 Susana Rivas1,2* and Stéphane Genin1,2
Susana Rivas1,2* and Stéphane Genin1,2
- 1 Institut National de la Recherche Agronomique, Laboratoire des Interactions Plantes-Microorganismes, UMR 441, Castanet-Tolosan, France
- 2 Centre National de la Recherche Scientifique, Laboratoire des Interactions Plantes-Microorganismes, UMR 2594, Castanet-Tolosan, France
Plant immune responses depend on the ability to couple rapid recognition of the invading microbe to an efficient response. During evolution, plant pathogens have acquired the ability to deliver effector molecules inside host cells in order to manipulate cellular and molecular processes and establish pathogenicity. Following translocation into plant cells, microbial effectors may be addressed to different subcellular compartments. Intriguingly, a significant number of effector proteins from different pathogenic microorganisms, including viruses, oomycetes, fungi, nematodes, and bacteria, is targeted to the nucleus of host cells. In agreement with this observation, increasing evidence highlights the crucial role played by nuclear dynamics, and nucleocytoplasmic protein trafficking during a great variety of analyzed plant–pathogen interactions. Once in the nucleus, effector proteins are able to manipulate host transcription or directly subvert essential host components to promote virulence. Along these lines, it has been suggested that some effectors may affect histone packing and, thereby, chromatin configuration. In addition, microbial effectors may either directly activate transcription or target host transcription factors to alter their regular molecular functions. Alternatively, nuclear translocation of effectors may affect subcellular localization of their cognate resistance proteins in a process that is essential for resistance protein-mediated plant immunity. Here, we review recent progress in our field on the identification of microbial effectors that are targeted to the nucleus of host plant cells. In addition, we discuss different virulence strategies deployed by microbes, which have been uncovered through examination of the mechanisms that guide nuclear localization of effector proteins.
Introduction
As sessile organisms living in a microbe-rich environment, plants have developed an intricate defense network to fight off invading pathogens. The first layer of plant defense involves recognition of pathogen-associated molecular patterns (PAMPs), defined as invariant epitopes within molecules that are fundamental to pathogen fitness and widely distributed among different microbes. Examples of PAMPs are flagellin from bacteria and chitin from fungi and their recognition, historically known as basal defense, is now referred to as PAMP-triggered immunity (PTI; Jones and Dangl, 2006). PTI is associated to the production of reactive oxygen species and antimicrobial compounds, the induction of mitogen-activated protein kinase (MAPK) cascades, the modulation of host gene transcription, and the deposition of lignin and callose at the plant cell wall (Asai et al., 2002; Torres et al., 2002; Hauck et al., 2003; Tao et al., 2003). Thriving pathogens evolved to suppress PTI and promote successful infection by delivering a plethora of small molecules, referred to as effectors, in the apoplastic space, and inside host cells (Gordeeva et al., 2003; Alfano and Collmer, 2004; Chisholm et al., 2006). In turn, plants have evolved to gain the ability to recognize directly or indirectly effectors through resistance (R) proteins. This recognition response, which leads to resistance of the plant, is associated with the long-standing gene-for-gene hypothesis and, more recently, with the guard hypothesis (van der Biezen and Jones, 1998), and is now known as effector-triggered immunity (ETI). ETI is frequently associated with development of the hypersensitive response (HR), a form of programmed cell death localized at the infection site, which prevents the spread of the pathogen inside the plant (Mur et al., 2008). In most cases, the onset of the HR results in the activation of systemic acquired resistance (SAR), which provides protection to the plant against a wide range of pathogens (Durrant and Dong, 2004). The co-evolutionary arms race between plants and pathogens has resulted in the generation of highly polymorphic repertoires of R proteins and microbial effectors.
