 Alexandre Bourles1†
Alexandre Bourles1† Guillaume Pierre2,3
Guillaume Pierre2,3 Hamid Amir1Alizée Le Floc’h1Eleftherios Chalkiadakis4Valérie Médevielle1
Hamid Amir1Alizée Le Floc’h1Eleftherios Chalkiadakis4Valérie Médevielle1 Philippe Jourand5Philippe Michaud2Valérie Burtet-Sarramégna1
Philippe Jourand5Philippe Michaud2Valérie Burtet-Sarramégna1 Linda Guentas1*
Linda Guentas1*- 1Institut de Sciences Exactes et Appliquées, Université de la Nouvelle-Calédonie, Noumea, New Caledonia
- 2Institut Pascal, Université Clermont Auvergne, Centre National de la Recherche Scientifique (CNRS), Clermont Auvergne Institut National Polytechnique (INP), Clermont-Ferrand, France
- 3Institut Universitaire de France (IUF), Paris, France
- 4BIOTECAL, Marine Biotechnology Company, Nouméa, New Caledonia
- 5Institute of Research for Development (IRD), UMR Entropie, Université de la Réunion, Saint Denis, France
Paraburkholderia ultramafica STM10279T is a metal-tolerant rhizobacterium that promotes plant growth. It was isolated from the roots of Tetraria arundinaceae, a pioneer endemic tropical herb growing on ultramafic soils in New Caledonia. We have recently shown that the main mechanism of metal tolerance of P. ultramafica is related to the production of an acidic exopolysaccharide (EPS). To explore the potential role of this EPS in the plant’s environmental adaptation, we first elucidated its structure by employing a combination of chromatography and mass spectrometry techniques. These analyses revealed that the EPS is highly branched and composed of galactosyl (35.8%), glucosyl (33.2%), rhamnosyl (19.5%), mannosyl (7.2%), and glucuronosyl residues (4.4%), similar to the EPS of the Burkholderia cepacia complex known as cepacian. We subsequently conducted greenhouse experiments on Tetraria comosa plantlets inoculated with P. ultramafica or a solution of its EPS during transplanting onto ultramafic substrate. The data showed that the dry weight of T. comosa shoots was 2.5 times higher in the plants treated with the EPS compared to the unexposed plants. In addition, inductively coupled plasma–optical emission spectrometry (ICP-OES) analysis revealed that exposure to the EPS significantly increased Ca, Mg, K, and P uptake as well as K content in roots. In vitro experiments using the Pikovskaya method showed that the EPS was able to solubilize phosphorus. Consistent with the retention of metals in roots and a reduction in shoots, our data revealed a significant decrease in metal translocation factors (TFs) in the plants inoculated with the EPS. These results suggest a beneficial effect of the rhizobacterial EPS on plant growth and abiotic stress mitigation. In addition, the data suggest that the reduced levels of trace metals in plants exposed to P. ultramafica STM10279T are due to metal chelation by the EPS. Further investigations are needed to firmly demonstrate whether this EPS could be used as a biostimulant for plant growth and adaptation to ultramafic soils.
1 Introduction
Plant-associated microorganisms are adapted to a wide range of environments, and most have been shown to improve plant growth or health (de la Fuente Cantó et al., 2020). Among these, plant growth-promoting rhizobacteria (PGPRs) can directly enhance plant growth by producing phytohormones and increasing nutrient availability in the rhizosphere (Khatoon et al., 2020). PGPRs are also known to increase tolerance to biotic and abiotic stresses (Zahedi, 2021). Many PGPRs secrete mixtures of exopolysaccharides (EPSs) of high molecular weight in response to physiological stress (Morcillo and Manzanera, 2021). EPS plays an important role in cell protection and bacterial adhesion to solid surfaces (i.e., biofilm formation) and is involved in cell–cell interactions (Netrusov et al., 2023). Recently, Naseem et al. (2018) reviewed the role of EPS-producing rhizobacteria on drought tolerance, plant growth, root and shoot biomass, and the physico-chemical properties of soil. In addition, it has been shown that some EPSs bind metals in soil (Prabhu et al., 2019), which affects the solubility of metal phosphates and plant nutrition (Kumar and Shastri, 2017). Moreover, metal-resistant PGPRs have been shown to mitigate metal stress in plants by reducing the bioavailability of trace metals through complexation by their secreted EPSs (Gupta and Diwan, 2017; Etesami, 2018). This process is of particularly high relevance to the ultramafic soils of New Caledonia where PGP bacteria play a crucial role in providing metal resistance to endemic plants, enabling their survival and growth in soils that would otherwise be inhospitable (Gonin et al., 2013; Guentas et al., 2016; Bourles et al., 2019; Bourles et al., 2020a).
New Caledonia is a unique biodiversity hotspot (Norman, 2003), with one-third of the main island covered by ultramafic soils. These soils, also known as serpentine soils, result from the weathering and pedogenesis of ultramafic rocks. They are characterized by i) the dominance of iron oxides with prominent levels of potentially toxic metals such as Ni, Co, Cr, and Mn; ii) low levels of key plant nutrients such as N, P, and K; and iii) a highly unbalanced Ca/Mg ratio (Brooks, 1987). Edaphic conditions led to the development of a specific flora characterized by 96.7% endemism (Isnard et al., 2016). In these extreme conditions, plants have developed several adaptive mechanisms, including i) slow growth, ii) the ability to limit Mg uptake, and iii) strategies to tolerate relatively high metal concentrations (Kazakou et al., 2008; Jaffré, 2023).
Ultramafic environments are exploited for economically valuable ores such as Ni, Co, and Cr (Pascal et al., 2008), and several strategies are used for ecological restoration (Losfeld et al., 2015). Recent studies have shown that the use of microorganisms appears to be a promising approach, regardless of the richness and phylogenetic diversity of the bacterial species found in New Caledonian ultramafic soils (Chaintreuil et al., 2007; Herrera et al., 2007; Klonowska et al., 2012; Gonin et al., 2013; Bourles et al., 2019; Bourles et al., 2020a).
Recently, we investigated the mechanisms underlying stress metal adaptation of the PGPR species Paraburkholderia ultramafica STM10279T. This bacterium was isolated from the roots of Tetraria arundinaceae (Cyperaceae), a pioneer species growing on ultramafic soils in New Caledonia (Guentas et al., 2016; Bourles et al., 2020b). We proposed that the EPS produced by this species may be responsible for the high tolerance of P. ultramafica STM10279T to the extreme edaphic constraints of ultramafic soils (Bourles et al., 2020b) by binding metal cations. EPS production has been reported in the genus Paraburkholderia (Achouak et al., 1999; Serrato et al., 2006; Silipo et al., 2008; Suárez-Moreno et al., 2012), and the importance of the corresponding EPS in plant adaptation has been reviewed (Ferreira et al., 2011). Notably, EPS has been shown to play a role in plant–bacteria interactions and tolerance to environmental stress (Liu et al., 2020; Fu and Yan, 2023).
Given the potential role of the EPS of P. ultramafica STM10279T in plant growth, we investigated the effects of EPS inoculation on the growth of Tetraria comosa. T. comosa ex Costularia comosa (Larridon et al., 2018) is a pioneer herbaceous plant belonging to the Cyperaceae family, the dominant family of herbaceous strata in New Caledonian ultramafic soils. T. comosa is used for restoration programs (Wulff et al., 2010), but it is characterized by slow growth. We hypothesize that EPS is involved in the interaction of P. ultramafica STM10279T with plant tissues, which is also expected to occur under edaphic constraints, thereby conferring metal resistance to endemic plant hosts. Our data suggest that plant inoculation with the EPS produced by P. ultramafica STM10279T offers significant advantages over inoculation with intact bacterial cells.
2 Materials and methods
2.1 Paraburkholderia strain
Plant sampling and isolation of the bacterial strains used in this work were as previously described (Gonin et al., 2013). The site used for this purpose was located in a ligno-herbaceous scrubland ecosystem on ultramafic cambisol at an altitude of 534 m. The geochemical soil characteristics are reported in Supplementary Table S1. The characterization of P. ultramafica STM10279T, including morphological, biochemical, and molecular traits, was performed as previously reported (Guentas et al., 2016). P. ultramafica STM10279T isolated from ultramafic soils tolerates edaphic constraints such as low pH (range 4–8), unbalanced Ca/Mg ratios (1/19), and high concentrations of trace metals. P. ultramafica STM10279T was also shown to exhibit specific properties relevant to the promotion of plant growth, such as the production of the phytohormones ACC and IAA, NH3, and siderophores. All details concerning its tolerance to edaphic constraints and PGP activity are reported in Supplementary Table S1.
2.2 Preparation of the bacterial inoculum
A primary culture of bacteria was established by inoculating 10 mL of Lysogenic Broth Lennox medium (LB; tryptone: 10 g/L, yeast extract: 5 g/L, and NaCl: 5 g/L; CONDA, Madrid, Spain) with one LB plate agar colony and incubating the culture at 28°C under orbital shaking at 125 rpm for 48 h. The primary culture was then used to inoculate 100 mL of LB medium at OD600 0.1, and the culture was incubated for 72 h in the above conditions. The cells were then pelleted by a 20-min centrifugation at 5,000 g (Sigma, 2K15) and washed twice with sterile deionized water. The pellets were resuspended in sterile deionized water to reach an OD600 of 0.5 corresponding to a bacterial concentration of 107 to 108 CFU/mL, as suggested by Larcher et al. (2003). The cell suspensions were further diluted and plated on LB agar plates (three replicates) to obtain isolated colonies, and the cultures were incubated at 28°C.
2.3 EPS production and purification
The EPS of P. ultramafica STM10279T was produced as described by Bourles et al. (2020b). To this end, 96-h cultures were grown at 28°C under 125 rpm orbital shaking in minimum broth medium (MB; KH2PO4 ·3H2O, 0.66 g/L; NaCl, 50 mg/L; MgSO4 ·7 H2O,10 mg/L; FeCl3, 4 mg/L; CaCl2,40 mg/L) supplemented with glucose (30 g/L). Eight liters of LB was inoculated using a 48-h grown culture (800 mL; 28°C, 125 rpm orbital shaking) to produce enough biomass. After 72 h of incubation (28°C, 125 rpm orbital shaking), the biomass was recovered by centrifugation at 9,000 rpm for 25 min (Sorvall RC5C Centrifuge, DuPont, Wilmington, DE, USA) and washed once with sterile deionized water. Cells were transferred at 28°C in 10-L reactors containing 8 L of MB medium supplemented with glucose (3% w:v). The bacteria were removed from the medium by centrifugation at 9,000 rpm for 25 min. The supernatants were collected, and the EPS was purified by ultrafiltration on a 100,000 cut-off Sartorius nominal molecular weight cutoff (NMWCO) membrane and immediately freeze-dried. The EPS powders were collected and used for structural analyses and to prepare plant inoculum. In the latter case, the EPS was solubilized in sterile water at a concentration of 10 mg/mL.
