 Christian Falter†
Christian Falter† Dorothea Ellinger
Dorothea Ellinger René Heim
René Heim Christian A. Voigt
Christian A. Voigt- Phytopathology and Biochemistry, Biocenter Klein Flottbek, University of Hamburg, Hamburg, Germany
The outwardly directed cell wall and associated plasma membrane of epidermal cells represent the first layers of plant defense against intruding pathogens. Cell wall modifications and the formation of defense structures at sites of attempted pathogen penetration are decisive for plant defense. A precise isolation of these stress-induced structures would allow a specific analysis of regulatory mechanism and cell wall adaption. However, methods for large-scale epidermal tissue preparation from the model plant Arabidopsis thaliana, which would allow proteome and cell wall analysis of complete, laser-microdissected epidermal defense structures, have not been provided. We developed the adhesive tape – liquid cover glass technique (ACT) for simple leaf epidermis preparation from A. thaliana, which is also applicable on grass leaves. This method is compatible with subsequent staining techniques to visualize stress-related cell wall structures, which were precisely isolated from the epidermal tissue layer by laser microdissection (LM) coupled to laser pressure catapulting. We successfully demonstrated that these specific epidermal tissue samples could be used for quantitative downstream proteome and cell wall analysis. The development of the ACT for simple leaf epidermis preparation and the compatibility to LM and downstream quantitative analysis opens new possibilities in the precise examination of stress- and pathogen-related cell wall structures in epidermal cells. Because the developed tissue processing is also applicable on A. thaliana, well-established, model pathosystems that include the interaction with powdery mildews can be studied to determine principal regulatory mechanisms in plant–microbe interaction with their potential outreach into crop breeding.
Introduction
The plant cell wall and its underlying plasma membrane are required to perceive environmental changes like biotic and mechanic stresses and also represent the first layers of defense to invading pathogens. In this regard, cells of the epidermal tissue have a predominant function as they form the outer barrier and are first in interacting with the surrounding environment. Epidermal plant cells usually differ in shape, function and physiological reactions to other plant tissues. Therefore, a specific analysis of separated epidermal tissue can provide new insights especially into the early regulation and organization of stress-related changes and adaption. This is of special interest in the field of plant–microbe interactions where cell wall thickenings, so-called papillae, are one of the most prominent and long-studied responses to invading pathogens in epidermal cells (de Bary, 1863). They contain the (1,3)-β-glucan callose as one of the most common chemical constituent, but also proteins (e.g., peroxidases and thionins), phenolics, and other putative antimicrobial and antifungal constituents (Aist and Williams, 1971; Sargent et al., 1973; Mercer et al., 1974; Sherwood and Vance, 1976; Mims et al., 2000). A precise, quantitative analysis of the composition of these stress-related structures and associated proteins would help to elucidate regulatory mechanisms involved in their organization and components required for establishing resistance to pathogens. However, conventional sample preparation from infected tissues, e.g., complete leaves after fungal infection, results in a relative strong dilution of the targeted stress-related defense structures because they only affect a relatively small part of the whole tissue. This is a major restriction in a time-resolved and plant line- or mutant-specific quantification of altered cell wall components, papillae constituents, and associated proteins.
To overcome these general limitations in sample preparation from tissues, laser microdissection (LM) techniques have been developed to separate specific cells or cellular structures from the surrounding tissue by using a focused laser (Emmert-Buck et al., 1996). After its initial application in medical research (Emmert-Buck et al., 1996; Fend and Raffeld, 2000; Wulfkuhle et al., 2001), LM has also been established in plant research to perform gene expression analyses from specific plant cell types (Asano et al., 2002; Nakazono et al., 2003; Casson et al., 2005), including epidermal cells (Inada and Wildermuth, 2005), and tissue selection for down-stream metabolic profiling (Schad et al., 2005b; Peukert et al., 2012), fluorometric assays (Jasik et al., 2011), or chemical analysis (Nakashima et al., 2008). Also in the area of plant–microbe interactions, an application of LM has been successfully demonstrated for the isolation of cells at infection sites and subsequent gene expression analyses (Chandran et al., 2010); and a growing number of reports about proteome analyses from LM-collected plant samples has been published (Fiorilli et al., 2012).
Despite the relatively large number of published LM studies including plant tissues, none of them provided a suitable protocol for our approach to separate stress-induced cell wall structures in epidermal cells for subsequent proteome and carbohydrate analysis in an efficient way. Methods described for LM of plant samples predominately include cross sections of either cryofixed of paraffin embedded tissue. Whereas paraffin embedding can be problematic for subsequent protein analysis (Ahram et al., 2003), cryofixation was shown to be compatible with subsequent protein analysis (Schad et al., 2005a), but is a relative elaborate method and difficult to apply on relative soft tissue, like leaf tissue from Arabidopsis thaliana. Therefore, we aimed to establish a LM method that would be relatively easy to operate and would not require tissue fixation. In addition, we wanted to avoid tissue cross section as early stage epidermal cell wall alterations, like papillae, would be difficult to identify, extending sample acquisition, and increasing possible protein degradation. A main prerequisite of our LM method development was to ensure an application on papillae generated in host epidermal leaf cells of the model pathosystem A. thaliana – powdery mildew, which would allow to study different mutants showing papillae formation and penetration resistance (Jacobs et al., 2003; Nishimura et al., 2003; Ellinger et al., 2013). Therefore, a primary target of our study was to develop an efficient method to isolate the plant’s adaxial epidermal cell layer containing localized, pathogen-induced cell wall structures. To reduce of sample contamination, we used LM coupled with laser pressure catapulting (LMPC) for sample collection (Meimberg et al., 2003) as a basic method in our study.
Materials and Methods
Biological Material
Arabidopsis thaliana (wild-type Columbia) as well as the powdery mildew Golovinomyces cichoracearum (strain UCSC1) used for leaf inoculation were cultivated and used as described by Stein et al. (2006). The cultivation of Brachypodium distachyon (inbred line Bd21) followed the description from Blümke et al. (2014). For all experiments, leaves from 4-weeks-old plants were used.