Plant pathogenic bacteria, viruses, fungi, oomycetes, and nematodes engage varied mechanisms to deliver effector proteins inside host cells. To achieve this goal, bacteria use specialized secretion systems, such as the type III and type IV secretion systems (Galan and Wolf-Watz, 2006; Block et al., 2008; McCann and Guttman, 2008). Biotrophic fungi and oomycetes have evolved specialized structures called haustoria that penetrate the host tissue, invaginating the host plasma membrane. Haustoria probably play a dual role as they have been proposed to enable effector secretion and uptake of nutrients, although this idea remains to be demonstrated (Panstruga and Dodds, 2009; de Jonge et al., 2011). In contrast, several fungal and oomycete effectors can enter plant cells independently of the pathogen, probably via receptor-mediated endocytosis (Kale and Tyler, 2011). Finally, plant parasitic nematodes use a specialized feeding organ, referred to as the stylet, to inject their effector proteins into parasitized plant vascular cells (Davis et al., 2008; Gheysen and Mitchum, 2011).
Following their translocation into plant cells, microbial effectors may be addressed to different subcellular compartments where they may manipulate a variety of host cellular functions. A particularly important role in plant defense responses has been attributed to nuclear dynamics since a growing number of reports has revealed that nuclear localization of not only pathogen effectors, but also of R proteins and key host components, including transcription factors (TFs) and regulators, is essential for plant immunity (Wiermer et al., 2007; Liu and Coaker, 2008; Deslandes and Rivas, 2011; Rivas, 2011). The fact that a significant number of effector proteins is translocated into the host cell nucleus (Table 1) suggests that effectors may manipulate host transcription or directly target essential nuclear host components for the benefit of the pathogen. Along these lines, it has been additionally proposed that some effectors may affect histone modification and chromatin remodeling (Kay and Bonas, 2009). Indeed, chromatin configuration allows or prevents protein access to specific DNA regions and regulates essential cellular processes such as DNA replication, DNA repair, and transcription (Clapier and Cairns, 2009). Although chromatin remodeling has only been formally demonstrated in the case of effector proteins from Agrobacterium tumefaciens (Citovsky et al., 2007), it is now well accepted that modulation of chromatin configuration is a strategy employed by bacterial virulence proteins to subvert plant immunity (Ma et al., 2011). Alternatively, nuclear translocation of effectors may affect subcellular localization of their cognate R proteins in a process that is essential for R protein-mediated plant immunity (Burch-Smith and Dinesh-Kumar, 2007; Shen and Schulze-Lefert, 2007).
