 Artur Czeszumski
Artur Czeszumski Benedikt V. Ehinger
Benedikt V. Ehinger Basil Wahn
Basil Wahn Peter König
Peter König- 1Institute of Cognitive Science, Universität Osnabrück, Osnabrück, Germany
- 2Department of Psychology, University of British Columbia, Vancouver, BC, Canada
- 3Institut für Neurophysiologie und Pathophysiologie, Universitätsklinikum Hamburg-Eppendorf, Hamburg, Germany
Humans achieve their goals in joint action tasks either by cooperation or competition. In the present study, we investigated the neural processes underpinning error and monetary rewards processing in such cooperative and competitive situations. We used electroencephalography (EEG) and analyzed event-related potentials (ERPs) triggered by feedback in both social situations. 26 dyads performed a joint four-alternative forced choice (4AFC) visual task either cooperatively or competitively. At the end of each trial, participants received performance feedback about their individual and joint errors and accompanying monetary rewards. Furthermore, the outcome, i.e., resulting positive, negative, or neutral rewards, was dependent on the pay-off matrix, defining the social situation either as cooperative or competitive. We used linear mixed effects models to analyze the feedback-related-negativity (FRN) and used the Threshold-free cluster enhancement (TFCE) method to explore activations of all electrodes and times. We found main effects of the outcome and social situation, but no interaction at mid-line frontal electrodes. The FRN was more negative for losses than wins in both social situations. However, the FRN amplitudes differed between social situations. Moreover, we compared monetary with neutral outcomes in both social situations. Our exploratory TFCE analysis revealed that processing of feedback differs between cooperative and competitive situations at right temporo-parietal electrodes where the cooperative situation elicited more positive amplitudes. Further, the differences induced by the social situations were stronger in participants with higher scores on a perspective taking test. In sum, our results replicate previous studies about the FRN and extend them by comparing neurophysiological responses to positive and negative outcomes in a task that simultaneously engages two participants in competitive and cooperative situations.
1. Introduction
In every day life, humans frequently commit errors. For example, they are prone to press incorrect buttons, trip over household objects or make typing mistakes. These errors often influence not only the person committing the mistake but also other people. Such erroneous actions may have a negative impact on others if people are cooperating in a task (e.g., moving furniture together). Conversely, they may have a positive impact on others if people are competing in a task (e.g., in a game of table tennis). These mistakes that involve others frequently require external feedback to find out about the impact of one's own and others' performed actions. Thus, it is likely that the human brain has mechanisms that distinguish between positive and negative outcomes of one's own and others' actions.
Earlier research on error processing in tasks performed individually shows that humans have a fast and efficient error detection mechanism (Coles et al., 2001; Yeung et al., 2004). In particular, studies using electroencephalography (EEG) identified event-related-potential (ERP) components instantly following one's own errors awareness, or feedback regarding the outcome of one's own actions (Falkenstein et al., 1991). These components are known as error-related-negativity (ERN) and feedback-related-negativity (FRN). The ERN is evoked 50–70 ms after an erroneous action is carried out (e.g., an incorrect button press) and it originates from the anterior cingulate cortex and the pre-supplementary motor area in the posterior medial frontal cortex (Holroyd et al., 2004; Ridderinkhof et al., 2004; de Bruijn et al., 2009). The FRN is elicited approximately 200–350 ms after performance feedback is received and is considered to have a similar origin as the ERN (Miltner et al., 1997). Holroyd and Coles (2002) proposed that the ERN/FRN component is elicited as soon as the outcome of an action can be detected by proprioceptive, motor or external feedback. They also proposed a direct relationship between a negative outcome detection and reward processing. In essence, whenever the result of an action is worse than expected, which results in a loss of reward, the ERN/FRN is elicited.
While these components have been widely studied in individuals, little research has investigated how humans process feedback about actions that involve others. A first step in this direction was made by van Schie et al. (2004). They found that the FRN component occurs after observing an error committed by others. Given the sensitivity of the FRN to mistakes of others, researchers suggest that it might reflect the processing of socially relevant stimuli. Further studies explored this idea by manipulating the social situation (i.e., either cooperative or competitive) while participants performed or observed actions and received feedback about monetary rewards (Itagaki and Katayama, 2008; Marco-Pallarés et al., 2010). Results showed that the FRN was elicited by losses of others in a cooperative situation. In a competitive situation, conversely, others' gains elicited the FRN. These results indicate that the FRN reflects the valence of an outcome, which in turn depends on the current social situation.
In contrast to studies of the FRN discussed above, the ERN, which is elicited for self-generated errors, appears to be not influenced by the social situation (de Bruijn and von Rhein, 2012). In another study self-generated errors elicited ERN in both cooperative and competitive situations, however, observed errors of others elicited the observed ERN (oERN) only in a cooperative situation (Koban et al., 2010). These studies focused on outcome processing in cooperative and competitive situations. However, the tasks used in these studies involved actions that are performed in turns and there was always either a division between a performer and observer participant (Koban et al., 2010; Marco-Pallarés et al., 2010; de Bruijn and von Rhein, 2012) or the partner was virtual (Itagaki and Katayama, 2008). Hence, it is not clear whether these findings would also generalize to designs in which co-actors perform a task that requires simultaneous responses to identical stimuli from both participants in contrast to turn-taking tasks such as for instance, joint Simon tasks (Sebanz et al., 2003, 2005; Dolk et al., 2014), in which co-actors respond to different stimuli at different time points.
To close this gap in the literature, a set of recent studies also investigated the FRN in situations in which humans perform tasks together. Humans in real life often perform actions together with others, instead of observing another human performing an action alone. Thus, studying the social aspect of outcome processing requires paradigms, in which co-actors perform tasks jointly (Hari and Kujala, 2009; Schilbach et al., 2013). In line with this idea Picton et al. (2012) tested dyads of participants in a cooperative joint choice reaction time task. In their study, participants were able to realize their own mistakes without feedback, which elicited the ERN, while mistakes of a partner had to be inferred from visual feedback, which elicited the FRN (Picton et al., 2012). In an even more naturalistic set-up, Loehr and colleagues tested piano duets (Loehr et al., 2013, 2015). Such a music paradigm allowed for a clear division between one's own, other's and joint errors. Results of both Picton et al's and Loehr et al's experiments confirmed that the FRN monitors both one's own and other's errors in joint situations. Interestingly, the FRN is stronger for one's own than joint mistakes, and stronger for joint mistakes than others' mistakes (Loehr et al., 2015). These studies focused on the monitoring of actions in cooperative joint set-ups. However, according to our knowledge there are no studies that involve two participants performing actions and receiving feedback about their individual and joint actions in both cooperative and competitive situations.
To fill this gap in the literature, in the present study we focused on two aspects: First, in our experiment both participants were actively performing a task. That is, in contrast to previous research there was no distinction between an active co-actor and a passively observing co-actor (Itagaki and Katayama, 2008; Marco-Pallarés et al., 2010). Instead, each of the participants performed their individually assigned task in parallel and observed their own and the co-actor's errors. Second, rewards (positive, negative, and neutral) associated with errors depended on whether the assigned task was performed in a cooperative or competitive situation. With this design, the main question we addressed was whether the FRN is influenced by different social situations when both co-actors actively perform a task. Additionally, by including neutral conditions (i.e., condition without any monetary rewards) in the design, we were able to investigate whether FRN amplitudes differed between errors that are associated with monetary outcomes (positive and negative) and errors that are not associated with any monetary rewards (neutral). Such comparisons were only rarely addressed in previous research (Holroyd et al., 2006). We also aimed to relate FRN amplitudes to personality traits measured with a questionnaire. Namely, we focused on the Perspective taking subscale of the Interpersonal reactivity index (IRI, Davis, 1983) that measures the tendency to spontaneously adopt the psychological point of view of others. We chose this subscale because it was already shown that FRN amplitudes correlate with the Perspective taking scores (Koban et al., 2012). Finally, we performed exploratory analysis to explore the time course of processing feedback about self-produced actions and co-actors' actions depending on the social situation.