Preparation of Single-Layered Epidermal Tissue by the Adhesive Tape – Liquid Cover Glass Technique
For the preparation of single layered epidermal leaf tissue, we developed a combined adhesive tape – liquid cover glass technique (ACT). First, microscope slides were coated with liquid cover glass (Carl Zeiss, Austria). To optimize adhesive surface capacity, slides were spray-coated with liquid cover glass, excess liquid removed, and after 2 min of drying again spray-coated. In case of a non-uniform drying of the liquid cover glass coat, which might impair epidermis preparation quality, a short heating of the coated slide at 30°C promoted the formation of an optimal adhesive surface. After a final drying period of 2–3 min, A. thaliana and B. distachyon leaf sections were placed with their adaxial side on the coated microscope slide. For the preparation of A. thaliana leaf sections, the petiole and about 1–2 mm of the adjacent part of the lamina were cut off; B. distachyon leaves were cut into 1 cm sections. Leaf sections on coated microscope slides were then laminated with a self-adhesive tape (Tesafilm transparent, product no. 57370, solvent-free adhesive on polypropylene foil, Tesa, Germany). During this procedure, the formation of air pockets should be avoided to obtain optimal results of epidermis preparation, which was achieved by a fast tear-off of the tape (Figure 1A). In case of an incomplete epidermis preparation, additional tear-offs of the adhesive tape containing the remaining leaf tissue on previously unused positions of the coated microscope slide allowed a preparation of the remaining epidermis (Figure 1B). ACT-prepared epidermal tissue layers were dried in a desiccator for 24 h to prepare samples for a long-term storage.
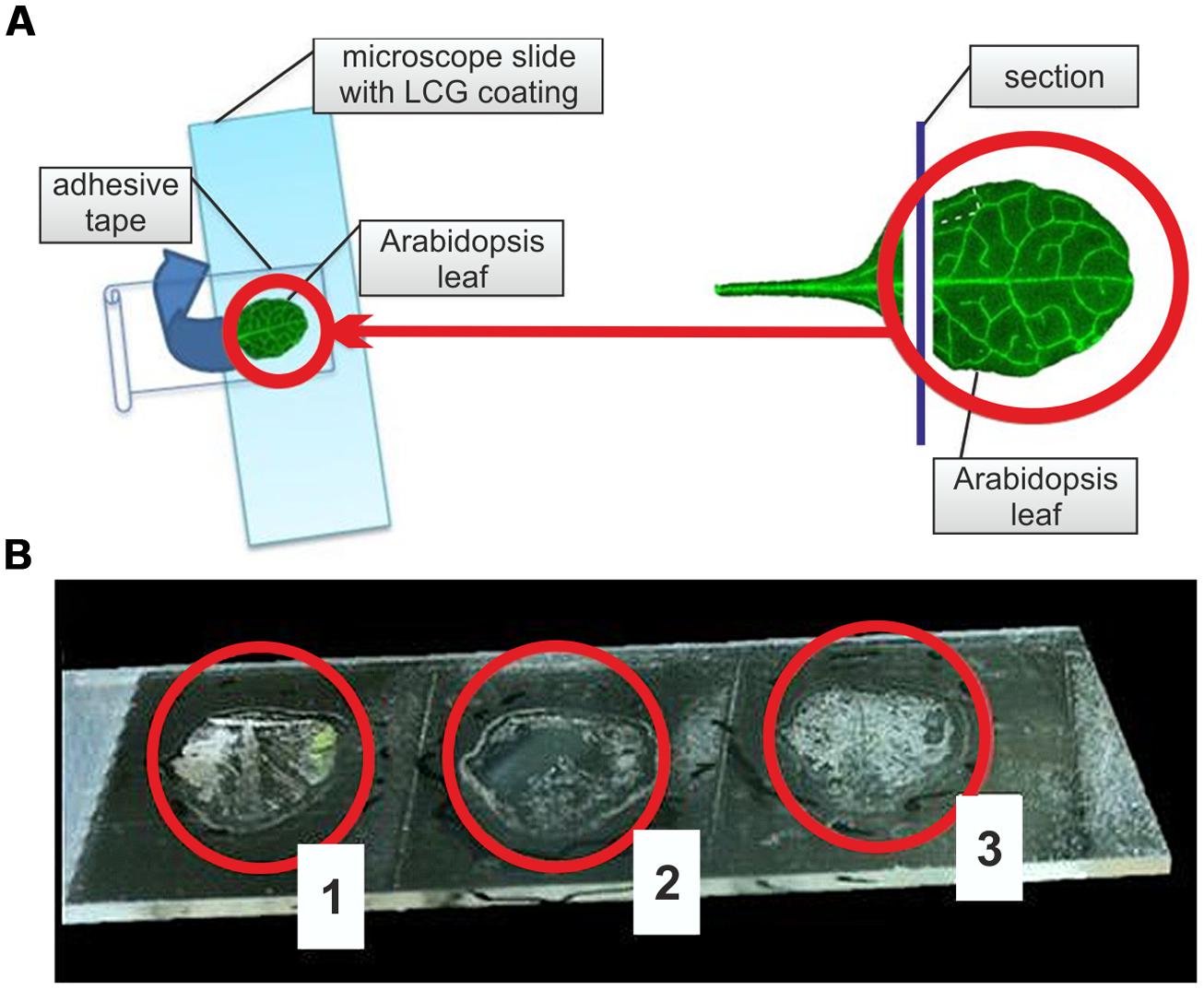
FIGURE 1. Adhesive tape – liquid cover glass technique (ACT) for adaxial leaf epidermis preparation. (A) Schematic overview of the application of ACT on Arabidopsis thaliana rosette leaves by sandwiching sectioned leaves between a microscope slide coated with liquid cover glass (LCG) and an adhesive tape. Blue arrow indicates direction of adhesive tape tear-off. Right panel demonstrates leaf sectioning before ACT. (B) Result of ACT preparation of adaxial A. thaliana leaf epidermis after three consecutive tear-offs of the same sample (1–3) on a LCG-coated microscope slide.
Staining of Epidermal Tissue
To visualize pathogen-induced cell wall modifications, which contained the (1,3)-β-glucan callose, ACT-prepared epidermal tissue was stained with aniline blue [0.01% (w/v) in 150 mM K2HPO4, pH 9]. Small droplets of the aniline blue solution were directly placed on the tissue. During pipetting of the staining solution, the tissue should not float off because this would prevent a subsequent laser capture microdissection. Evaporation of water and drying of the tissue was done in a dark compartment for 16 h at room temperature (22–24°C) without humidity control (room humidity ranging from 40 to 60%). Aniline blue stained-cell wall structures of ACT-prepared epidermis were visualized via epifluorescence microscopy (Stein et al., 2006). Additional washing of the stained tissue samples was not required.