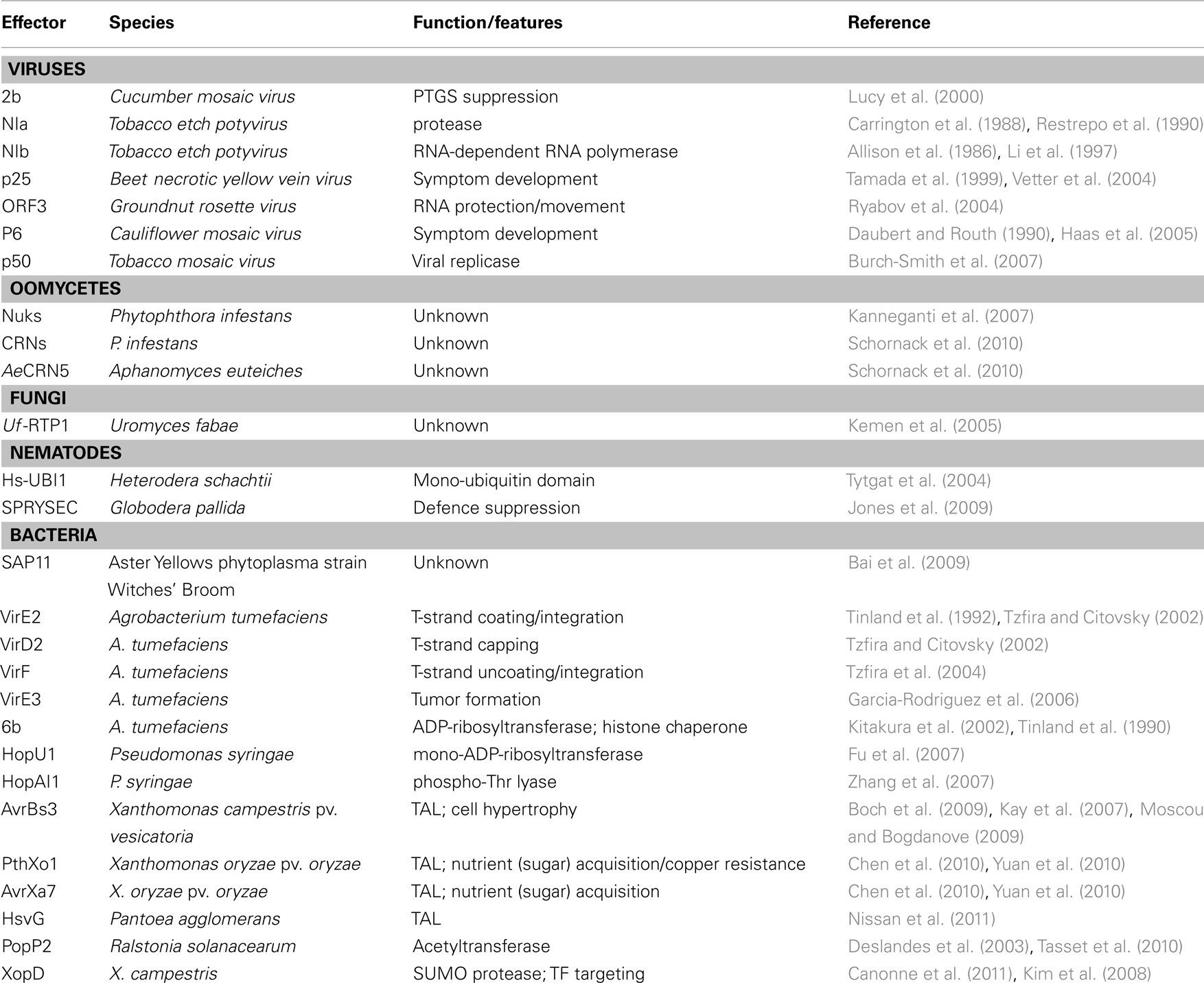
Table 1. Examples of effector proteins with demonstrated nuclear localization from different phytopathogenic microorganisms.
In eukaryotic cells, trafficking of protein and RNA molecules between the cytoplasm and the nucleus occurs through nuclear pore complexes (NPCs). NPCs are composed of multiple subunits of nucleoporins (Nups) organized in a donut-shaped complex of eightfold symmetry that spans the double membrane of the nuclear envelope (Meier, 2007; Figure 1). Nucleocytoplasmic transport of macromolecules depends on import and export receptors (importins and exportins), which translocate cargo through the nuclear pore following their respective recognition of exposed nuclear localization signals (NLSs) and nuclear export signals (NESs) on cargo proteins (Merkle, 2001; Meier, 2007; Patel et al., 2007). NLSs are typically classified as either monopartite NLSs composed of a continuous stretch of basic amino acids (Kalderon et al., 1984a,b), or bipartite NLSs composed of two sets of two to three positively charged amino acids separated by a 10-amino acid linker region (Dingwall et al., 1982; Robbins et al., 1991). α-importins are adapter proteins that bind to NLS-containing cargo proteins and bridge their interaction with importin β. The trimeric complex importin-α/importin-β/cargo protein is translocated into the nucleus across the NPC, thanks to the interaction between Nups and importin β (Figure 1). The directionality of the nuclear transport is maintained by the ratio of additional import factors such as Ran-GDP (cytoplasmic side) and Ran-GTP (nuclear side). Upon binding of Ran-GTP in the nucleus, the complex is disassembled and importin-α and β shuttle back to the cytoplasm to allow additional rounds of transport (Merkle, 2001; Meier, 2007; Figure 1). Nuclear export of proteins classically occurs through the nuclear export pathway mediated by an evolutionarily conserved CRM1/exportin protein that belongs to the importin-β family. The CRM1–Ran-GTP complex binds directly to the leucine-rich NES contained in cargo and directs the export of the ternary complex from the nucleus. The cargo is released from the complex by hydrolysis of Ran-GTP to Ran-GDP in the cytoplasm (Ossareh-Nazari et al., 2001; Figure 1).
