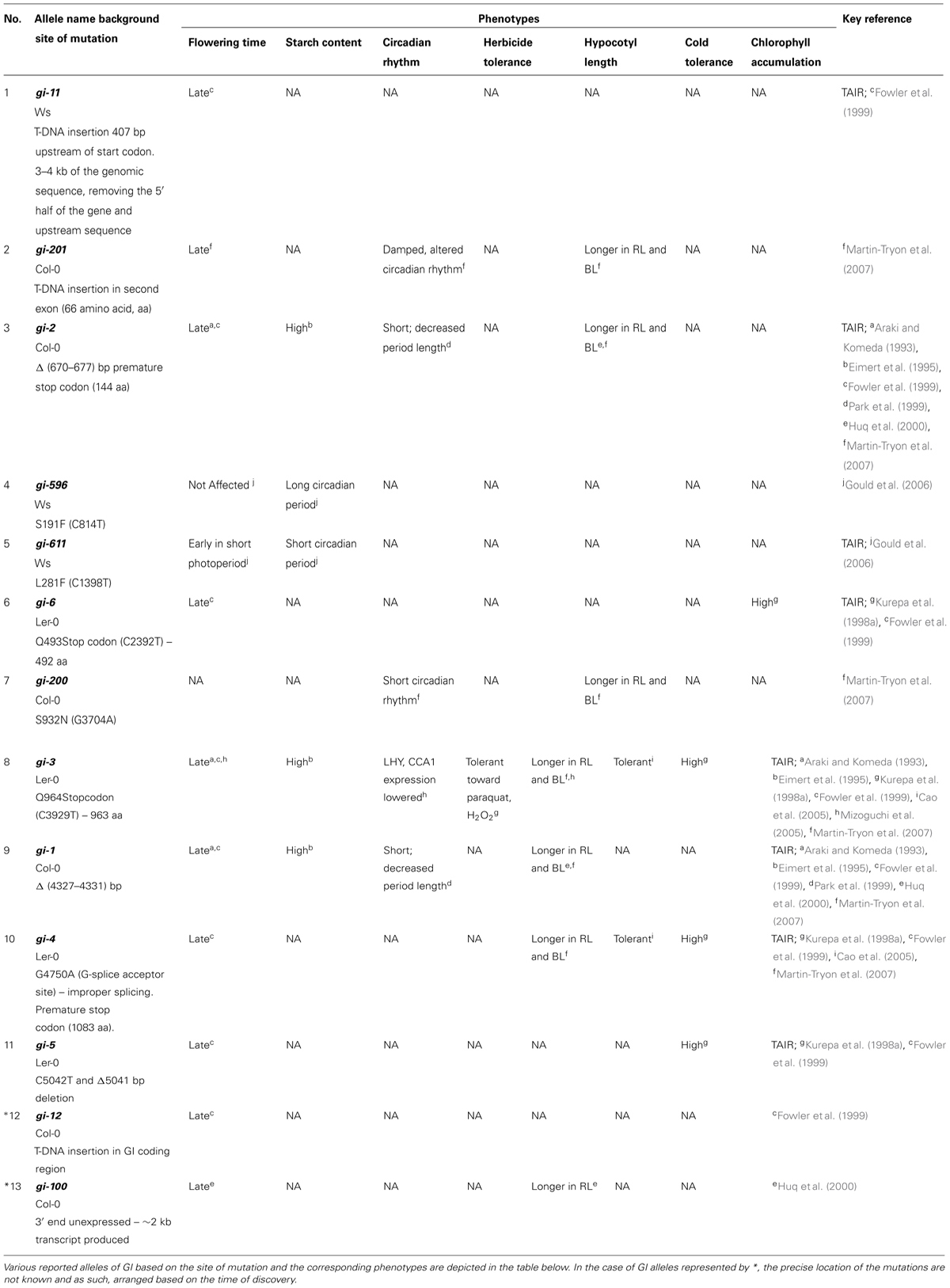
TABLE 1. List of known GI alleles and their phenotypes.
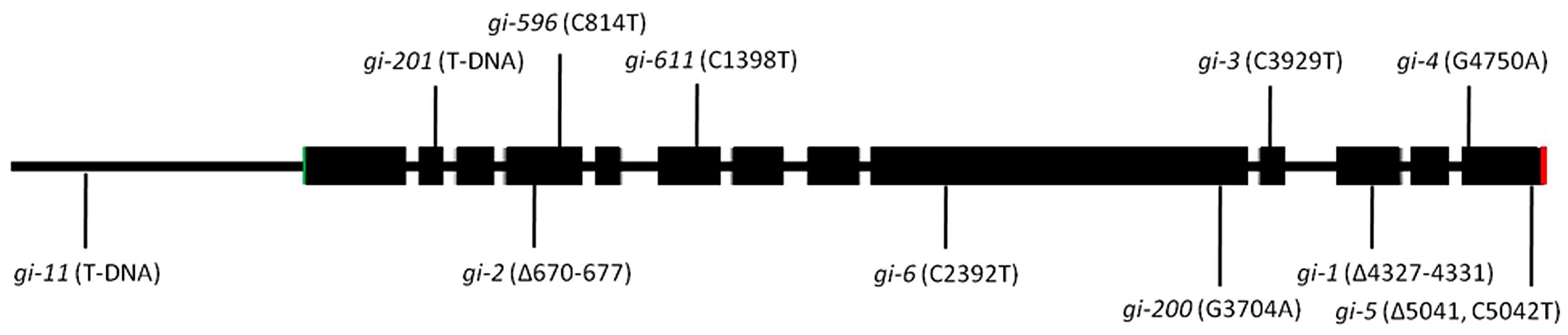
FIGURE 2. The schematic presentation of the mutations in the GI locus. The relative position of the available mutations in the genomic sequence of GI has been depicted. The start codon is indicated by green color while the stop position is represented by red. Coding regions are represented in black boxes while the non-coding regions are represented as lines.
The gi-2 mutant at higher temperature of about 28°C showed longer hypocotyl and flowered earlier in comparison to the plants grown at temperatures of 18 and 22°C (Araki and Komeda, 1993). Even though higher temperature were shown to regulate flowering (at 18, 22, 28°C) and hypocotyl elongation (at 22, 28°C) in gi-2 mutant, it was almost equally sensitive toward vernalization as in WT. Vernalization is the exposure of plants to prolonged cold temperature that leads to earlier flowering cue in Arabidopsis. This implies that probably GI regulates flowering using a vernalization-independent pathway (Martinez-Zapater and Somerville, 1990; Koornneef et al., 1991; Araki and Komeda, 1993).
The alleles of GI are the result of random mutagenesis or T-DNA insertion which have aided in understanding its various functions. Alleles such as gi-1, gi-2, gi-3, and gi-6 introduce premature stop codon whereas gi-4 and gi-5 most probably alter the C-terminus of the protein due to frame-shift mutations (Fowler et al., 1999). No GI expression was detected in the gi-11 and gi-201 alleles carrying T-DNA insertion (Richardson et al., 1998; Martin-Tryon et al., 2007). The gi-100 mutation, originally identified in a red light screen, also contained a T-DNA insertion, but produced a truncated transcript of about 2 kb due to the absence of the 3′ end of GI (Huq et al., 2000). The transcript level in gi-1, gi-2, and gi-3 is lower compared to that of gi-4, gi-5, gi-6, and gi-100, which show similar or higher levels compared to their respective WT (Fowler et al., 1999; Huq et al., 2000). The role of GI in blue light dependent hypocotyl elongation was revealed using the gi-200 allele, consisting of a substitution of the serine 932 (Martin-Tryon et al., 2007).
Various deletions in GI sequences and its phenotypes are summarized in Table 1. After analyzing the data depicted in Table 1, it is evident that any deletion in GI mostly causes defects in the flowering time, circadian clock, and control of hypocotyl elongation. In the gi-4 mutant, improper splicing leads to a loss of 90 amino acids from the C-terminus causing late flowering. This deletion also causes the over-expression of its own transcript suggesting that the C-terminal 90 amino acids are required for its auto-regulation and flowering time (Fowler et al., 1999). The abundance of the gi-4 transcript could be due to increased stability or decreased decay which needs to be verified. Since GI stimulates CO transcription, this C-terminal domain of GI might be acting as an enhancer of CO transcription or involved in the recruitment of activators to the CO promoter.
The seeds of Wassilewskija (Ws) ecotype expressing CAB:LUC were mutagenized and screened for altered period length. Two novel alleles gi-596 and gi-611 were identified in this screen (Gould et al., 2006). In the gi-596 allele, mutation caused by the substitution of the serine residue at 191 position to phenylalanine (S191F) did not affect the flowering-time although the period length of the circadian clock is lengthened and longer hypocotyl was observed under both red and blue light conditions. This suggests that the serine 191 residue might have an important role in photoreceptor signaling. On the contrary, the mutation in gi-611 allele was mapped to the lysine 281. This allele showed significantly early flowering in SDs suggesting that this lysine in WT is involved in decelerating the flowering time (Gould et al., 2006). Since the Ws ecotype is a natural null for the high light sensor Phytochrome D, the phenotype observed could be a combinatorial effect of the lack of this photoreceptor and the respective mutations in GI allele (Aukerman et al., 1997). It would be interesting to evaluate if these alleles in Col background would show the similar light dependent effect to rule out the involvement and interaction of PHYD in this process. Both the positions, Lys281 and Ser191 are conserved in the Col-0 and Ler-0 ecotypes and thus, the role of these residues could be confirmed by the expression of the respective GI alleles containing the substitutions in these ecotypes to determine the importance of these mutated residues.
Transcriptional Regulation of GIGANTEA
Defects in the circadian clock components have been found to affect the GI transcription. CIRCADIAN CLOCK ASSOCIATED 1 (CCA1), a core component of the circadian clock, reduces the GI expression by binding to CCA1-binding site on GI promoter (Lu et al., 2012). GI transcript, thus accumulates toward the middle of the day, when CCA1 expression is repressed by TIMING OF CAB EXPRESSION 1 (TOC1). The rhythmicity of GI transcript level is lost in elf3 mutant in continuous light (LL) suggesting that ELF3 might also regulate the GI mRNA abundance (Fowler et al., 1999). Since CCA1 and ELF3 have been proposed to physically interact to control flowering time and hypocotyl elongation, it would be interesting to investigate the coordinated involvement of these two proteins in the regulation of GI transcription. Clock proteins, such as, LIGHT-REGULATED WD 1 and 2 (LWD1 and LWD2) also affect the GI expression pattern, since in lwd1lwd2 double mutant GI transcript is most abundant at ZT 6 instead of ZT 10 (Wu et al., 2008). The two proteins being very similar (∼90% identity) possess functional redundancy, evident from single mutants being phenotypically similar to WT. Another clock associated gene, TIME FOR COFFEE (TIC) is also known to regulate the rhythmicity of GI in Arabidopsis. In tic mutants, GI transcript level is lower and the peak is shifted ∼4 h earlier than in WT plants (Hall et al., 2003). Since in both the lwd1lwd2 and tic mutants the GI expression is shifted to ZT6, it suggests that the activities of both the proteins might be required for the repression of the GI transcription in the morning. pseudo-response regulators (PRRs), namely, PRR5, PRR7, and PRR9 also have been proposed to regulate GI expression and therefore, flowering time via the CO-FT module (Nakamichi et al., 2007; Kawamura et al., 2008). Epistatic analysis and mutant combinations between LWD1/2, PRRs, and TIC would be beneficial to explain the additive roles of these genes products and the genetic hierarchy of the genes regulating the inhibition of GI expression. The expression of GI at the wrong time of the diurnal cycle is known to cause flowering time defects in At (Fornara et al., 2009). These mutants might behave as late flowering due to the untimely expression of GI. Although a lot is known from the transcript analysis, the work at the protein level is far from being understood due to the unavailability of a GI anti-serum that could detect the endogenous GI protein. The detailed post-translational regulation of GI is explained in the Section “Post-Translational Regulation of GIGANTEA.”