2.4 EPS chemical analysis
2.4.1 Polymer hydrolysis
Ten milligrams of polysaccharides was hydrolyzed in trifluoroacetic acid (TFA) (1 mL; 2 M) for 90 min at 120°C under gentle agitation (600 rpm).
2.4.2 HPAEC-PAD analysis
The hydrolyzed sample (from 2.4.1) was neutralized using a 33% NH3 solution, centrifuged at 4°C (13,000 × g; 15 min, room temperature), diluted (1/10, 1/100, and 1/1,000), and filtered on 0.22-µm filters. Twenty-five microliters was injected on a Carbopac PA-1 column (Dionex Corporation, Sunnyvale, CA, USA; 4 × 250 mm) equipped with a pre-column (Dionex Corporation, 4 × 50 mm). The elution was performed at a flow rate of 1 mL/min using an isocratic gradient of NaOH (18 mM) for 30 min followed by a linear gradient of sodium acetate (NaOAc; from 0 to 1 M) in NaOH (200 mM) for 20 min. A pulsed amperometric detector was used for the analysis (Dionex Corporation, ICS 3000). The acquisition and processing software used was Chromeleon version 6.8.
2.4.3 GC/MS-EI analysis
The EPS hydrolysate (from 2.4.1) was dried under nitrogen, washed twice with methanol (1 mL), and dried again. The derivatization was carried out as described by Pierre et al. (2012); Pierre et al. (2014) using BSTFA : TMCS (99:1) (2 h, room temperature, 600 rpm). The solvent was then evaporated under nitrogen, and the trimethylsilyl-O-glycosides were resuspended into dichloromethane (10 g/L) and diluted prior to analysis. Standard monosaccharides (Man, Glc, Ara, Rha, Rib, Fuc, Xyl, Fruc, ManA, GalA, and GlcA) were prepared under the same experimental conditions. The analyses were performed by gas chromatography/mass spectrometry–electron ionization (GC/MS-EI) using an Agilent 6890 Series GC System coupled to an Agilent 5973 Network Mass Selective Detector. The derivatives were injected on an OPTIMA-1MS (30 m, 0.32 mm, 0.25 μm) from Macherey-Nagel (Düren, Germany) with a helium flow rate of 2.3 mL/min. The helium pressure was set at 8.8 psi and the split ratio at 50:1. The following temperature program was used for the separation: 8°C/min to reach 100°C, 3 min at 100°C, 8°C/min to 200°C and 200°C for 1 min, and 5°C/min to >215°C (runtime 19 min 50 s). The ionization was carried out by EI (70 eV), with the trap temperature fixed at 150°C and the target ion set at 40–800 m/z. The injector temperature was 250°C.
2.5 Glycosidic linkage analysis
Pellets of solid NaOH were crushed in dimethyl sulfoxide (DMSO) as described by Ciucanu and Kerek (1984). Methylation was performed by a method adapted from Peña et al. (2012). Briefly, 2 mg of P. ultramafica EPS was resuspended in DMSO (200 to 500 μL), methylated using iodomethane, and hydrolyzed in TFA (2 M, 90 min, 120°C). The methylated monosaccharides were converted into partially O-methylated alditol acetates (PMAAs) using sodium borodeuteride and ethyl acetate. PMAAs were finally solubilized in dichloromethane (200 μL) and analyzed by GC/MS-EI using the same conditions and apparatus as described above.
2.6 Phosphate solubilization by the P. ultramafica EPS
Phosphate solubilization by P. ultramafica EPS was evaluated in vitro using the EPS alone or the EPS in citric acid as suggested by Yi et al. (2008). The freeze-dried EPS was sterilized by cold ethanol, centrifuged, and freeze-dried again. A solution of 10 mg of the EPS in 10 mL sterile deionized water and a stock solution of citric acid consisting of 19.2 mg in 10 mL sterile H2O were prepared. One milliliter of the EPS solution and 10 µL of citric acid solution were mixed and freeze-dried prior to the addition of 1 mL of Pikovskaya medium (PVK) (yeast extract: 0.5 g/L; glucose: 10 g/L; Ca3(PO4)2: 5 g/L; MgSO4, H2O: 0.1 g/L; MnSO4, H2O: 0.002; KCl: 0.2 g/L; NH4SO4: 0.5 g/L; FeSO4, 7H2O: 0.002 g/L; NaCl: 0.2 g/L). The PVK medium and a KH2PO4 solution (2 mg/L) were used as negative and positive controls, respectively. The mixtures were incubated at 28°C under lateral agitation (30 rpm). After 48 h, 5 mL H2Od, 1 mL chloromolybdic acid, 1 mL hydroquinone (10 g/L), and 1 mL sodium sulfite (200 g/L) were added. The appearance of a blue color indicates a positive result, i.e., phosphate solubilization (Pikovskaya, 1948). The concentration of the solubilized phosphate was estimated using a standard curve of KH2PO4 (Supplementary Figure S1; dilutions: 25, 50, 100, 150, 200, and 250 mg/L of KH2PO4; λ = 700 nm).
2.7 Greenhouse experiments
2.7.1 Substrate preparation
The substrate used was a mixture of i) 2-mm sieved colluvial lateritic soil (ferralsol) sampled from the Plum area in New Caledonia (22°16′59″S, 166°39′12″E) with the following characteristics: coarse sand, 39.4%; fine sand, 22.1%; silt-clay, 37.2%; pH H2O, 5.9; pH KCl, 5.6; total C, 42.1 g/kg; total N, 2.2 g/kg; total P, 147 mg/kg; available P (Mehlich), 3 mg/kg; total Ca, 1.06 g/kg; total Mg, 5.08 g/kg; Ca/Mg, 0.207; and ii) commercial compost (4:1 v/v). The composition of commercial compost was as follows: N, 1.7 mg/g; Ptotal, 150 mg/kg; POlsen, 7 mg/kg; K, 139 mg/kg (Terreau universel, Agrofino, France). Metal contents of the ferralsol were as follows: Co, 0.87 g/kg; Cr, 22 g/kg; Fe, 348 g/kg; Mn, 10 g/kg; and Ni, 5.6 g/kg. Available metal extracted by dimethylene triaminopentaacetic acid (DTPA) were as follows: CoDTPA, 68 mg/kg; CrDTPA, 0.17 mg/kg; FeDTPA, 71 mg/kg; MnDTPA, 1.1 g/kg; and NiDTPA, 130 mg/kg. The addition of this commercial compost to the substrate for the greenhouse experiments was necessary because the growth of the plantlets of T. comosa is known to be inhibited in pure lateritic topsoil (Lagrange et al., 2011). The soil/compost mixture was autoclaved three times at 120°C for 1 h, with an interval of 24 h, to eliminate the soil’s original microorganisms.
2.7.2 Plant production in nursery and plant inoculation/amendment
The seeds of T. comosa were provided by SIRAS Pacifique (Nouméa, New Caledonia). Germination was carried out on the soil/compost mixture described above. After 4 months, two- or three-leaf stage T. comosa plantlets were harvested as indicated elsewhere (https://www.agripedia.nc/sites/default/files/pdf/fiche_costularia-comosa.pdf) and washed with sterile water to remove any contaminants. The seedlings were then transplanted into 1-L plastic containers filled with approximately 600 g of the soil/compost mixture described above.
A comparison was made between non-inoculated plants, plants inoculated with P. ultramafica STM10279T, and plants exposed to the EPS produced by P. ultramafica STM10279T. A 5-mL bacterial suspension of 107 and 108 CFU/mL (corresponding to a 0.5 OD600) or EPS solution (10 mg/mL) or sterilized water (control) was applied once on the roots of each plantlet at transplanting (15 plants per inoculation treatment), and the plants were grown under greenhouse conditions for 8 months as described elsewhere (Lagrange et al., 2011). To control water supply and better reproduce environmental conditions, watering (100 mL) was manually conducted every 2 days. Plant growth was assessed over time by measuring the size of the largest leaf every 2 months. After harvesting the plants, the soil was removed from the roots by extensive washing, the shoots and roots were separated, and their mass was measured after drying at 60°C for 72 h (Bashan et al., 2017).
2.7.3 Chemical analysis of plants
Plant tissue samples were pooled in groups of three replicates and ground to powder. The Ca, Mg, Na, K, P, Co, Cr, Fe, Mn, and Ni contents were measured in shoots and roots by inductively coupled plasma–optical emission spectrometry (ICP-OES; Varian®, Varian 730-ES, Palo Alto, CA, USA) after hydrolysis with a mixture of HNO3 (69%) and H2SO4 (37%) (4:1, v/v) at the “Laboratoire des Moyens Analytiques” (LAMA-US IMAGO-IRD, New Caledonia). The concentration of the different elements in the plant tissues was used to estimate the percentage of gain or loss in response to bacterial inoculation, as well as the translocation factor (TF). The gain or loss of a trace metal was calculated as [(Concentration in the tissue of inoculated plant − Concentration in the tissue of control plant) * 100)/Concentration in the tissue of control plant] (Gonin et al., 2013). The TF was calculated as [Concentration in the shoots of inoculated plant/Concentration in the roots of inoculated plant] (Galal and Sheata, 2015).
2.7.4 Data and statistical analyses
All statistical analyses were performed using the R software version 3.3.1 (R Core Team, 2021) (version 1.2–4, available at http://CRAN.R-project.org/package=agricolae). All data were analyzed using ANOVA parametric tests, followed by Tukey’s honestly significant difference (HSD) test (p < 0.05) or with non-parametric tests (Kruskal–Wallis test followed by Fisher’s least significant difference (LSD) test (p < 0.05) when data did not follow the assumptions of a parametric test. Methods were applied independently for each plant species and each variable. For the greenhouse experiments, a principal component analysis (PCA) was carried out followed by a hierarchical clustering on principal components (HCPC), which allowed classification of the treatments according to the chosen variables.
3 Results
3.1 Chemical composition of the EPS of P. ultramafica STM10279T
The chemical composition of the EPS is shown in Table 1. High-performance anion-exchange chromatography (HPAEC) analysis showed the presence of galactosyl (Gal; 35.8%), glucosyl (Glc; 33.2%), rhamnosyl (Rha; 19.5%), mannosyl (Man; 7.2%), and glucuronosyl (GlcA; 4.4%) residues. The composition was verified by GC/MS analysis of O-trimethylsilylated derivatives, which indicated the following molar ratios (%): Gal (30.6%), Rha (25.0%), Glc (22.9%), Man (12.1%), and GlcA (9.4%).

Table 1 Biochemical and monosaccharide composition of the EPS produced by Paraburkholderia ultramafica STM10279T.