Laser Capture Microdissection
For LM of ACT-prepared unstained or stained epidermal tissue, a PALM MicroBeam LM system (Carl Zeiss, Germany) was used, which was coupled with LMPC for sample collection. Typical settings for microdissection were a laser focus of 66% and a laser power of 32%, whereas the focus was re-adjusted to 64%, and the power to 62% during LMPC using the system’s operating software PALM Robosoftware V4.3 (Carl Zeiss, Germany). Optimal results were achieved by calibrating laser focus, laser power, and LMPC prior each single application. To establish LMPC from ACT-prepared epidermal tissue samples, disks with a diameter of 30 μm were randomly marked with the “close cut” function of the PALM Robosoftware. In case of aniline blue-stained samples, disks contained fluorescent regions that indicated pathogen-induced papillae. For optimal LMPC results, catapulting points were defined on a virtual circle with a diameter of 15 μm within the 30 μm disk using the “line auto-LPC” function of the PALM Robosoftware. Catapulted samples were collected in 40 μl denaturizing buffer [8 M urea, 2 M thiourea, 1 mM PMSF, 2% (v/v) protease inhibitor cocktail for plant cell and tissue extracts (Sigma–Aldrich, USA)], placed in the lid of collection tubes (Carl Zeiss). Collected samples were stored at -20°C until subsequent processing.
Protein Purification of Dissected Epidermal Tissue Samples
Laser pressure catapulting-prepared epidermal tissue samples were sonicated in denaturizing buffer in the ultrasonic bath Elmasonic S40 (Elma Hans Schmidtbauer, Germany) for 1 min at 37 kHz, which was repeated three-times, including a short cooling of the sample on ice between each application. Proteins were separated from cell wall material and cell fragments by ultra-centrifugation at 4°C, 100,000 g for 60 min. The supernatant was used for protein analysis, the pellet for cell wall analysis. Before protein analysis by liquid chromatography coupled with tandem spectrometry (LC-MS/MS) using a nanoLC-electrospray iontrap MS/MS system [ion-trap, XCT, Agilent Technologies, Core Facility for Mass Spectrometric Proteomics, University Medical Center Hamburg-Eppendorf, Germany; mass spectrometric peptide identification followed the description in Hildebrand et al. (2013)], protein concentration of the sample solution was increased by methanol/chloroform precipitation (Wessel and Flügge, 1984), followed by conventional enzymatic digestion with trypsin. Mascot MS/MS Ion Search (Matrix Science, USA) was used for LC-MS/MS data analysis with a peptide mass tolerance of ±10 Da and a fragment mass tolerance of ±4 Da.
Determination of Protein Concentration
After protein purification, protein concentrations of LMPC-prepared epidermal tissue samples were quantified using the fluorescence-based NanoOrange Protein Quantitation Kit (Life Technologies, USA) and the Synergy HT platereader (Biotek, USA) for detection. Procedures followed the manufacturer’s instructions.
Mono-carbohydrate Composition by HPAEC-PAD
For cell wall analysis of ACT- and LCM-prepared epidermal tissue, the non-cellulosic mono-carbohydrate composition was determined via high-performance anion exchange chromatography with pulsed amperometric detection (HPAEC-PAD). The alcohol insoluble residue (AIR) was prepared from pellets deriving from previous ultra-centrifugation of sonicated epidermis samples. After destarching, samples were treated with trifluoro acetic acid (TFA) before HPAEC-PAD analysis. All procedures for AIR preparation, destarching, TFA treatment, and HPAEC-PAD application followed the description in Ellinger et al. (2013).
Results and Discussion
Leaf Epidermis Preparation by Adhesive Tape – Liquid Cover Glass Technique
One of the main targets of our study was the establishment of a method for an easy preparation of the adaxial epidermis of plant leaves with a special focus on A. thaliana, characterized by relatively soft leaf tissue, which prevented an application of simple peeling techniques known from preparations of grass’ epidermal leaf tissue (Schultheiss et al., 2002). The developed ACT fulfilled our targets of an easy method of complete adaxial epidermis preparation without previous tissue fixation and applicability on soft tissues. The ACT method can be seen as further development of the perforated-tape epidermal detachment method, which was also applicable on A. thaliana adaxial epidermal tissue (Ibata et al., 2013), but not compatible with downstream LM applications. For epidermis preparation by ACT, we spray-coated microscope slides with liquid cover glass to generate an adhesive surface that is compatible with downstream LMPC applications (Moelans et al., 2011). After cutting off the petiole and lower part of the lamina, A. thaliana leaves were placed with their adaxial side on the coated microscope slide (Figure 1A). We used 1 cm leaf sections from the model grass B. distachyon as an additional reference for ACT epidermis preparation. Leaf sections were then laminated with an adhesive tape, sandwiching the plant sample between the coated microscope slide and the tape. A fast tear-off of the tape resulted in an epidermal cell layer, which was attached to the coated microscope slide. An additional tear-off of the adhesive tape containing the remaining leaf tissue on previously unused positions of the coated microscope slide allowed complete preparation of the epidermis (Figure 1B). However, three or more tear-offs of the same leaf sample increased the chance of a sample contamination with mesophyll tissue. Therefore, a microscopic examination of prepared epidermis sample was required to ensure sample collection only from regions with single epidermal cell layers. Figure 2 provides an overview about successful single-layered, adaxial epidermis preparation from A. thaliana and B. distachyon leaf sections, showing the typical lobed profile of A. thaliana and the rectangular profile of B. distachyon epidermal cells. In addition, the result of epidermis preparation revealed that not the complete epidermal cell layer was attached to the coated microscope slide, but predominantly the outward-oriented part of the cells (Figures 2A,C and 3A). This suggests that the cytosolic part of the epidermal cells was mainly removed during the tear-off of the adhesive tape. The observed effect of ACT promoted our efforts in analyzing the proteome of only stress-induced cell wall structures and associated plasma membrane fractions.