Several studies have demonstrated that light quality and quantity influence GI transcription, although systematic studies involving changes in the light fluence and wavelength to evaluate GI expression is yet to be carried out. In Arabidopsis, upon transition to night, GI mRNA level decreases with a half-life of about 1 h irrespective of the photoperiod (Fowler et al., 1999). A significant light dependent down-regulation is also detected in the legume Medicago truncatula suggesting a similar mechanism might coordinate light sensing with transcriptional activity (Paltiel et al., 2006). GI mRNA accumulation pattern in both Arabidopsis and M. truncatula showed a secondary peak at ZT 2 under SDs as well as LDs (Paltiel et al., 2006). This peak could be the result of an acute response to light. A similar peak of GI mRNA at ZT 2 has also been documented in plants grown under blue light. The role of blue light in the regulation of this early secondary peak of GI needs to be thoroughly examined using mutants that are affected in blue light signaling. This would clarify if the peak at ZT 2 is due to photoreceptors or secondary signaling components involved in blue light signaling. The peak expression of GI is delayed by approximately 4 h in plants grown in low red:far-red (R:FR) light conditions in comparison to plants grown in white light condition (Wollenberg et al., 2008). This indicated that the photoreceptors and their activity are fine-tuning the timing and quantity of the GI transcript. The accumulation of the GI protein in the morning around ZT 3–4 and its consequence in plant development has not been studied yet, that needs to be evaluated in depth.
Besides light, temperature too has been found to regulate GI expression. Warmer temperature of 28°C up-regulates GI mRNA level as compared to the cooler temperatures of 12°C at dawn (Paltiel et al., 2006). The night time repression of GI transcription was shown to be temperature dependent and regulated by evening complex (EC) night time repressor constituted of ELF3, ELF4, and LUX ARRHYTHMO (LUX; Nusinow et al., 2011; Mizuno et al., 2014). The EC night time repressor was revealed to bind to the GI promoter through LUX binding site (LBS).
GIGANTEA has been proposed to regulate its own expression, since the mutants, gi-1 and gi-2, have lower expression of the GI alleles, approximately 40 and 20% of the WT transcript, respectively (Fowler et al., 1999; Park et al., 1999). But this auto-regulatory role of GI transcription is contradictory, since, gi-4 and gi-6 lines show ∼30% higher expression of GI compared to its WT (Fowler et al., 1999). This effect could be either due to the difference in the ecotypes or differential regulation of the transcript stability. Another question worth investigating would be the abundance of the mutant proteins produced in each mutant, which would require a functional GI antiserum. The positive or negative auto-regulatory role suggests that mutations at different residues in the coding sequence can influence the abundance of transcriptional enhancers or repressors, affecting GI expression.
Post-Translational Regulation of GIGANTEA
Over-expression of GI leads to the constitutive accumulation of the GI transcript throughout the photoperiod. Despite its constant expression level, GI protein follows a cyclic pattern of accumulation in both LDs and SDs. This is suggestive of the degradation of the GI protein (David et al., 2006). GI was found to be ubiquitinated upon dusk, a pre-requisite for its degradation via the 26S proteasome mechanism (David et al., 2006). In the dark phase, nuclear GI abundance has been shown to be regulated by the E3 ubiquitin ligase activity of COP1 and ELF3 (Yu et al., 2008). The interaction between COP1 and GI is ELF3 dependent, where ELF3 serves as an adaptor protein (Yu et al., 2008). The shuttling of COP1 between the nucleus and the cytoplasm is regulated by light (von Arnim and Deng, 1994). COP1 being nuclear localized in the night phase makes it competent for COP1–ELF3 mediated degradation of GI through 26S proteasome.
Upon heat shock GI is SUMOylated (López-Torrejón et al., 2013). It has been proposed that SUMOylation prevents the degradation of GI, thus enhancing its abundance. GI accumulation has been correlated with earlier flowering under heat stress. The identification of SUMOylation and ubiquitination sites in GI that alter its stability and degradation could be of pivotal importance in manipulating flowering time of crop plants. Current knowledge on the transcriptional and post-translational regulation of GI is presented schematically in Figure 3.
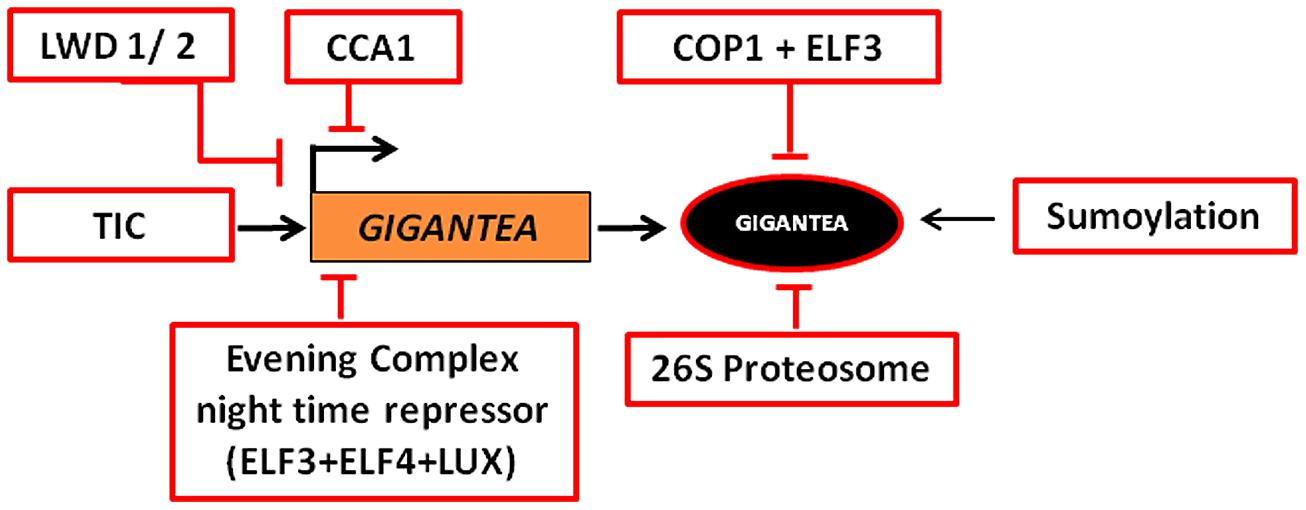
FIGURE 3. Transcriptional and post-translational regulation of GI. The expression of GI is regulated by LIGHT-REGULATED WD1/2 (LWD1/2), TIME FOR COFFEE (TIC), and CIRCADIAN CLOCK ASSOCIATED 1 (CCA1) in the day and EVENING COMPLEX (EC) night time repressor during night, such that the transcript peaks at around ZT 10 in LD and ZT 8 in SD. In the dark phase, 26S Proteasome, CONSTITUTIVE PHOTOMORPHOGENESIS 1 (COP1), and EARLY FLOWERING 3 (ELF3) regulate the abundance of the protein. SUMOylation of GI stabilizes the protein.
Roles of GIGANTEA
GIGANTEA plays multiple roles throughout plant development. Its functions in processes such as light signaling, circadian clock regulation, flowering time control, chlorophyll accumulation, sugar metabolism, and stress tolerance have been discussed below.
Light Signaling
Photoreceptors such as phytochromes, cryptochromes, UV-light receptor, and phototropins help plants to sense variations in the light quality, quantity, and direction. The red and far-red light photoreceptors, phytochromes, are encoded by a multigene family, PhyA–E in Arabidopsis. While PhyA is the far-red light receptor, PhyB–E function as red light receptors with PhyB playing a predominant role. They mediate very-low-fluence responses (VLFRs), low-fluence responses (LFRs), and the high-irradiance responses (HIRs), with reference to the photon flux density (Casal et al., 1998). Like phyB-9 mutant, gi-100 also shows elongated hypocotyl when grown under saturated red light (Huq et al., 2000). Neither the genes nor the proteins abundance of PhyA and PhyB are influenced in gi-100 (Huq et al., 2000). Therefore, GI was suggested to function downstream of PhyA and PhyB. Mutation in GI leads to decreased VLFR under FR light suggesting its role in PhyA signaling (Oliverio et al., 2007). The gi mutants showed reduced seed germination and cotyledon unfolding in FR light conditions. These phenotypes are rescued by over-expression of GI. This suggested that GI might have a positive role specifically in PhyA mediated VLFR. GI also has a role in regulating flowering in low R:FR ratio which might be attributed to PhyA signaling (Wollenberg et al., 2008). Both PhyA and PhyB form NBs like GI. It would be interesting to determine if Phys and GI are present in the same sub-nuclear complexes and the localization of GI in the NBs alters the Phy-mediated functions.
The gi mutants showed longer hypocotyl in comparison to WT under blue light (Martin-Tryon et al., 2007). Earlier, it had been suggested that GI may be either a positive regulator of TOC1 or act parallel to it for the regulation of hypocotyl elongation. Since only gi not toc1 mutants show the longer hypocotyl in blue light, it can be inferred that GI does not regulate TOC1 for hypocotyl elongation (Martin-Tryon et al., 2007).
Circadian Clock Control
The circadian clock controls many processes depending on the length of the day and night cycle in an organism. In plants, the rhythmic expressions of various genes are influenced by the circadian clock, thereby regulating functions such as elongation of hypocotyl, petioles and inflorescence stem, movement of cotyledon and leaf, and flowering time. CCA1, LATE ELONGATED HYPOCOTYL (LHY), and TOC1 are the core components of circadian oscillator in plants (Somers, 1999). In 2005, the clock was proposed to be an interlocking network of proteins working in a feedback loop (Locke et al., 2005). According to the new model of clock, while the morning elements LHY and CCA1 repress TOC1 transcription, the evening element TOC1 down-regulates LHY/CCA1 accumulation, differing with the earlier observations (Alabadí et al., 2001; Gendron et al., 2012; Huang et al., 2012).
To understand the circadian rhythm in Arabidopsis, the ESPRESSO Quantitative Trait Loci (QTL) was generated from the cross between Ler and Cvi ecotypes (Swarup et al., 1999). Ler and Cvi ecotypes were suggested to comprise of an even distribution of alleles involved in the shortening and lengthening of period, since the progeny of their cross generated lines which had period length both longer and shorter than the parents. GI was identified as one of the genes that could be responsible for regulating the rhythms of cotyledon movement (Park et al., 1999; Swarup et al., 1999). The gi mutants have diverse circadian periods than WT concluding that GI has a role in period length regulation. Mutation in GI affects CHLOROPHYLL A/B-BINDING PROTEIN 2 (CAB2) gene expression which is also under the control of circadian clock (Park et al., 1999).
Soon after a day of imbibition of seeds, GI is required for initiating the rhythmicity of the circadian clock (Salomé et al., 2008). Mutations in the GI locus affect the CCA1 and LHY gene expression in both LDs and SDs conditions (Fowler et al., 1999). A recent study proposed that both the nuclear and cytosolic GI are required to positively and negatively regulate LHY expression, respectively, that fine-tunes the clock function (Kim et al., 2013b). Over-expression or mutations of CCA1 and LHY disrupted the GI expression (Fowler et al., 1999). Accordingly, the double mutant of LHY and CCA1 showed early abundance of GI transcript (Mizoguchi et al., 2002, 2005). It suggests that GI operates in a feedback loop as a component to maintain the rhythmicity and period length of the clock.
The established LHY/CCA1-TOC1 module of the clock could not explain the experimental data like the time difference of about 12 h between LHY/CCA1 abundance in morning and TOC1 accumulation in evening (Alabadí et al., 2001; Locke et al., 2005). It was therefore proposed that LHY/CCA1-TOC1 module comprises of other components. One of the components was predicted to be GI, whose expression followed the same pattern as predicted by the in silico analysis and was subsequently experimentally confirmed (Locke et al., 2006). Further work suggested that GI alone would not be able to regulate the observed time difference (Kawamura et al., 2008). TOC1 in turn is regulated by GI along with ZEITLUPE (ZTL), an F-box protein (Kim et al., 2011). ZTL is a blue light photoreceptor which is stabilized by its interaction with GI and Heat Shock Protein 90 (HSP90). Together the ZTL-GI complex control TOC1 level (Kim et al., 2007).