The data obtained from the GC/MS analysis of the PMAAs and corresponding glycosidic linkages are summarized in Table 2. The results show the presence of terminal Galp (34.4%), (1,3)-Glcp (14.1%), (1,2,3)-Glcp (10.3%), (1,2,3)-Rhap (8.4%), (1,2)-Rhap (6.6%), (1,3)-Manp (6.4%), (1,3,6)-Manp (5.8%), and terminal Glcp (2.5%). These assessments were in keeping with the monosaccharide composition obtained after trimethylsilyl (TMS) derivatization (Table 1), except for Rha (15% versus 19.5%–25%). The GC/MS column and temperature program used for methylation analysis of the EPS did not allow the separation of Glcp from GlcAp. However, with the assistance of TMS analysis, we were able to estimate that 11.6% of the (1,2,3,4)-linked Glcp was in fact assignable to GlcAp. The proportion of Gal (34.4%):Glc (26.9%):Rha (15%):Man (12.1%):GlcA (11.6%) was close to 3:2.3:1.3:1:1. Furthermore, terminal and branched total residues % were close (36.98% and 36.02%, respectively).
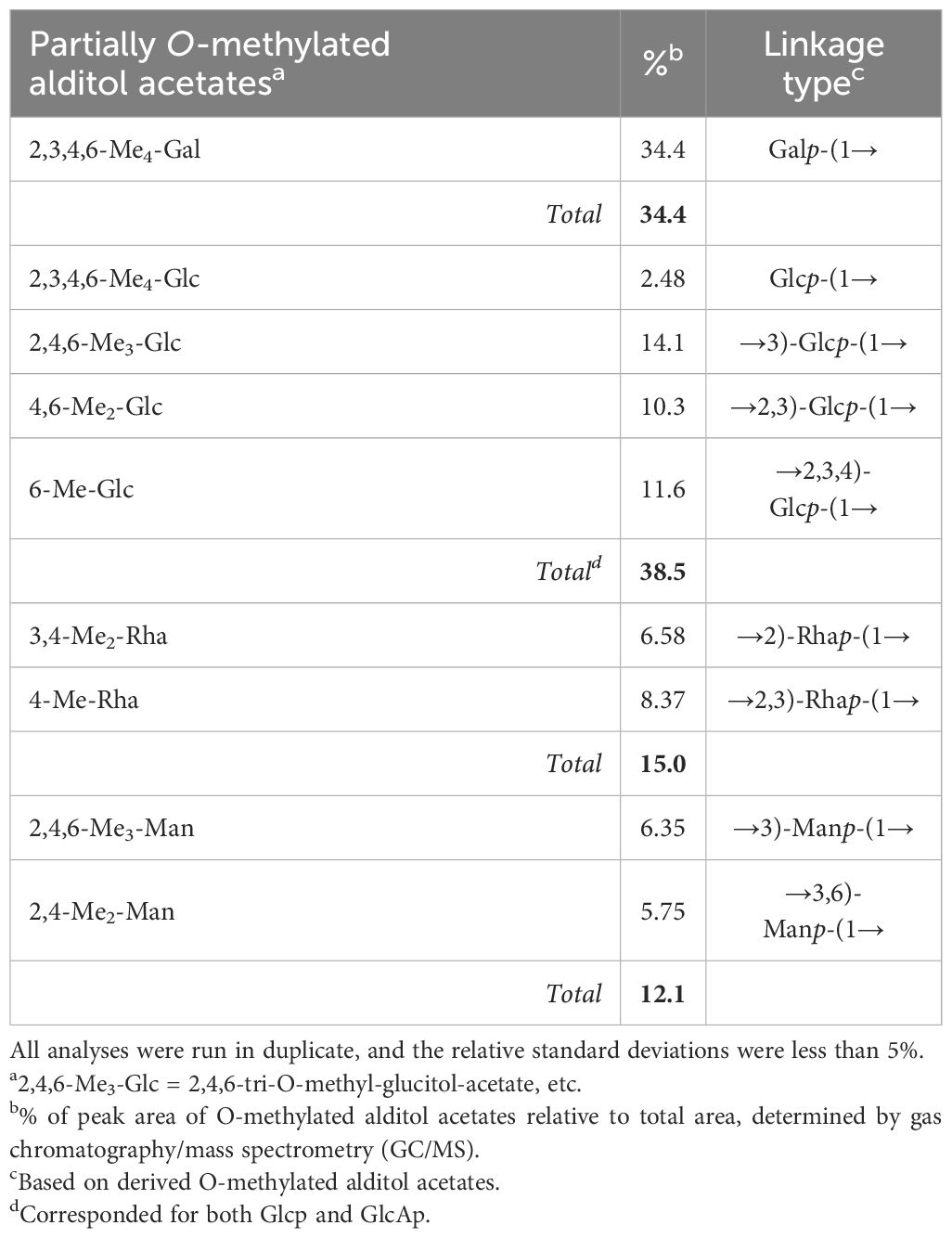
Table 2 Partially methylated alditol acetates (PMAAs) and glycosidic linkages of the EPS produced by Paraburkholderia ultramafica STM10279T.
The main backbone (48.1%) of the EPS could thus be composed of (1,3)-linked Glcp (and/or GlcAp) and (1,3)-linked Manp. The presence of primary branched sugar residues (1,2,3)-linked Glcp (10.3%), (1,3,6)-linked Manp (5.75%), and (1,2,3,4)-linked GlcA (11.6%) suggests that the main chain is highly branched (>60%) throughout the O-4 of (1,3)-linked GlcAp, probably by (1,2)-Rhap. The ramifications are probably composed of [1-Rhap-(2→1)-Galp] substituted with putative Glcp through the O-3 position of the (1,2)-Rhap. The core-chain unit may be also substituted by Galp through the O-2 position of (1,3)-linked Glcp or (1,2,3,4)-linked GlcAp and the O-6 positions of the (1,3)-linked Manp. The non-reducing ends in the EPS of P. ultramafica most likely correspond to t-Galp (34.4%) and t-Glcp (2.48%).
3.2 Phosphorus solubilization
The experiment showed a phosphorus-solubilizing effect of the P. ultramafica STM10279T EPS. A semi-quantitative assessment of phosphorus solubilization was made by measuring absorbance at 700 nm (Supplementary Figure S1; Supplementary Table S2). The results showed a positive effect of P. ultramafica STM10279T EPS on phosphate solubilization, with an average solubilization of 74.9 ± 32.1 mg phosphorus (expressed in KH2PO4 equivalents) per 10 mg EPS (n = 3). The addition of citric acid was not accompanied by an increase in phosphate solubilization (Supplementary Table S3).
3.3 Plant growth, and shoot and root biomass
Photographs of T. comosa inoculated with bacteria or EPS, removed from pots after 8 months of growth in ultramafic soil, are shown in Supplementary Figure S3. Throughout the experimental period, no significant differences were observed between the different treatments. At the end of the experiment, T. comosa plants had grown to an average height of 16 ± 0.4 cm. The effect of bacterial and EPS inoculation on shoot and root dry weight is shown in Figure 1. The shoot dry weight of T. comosa inoculated with the EPS of P. ultramafica STM10279T was the highest (549 ± 117 mg) with a significant difference compared to the control (2.5 times higher). With regard to the root, no significant differences were observed between the treatments. However, T. comosa plants inoculated with the P. ultramafica STM10279T EPS also showed the highest value of root dry weight (281 ± 80 mg, 2.5 times higher than the control, Supplementary Table S4).
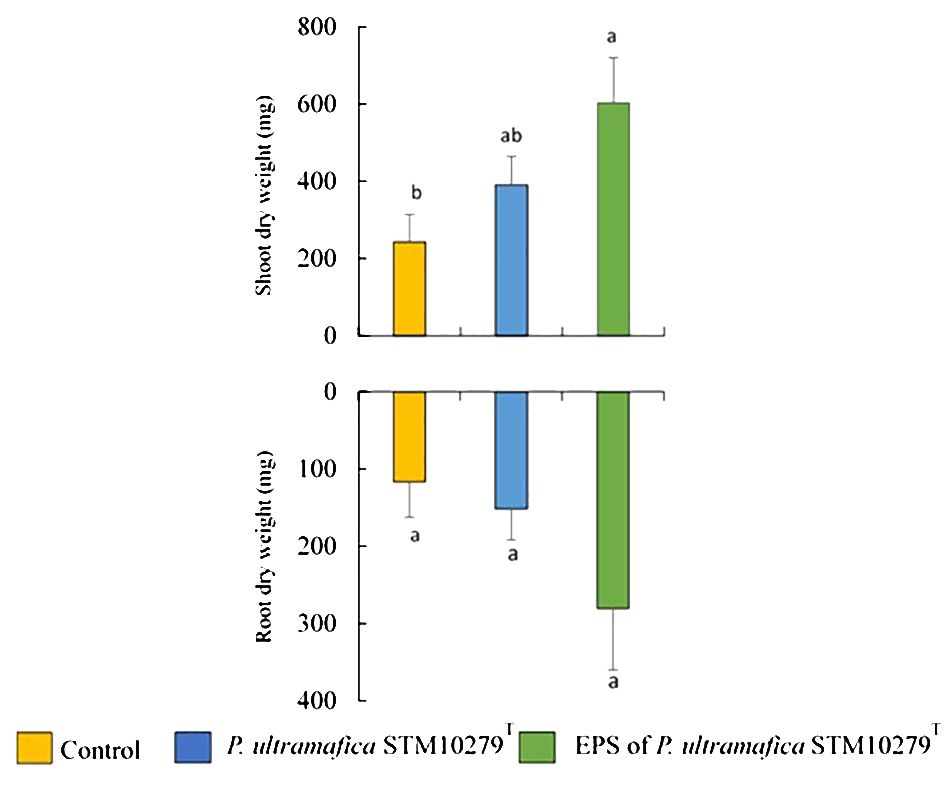
Figure 1 Influence of bacterial or exopolysaccharide (EPS) inoculation on Tetraria comosa on dry weight (mean values of height with standard errors). For each measure part of the plant, different letters above columns indicate significant differences at p < 0.05.
3.4 Plant nutrient content and uptake
Inoculation of the P. ultramafica STM10279T EPS (Supplementary Figure S2; Supplementary Table S5) significantly enhanced P concentration (455 ± 24 mg/kg; + 18%) in shoots compared to the control. In root tissues, K concentration was significantly higher in the plant inoculated with the EPS (6.8 ± 0.0 mg/kg; +13%) compared to the control. No significant differences were observed in either tissue for the Ca/Mg ratio (Table 3).
