Abstract
We address the puzzle of “unity in diversity” in human languages by advocating the (minimal) common denominator for the diverse expressions of transitivity across human languages, consistent with the view that early in language evolution there was a modest beginning for syntax and that this beginning provided the foundation for the further elaboration of syntactic complexity. This study reports the results of a functional MRI experiment investigating differential patterns of brain activation during processing of sentences with minimal versus fuller syntactic structures. These structural layers have been postulated to represent different stages in the evolution of syntax, potentially engaging different brain networks. We focused on the Serbian “middles,” analyzed as lacking the transitivity (vP) layer, contrasted with matched transitives, containing the transitivity layer. Our main hypothesis was that transitives will produce more activation in the syntactic (Broca's–Basal Ganglia) brain network, in comparison to more rudimentary middles. The participants (n = 14) were healthy adults (Mean age = 33.36; SD = 12.23), native speakers of Serbo-Croatian. The task consisted of reading a series of sentences (middles and transitives; n = 64) presented in blocks of 8, while being engaged in a detection of repetition task. We found that the processing of transitives, compared to middles, was associated with an increase in activation in the basal ganglia bilaterally. Although we did not find an effect in Broca's area, transitives, compared to middles, evoked greater activation in the precentral gyrus (BA 6), proposed to be part of the “Broca's complex.” Our results add to the previous findings that Broca's area is not the sole center for syntactic processing, but rather is part of a larger circuit that involves subcortical structures. We discuss our results in the context of the recent findings concerning the gene-brain-language pathway involving mutations in FOXP2 that likely contributed to the enhancement of the frontal-striatal brain network, facilitating human capacity for complex syntax.
Introduction
One of the most challenging puzzles about human languages is how to account for the immense diversity in their form and the equal ease with which all languages are learned by children, including those that are simultaneously acquiring two languages with very different grammatical properties (Aboh and Ansaldo, this volume). In this paper, we shed new light on this puzzle of “unity in diversity” by examining transitivity, i.e., a clause-structure phenomenon controlling how grammatical relations encode distinct participants in an event denoted by the verb. We report the results of an fMRI study investigating differential patterns of brain activation during processing of well-formed sentences with minimal versus fuller clause structural complexity. We look at this phenomenon from a language evolution perspective, conceptualizing sentences with different degrees of syntactic complexity as reflecting distinct stages in the evolution of language.
Relying on accepted postulates of theoretical syntax, Progovac (2015a, 2016a) proposed a gradual emergence of syntactic layers in language evolution, starting from the minimally complex intransitive small clause, gradually adding others, to accommodate transitivity, and verb finiteness. This precise reconstruction is achieved by peeling off the syntactic layers in (1), widely accepted (Adger, 2003; Carnie, 2013) to constitute the basic skeleton of the modern sentence1:
(1) VP/SC < vP < TP < CP
The lowest layer is the small clause (SC/VP), which accommodates a verb with one noun phrase (NP). The SC/VP is subsumed by the “little v” phrase (vP), an additional layer into which the traditional verb phrase is split (syntactic argumentation for this split is presented in the references above). This layer supports transitivity (i.e., addition of another argument, permitting a syntactic differentiation between agents and patients). The phrase that dominates the vP is Tense Phrase (TP), which accommodates the expression of tense and finiteness. TP can be dominated by the Complementizer Phrase (CP), which accommodates sentence embedding and wh-movement.
This approach offers a probe for investigating the sources of unity and diversity of human languages. Our argument is that the unity comes from the common (SC) core, which to a large extent determines further elaboration of structure, including its binary and its hierarchical nature. The diversity, on the other hand, comes from the options regarding which of the layers are utilized in which languages and in which constructions. This, for example, allows for a different sub-set of projections to be built in different languages, such as AspP instead of a TP. It also allows for coexistence of structures of varying degrees of syntactic complexity within a language, as per the analysis of Serbian transitive sentences (with a vP), and the corresponding middles (without a vP).
We seek support for this approach by using functional neuroimaging. In particular, we propose to capitalize on the coexistence of proto-syntactic “fossils” (i.e., constructions consisting of fewer and more basic layers of syntactic structure) alongside more modern structures (i.e., those operating with a fuller set of syntactic layers). We hypothesize that processing of structures with fewer syntactic layers relies on more ancient and more diffused neural networks, while highly layered structures require more recently evolved, densely interconnected and more specialized neural networks for quick and automatic processing (see also Ansaldo et al., 2015). We report the results of an fMRI experiment contrasting between the processing of constructions of differing structural complexity against each other in a reading recognition paradigm. We argue that the approach explored here brings together theories of syntax with theories of evolution, gives rise to specific testable neuroscientific hypotheses, and sheds novel light on the unity and diversity of human languages.
Scientific premises and hypotheses
Language variation: is there a common denominator?
The “unity in diversity” that underlies children's adaptability to various linguistic ecologies/environments is certainly of evolutionary significance, and we propose that the nature and trajectory of language evolution provides a key to understanding both the diversity and equal learnability of human languages. As previously noted, “language cannot be fully understood without reference to its evolution, whether proven or hypothesized” (Greenberg, 1966; Givón, 2002)2. The position that we advance here concurs with this statement: while we acknowledge that there are no definitive answers regarding how language evolved, there are specific hypotheses that can be used as tools to probe this question.
Our study focuses on transitivity: its emergence in language evolution, its typological variation in today's languages, and its representation in the brain. There is substantial diversity in the syntactic realization of transitivity across different languages. The conceptualization of sentence structure as described above provides a common denominator—the minimal intransitive clause—for the appearance of the various types of transitivity attested across languages: not only the nominative-accusative type (characterized by subjects receiving distinct case-marking irrespective of the presence of additional arguments, and in opposition to objects), but also ergative-absolutive type [in which agents of transitive verbs are marked distinctly (ergative case) from objects and single arguments of intransitive verbs (absolutive case)]. It also accounts for serial verb patterns, where two intransitive clauses, rather than just one, provide a foundation for constructing a single transitive sentence.
As discussed above, Progovac's (2015a; 2016a) reconstruction of the gradual emergence of syntactic layers in language evolution is based on certain well-established theoretical postulates of syntactic theory (e.g., Chomsky, 1995; Adger, 2003; Carnie, 2013), which include the (partial) hierarchy of syntactic layers for the modern sentence, as introduced in section 13:
In Figure 1 we present two sentences from English of varying degrees of syntactic elaboration. The example in (2) from English illustrates a typical transitive TP (Tense Phrase), where each argument is generated in its own layer: the agent (Maria) in the vP layer, and the patient (corn) in the VP/SC layer. Being the highest noun phrase in the structure, the agent moves to the TP layer to become the sentence subject. While the TP in (2) is a highly hierarchical structure, its bottom layer (VP/SC) is not hierarchical. Somewhere between the two extremes, one can construct a hierarchically less complex sentence in (3), which lacks the agent and the vP layer, and in which it is the patient (corn) that moves to TP to become the subject.
Figure 1
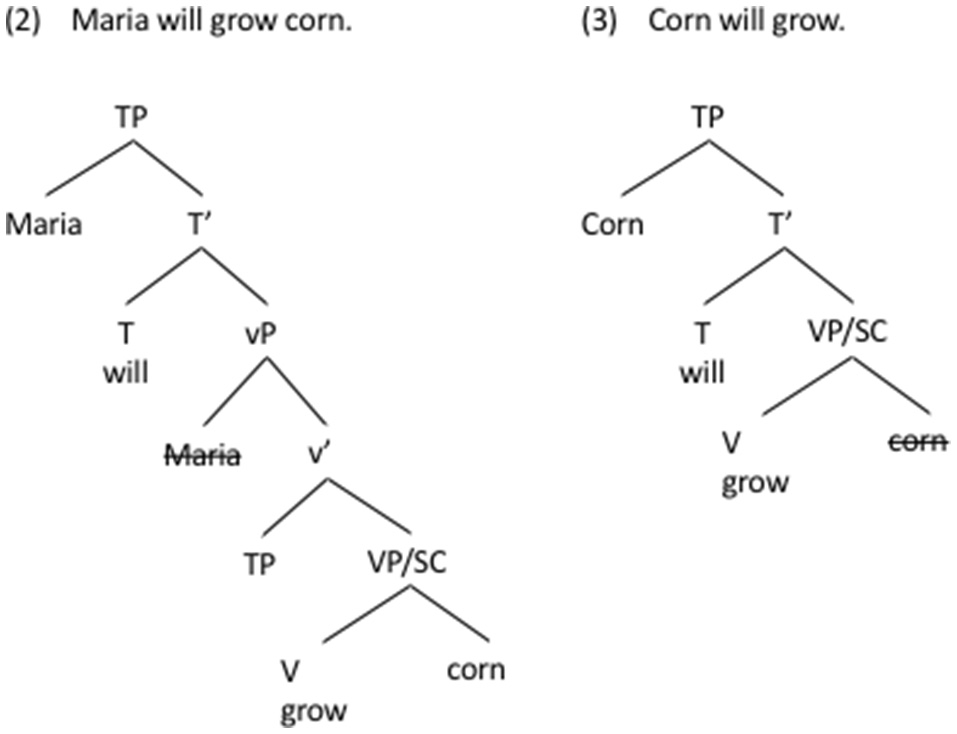
We present two sentences from English of varying degrees of syntactic elaboration. (2) illustrates a typical transitive TP (Tense Phrase), where each argument is generated in its own layer: the agent (Maria) in the vP layer, and the patient (corn) in the VP/SC layer. (3) is a hierarchically less complex, intransitive sentence, which lacks the agent and the vP layer.