Temperature compensation is an important characteristic of the circadian clock to maintain the rhythm over a range of environmental temperature. GI was recognized as a candidate regulating temperature compensation effect, especially at higher temperatures (Edwards et al., 2005; Gould et al., 2006). Since fluctuations in the temperature regulate the abundance of GI transcript, it could be plausible that GI and temperature sensing mechanism crosstalk and feedback each other.
Arabidopsis thaliana dawn and dusk, GI regulates the clock rhythm along with ELF4 (Kim et al., 2012). GI was also required for iron-deficiency induced long circadian clock rhythm (Chen et al., 2013). Reduced depolymerization of actin filament caused the period of the circadian clock to shorten, as evident from the shortened period of GI expression (Tóth et al., 2012). Since GI expression is under the control of the circadian clock, GI accumulation pattern has been exploited to screen for novel clock mutants (Onai et al., 2004). Many components that mediate between GI and the clock are still to be unraveled. The role of GI in the regulation of the clock documented till date is summarized in Figure 4.
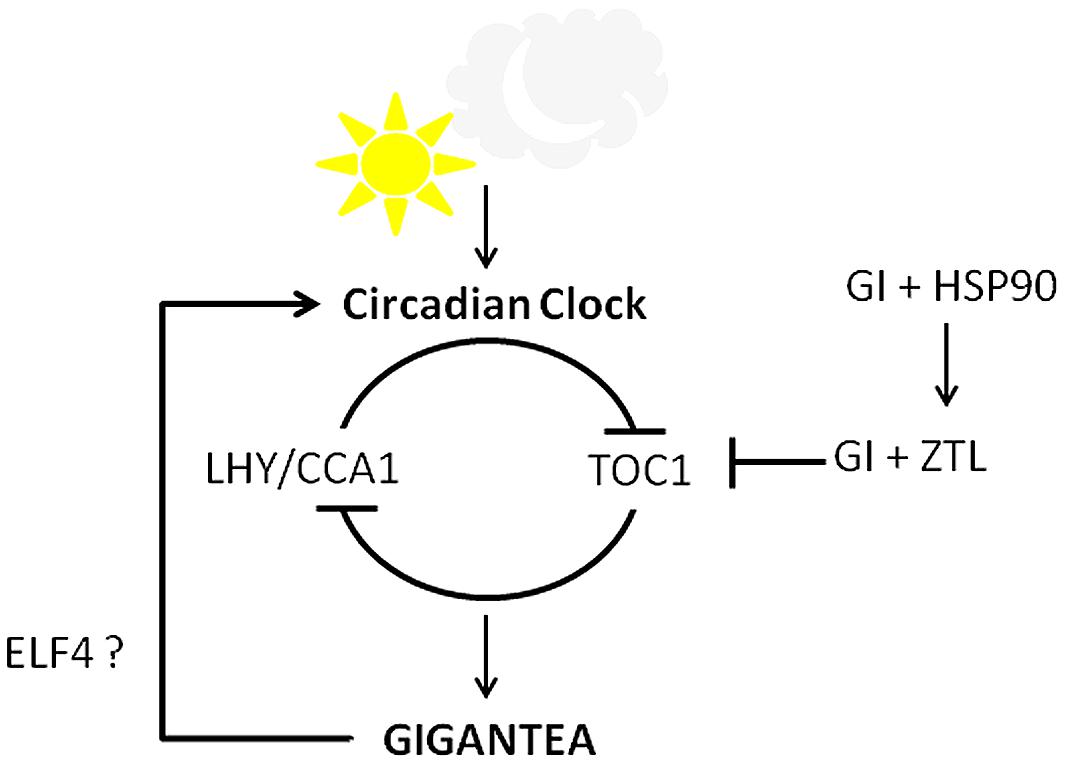
FIGURE 4. Circadian clock control by GI. GI and the central clock components work in a feedback loop. GI along with ELF4 positively regulates the clock while GI and ZTL form a complex to degrade TOC1 in evening. GI and HSP90 regulate the stability of ZTL, thereby influencing clock.
Photoperiodic Flowering-Time Regulation
GIGANTEA is a major mediator between the circadian clock and the master regulator of photoperiodic flowering time control, CO. GI upregulates CO transcription, thereby accelerating time required to flower. Koornneef et al. (1998) showed that a novel mutant, gi-3, is epistatic to CO and FLOWERING LOCUS T (FT) way back. Mutation in GI led to a decrease in the accumulation of CO mRNA without affecting its cycling phase compared to its WT that led to delayed flowering (Suárez-López et al., 2001). Mutants in the GI locus or over-expressors of GI did not discriminate day-length for flowering. Accordingly, gi mutants were later flowering and over-expressors were earlier in both LDs and SDs (Rédei, 1962; Araki and Komeda, 1993; Mizoguchi et al., 2005).
During dawn, CO transcription is repressed by the combinatorial activity of the DOF transcription repressors bound to the CO promoter. In LDs, the expression of FKF1 and GI coincide at ZT10. Therefore, toward the middle of the day the accumulation of GI along with FKF1 forms a complex competent to degrade the DOF factors. This elevates the CO transcription, thereby leading to FT expression (Imaizumi et al., 2003, 2005; Sawa et al., 2007). While in SDs, since FKF1 accumulates 3 h after GI peaks, it does not allow the formation of the degradation complex, therefore leading to a low abundance of CO transcript. This photoperiod pathway where GI regulates FT expression in a CO-dependent manner is schematically depicted in Figure 5. GI regulates the abundance of FKF1, which is involved in the proteasomal degradation of proteins (Fornara et al., 2009). Post-transcriptionally, GI also controls the sub-cellular level of CYCLING DOF FACTOR 2 (CDF2; Fornara et al., 2009). FKF1 belongs to a family of F-Box proteins containing two other candidates – LOV KELCH Protein 2 (LKP2) and ZTL. The blue light dependent interaction between GI and FKF1 is mediated by the LOV (Light, Oxygen, or Voltage) domain of FKF1 and the amino-terminal of GI in vivo (Sawa et al., 2007). The gi-100 mutant is later flowering than the F-Box triple mutant fkf1 ztl-4 lkp2-1. This might be due to the presence of GI in fkf1 ztl-4 lkp2-1, which down-regulates the abundance of CDF transcripts, or the presence of an additional layer of control by GI bypassing the triple F-Box module.
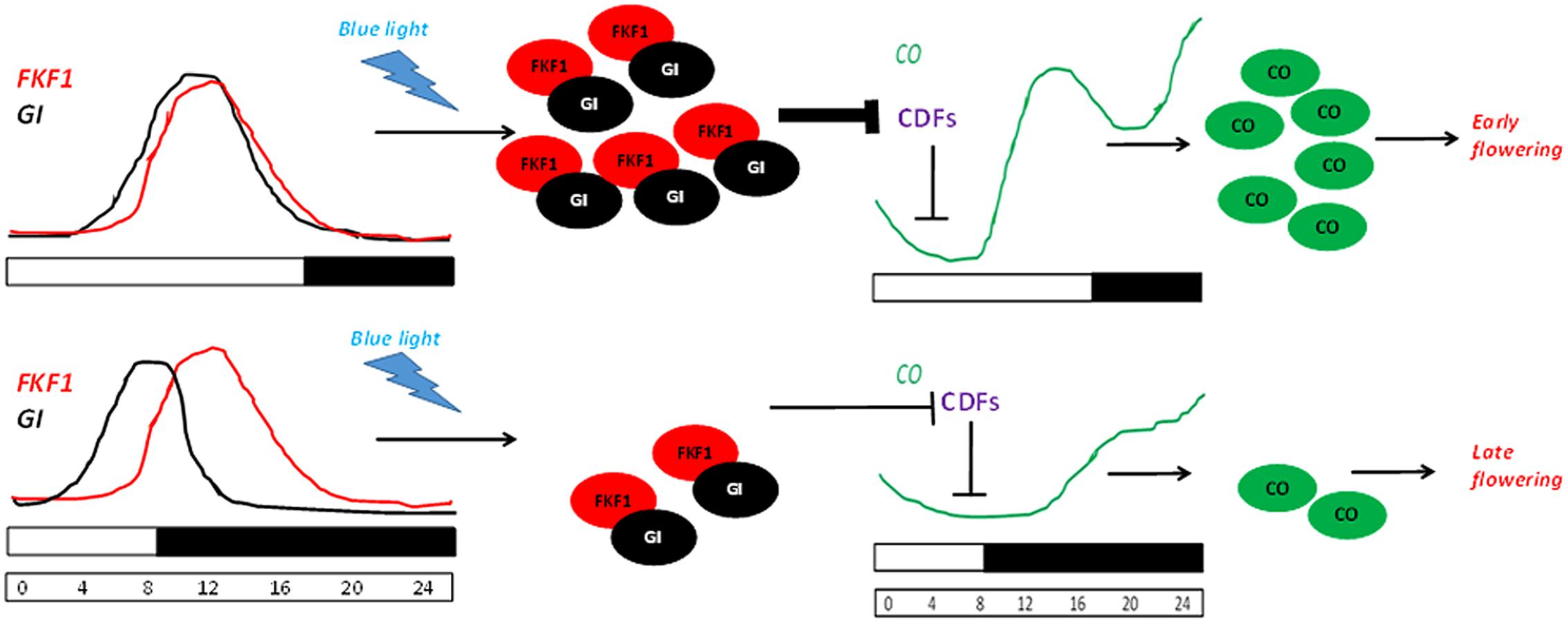
FIGURE 5. GI-FKF1 complex regulates the photoperiodic flowering time. In LD, the peak expression of GI and FLAVIN-BINDING KELCH REPEAT F-BOX 1 (FKF1) coincide, leading to the accumulation of GI-FKF1 complex. The GI-FKF1 complex regulates the degradation of Cycling DOF factors (CDFs) bound to the CONSTANS (CO) promoter. In the absence of the inhibitor CO transcription occurs resulting in the accumulation of CO protein that promotes flowering. While in SDs, less abundance of GI-FKF1 complex does not degrade CDFs repressing CO transcription.
There are at least two independent mechanisms through which GI regulates FT expression independent of CO. While the first mechanism involves microRNA, the second mechanism is through the binding of GI to the FT promoter. The microRNA based control involves miRNA172, which is positively regulated in the presence of GI. The miR172 inhibits the expression of TARGET OF EAT1 (TOE1), an APETALA 2 (AP2)-related transcriptional repressor of FT (Jung et al., 2007). In the recent past, expression of GI specifically in the mesophyll or vascular tissue was carried out. This rescued the late-flowering phenotype of gi-2 under both day length conditions and two different temperatures of 16 and 23°C (Sawa and Kay, 2011). The expression of GI in mesophyll and vascular tissue was done using tissue specific promoters LIGHT-HARVESTING COMPLEX B2.1 (pLhCB2.1) and SUCROSE TRANSPORTER 2 (pSUC2), respectively. While expression pattern of GI under the control of pLhCB2.1 is altered and peaked at ZT 0, GI expressed under the phloem specific promoter led to the over-expression of the transcript with peak at ZT 10. The FT transcript level was up-regulated without an increase in CO mRNA in both day-length conditions. GI was shown to binds to the FT transcriptional repressors such as SHORT VEGETATIVE PHASE (SVP), TEMPRANILLO 1 (TEM1), and TEMPRANILLO 2 (TEM2), and their specific target regions within the FT promoter in the mesophyll, thereby relieving the repression and promoting FT transcription (Sawa and Kay, 2011). The degradation of the FT transcriptional repressors or the unavailability of their binding sites on the FT promoters due to the presence of GI could lead to the abundance of the FT transcript. FT expressed in the vascular tissue normally triggers flowering. Since GI expressed in mesophyll accelerated flowering, elevating the FT level in vasculature, the signal from mesophyll GI most likely induces CO transcription in vasculature. Alternatively, the GI could be transported to the vascular tissues and induce the photoperiod module which needs to be investigated.