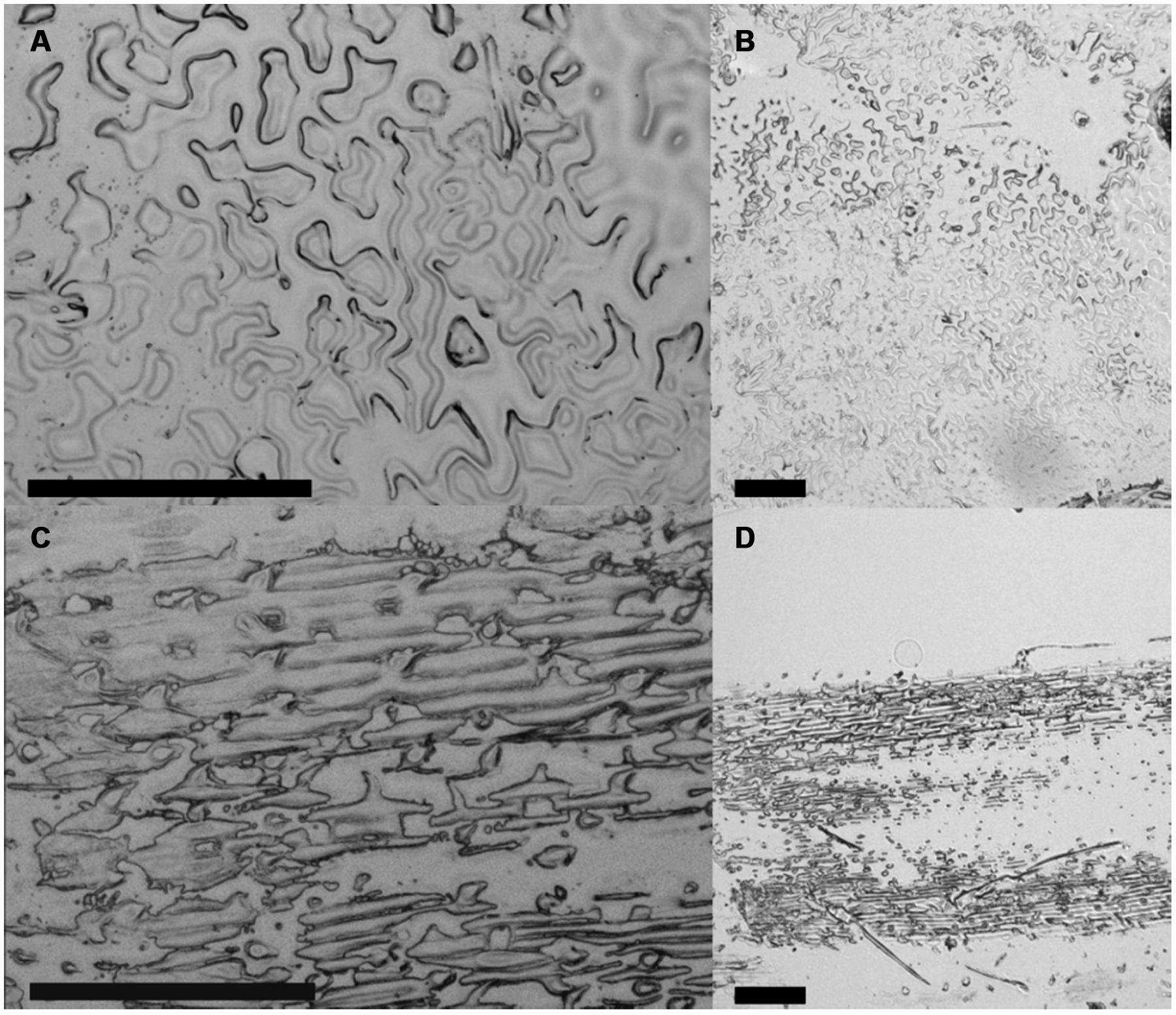
FIGURE 2. Microscopic analysis of ACT-prepared adaxial leaf epidermis. Epidermis preparation from 4-weeks-old (A,B) A. thaliana and (C,D) B. distachyon leaves using the ACT. Micrographs taken by the PALM MicroBeam laser microdissection (LM) system. Scale bars = 200 μm.

FIGURE 3. Dissection and catapulting of 30 μm-disks from ACT-prepared adaxial leaf epidermis. Application of LM coupled with laser pressure catapulting (LMPC) on ACT-prepared A. thaliana adaxial leaf epidermis. Epidermis (A) before and (B) after microdissection of a 30 μm disk; (C) removal of 30 μm-disk from epidermal tissue by LMPC. (D) ACT-prepared adaxial leaf epidermis 3 days post-inoculation with the powdery mildew Golovinomyces cichoracearum, additionally stained with aniline blue to visualize callose deposition. Red arrow indicates 30 μm-disk including pathogen-induced, callosic papillae (spots with light-blue fluorescence). (E,F) Bright-field micrographs of the same epidermis area as in (D,E) after microdissection and (F) after LMPC of a 30 μm-disk. PALM MicroBeam LM system used for LMPC and micrograph acquisition. Scale bars = 50 μm.
In the next step, we tested LMPC from ACT-prepared epidermal tissue choosing a disk size of 30 μm in diameter for dissection because this area would cover also enlarged papillae with extended callose deposition from powdery mildew-resistant A. thaliana lines (Ellinger et al., 2013). As expected from the relatively low thickness of the ACT-prepared epidermal samples (Figure 2), LMPC was not problematic (Figures 3A–C). Also additional staining of ACT-prepared epidermal samples with aniline blue, which is specific for (1,3)-β-glucan polymers (Evans et al., 1984), to visualize powdery mildew- induced, callose-containing papillae (Figure 3D) did not affect LMPC (Figures 3E,F). This proved the applicability of the developed ACT for isolating entire and stained stress-induced cell wall structures by LMPC from adaxial plant epidermal tissue.
Optimized laser settings for microdissection allowed a collection of up to 5000 disks per day if all targeted sample disks were first dissected and then catapulted and collected in the denaturizing sample buffer.
Proteome Analysis of LMPC-Prepared Epidermis Samples
After successfully establishing ACT preparation of epidermal tissue with subsequent LMPC, we tested whether the collected samples would generally allow a proteome analysis. Moreover, we also wanted to evaluate the appropriate amount of dissected disks, which would be required to perform significant comparable proteome analysis of different samples. Therefore, we randomly dissected 11,000, 35,000, and 50,000 disks from ACT-prepared A. thaliana adaxial epidermal leaf tissue (Figures 3A–C), which was equivalent to a total area of 7.8, 24.7, and 25.4 mm2, respectively. Total protein extracts from these samples yielded 0.69 [ ± 0.19] μg protein per 10,000 disks and were analyzed by liquid chromatography coupled with tandem spectrometry (LC-MS/MS). Results from ACT- and LMPC-prepared epidermal leaf samples were compared with control protein samples from whole A. thaliana leaf tissue.