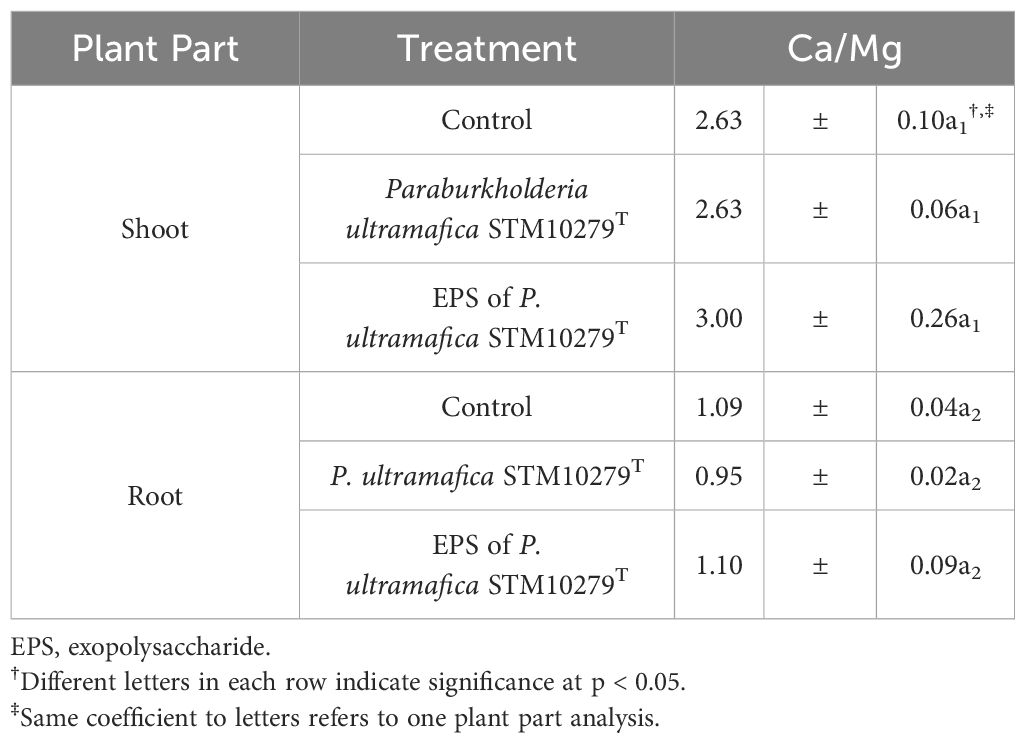
Table 3 Ca/Mg ratio in Tetraria comosa as a function of various treatments (mean ± SE, n = 3).
Variations of the Ca, K, and P uptake in the roots and shoots of the T. comosa inoculated plants in comparison to the non-inoculated plants are presented in Figure 2, while full data are reported in Supplementary Table S7. Consistent with biomass increase, inoculation of the EPS (Figure 2; Supplementary Table S6) significantly enhanced Ca uptake (shoot: 2.4 mg/plant, 2.4 times higher; root: 0.5 mg/plant, 2.8 times higher), K uptake (shoot: 6.2 mg/plant, 2.7 times higher; root: 1.9 mg/plant, 2.7 times higher), and P. uptake (shoot: 249 µg/plant, 2.7 times higher; root: 94 µg/plant, 3.1 times higher) compared to the control.

Figure 2 Effect of inoculation on essential mineral uptake of Tetraria comosa grown on ultramafic substrate. Relative variations of element content in shoots and roots are expressed as % of element content in dry mass of tissues of inoculated plants compared with controls. Bars represent means, and error bars represent standard deviation of means (n = 3). Different letters above columns indicate significant differences at p < 0.05. The same coefficient to letters refers to one element analysis. The letters and coefficients with an asterisk indicate significant differences with controls at p < 0.05.
3.5 Metal content
In shoot tissues, inoculation with P. ultramafica STM10279T and exposure to the EPS led to a reduction in all trace metals, but no significant differences were observed compared to the control (Figure 3; Supplementary Table S7). In roots, an increase in trace metal content was observed with both inoculations, but also no significant differences were observed compared to the control. Metal translocations (Table 4) were significantly reduced with the inoculation of P. ultramafica STM10279T and with the EPS, but significant differences were observed only for the translocation of Co, Cr, and Fe compared to the control. No significant differences were observed between plants inoculated with P. ultramafica STM10279T or exposed to the P. ultramafica STM10279T EPS.
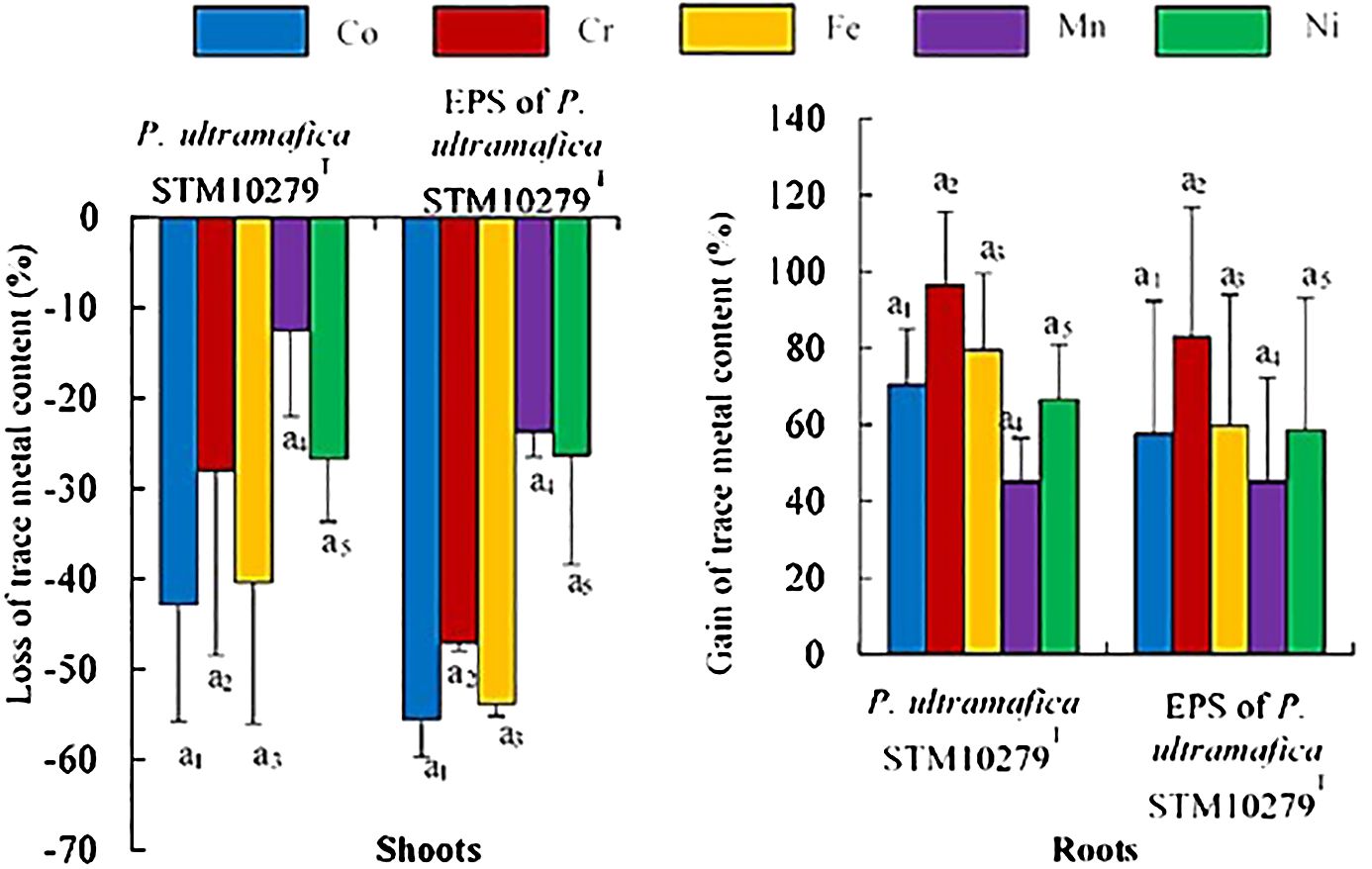
Figure 3 Effect of inoculation on essential mineral content of Tetraria comosa grown on ultramafic substrate. Relative variations of element content in shoots and roots are expressed as % of element content in dry mass of tissues of inoculated plants compared with controls. Bars represent means, and error bars represent the standard deviation of means (n = 3). Different letters above columns indicate significant differences at p < 0.05. The same coefficient to letters refers to one element analysis. The letters and coefficients with an asterisk indicate significant differences with controls at p < 0.05.
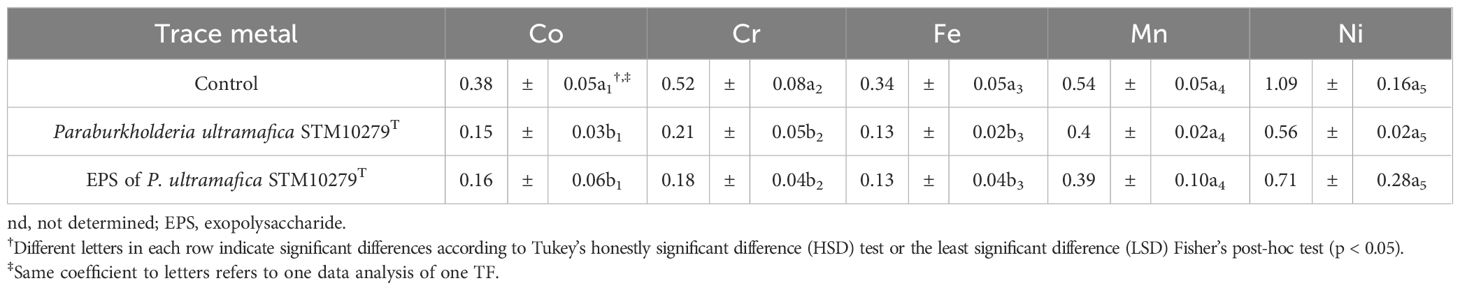
Table 4 Trace metal translocation factor (TF) (mean ± SE, n = 3) as a function of various treatments.
3.6 PCA
A PCA was carried out to analyze the effect of the multiple variables arising from the experiments described above (Figure 4). The analysis showed that two principal components (PC1 and PC2) explained 72.9% of the total variation. The variables involved in the clustering of each group are presented in Table 5. Briefly, control plants are characterized by significantly higher i) translocation of Co, Cr, and Fe and by significantly lower (i) contents of Mg, P in the root; ii) uptake of P, K, Ca, and Mg in shoots; iii) uptake of P and Mg in roots; and iv) root biomass compared to the other treatments. T. comosa inoculated with P. ultramafica STM10279T are characterized by significantly higher i) Na content in shoot and root, ii) Mg content in root, and iii) Na uptake in shoot compared to the other treatments. Inoculation of the P. ultramafica STM10279T EPS significantly enhanced the uptake of Ca, Mg, K, and P and K contents in roots.

Figure 4 Principal component analysis (PCA) and hierarchical clustering on principal components (HCPC). PCA and HCPC were performed with 29 variables: biomass [dry weight tissue of shoots (BA) and roots (BR)], mineral and metal content in shoots (i.e., Ca_T) and roots (i.e., Ca._R) (Ca, Mg, K, Na, and P), metal translocation factor (i.e., Co_FT) (Co, Cr, Fe, Mn, and Ni), mineral element uptake in shoots (i.e., Ca_uptT) and roots (i.e., Ca_uptR) (Ca, Mg, Na, K, and P), and Ca/Mg ratio in shoots (CaMg_T) and roots (CaMg_R). Analyses were carried out using R software with the “FactoMineR” package. HCPC used Euclidean distances for calculating dissimilarities between observations and average method to define clusters.