2. Methods
2.1. Participants
Fifty-two students (37 females, mean age = 24.1, standard deviation = 4 years) randomly grouped into 26 dyads (15 female-male and 11 female-female dyads) participated in the experiment. Twenty-six participants were measured with EEG (16 females, mean age = 24.5, standard deviation = 3.3 years). Prior to the experiment we asked all participants whether they knew each other and paired only strangers in a dyad. The ethics committee of the University of Osnabrück approved the experiment. We informed participants about their rights and all participants signed a written consent form. The study was conducted in Osnabrück and all participants were students in an international study program. Therefore, all instructions and questionnaires were provided in English. Participants could chose either a monetary reward or course credits in exchange for their participation. All participants that were measured EEG opted for the monetary reward.
2.2. General Apparatus
We tested participants in dyads. They sat next to each other on the same side of a table in the same room. To avoid interference and communication during the experiment, we separated them with a cardboard screen (Figure 1B). We presented stimuli on two identical computer monitors (BenQ 24 inches, 1920 x 1080 pixels, refresh rate 120 Hz). We used two separate keyboards (Cherry RS 6000) to collect behavioral responses, one for each participant. The experiment was programmed using the Python library PsychoPy (Peirce, 2007) and the experimental procedure and data collection were implemented in Python 2.7.3 [code available (https://osf.io/c4wkx/)]. The experiment was run on an Intel Xeon CPU.
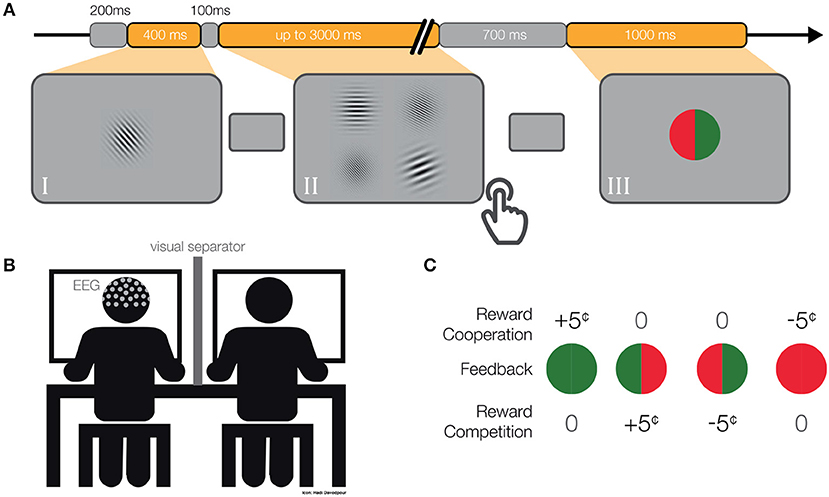
Figure 1. (A) Single-trial. We presented a gray mask for 200 ms, followed by (I) a target Gabor patch for 400 ms. Then again, a gray mask was displayed for 100 ms, followed by (II) four Gabor patches until both participants responded (maximum 3,000 ms). Subsequently, we displayed a gray mask for 700–800 ms, followed by (III) the feedback for 1,000 ms. (B) Schematic depiction of the experimental set-up. (C) Pay-off matrix. Participants received rewards differently in cooperative and competitive situations.
2.3. Experimental Design
Each member of a dyad performed a four-alternative forced-choice (4-AFC) visual task (Figure 1A) and later received feedback about their performance and associated monetary rewards (Figure 1C). In each dyad, one participant performed an orientation discrimination task and the other participant a spatial frequency discrimination task. The assignment of the participants to both tasks was randomized and counterbalanced. First, we presented a target object in the middle of the screen for 400 ms. The target object was a single Gabor Patch of size 9.95° x 9.95° visual angle, oriented at a randomly chosen angle (between 20°–80° and 100°–160°) and with a randomly chosen spatial frequency (between 10 and 20 cycles/stimulus size). Subsequently, we displayed a gray mask with a fixation cross in the middle (linewidth of 0.13° visual angle) for 100 ms followed by four Gabor patches arranged in a 2 x 2 grid, each patch separated from neighboring patches by 0.41° visual angle on each side. Each of the four Gabor patches was of the same size as the target object. One Gabor Patch always had the same orientation as the target object while the other three patches were manipulated according to a QUEST staircase procedure (Watson and Pelli, 1983). A different Gabor patch had the same spatial frequency as the target object and the other three patches again had different spatial frequencies according to a second QUEST staircase procedure (for more details about the QUEST procedure, see section 2.5). Therefore, both participants simultaneously had to respond to identical visual stimuli, however, their tasks were independent. This means, participants could not influence each other's performance while performing the task. Participants were informed that their partners had different tasks and they were familiar with the partner's instructions. The location of the correct answer for each of the participants was randomized between four possible locations. Participants responded with key presses (“Q,”“W,”“A,”“S,”or “7,”“8,”“4,”“5” on the num-pad, for the participants seated on the left or right respectively). The key corresponded spatially with the displayed Gabor patches. We displayed the Gabor patches until both of the participants gave their responses or 3,000 ms passed. In the case of no response, the answer was considered as incorrect. We instructed participants to give their answers as accurately and as quickly as possible. Subsequently, a gray mask with a fixation cross was displayed for 700–800 ms and then feedback appeared on the screen. We used a colored circle (radius: 3.94° visual angle) vertically divided in halves to inform participants about the performance of both participants. The color of the feedback was dependent on the participants' answers. The green color indicated correct answers and red incorrect answers. The left semicircle and right semicircle gave feedback to the left and right participants, respectively. Additionally, we presented individually a letter (0.8° visual angle, “W” for wins, “L” for losses and “T” for ties; for more details, see section 2.4 below) in the middle of a circle. Feedback was displayed for 1,000 ms and was followed by a gray mask for 200 ms before moving on to the next trial (Figure 1A).
2.4. Social Manipulation and Monetary Rewards
The feedback included information about individual and joint errors as well as the resulting positive, negative or neutral monetary rewards. Note, the schema of monetary rewards, as given in the pay-off matrix, defined the social situation as cooperative or competitive. The gain or loss of 5 cents was dependent on the particular social situation as follows (Figure 1C):
In the cooperative situation the trial was considered as a win, and consequently positively rewarded, only in the case in which both of the participants responded correctly (one green semi-circle for each of the two participants). In the case that both participants were wrong, it was considered a loss and as a negative reward five cents were subtracted from their budgets (one red semi-circle for each of the two participants). In the case that one participant was correct and the other was incorrect, no money was added to or subtracted from either budget (half green and half red circle).
In the competitive situation both participants answering correctly or incorrectly resulted in a tie (full green or red circle). Thus, no money was added to or subtracted from either budget. A reward was achieved when one participant was correct and the other was incorrect (half green and half red circle). In this case the reward was added to the correct participant's budget and subtracted from the incorrect participant's budget. At the end of each block the participants' respective budgets were calculated and displayed on the screen.
Social situations alternated between blocks (16 blocks in total, 8 cooperation and 8 competition). The order of blocks was counterbalanced across participants and randomly chosen for each dyad, with never more than three repetitions. To ensure that participants know and understand both social situations, we provided information regarding the block number, the social situation, and rewards associated with each feedback at the beginning of each block. In addition, “win” or “lose” was shown as text inside the feedback stimulus (Figure 1C). Each participant had an initial budget of 10 Euro that could increase or decrease by 5 cents based on their performances in each trial.
2.5. Experimental Procedure
One participant of each dyad was invited one hour earlier than the other and was prepared for the EEG recordings outside of the recording chamber. Thus, the participant assignment for EEG recordings was done prior to the experiment. After around 45 min, when preparation was finished and the second participants arrived, both participants were seated side-by-side in a room at a 60 cm distance to their screen. For technical reasons, the participant measured with the EEG sat on the left side. The experimental session lasted approximately 90 min and was structured as follows: After detailed written and oral instructions, a QUEST staircase procedure (Watson and Pelli, 1983) was performed for each participant separately (one after another) for the assigned task with the goal to home in on 50% performance, i.e., well above the chance level of 25%. To achieve this, we used the PsychoPy QuestHandler function with the threshold set to 0.63 and a gamma 0.01. Both participants performed 100 training trials. For the participant performing the orientation discrimination task we varied the degree of orientation between 1° and 45° with a starting value of 15° and a standard deviation 10°. For the other participant, who performed the spatial frequency discrimination task, we varied the spatial frequency between 1 and 25 cycles/stimulus size with starting value of 3 cycles/stimulus with a standard deviation of 3 cycles/stimulus. Subsequently, participants proceeded to the actual experiment, which consisted of a total of 640 trials grouped in 16 blocks of 40 trials each. After 20 trials in each block, participants were asked to answer in which social situation they were currently in. Namely, they were asked to indicate whether the current block was a cooperative or competitive situation, in order to check whether the participants remembered the social situation manipulation correctly. Blocks were separated by short rests and the overall experiment was divided into three parts with short breaks. In these breaks experimenters made sure that participants were not exchanging any information about the experiment. When the tasks were completed, participants filled out the Interpersonal reactivity index (IRI, 28 questions) questionnaire (Davis, 1983).