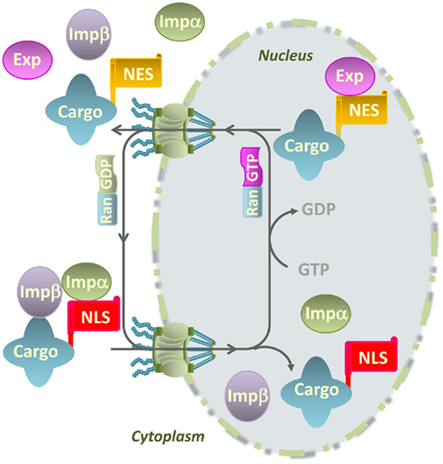
Figure 1. Model for nucleocytoplasmic transport of macromolecules through the nuclear pore complex. Cytoplasmic proteins with a nuclear localization signal (NLS) are translocated into the nucleus through nuclear pore complexes that are composed of nucleoporins and span the double membrane of the nuclear envelope. The NLS in cargo proteins is recognized by importin-α (Impα) that bridges the interaction of cargo proteins with importin-β (Impβ) and promote their transport into the nucleus. The directionality of transport is maintained by the Ras-related nuclear (Ran) protein through its binding to GDP (cytoplasmic side) or GTP (nuclear side). Export receptors or exportins (Exp) recognize nuclear export signals (NES) in cargo proteins, promoting their nuclear export to the cytoplasm.
Mounting evidence suggests that pathogen effectors co-opt the host cell nuclear transport machinery, including α-importins, to target plant cell nuclei (Vetter et al., 2004; Kay and Bonas, 2009; Schornack et al., 2010). In other cases, α-importin-independent translocation of effector proteins into the nucleus has been reported (Kanneganti et al., 2007). In addition, mutations in cellular factors involved in the transport of macromolecules through the nuclear envelope, compromise plant resistance signaling, underlining the importance of nucleocytoplasmic trafficking during plant innate immunity (Kinkema et al., 2000; Mou et al., 2003; Zhang and Li, 2005).
Here, we provide an overview on our current knowledge about translocation of microbial effectors into the host cell nucleus. We first discuss nuclear-targeted effectors from bacteria and viruses, whose functional characterization is rather well documented. Second, we summarize recent data involving nuclear effectors from nematodes and filamentous pathogens, for which further functional studies are required before concluding on the specificities and commonalities of the virulence strategies that are hidden behind nuclear targeting of effector proteins.
Agrobacterium tumefaciens
Agrobacterium tumefaciens is a soil phytopathogenic bacterium with the unique ability to transfer a segment of its Ti plasmid (T-DNA) into plant cells and integrate it into the chromosomal DNA. As a result, Agrobacterium infection induces the formation of crown gall tumors on dicotyledonous plants (Gelvin, 1998). T-DNA is transported from the bacterium to the host cell as a single-stranded DNA (ssDNA) molecule, named the T-strand, via the so-called type IV secretion system (T4SS), which is ancestrally related to bacterial conjugation machines (Christie, 2004; Christie et al., 2005). Conjugation systems enable bacteria to adapt to changing environments through acquisition of fitness traits. Throughout evolution, conjugation has substantially contributed to genome rearrangement and plasticity. The genes responsible for the processing and transfer of the T-DNA to plant cells are present in the Virulence region of the Ti plasmid and are called vir genes. The translocated virulence proteins are effector proteins, also delivered into the host cell through the T4SS. Once in the plant cell, Vir proteins form a nucleoprotein complex (T-complex) with the T-strand, as well as with host proteins, to mediate the transformation of a normal plant cell into a crown gall tumor cell (Lacroix and Citovsky, 2009).