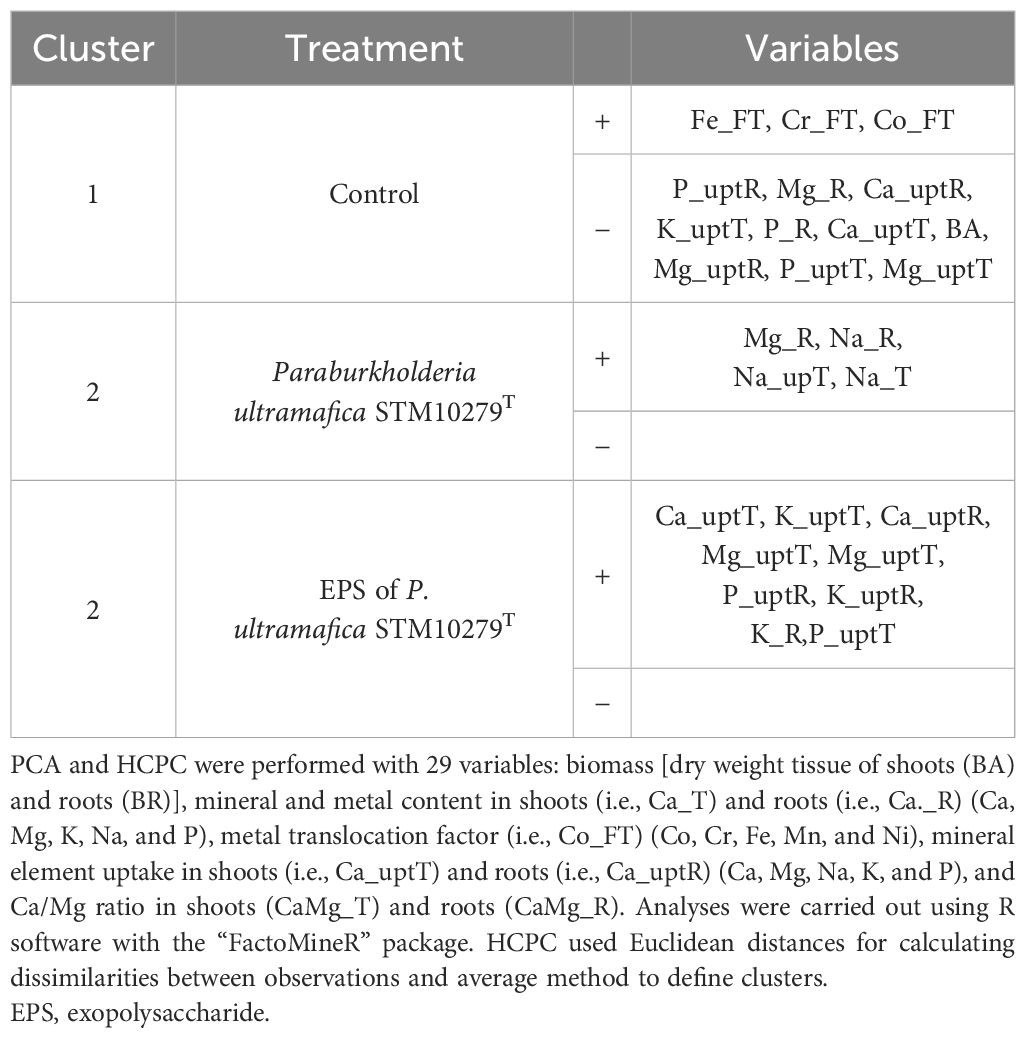
Table 5 Variables that defined each cluster (p < 0.05) using a principal component analysis (PCA) and hierarchical clustering on principal components (HCPC).
4 Discussion
EPS plays important roles in plant–bacteria interaction and tolerance to environmental stresses (Bhagat et al., 2021) and has also been found to be important in facilitating plant–endophyte interactions, especially under stress conditions (Liu et al., 2017, 2020). A recent study by Fu and Yan (2023) has shown that EPS production is necessary for optimal colonization of the endophytic bacterium Paraburkholderia phytofirmans PsJN in plant tissues under drought-stress conditions. Given the numerous beneficial properties of EPS under varied stress conditions, as discussed in previous literature (Bhagat et al., 2021), our aim is to study the specific plant growth-promoting and metal resistance properties of the cepacian-like EPS produced by P. ultramafica STM10279T, a bacterium previously isolated in our laboratory (Bourles et al., 2020b).
4.1 P. ultramafica STM10279T exopolysaccharide
The most common exopolysaccharide produced by Burkholderia gender is cepacian. Cepacian is a well-known acetylated EPS whose repeating unit consists of a heptasaccharide repeating unit comprising d-Glc, d-Rha, d-Man, d-Gal, and d-GlcA in a ratio of 1:1:1:3:1 (Cescutti et al., 2010). It has been found in different species belonging to environmental or clinical isolates (Ferreira et al., 2011). Ferreira et al. (2010) showed that the proteins involved in cepacian production are well conserved among the Burkholderia genus. External factors directly influence the type and number of EPS produced; indeed, some strains produced a single EPS, while others produced a mixture. Hallack et al. (2010) described the EPS “B” for Paraburkholderia kururiensis, a mixture of cepacian and an octosaccharide polymer. Bourles et al. (2020b) demonstrated the ability of P. ultramafica STM10279T to produce an EPS when the strain was cultivated in a glucose-rich medium. The recovered EPS is composed of 17% uronic acid, and the carbohydrate-to-protein (C/P) ratio was almost 15, with values of 61% carbohydrate to 4% protein residues. Infrared spectroscopy analysis using a Frontier™ (PerkinElmer, Waltham, MA, USA) Fourier transform infrared (FT-IR) spectrometer revealed that the monosaccharides have pyranose rings and the presence of uronic acid groups. Bourles et al. (2020b) showed that the main mechanism involved in the Ni tolerance of the metal-resistant PGP P. ultramafica STM10279T strain was related to the production of an acidic exopolysaccharide.
In this study, we examined the composition of the EPS of P. ultramafica STM10279T and found that it is an anionic EPS containing glucuronic acid. Its structure is highly branched with over 60% branching. The structure is very similar to cepacian (Cérantola et al., 1999), but differs in two ways: first, there is a terminal glycosyl residue (2.48%) instead of the lateral chain throughout the O-3 position of the (1,2)-Rhap as in the structure of the EPS “B” of P. kururiensis (Hallack et al., 2010); second, the main chain of the EPS contains and unsubstituted (1,3)-linked Manp and is substituted throughout the O-2 position of the (1,3)-Glcp probably by Galp. Table 6 shows the hypothetical structural formula of the repeating unit of the EPS of P. ultramafica and a comparison with the structures of cepacian and EPS “B” of P. kururiensis.
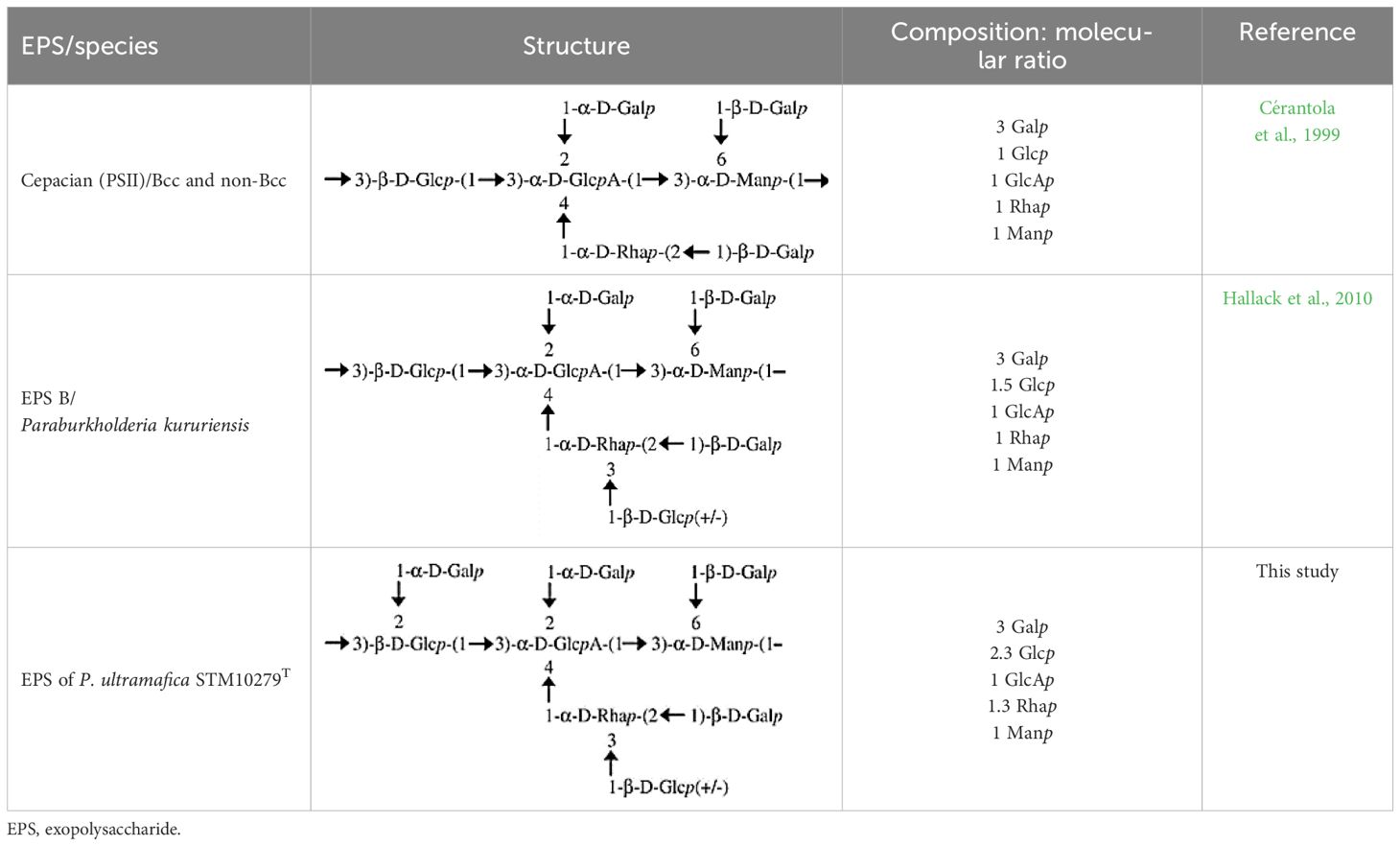
Table 6 Hypothetic formula of the exopolysaccharide repeated units of the Paraburkholderia ultramafica STM10279T and comparison with the closest EPS repeated unit described in the literature.