Expression of 35S::GI:GFP in gi-3 plants complemented the late flowering phenotype of gi-3. On the contrary, expressing the 35S::GFP:GI in gi-3 caused later flowering compared to the background lines indicating that the N-terminal fusion of GI might be either non-functional or might not be imported into the nucleus. In the transgenic line expressing C-terminal fusion, the fusion protein was localized to the nucleus and formed NBs (Mizoguchi et al., 2005). In an independent study, transgenic plants expressing glucocorticoid receptor (GR) fusion of GI flowered with ∼20 leaves less when treated with dexamethasone, compared to its untreated control which flowered with ∼55 leaves under LDs (Günl et al., 2009). In 15 day old seedlings, the induction of flowering time genes like CO and FT took place ∼28 h after dexamethasone treatment causing early flowering. This indicates that cytoplasmic retention of GI most probably delays time to flower. Mutation in GI is epistatic to mutation in ELF4 and together regulate CO expression (Kim et al., 2012). Recent studies showed that ELF4 sequesters GI into nuclear bodies, thereby preventing GI to associate with the CO promoter (Kim et al., 2013b). It would be interesting to know the nature of the GI nuclear bodies and the components there in, using biochemical approach followed by mass spectrometric analysis.
GIGANTEA interacts with N-terminal tetracopeptide domains of SPINDLY (SPY), a plant O-linked β-N-acetylglucosamine transferase, and antagonizes its activity, thereby, promoting flowering (Tseng et al., 2004). Acetylglucosamine transferases have role in the addition of acetylglucosamine residues to proteins, which often competes with phosphorylation. This suggests that sugar modification may function as an important event in flowering time regulation. The known pathways through which GI regulates flowering are summarized in the Figure 6.
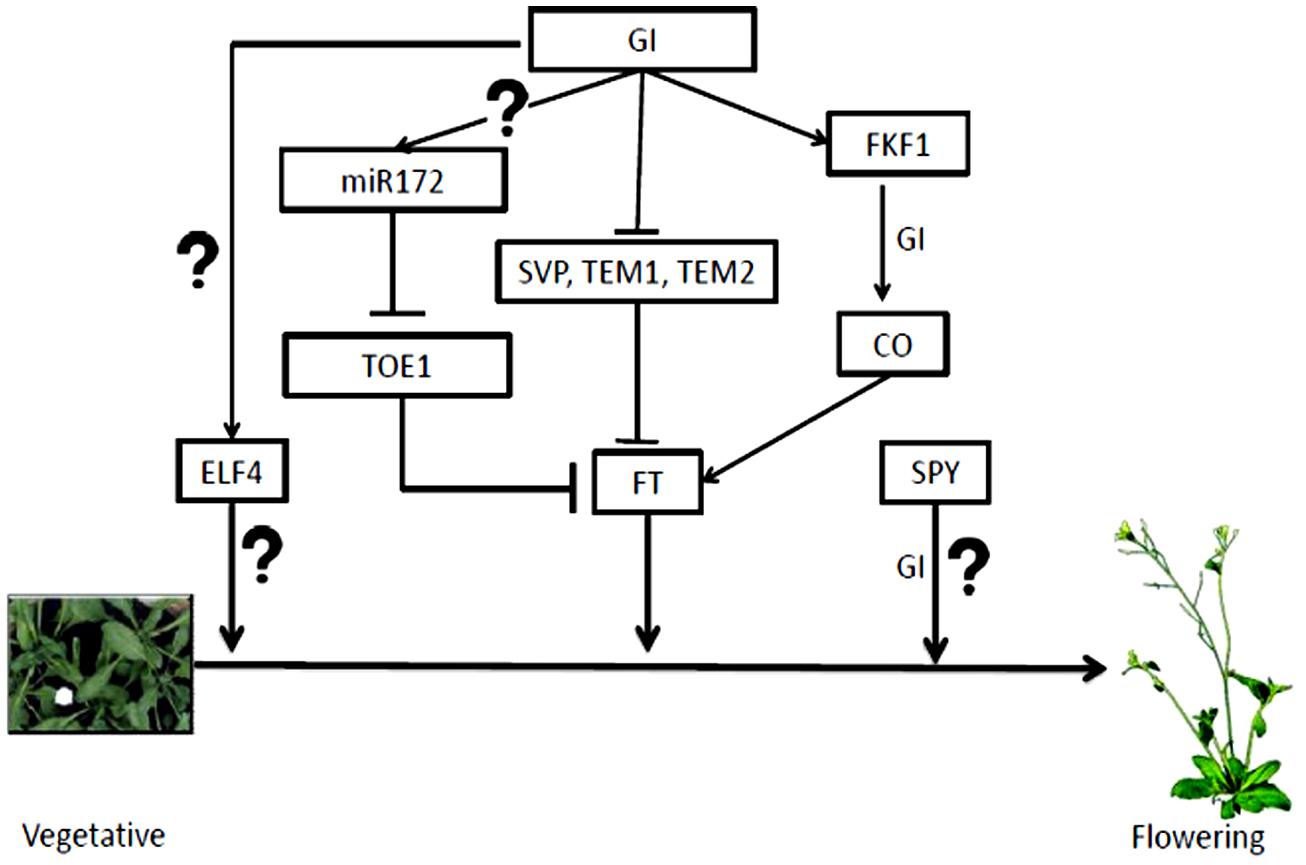
FIGURE 6. Flowering time regulation by GI. GI regulates flowering time through many pathways mostly up-regulating FT. The mechanism of flowering time control by GI along with ELF4 and SPY is unknown. The miR172 processed by GI inhibits TOE1/2 that up-regulates FT transcription. GI also degrades inhibitors of FT transcription like SVP, TEM1 and 2. GI-FKF1 complex tunes CO transcription, which in turn controls FT accumulation.
Pleotropic Functions of GIGANTEA
Besides flowering time, circadian clock, and light signaling regulation, GI has been implicated in other processes such as, sucrose signaling (Dalchau et al., 2011), starch accumulation (Eimert et al., 1995), and stress tolerance (Kurepa et al., 1998a; Fowler and Thomashow, 2002; Kim et al., 2013a; Riboni et al., 2013). The control of cotyledon movement, transpiration, and hypocotyl elongation responses have been shown to be attributed to the concerted activity of SPY and GI (Sothern et al., 2002; Tseng et al., 2004). The precise nature of this interaction is still unclear. However, GI functions antagonistically to SPY. The interaction of GI with SPY and ELF4 independently regulates hypocotyl length, where mutation in ELF4 and SPY are epistatic to gi-2.
GIGANTEA has been demonstrated to play a role between sucrose signaling and the circadian clock while grown in DD (Dalchau et al., 2011). Plants entrained in LD when shifted to DD, maintained the rhythmic GI expression exclusively in the presence of sucrose suggesting light independent control of GI rhythmicity. Although contradictory evidence on role of sucrose on GI expression has been reported, sucrose seems to affect the GI expression through SENSITIVE TO FREEZING6 (SFR6) locus (Knight et al., 2008; Usadel et al., 2008). More precise experiments are required to unravel this mechanism. In the leaves of Arabidopsis, starch accumulation is elevated in the gi mutants (Eimert et al., 1995). On the contrary, presence of multiple copies of GI led to starch accumulation in the progeny of a cross between A. thaliana and A. arenosa, suggesting the antagonistic role of GI in these plants (Ni et al., 2009).
The gi-3 mutants showed higher tolerance capacity to redox cycling agent, paraquat, and H2O2 (Kurepa et al., 1998a). Tolerance against paraquat is counteracted by the exogenously applied polyamines such as spermidine, spermine, and putrescine (Kurepa et al., 1998b). Paraquat treatment upregulated endogenous levels of putrescine in gi-3 and WT. Since exogenous application of polyamines is effective for the resistance, the mechanism of the transporters during this stress needs attention. Oxidative stress due to herbicide imazethapyr has been shown to increase GI abundance and cause earlier flowering by ∼4 days (Qian et al., 2014). The mechanism behind higher tolerance to oxidative stress mediated by GI is still unclear.
Kurepa et al. (1998a) showed that gi mutants, gi-3, gi-4, gi-5, and gi-6, have more chlorophyll accumulation in comparison to WT in presence of paraquat. Even treatment with nitric oxide (NO) reduces the GI mRNA abundance and increases the chlorophyll content (He et al., 2004). In both the cases above, lower abundance of functional GI can be correlated to higher accumulation of chlorophyll. The role of GI in regulating the chlorophyll content needs to be studied in mutants and over-expressors of GI. Chlorophyll accumulation in allotetraploid, obtained by a cross between A. thaliana and A. arenosa, is higher than the WT individuals (Ni et al., 2009). The starch and chlorophyll accumulation in allotetraploids is exactly opposite in comparison to that seen in A. thaliana. The reverse trend might be due to post-transcriptional silencing posed by the presence of multiple homologous sequences of GI transcript, essentially a co-suppression phenomenon.
Dynamin, a GTPase having role in vesicle recycling during endocytosis, was found to interact with TAP tagged GI in rice (Abe et al., 2008). Although mutation in dynamin gene did not have any effect on the flowering time, it showed aerial rosette phenotype in Arabidopsis. In Arabidopsis, GI has been found to be involved in setting of fruits (Brock et al., 2007). No significant association of the GI haplogroup was detected with days to flower, petiole length, and inflorescence height. A significant association was observed between one haplogroup with fruit set, producing 14% more fruit than other haplogroups. Such studies in the crop plants could help in increasing the yield.
GIGANTEA mRNA levels increases about five- to eightfold in the cold treated Arabidopsis plants suggesting that GI is a cold-responsive gene (Fowler and Thomashow, 2002). The flowering time of gi mutants was further delayed when exposed to low temperature compared to WT (Cao et al., 2005). C-repeat Binding proteins (CBFs) have been known to regulate various genes responsive to cold and are implicated in cold stress tolerance. On the contrary, Cao et al. (2005), it was revealed that GI regulates cold acclimation through CBF-independent pathway. The ability to tolerate and acclimatize toward cold is reduced in gi mutants suggesting the protective role of GI in cold tolerance.
Recently, the role of GI under salt stress was documented (Kim et al., 2013a). Although, salt stress did not affect the GI expression, it affected the GI protein stability in pGI::GI-HA transgenics (Cao et al., 2005; Kim et al., 2013a). It seems plausible that there is a mechanism at the post-translational level that regulates GI abundance. GI also regulated the activities of the proteins involved in the salt stress tolerance. It interacts with Salt Overly Sensitive 2 (SOS2) directly and inhibits the activity of SOS1, a Na+/H+ anti-porter. Therefore, GI is a negative regulator of salt tolerance and is degraded during salt stress. According to a recent model, plants under salt stress would flower later than when grown in normal growth conditions reasoned for the degradation of GI (Park et al., 2013).
In At and other plants, the tolerance to higher salinity, enhanced cold, and sustained drought were manifested by the increase of sub-cellular level of abscisic acid (ABA). Recent reports indicated that GI has role in ABA-dependent drought escape tolerance. It suggests that the GI regulation of salt and cold stress tolerance could very likely be ABA-mediated (Riboni et al., 2013). Drought stress up-regulates GI transcription and in turn, increases the abundance of miR172E variant (Han et al., 2013). WRKY DNA binding protein 44 (WRKY44) was found to be suppressed by GI in drought stress and interact with TOE1. GI-miR172-WRKY44 were proposed to be in the same pathway possibly associated with drought stress tolerance. On the same line of thinking, the light dependent GI-mediated stomatal opening response could be ABA mediated (Ando et al., 2013). GI also has a role in wall in-growth deposition in phloem parenchyma transfer cells in A. thaliana in response to high light and cold stress (Edwards et al., 2010; Chinnappa et al., 2013).