Whereas LC-MS/MS only identified 14 individual proteins in the sample deriving from 11,000 disks (0.38 μg protein analyzed), which we considered not sufficient for significant comparative analysis, 47 proteins derived from the 35,000-disks and 54 proteins from the 50,000-disks protein samples (1.21 and 1.73 μg protein, respectively; Figure 4A). Even though these numbers of proteins were only about a sixth of the amount detected in the control protein samples (5 μg protein; Figure 4A), they allowed a comparative analysis to evaluate the efficiency of the ACT epidermis preparation in accumulating proteins from the plasma membrane. A qualitative analysis of the subcellular localization of the detected proteins revealed that this was especially effective in the 50,000-disks sample. We observed a relative reduction of proteins originating from the cytosol, mitochondria, and plastids, whereas the relative fraction of proteins deriving from membranes, and especially the plasma membrane, was strongly enriched (Figure 4B). These results supported our microscopic observation that ACT of epidermal tissue results in a relatively efficient removal of the cytosol and cellular organelles. However, this method was not as effective in removing the nucleus from ACT-prepared epidermal samples as the relative fraction of nucleus-localized proteins was expanded in ACT-prepared epidermis samples (Figure 4B). Because the requested effects in accumulating plasma membrane-localized proteins and reducing proteins from the cytosol and organelles was not as distinct in the 35,000-disks sample as in the 50,000-disks sample, we propose sample sizes of about 50,000 disks (∼25 mm2) to be appropriate for downstream proteome analysis by LC-MS/MS. The observed effect of ACT epidermis preparation in accumulating membrane proteins was not only restricted to a qualitative analysis but also reflected in a quantitative analysis. A Top 15 list of protein abundance (Table 1) revealed a quantitative predominance of chloroplast localized proteins like ribulose bisphosphate carboxylase in the control sample from whole leaf protein extracts whereas the LMPC-prepared 50,000-disks epidermis sample was enriched in membrane and nuclear proteins. Remarkably, a (1,3)-β-glucosidase, which is known to be anchored to the plasma membrane (Elortza et al., 2003), was most abundant (Table 1).
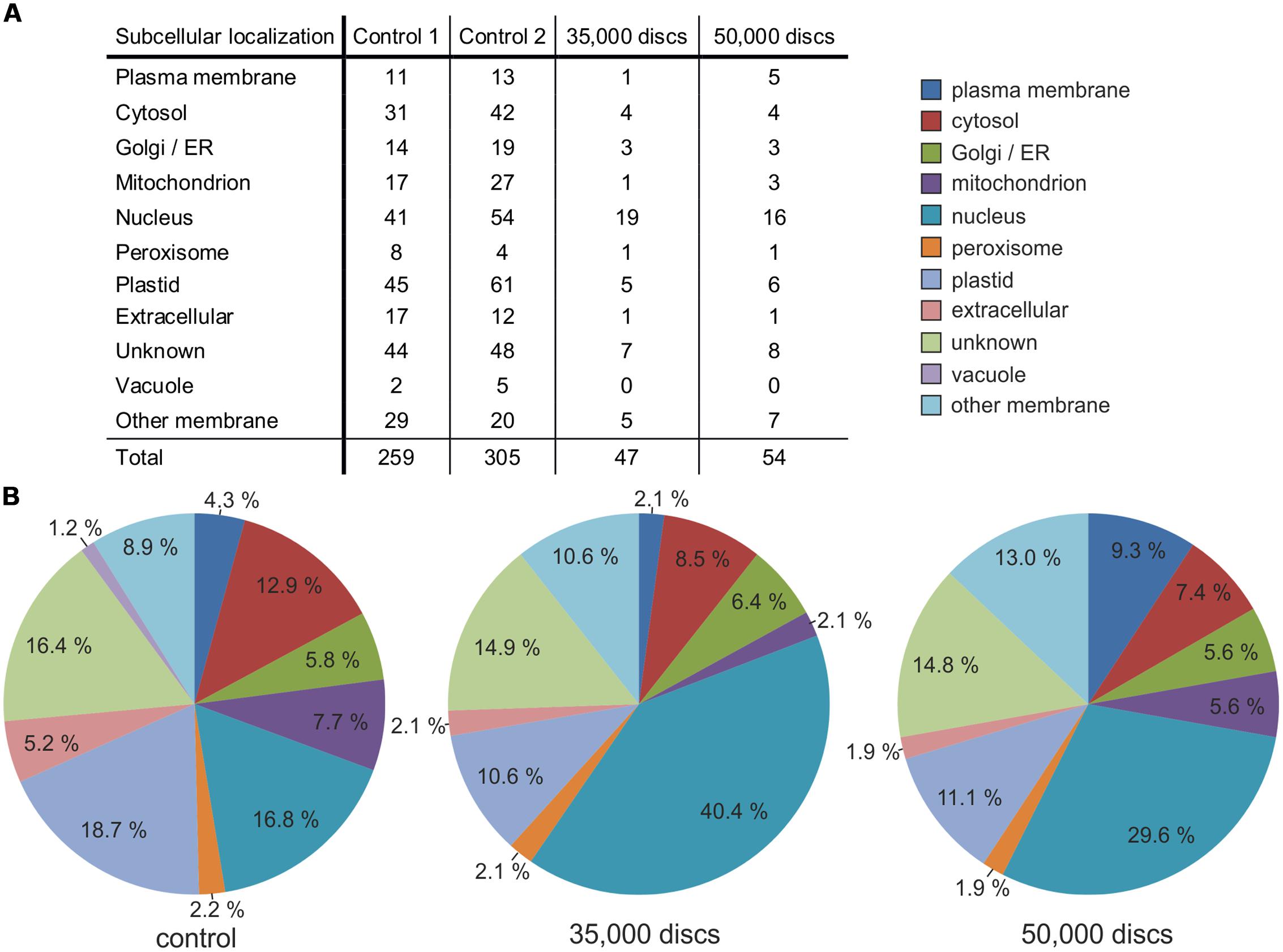
FIGURE 4. Subcellular localization of extracted proteins from ACT-/LMPC-prepared adaxial A. thaliana leaf epidermis. Epidermis preparation from 4-weeks-old A. thaliana leaves using the ACT with subsequent laser capture microdissection coupled with LMPC of 30 μm-disks; sample sizes: 35,000 and 50,000 disks. Protein extracts from whole leaf tissue served as control. Protein identity determined by LC-MS/MS. (A) Total number of different proteins and their subcellular localization in control and disk samples. (B) Relative share of each subcellular localization class based on data as shown in (A). Control indicates the mean value of controls 1 and 2. A biological replicate gave similar results.