This hierarchy, widely adopted in theoretical syntax, allows a precise reconstruction of the initial stages of syntax. By peeling off the syntactic layers postulated for the modern sentence down to the lowest layer, one arrives at the most basic clausal construct, the universal minimal structure—a VP/SC (Small Clause). The novelty of this proposal is that it isolates an intransitive absolutive-like two-slot grammar as a plausible precursor for all syntactic elaboration in the evolution of human language (and language acquisition)4. The absolutive-like stage exhibits an underspecified type of grammar capable of accommodating only a single argument, without grammatical means to distinguish subjects from objects. This is reminiscent of the modern absolutive case (contrasting with ergative case), (in) famous for blurring the distinction between subject-like and object-like arguments, as discussed in more detail below. Progovac (2016a) has proposed that this intransitive one-argument foundation is the common denominator for attested language variation in the expression of transitivity/argument structure, including ergative absolutive, nominative-accusative, and serial verb patterns (see also Mithun, 1991, for additional but much rarer possibilities).
In other words, the SC/VP provides the common core that languages share, and a starting point for syntactic elaboration, including for the various ways transitivity is expressed cross-linguistically. By itself simple and straightforward, this foundational structure can be tinkered with by enhancing it and/or reduplicating it in a variety of ways, leaving it up to the creativity of cultures to converge on a solution for transitivity. This is in the spirit of Mufwene's (2013) proposal that language variation in general boils down to different solutions to similar problems. This reasoning dates back to now famous evolutionary scholar, François Jacob, and his claim (1977) that different tinkerers likely develop different solutions to similar problems, such as different types of eyes. Natural selection tends to use materials at its disposal to form a variety of adaptations to similar challenges. Language variation in transitivity, we contend, can be understood in the same light. With rare exceptions, transitive structures typically add only one extra argument. In case of ergative-absolutive languages, the second, ergative-marked argument is “higher” than the first argument, while in nominative-accusative languages, the second, accusative-marked argument is “lower” than the first argument. In languages that have serial verb constructions, one finds multiple intransitive (small) clauses strung together, often just two. It is noteworthy that the enhancements of the foundational pattern are neither radical nor unrestricted, but rather modest and circumscribed. In this respect, they are consistent with typical recombination and replication processes that characterize evolution more generally.
Thus, this proposal provides a way to deconstruct the emergence of transitivity into specific small steps, consistent with typological variation, which in turn reveal communicative pressures and benefits for evolving transitivity. Importantly, we do not propose that the evolution of syntax is a sequence of discrete stages, where each stage limits one to only certain, endemic to that stage, grammatical structures. It is rather a patchwork quilt of partially overlapping, coexisting grammars, accrued throughout language evolution. We propose that the relatively uniform and modest beginnings for grammar, supported by more ancient and more diffuse neurocognitive processing strategies, were followed by gradual enhancements and enrichments of both the grammars and the neural networks that support them. We test this hypothesis in an fMRI experiment reported below.
It is fair to point out that gradualist approaches to evolution have been argued against in biology with a reference to the idea of punctuated equilibrium by Gould and Eldredge (Eldredge and Gould, 1972; Gould and Eldredge, 1977) which was interpreted by some as a refutation of the gradualist, incremental evolution of complex adaptation. However, contrary to this interpretation, Gould and Eldredge did not refute Darwinian adaptationism, nor proposed instantaneous evolutionary change. As discussed at length in Dawkins (1996) and Fitch (2010, pp. 46–53), situating Darwin's writings within the context of the debates of his own time, one can clearly see that Darwin was not a constant-rate gradualist, as is sometimes alleged by punctuated equilibrium advocates, but that his proposal merely allowed incremental changes. As Fitch (2010, pp. 48–49) explained, “gradual speciation and discrete mutation are not in conflict, but offer explanations at a different level of analysis,” the former at the level of the population, and the latter at the level the individual. Thus, we maintain that there are no obstacles coming from biology for studying language evolution as proceeding in small, incremental steps. This approach to the evolution of language is consistent with the many-genes-of-small-effects approach, pursued more recently by Fisher (2017); cf. also Dediu (2015) and Dediu and Ladd (2007). We also contend that there are no such obstacles coming from linguistics either, even though, understandably, there are opposing views5. The syntactic analysis we adopt as our starting point here is also in sync with a recent development in the field of linguistic typology. Interestingly, typological research has found it useful to steer away from formulating implicational universals based on transitive structures, such as SOV, SVO, VSO, etc., as originally formulated in Greenberg (1966). Instead, more precise and more insightful universals can be formulated by breaking down transitivity into binary parameters, SV/VS and OV/VO, as they are better predictors of various correlations (Dryer, 1992, 1997; Croft, 2003). This is another argument in favor of decomposing transitivity into smaller structures, with verb and only one argument at a time. We believe that our approach brings together insights and analyses from a variety of different frameworks and approaches to language, each illuminating its different facets.
The co-existence of “linguistic fossils” with more modern syntax
Jackendoff (1999, 2002) introduced the influential idea of linguistic fossils, i.e., structures occurring in present-day languages, hypothesized to approximate ancient language. Jackendoff considers compounds (e.g., snowman, scarecrow), as well as other “loose” combinations, such as adjunction processes (e.g., the attachment of adjectives and adverbs), as good candidates for syntactic fossils. Progovac (2015a, 2016a) builds on this idea to argue that intransitive absolutive-like structures are available in various guises as “living fossils” in modern languages, and that these structures also serve as the foundation upon which a modern sentence gets constructed. The relevant fossils include, but are not limited to, sentences with one absolutive argument in an ergative language such as Tongan (4), as well as the so-called “middles” in an accusative language such as Serbian (6). The fossils in (4, 6) are argued not to project the specialized transitivity layer (such as a vP), rendering them “flatter” than their transitive counterparts, in the sense that they project fewer layers of syntactic structure. In other words, the structures of (4, 6) are comparable to the structure in (3) in Figure 1, lacking a vP, and not to the structure in (2) in Figure 1, which projects a vP. Moreover, the only argument in (4, 6) is not thematically specified as either agent or patient, often resulting in vagueness, as indicated by multiple translations, and as captured semantically in (5, 7).
(4) Oku ui ‘a Mele. (Tchekhoff, 1973, p. 283).
pres call abs Mary
‘Mary calls.'/ ‘Mary is called.'
(5) ∃ e [C(e) ∧ Participant (Mary,e)]
[i.e.: There is an event e such that e is Calling, and
Mary is a Participant in e.]
(6) Deca se tuku.
childrense hit
‘Children hit/are hitting each other.'
‘Children hit/are hitting themselves.'
‘Children hit/are hitting somebody (else)/me/us.'
‘One hits/is hitting children.'
(7) ∃e [H(e) ∧ Participant (Children, e)]
[i.e: There is an event e such that e is Hitting, and
Children is the Participant in e.]
Gil (2005) has made a comparable argument for certain one-argument sentences in Riau Indonesian, where there is no case marking (8). He has proposed that their interpretation is simply achieved by an association operator, which associates the meanings of the two words/concepts6.
(8) Ayam makan
chicken eat
‘The chicken is eating.'
‘Somebody is eating the chicken.'
What is surprising about these one-argument sentences/clauses is that there is no structural/grammatical differentiation between the agent (subject) and the patient (object). As pointed out in Tchekhoff (1973) for Tongan, it would be erroneous to analyze the distinct interpretations in (4) as involving two distinct syntactic structures, one in which Mary is the agent/subject, and the other in which Mary is the patient/object. Absolutive agents and absolutive patients in Tongan are clearly unified grammatically into a single, absolutive role. Instead, Tchekhoff (1973, p. 283) characterizes the pattern in (4) as vague/unspecified, where Mary is neither an agent nor a patient, and the two available translations in (4) just reflect a nominative-accusative bias. In his own words, in (4) “Mary is the only determiner [i.e., argument, L.P.], and the whole utterance gives us only the following information: present tense, verb call, Mary. And we don't know whether Mary was the agent of the calling, or the recipient of it. Nothing in the Tongan original informs us as to this particular point.” If so, then (4) should be analyzed semantically as in (5), where Mary is just an underspecified proto-participant role in the event of calling. Dowty (1991) has advanced the idea of proto-roles for proto-agents and proto-patients. The idea is that proto-agents range over a variety of roles along the agent (volitional) continuum, and that proto-patients range over a variety of roles on the patient (affected) continuum, with the boundary between the two macro roles not rigidly demarcated. The data and analysis presented above lead to conclusion that language can operate with an even more general proto-role: proto-participant role.
Similar considerations hold of Serbian middles, as per the discussion and analysis in Progovac (2015a; 2015b). These se constructions involving just one argument are thematically unspecified, and (6) can be interpreted in a variety of ways that involve an event of hitting, and children as participants in that event, as semantically captured in (7). Which interpretation will be salient depends on the context and the pragmatic plausibility, but is not determined by the grammar (see Marelj 2004, who also discusses some aspectual restrictions on the interpretation of middles). The only expressed argument can be either agent-like or patient-like, or even both at the same time, in the case of the reflexive interpretation.