2.6. Methods of EEG Data Acquisition and Preprocessing
Electrophysiological data were recorded using a 64-Ag/ AgCl electrode system (ANT Neuro, Enschede, Netherlands), using a REFA-2 amplifier (TMSi, Enschede, Netherlands) with electrodes placed on a Waveguard cap according to the 5% electrode system (Oostenveld and Praamstra, 2001). The data was recorded using average reference electrode at a sampling rate of 1,024 Hz. Impedances of all electrodes were manually checked to be below 10 kΩ before each experiment. We used R and MATLAB to preprocess and analyze the data. All analysis scripts and data are available online (https://osf.io/c4wkx/). We used the eegvis toolbox (Ehinger, 2018) to visualize the exploratory analyses. Data were preprocessed using the EEGLAB toolbox (Delorme and Makeig, 2004) in the following order: First, the data were downsampled to 512 Hz and subsequently filtered using a 0.1 Hz high-pass filter and a 120 Hz low pass filter (6 dB cutoff at 0.5Hz, 1 Hz transition bandwidth, FIRFILT, EEGLAB plugin). Channels exhibiting either excessive noise or strong drifts were manually detected and removed (2.1 ± 2.5, mean and standard deviation, respectively). After this, the continuous data were manually cleaned, rejecting data sequences including jumps, muscle artifacts, and other sources of noise. To remove eye and muscle movement-related artifacts, an independent component analysis based on the AMICA algorithm (Palmer et al., 2008) was computed on the cleaned data. The independent components (ICs) corresponding to eye, heart, or muscle activity were manually selected based on their timecourse, spectra and topography, and removed before transforming the data back into the original sensor space (number of removed ICs 8.3 ± 5.2, mean and standard deviation, respectively). The initially removed channels were interpolated based on the activity of their neighboring channels (spherical interpolation). Subsequently, the continuous data were divided into epochs for each trial by including data from 200 ms pre-stimulus to 1,000 ms post stimulus, using the time window between –200 ms and stimulus onset for baseline correction. For the exploratory analysis we used 62 electrodes (Fp1, FPz, Fp2, F7, F3, Fz, F4, F8, FC5, FC1, FC2, FC6, T7, C3, Cz, C4, T8, CP5, CP1, CP2, CP6, P7, P3, Pz, P4, P8, POz, O1, Oz, O2, AF7, AF3, AF4, AF8, F5, F1, F2, F6, FC3, FCz, FC4, C5, C1, C2, C6, CP3, CPz, CP4, P5, P1, P2, P6, PO5, PO3, PO4, PO6, FT7, FT8, TP7, TP8, O7, PO8).
3. Results
3.1. Behavioral Analysis
3.1.1. Social Situation Awareness
To assure that participants payed attention to the different social situations in the experiment we asked them in the middle of each block whether the current block was a cooperative or competitive situation. Answering this question participants achieved a high accuracy (mean correct answers = 97%, standard deviation = 7%), suggesting that participants consistently understood and memorized the instructions about differences between social situations.
3.1.2. Accuracy
Prior to running the actual experiment, we used a QUEST staircase procedure to adjust a difficulty in each task for participants such that participants were expected to attain a 50% accuracy. Confirming this expectation, the mean accuracy in the task was 53% (standard deviation = 9%) and the mean difference between paired participants was 8% (standard deviation = 6%). It was important that both paired participants performed with comparable accuracy to avoid that the analyzed ERPs are influenced by differences at the behavioral level. Further, it results in an even distribution of performance data in correct-correct, correct-false, false-correct, and false-false.
3.1.3. Response Time
We analyzed response times to test whether our experimental manipulations influenced behavioral responses. Prior to analysis, we excluded all trials with response times faster than 50 ms (2 trials) because such fast responses are likely due to premature responses. Then, we used a linear mixed model (LMM) to analyze response times. The LMM was calculated with the lme4 package (Bates et al., 2015) and p-values were based on Walds-T test using the lmerTest package. Degrees of freedoms were calculated using the Satterthwaite approximation. We modeled responses times by task, social situation, and correctness as fixed effects and interactions between them. As random effects, we used random intercepts for grouping variables participants and dyads. In addition, we used random slopes for all fixed effects, including interactions, in the participant grouping variable. For all predictors, we used an effect coding scheme with binary factors coded as –0.5 and 0.5. Thus, the resulting estimates can be directly interpreted as the main effects. The advantage of this coding scheme is that the fixed effect intercept is estimated as the grand average across all conditions and not the average of the baseline condition. We found a main effect of correctness [t(50.18) = −8.1, p < 0.0001]. Correct answers were on average 80 ms faster than incorrect answers. The main effects for the two other predictors (tasks and social situations) and all possible interactions were not significant (p >0.17). These results suggest that different tasks (orientation and spatial frequency) and social situations (cooperative and competitive) are of comparable level of difficulty and engage two participants to similar degrees.
3.2. Electrophysiological Data
To analyze EEG data in form of ERPs, we applied a preselected single-trial based LMM analysis (Frömer et al., 2018). We defined the FRN as the mean amplitude over six electrodes (Fz, F1, F2, FCz, FC1, FC2) between 200 and 300 ms after the feedback of each trial. Our choice of electrodes and time window was based on previous research and were pre-specified before any analysis (Ullsperger et al., 2014). We modeled the FRN using outcomes (win and lose) and social situations (cooperative and competitive) as fixed effects and an interaction between them. As random effects, we modeled random intercepts for participants and random slopes for both predictors (outcomes and social situations) and interaction between them. For the same reason as above, predictors were effect coded, i.e., binary factors are coded as –0.5 and 0.5. The result of this analysis are presented in Table 1 and ERPs in Figure 2. We found main effects for the outcome [t(26.02) = −5.85, p < 0.001] and the social situation [t(26.01) = 4.4, p < 0.001]. The FRN amplitudes were on average 1.03 (standard deviation = 0.23) μV higher in lose than win trials and 1.54 (standard deviation = 0.26) μV higher in competitive than cooperative trials. The interaction between these factors was not significant [t(27.15) = −0.93, p = 0.36]. These results suggest that the FRN differs between positive and negative outcomes and between cooperative and competitive social situations and that these two effects are independent of each other.
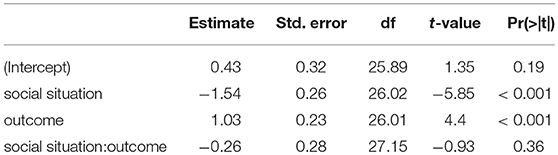
Table 1. LMM Effects of outcome and social situation on the FRN [mean amplitude (200–300 ms)] (effect coding: –0.5,0.5, maximal LMM).
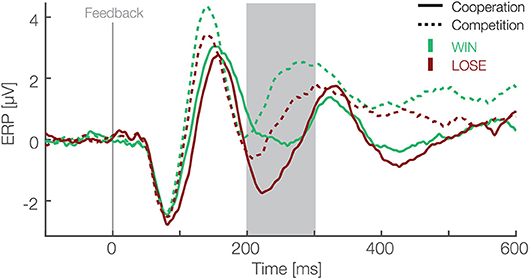
Figure 2. Feedback locked ERP waveforms at pooled electrode sites (F1, Fz, F2, FC1, FCz, FC2). Data are averaged referenced. Green and red colors represent the outcome, i.e., win and lose trials respectively. Solid and dashed lines represent cooperative and competitive situations. The gray box shows the preselected time window used for the confirmatory statistical analysis (200–300 ms).