4.2 Effects in planta of P. ultramafica STM10279T and its EPS
ETMs (trace metal elements), or essential trace metals, interact with plants through various metabolic and physiological pathways. When soils have high ETM concentrations, it can result in reduced shoot and root biomass. This can cause seedlings to have poorly developed root systems, which can make it difficult for them to absorb nutrients that are essential for proper plant development. In fact, seedlings growing on soils contaminated with ETMs may not be able to absorb the necessary nutrients (Hameed et al., 2015).
High concentrations of ETMs in the aerial part of the plant can have a negative effect on chlorophyll content and can inhibit the activity of certain enzymes, which can slow down metabolic pathways. While certain ETMs, such as Mn, Mo, and Ni, are essential for proper cell functioning, an excess of these metals can inhibit a large number of plant enzymes, particularly those involved in the Calvin cycle and chlorophyll biosynthesis. The real danger with most ETMs lies in their oxidation state and their ability to interact with the cellular components, leading to the formation of reactive oxygen species (ROS) that disrupt cellular metabolism. This can cause symptoms like chlorosis and necrosis, leading to a reduction in the plant’s growth and vitality and even death (Hossain et al., 2012; Evlard and Campanella, 2013). Several studies have been conducted to explore the impact of inoculating P. ultramafica STM10279T on sorghum as well as on two native plants of New Caledonian ultramafic soils: Metrosideros laurifolia and T. comosa (Gonin et al., 2013; Gensous, 2014; Guentas et al., 2016; Bourles et al., 2020b). The primary effect of inoculating P. ultramafica STM10279T was observed in the reduction of trace metals in tissues. Bourles et al. (2020b) proposed that this effect could be attributed to the bacteria’s ability to produce EPSs. EPS production can lead to biomineralization of metal ions, which can enhance metal stress tolerance in plants (Bhagat et al., 2021). Several studies have reported that EPS can bind with cationic ETMs owing to the presence of anionic functional groups, which results in the formation of EPS–metal complexes (Mishra et al., 2017). These complexes restrict the mobility of the metal ions to the plants (Bhagat et al., 2021). The functional groups, such as hydroxyl and carboxyl, present in the polysaccharides, play a crucial role in the metal binding process (Gupta and Diwan, 2017; Etesami, 2018). To test this hypothesis, we inoculated T. comosa with EPS produced by P. ultramafica STM10279T. To the best of our knowledge, this is the first study to demonstrate the effect of a microbial EPS on plant growth in ultramafic soils. Indeed, although the role of EPS in plant interactions is well established (Naseem et al., 2018), especially in Burkholderia species (Ferreira et al., 2011), no study has directly demonstrated the effects of EPS inoculation on host plants.
In our experiment where plants were inoculated with P. ultramafica STM10279T or amended with its EPS, it was observed that metal translocation factors decreased significantly. This suggests that metals are immobilized at the root level and not translocated to the aerial parts. However, no significant differences in root physiology or shoot symptoms were observed. Plants amended with EPS of P. ultramafica STM10279T had the highest values of shoot dry weight, indicating that supplementing the ETM roots with P. ultramafica’s EPS could improve the plant’s status. Additionally, metal accumulation at the root level corresponds to a phytostabilization process, which reduces the mobility and availability of contaminants in the soil (Alkorta et al., 2010). This helps to stabilize the contaminated site and prevent the dispersion of contaminants by erosion, reducing their impact on the food chain. Our experiment found higher concentrations of ETMs in the roots of inoculated plants compared to non-inoculated plants. However, a previous study by Bourles et al. (2020b) showed a significant decrease in metal concentrations in the roots after inoculation with P. ultramafica STM10279T. The difference in results could be due to the duration of the experiment (8 months in our study compared to the study of Bourles et al.). Despite the difference in results, we observed a similar effect in plants inoculated with P. ultramafica STM10279T or amended with its EPS, which underlines the involvement of EPS in the alleviation of metals. This finding supports the hypothesis of Bourles et al.
Indeed, EPS is known to improve soil aggregation, water permeability, and nutrient uptake at the root level, as reported in studies by Vejan et al. (2016) and Upadhyay et al. (2012). Different watering methods were observed in various experiments, which could explain the varying effects seen with P. ultramafica STM10279T inoculation. Rubin et al. (2017) demonstrated that PGPRs are more effective in mitigating water stress, while Bourles et al. (2020b) used an automatic watering system that irrigated the plants twice a day, but this method did not replicate the natural conditions (L'Huillier et al., 2010). In our study, watering was reduced to a manual process (100 mL) every 2 days. These differences highlight the fact that there are still parameters in the rhizosphere that remain to be discovered. Bourles et al. (2020b) found that the inoculation with P. ultramafica STM10279T did not have a positive effect on the biomass of T. comosa; nor did it have any significant effects on plant nutrition. Our study, in contrast, showed that shoot and root dry weight increased when the plantlet was amended with P. ultramafica EPS, although this result was not statistically significant at the time of sampling. Our study also highlighted a significant effect of P. ultramafica EPS on trace metal contents compared to non-inoculated plants. However, the effect of metals does not explain the differences observed in growth. Bourles et al. (2020b) suggested that the lack of effect on growth could be explained by the lack of effect on concentrations of essential minerals.
One of the main nutrients limiting the plant growth is phosphorus. On ultramafic soils, the lack of essential plant nutrients is a major limitation for plants (Brooks, 1987). On ultramafic soils in New Caledonia, the amount of phosphorus is limiting for plant growth (L'Huillier et al., 2010). Phosphorus is adsorbed by the abundant iron and aluminum oxides and hydroxides found in these soils, making it unavailable to plants (Gobat et al., 2010). Plants use various strategies to acquire phosphorus from their surroundings, including modification of the root system (López-Bucio et al., 2003), establishment of mycorrhizal symbiosis (Amir and Ducousso, 2010), or interaction with phosphate-solubilizing bacteria (Gonin et al., 2013; Guentas et al., 2016).
Arbuscular mycorrhizal fungi (AMF) are known to enhance plant growth by improving mineral nutrition, particularly phosphorus and potassium (Smith and Read, 2008). The ability of some rhizobacteria to convert insoluble forms of phosphorus into soluble forms is a sought-after trait in PGPRs, particularly in ultramafic soils. The release of low-molecular-weight organic acids is the most common mechanism for P solubilization by phosphate-solubilizing bacteria (PSBs) (Goldstein, 1995). In fact, 33 out of 36 rhizobacterial strains isolated by Chen et al. (2006) from subtropical soils were found to excrete organic acid for P solubilization. Three of their P-solubilizing isolates did not produce organic acid, suggesting that another type of molecule is involved.
Yi et al. (2008) investigated the role of exopolysaccharides in the microbial-mediated solubilization of P. Bacterial strains belonging to the genera Enterobacter, Arthrobacter, and Azotobacter and having the ability to solubilize tri-calcium phosphate were used to evaluate the role of EPS in the solubilization of P. The authors demonstrated that EPS altered P solubilization homeostasis, favoring P dissolved by retaining free phosphorus in the medium, leading to a greater release of phosphorus from insoluble phosphate. In New Caledonia, particular attention has been paid to phosphorous in Cyperaceae (Lagrange, 2009; Lagrange et al., 2011, 2013; Gensous, 2014). Plants in the Cyperaceae family are generally considered to be non-mycorrhizal or very weakly mycorrhizal (Brundrett and Tedersoo, 2019). However, Lagrange et al. (2011) showed that mycorrhizal colonization is functional under greenhouse conditions in T. comosa, one of the most common sedges in the ultramafic maquis.
Bourles et al. (2020a) have demonstrated synergistic microbial interactions between Curtobacterium citreum BE strains, rhizobacteria, and AMF, particularly for P nutrition. C. citreum BE is a PSB capable of producing exopolysaccharide as P. ultramafica STM10279T. Gonin et al. (2013) showed that P. ultramafica STM10279T could solubilize phosphate, but there were no significant increases in phosphorus content when T. comosa was inoculated with P. ultramafica STM10279T. In our study, an improvement in growth was only observed after inoculation with the EPS of P. ultramafica STM10279T. In the aerial parts of these plants, there was a significantly higher concentration of phosphorus compared to both non-inoculated and P. ultramafica inoculated plants. The improvement in growth could be explained by this improvement in P concentration driven by the EPS. Yi et al. (2008) showed the great importance of EPS in the dissolution of tricalcium phosphate, apart from organic acids and protons. Therefore, it is likely to be indirectly involved in phosphate solubilization if a large amount of organic acid (e.g., citric acid) is produced by the PSB. Our experiment showed that P. ultramafica STM279T EPS alone was capable of solubilizing phosphate. Contrary to the results of Yi et al. (2008), the addition of citric acid did not lead to an increase in phosphate solubilization. We found an antagonistic effect of citric acid on phosphate solubilization by EPS.
5 Conclusion
A study was conducted on the EPS extracted from P. ultramafica STM10279T, which revealed that it is a highly branched polymer that is similar to cepacian and EPS B from P. kururiensis. The presence of glucuronic acids in the EPS was confirmed, which could potentially help in reducing metal stress. Greenhouse experiments were conducted involving T. comosa. The experiments used EPS at the root level or inoculated with P. ultramafica STM10279T. The results showed that the EPS not only alleviates metal stress but also stimulates growth. This growth stimulation may be due to improved nutrition. Since T. comosa has little or no mycorrhizae, phosphate nutrition is crucial for its growth, especially in ultramafic soils. The in vitro study on the effect of P. ultramafica EPS on phosphate solubilization highlighted its ability to solubilize phosphate. This ability could explain the growth stimulation of T. comosa on ultramafic soil when amended with EPS. The findings indicate that EPS plays a crucial role in P. ultramafica STM10279T in decreasing the metal levels in plants. However, additional research is required to ascertain whether the EPS generated by P. ultramafica is responsible for the observed impact on T. comosa growth. At the end of the experiment, we need to verify or quantify the presence of P. ultramafica in the roots or rhizosphere of T. comosa. Research will be conducted to understand how EPS chelates with metal ions. The aim is to use this knowledge to develop environmental metal remediation processes. The effectiveness of P. ultramafica STM10279T EPS in reducing the harmful effects of metals on metal-sensitive plants should be studied. Our research indicates that amending P. ultramafica STM10279T EPS directly to the soil is more effective than inoculating the plant with bacterial suspension for improving plant growth. Inoculating the soil with EPS has practical advantages such as easy storage and preparation. It is also an inert, non-living product, which reduces biological risks, making it marketable and safer for use in greenhouses. Direct inoculation of EPS could be a promising approach to improve plant growth and metal tolerance in ultramafic soils. It could also be a useful tool in global strategies for successfully restoring ultramafic areas after mining.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
AB: Data curation, Formal analysis, Investigation, Methodology, Writing – original draft. GP: Formal analysis, Methodology, Writing – review & editing. HA: Supervision, Writing – review & editing, Validation. AL: Data curation, Formal analysis, Methodology, Writing – original draft. EC: Resources, Writing – review & editing. VM: Formal analysis, Methodology, Writing – review & editing. PJ: Writing – review & editing. PM: Writing – review & editing. VB-S: Funding acquisition, Supervision, Writing – review & editing. LG: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing, Validation.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Province Sud of New Caledonia (Prix d’Encouragement à la Recherche: Thesis Grant).