TABLE 1. Protein abundance in control and ACT-/LMPC-prepared epidermal tissue samples (50,000 disks) from Arabidopsis thaliana leaves.
These results from LC-MS/MS analysis ACT- and LMPC-prepared epidermal leaf samples from A. thaliana clearly showed that this method approach is appropriate for proteome analysis of selected cell wall structures and associated membranes. However, whereas the total number of proteins detected in control samples was in a range that can be expected in complex proteome analyses (∼280–300 different proteins in 5 μg total protein sample; Mallick and Kuster, 2010) with the system used in this study, protein numbers were below this expectation in ACT- and LMPC-prepared epidermal leaf samples. This suggests possible protein degradation during ACT-preparation and LM of epidermal leaf samples, when proteases were not specifically inhibited by protease inhibitors. To avoid possible protein degradation at this stage of sample preparation, addition of protease inhibitors to the liquid cover glass might provide an adhesive film with protease-inhibiting properties.
Mono-carbohydrate Composition of LMPC-prepared Epidermis Samples
Besides proteome analysis, our target for developing a simple epidermis preparation method was to facilitate quantitative non-cellulosic mono-carbohydrate analysis of defined cell wall structures using high-performance anion exchange chromatography with pulsed amperometric detection (HPAEC-PAD). To test HPAEC-PAD on ACT-prepared epidermal leaf cell layers, we dissected 6,000 disks and extracted the mono-carbohydrates from the hydrolyzed hemicellulosic cell wall fraction. As a control, we applied the same method on LMPC samples from liquid cover glass-coated microscope slides without epidermis preparation.
In the 6,000-disks sample, we detected the mono-carbohydrates galactose, glucose, mannose, galacturonic, and glucoronic acid that are typical for A. thaliana cell walls (Ellinger et al., 2013) in such amounts that would allow quantification. Only residual amounts were detectable for rhamnose and arabinose (Figure 5). We also detected residual amounts of glucose as well as galacturonic and glucoronic acid in the control liquid cover glass sample (Figure 5). Because it is most likely that these mono-carbohydrate contaminations derive from liquid cover glass, it would not be possible to avoid this contaminations based on the developed ACT for epidermis preparation. Hence, it would be required to analyze the same sample size of liquid cover glass-coating in parallel to plant samples to facilitate precise mono-carbohydrate quantification. In this regard, we consider a sample size of 6,000 disks (∼8 mm2) as a lower limit for significant and precise mono-carbohydrate quantification.
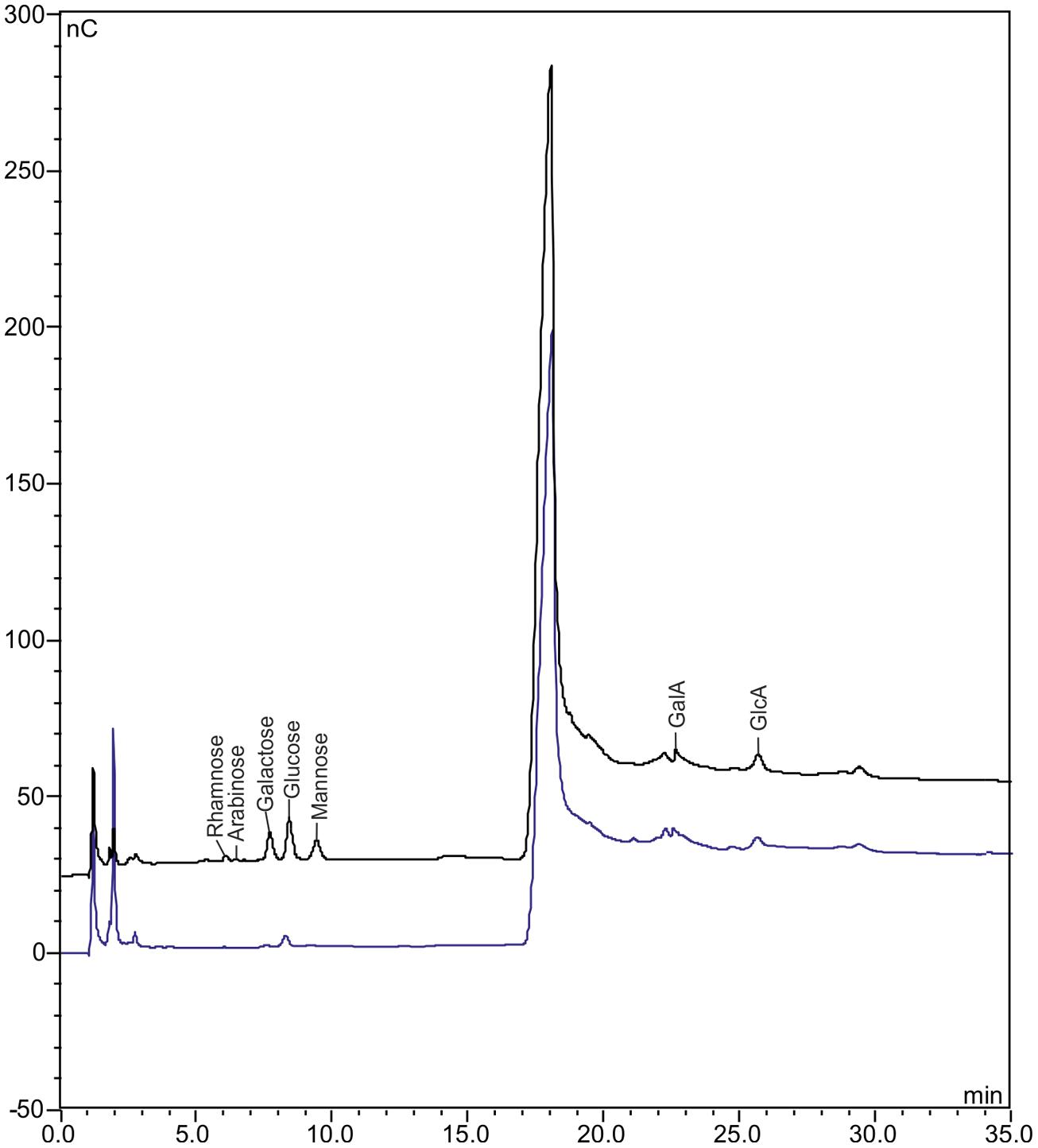
FIGURE 5. Chromatograms obtained by HPAEC-PAD of ACT-/LMPC-prepared adaxial A. thaliana leaf epidermis. Epidermis preparation from 4-weeks-old A. thaliana leaves using the ACT with subsequent laser capture microdissection coupled with LMPC of 30 μm-disks; sample size: 6,000 disks. Non-cellulosic mono-carbohydrate composition determined via high-performance anion exchange chromatography with pulsed amperometric detection (HPAEC-PAD). Black line: chromatogram of epidermal disk sample; blue line: chromatogram of control sample (6,000 liquid cover class disks, same treatment as epidermal disk sample). GalC, galacturonic acid; GlcA, glucuronic acid. A biological replicate gave similar results.