Importantly, Serbian (6) sharply contrasts with the transitive accusative counterpart in (9), which has only one interpretation, and which is standardly analyzed as projecting the transitivity (vP) layer. In this respect, the example in (9) corresponds to the structure in (2) in Figure 1, rather than to the structure in (3) in Figure 1, which lacks the vP.
(9) Deca me/ga tuku.
children me/him hit
‘The children are hitting me/him.'
(10) ∃e [H(e) ∧ Participant (Children,e) ∧ Patient
(Me/Him,e)]
[i.e., There is an event e such that e is Hitting, and
Children is the Participant in e, and Me/Him is the
patient in e.]
Here the accusative argument (me/him) is added from below, to instantiate a patient/object, which then renders the other participant, the unmarked (nominative) argument, as a thematically higher argument, in this case agent/subject. This analysis allows the proto-layer to retain its underspecified character, with one proto-participant argument, given that the specification of the second, added argument is sufficient to disambiguate.
A comparable (but opposite) situation holds in Tongan, where an extra (ergative) argument is a higher argument (in this case agent), added on the top, while the other participant, the unmarked (absolutive) argument, is rendered as a thematically lower argument (in this case patient). Again, the analysis allows for the proto-layer to be preserved as essentially intact, with one proto-participant argument, given that the added (ergative) argument is sufficient for disambiguation.
(11) Oku ui ‘e Sione ‘a Mele
pres call erg John abs Mary
‘John calls Mary.'
(12) ∃e [C(e) ∧ Agent (John,e) ∧ Participant (Mary,e)]
[i.e.,: There is an event e such that e is Calling, and
John is the Agent in e, and Mary is a Participant
in e.]
It is significant that both patterns (ergative and accusative) have as their foundation an intransitive small clause with one verb and one unmarked argument, a proto-participant in the event, which reflects the initial stage of syntax as reconstructed in Progovac (2015a, 2016a)7.
This evolutionary characterization of ergative and accusative cases as secondary, tacked-on cases, is well aligned with the Dependent Case Theory explored in Yip et al. (1987), and Marantz (1991), the approach that has recently been revived in the work of McFadden (2004) and Baker (2015), among others. According to Marantz (1991, p. 24), “dependent case is what we will call accusative and ergative… Accusative is the name for the dependent case that is assigned downward to an NP position…Ergative is the name for the dependent case assigned upward to the subject position….” This same insight was adopted in Baker (2015, pp. 48–49). In other words, accusative and ergative are cases dependent on the presence of another (first) argument. The evolutionary approach explored here can shed light on various otherwise unexplained phenomena, including on the small clause foundation for all sentences, and on the nature of language variation in the expression of transitivity.
The following question arises in this respect: why would this small clause foundation be there at all, and why would it still be there even today, after many millennia of evolution? There are two reasons. First, evolution of complex adaptations typically takes place gradually, in incremental small steps, which would explain why the small clause step was useful, or even necessary, in the evolution of human language. Gradual evolution is often described as a “tinkering” process, where new, slightly more complex elements are only created by tinkering with the existing elements. In his highly influential and insightful essay, Jacob (1977) maintains that natural selection's creative force lies in its ability to combine and recombine old material into novelties, in a process that resembles imperfect tinkering, rather than engineering (see also Fitch, 2010, p. 55, who reinforces this view). Crucially, as Jacob points out, evolution hardly ever produces complex traits from scratch (see also Pinker and Bloom, 1990), which is why the small clause one-argument beginnings may have provided necessary scaffolding for the assembly of more complex syntactic patterns. Second, if the basic small clause was a useful and robust stage in the evolution of human language, then rather than being discarded, it has persisted as a necessary foundation/scaffolding that facilitates the elaboration/tinkering of additional structure, including in language acquisition.
Neuroscientific hypotheses: transitivity and the basal ganglia in an fMRI experiment
The coexistence of proto-syntactic fossils with more modern syntactic structures in present-day languages provides a fertile ground for testing evolutionary hypotheses using neuroimaging. Our study focused on the Serbian middles contrasted with canonical transitive structures; both are highly productive in Serbian. As discussed above, in contrast to the flatter middles, the transitives in Serbian are analyzed as projecting an additional layer of structure, an active vP layer, which renders them more hierarchical. We hypothesized that Serbian middles, relative to matched transitive accusative structures, will result in reduced activation in regions associated with syntactic processing, specifically in the Broca's–Basal Ganglia network. Our main hypothesis ties directly into the findings that the connectivity of the Broca's–Basal Ganglia network was bolstered relatively recently in evolution, in the line of descent of humans (see e.g., Enard et al., 2009; Hillert, 2014; Dediu, 2015; for the role of FOXP2 and other genes in this respect). We also hypothesized possible increased activation in regions not typically associated with syntactic processing when examining the reverse, that is, the activation in middles relative to full transitive structures. This would be consistent with Bisang (2009) and Ansaldo et al. (2015)'s claims that syntactically simpler structures may activate more semantic processing.
The basic assumptions behind these predictions are the following. On the one hand, complex grammatical patterns, which arguably only human brains can process, are supported by the most recently evolved/enhanced neural networks. To put it differently, complex hierarchical structures require more support from these recently evolved networks than their flatter proto-syntactic counterparts. On the other hand, the vague/underspecified structures of proto-syntax, which need less support from these most recently evolved networks, may show a more diffuse and less streamlined distribution across the brain, as well as possibly more individual variability.
Whereas middles in Serbian show ambiguity/underspecification with respect to the thematic role of their only expressed argument (cf. e.g., Deca se tuku “Children SE hit”) and canonical transitives are unambiguous in this respect (cf. Deca me tuku “Children me hit”), it is possible to skew the semantic interpretation toward the “me” readings in middles by selecting exclamatives (13, 14) or imperatives (15, 16), rendering them as close semantic and phonological matches to transitives, differing primarily in the level of syntactic layering/elaboration. The examples in (13, 15) are middles, and the examples in (14, 16) are the corresponding transitives.
(13) a. Mama, Milanseudara!
Mom, Milan se hits
‘Mom, Milan is hitting (me)!
b. Mama, ovajpasseujeda!
Mom, this dog se bites!
‘Mom, this dog is biting (me)!'
(14) a. Mama, Milanmeudara!
Mom, Milan me hits
‘Mom, Milan is hitting me!'
b. Mama, ovajpasmeujeda!
Mom, this dog me bites!
‘Mom, this dog is biting me!'
(15) a. Neudarajse!
not hit.impse
‘Don't hit (me)!
b. Negurajse!
not push.impse
‘Don't push (me)!'
(16) a. Neudarajme!
not hit.imp me
‘Don't hit me!'
b. Negurajme!
not push.imp me
‘Don't push me!'
Method
Participants
We tested 14 healthy adults, native Serbo-Croatian speakers (6 Female, age range 21–62, Mean = 33.36, SD = 12.23) residing in the US. Participants' self-reported native fluency in Serbo-Croatian was verified by the first author, a Serbian native speaker, who conducted both oral and written interviews with each participant. All the participants were also fluent in English, with 11 participants out of 14 clearly exhibiting non-native command of English8. Participants were screened prior to the study and reported no language impairments and no abnormal psychiatric or neurological history. All participants were determined to be right-handed by the Edinburgh Handedness Inventory (Oldfield, 1971). Written informed consent was obtained from all participants, as required by Wayne State University IRB.
Stimuli and procedure
Our main hypotheses concerned neural correlates of processing middles compared to transitives in Serbian. Each of the two conditions (Middles and Transitives) consisted of two types of sentences: two types of middles, exclamative (see 13 above) and imperative (see 15 above), as constructions most semantically matched to their transitive counterparts, and two types of transitives, also exclamative (see 14 above) and imperative (see 16 above), with a total of 64 unique stimuli used across the two conditions (see Appendix for complete list of stimuli used in this study). Stimuli were presented visually as centered white text on a black background (Times New Roman, 80-point font). Each stimulus was presented for 1,500 ms followed by a 250 ms fixation cross. Stimuli were presented in a block-design, with each block consisting of 8 unique stimuli. There were 4 blocks per each of the 2 conditions of interest. To ensure that participants adequately engaged with the stimuli, we embedded a simple repetition detection task (1-back) in each block, so that one of the stimuli was presented twice in succession and participants were asked to indicate with a button press when such repetition occurred. Each block lasted a total of 15.75 s, and was followed by 10 s inter-block-interval, during which a fixation crosshair was presented in the middle of the screen. Additional conditions were included in the imaging design but are not the focus of this study. In these other conditions, 144 other unique stimuli were presented in 18 blocks with stimuli consisting of either small clauses, full sentences, simple compounds, or complex compounds. The order of blocks presentation was pseudorandomized to minimize condition order effects. Two pseudorandom orders were created and assigned to participants based on whether they had an even or an odd numbered study ID.
Presentation of the stimuli and recording of responses was conducted with PsychToolbox in MATLAB. Accuracy and reaction times were collected and averages were calculated per condition and across participants. Due to technical difficulties accuracy and reaction times could not be calculated for 3 participants; these participants provided complete fMRI data sets and therefore data from these participants were included in all neuroimaging analyses. In addition, data form one other participant was excluded due to noncompliance, as it was evident the participant was sleeping during the scanning session.