Additionally, we used individual estimates of the difference between the FRN in the two social situations to correlate them with the Perspective Taking Score. We calculated the Spearman's Rho to quantify the association of the Perspective taking score and individual participant's mixed model best linear unbiased prediction of the factor social situation from the mean amplitude analysis. We chose Spearman's correlation because our questionnaire data was rank data. We found a significant negative correlation (r = –0.54, p = 0.005, Figure 3). This result suggests that on average the effect of the social situation is stronger on the characteristic ERPs in participants with personality traits related to high perspective taking abilities.
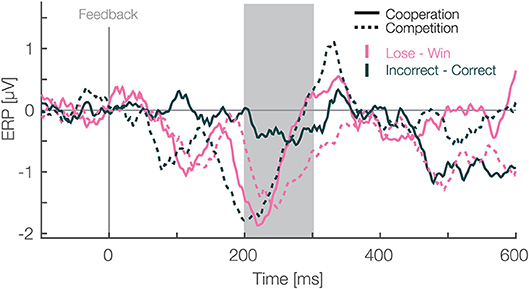
Figure 3. Feedback locked difference waveforms at pooled electrode sites (F1, Fz, F2, FC1, FCz, FC2). Data are average referenced. Pink and green colors represent the monetary outcome, i.e., lose-win (monetary) and incorrect-correct (neutral) trials respectively. Solid and dashed lines represent cooperative and competitive situations. The gray box shows the preselected time window used for the confirmatory statistical analysis (200–300 ms). All ERP waveforms that were used to make difference waves are presented in Supplementary Materials.
Furthermore, after visual inspection of the grand average ERPs (Figure 2), we decided to also apply a peak to peak amplitude analysis because the FRN peaked earlier than expected (Ferdinand et al., 2012). For the peak to peak analysis we used the same electrodes as for the mean amplitude analysis (Fz, F1, F2, FCz, FC1, FC2). We used the grand average to identify the maximum positive peak between 140 and 200 ms and the maximum negative peak between 200 and 270 ms after feedback presentation. We subtracted the average maximum negative peak amplitude from the average maximum positive peak over these time windows. This is equivalent to applying directly a peak-to-peak analysis on data low-pass filtered by a boxcar kernel. Compared to the plain peak-to-peak analysis it is, however, less susceptible to high frequency noise and therefore more robust. Then, we applied exactly the same LMM analysis as with the mean amplitude (details above). The result of this analysis are presented in the Table 2. We found main effects for the outcome [t(26) = 3.55, p = 0.001] and the social situation [t(26.09) = −3.04, p = 0.005]. The peak amplitudes were on average 0.71 (standard deviation = 0.23) μV higher in win than lose trials and 1.3 (standard deviation = 0.36) μV higher in cooperative than competitive trials. The interaction between these factors was not significant [t(25.98) = −0.98, p = 0.34]. These results are in line with results of mean amplitude analysis, further corroborating that FRN amplitudes differ between positive and negative outcomes and between cooperative and competitive situations.
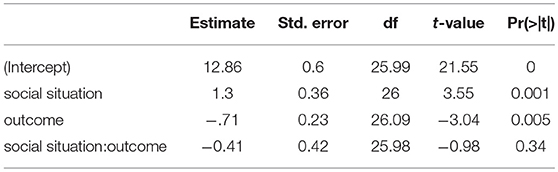
Table 2. Effects of outcome and social situation on the FRN [peak to peak amplitude (140–270 ms)] (effect coding: –0.5,0.5, maximal LMM).
For the exploratory analysis, we used the Threshold-Free-Cluster-Enhancement method (TFCE) and permutation analysis (Smith and Nichols, 2009; Mensen and Khatami, 2013; Ehinger et al., 2015). This method allows for comparisons between experimental conditions over all electrodes and time points of ERPs while at the same time controlling for the multiple comparison. We analyzed the EEG data with a two-way repeated measures ANOVA with outcome (win vs. lose) and the social situation (cooperative vs. competitive) as within participants factors and taking into account 62 electrodes, and all time points between 0 and 600 ms. We enhanced the signal with the TFCE method and used permutation tests to account for multiple comparisons. We used 5,000 permutations and for each permutation we randomized the assignment to different experimental conditions of each data point within each participant. For each of these TFCE permutations, a repeated measures ANOVA was calculated. The maximum F-value across chosen samples in time and space were used to construct a max F-value distribution, against which the actual F-values were compared. We considered F-values above the 95th percentile to be significant. The results of this analysis are presented in Figure 5. We found two separate clusters of significant activity for the main effect of outcome. One cluster spans from 88 to 152 ms (median p value: p = 0.01, min p value: p = 0.001) with a peak at C1 electrode 121 ms after the feedback and was more negative for lose than win outcomes. The other cluster ranges between 172 and 340 ms with a peak at Fz electrode 240 ms following the feedback (median p value: p = 0.01, min p value = 0.0006). This cluster resembles spatially and temporally the FRN and it was more negative for lose than win outcomes. Please note that in contrast to the conventional analysis above that makes assumptions on the timing of the relevant signals, the TFCE approach gives the intervals with significant differences as a result. Thus, the present analysis validates and makes the assumptions of the analysis above more precise. Moreover, we found that there is a main effect of the social situation. This cluster stretched from 68 till 600 ms (median p value: p = 0.0004, min p value: p = 0.0002) and encompassed all electrodes at different time points, suggesting a robust difference in processing of feedback between cooperative and competitive situations. The peak significant value was at FC5 electrode 143 ms after the feedback. Overall, these results support the observations above of large differences between processing of feedback between cooperative and competitive situations and suggest that the difference in processing positive and negative feedback starts earlier than classically considered time window for the FRN.
Next, to fully explore our design we analyzed differences in the FRN amplitudes between monetary vs. neutral outcomes crossed with social situations. For this, we utilized a difference wave approach (Li et al., 2018). In each of the social situations, we subtracted ERPs of negative from positive monetary outcomes. In addition we subtracted incorrect responses from correct ones in the neutral monetary outcomes. Then, we quantified the FRN, likewise as in the LMM analysis above, as the mean amplitude between 200 and 300 ms after the feedback presentation for each condition. We used a two-way repeated measures ANOVA with social situation (cooperative vs. competitive) and type of outcome (monetary vs. neutral) as within participant factors. We applied a different statistical method than in above mean amplitude analysis to analyze difference waves as it is unclear how one would pair trials for subtraction on a single trial level. Thus, we used grand averages for each condition to calculate difference waves. Moreover, we used a difference wave approach to simplify the necessary statistical model and answer a different question. Namely, whether types of outcomes (monetary vs. neutral), without considering whether it's positive or negative, are different. In the time window from 200 to 300 ms we found a main effect of social situation [F(1,25) = 6.17, p = 0.02, η2 = 0.022, Figure 4)], a main effect of type of outcome [F(1,25) = 4.55, p = 0.04, η2 = 0.026], and no interaction between these factors [F(1,25) = 0.34, p = 0.56, η2 = 0.003]. The amplitudes were more negative for monetary than neutral outcomes and more negative in competitive than cooperative situations. These results suggest that the effect of the social situation on the FRN reported above extends to neutral outcomes. Furthermore, the significant difference between monetary and neutral outcomes suggest that is sensitive to both monetary rewards and task performance. Furthermore, after we observed significant cluster resembling the FRN in our exploratory analysis (172 to 340 ms after the feedback presentation, see above) we analyzed this later time window as well. In particular, we calculated the mean amplitude between 300 and 340 ms for each difference wave and analyzed it with a two-way repeated measures ANOVA with social situation (cooperative vs. competitive) and type of outcome (monetary vs. neutral) as within participant factors. The main effect of social situation [F(1,25) = 0.05, p = 0.83, η2 = 0.0002, Figure 4] was not significant. However, we found a main effect of type of outcome [F(1,25) = 8.32, p = 0.007, η2 = 0.02] and an interaction effect between the factors [F(1,25) = 4.91, p = 0.036, η2 = 0.06]. Thus, in contrast to the time window predefined based on earlier literature, we do not observe a main effect of the social situation in the late window, but an interaction arises between the factors of social situation and outcome. Thus, the complete time window reaching up to 340 ms contains dynamics and is not a completely homogeneous block. Specifically, these results further corroborate that feedback processing is sensitive to both monetary rewards and task performance. Moreover, the social situation modulates amplitudes for both of them.