Acknowledgments
We are very grateful to the South Province of New Caledonia for providing a Ph.D. grant to AB. We are also grateful to the LAMA laboratory for chemical analysis (LAMA-US IMAGO-IRD, New Caledonia). Thanks to V. Malisan and L. Carron for the technical support. Many thanks to Professor Vincent Bulone (Flinders University, Adelaide, Australia) for his English proofreading.
Conflict of interest
Author EC is employed by BIOTECAL, Marine Biotechnology Company, New Caledonia.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpls.2024.1349724/full#supplementary-material
References
Achouak, W., Christen, R., Barakat, M., Martel, M. H., Heulin, T. (1999). Burkholderia caribensis sp. nov., an exopolysaccharide-producing bacterium isolated from vertisol microaggregates in Martinique. Int. J. Syst. Bacteriol. 49 (2), 787–794. doi: 10.1099/00207713-49-2-787
Alkorta, I., Becerril, J. M., Garbisu, C. (2010). Phytostabilization of metal contaminated soils. Rev. Environ. Health 25, 135–146. doi: 10.1515/REVEH.2010.25.2.135
Amir, H., Ducousso, M. (2010). “Les bactéries et les champignons du sol sur roches ultramafiques,” in Mines et environnement en Nouvelle-Calédonie : les milieux sur substrats ultramafiques et leur restauration. (Nouméa: IAC), 129–145. Available at: http://www.documentation.ird.fr/hor/fdi:010057561.
Bashan, Y., Huang, P., Kloepper, J. W., de-Bashan, L. (2017). A proposal for avoiding fresh-weight measurements when reporting the effect of plant growth-promoting (rhizo) bacteria on growth promotion of plants. Biol. Fertil. Soils 53, 1–2. doi: 10.1007/s00374-016-1153-1
Bhagat, N., Raghav, M., Dubey, S., Bedi, N. (2021). Bacterial exopolysaccharides: Insight into their role in plant abiotic stress tolerance. J. Microbiol. Biotechnol. 31 (8), 1045–1059. doi: 10.4014/jmb.2105.05009
Bourles, A., Amir, H., Gensous, S., Cussonneau, F., Medevielle, V., Juillot, F., et al. (2020b). Investigating some mechanisms underlying stress metal adaptations of two Burkholderia sensu lato species isolated from New Caledonian ultramafic soils. Eur. J. Soil Biol. 97, 103–166. doi: 10.1016/j.ejsobi.2020.103166
Bourles, A., Guentas, L., Chalkiadakis, E., Majorel, C., Juillot, F., Cavaloc, Y., et al. (2019). New caledonian ultramafic conditions structure the features of curtobacterium citreum strains that play a role in plant adaptation. Can. J. Microbiol. 65 (12), 880–894. doi: 10.1139/cjm-2019-0283
Bourles, A., Guentas, L., Charvis, C., Gensous, S., Majorel, C., Crossay, T., et al. (2020a). Co-inoculation with a bacterium and arbuscular mycorrhizal fungi improves root colonization, plant mineral nutrition, and plant growth of a Cyperaceae plant in an ultramafic soil. Mycorrhiza 30 (1), 121–131. doi: 10.1007/s00572-019-00929-8
Brooks, R. R. (1987). Serpentine and its vegetation. A multidisciplinary approach., Brittonia. (London: Dioscorides Press, U.S). doi: 10.2307/2807470
Brundrett, M., Tedersoo, L. (2019). Viewpoints Misdiagnosis of mycorrhizas and inappropriate recycling of data can lead to false conclusions. New Phytol. 221), 18–24. doi: 10.1111/nph.15440
Cérantola, S., Lemassu-Jacquier, A., Montrozier, H. (1999). Structural elucidation of a novel exopolysaccharide produced by a mucoid clinical isolate of Burkholderia cepacia: Characterization of a trisubstituted glucuronic acid residue in a heptasaccharide repeating unit. Eur. J. Biochem. 260, 373–383. doi: 10.1046/j.1432-1327.1999.00171.x
Cescutti, P., Foschiatti, M., Furlanis, L., Lagatolla, C., Rizzo, R. (2010). Isolation and characterisation of the biological repeating unit of cepacian, the exopolysaccharide produced by bacteria of the Burkholderia cepacia complex. Carbohydr. Res. 345 (10), 1455–1460. doi: 10.1016/j.carres.2010.03.029
Chaintreuil, C., Rigault, F., Moulin, L., Jaffré, T., Fardoux, J., Giraud, E., et al. (2007). Nickel resistance determinants in Bradyrhizobium strains from nodules of the endemic New Caledonia legume Serianthes calycina. Appl. Environ. Microbiol. 73 (24), 8018–8022. doi: 10.1128/AEM.01431-07
Chen, Y. P., Rekha, P. D., Arun, A. B., Shen, F. T., Lai, W. A., Young, C. C. (2006). Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 34, 33–41. doi: 10.1016/j.apsoil.2005.12.002
Ciucanu, I., Kerek, F. (1984). A Simple and rapid method for the permethylation of carbohydrates. Carbohydr. Res. 131, 209–217. doi: 10.1016/0008-6215(84)85242-8
de la Fuente Cantó, C., Simonin, M., King, E., Moulin, L., Bennett, M. J., Castrillo, G., et al. (2020). An extended root phenotype: the rhizosphere, its formation and impacts on plant fitness. Plant J. 103 (3), 951–964. doi: 10.1111/tpj.14781
Etesami, H. (2018). Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues: Mechanisms and future prospects. Ecotoxicology Environ. Saf. 147, 175–191. doi: 10.1016/j.ecoenv.2017.08.032
Evlard, A., Campanella, B. (2013). Impact des éléments-traces métalliques sur les plantes et les techniques de phytoremédiation. Presses agronomiques Gembloux 1121, 59–75.
Ferreira, A. S., Leitao, J. H., Silva, I. N., Pinheiro, P. F., Sousa, S. A., Ramos, C. G., et al. (2010). Distribution of cepacian biosynthesis genes among environmental and clinical Burkholderia strains and role of cepacian exopolysaccharide in resistance to stress conditions. Appl. Environ. Microbiol. 76 (2), 441–450. doi: 10.1128/AEM.01828-09
Ferreira, A. S., Silva, I. N., Oliveira, V. H., Cunha, R., Moreira, L. M. (2011). Insights into the role of extracellular polysaccharides in Burkholderia adaptation to different environments. Front. Cell. Infect. Microbiol. 1, 16. doi: 10.3389/fcimb.2011.00016
Fu, B., Yan, Q. (2023). Exopolysaccharide is required for motility, stress tolerance, and plant colonization by the endophytic bacterium Paraburkholderia phytofirmans PsJN. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1218653
Galal, T. M., Sheata, H. S. (2015). Bioaccumulation and translocation of heavy metals by Plantago major L. grown in contaminated soils under the effect of traffic pollution. Ecol. Indic. 48, 244–251. doi: 10.1016/j.ecolind.2014.08.013
Gensous, S. (2014). Les champignons mycorhiziens à arbuscules des maquis miniers de la Nouvelle Calédonie : Diversité, rôle dans l'adaptation des plantes à la contrainte ultramafique et interaction avec des rhizobactéries promotrices de la croissance (Doctoral dissertation, Université de Nouvelle Calédonie). doi: 10.6098/2014NCAL0062
Gobat, J., Aragno, M., Matthey, W. (2010). Le sol vivant : bases de pédologie, biologie des sols (Polytechniques). P.P.
Goldstein, A. H. (1995). Recent progress in understanding the molecular genetics and biochemistry of calcium phosphate solubilization by gram negative bacteria. Biol. Agric. Horticulture 12, 185–193. doi: 10.1080/01448765.1995.9754736
Gonin, M., Gensous, S., Lagrange, A., Ducousso, M., Amir, H., Jourand, P. (2013). Rhizosphere bacteria of Costularia spp. from ultramafic soils in New Caledonia: Diversity, tolerance to extreme edaphic conditions, and role in plant growth and mineral nutrition. Can. J. Microbiol. 59 (3), 164–174. doi: 10.1139/cjm-2012-0570
Guentas, L., Gensous, S., Cavaloc, Y., Ducousso, M., Amir, H., de Ledenon, B. D. G., et al. (2016). Burkholderia novacaledonica sp. nov. and B. ultramafica sp. nov. isolated from roots of Costularia spp. pioneer plants of ultramafic soils in New Caledonia. Syst. Appl. Microbiol. 39 (3), 151–159. doi: 10.1016/j.syapm.2016.03.008
Gupta, P., Diwan, B. (2017). Bacterial Exopolysaccharide mediated heavy metal removal: A Review on biosynthesis, mechanism and remediation strategies. Biotechnol. Rep. 13, 58–71. doi: 10.1016/j.btre.2016.12.006
Hallack, L. F., Passos, D. S., Mattos, K. A., Agrellos, O. A., Jones, C., Mendonça-Previato, L., et al. (2010). Structural elucidation of the repeat unit in highly branched acidic exopolysaccharides produced by nitrogen fixing Burkholderia. Glycobiology 20 (3), 338–347. doi: 10.1093/glycob/cwp181
Hameed, A., Rasool, S., Azooz, M. M., Hossain, M. A., Ahanger, M. A., Ahmad, P. (2015). Heavy metal stress: plant responses and signaling. Plant metal interaction, 557–583. doi: 10.1016/B978–0-12–803158–2.00024–2
Herrera, A., Héry, M., Stach, J. E., Jaffré, T., Normand, P., Navarro, E. (2007). Species richness and phylogenetic diversity comparisons of soil microbial communities affected by nickel-mining and revegetation efforts in New Caledonia. Eur. J. Soil Biol. 43 (2), 130–139. doi: 10.1016/j.ejsobi.2006.12.001
Hossain, M. A., Piyatida, P., da Silva, J. A. T., Fujita, M. (2012). Molecular mechanism of heavy metal toxicity and tolerance in plants: central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 1–37. doi: 10.1155/2012/872875
Isnard, S., L’huillier, L., Rigault, F., Jaffré, T. (2016). How did the ultramafic soils shape the flora of the New Caledonian hotspot? Plant Soil 403, 53–76. doi: 10.1007/s11104-016-2910-5
Jaffré, T. (2023). Plant mineral nutrition on ultramafic rocks of New Caledonia. Bot. Lett. 170, 375–411. doi: 10.1080/23818107.2022.2080112
Kazakou, E., Dimitrakopoulos, P. G., Baker, A. J. M., Reeves, R. D., Troumbis, A. Y. (2008). Hypotheses, mechanisms and trade-offs of tolerance and adaptation to serpentine soils: From species to ecosystem level. Biol. Rev. 83 (4), 495–508. doi: 10.1111/j.1469-185X.2008.00051.x
Khatoon, Z., Huang, S., Rafique, M., Fakhar, A., Kamran, M. A., Santoyo, G. (2020). Unlocking the potential of plant growth-promoting rhizobacteria on soil health and the sustainability of agricultural systems. J. Environ. Manage. 273, 111118. doi: 10.1016/j.jenvman.2020.111118
Klonowska, A., Chaintreuil, C., Tisseyre, P., Miché, L., Melkonian, R., Ducousso, M., et al. (2012). Biodiversity of Mimosa pudica rhizobial symbionts (Cupriavidus Taiwanensis, Rhizobium mesoamericanum) in New Caledonia and their adaptation to heavy metal-rich soils. FEMS Microbiol. Ecol. 81 (3), 618–635. doi: 10.1111/j.1574-6941.2012.01393.x
Kumar, R., Shastri, B. (2017). “Role of phosphate-solubilising microorganisms in sustainable agricultural development,” in Agro-Environmental Sustainability: Volume 1: Managing Crop Health., (Cham: Springer International Publishing), 271–303. doi: 10.1007/978–3-319–49724-2
Lagrange, A. (2009). Etudes écologique et microbiologique des espèces du genre Costularia (Cyperaceae) pionnières des sols ultramafiques de Nouvelle-Calédonie: applications à la restauration écologique. (Doctoral dissertation, Université de Nouvelle Calédonie).