MRI data acquisition and analyses
MRI data were acquired in a 3T Siemens Verio scanner at the Wayne State University MR Research Facility located in Harper University Hospital in Detroit, MI. T1-weighted whole-brain structural images were acquired using an MP-RAGE sequence (176 coronal slices, repetition time (TR) = 1,680 ms, echo time (TE) = 3.51 ms, flip angle = 9°, field of view = 256 mm, 176 × 256 voxels, and voxel size = 0.7 mm × 0.7 mm × 1.3 mm). Functional images during the experimental task were acquired in a single run using a T2*-weighted multiband accelerated EPI pulse sequence: 75 slices parallel to the AC-PC plane (TR = 2,000 ms, TE = 30 ms, flip angle = 90° voxel size = 2 × 2 × 2 mm3, multiband factor = 3, duration = 11 min and 18 s, total 339 volumes).
Functional data were analyzed using SPM12 package (Wellcome Department of Imaging Neuroscience, London, UK). Images were motion corrected, normalized to the Montreal Neurological Institute (MNI) template, and smoothed with a 6-mm full-width half-maximum Gaussian kernel. Statistical analyses of fMRI data were conducted using general linear modeling (GLM), as implemented in SPM12. First-level analyses included conditions of interest: Middles, Transitives and other conditions not of interest in this study (Small Clauses, Full Sentences, Simple Compounds, Complex Compounds). All conditions were modeled with separate regressors. The BOLD response was modeled by convolving a canonical hemodynamic response (HDR) function with a boxcar function spanning the duration of the block (15.75 s) and temporal derivatives of each block were included in the GLM to account for temporal shifts in the response of the stimuli (Friston et al., 1998).
We assessed activation associated with the processing of Middles compared to Transitives in a priori anatomically-defined regions of interest (ROIs) that are part of the known specialized language network involved in syntax-related processing (i.e., Broca's area and the basal ganglia). Four anatomically defined ROIs were generated using the Wake Forest University Pickatlas tool (TD Brodmann areas) spanning the left and right inferior frontal cortex regions identified as Brodmann Area (BA) 44 and 45, and the left and right basal ganglia (combined caudate and putamen). These ROIs were used to extract average activation per condition. Planned comparisons were conducted with the mean extracted contrast values from each of the four ROIs described above (all reported findings from these analyses were significant at p < 0.05). We tested significant effects using a paired t-test (2-tailed), Bonferonni-corrected for multiple comparisons.
In addition, with group-level whole-brain analyses we set to identify the regions in which there was an increase in activation for Transitives compared to Middles, and to examine whether regions evinced the opposite trend, an increase associated with the processing of flatter structures, which may rely on more general cognitive processing strategies available prior to the emergence of complex human language, and the concomitant modification of the human brain. Thus, the contrasts of interest—Middles versus Transitives—were computed on the individual level and combined in a whole brain group-level analysis. Group-level activation maps were threshold at p < 0.001 at the voxel level, with an extent threshold of 100 contiguous voxels for the cluster level, minimizing the possibility of the findings being false positives.
Results
Behavior
Overall, participants were highly accurate in responding to repeated stimuli (mean ± SD: 0.98 ± 0.03), suggesting that participants were engaged and processing the presented stimuli. Accuracy rate in this detection task did not differ between Middles (1.00 ± 0.00) and Transitives (0.98 ± 0.08), t(9) = 1.00, p = 0.34. Mean reaction time in the detection task was slightly longer [t(9) = 2.43, p = 04] in the Middles (640 ± 160 ms) compared to the Transitives condition (590 ± 140 ms), suggesting the possibility of increased demands for processing Middles compared to Transitives.
Reduced brain activation for middles compared to transitives: syntax-related rois
Parameter estimates from a priori anatomically identified regions known to be involved in the processing of syntax were used in the main analyses. Specifically, we extracted parameter estimates for Middles and Transitives for 4 ROIs: bilateral BA 44/45, and bilateral basal ganglia. Extracted values per each condition are presented in Figure 2 to allow comparisons across conditions. Statistical tests were only conducted in predefined comparisons using t-test in bilateral basal ganglia, and bilateral BA 44/45.
Figure 2
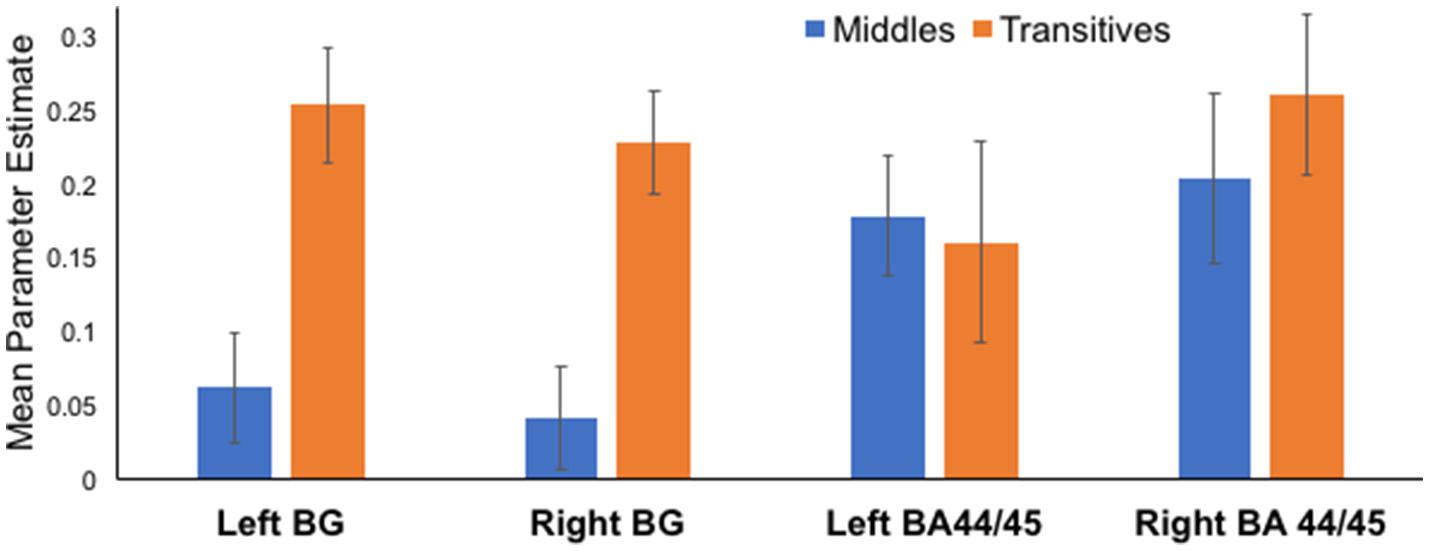
Differential activation for Middles compared to Transitives in basal ganglia and BA 44/45 ROIs. In both left and right basal ganglia ROIs there was marked increase in activation for Transitive compared to Middles. Activation in left and right BA44/45 ROIs was associated with the processing of both types of stimuli.
Processing of Transitives, compared to Middles, was associated with an increase in activation in the basal ganglia ROIs both on the left [Transitives: 0.25 ± 0.14, Middles: 0.06 ± 0.13, t(12) = 5.64, p < 0.001] and the right [Transitive: 0.23 ± 0.12, Middles: 0.04 ± 0.12, t(12) = 4.57, p = 0.001]. In contrast, in BA 44/45 ROIs activation was evidenced for both Transitives and Middles with no significant differences between conditions in the left [Transitives: 0.18 ± 0.20, Middles: 0.18 ± 0.14, t(12) = 0.25, p = 0.83], and a non-significant trend in the right [Transitives: 0.20 ± 0.17, Middles: 0.13 ± 0.18, t(12) = 1.87, p = 0.09 uncorrected].
Reduced brain activation for middles compared to transitives: whole-brain analyses
To confirm the involvement of language related regions in the processing of Transitives compared to Middles, we used a planned exploratory analysis conducted at the whole-brain level by created group-level model of the first-level, individual subject contrasts of activation associated with the processing of Transitives compared to Middles. Brain regions in bilateral basal ganglia, as well as regions in the precentral gyrus (BA 6) and visual cortex including bilateral lingual gyrus (BA 30) showed more activation for Transitives compared to Middles (Table 1; Figure 3). BA 6 plays a role in planning coordinated movements, and has also recently been linked to language related processing (Kambara et al., 2017; Nishida et al., 2017). Indeed, according to Ardila et al. (2016b), “‘Broca's complex’ includes not only left BA44 and BA45, but also BA46, BA47, partially BA6 (mainly its mesial supplementary motor area) and extending subcortically toward the basal ganglia and the thalamus.”
Table 1
| Hemisphere | Region | BA | MNI coordinates | T Values | Number of voxels | ||
|---|---|---|---|---|---|---|---|
| x | y | z | |||||
| A. TRANITIVES > MIDDLES | |||||||
| Right | Putamen | NA | 24 | 16 | 2 | 7.33 | 103 |
| Caudate | 12 | 10 | 4 | 5.25 | |||
| Left | Caudate | NA | −6 | 6 | 4 | 7.06 | 281 |
| Putamen | −18 | 10 | 4 | 6.82 | |||
| Right | Precentral gyrus | 6 | 36 | −6 | 42 | 11.20 | 308 |
| Left | Precentral gyrus | 6 | −30 | −4 | 62 | 6.84 | 383 |
| Left | Medial frontal gyrus | 6 | 0 | 10 | 60 | 8.89 | 459 |
| Right | Middle temporal gyrus | 39/19 | 42 | −60 | 10 | 7.97 | 132 |
| Left/Right | Lingual syrus | 30 | −4 | −72 | 4 | 10.61 | 1511 |
| Right | Cerebellum | NA | 42 | −52 | −30 | 7.51 | 139 |
| B. MIDDLES > TRANSITIVES | |||||||
| No voxels survived the threshold | |||||||
Brain activations associated with processing Serbian Transitives compared to Middles.
p < 0.001, 100 contiguous voxels.