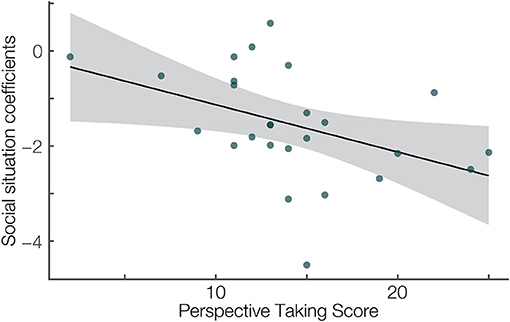
Figure 4. Correlation between the Perspective taking subscale of the Interpersonal Reactivity Index (Davis, 1983) (x-axis) and the single-subject linear mix model estimates of the social situation effect (y-axis). Spearman's r = –0.54, p = 0.005. Linear fit with 95% confidence interval.
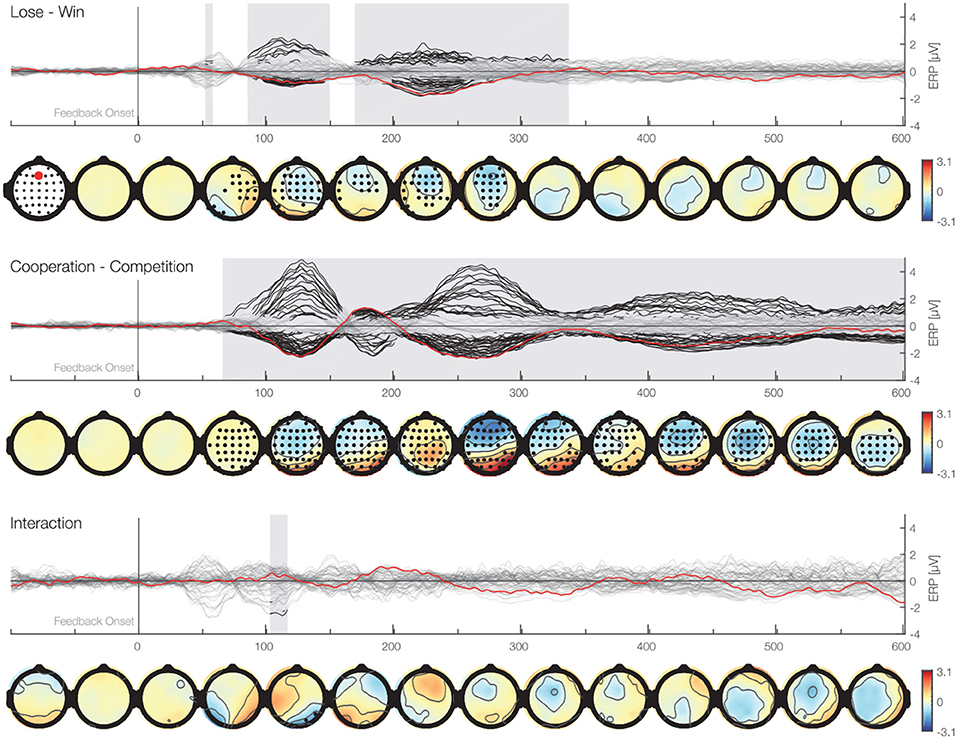
Figure 5. Time-series plots of the EEG amplitudes of the main factors and interaction for each electrode aligned to the feedback stimulus. First row (butterfly plot) shows time against activity of all electrodes. In addition, the Fz electrode is marked in red. Black marked clusters are significant under a TFCE permutation p-value of 0.05. TFCE corrects for multiple comparisons over time and electrodes. Second row shows topographical plots representing the mean amplitudes averaged over 50 ms bins. Black marked electrodes represent significant channels.
Lastly, we address a potential visual confound in our design. As we used four different visual stimuli to inform our participants about their performance and associated rewards, results potentially reflect differences of the visual feedback. To address this potential perceptual confound, we invited five participants again, who previously completed the experiment, for a control experiment. In this version of the experiment, the Gabor patches were not displayed and random feedback was provided. Thus, this experiment controls for the pure visual effect of the feedback. To assess this potential confound, we calculated grand average ERPs for experimental and control data. We visually inspected the ERPs and found no difference between the different visual feedback displays, including the early visual components. Then, we subtracted these control data from the experimental data. Again, visual inspection suggests that differences in the visual appearance of the feedback information did not influence the FRN (Figure 6). This is in line with previous research that shows only early components e.g., C1, P1, N1, in the first 150 ms are modulated by such low-level visual stimuli properties (Wijers et al., 1989; Hillyard and Anllo-Vento, 1998). Hence, we are reassured that our results represent differences between outcomes and social situations and not due to differences in the visual stimuli.
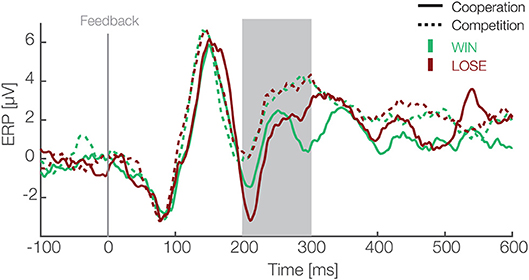
Figure 6. Feedback locked averaged difference waves between experimental and control data for 5 participants pooled at electrode sites (F1, Fz, F2, FC1, FCz, FC2) are shown. In experimental data green and red colors represent the outcome, i.e., win and lose trials respectively, while solid and dashed lines represent cooperative and competitive situations. Difference between social situations and outcomes is visible after subtraction of electrophysiological response to identical visual stimuli without any content. This suggests that our results represent differences in experimental manipulations but not visual properties of stimuli.
4. Discussion
The goal of the present study was to compare reward processing between different social situations as well as to test whether earlier results (Picton et al., 2012) generalize to a setting which actively involves two participants. For this purpose, we designed a joint 4-AFC visual task, in which two co-actors both concurrently perform a task and receive rewards depending on the social situation. We were able to replicate the difference in FRN amplitudes between positive and negative outcomes in the cooperative situation (Picton et al., 2012). Moreover, we extended these earlier results by observing a significant difference between win and lose outcomes in the competitive situation. We also found that the FRN significantly differs between social situations, suggesting that reward processing is modulated by the social situation. However, we did not observe an interaction between these two factors. Further, the difference induced by the social situations were stronger in participants with higher perspective taking scores, which were obtained using a perspective taking questionnaire. Finally, we compared feedbacks with and without monetary outcomes (win/lose vs. neutral) in both social situations. We found that our reported effect, that the social situation affects the FRN, also extends to the processing of neutral outcomes. Moreover, we found a significant difference between feedbacks with and without monetary outcomes, suggesting that the FRN is sensitive to both monetary rewards and task performance.
Earlier behavioral findings support the idea that humans co-represent co-actors actions even if they are irrelevant to one's own goals (Atmaca et al., 2011), for a recent general review,(see, Vesper et al., 2017). Such representations may also influence how humans process feedback about actions and associated monetary rewards while performing joint actions with another person. Therefore, our experiment involved two participants performing their tasks simultaneously and hence differs from previous studies that utilized a virtual partner to investigate differences between social situations (Itagaki and Katayama, 2008). Moreover, the design allows for concurrent actions from both participants–an aspect that it is not present in designs that employ turn-taking tasks which create a division between a performer and observer (Koban et al., 2010; Marco-Pallarés et al., 2010; de Bruijn and von Rhein, 2012). Thus, with the results of the present study, we extended earlier findings by demonstrating that they also generalize to a setting involving co-actors that both actively and simultaneously perform a task.
Our result that the outcome (positive vs. negative reward) affects the FRN in both social situations is in line with a great body of earlier research (Ullsperger et al., 2014). We quantified the FRN in two different ways (mean and peak to peak amplitude) and applied additional exploratory analyses. Results of all three analyses provide strong evidence that negative outcomes elicit more negative amplitudes at mid-line electrodes around 200 to 300 ms after the feedback presentation. Such an outcome of our study suggests that the FRN component is robust and it generalizes from individual to joint set-ups and different social situations. In contrast, our results are not compatible with the theory that the FRN represents differences in expectancies and probabilities (Alexander and Brown, 2010, 2011). In our task the probabilities for each outcome were nearly equal, therefore, there are no differences in probabilities or expectancies. Future studies could investigate whether reward processing is also affected by the outcome in tasks, in which both co-actors actively perform a task collaboratively as, for instance, in joint perceptual tasks (e.g., Brennan et al., 2008; Brennan and Enns, 2015; Wahn et al., 2016b, 2017a,b, 2018c; for a recent review, see Wahn et al., 2018a) or in joint motor tasks (e.g., Knoblich and Jordan, 2003; Wahn et al., 2016a, for a recent review, see Wahn et al., 2018b).