Role of GIGANTEA Homologs
GIGANTEA homologs in prokaryotes, fungi, mosses, or animals have not been reported as yet (Holm et al., 2010). GI homolog has been shown to be absent in the green unicellular alga Ostreococcus tauri (Corellou et al., 2009). Evolution of GI has been correlated with the evolution of higher plants from liverwort onward, although being absent in mosses. The evolution of GI can be proposed to have taken place alongside the origin of land plants. The role of GI in light signaling, circadian clock control, and flowering time regulation seems to be conserved across the plant kingdom, as inferred from the various studies to understand the role of GI homologs in Arabidopsis. GI homologs from the non-flowering and flowering plants have been summarized below.
The GI-FKF1 interaction and function has been recently shown to be conserved in the liverwort Marchantia polymorpha (Kubota et al., 2014). The LOV domain of FKF1, which has been found to be required for the interaction with GI, contains a conserved cysteine residue in AtGI and MpGI important for its blue light dependent functions (Sawa et al., 2007). GI-FKF1 module has been proposed to be important for the transition of plants from water to land and the evolution of vascular system. The Marchantia polymorpha ortholog of GI, MpGI, has been shown to partially rescue the late flowering phenotype of Arabidopsis gi mutant suggesting the functional conservation of GI across the plant kingdom. The FKF1 homologs have been shown to be present in A. thaliana (AtFKF1, AtZTL, and AtLKP2), Oryza sativa (OsFKF1, OsZTL1, and OsZTL2), Glycine max (GmFKF1, GmZTL1, and GmZTL2), Triticum aestivum (TaFKF1 and TaZTL), Allium cepa (AcFKF1 and AcZTL), Mesembryanthemum crystallinum (McFKF1 and McZTL), and Selaginella moellendorffii (SmFKF; Kubota et al., 2014). The GI counterparts in the above mentioned species are also conserved. This shows that GI-FKF1 module has been conserved since the primitive time and thus might be have been important in shaping the development of higher plant. This light perceiving module needs to be studied in detail to understand the evolution of various functions and residues along with putative domains required to carry out these functions in plants. The conserved interaction of GI with FKF1 has been shown to be conserved in soybean.
Gymnosperm
Norway Spruce (Picea abies)
GIGANTEA ortholog of Picea abies, PaGI and AtGI share 58% identity and 72% similarity. Natural variations in GI have been correlated to clinal variations in the different populations of close relative of the Scandinavian Norway spruce (Chen et al., 2014). Over-expression of PaGI in WT Arabidopsis did not show any phenotypical changes (Karlgren et al., 2013). However, when PaGI was over-expressed in gi-2 mutant, it partially rescued the late flowering phenotype and flowered at the same time as WT plants suggesting that PaGI and AtGI are functionally conserved to large extent. The strength of the over-expression has neither been verified at the gene expression level nor the protein accumulation level and therefore needs to be confirmed.
Angiosperms (Monocots)
Barley (Hordeum vulgare)
GIGANTEA homolog in Barley was identified using BLAST searches and later confirmed by Southern hybridization analyses. Only one homolog was detected in barley. Barley GI (HvGI) has ∼94 and ∼79% similarity with OsGI and AtGI, respectively (Dunford et al., 2005). Barley, being a long-day plant, its GI expression followed the pattern documented for AtGI. Characteristically, in SDs, the peak of expression was noticed about 6–9 h after dawn whereas, in LDs, the peak is shifted to 15 h after dawn (Dunford et al., 2005). The mutation in HvELF3 (mat-a.8), a 4 bp deletion causing frame shift and premature stop codon, was found in the barley cultivar Mari (Zakhrabekova et al., 2012). This mutation led to the up-regulation of HvGI transcription and was found to be the reason for early flowering phenotype in this cultivar. Although, post-translational interaction between AtELF3 and AtGI is known, no evidence is there in Arabidopsis suggesting the transcriptional regulation of GI by ELF3.
Duckweed (Lemna gibba)
The AtGI homolog of L. gibba, LgGIH1, a LD plant, plays a pivotal role in its circadian clock control, since the LgGIH1 knockdown resulted in the arrhythmic gene expression phenotype in plants (Serikawa et al., 2008). Earlier reports suggested that AtGI and LgGIH1 followed similar expression pattern in both LD and LL conditions (Miwa et al., 2006). The function of GI and ELF3 homologs are shown to be conserved between Arabidopsis and L. gibba.
Maize (Zea mays)
Maize is a SD plant, which has two diurnally regulated GI homologs called gigantea of Z. mays 1a and 1b (gi1 and 2) due to tetraploidy events and genome evolution expressed in leaves (Gaut and Doebley, 1997; Swigonová et al., 2004; Miller et al., 2008; Hayes et al., 2010; Khan et al., 2010; Schnable et al., 2011). Among the two homologs, the gigantea1 transcript was highly expressed. Mutation in gi1 caused early flowering in LD but had lesser effects in SD. The gi1 mutation also increases plant height and alters the timing of the vegetative phase (Bendix et al., 2013). The early flowering phenotype of gi1 mutant was due to the conserved pathway involving the up-regulation of CO-like flowering regulatory gene called CO of Z. mays1 (conz1) and FT-like floral activator gene named Z. mays centroradialis8 (zcn8).
Purple False Brome (Brachypodium distachyon)
GIGANTEA ortholog of B. distachyon (BdGI) is rhythmically regulated by the circadian clock and up-regulated by both cold and dark (Hong et al., 2010). BdGI was identified by BLAST search followed by Southern hybridization analysis. The BdGI transcript level was found to be oscillating in both SD and LD conditions, like AtGI. While the lowest transcript level in both SD and LD was at ZT 0, the peak in SD was at ZT 8 and in LD was at ZT 12. BdGI shares 65% identity with AtGI. BdGI, like AtGI, is a nuclear localizing protein and interacts with COP1 and ZTL proteins as evident from the yeast two-hybrid assays. BdGI complements the late flowering phenotype of Arabidopsis gi-2 mutant suggesting the conserved function of GI in monocots and dicots. While PhyC does not show a pronounced effect in the LD model Arabidopsis, it causes late flowering in this temperate grass (Woods et al., 2014). In phyC mutants, GI expression is almost undetectable. The low GI expression could explain the lower abundance of the homologs of CO and FT. The delayed flowering phenotype suggests that the photoperiodic flowering pathway through GI is conserved in grasses as in Arabidopsis.
Rice (Oryza sativa)
Rice and Arabidopsis GI share 67% similarity and the NLS are quite conserved between OsGI and AtGI (Hayama et al., 2002). GI expression pattern was similar in both rice and At (Hayama et al., 2002) and similarly, OsGI acts as a positive regulator of Hd1 (CO homolog of rice; Hayama et al., 2003). It controls the rhythm of nearly 27000 genes in rice (Izawa et al., 2011). When gi mutants are grown in field conditions, sucrose, and starch content increases, chlorophyll content decreases, stomatal conductance increases, panicle, and spikelet number increases and fertility was reduced. OsGI was shown to be involved in ETR2 (ethylene receptor)-dependent late flowering phenotype and starch accumulation thus, regulating the developmental transition based on the availability of energy (Wuriyanghan et al., 2009).
Tulip (Liriodendron tulipifera)
GIGANTEA ortholog was shown to be closer to eudicot GI sequence than the monocot sequences (Liang et al., 2010).
Wheat (Triticum aestivum L.)
Wheat is a LD plant and has been shown to have an ortholog of AtGI, referred as TaGI1 (Zhao et al., 2005). TaGI1 has ∼81 and 63% identity with OsGI and AtGI, respectively. The TaGI1 follow rhythmic pattern of expression similar to that of Arabidopsis and over-expression of TaGI1 complements late flowering phenotype of gi-2 mutant Arabidopsis. TaGI was also associated with “earliness phenotype” of wheat which helps in its adaptation and increase in yield in varied environmental conditions (Rousset et al., 2011).
Angiosperms (Dicots)
Common Ice Plant (Mesembryanthemum crystallinum)
A crassulacean acid metabolism plant, Mesembryanthemum crystallinum, also showed a rhythmic expression of the orthologs of GI, McGI (Boxall et al., 2005). The ortholog was identified using BLAST search and later isolated and sequenced. McGI expression peaks at ZT 9 similar to AtGI.
Morning Glory (Pharbitis nil)
PnGI protein shares 70 and 67% identity with AtGI and OsGI protein, respectively (Higuchi et al., 2011). PnGI mRNA is also circadian regulated like the other GI orthologs. Over-expression of PnGI led to altered period length affecting the expression pattern of downstream genes. Pharbitis nil is a SD plant, and like OsGI, PnGI inhibits the expression of PnFT (FT homolog of morning glory).
Pea (Pisum sativum)
LATE BLOOMER 1 (LATE1) is the AtGI ortholog in pea, a LD plant, and follows a rhythmic pattern of expression as seen in Arabidopsis (Hecht et al., 2007). LATE 1 was shown to be regulating the pea homologs of Arabidopsis circadian clock genes. Apart from its role in flowering time and circadian clock regulation, LATE1 has been implicated in Phy-B dependent seed de-etiolation in red light. LATE1 was found to regulate circadian clock gene expression in constant light and dark (Liew et al., 2009). In LD and SD, LATE1 was shown to control a mobile signal that regulates the flowering time.
Radish (Raphanus sativa)
In another instance, expression of antisense AtGI gene, under the constitutive 35S promoter, led to delayed bolting in LDs, proving that GI has an important role in photoperiodic flowering time control in this plant (Curtis et al., 2002). The bolting and flowering time was delayed by 17 and 18 days, respectively, with respect to WT plants.
Soybean (Glycine max)
Glycine max, a SD plant, has two GI orthologs – GmGIa and GmGIb (Watanabe et al., 2011). Both the GmGI sequences have nearly 70–91% identity to eudicot and monocot genes. Like OsGI, GmGI regulated GmFT paralogs. GmGI has been shown to have role in soybean seed maturity. GmGI loss of function leads to early flowering as in the model SD rice plant. Interestingly, a recent study in soybean suggested that there are three AtGI homologs in the soybean genome unlike previously suggested two orthologs GmGIa and GmGIb (Li et al., 2013). The third form is a result of alternative splice form of GmGIa, resulting in GmGIα and GmGIβ. The GI orthologs were diurnally regulated and differentially expressed in different tissues adding up to a more complex regulation. GmGI proteins have the conserved NLS and localize to nucleus. GmGI proteins have been shown to interact with orthologs of FKF1 in soybean suggesting that function most likely is conserved.
Tomato (Solanum lycopersicum)
Tomato is a day neutral plant. GI was shown to be up-regulated and inhibit tomato seed germination thereby promoting seed dormancy under FR condition in the presence of functional PhyA (Auge et al., 2009). On the contrary, in Arabidopsis, loss of function of GI led to elevated dormancy (Penfield and Hall, 2009). In other members of the Solanaceae such as potato and tobacco, photoperiodic control of GI was also shown to be operational (Rutitzky et al., 2009).
The conserved diurnal regulation of GI in different plants described above suggests the prevalence of an important transcriptional machinery as well as the GI promoter. The availability of GI antiserum would help to understand the regulation of GI in these crop plants. The localization and the stability of GI in most of these plants are still to be addressed. While few of the interaction with proteins such as orthologs of ELF3, COP1, ZTL, and FKF1 are shown to be conserved, the function of these complexes needs to be disclosed in various species.