Figure 3
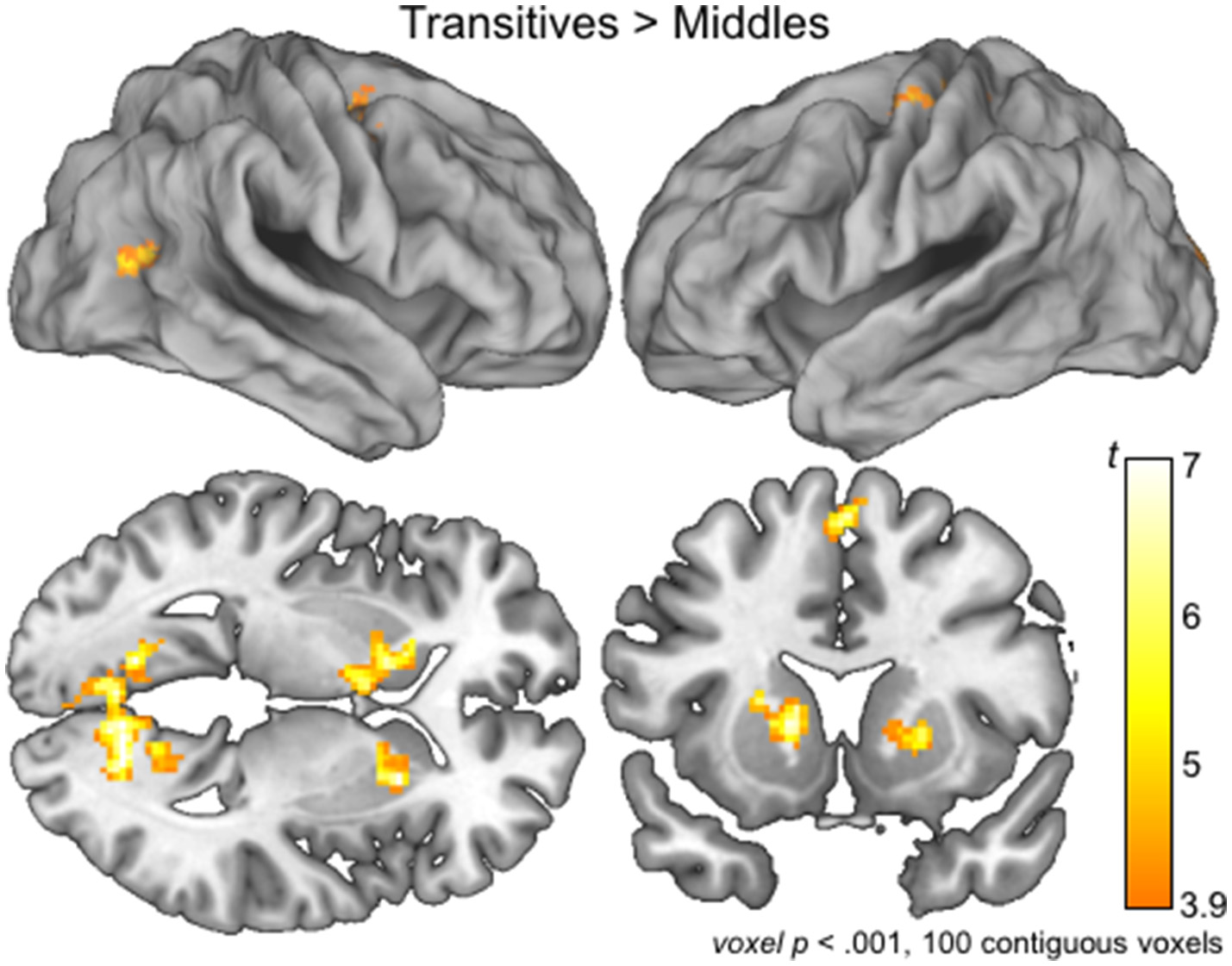
Brain activations associated with the processing of Serbian Transitives compared to Middles. Group level whole brain activations maps for the contrast Transitive > Middles are overlaid on lateral view of the right (top, left) and left (top, right) hemispheres, and on a sagittal (bottom, left) and coronal (bottom, right) slices at the level of the basal ganglia. See Table 1 for detailed list of activations.
Whole-brain group-level analysis allowed us to assess the possibility of unique regions supporting the processing of Middles when compared to Transitive syntactic structures. To test this possibility, we used the contrast to identify increased activations associated with the processing of Middles compared to Transitives. This contrast did not identify any brain regions in which activation was increased for Middles compared to Transitives. These findings are consistent with a possibility that processing of middles relies on strategies that may be different in different individuals and therefore no consistent findings can be identified at the group level.
Discussion
We found support for our main hypothesis that processing of less hierarchical structures would be related to reduced activation in the brain networks associated with syntactic processing, namely in basal ganglia. Although we did not find an effect in Broca's area, transitives, compared to middles, evoked greater activation in the precentral gyrus (BA 6), proposed to be part of the “Broca's complex” (e.g., Ardila et al., 2016b). Our finding of the role of basal ganglia is consistent with a number of previous studies, which found that Broca's area is not the sole center for syntactic processing, but rather is part of a larger circuit that involves subcortical structures (Gibson, 1996; Lieberman, 2000, 2009; Vargha-Khadem et al., 2005; Ullman, 2006; Ardila et al., 2016a,b). While in this study we did not find differential activation in the Broca's region, in a related study we found greater activation in both Broca's region and basal ganglia in the more complex syntax condition (Progovac et al., in preparation). There we investigated the processing of English small clauses (SCs) (e.g., Point taken; Problem solved) relative to their tensed sentential counterparts (TPs) (e.g., The point is taken; The problem is solved). Increased activation was found in the left BA 44 and in the right basal ganglia for more hierarchical tensed clauses relative to the flatter small clauses, even after controlling for the sentence length effect, indicating that greater activation in these regions was not merely due to differences in the length of word strings, but rather was likely related to the presence of extra layers of syntactic structure9.
The key difference between that study and the experiment reported in the current article was that the latter tested the impact of the vP (transitivity) layer, while the experiment with English clauses tested the impact of the TP layer, as well as the DP layer, which is required by the TP in English. In other words, unlike the small clauses in (17), the TP sentences require their subjects to be DPs, as indicated in the contrast between (18) and (19) below (see Progovac, 2013 and references cited there for the details of the analysis of these clauses).
(17) Problem solved. Point taken. Mission accomplished.
(18) The problem is solved. The point is taken. The mission is accomplished.
(19) *Problem is solved. *Point is taken. *Mission is accomplished.
We offer the following tentative suggestions for the difference in the findings between the two experiments regarding the Broca's area. While the two findings overlap in their isolation of the involvement of basal ganglia, only the TP/DP condition has also identified an effect in the Broca's area. One possible explanation may be that even though the vP layer is hierarchical, it is more basic and less abstract than the TP or DP layers, as it still deals with content words, nouns and verbs. The TP and DP layers involve abstract, purely grammatical categories, such as tense auxiliaries (is, was) and articles (a, the). While transitivity can be seen as instantiating hierarchy, but not a high level of functional abstraction, the TP and DP layers instantiate both hierarchy and a high level of functional abstraction, potentially resulting in higher processing demands. Thus, it is plausible that the transitivity (vP) layer, being a more basic (lower) layer of structure and less functional/less abstract in nature than TP/DP layers, is less reliant on Broca's region and/or requires less dense connectivity between the Broca's region and basal ganglia.
Another possible explanation for the different findings may be the degree to which the constructions that we examined in the two experiments differed from each other. Namely, while the Serbian vP condition (transitives vs. middles) differed only in one layer of structure (the vP layer), the TP/DP condition in English differed in two layers of structure (TP and DP). Perhaps the layering of functional projections has a cumulative effect on processing, with more such layers requiring more neuronal interconnectivity (see also Footnote 10). Future experiments can be designed to tease these options apart, and we suspect that a variety of languages, as well as a variety of constructions, will need to be tested before a clear picture emerges.
Our finding of the differential involvement of basal ganglia in the processing of more complex syntax is especially significant in light of the recent studies that have identified the BA 44–Basal Ganglia network as a syntactic processing network with very strong neural interconnectivity (Ardila et al., 2016a). The basal ganglia are a collection of subcortical structures on both sides of the thalamus, outside and above the limbic system, below the cingulate gyrus and within the temporal lobes. The largest group of these structures is the striatum, consisting largely of the caudate nucleus and the putamen. The basal ganglia are highly interconnected to cortical regions, especially in the frontal lobes, including Broca's, via parallel anatomically and functionally segregated “loops” (Draganski et al., 2008; Frey et al., 2008; Ford et al., 2013). The involvement of the striatum in syntactic processing was demonstrated across different languages in lesion studies, including in patients with Parkinson's and Huntington's disease (Moro et al., 2001; Teichmann et al., 2005, 2008; Newman et al., 2010). Furthermore, studies produced experimental evidence for a language-basal-ganglia-gene pathway using animal models. For example, in one study (Enard et al., 2009), two amino acid substitutions, thought to have been positively selected for in the course of human evolution because of their enhancing effect on language, were introduced in the FOXP2 gene of mice. These genetic mutations did not result in alterations in the general health of mice, but triggered changes in their ultrasonic vocalizations, exploratory behavior and decreased dopamine concentrations in the brain. Most relevant for our study, the humanized FOXP2 alleles affected the basal ganglia, increasing dendrite lengths and synaptic plasticity of the medium spiny neurons in the striatum.