The main question, namely, whether reward processing differs between social situations was addressed in three ways. First, we analyzed the FRN as mean as well as peak to peak amplitude and found a main effect of social situation. Second, we also found a main effect of social situation when analyzing the difference waves. Third, using an exploratory analysis, we again found a main effect of social situation. Taken together, these results suggest that the FRN amplitudes are affected by the social situation, although the lack of interaction in a pre-specified time window (200 to 300 ms) implies that positive and negative outcomes are equally affected. Additionally, analysis of a later time window (300 to 340 ms) in difference waves revealed a main effect of different types of outcome and an interaction effect. This raises the question which aspect of the change in social situation affects the FRN. Potentially, the social situations might differ with respect to arousal state and the amount of attentional resources utilized. Previous research points in the direction of such an interpretation (Cui et al., 2015). However, we did not observe differences in the level of performance as a function of the social situation. This makes an influence on the FRN by variations of arousal or attentional resources unlikely. Therefore, our study provides evidence that reward processing is affected by social situations, however, further research is needed to unravel details of involved processes. Given, that we find that the social situation (cooperative or competitive) modulates processing of feedback about our actions, an interesting research direction would be to test how different social situations may also affect how co-actors monitor actions joint actions (Keller, 2008; Vesper et al., 2010), representations of co-actors in a dyad (Sebanz et al., 2005), and the prediction of co-actors actions of co-actors (Keller et al., 2007). Moreover, our results are in line with EEG hyperscanning studies suggesting that different cognitive processes are involved in cooperative and competitive situations (Astolfi et al., 2010; Sinha et al., 2016).
A previous study suggested that that the FRN is only sensitive to the outcome, but not task performance as such (Itagaki and Katayama, 2008). As studying the FRN in response to neutral outcomes is mostly neglected in literature (but see Holroyd et al., 2006), this is difficult to disentangle. Due to our design that included neutral outcomes, we were in a better position. Specifically, the comparison of FRN amplitudes between feedbacks with and without monetary outcomes in combination with correct or incorrect individual performance, results in a significant difference between feedbacks with and without monetary outcomes. This result suggests that different neural processes are involved in processing outcomes and task performance. Given that we find that the FRN is present for neutral outcomes, this result suggests that the FRN is sensitive to outcome as well as task performance.However, we have to be cautious with interpretation of these results because both performance and monetary feedback was delivered at once and it is not clear how to disentangle them in our design. In future research, one could manipulate chance of winning (25, 50, and 75%) for individual participants to extend current results.
In this study, we used state of the art EEG analysis methods, namely Linear Mixed Models for hierarchical analysis of single trial activity (Frömer et al., 2018) and TFCE to control for multiple comparisons (Smith and Nichols, 2009; Mensen and Khatami, 2013). In the following, we first provide a discussion of the benefits using these analysis techniques and then further discuss the obtained results of our exploratory analysis. We quantified the FRN on a single trial basis and used the LMM to model the FRN. This approach helps to account for a multitude of problems. For instance it handles unequal number of observations per cell, allows for between participant variability in effect sizes and combines single participant variability and group level variability (Pinheiro and Bates, 2000; Baayen et al., 2008; Barr et al., 2013; Matuschek et al., 2017). In our experiment, we tried to reduce the first problem of unequal cell size by using the QUEST procedure to obtain almost equal number of trials. Nevertheless, EEG data has to be cleaned and depending on the noise level the number of rejected trials varies between participants. However, the issue of high variability between participants in cognitive neuroscience field is prevalent and has to be accounted for (Seghier and Price, 2018). The LMM approach is suitable to address this problem. Our additional motivation to use this method was related to its capability of estimating effect sizes for individual participants. We used those to correlate them with information about personality traits of participants to test a possible association between neurophysiological and questionnaire data. We also made use of the TFCE permutation analysis to perform the exploratory analysis (Mensen and Khatami, 2013) without specifying electrode sites or time window. This approach circumvents the need to preselect time points and electrodes (Bishop, 2007), which is an additional benefit as making these decisions may not always be straightforward, especially in the absence of clear guidelines.
Using this exploratory analysis, we found the same pattern of results as above in our confirmatory analysis. Namely, we found a main effect of the outcome and social situation in both the LMM and the permutation analysis, further corroborating earlier results that the FRN is sensitive to positive and negative outcomes and the social situation (Ullsperger et al., 2014). In addition, our exploratory analysis showed that these differences for the FRN preceded the time window typically defined for the FRN, suggesting that the human brain differentiates the valence of the outcome and the social situation earlier than previously suggested (Koban et al., 2010; Marco-Pallarés et al., 2010; Rigoni et al., 2010; de Bruijn and von Rhein, 2012; Picton et al., 2012; Loehr et al., 2013, 2015). Our results (Figure 5, second row), suggest that there are stronger positive activation in cooperative than competitive situation in two stages of processing of the feedback. Namely, around 160 and 280 ms after the feedback presentation. The social situation main effect might arise from a source close to CP6 and P6 electrode. Because Superior temporal sulcus (STS) and Temporoparietal junction (TPJ) are close to these electrodes and earlier fMRI research suggests these areas are involved in differentiating the self from others, this might be the origin (Saxe and Kanwisher, 2003). Thus, it might be interpreted that while people receive feedback and process them simultaneously in a cooperative situation they merge their own and their co-actor positive outcomes and process them as simultaneously while the competitive situation requires distinct processing of rewards. However, this interpretation have to be taken cautiously due to the inverse problem.
Moreover, we investigated the relation between the Perspective taking score and mixed model best linear unbiased prediction of the factor social situation. We found that the higher the Perspective taking score, the stronger is the difference in FRN amplitudes between social situations. This result suggests that personality traits related to perceiving and understanding others might be related to the strength of the neurophysiological response to rewards. Thus, brain mechanisms involved in reward processing in people showing more consideration for others, might be more sensitive for different social situations. However, this result and interpretation should be treated with caution, as using mixed model best linear unbiased prediction in combination with a correlation analysis is a new approach and still has to be fully validated (Houslay and Wilson, 2017).
Taken together, we investigated neural underpinnings of feedback processing in cooperative and competitive situations. We find that the FRN component is sensitive not only to positive and negative outcomes but also to the social situation in a design, in which both co-actors in dyad actively perform a task.
Author Contributions
AC, BW, and PK: Study design; AC: data collection; AC and BE: data analysis; AC, BE, BW, and PK: draft and revisions of manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully acknowledge the support by the European Commission Horizon H2020-FETPROACT-2014 641321-socSMCs, DFG-funded Research Training Group Situated Cognition (GRK 2185/1) and the Deutsche Forschungsgemeinschaft (DFG) Open Access Publishing Fund of Osnabrück University. Moreover, we would like to thank Anna Lisa Gert for her help with preprocessing EEG data as well as following students, who collected the data: Chiara Carrera, Marketa Becevova, Maria Sokotushchenko, Greta Häberle and Susanne Schuberth.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyg.2019.00361/full#supplementary-material
References
Alexander, W. H., and Brown, J. W. (2010). Computational models of performance monitoring and cognitive control. Top. Cogn. Sci. 2, 658–677. doi: 10.1111/j.1756-8765.2010.01085.x
Alexander, W. H., and Brown, J. W. (2011). Medial prefrontal cortex as an action-outcome predictor. Nat. Neurosci. 14:1338. doi: 10.1038/nn.2921
Astolfi, L., Cincotti, F., Mattia, D., Fallani, F. D. V., Salinari, S., Vecchiato, G., et al. (2010). “Imaging the social brain: multi-subjects eeg recordings during the “chicken's game,” in Engineering in Medicine 505 and Biology Society (EMBC), 2010 Annual International Conference of the IEEE (Buenos Aires: IEEE), 1734–1737.