Lagrange, A., Ducousso, M., Jourand, P., Majorel, C., Amir, H. (2011). New insights into the mycorrhizal status of Cyperaceae from ultramafic soils in New Caledonia. Can. J. Microbiol. 57 (1), 21–28. doi: 10.1139/W10-096
Lagrange, A., Huillier, L. L., Amir, H. (2013). Mycorrhizal status of Cyperaceae from New Caledonian ultramafic soils : effects of phosphorus availability on arbuscular mycorrhizal colonization of Costularia comosa under field conditions. Mycorrhiza 23, 655–661. doi: 10.1007/s00572-013-0503-1
Larcher, M., Muller, B., Mantelin, S., Rapior, S., Cleyet‐Marel, J. C. (2003). Early modifications of Brassica napus root system architecture induced by a plant growth-promoting Phyllobacterium strain. New Phytol. 160 (1), 119–125. doi: 10.1046/j.1469-8137.2003.00862.x
Larridon, I., Verboom, G. A., Muasya, A. M. (2018). Revised delimitation of the genus Tetraria, nom. cons. prop. (Cyperaceae, tribe Schoeneae, Tricostularia clade). South Afr. J. Bot. 118, 18–22. doi: 10.1016/j.sajb.2018.06.007
L'Huillier, L., Jaffré, T., Wulff, A., Lebrun, M., Maggia, L., Barré, N., et al. (2010). Mines et environnement en Nouvelle-Calédonie: les milieux sur substrats ultramafiques et leur restauration. (Institut Agronomique Néo-Calédonien), 411.
Liu, J., Wang, X., Pu, H., Liu, S., Kan, J., Jin, C. (2017). Recent advances in endophytic exopolysaccharides: Production, structural characterization, physiological role and biological activity. Carbohydr. Polymers 157, 1113–1124. doi: 10.1016/j.carbpol.2016.10.084
Liu, Y., Bellich, B., Hug, S., Eberl, L., Cescutti, P., Pessi, G. (2020). The Exopolysaccharide Cepacian Plays a Role in the Establishment of the Paraburkholderia phymatum – Phaseolus vulgaris Symbiosis. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.01600
López-Bucio, J., Cruz-Ramírez, A., Herrera-Estrella, L. (2003). The role of nutrient availability in regulating root architecture. Curr. Opin. Plant Biol. 6, 280–287. doi: 10.1016/S1369-5266(03)00035-9
Losfeld, G., L’huillier, L., Fogliani, B., Jaffré, T., Grison, C. (2015). Mining in New Caledonia: environmental stakes and restoration opportunities. Environ. Sci. pollut. Res. 22, 5592–5607. doi: 10.1007/s11356-014-3358-x
Mishra, J., Singh, R., Arora, N. K. (2017). Alleviation of heavy metal stress in plants and remediation of soil by rhizosphere microorganisms. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.01706
Morcillo, R. J. L., Manzanera, M. (2021). The effects of plant-associated bacterial exopolysaccharides on plant abiotic stress tolerance. Metabolites 11 (6), 337. doi: 10.3390/metabo11060337
Naseem, H., Ahsan, M., Shahid, M. A., Khan, N. (2018). Exopolysaccharides producing rhizobacteria and their role in plant growth and drought tolerance. J. Basic Microbiol. 58 (12), 1009–1022. doi: 10.1002/jobm.201800309
Netrusov, A. I., Liyaskina, E. V., Kurgaeva, I. V., Liyaskina, A. U., Yang, G., Revin, V. V. (2023). Exopolysaccharides producing bacteria: A review. Microorganisms 11 (6), 1541. doi: 10.3390/microorganisms11061541
Norman, M. (2003) Biodiversity hotspots revisited. BioScience 53 (10), 916–917. doi: 10.1641/0006-3568(2003)053[0916:BHR]2.0.CO;2
Pascal, M., Deforges, B., Leguyader, H., Simberloff, D. (2008). Mining and other threats to the New Caledonia biodiversity hotspot. Conserv. Biol. 22 (2), 498–499. doi: 10.1111/j.1523-1739.2008.00889.x
Peña, M. J., Tuomivaara, S. T., Urbanowicz, B. R., O'Neill, M. A., York, W. S. (2012). “Methods for structural characterization of the products of cellulose-and xyloglucan-hydrolyzing enzymes,” in Methods in enzymology vol. 510 (Academic Press), 121–139. doi: 10.1016/B978–0-12–415931–0.00007–0
Pierre, G., Graber, M., Rafiliposon, B. A., Dupuy, C., Orvain, F., De Crignis, M., et al. (2012). Biochemical composition and changes of extracellular polysaccharides (ECPS) produced during microphytobenthic biofilm development (Marennes-Oléron, France). Microbial Ecol. 63, 157–169. doi: 10.1007/s00248-011-9959-8
Pierre, G., Zhao, J. M., Orvain, F., Dupuy, C., Klein, G. L., Graber, M., et al. (2014). Seasonal dynamics of extracellular polymeric substances (EPS) in surface sediments of a diatom-dominated intertidal mudflat (Marennes-Oléron, France). J. Sea Res. 92, 26–35. doi: 10.1016/j.seares.2013.07.018
Pikovskaya, R. I. (1948). Mobilization of phosphorus in soil in connection with the vital activity of some microbial species. Microbiologiya 17, 362–370.
Prabhu, N., Borkar, S., Garg, S. (2019). Phosphate solubilization by microorganisms: overview, mechanisms, applications and advances. Advances in biological science research, 161–176. doi: 10.1016/B978–0-12–817497–5.00011–2
R Core Team. (2021). R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing. Available at: https://www.R-project.org/.
Rubin, R. L., van Groenigen, K. J., Hungate, B. A. (2017). Plant growth promoting rhizobacteria are more effective under drought: a meta-analysis. Plant Soil 416, 309–323. doi: 10.1007/s11104-017-3199-8
Serrato, R. V., Sassaki, G. L., Cruz, L. M., Pedrosa, F. O., Gorin, P. A., Iacomini, M. (2006). Culture conditions for the production of an acidic exopolysaccharide by the nitrogen-fixing bacterium Burkholderia tropica. Can. J. Microbiol. 52 (5), 489–493. doi: 10.1139/w05-155
Silipo, A., Ierano, T., Lanzetta, R., Molinaro, A., Parrilli, M. (2008). The structure of the O-chain polysaccharide from the gram-negative endophytic bacterium Burkholderia phytofirmans strain PsJN. Eur. J. Organic Chem. 13), 2303–2308. doi: 10.1002/ejoc.200800135
Suárez-Moreno, Z. R., Caballero-Mellado, J., Coutinho, B. G., Mendonça-Previato, L., James, E. K., Venturi, V. (2012). Common features of environmental and potentially beneficial plant-associated burkholderia. Microbial Ecol. 63, 249–266. doi: 10.1007/s00248-011-9929-1
Upadhyay, S. K., Singh, J. S., Saxena, A. K., Singh, D. P. (2012). Impact of PGPR inoculation on growth and antioxidant status of wheat under saline conditions. Plant Biol. 14, 605–611. doi: 10.1111/j.1438-8677.2011.00533.x
Vejan, P., Abdullah, R., Khadiran, T., Ismail, S., Nasrulhaq Boyce, A. (2016). Role of plant growth promoting rhizobacteria in agricultural sustainability—a review. Molecules 21, 573. doi: 10.3390/molecules21050573
Wulff, A., LHuillier, L., Jaffré, T. (2010). “Espèces indigènes utilisables en revégétalisation,” in Mines et Environnement en Nouvelle-Calédonie : Les milieux sur substrats ultramafiques et leur restauration. (Nouméa: Institut Agronomique Néo-Calédonien), 231–344.
Yi, Y., Huang, W., Ge, Y. (2008). Exopolysaccharide: A novel important factor in the microbial dissolution of tricalcium phosphate. World J. Microbiol. Biotechnol. 24, 1059–1065. doi: 10.1007/s11274-007-9575-4
Keywords: PGPR, exopolysaccharides, biostimulant, Paraburkholderia, plant metal alleviation, ultramafic soil, phosphate solubilization
Citation: Bourles A, Pierre G, Amir H, Le Floc’h A, Chalkiadakis E, Médevielle V, Jourand P, Michaud P, Burtet-Sarramégna V and Guentas L (2024) The cepacian-like exopolysaccharide of Paraburkholderia ultramafica STM10279T enhances growth and metal adaptation of Tetraria comosa on New Caledonian ultramafic soil. Front. Plant Sci. 15:1349724. doi: 10.3389/fpls.2024.1349724
Received: 05 December 2023; Accepted: 13 May 2024;
Published: 05 June 2024.
Edited by:
Luz E. Bashan, Bashan Institute of Science, United StatesReviewed by:
Maria Elena Antinori, Catholic University of the Sacred Heart, ItalyLuis Ernesto Fuentes-Ramírez, Meritorious Autonomous University of Puebla, Mexico
Copyright © 2024 Bourles, Pierre, Amir, Le Floc’h, Chalkiadakis, Médevielle, Jourand, Michaud, Burtet-Sarramégna and Guentas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Linda Guentas, linda.guentas@unc.nc
†Present address: Alexandre Bourles, Pôle de Bactériologie, Groupe de Bactériologie Médicale et Environnementale, Institut Pasteur de Nouvelle-Calédonie, Nouméa, New Caledonia