Our study offers some encouraging preliminary evidence, in need of further confirmation, for the proposal that relatively uniform and modest beginnings for human language (i.e., proto-syntax prior to the emergence of more hierarchical grammar), supported by more ancient brain processing strategies, were followed by a (gradual) enrichment of the grammar, accompanied by more streamlined processing strategies and more densely interconnected neural networks needed to support them (see also Ansaldo et al., 2015). Our results and the approach we explore here are compatible with the idea that recent genetic mutations, including in FOXP2, in the line of descent of humans, increased synaptic plasticity and neuronal connectivity of the human brain (e.g., Hillert, 2014; Dediu, 2015), particularly in the frontal-striatal network, enabling human capacity for more complex language. This is where our proposal addresses the language-brain-gene connections.
The involvement of FOXP2 in language and frontal-striatal brain network was directly documented by a discovery that showed that individuals heterozygous for FOXP2 alleles with a certain mutation affecting the forkhead DNA binding domain of the protein suffered from a developmental impairment affecting speech and language (Lai et al., 2001). In the widely publicized case of the KE family, whose congenital language impairment was caused by a mutation in the FOXP2 gene (Fisher et al., 1998), language deficits included impaired understanding of complex syntax and grammatical morphology (Gopnik, 1990) and difficulties with articulation, among several other symptoms. The morpho-syntactic deficits included subject drop and the nonsystematic use of plural forms and tense (e.g., Gopnik and Crago, 1991; see also Piattelli-Palmarini and Uriagereka, 2011). This implicates problems with higher functional categories, including tense and TP. Liégeois et al. (2003) showed that the affected KE members not only exhibited under-activation in the Broca's area and its right homolog, but that both the caudate nucleus and putamen were sites of morphological abnormality in the affected members. Impairments in the members of the KE family were not restricted to language. In addition, the affected family members had symptoms of verbal apraxia and lowered nonverbal cognition (Vargha-Khadem et al., 1995), suggesting complex wide-ranging effects of the FOXP2 gene.
Recent comparative work that examined linguistic phenotypes across developmental disorders affecting language acquisition converged on the observation that narrow syntax (i.e., the internal computational system) tends to be more resilient than usually thought, i.e., likely to be preserved in many forms of developmental pathology, which by and large affect linguistic phenomena related to externalization, i.e., morphosyntax and morphophonology, linguistic domains that interface with the Intentional-Conceptual and Sensory-Motor systems (The Locus Preservation hypothesis; Leivada et al., 2017; Lorenzo and Vares, 2017)10. We would argue that dissociation in the patterns of language breakdown may reflect distinct underlying brain network organization corresponding to distinct evolutionary stages during which these linguistic domains emerged.
Further investigations in the function of FOXP2 and developmental language disorders revealed that FOXP2, a regulator gene, is not the “grammar” or even the “language gene”, given that (i) it is expressed not only in multiple areas of the brain, but also in other organs, serving a variety of functions, and, that (ii) most common cases of developmental language disorders that entail grammatical impairment do not involve mutations in FOXP2 (Rakhlin and Grigorenko, 2015). However, it is clearly part of the genetic network relevant to language. Since the discovery of FOXP2, it has become increasingly clearer that genetic bases of language are complex and involve many genes of small effect, including genes that, like FOXP2, act pleiotropically (i.e., serve multiple functions). The case of the KE family is one of the rare cases of a monogenetic language disorder: the majority of cases of developmental language disorders do not involve a major single causal gene, but rather a constellation of genes, each exerting only a modest effect (raising the risk of a language disorder in an individual by a small percent) and interacting with environmental effects in a complex way. Notably, one of the implicated genes is a gene down-regulated by FOXP2, CNTNAP2. This gene has not only been linked to language impairment in children (Vernes et al., 2008), but also to normal variability in language development at the age of 2 years (Whitehouse et al., 2011), and to distinct patterns of neural activation on ERP language tasks (Kos et al., 2012) and fMRI (Whalley et al., 2011) in healthy adults.
A gradualist approach to the evolution of syntax we subscribe to is entirely consistent with the multiple-genes-of-small-effect view and does not expect there to be a grammar gene, i.e., a major causal gene of large effect, such that a single mutation in this gene would give rise to complex hierarchical syntax. Instead, a gradualist approach lends itself well to the modern understanding that pathways connecting genes to complex phenotypes, such as language, are complex and nonlinear, as articulated in Dediu (2015, p. 10944) and Dediu and Ladd (2007). While the outline of the puzzle is becoming increasingly clearer, more studies combining insights from theoretical linguistics, neuroscience, and developmental language disorders will be essential for advancing this line of research. The case of KE family is exceptional but consistent with this view: a single mutation in the gene caused severe disturbances in language processing and language use, but not complete loss of language), as well as disturbances in functions outside of language. This case also distills the essence of our evolutionary proposal, with all the components of an evolutionary approach to language coming together: a genetic change (a harmful or beneficial mutation), a linguistic change (impairment or development), and a distinct processing path associated with evolutionary development.
Conclusions
The knowledge generated in the fields of genetics, neurophysiology, developmental cognitive neuroscience, and linguistics is rapidly growing, and it is difficult to predict how our understanding of the evolution of language may shift in light of future discoveries. However, our present knowledge is consistent with the conclusion that genetic mutations in FOXP2 and other genes played an important role in the enhancement of the frontal-striatal brain network, densely connecting Broca's area to the basal ganglia, which in turn facilitated effortless processing of ever more and more complex syntax. The approach outlined here provides a good model for probing these questions further, and for shedding direct light on both language commonalities and language variation, a model that enables sustained cross-pollinating influences among the fields of linguistics, neuroscience, and genetics.
Statements
Ethics statement
This study was carried out in accordance with the recommendations of Wayne State University IRB Committee guidelines, with written informed consent from all subjects. All subjects gave written informed consent in accordance with the Declaration of Helsinki. The protocol was approved by Wayne State University IRB Committee.
Author contributions
LP provided theoretical framework and linguistics background. NR provided speech language pathology and genetics background. WA and RL ran the experiments and did statistical calculations. LT provided help with statistical and other analyses. NO provided the fMRI expertise and oversight.
Funding
This project was supported by funding to LP from Endowed Distinguished Faculty Fellowship awarded through Wayne State University Humanities Center; funding to NO through Wayne State University Institute of Gerontology; and additional funding through the Office of the Vice President of Research. We are also grateful to all the participants in this project, and to Nela Popadić for her help and support in recruiting participants. Our special thanks go to the reviewers, as well as to the editors, Enoch Aboh and Umberto Ansaldo, for putting together this project, and for providing both logistical and intellectual guidance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyg.2018.00278/full#supplementary-material
Footnotes
1.^There are additional projections that have been postulated for modern sentences in syntactic literature, such as Aspect Phrases (AspPs), Agreement Phrases (AgrPs), Topic Phrases (TopPs), etc. Our focus is on the syntactic layers in (1) because they meet with wide acceptance, and have the widest utility in the syntactic analysis of a variety of languages, as they relate to: transitivity, finiteness, and subordination.
2.^A related claim is found in the title of Dobzhansky's (1973) article “Nothing in biology makes sense except in the light of evolution.” On the other hand, as pointed out by a reviewer, there are several linguists who do not adopt this view, and who argue that language does not evolve but only changes (see e.g., Lightfoot's (in press); article titled “Nothing makes sense except in the light of change”). That language constantly changes is undeniable, and easily observable, but that does not mean that there are no selection forces acting upon language. As stated in Fitch (2008, p. 522), “language change does not entail a cessation of selection” (his article is subtitled “There is no ‘logical problem of language evolution”’). Steels (2011), who advocates studying cultural evolution (language change), also considers that both biological and cultural evolution are necessary to explain language, as these two processes propel each other through a self-reinforcing spiral process. In other words, we do not deny the relevance of language change—on the contrary, we believe that language change is the driving force behind evolution. But we do not subscribe to the view that language change, and language in general, are exempt from evolutionary processes, and we do not see the studies of language change as alternatives to the studies of language evolution, but rather we see them as two different facets of the same phenomenon.
3.^The CP layer is especially relevant for embedded clauses/subordination, or for sentences with wh-movement, neither of which are discussed in this article.
4.^Arguably, the intransitive small clause constitutes not only the foundation of syntactic structure elaboration in language evolution, but also in language acquisition. Thus, majority of verbal utterances in the speech of children learning a variety of languages at the onset of syntactic development (early verbal combinations) are analyzable as absolutive-like small clauses, with the additional layers emerging gradually (e.g., Rakhlin and Progovac, 2017).
5.^Many arguments used by saltationist approaches to the evolution of language, such as those in Lightfoot (1991) and Berwick and Chomsky (2011; 2016) have been responded to in e.g., Progovac (2009, 2015a, 2016b).