Atmaca, S., Sebanz, N., and Knoblich, G. (2011). The joint flanker effect: sharing tasks with real and imagined co-actors. Exp. Brain Res. 211, 371–385. doi: 10.1007/s00221-011-2709-9
Baayen, R. H., Davidson, D. J., and Bates, D. M. (2008). Mixed-effects modeling with crossed random effects for subjects and items. J. Mem. Lang. 59, 390–412. doi: 10.1016/j.jml.2007.12.005
Barr, D. J., Levy, R., Scheepers, C., and Tily, H. J. (2013). Random effects structure for confirmatory hypothesus testing: keep it maximal. J. Mem. Lang. 68, 255–278. doi: 10.1016/j.jml.2012.11.001
Bates, D., Machler, M., Bolker, B. M., and Walker, S. C. (2015). Fitting linear mixed-effects models using lme4. J. Stat. Softw. 67, 1–48. doi: 10.18637/jss.v067.i01
Bishop, D. V. M. (2007). Using mismatch negativity to study central auditory processing in developmental language and literacy impairments: where are we, and where should we be going? Psychol. Bull. 133, 651–672. doi: 10.1037/0033-2909.133.4.651
Brennan, A. A., and Enns, J. T. (2015). When two heads are better than one: interactive versus independent benefits of collaborative cognition. Psychon. Bull. Rev. 22, 1076–1082. doi: 10.3758/s13423-014-0765-4
Brennan, S. E., Chen, X., Dickinson, C. A., Neider, M. B., and Zelinsky, G. J. (2008). Coordinating cognition: the costs and benefits of shared gaze during collaborative search. Cognition 106, 1465–1477. doi: 10.1016/j.cognition.2007.05.012
Coles, M. G., Scheffers, M. K., and Holroyd, C. B. (2001). Why is there an ERN/Ne on correct trials? Response representations, stimulus-related components, and the theory of error-processing. Biol. Psychol. 56, 173–189. doi: 10.1016/S0301-0511(01)00076-X
Cui, F., Zhu, X., Duan, F., and Luo, Y. (2015). Instructions of cooperation and competition influence the neural responses to others ' pain : an ERP study. Soc. Neurosci. 11, 289–296. doi: 10.1080/17470919.2015.1078258
Davis, M. H. (1983). Measuring individual differences in empathy: Evidence for a multidimensional approach. J. Pers. Soc. Psychol. 44, 113–126. doi: 10.1037/0022-3514.44.1.113
de Bruijn, E. R., and von Rhein, D. T. (2012). Is your error my concern? An event-related potential study on own and observed error detection in cooperation and competition. Front. Neurosci. 6:8. doi: 10.3389/fnins.2012.00008
de Bruijn, E. R. A., de Lange, F. P., von Cramon, D. Y., and Ullsperger, M. (2009). When errors are rewarding. J. Neurosci. 29, 12183–12186. doi: 10.1523/JNEUROSCI.1751-09.2009
Delorme, A., and Makeig, S. (2004). EEGLAB: an open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 134, 9–21. doi: 10.1016/j.jneumeth.2003.10.009
Dolk, T., Hommel, B., Colzato, L. S., Schutz-Bosbach, S., Prinz, W., and Liepelt, R. (2014). The jointsimon effect: a review and theoretical integration Front. Psychol. 5:974. doi: 10.3389/fpsyg.2014.00974
Ehinger, B. V., Konig, P., and Ossandon, J. P. (2015). Predictions of visual content across eye movements and their modulation by inferred information. J. Neurosci. 35, 7403–7413. doi: 10.1523/JNEUROSCI.5114-14.2015
Falkenstein, M., Hohnsbein, J., Hoormann, J., and Blanke, L. (1991). Effects of crossmodal divided attention on late ERP components. II. Error processing in choice reaction tasks. Electroencephalogr. Clin. Neurophysiol. 78, 447–455. doi: 10.1016/0013-4694(91)90062-9
Ferdinand, N. K., Mecklinger, A., Kray, J., and Gehring, W. J. (2012). The processing of unexpected positive response outcomes in the mediofrontal cortex. J. Neurosci. 32, 12087–12092. doi: 10.1523/JNEUROSCI.1410-12.2012
Frömer, R., Maier, M., and Rahman, R. A. (2018). Group-level EEG-processing pipeline for flexible single trial-based analyses including linear mixed models. Front. Neurosci. 12:48. doi: 10.3389/fnins.2018.00048
Hari, R., and Kujala, M. V. (2009). Brain basis of human social interaction: from concepts to brain imaging. Physiol. Rev. 89, 453–479. doi: 10.1152/physrev.00041.2007
Hillyard, S. A., and Anllo-Vento, L. (1998). Event-related brain potentials in the study of visual selective attention. Proc. Natl. Acad. Sci. U.S.A. 95, 781–787.
Holroyd, C. B., and Coles, M. G. (2002). The neural basis of human error processing: reinforcement learning, dopamine, and the error-related negativity. Psychol Rev. 109, 679–709. doi: 10.1037/0033-295X.109.4.679
Holroyd, C. B., Hajcak, G., and Larsen, J. T. (2006). The good, the bad and the neutral: electrophysiological responses to feedback stimuli. Brain Res. 1105, 93–101. doi: 10.1016/j.brainres.2005.12.015
Holroyd, C. B., Nieuwenhuis, S., Yeung, N., Nystrom, L., Mars, R. B., Coles, M. G. H., et al. (2004). Dorsal anterior cingulate cortex shows fMRI response to internal and external error signals. Nat. Neurosci. 7, 497–498. doi: 10.1038/nn1238
Houslay, T. M., and Wilson, A. J. (2017). Avoiding the misuse of BLUP in behavioural ecology. Behav. Ecol. 28, 948–952. doi: 10.1093/beheco/arx023
Itagaki, S., and Katayama, J. (2008). Self-relevant criteria determine the evaluation of outcomes induced by others. Neuroreport 19, 383–387. doi: 10.1097/WNR.0b013e3282f556e8
Keller, P. E. (2008). “Joint action in music performance,” in Emerging Communication: Studies on New Technologies and Practices in Communication, Vol. 10, Enacting Intersubjectivity: A Cognitive and Social Perspective on the Study of Interactions, eds F. Morganti, A. Carassa, and G. Riva (Amsterdam: IOS Press), 205–221.
Keller, P. E., Knoblich, G., and Repp, B. H. (2007). Pianists duet better when they play with themselves: on the possible role of action simulation in synchronization. Conscious. Cogn. 16, 102–111. doi: 10.1016/j.concog.2005.12.004
Knoblich, G., and Jordan, J. S. (2003). Action coordination in groups and individuals: learning anticipatory control. J. Exp. Psychol. Learn. Mem. Cogn. 29, 1006–1016. doi: 10.1037/0278-7393.29.5.1006
Koban, L., Pourtois, G., Bediou, B., and Vuilleumier, P. (2012). Effects of social context and predictive relevance on action outcome monitoring. Cogn. Affect. Behav. Neurosci. 12, 460–478. doi: 10.3758/s13415-012-0091-0
Koban, L., Pourtois, G., Vocat, R., and Vuilleumier, P. (2010). When your errors make me lose or win: event-related potentials to observed errors of cooperators and competitors. Soc. Neurosci. 5, 360–374. doi: 10.1080/17470911003651547
Li, P., Peng, W., Li, H., and Holroyd, C. B. (2018). Electrophysiological measures reveal the role of anterior cingulate cortex in learning from unreliable feedback. Cogn. Affect. Behav. Neurosci. 18, 949–963. doi: 10.3758/s13415-018-0615-3
Loehr, J. D., Kourtis, D., and Brazil, I. A. (2015). It's not just my fault: Neural correlates of feedback processing in solo and joint action. Biol. Psychol. 111, 1–7. doi: 10.1016/j.biopsycho.2015.08.004
Loehr, J. D., Kourtis, D., Vesper, C., Sebanz, N., and Knoblich, G. (2013). Monitoring individual and joint action outcomes in duet music performance. J. Cogn. Neurosci. 25, 1049–1061. doi: 10.1162/jocna00388
Marco-Pallarés, J., Krämer, U. M., Strehl, S., Schröder, A., and Münte, T. F. (2010). When decisions of others matter to me: an electrophysiological analysis. BMC Neurosci. 11:86. doi: 10.1186/1471-2202-11-86
Matuschek, H., Kliegl, R., Vasishth, S., Baayen, H., and Bates, D. (2017). Balancing Type I error and power in linear mixed models. J. Mem. Lang. 94, 305–315. doi: 10.1016/j.jml.2017.01.001
Mensen, A., and Khatami, R. (2013). Advanced EEG analysis using threshold-free cluster-enhancement and non-parametric statistics. Neuroimage 67, 111–118. doi: 10.1016/j.neuroimage.2012.10.027
Miltner, W. H., Braun, C. H., and Coles, M. G. (1997). Event-related brain potentials following incorrect feedback in a time-estimation task: evidence for a ‘generic’ neural system for error detection. J. Cogn. Neurosci. 9, 788–798. doi: 10.1162/jocn.1997.9.6.788
Oostenveld, R., and Praamstra, P. (2001). The five percent electrode system for high-resoluti EEEG and ERP measurements. Clin. Neurophys. 112, 713–719. doi: 10.1016/S1388-2457(00)00527-7
Palmer, J. A., Makeig, S., Kreutz-Delgado, K., and Rao, B. D. (2008). “Newton method for the ica mixture model,” in ICASSP, IEEE International Conference on Acoustics, Speech and Signal Processing - Proceedings (Las Vegas, NV), 1805–1808.