Conclusion and Perspective
GIGANTEA seems to be a very important plant protein involved in various processes, from developmental regulation to metabolic flux. Despite its pivotal roles, it is surprising that GI null mutants are not lethal. Being a large protein, it might satisfy to function in several pathways summarized, yet to be fully understood. It would be a great challenge to understand and connect the functional roles of GI at different developmental stages. Although GI is a multifunctional protein, the role of its various functional domains are still in darkness. A functional antiserum against a conserved domain of GI that would detect the endogenous level of protein across species and in multiple mutational background would be very useful. The lack of such an antiserum possesses a serious bottleneck delaying the understanding of its abundance, regulation at the protein level and regulatory functions like the GI-FKF1 module across the plant kingdom. Despite this problem, several elegant experiments have been published where researchers have attempted to understand its role using transgenic plants expressing tagged versions of GI. Although time-consuming, these are the impressive feats that place GI mechanistically in a network of photoperiod control pathway.
The role of GI in flowering time regulation, circadian clock control, and light signaling is still being pursued. But less-known functions such as sucrose signaling, chlorophyll accumulation, oxidative stress resistance demand more attention. More functions of GI are beginning to be documented. Recently, the emerging role of GI in salt tolerance has been demonstrated, which indicates that we are still not saturated in understanding the various functions GI. It would be interesting to understand how GI regulates so many functions before going into the complex cross talk between them it can fine tune. The lower plant moss Physcomitrella patens does not have a GI ortholog but still carries out most of the developmental aspects except flowering. It is very interesting to note that they do have CO-like genes, therefore the evolution of GI function is still an interesting area and demands further attention (Zobell et al., 2005).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors acknowledge the Department of Biotechnology, India and National Institute of Science Education and Research, India for its funding and support. The authors would like to thank Dr. Chandan Goswami and Ms. Anamika Singh to help in editing the manuscript.
References
Abe, M., Fujiwara, M., Kurotani, K., Yokoi, S., and Shimamoto, K. (2008). Identification of dynamin as an interactor of rice GIGANTEA by tandem affinity purification (TAP). Plant Cell Physiol. 49, 420–432. doi: 10.1093/pcp/pcn019
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Alabadí, D., Oyama, T., Yanovsky, M. J., Harmon, F. G., Más, P., and Kay, S. A. (2001). Reciprocal regulation between TOC1 and LHY/CCA1 within the Arabidopsis circadian clock. Science 293, 880–883. doi: 10.1126/science.1061320
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ando, E., Ohnishi, M., Wang, Y., Matsushita, T., Watanabe, A., Hayashi,Y.,et al. (2013). TWIN SISTER OF FT, GIGANTEA and CONSTANS have a positive but indirect effect on blue light-induced stomatal opening in Arabidopsis. Plant Physiol. 162, 1529–1538. doi: 10.1104/pp.113.217984
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Araki, T., and Komeda, Y. (1993). Analysis of the role of the late-flowering locus, GI, in the flowering of Arabidopsis thaliana. Plant J. 3, 231–239. doi: 10.1046/j.1365-313X.1993.t01-15-00999.x
Auge, G. A., Perelman, S., Crocco, C. D., Sánchez, R. A., and Botto, J. F. (2009). Gene expression analysis of light-modulated germination in tomato seeds. New Phytol. 183, 301–314. doi: 10.1111/j.1469-8137.2009.02867.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Aukerman, M. J., Hirschfeld, M., Wester, L., Weaver, M., Clack, T., Amasino, R. M.,et al. (1997). A deletion in the PHYD gene of the Arabidopsis Wassilewskija ecotype defines a role for phytochrome D in red/far-red light sensing. Plant Cell 9, 1317–1326. doi: 10.1105/tpc.9.8.1317
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bendix, C., Mendoza, J. M., Stanley, D. N., Meeley, R., and Harmon, F. G. (2013). The circadian clock-associated gene gigantea1 affects maize developmental transitions. Plant Cell Environ. 36, 1379–1390. doi: 10.1111/pce.12067
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Black, M. M., Stockum, C., Dickson, J. M., Putterill, J., and Arcus, V. L. (2011). Expression, purification and characterisation of GIGANTEA: a circadian clock-controlled regulator of photoperiodic flowering in plants. Protein Expr. Purif. 76, 197–204. doi: 10.1016/j.pep.2010.11.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Boxall, S. F., Foster, J. M., Bohnert, H. J., Cushman, J. C., Nimmo, H. G., and Hartwell, J. (2005). Conservation and divergence of circadian clock operation in a stress-inducible Crassulacean acid metabolism species reveals clock compensation against stress. Plant Physiol. 137, 969–982. doi: 10.1104/pp.104.054577
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Brock, M. T., Tiffin, P., and Weinig, C. (2007). Sequence diversity and haplotype associations with phenotypic responses to crowding: GIGANTEA affects fruitset in Arabidopsis thaliana. Mol. Ecol. 16, 3050–3062. doi: 10.1111/j.1365-294X.2007.03298.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cao, S., Ye, M., and Jiang, S. (2005). Involvement of GIGANTEA gene in the regulation of the cold stress response in Arabidopsis. Plant Cell Rep. 24, 683–690. doi: 10.1007/s00299-005-0061-x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Casal, J. J., Sánchez, R. A., and Botto, J. F. (1998). Modes of action of phytochromes. J. Exp. Bot. 49, 127–138. doi: 10.1093/jxb/49.319.127
Chen, J., Tsuda, Y., Stocks, M., Källman, T., Xu, N., Kärkkäinen, K.,et al. (2014). Clinal variation at phenology-related genes in Spruce: parallel evolution in FTL2 and Gigantea? Genetics 197, 1025–1038. doi: 10.1534/genetics.114.163063
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, Y., Wang, Y., Shin, L., Wu, J., Shanmugam, V., Tsednee, M.,et al. (2013). Iron is involved in the maintenance of circadian period length in Arabidopsis. Plant Physiol. 161, 1409–1420. doi: 10.1104/pp.112.212068
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chinnappa, K. S. A., Nguyen, T. T. S., Hou, J., Wu, Y., and McCurdy, D. W. (2013). Phloem parenchyma transfer cells in Arabidopsis – an experimental system to identify transcriptional regulators of wall ingrowth formation. Front. Plant Sci. 4:102. doi: 10.3389/fpls.2013.00102
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Corellou, F., Schwartz, C., Motta, J., Djouani-Tahri, E. B., Sanchez, F., and Bouget, F. (2009). Clocks in the green lineage: comparative functional analysis of the circadian architecture of the picoeukaryote Ostreococcus. Plant Cell 21, 3436–3449. doi: 10.1105/tpc.109.068825
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Curtis, I. S., Nam, H. G., Yun, J. Y., and Seo, K. H. (2002). Expression of an antisense GIGANTEA gene fragment in transgenic radish causes delayed bolting and flowering. Transgenic Res. 11, 249–256. doi: 10.1023/A:1015655606996
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dalchau, N., Baek, S. J., Briggs, H. M., Robertson, F. C., Dodd, A. N., Gardner, M. J.,et al. (2011). The circadian oscillator gene GIGANTEA mediates a long-term response of the Arabidopsis thaliana circadian clock to sucrose. Proc. Natl. Acad. Sci. U.S.A. 108, 5104–5109. doi: 10.1073/pnas.1015452108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
David, K. M., Armbruster, U., Tama, N., and Putterill, J. (2006). Arabidopsis GIGANTEA protein is post-transcriptionally regulated by light and dark. FEBS Lett. 580, 1193–1197. doi: 10.1016/j.febslet.2006.01.016
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
de Montaigu, A., Tóth, R., and Coupland, G. (2010). Plant development goes like clockwork. Trends Genet. 26, 296–306. doi: 10.1016/j.tig.2010.04.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dunford, R. P., Griffiths, S., Christodoulou, V., and Laurie, D. A. (2005). Characterisation of a barley (Hordeum vulgare L.) homologue of the Arabidopsis flowering time regulator GIGANTEA. Theor. Appl. Genet. 110, 925–931. doi: 10.1007/s00122-004-1912-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Edwards, J., Martin, A. P., Andriunas, F., Offler, C. E., Patrick, J. W., and McCurdy, D. W. (2010). GIGANTEA is a component of a regulatory pathway determining wall ingrowth deposition in phloem parenchyma transfer cells of Arabidopsis thaliana. Plant J. 63, 651–661. doi: 10.1111/j.1365-313X.2010.04269.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Edwards, K. D., Lynn, J. R., Gyula, P., Nagy, F., and Millar, A. J. (2005). Natural allelic variation in the temperature-compensation mechanisms of the Arabidopsis thaliana circadian clock. Genetics 170, 387–400. doi: 10.1534/genetics.104.035238
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Eimert, K., Wang, S., Lue, W., and Chen, J. (1995). Monogenic recessive mutations causing both late floral lnitiation and excess starch accumulation in Arabidopsis. Plant Cell 7, 1703–1712. doi: 10.1105/tpc.7.10.1703
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fornara, F., Panigrahi, K. C. S., Gissot, L., Sauerbrunn, N., Rühl, M., Jarillo, J. A.,et al. (2009). Arabidopsis DOF transcription factors act redundantly to reduce CONSTANS expression and are essential for a photoperiodic flowering response. Dev. Cell 17, 75–86. doi: 10.1016/j.devcel.2009.06.015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fowler, S., Lee, K., Onouchi, H., Samach, A., Richardson, K., Morris, B.,et al. (1999). GIGANTEA: a circadian clock-controlled gene that regulates photoperiodic flowering in Arabidopsis and encodes a protein with several possible membrane-spanning domains. EMBO J. 18, 4679–4688. doi: 10.1093/emboj/18.17.4679
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fowler, S., and Thomashow, M. F. (2002). Arabidopsis transcriptome profiling indicates that multiple regulatory pathways are activated during cold acclimation in addition to the CBF Cold Response Pathway. Plant Cell 14, 1675–1690. doi: 10.1105/tpc.003483
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gaut, B. S., and Doebley, J. F. (1997). DNA sequence evidence for the segmental allotetraploid origin of maize. Proc. Natl. Acad. Sci. U.S.A. 94, 6809–6814. doi: 10.1073/pnas.94.13.6809
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gendron, J. M., Pruneda-Paz, J. L., Doherty, C. J., Gross, A. M., Kang, S. E., and Kay, S. A. (2012). Arabidopsis circadian clock protein, TOC1, is a DNA-binding transcription factor. Proc. Natl. Acad. Sci. U.S.A. 109, 3167–3172. doi: 10.1073/pnas.1200355109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gould, P. D., Locke, J. C. W., Larue, C., Southern, M. M., Davis, S. J., Hanano, S.,et al. (2006). The molecular basis of temperature compensation in the Arabidopsis circadian clock. Plant Cell 18, 1177–1187. doi: 10.1105/tpc.105.039990
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Günl, M., Liew, E. F., David, K., and Putterill, J. (2009). Analysis of a post-translational steroid induction system for GIGANTEA in Arabidopsis. BMC Plant Biol. 9:141. doi: 10.1186/1471-2229-9-141
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hall, A., Bastow, R. M., Davis, S. J., Hanano, S., McWatters, H. G., Hibberd, V.,et al. (2003). The TIME FOR COFFEE gene maintains the amplitude and timing of Arabidopsis circadian clocks. Plant Cell 15, 2719–2729. doi: 10.1105/tpc.013730
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Han, Y., Zhang, X., Wang, Y., and Ming, F. (2013). The suppression of WRKY44 by GIGANTEA-miR172 pathway is involved in drought response of Arabidopsis thaliana. PLoS ONE 8:e73541. doi: 10.1371/journal.pone.0073541