6.^Gil's association operator allows other possible interpretations for (8), including “the chicken that is eating,” which are not available in the sentences in (4) or (6). This may be due to the presence of specific functional categories in the latter, such as tense/agreement marking, which flag these as finite sentences. In general, the presence of functional categories tends to delimit the range of possible interpretations, leading to more precision/less vagueness.
7.^For completeness, we offer a representative example of the third transitivity type, a serial verb construction from Gungbe (Aboh, 2009), the structure that is not the focus of this paper:(i) Àsíbá bέ lέsì έù Asiba collect rice eat ‘Asiba ate a lot of rice.'We observe that in (i) each verb has only one argument, but this seemingly bi-clausal structure results in a single transitive clause interpretation. As Aboh (2009) makes it clear, the verb series in today's languages involves various grammatical complexities, and variation across languages, and this is also the case with ergative-absolutive (e.g., Aldridge, 2008) and nominative-accusative patterns.
8.^We recognize that using bilingual speakers is a limitation of the study and a replication of this experiment would benefit from being conducted with monolingual speakers of Serbo-Croatian.
9.^However, our experiment with Serbian small clauses vs. full TPs did not yield a significant result. While the issue deserves further attention, it is possible that the distinction between a small clause and a sentence is more subtle/more fluid in Serbian, given that it is only marked by the absence vs. presence of an auxiliary verb, whereas in English the presence of the tense auxiliary is necessarily accompanied by the presence of a determiner, as discussed below in the text. This indicates that English requires a DP (Determiner Phrase) projection with full TPs, while Serbian does not (see e.g., Bošković, 2008).
10.^We thank an anonymous reviewer for bringing this literature to our attention.
References
1
AbohE. O. (2009). Clause structure and verb series. Linguist. Inq.40, 1–33. 10.1162/ling.2009.40.1.1
2
AdgerD. (2003). Core Syntax: A Minimalist Approach. Oxford: Oxford University Press.
3
AldridgeE. (2008). Generative approaches to ergativity. Lang. Linguist. Compass2, 966–995. 10.1111/j.1749-818X.2008.00075.x
4
AnsaldoU.LaiJ.JiaF.SiokW. T.TanL. H.MatthewsS. (2015). Neural basis for processing hidden complexity indexed by small and finite clauses in Mandarin Chinese. J. Neurolinguist.33, 118–127. 10.1016/j.jneuroling.2014.08.005
5
ArdilaA.BernalB.RosselliM. (2016a). How localized are language brain areas? a review of brodmann areas involvement in oral language. Arch. Clin. Neuropsychol.31, 112–122. 10.1093/arclin/acv081
6
ArdilaA.BernalB.RosselliM. (2016b). Why Broca's area damage does not result in classical Broca's aphasia?Front. Hum. Neurosci.10:249. 10.3389/fnhum.2016.00249
7
BakerM. C. (2015). Case: Its Principles and Its Parameters.Cambridge: Cambridge University Press.
8
BerwickR.ChomskyN. (2011). The Biolinguistic Program. The current state of its development, in The Biolinguistic Enterprise: New Perspectives on the Evolution and Nature of the Human Language Faculty, eds Di SciulloA. M.BoeckxC. (Oxford: Oxford University Press), 19–41.
9
BerwickR.ChomskyN. (2016). Why Only Us? Language and Evolution. Cambridge, MA; London, UK: The MIT Press.
10
BisangW. (2009). On the evolution of complexity – sometimes less is more in East and mainland Southeast Asia, in Language Complexity as an Evolving Variable, eds SampsonG.GilD.TrudgillP. (Oxford: Oxford University Press), 34–49.
11
BoškovićŽ. (2008). What will you have, DP or NP? in Proceedings of NELS 37, eds WalkowM.ElfnerE. (Amherst: GLSA), 101–114.
12
CarnieA. (2013). Syntax: a Generative Introduction, 3rd Edn. Oxford, UK: Wiley-Blackwell.
13
ChomskyN. (1995). Minimalist Program. Cambridge, MA: MIT Press.
14
CroftW. (2003). Typology and Universals, 2nd Edn. Cambridge Textbooks in Linguistics. Cambridge: Cambridge University Press.
15
DawkinsR. (1996). The Blind Watchmaker: Why the Evidence of Evolution Reveals a Universe withtout Design. New York, NY: W. W. Norton and Company.
16
DediuD. (2015). An Introduction to Genetics for Language Scientists. Cambridge: Cambridge University Press.
17
DediuD.LaddD. R. (2007). Linguistic tone is related to the population frequency of the adaptive haplogroups of two brain size genes, ASPM and microcephalin. Proc. Natl. Acad. Sci. U.S.A.104, 10944–10949. 10.1073/pnas.0610848104
18
DobzhanskyT. (1973). Nothing in biology makes sense except in the light of evolution. Am. Biol. Teacher35, 125–129. 10.2307/4444260
19
DowtyD. (1991). Thematic proto-roles and argument selection. Language67, 547–619. 10.1353/lan.1991.0021
20
DraganskiB.KherifF.KlöppelS.CookP. A.AlexanderD. C.ParkerG. J.et al. (2008). Evidence for segregated and integrative connectivity patterns in the human basal ganglia. J. Neurosci.28, 7143–7152. 10.1523/JNEUROSCI.1486-08.2008
21
DryerM. (1992). The Greenbergian word order correlations. Language68, 81–138. 10.1353/lan.1992.0028
22
DryerM. (1997). On the six-way word order typology. Stud. Lang.21, 69–103. 10.1075/sl.21.1.04dry
23
EldredgeN.GouldS. J. (1972). Punctuated equilibria: an alternative to phyletic gradualism, in Models in Paleobiology, ed SchopfT. J. M. (San Francisco, CA: Freeman Cooper), 82–115.
24
EnardW.GehreS.HammerschmidtK.HölterS. M.BlassT.SomelM.et al. (2009). A humanized version of FOXP2 affects cortico-basal ganglia circuits in mice. Cell137, 961–971. 10.1016/j.cell.2009.03.041
25
FisherS. E. (2017). Evolution of language: Lessons from the genome. Psychon. Bull. Rev.24, 34–40. 10.3758/s13423-016-1112-8
26
FisherS. E.Vargha-KhademF.WatkinsK. E.MonacoA. P.PembreyM. E. (1998). Localisation of a gene implicated in a severe speech and language disorder. Nat. Genet.18, 168–170. 10.1038/ng0298-168
27
FitchW. T. (2008). Co-evolution of phylogeny and glossogeny: there is no logical problem of language evolution. Behav. Brain Sci.31, 521–522. 10.1017/S0140525X08005128
28
FitchW. T. (2010). The Evolution of Language. Cambridge: Cambridge University Press.
29
FordA. A.TriplettW.SudhyadhomA.GullettJ.McGregorK.FitzGeraldD. B.et al. (2013). Broca's area and its striatal and thalamic connections: a diffusion-MRI tractography study. Front. Neuroanat.7:8. 10.3389/fnana.2013.00008
30
FreyS.CampbellJ. S.PikeG. B.PetridesM. (2008). Dissociating the human language pathways with high angular resolution diffusion fiber tractography. J. Neurosci.28, 11435–11444. 10.1523/JNEUROSCI.2388-08.2008
31
FristonK. J.FletcherP.JosephsO.HolmesA.RuggM. D.TurnerR. (1998). Event-related fMRI: characterizing differential responses. Neuroimage7, 30–40. 10.1006/nimg.1997.0306
32
GibsonK. R. (1996). The ontogeny and evolution of the brain, cognition, and language, in Handbook of Human Symbolic Evolution, eds LockA.PetersC. R. (New York, NY: Oxford University Press), 409–433.
33
GilD. (2005). Isolating-monocategorial-associational language, in Handbook of Categorizaton in Cognitive Science, eds CohenH.LefebvreC. (Amsterdam: Elsevier), 347–379.
34
GivónT. (2002). Bio-linguistics: the Santa Barbara lectures. Amsterdam: John Benjamins.
35
GopnikM. (1990). Feature-blind grammar and dysphasia. Nature344:715. 10.1038/344715a0
36
GopnikM.CragoM. B. (1991). Familial aggregation of a developmental language disorder. Cognition39, 1–50. 10.1016/0010-0277(91)90058-C
37
GouldS. J.EldredgeN. (1977). Punctuated equilibria: the tempo and mode of evolution reconsidered. Paleobiology3, 115–151. 10.1017/S0094837300005224
38
GreenbergJ. H. (1966). Some universals of grammar with particular reference to the order of meaningful elements, in Universals of Grammar, 2nd Edn, ed GreenbergJ. H. (Cambridge, MA: MIT Press), 73–113.
39
HillertD. (2014). The Nature of Language: Evolution, Paradigms and Circuits. New York, NY: Springer.
40
JackendoffR. (1999). Possible stages in the evolution of the language capacity. Trends Cogn. Sci.3, 272–279. 10.1016/S1364-6613(99)01333-9
41
JackendoffR. (2002). Foundations of Language: Brain, Meaning, Grammar, Evolution.Oxford: Oxford University Press.