Peirce, J. W. (2007). PsychoPy-Psychophysics software in Python. J. Neurosci. Methods 162, 8–13. doi: 10.1016/j.jneumeth.2006.11.017
Picton, L., Saunders, B., and Jentzsch, I. (2012). “I will fix only my own mistakes”: an ERP study investigating error processing in a joint choice-RT task. Neuropsychologia 50, 777–785. doi: 10.1016/j.neuropsychologia.2012.01.011
Pinheiro, J. C., and Bates, D. M. (2000). Mixed-Effects Models in S and S-PLUS. New York, NY: Springer.
Ridderinkhof, K. R., Ullsperger, M., Crone, E. A., and Nieuwenhuis, S. (2004). The role of the medial frontal cortex in cognitive control. Science 306, 443–447. doi: 10.1126/science.1100301
Rigoni, D., Polezzi, D., Rumiati, R., Guarino, R., and Sartori, G. (2010). When people matter more than money: an ERPs study. Brain Res. Bull. 81, 445–452. doi: 10.1016/j.brainresbull.2009.12.003
Saxe, R., and Kanwisher, N. (2003). People thinking about thinking peopleThe role of the temporo-parietal junction in “theory of mind”. Neuroimage 19, 1835–1842. doi: 10.1016/S1053-8119(03)00230-1
Schilbach, L., Timmermans, B., Reddy, V., Costall, A., Bente, G., Schlicht, T., et al. (2013). A second-person neuroscience in interaction. Behavioral and Brain Sciences, 36:441–462. doi: 10.1017/S0140525X12002452
Sebanz, N., Knoblich, G., and Prinz, W. (2003). Representing others' actions: just like one's own? Cognition 88, B11–B21. doi: 10.1016/S0010-0277(03)00043-X
Sebanz, N., Knoblich, G., and Prinz, W. (2005). How two share a task: corepresenting stimulus-response mappings. J. Exp. Psychol. Hum. Percept. Perform. 31, 1234. doi: 10.1037/0096-1523.31.6.1234
Seghier, M. L., and Price, C. J. (2018). Interpreting and utilising intersubject variability in brain function. Trends Cogn. Sci. 22, 517–530. doi: 10.1016/j.tics.2018.03.003
Sinha, N., Maszczyk, T., Wanxuan, Z., Tan, J., and Dauwels, J. (2016). EEG hyperscanning study of inter-brain synchrony during cooperative and competitive interaction. in 2016 IEEE International 630 Conference on Systems, Man, and Cybernetics (SMC) (Budapest: IEEE), 004813-004818.
Smith, S. M., and Nichols, T. E. (2009). Threshold-free cluster enhancement: addressing problems of smoothing, threshold dependence and localisation in cluster inference. Neuroimage 44, 83–98. doi: 10.1016/j.neuroimage.2008.03.061
Ullsperger, M., Fischer, A. G., Nigbur, R., and Endrass, T. (2014). Neural mechanisms and temporal dynamics of performance monitoring. Trends Cogn. Sci. 18, 259–267. doi: 10.1016/j.tics.2014.02.009
van Schie, H. T., Mars, R. B., Coles, M. G., and Bekkering, H. (2004). Modulation of activity in medial frontal and motor cortices during error observation. Nat. Neurosci. 7, 549–554. doi: 10.1038/nn1239
Vesper, C., Abramova, E., Bütepage, J., Ciardo, F., Crossey, B., Effenberg, A., et al. (2017). Joint action: mental representations, shared information and general mechanisms for coordinating with others. Front. Psychol. 7:2039. doi: 10.3389/fpsyg.2016.02039
Vesper, C., Butterfill, S., Knoblich, G., and Sebanz, N. (2010). A minimal architecture for joint action. Neural Netw. 23, 998–1003. doi: 10.1016/j.neunet.2010.06.002
Wahn, B., Czeszumski, A., and König, P. (2018a). Performance similarities predict collective benefits in dyadic and triadic joint visual search. PLoS ONE 13:e0191179. doi: 10.1371/journal.pone.0191179
Wahn, B., Karlinsky, A., Schmitz, L., and König, P. (2018b). Let's move it together: a review of group benefits in joint object control. Front. Psychol. 9:918. doi: 10.3389/fpsyg.2018.00918
Wahn, B., Keshava, A., Sinnett, S., Kingstone, A., and König, P. (2017a). “Audiovisual integration is affected by performing a task jointly,” in Proceedings of the 39th Annual Conference of the Cognitive Science Society (Austin, TX), 1296–1301.
Wahn, B., Kingstone, A., and König, P. (2017b). Two trackers are better than one: information about the co-actor's actions and performance scores contribute to the collective benefit in a joint visuospatial task. Front. Psychol. 8:669. doi: 10.3389/fpsyg.2017.00669
Wahn, B., Kingstone, A., and Peter, K. (2018c). Group benefits in joint perceptual tasks–a review. Ann. N. Y. Acad. Sci. doi: 10.1111/nyas.13843. [Epub ahead of print].
Wahn, B., Schmitz, L., König, P., and Knoblich, G. (2016a). “Benefiting from being alike: interindividual skill differences predict collective benefit in joint object control,” in Proceedings of the 38th Annual Conference of the Cognitive Science Society (Austin, TX), 2747–2752.
Wahn, B., Schwandt, J., Krüger, M., Crafa, D., Nunnendorf, V., and König, P. (2016b). Multisensory teamwork: using a tactile or an auditory display to exchange gaze information improves performance in joint visual search. Ergonomics 59, 781–795. doi: 10.1080/00140139.2015.1099742
Watson, A. B., and Pelli, D. G. (1983). Quest: a Bayesian adaptive psychometric method. Percept. Psychophys. 33, 113–120. doi: 10.3758/BF03202828
Wijers, A. A., Mulder, G., Okita, T., and Mulder, L. J. (1989). Event-related potentials during memory search and selective attention to letter size and conjunctions of letter size and color. Psychophysiology 26, 529–547. doi: 10.1111/j.1469-8986.1989.tb00706.x
Keywords: social cognition, joint action, EEG, feedback related negativity, cooperation, competition
Citation: Czeszumski A, Ehinger BV, Wahn B and König P (2019) The Social Situation Affects How We Process Feedback About Our Actions. Front. Psychol. 10:361. doi: 10.3389/fpsyg.2019.00361
Received: 17 July 2018; Accepted: 05 May 2019;
Published: 25 February 2019.
Edited by:
Karl Christoph Klauer, University of Freiburg, GermanyReviewed by:
Benjamin Ernst, Katholische Universität Eichstätt-Ingolstadt, GermanyAnnelie Rothe-Wulf, University of Freiburg, Germany
Copyright © 2019 Czeszumski, Ehinger, Wahn and König. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Artur Czeszumski, YWN6ZXN6dW1za2lAdW9zLmRl