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hayama, R., Izawa, T., and Shimamoto, K. (2002). Isolation of rice genes possibly involved in the photoperiodic control of flowering by a fluorescent differential display method. Plant Cell Physiol. 43, 494–504. doi: 10.1093/pcp/pcf059
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hayama, R., Yokoi, S., Tamaki, S., Yano, M., and Shimamoto, K. (2003). Adaptation of photoperiodic control pathways produces short-day flowering in rice. Nature 422, 719–722. doi: 10.1038/nature01549
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hayes, K. R., Beatty, M., Meng, X., Simmons, C. R., Habben, J. E., and Danilevskaya, O. N. (2010). Maize global transcriptomics reveals pervasive leaf diurnal rhythms but rhythms in developing ears are largely limited to the core oscillator. PLoS ONE 5:e12887. doi: 10.1371/journal.pone.0012887
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
He, Y., Tang, R. H., Hao, Y., Stevens, R. D., Cook, C. W., Ahn, S. M.,et al. (2004). Nitric oxide represses the Arabidopsis floral transition. Science 305, 1968–1971. doi: 10.1126/science.1098837
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hecht, V., Knowles, C. L., Vander Schoor, J. K., Liew, L. C., Jones, S. E., Lambert, M. J.,et al. (2007). Pea LATE BLOOMER1 is a GIGANTEA ortholog with roles in photoperiodic flowering, deetiolation, and transcriptional regulation of circadian clock gene homologs. Plant Physiol. 144, 648–661. doi: 10.1104/pp.107.096818
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Higuchi, Y., Sage-Ono, K., Sasaki, R., Ohtsuki, N., Hoshino, A., Iida, S.,et al. (2011). Constitutive expression of the GIGANTEA ortholog affects circadian rhythms and suppresses one-shot induction of flowering in Pharbitis nil, a typical short-day plant. Plant Cell Physiol. 52, 638–650. doi: 10.1093/pcp/pcr023
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Holm, K., Källman, T., Gyllenstrand, N., Hedman, H., and Lagercrantz, U. (2010). Does the core circadian clock in the moss Physcomitrella patens (Bryophyta) comprise a single loop? BMC Plant Biol. 10:109. doi: 10.1186/1471-2229-10-109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hong, S. Y., Lee, S., Seo, P. J., Yang, M. S., and Park, C. M. (2010). Identification and molecular characterization of a Brachypodium distachyon GIGANTEA gene: functional conservation in monocot and dicot plants. Plant Mol. Biol. 72, 485–497. doi: 10.1007/s11103-009-9586-7
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Huang, W., Pérez-García, P., Pokhilko, A., Millar, A. J., Antoshechkin, I., Riechmann, J. L.,et al. (2012). Mapping the core of the Arabidopsis circadian clock defines the network structure of the oscillator. Science 336, 75–79. doi: 10.1126/science.1219075
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Huq, E., Tepperman, J. M., and Quail, P. H. (2000). GIGANTEA is a nuclear protein involved in phytochrome signaling in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 97, 9789–9794. doi: 10.1073/pnas.170283997
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Imaizumi, T., Schultz, T. F., Harmon, F. G., Ho, L. A., and Kay, S. A. (2005). FKF1 F-box protein mediates cyclic degradation of a repressor of CONSTANS in Arabidopsis. Science 309, 293–297. doi: 10.1126/science.1110586
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Imaizumi, T., Tran, H. G., Swartz, T. E., Briggs, W. R., and Kay, S. A. (2003). FKF1 is essential for photoperiodic-specific light signalling in Arabidopsis. Nature 426, 302–306. doi: 10.1038/nature02090
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Izawa, T., Mihara, M., Suzuki, Y., Gupta, M., Itoh, H., Nagano, A. J.,et al. (2011). Os-GIGANTEA confers robust diurnal rhythms on the global transcriptome of rice in the field. Plant Cell 23, 1741–1755. doi: 10.1105/tpc.111.083238
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jung, J., Seo, Y., Seo, P. J., Reyes, J. L., Yun, J., Chua, N., etal. (2007). The GIGANTEA-regulated microRNA172 mediates photoperiodic flowering independent of CONSTANS in Arabidopsis. Plant Cell 19, 2736–2748. doi: 10.1105/tpc.107.054528
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karlgren, A., Gyllenstrand, N., Källman, T., and Lagercrantz, U. (2013). Conserved function of core clock proteins in the gymnosperm Norway spruce (Picea abies L. Karst). PLoS ONE 8:e60110. doi: 10.1371/journal.pone.0060110
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kawamura, H., Ito, S., Yamashino, T., Niwa, Y., Nakamichi, N., and Mizuno, T. (2008). Characterisation of genetic links between two clock-associated genes, GI and PRR5 in the current clock model of Arabidopsis thaliana. Biosci. Biotechnol. Biochem. 72, 2770–2774. doi: 10.1271/bbb.80321
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Khan, S., Rowe, S. C., and Harmon, F. G. (2010). Coordination of the maize transcriptome by a conserved circadian clock. BMC Plant Biol. 10:126. doi: 10.1186/1471-2229-10-126
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, T. S., Kim, W. Y., Fujiwara, S., Kim, J., Cha, J. Y., Park, J. H.,et al. (2011). HSP90 functions in the circadian clock through stabilization of the client F-box protein ZEITLUPE. Proc. Natl. Acad. Sci. U.S.A. 108, 16843–16848. doi: 10.1073/pnas.1110406108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, W., Ali, Z., Park, H. J., Park, S. J., Cha, J., Perez-Hormaeche, J.,et al. (2013a). Release of SOS2 kinase from sequestration with GIGANTEA determines salt tolerance in Arabidopsis. Nat. Commun. 4:1352. doi: 10.1038/ncomms2357
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, Y., Han, S., Yeom, M., Kim, H., Lim, J., Cha, J. Y.,et al. (2013b). Balanced nucleocytosolic partitioning defines a spatial network to coordinate circadian physiology in plants. Dev. Cell 26, 73–85. doi: 10.1016/j.devcel.2013.06.006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, Y., Lim, J., Yeom, M., Kim, H., Kim, J., Wang, L.,et al. (2013c). ELF4 regulates GIGANTEA chromatin access through subnuclear sequestration. Cell Rep. 3, 671–677. doi: 10.1016/j.celrep.2013.02.021
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, W., Fujiwara, S., Suh, S., Kim, J., Kim, Y., Han, L.,et al. (2007). ZEITLUPE is a circadian photoreceptor stabilized by GIGANTEA in blue light. Nature 449, 356–360. doi: 10.1038/nature06132
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, Y., Yeom, M., Kim, H., Lim, J., Koo, H. J., Hwang, D.,et al. (2012). GIGANTEA and EARLY FLOWERING 4 in Arabidopsis exhibit differential phase-specific genetic influences over a diurnal cycle. Mol. Plant 5, 678–687. doi: 10.1093/mp/sss005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Knight, H., Thomson, A. J. W., and McWatters, H. G. (2008). SENSITIVE TO FREEZING6 integrates cellular and environmental inputs to the plant circadian clock. Plant Physiol. 148, 293–303. doi: 10.1104/pp.108.123901
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Koornneef, M., Alonso-Blanco, C., Blankestijn-de Vries, H., Hanhart, C. J., and Peeters, A. J. (1998). Genetic interactions among late-flowering mutants of Arabidopsis. Genetics 148, 885–892.
Koornneef, M., Hanhart, C. J., and van der Veen, J. H. (1991). A genetic and physiological analysis of late flowering mutants in Arabidopsis thaliana. Mol. Gen. Genet. 229, 57–66. doi: 10.1007/BF00264213
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kubota, A., Kita, S., Ishizaki, K., Nishihama, R., Yamato, K. T., and Kohchi, T. (2014). Co-option of a photoperiodic growth-phase transition system during land plant evolution. Nat. Commun. 5:3668. doi: 10.1038/ncomms4668
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kurepa, J., Smalle, J., Mantagu, M. V., and Inzé, D. (1998a). Oxidative stress tolerance and longevity in Arabidopsis: the late flowering mutant GIGANTEA is tolerant to paraquat. Plant J. 14, 759–764. doi: 10.1046/j.1365-313x.1998.00168.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kurepa, J., Smalle, J., Van Montagu, M., and Inzé, D. (1998b). Polyamines and paraquat toxicity in Arabidopsis thaliana. Plant Cell Physiol. 39, 987–992. doi: 10.1093/oxfordjournals.pcp.a029463
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Li, F., Zhang, X., Hu, R., Wu, F., Ma, J., Meng, Y.,et al. (2013). Identification and molecular characterization of FKF1 and GI homologous genes in soybean. PLoS ONE 8:e79036. doi: 10.1371/journal.pone.0079036
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Liang, H., Barakat, A., Schlarbaum, S. E., Mandoli, D. F., and Carlson, J. E. (2010). Comparison of gene order of GIGANTEA loci in yellow-poplar, monocots, and eudicots. Genome 53, 533–544. doi: 10.1139/g10-031
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Liew, L. C., Hecht, V., Laurie, R. E., Knowles, C. L., Vander Schoor, J. K., Macknight, R. C.,et al. (2009). DIE NEUTRALIS and LATE BLOOMER 1 contribute to regulation of the pea circadian clock. Plant Cell 21, 3198–3211. doi: 10.1105/tpc.109.067223
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Locke, J. C., Kozma-Bognár, L., Gould, P. D., Fehér, B., Kevei, E., Nagy, F.,et al. (2006). Experimental validation of a predicted feedback loop in the multi-oscillator clock of Arabidopsis thaliana. Mol. Syst. Biol. 2, 1–6. doi: 10.1038/msb4100102
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Locke, J. C., Southern, M. M., Kozma-Bognár, L., Hibberd, V., Brown, P. E., Turner, M. S.,et al. (2005). Extension of a genetic network model by iterative ecperimentation and mathematical analysis. Mol. Syst. Biol. 1:2005.0013. doi: 10.1038/msb4100018
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
López-Torrejón, G., Guerra, D., Catalá, R., Salinas, J., and del Pozo, J. C. (2013). Identification of SUMO targets by a novel proteomic approach in plants. J. Integr. Plant Biol. 55, 96–107. doi: 10.1111/jipb.12012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lu, S. X., Webb, C. J., Knowles, S. M., Kim, S. H. J., Wang, Z., and Tobin, E. M. (2012). CCA1 and ELF3 interact in the control of hypocotyl length and flowering time in Arabidopsis. Plant Physiol. 158, 1079–1088. doi: 10.1104/pp.111.189670
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Martin-Tryon, E. L., Kreps, J. A., and Harmer, S. L. (2007). GIGANTEA acts in blue light signaling and has biochemically separable roles in circadian clock and flowering time regulation. Plant Physiol. 143, 473–486. doi: 10.1104/pp.106.088757
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Martinez-Zapater, J. M., and Somerville, C. R. (1990). Effect of light quality and vernalization on late-flowering mutants of Arabidopsis thaliana. Plant Physiol. 92, 770–776. doi: 10.1104/pp.92.3.770
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Miller, T. A., Muslin, E. H., and Dorweiler, J. E. (2008). A maize CONSTANS-like gene, conz1, exhibits distinct diurnal expression patterns in varied photoperiods. Planta 227, 1377–1388. doi: 10.1007/s00425-008-0709-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Miwa, K., Serikawa, M., Suzuki, S., Kondo, T., and Oyama, T. (2006). Conserved expression profiles of circadian clock-related genes in two Lemna species showing long-day and short-day photoperiodic flowering responses. Plant Cell Physiol. 47, 601–612. doi: 10.1093/pcp/pcj027
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mizoguchi, T., Wheatley, K., Hanzawa, Y., Wright, L., Mizoguchi, M., Song, H. R.,et al. (2002). LHY and CCA1 are partially redundant genes required to maintain circadian rhythms in Arabidopsis. Dev. Cell 2, 629–641. doi: 10.1016/S1534-5807(02)00170-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mizoguchi, T., Wright, L., Fujiwara, S., Cremer, F., Lee, K., Onouchi, H.,et al. (2005). Distinct roles of GIGANTEA in promoting flowering and regulating circadian rhythms in Arabidopsis. Plant Cell 17, 2255–2270. doi: 10.1105/tpc.105.033464