In addition to proteome analysis, the successful detection of hemicellulose-derived mono-carbohydrates clearly revealed that ACT- and LMPC-prepared epidermal leaf samples can be used for the quantitative analysis of the cell wall composition of defined cell wall structures using HPAEC-PAD.
Conclusion
The developed ACT allows a simple preparation of complete adaxial epidermal tissue layers from large-scale leaf sections. Because the developed method is also applicable on soft tissue, preparations from the most important model plant A. thaliana have been facilitated. ACT epidermis preparation neither requires tissue embedding nor cross sections. Therefore, complete, defined cell wall structures or areas of modification can be readily identified and isolated using LMPC. This is of special importance in plant–microbe interactions where pathogen-induced cell wall modifications are decisive for plant defense. We successfully demonstrated further processing of ACT- and LMPC-prepared epidermis samples in downstream applications for proteome analysis and carbohydrate composition, opening new perspectives in the quantitative analysis of stress-induced cell wall modification.
Author Contributions
CF, DE, BvH, RH, CV: conception and design of the research; CF, DE, BvH, RH: acquisition and analysis of the data; CF, DE, CV: interpretation of data; CF, DE, BvH, RH, CV: drafting the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
This work was supported by the German Federal Ministry of Education and Research (grant no. FKZ 0315521A, to CV).
References
Ahram, M., Flaig, M. J., Gillespie, J. W., Duray, P. H., Linehan, W. M., Ornstein, D. K., et al. (2003). Evaluation of ethanol-fixed, paraffin-embedded tissues for proteomic applications. Proteomics 3, 413–421. doi: 10.1002/pmic.200390056
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Aist, J. R., and Williams, P. H. (1971). The cytology and kinetics of cabbage root hair penetration by Plasmodiophora brassicae. Can. J. Bot. 49, 2023–2034. doi: 10.1139/b71-284
Asano, T., Masumura, T., Kusano, H., Kikuchi, S., Kurita, A., Shimada, H., et al. (2002). Construction of a specialized cDNA library from plant cells isolated by laser capture microdissection: toward comprehensive analysis of the genes expressed in the rice phloem. Plant J. 32, 401–408. doi: 10.1046/j.1365-313X.2002.01423.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Blümke, A., Sode, B., Ellinger, D., and Voigt, C. A. (2014). Reduced susceptibility to Fusarium head blight in Brachypodium distachyon through priming with the Fusarium mycotoxin deoxynivalenol. Mol Plant Pathol. doi: 10.1111/mpp.12203 [Epub ahead of print].
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Casson, S., Spencer, M., Walker, K., and Lindsey, K. (2005). Laser capture microdissection for the analysis of gene expression during embryogenesis of Arabidopsis. Plant J. 42, 111–123. doi: 10.1111/j.1365-313X.2005.02355.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chandran, D., Inada, N., Hather, G., Kleindt, C. K., and Wildermuth, M. C. (2010). Laser microdissection of Arabidopsis cells at the powdery mildew infection site reveals site-specific processes and regulators. Proc. Natl. Acad. Sci. U.S.A. 107, 460–465. doi: 10.1073/pnas.0912492107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
de Bary, A. (1863). Recherches sur le développement de quelques champignons parasites. Ann. Sci. Nat. Bot. Biol. Vég. 20, 5–148.
Ellinger, D., Naumann, M., Falter, C., Zwikowics, C., Jamrow, T., Manisseri, C., et al. (2013). Elevated early callose deposition results in complete penetration resistance to powdery mildew in Arabidopsis. Plant Physiol. 161, 1433–1444. doi: 10.1104/pp.112.211011
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Elortza, F., Nuhse, T. S., Foster, L. J., Stensballe, A., Peck, S. C., and Jensen, O. N. (2003). Proteomic analysis of glycosylphosphatidylinositol-anchored membrane proteins. Mol. Cell. Proteomics 2, 1261–1270. doi: 10.1074/mcp.M300079-MCP200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Emmert-Buck, M. R., Bonner, R. F., Smith, P. D., Chuaqui, R. F., Zhuang, Z., Goldstein, S. R., et al. (1996). Laser capture microdissection. Science 274, 998–1001. doi: 10.1126/science.274.5289.998
Evans, N. A., Hoyne, P. A., and Stone, B. A. (1984). Characteristics and specificity of the interaction of a fluorochrome from aniline blue (sirofluor) with polysaccharides. Carbohydr. Polym. 4, 215–230. doi: 10.1016/0144-8617(84)90012-2
Fend, F., and Raffeld, M. (2000). Laser capture microdissection in pathology. J. Clin. Pathol. 53, 666–672. doi: 10.1136/jcp.53.9.666
Fiorilli, V., Klink, V. P., and Balestrini, R. (2012). “Proteomic analyses of cells isolated by laser microdissection, integrative proteomics,” in Integrative Proteomics, ed. H.-C. Leung (Rijeka: InTech).