42
JacobF. (1977). Evolution and tinkering. Science196, 1161–1166. 10.1126/science.860134
43
KambaraT.BrownE. C.JeongJ.-W.OfenN.NakaiY.AsanoE. (2017). Spatio-temporal dynamics of short-term working memory maintenance and scanning of verbal information. Clin. Neurophysiol.128, 882–891. 10.1016/j.clinph.2017.03.005
44
KosM.van den BrinkD.SnijdersT. M.RijpkemaM.FrankeB.FernandezG.et al. (2012). CNTNAP2 and language processing in healthy individuals as measured with ERPs. PLoS ONE7:e46995. 10.1371/journal.pone.0046995
45
LaiC. S.FisherS. E.HurstJ. A.Vargha-KhademF.MonacoA. P. (2001). A forkhead-domain gene is mutated in a severe speech and language disorder. Nature413, 519–523. 10.1038/35097076
46
LeivadaE.KambanarosM.GrohmannK. (2017). The locus preservation hypothesis: shared linguistic profiles across developmental disorders and the resilient part of the human language faculty. Front. Psychol.8:1765. 10.3389/fpsyg.2017.01765
47
LiebermanP. (2000). Human Language and Our Reptilian Brain: The Subcortical Bases of Speech, Syntax, and Thought. Cambridge, MA: Harvard University Press.
48
LiebermanP. (2009). FOXP2 and human cognition. Cell137, 801–802. 10.1016/j.cell.2009.05.013
49
LiégeoisF.BaldewegT.ConnellyA.GadianD. G.MishkinM.Vargha-KhademF. (2003). Language fMRI abnormalities associated with FOXP2 gene mutation. Nat. Neurosci.6, 1230–1237. 10.1038/nn1138
50
LightfootD. (1991). Subjacency and sex. Lang. Commun.11, 67–69. 10.1016/0271-5309(91)90020-V
51
LightfootD. (in press). Nothing makes sense except in the light of change, in Language, Syntax, and the Natural Sciences, eds GallegoA. J.MartinR. (Cambridge: Cambridge University Press).
52
LorenzoG.VaresE. (2017). The externalization component as the locus of specific impairments. Biolinguistics11, 001–019.
53
MarantzA. (1991). Case and licensing, in Proceedings of the 8th Eastern States Conference on Linguistics, eds WestphalG.AoB.ChaeH.-R. (Ithaca, NY: CLC Publications).
54
MareljM. (2004). Middles and Argument Structure across Languages. Ph.D. thesis, Utrecht University, Published by LOT.
55
McFaddenT. (2004). The Position of Morphological Case in the Derivation: A Study on the Syntax-Morphology Interface. Ph.D. thesis, University of Pennsylvania, Philadelphia.
56
MithunM. (1991). Active/Agentive case marking and its motivations. Language67, 510–546. 10.1353/lan.1991.0015
57
MoroA.TettamantiM.PeraniD.DonatiC.CappaS. F.FazioF. (2001). Syntax and the brain: disentangling grammar by selective anomalies. Neuroimage13, 110–118. 10.1006/nimg.2000.0668
58
MufweneS. (2013). The origins and the evolution of language, in The Oxford Handbook of the History of Linguistics, ed AllanK. (Oxford: Oxford University Press), 13–52.
59
NewmanA. J.SupallaT.HauserP.NewportE. L.BavelierD. (2010). Dissociating neural subsystems for grammar by contrasting word order and inflection. Proc. Natl. Acad. Sci. U.S.A.107, 7539–7544. 10.1073/pnas.1003174107
60
NishidaM.KorzeniewskaA.CroneN. E.ToyodaG.NakaiY.OfenN.et al. (2017). Brain network dynamics of the human articulatory loop. Clin. Neurophysiol.128, 1473–1487. 10.1016/j.clinph.2017.05.002
61
OldfieldR. C. (1971). The assessment and analysis of handedness: the edinburgh inventory. Neuropsychologia9, 97–113. 10.1016/0028-3932(71)90067-4
62
Piattelli-PalmariniM.UriagerekaJ. (2011). A geneticist's dream, a linguist's nightmare: the case of FOXP2 gene, in The Biolinguistic Enterprise: New Perspectives on the Evolution and Nature of the Human Language Faculty, eds Di SciulloA. M.BoeckxC. (Oxford: Oxford University Press), 100–125.
63
PinkerS.BloomP. (1990). Natural language and natural selection. Behav. Brain Sci.13, 707–784. 10.1017/S0140525X00081061
64
ProgovacL. (2009). Sex and syntax: subjacency revisited. Biolinguistics3, 2–3, 305–336.
65
ProgovacL. (2013). Nonsentential vs. ellipsis approaches: review and extensions. Lang. Linguist. Compass 7/11, 597–617. 10.1111/lnc3.12044
66
ProgovacL. (2015a). Evolutionary Syntax. Oxford: Oxford University Press.
67
ProgovacL. (2015b). The absolutive basis of “middles” and the status of vP and UTAH, in Proceedings of FASL23 (Formal Approaches to Slavic Linguistics 23), Berkeley Meeting (Ann Arbor, MI: Michigan Slavic Publications), 242–261.
68
ProgovacL. (2016a). A Gradualist scenario for language evolution: precise linguistic reconstruction of early human (and Neandertal) grammars. Front. Psychol.7:1714. 10.3389/fpsyg.2016.01714
69
ProgovacL. (2016b). Review of Robert Berwick and Noam Chomsky's 2016 book Why Only Us: Language and Evolution. Cambridge, MA: Cambridge University Press. Language 92, 992–996.
70
RakhlinN.GrigorenkoE. (2015). (A)typical language development: genetic and environmental influences, in Handbook of Communication Disorders, eds BahrR. A.SillimanE. (London, UK; New York, NY: Routledge), 11–21.
71
RakhlinN.ProgovacL. (2017). Acquisition of clausal structure: the ‘Weakest Continuity’ view, in Proceedings of the 19th Annual International Conference of the Japanese Society for Language Sciences (JSLS), (Kyoto: Kyoto Women's University), 54–57.
72
SteelsLuc. (2011). Modeling the cultural evolution of language. Phys. Life Rev. 8, 339–356. 10.1016/j.plrev.2011.10.014
73
TchekhoffC. (1973). Some verbal patterns in Tongan. J. Polyn. Soc. 82.3, 281–292.
74
TeichmannM.DupouxE.KouiderS.BrugièresP.BoisséM. F.BaudicS.et al. (2005). The role of the striatum in rule application: the model of Huntington's disease at early stage. Brain128, 1155–1167. 10.1093/brain/awh472
75
TeichmannM.GauraV.DémonetJ. F.SupiotF.DelliauxM.VernyC.et al. (2008). Language processing within the striatum: evidence from a PET correlation study in Huntington's disease. Brain131, 1046–1056. 10.1093/brain/awn036
76
UllmanM. (2006). Is Broca's area part of a basal ganglia thalamocortical circuit?Cortex42, 480–485. 10.1016/S0010-9452(08)70382-4
77
Vargha-KhademF.GadianD. G.CoppA.MishkinM. (2005). FOXP2 and the neuroanatomy of speech and language. Nat. Rev. Neurosci.6, 131–138. 10.1038/nrn1605
78
Vargha-KhademF.WatkinsK.AlcockK.FletcherP.PassinghamR. (1995). Praxic and nonverbal cognitive deficits in a large family with a genetically transmitted speech and language disorder. Proc. Natl. Acad. Sci. U.S.A.92, 930–933. 10.1073/pnas.92.3.930
79
VernesS. C.NewburyD. F.AbrahamsB. S.WinchesterL.NicodJ.GroszerM.et al. (2008). A functional genetic link between distinct developmental language disorders. N. Engl. J. Med.359, 2337–2345. 10.1056/NEJMoa0802828
80
WhalleyH. C.O'ConnellG.SussmannJ. E.PeelA.StanfieldA. C.Hayiou-ThomasM. E.et al. (2011). Genetic variation in CNTNAP2 alters brain function during linguistic processing in healthy individuals. Am. J. Med. Genet. B-Neuropsychiatr. Genet.156B, 941–948. 10.1002/ajmg.b.31241
81
WhitehouseA. J.BishopD. V.AngQ. W.PennellC. E.FisherS. E. (2011). CNTNAP2 variants affect early language development in the general population. Genes Brain Behav.10, 451–456. 10.1111/j.1601-183X.2011.00684.x
82
YipM.MalingJ.JackendoffR. (1987). Case in tiers. Language63, 217–250. 10.2307/415655
Summary
Keywords
evolution of syntax, transitivity, syntactic processing, functional MRI, broca's–basal ganglia network
Citation
Progovac L, Rakhlin N, Angell W, Liddane R, Tang L and Ofen N (2018) Diversity of Grammars and Their Diverging Evolutionary and Processing Paths: Evidence From Functional MRI Study of Serbian. Front. Psychol. 9:278. doi: 10.3389/fpsyg.2018.00278
Received
01 November 2017
Accepted
19 February 2018
Published
06 March 2018
Volume
9 - 2018
Edited by
Enoch Oladé Aboh, University of Amsterdam, Netherlands
Reviewed by
David W. Lightfoot, Georgetown University, United States; Evelina Leivada, UiT The Arctic University of Norway, Norway
Updates
Copyright
© 2018 Progovac, Rakhlin, Angell, Liddane, Tang and Ofen.
This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ljiljana Progovac progovac@wayne.edu
This article was submitted to Language Sciences, a section of the journal Frontiers in Psychology
Disclaimer
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.