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mizuno, T., Nomoto, Y., Oka, H., Kitayama, M., Takeuchi, A., Tsubouchi, M.,et al. (2014). Ambient temperature signal feeds into the circadian clock transcriptional circuitry through the EC night-time repressor in Arabidopsis thaliana. Plant Cell Physiol. 55, 958–976. doi: 10.1093/pcp/pcu030
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nakamichi, N., Kita, M., Niinuma, K., Ito, S., Yamashino, T., Mizoguchi, T.,et al. (2007). Arabidopsis clock-associated pseudo-response regulators PRR9, PRR7 and PRR5 coordinately and positively regulate flowering time through the canonical CONSTANS-dependent photoperiodic pathway. Plant Cell Physiol. 48, 822–832. doi: 10.1093/pcp/pcm056
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ni, Z., Kim, E. D., Ha, M., Lackey, E., Liu, J., Zhang, Y.,et al. (2009). Altered circadian rhythms regulate growth vigour in hybrids and allopolyploids. Nature 457, 327–331. doi: 10.1038/nature07523
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nusinow, D. A., Helfer, A., Hamilton, E. E., King, J. J., Imaizumi, T., Schultz, T. F.,et al. (2011). The ELF4-ELF3-LUX complex links the circadian clock to diurnal control of hypocotyl growth. Nature 475, 398–402. doi: 10.1038/nature10182
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Oliverio, K. A., Crepy, M., Martin-Tryon, E. L., Milich, R., Harmer, S. L., Putterill, J.,et al. (2007). GIGANTEA regulates phytochrome A-mediated photomorphogenesis independently of its role in the circadian clock. Plant Physiol. 144, 495–502. doi: 10.1104/pp.107.097048
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Onai, K., Okamoto, K., Nishimoto, H., Morioka, C., Hirano, M., Kami-ike, N.,et al. (2004). Large-scale screening of Arabidopsis circadian clock mutants by a high-throughput real-time bioluminescence monitoring system. Plant J. 40, 1–11. doi: 10.1111/j.1365-313X.2004.02191.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Paltiel, J., Amin, R., Gover, A., Ori, N., and Samach, A. (2006). Novel roles for GIGANTEA revealed under environmental conditions that modify its expression in Arabidopsis and Medicago truncatula. Planta 224, 1255–1268. doi: 10.1007/s00425-006-0305-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Park, D. H., Somers, D. E., Kim, Y. S., Choy, Y. H., Lim, H. K., Soh, M. S.,et al. (1999). Control of circadian rhythms and photoperiodic flowering by the Arabidopsis GIGANTEA gene. Science 285, 1579–1582. doi: 10.1126/science.285.5433.1579
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Park, H. J., Kim, W., and Yun, D. (2013). A role for GIGANTEA: keeping the balance between flowering and salinity stress tolerance. Plant Signal. Behav. 8:e24820. doi: 10.4161/psb.24820
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Penfield, S., and Hall, A. (2009). A role for multiple circadian clock genes in the response to signals that break seed dormancy in Arabidopsis. Plant Cell 21, 1722–1732. doi: 10.1105/tpc.108.064022
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Qian, H., Han, X., Peng, X., Lu, T., Liu, W., and Fu, Z. (2014). The circadian clock gene regulatory module enantioselectively mediates imazethapyr-induced early flowering in Arabidopsis thaliana. J. Plant Physiol. 171, 92–98. doi: 10.1016/j.jplph.2013.11.011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Riboni, M., Galbiati, M., Tonelli, C., and Conti, L. (2013). GIGANTEA enables drought escape response via abscisic acid-dependent activation of the florigens and SUPPRESSOR OF OVEREXPRESSION OF CONSTANS. Plant Physiol. 162, 1706–1719. doi: 10.1104/pp.113.217729
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Richardson, K., Fowler, S., Pullen, C., Skelton, C., Morris, B., and Putterill, J. (1998). T-DNA tagging of a flowering-time gene and improved gene transfer by in planta transformation of Arabidopsis. Aust. J. Plant Physiol. 25, 125–130. doi: 10.1071/PP97109
Rousset, M., Bonnin, I., Remoué, C., Falque, M., Rhoné, B., Veyrieras, J. B.,et al. (2011). Deciphering the genetics of flowering time by an association study on candidate genes in bread wheat (Triticum aestivum L.). Theor. Appl. Genet. 123, 907–926. doi: 10.1007/s00122-011-1636-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rutitzky, M., Ghiglione, H. O., Curá, J. A., Casal, J. J., and Yanovsky, M. J. (2009). Comparative genomic analysis of light-regulated transcripts in the Solanaceae. BMC Genomics 10:60. doi: 10.1186/1471-2164-10-60
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Salomé, P. A., Xie, Q., and McClung, C. R. (2008). Circadian timekeeping during early Arabidopsis development. Plant Physiol. 147, 1110–1125. doi: 10.1104/pp.108.117622
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sawa, M., and Kay, S. A. (2011). GIGANTEA directly activates flowering locus T in Arabidopsis thaliana. Proc. Natl. Acad. Sci. U.S.A. 108, 11698–11703. doi: 10.1073/pnas.1106771108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sawa, M., Nusinow, D. A., Kay, S. A., and Imaizumi, T. (2007). FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science 318, 261–265. doi: 10.1126/science.1146994
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schnable, J. C., Springer, N. M., and Freeling, M. (2011). Differentiation of the maize subgenomes by genome dominance and both ancient and ongoing gene loss. Proc. Natl. Acad. Sci. U.S.A. 108, 4069–4074. doi: 10.1073/pnas.1101368108
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Serikawa, M., Miwa, K., Kondo, T., and Oyama, T. (2008). Functional conservation of clock-related genes in flowering plants: overexpression and RNA interference analyses of the circadian rhythm in the monocotyledon Lemna gibba. Plant Physiol. 146, 1952–1963. doi: 10.1104/pp.107.114611
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Somers, D. E. (1999). The physiology and molecular bases of the plant circadian clock. Plant Physiol. 121, 9–19. doi: 10.1104/pp.121.1.9
Sothern, R. B., Tseng, T. S., Orcutt, S. L., Olszewski, N. E., and Koukkari, W. L. (2002). GIGANTEA and SPINDLY genes linked to the clock pathway that controls circadian characteristics of transpiration in Arabidopsis. Chronobiol. Int. 19, 1005–1022. doi: 10.1081/CBI-120015965
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Stoelzle, S., Kagawa, T., Wada, M., Hedrich, R., and Dietrich, P. (2003). Blue light activates calcium-permeable channels in Arabidopsis mesophyll cells via the phototropin signaling pathway. Proc. Natl. Acad. Sci. U.S.A. 100, 1456–1461. doi: 10.1073/pnas.0333408100
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Suárez-López, P., Wheatley, K., Robson, F., Onouchi, H., Valverde, F., and Coupland, G. (2001). CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 410, 1116–1120. doi: 10.1038/35074138
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Swarup, K., Alonso-Blanco, C., Lynn, J. R., Michaels, S. D., Amasino, R. M., Koornneef, M.,et al. (1999). Natural allelic variation identifies new genes in the Arabidopsis circadian system. Plant J. 20, 67–77. doi: 10.1046/j.1365-313X.1999.00577.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Swigonová, Z., Lai, J., Ma, J., Ramakrishna, W., Llaca, V., Bennetzen, J. L.,et al. (2004). Close split of Sorghum and maize genome progenitors. Genome Res. 14, 1916–1923. doi: 10.1101/gr.2332504
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tóth, R., Gerding-Reimers, C., Deeks, M. J., Menninger, S., Gallegos, R. M., Tonaco, I. A.,et al. (2012). Prieurianin/endosidin 1 is an actin-stabilizing small molecule identified from a chemical genetic screen for circadian clock effectors in Arabidopsis thaliana. Plant J. 71, 338–352. doi: 10.1111/j.1365-313X.2012.04991.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tseng, T., Salomé, P. A., McClung, C. R., and Olszewski, N. E. (2004). SPINDLY and GIGANTEA interact and act in Arabidopsis thaliana pathways involved in light responses, flowering, and rhythms in cotyledon movements. Plant Cell 16, 1550–1563. doi: 10.1105/tpc.019224
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Usadel, B., Bläsing, O. E., Gibon, Y., Retzlaff, K., Höhne, M., Günther, M.,et al. (2008). Global transcript levels respond to small changes of the carbon status during progressive exhaustion of carbohydrates in Arabidopsis rosettes. Plant Physiol. 146, 1834–1861. doi: 10.1104/pp.107.115592
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
von Arnim, A. G., and Deng, X. W. (1994). Light inactivation of Arabidopsis photomorphogenic repressor COP1 involves a cell-specific regulation of its nucleocytoplasmic partitioning. Cell 79, 1035–1045. doi: 10.1016/0092-8674(94)90034-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Watanabe, S., Xia, Z., Hideshima, R., Tsubokura, Y., Sato, S., Yamanaka, N.,et al. (2011). A map-based cloning strategy employing a residual heterozygous line reveals that the GIGANTEA gene is involved in soybean maturity and flowering. Genetics 188, 395–407. doi: 10.1534/genetics.110.125062
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wollenberg, A. C., Strasser, B., Cerdán, P. D., and Amasino, R. M. (2008). Acceleration of Flowering during shade avoidance in Arabidopsis alters the balance between FLOWERING LOCUS C-mediated repression and photoperiodic induction of flowering. Plant Physiol. 148, 1681–1694. doi: 10.1104/pp.108.125468
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Woods, D. P., Ream, T. S., Minevich, G., Hobert, O., and Amasino, R. M. (2014). PHYTOCHROME C is an essential light receptor for photoperiodic flowering in the temperate grass Brachypodium distachyon. Genetics 198, 397–408. doi: 10.1534/genetics.114.166785
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wu, J., Wang, Y., and Wu, S. (2008). Two new clock proteins, LWD1 and LWD2, regulate Arabidopsis photoperiodic flowering. Plant Physiol. 148, 948–959. doi: 10.1104/pp.108.124917
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wuriyanghan, H., Zhang, B., Cao, W. H., Ma, B., Lei, G., Liu, Y. F.,et al. (2009). The ethylene receptor ETR2 delays floral transition and affects starch accumulation in rice. Plant Cell 21, 1473–1494. doi: 10.1105/tpc.108.065391
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yu, J., Rubio, V., Lee, N., Bai, S., Lee, S., Kim, S.,et al. (2008). COP1 and ELF3 control circadian function and photoperiodic flowering by regulating GI stability. Mol. Cell 32, 617–630. doi: 10.1016/j.molcel.2008.09.026
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zakhrabekova, S., Gough, S. P., Braumann, I., Müller, A. H., Lundqvist, J., Ahmann, K.,et al. (2012). Induced mutations in circadian clock regulator Mat-a facilitated short season adaptation and range extension in cultivated barley. Proc. Natl. Acad. Sci. U.S.A. 109, 4326–4331. doi: 10.1073/pnas.1113009109
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhao, X. Y., Liu, M. S., Li, J. R., Guan, C. M., and Zhang, X. S. (2005). The wheat TaGI1, involved in photoperiodic flowering, encodes an Arabidopsis GI ortholog. Plant Mol. Biol. 58, 53–64. doi: 10.1007/s11103-005-4162-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zobell, O., Coupland, G., and Reiss, B. (2005). The family of CONSTANS-like genes in Physcomitrella patens. Plant Biol. (Stuttg.) 7, 266–275. doi: 10.1055/s-2005-865621
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: GIGANTEA, flowering time regulation, circadian clock control, GI mutants, light signaling
Citation: Mishra P and Panigrahi KC (2015) GIGANTEA – an emerging story. Front. Plant Sci. 6:8. doi: 10.3389/fpls.2015.00008
Received: 30 June 2014; Accepted: 06 January 2015;
Published online: 26 January 2015.
Edited by:
George Coupland, Max Planck Society, GermanyCopyright © 2015 Mishra and Panigrahi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Kishore C. Panigrahi, Plant Science Lab, School of Biological Sciences, National Institute of Science Education and Research, IOP campus, Sachivalaya Marg, P.O. Sainik School, Bhubaneshwar 751005, Orissa, India e-mail: panigrahi@niser.ac.in