A subset of the Vir proteins that are translocated into plant cells is targeted to the nucleus and historically Vir proteins were the first nuclear-localized effectors described (Tinland et al., 1992). Studies on nuclear-targeted proteins from Agrobacterium provide a fascinating illustration of the great diversity of molecular activities that bacterial effectors are able to display in the host nucleus to subvert basic cellular processes. Among the Vir proteins involved in the processing and/or transfer of the T-DNA to the plant cell, VirE2 is the most abundant protein produced after induction of the vir genes. It binds in an unspecific and highly cooperative manner to ssDNA, coating the T-strand to protect it from the attack of host nucleases (Figure 2). Another Vir protein, VirD2, is covalently attached to the 5′ end of the T-strand following generation of nicks at the border repeats surrounding the T-DNA in the Ti plasmid (Tzfira and Citovsky, 2002; Figure 2). VirE2 is translocated into plant cells independently of the T-strand-VirD2 complex (Vergunst et al., 2000). Both VirD2 and VirE2 have NLSs, which helps target the T-strand to the plant cell nucleus. However, nuclear import of the T-complex is mediated by VirD2, following its binding to the host importin-α (Ballas and Citovsky, 1997; Bako et al., 2003). VirE2, which has low binding affinity for importin-α (Citovsky et al., 2004), interacts with the nuclear protein VIP1 (VirE2-interacting protein 1; Tzfira et al., 2001) that then binds to importin-α and guides VirE2 nuclear import, and association with the host chromatin (Tzfira and Citovsky, 2002; Citovsky et al., 2004; Lacroix et al., 2008). VIP1 is a basic domain-leucine zipper (bZIP) protein that may act as a TF involved in plant defense (Tzfira et al., 2001; Djamei et al., 2007).
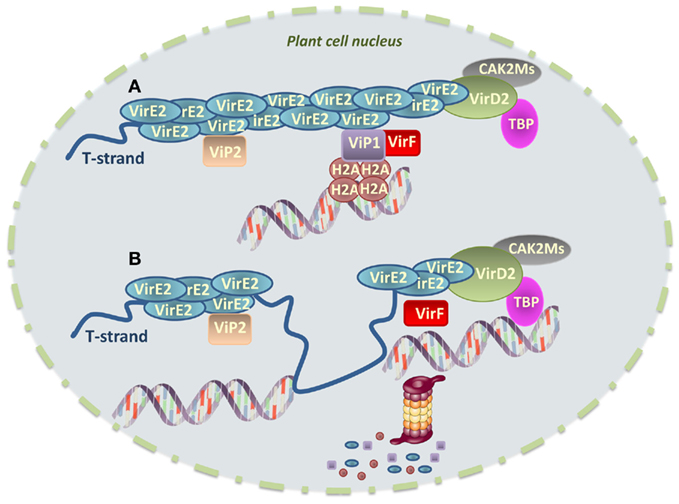
Figure 2. Model for integration of Agrobacterium tumefaciens T-DNA into the host cell chromatin. (A) The T-strand is transported into the host cell nucleus as an ssDNA molecule. VirE2 coats the T-strand to protect it from the attack of host nucleases, whereas VirD2 is covalently attached to the 5′ end of the T-strand. VirE2 interacts with the nuclear protein VIP1, which acts as a molecular bridge between VirE2 and nucleosomes thanks to its association with core histones, thereby facilitating T-DNA integration. The role of VIP2 in this process remains to be determined. VirD2-intercating proteins CAK2Ms and TBP are also represented. VirF interacts with VIP1 attached to both nucleosomes and the T-complex. (B) Before integration, VirF helps uncoat the T-strand promoting proteasomal degradation of VIP1, VirE2 and, very likely, VIP1-interacting core histones. This creates a chromatin environment favorable for T-DNA integration. Whether synthesis of the second-strand occurs before or after association of the T-strand with the chromatin remains unknown. It is possible that second-strand synthesis and integration represent coupled events.
VirE2 has been shown to modulate chromatin functions and facilitate T-DNA integration. In Arabidopsis, VIP1 directly interacts with various core histones, such as H2A, and it may therefore bridge the association of VirE2 with the plant nucleosome, facilitating T-DNA integration (Li et al., 2005; Lacroix et al., 2008; Figure 2). Furthermore, another VirE2-interacting protein, VIP2, may regulate histone gene transcription (Anand et al., 2007). In agreement with this observation, expression of several histone genes is induced upon Agrobacterium infection (Veena et al., 2003). It is thus tempting to speculate that VirE2 and/or other Agrobacterium effectors may modulate histone gene expression to facilitate infection.