Hildebrand, D., Merkel, P., Eggers, L. F., and Schluter, H. (2013). Proteolytic processing of angiotensin-I in human blood plasma. PLoS ONE 8:e64027. doi: 10.1371/journal.pone.0064027
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ibata, H., Nagatani, A., and Mochizuki, N. (2013). Perforated-tape Epidermal Detachment (PED): a simple and rapid method for isolating epidermal peels from specific areas of Arabidopsis leaves. Plant Biotechnol. 30, 497–502. doi: 10.5511/plantbiotechnology.13.0903b
Inada, N., and Wildermuth, M. C. (2005). Novel tissue preparation method and cell-specific marker for laser microdissection of Arabidopsis mature leaf. Planta 221, 9–16. doi: 10.1007/s00425-004-1427-y
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jacobs, A. K., Lipka, V., Burton, R. A., Panstruga, R., Strizhov, N., Schulze-Lefert, P., et al. (2003). An Arabidopsis callose synthase, GSL5, is required for wound and papillary callose formation. Plant Cell 15, 2503–2513. doi: 10.1105/tpc.016097
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jasik, J., Schiebold, S., Rolletschek, H., Denolf, P., Van Adenhove, K., Altmann, T., et al. (2011). Subtissue-specific evaluation of promoter efficiency by quantitative fluorometric assay in laser microdissected tissues of rapeseed. Plant Physiol. 157, 563–573. doi: 10.1104/pp.111.180760
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mallick, P., and Kuster, B. (2010). Proteomics: a pragmatic perspective. Nat. Biotechnol. 28, 695–709. doi: 10.1038/nbt.1658
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Meimberg, H., Thalhammer, S., Brachmann, A., Muller, B., Eichacker, L. A., Heckl, W. M., et al. (2003). Selection of chloroplasts by laser microbeam microdissection for single-chloroplast PCR. Biotechniques 34, 1238–1243.
Mercer, P. C., Wood, R. K. S., and Greenwood, A. D. (1974). Resistance to anthracnose of French bean. Physiol. Plant Pathol. 4, 291–306. doi: 10.1016/0048-4059(74)90016-2
Mims, C. W., Copes, W. E., and Richardson, E. A. (2000). Ultrastructure of the penetration and infection of pansy roots by Thielaviopsis basicola. Phytopathology 90, 843–850. doi: 10.1094/PHYTO.2000.90.8.843
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Moelans, C. B., De Weger, R. A., and Van Diest, P. J. (2011). Amplification testing in breast cancer by multiplex ligation-dependent probe amplification of microdissected tissue. Methods Mol. Biol. 755, 107–118. doi: 10.1007/978-1-61779-163-5_9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nakashima, J., Chen, F., Jackson, L., Shadle, G., and Dixon, R. A. (2008). Multi-site genetic modification of monolignol biosynthesis in alfalfa (Medicago sativa): effects on lignin composition in specific cell types. New Phytol. 179, 738–750. doi: 10.1111/j.1469-8137.2008.02502.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nakazono, M., Qiu, F., Borsuk, L. A., and Schnable, P. S. (2003). Laser-capture microdissection, a tool for the global analysis of gene expression in specific plant cell types: identification of genes expressed differentially in epidermal cells or vascular tissues of maize. Plant Cell 15, 583–596. doi: 10.1105/tpc.008102
Nishimura, M. T., Stein, M., Hou, B. H., Vogel, J. P., Edwards, H., and Somerville, S. C. (2003). Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 301, 969–972. doi: 10.1126/science.1086716
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Peukert, M., Matros, A., Lattanzio, G., Kaspar, S., Abadia, J., and Mock, H. P. (2012). Spatially resolved analysis of small molecules by matrix-assisted laser desorption/ionization mass spectrometric imaging (MALDI-MSI). New Phytol. 193, 806–815. doi: 10.1111/j.1469-8137.2011.03970.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sargent, J. A., Tommerup, I. C., and Ingram, D. S. (1973). The penetration of a susceptible lettuce variety by the downy mildew fungus Bremia lactucae Regel. Physiol. Plant Pathol. 3, 231–239. doi: 10.1016/0048-4059(73)90085-4
Schad, M., Lipton, M. S., Giavalisco, P., Smith, R. D., and Kehr, J. (2005a). Evaluation of two-dimensional electrophoresis and liquid chromatography – tandem mass spectrometry for tissue-specific protein profiling of laser-microdissected plant samples. Electrophoresis 26, 2729–2738. doi: 10.1002/elps.200410399
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schad, M., Mungur, R., Fiehn, O., and Kehr, J. (2005b). Metabolic profiling of laser microdissected vascular bundles of Arabidopsis thaliana. Plant Methods 1:2. doi: 10.1186/1746-4811-1-2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schultheiss, H., Dechert, C., Kogel, K. H., and Huckelhoven, R. (2002). A small GTP-binding host protein is required for entry of powdery mildew fungus into epidermal cells of barley. Plant Physiol. 128, 1447–1454. doi: 10.1104/pp.010805
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sherwood, R. T., and Vance, C. P. (1976). Histochemistry of papillae formed in reed canarygrass leaves in response to noninfecting pathogenic fungi. Phytopathology 66, 503–510. doi: 10.1094/Phyto-66-503
Stein, M., Dittgen, J., Sanchez-Rodriguez, C., Hou, B. H., Molina, A., Schulze-Lefert, P., et al. (2006). Arabidopsis PEN3/PDR8, an ATP binding cassette transporter, contributes to nonhost resistance to inappropriate pathogens that enter by direct penetration. Plant Cell 18, 731–746. doi: 10.1105/tpc.105.038372
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wessel, D., and Flügge, U. I. (1984). A method for the quantitative recovery of protein in dilute solution in the presence of detergents and lipids. Anal. Biochem. 138, 141–143. doi: 10.1016/0003-2697(84)90782-6
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: laser microdissection, proteome analysis, cell wall, epidermis, plant–microbe interaction, infection structures, Arabidopsis thaliana
Citation: Falter C, Ellinger D, von Hülsen B, Heim R and Voigt CA (2015) Simple preparation of plant epidermal tissue for laser microdissection and downstream quantitative proteome and carbohydrate analysis. Front. Plant Sci. 6:194. doi: 10.3389/fpls.2015.00194
Received: 24 January 2015; Paper pending published 24 February 2015;
Accepted: 11 March 2015 Published online: 27 March 2015.
Edited by:
Basil J. Nikolau, Iowa State University, USAReviewed by:
Vania Michelotti, Consiglio per la Ricerca e la Sperimentazione in Agricoltura, ItalyZhao Su, The Penn State University, USA
Copyright © 2015 Falter, Ellinger, vonHülsen, Heim and Voigt. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian A. Voigt, Phytopathology and Biochemistry, Biocenter Klein Flottbek, University of Hamburg, Ohnhorststrasse 18, 22609 Hamburg, GermanyY2hyaXN0aWFuLnZvaWd0QHVuaS1oYW1idXJnLmRl
† These authors have contributed equally to this work.