VirD2-interacting proteins have also been described. For example, the nuclear kinase CAK2Ms, a member of the conserved cyclin-dependent kinase-activating kinase family, interacts with, and phosphorylates VirD2, perhaps modulating its activity. In addition, a TATA box-binding protein (TBP) also interacts with VirD2, possibly guiding the T-DNA to transcription-prone sites in the host genome, which are preferential for T-DNA integration (Bako et al., 2003; Figure 2).
An additional group of Vir proteins is involved in hijacking the host cell metabolism to allow the integration and expression of the genes carried on the T-DNA. Before integration, VirF, an F-box protein translocated into the host cell, helps uncoat the T-DNA from VirE2 and VIP1 proteins (Schrammeijer et al., 2001). Indeed, as part of the Skp1-Cdc53-cullin-F-box (SCF) complex, VirF binds VIP1 that is attached both to nucleosomes and to the T-complex, promoting proteasomal degradation of both VIP1 and its associated VirE2 (Tzfira et al., 2004; Figure 2). It has been proposed that VirF may additionally induce degradation of the core histones that are bound to VIP1 (Li et al., 2005; Loyter et al., 2005; Lacroix et al., 2008), thereby inducing their local destabilization and creating a chromatin environment favorable for T-DNA integration (Figure 2). In agreement with the observation that some plant species do not require VirF for transformation (Hirooka et al., 1987), recent data show that a plant F-box protein, named VBF (VIP1-binding F-box protein), is able to functionally replace VirF. VBF is induced by Agrobacterium infection and, as VirF, regulates the protein levels of VIP1 and its associated VirE2 after binding to VIP1 (Zaltsman et al., 2010). VBF expression in and export from Agrobacterium lead to increased tumorigenesis, indicating that Agrobacterium subverts a component of the host cell proteasome, whose expression is induced during pathogen infection, to promote plant genetic transformation.
VirE3 is conserved among all the Agrobacterium Ti plasmids studied so far. Even the Agrobacterium rhizogenes Ri-plasmid that lacks the genes virE1 and virE2 contains a copy of virE3, suggesting that the VirE3 protein plays an important role during transformation (Garcia-Rodriguez et al., 2006). Indeed, mutations in virE3 diminish tumor formation on tobacco, tomato, and sunflower (Garcia-Rodriguez et al., 2006). The VirE3 protein has NLSs that mediate its interaction with importin-α and nuclear localization (Garcia-Rodriguez et al., 2006). VirE3 additionally interacts with pCsn5, a component of the COP9 signalosome and pBrp, a plant-specific general TF that belongs to the TFIIB family. When bound to DNA, VirE3 is able to promote gene transcription. These data suggest that nuclear VirE3 may act as a transcriptional activator to induce the expression of genes needed for tumor development (Garcia-Rodriguez et al., 2006).
Finally, the 6b gene in the T-DNA from A. tumefaciens has oncogenic activity in plant cells, inducing tumor formation, and alterations in leaf morphology (Tinland et al., 1990). The tobacco protein NtSIP1 was identified in a yeast two-hybrid screen as a 6b-interacting protein (Kitakura et al., 2002). NtSIP1 is localized in the nucleus and appears to act as a TF because its predicted amino acid sequence includes two predicted NLSs and a putative DNA-binding motif, which is similar to the triple helix motif of rice TF GT-2 (Dehesh et al., 1992). Nuclear localization of 6b was enhanced by co-expression with NtSIP1 in tobacco cells. In addition, a fusion protein between the DNA-binding domain of yeast GAL4 and 6b activated the transcription of a reporter gene in tobacco (Kitakura et al., 2002). Finally, an acidic C-terminal domain of 6b is required for its nuclear localization and transactivation as well as for hormone-independent proliferation of tobacco cells. These data suggest that 6b may affect transcription of plant genes controlled by NtSIP1 and function in the proliferation of plant cells through an association with NtSIP1. In addition to NtSIP1, 6b associates with other Arabidopsis proteins in the nucleus, including key components of the microRNA pathway and the core histone H3 (Terakura et al., 2007; Wang et al., 2011). Based on this finding, 6b has been proposed to act as a histone chaperone, which works together with other chromatin remodelers to affect nucleosome assembly, histone displacement and transcription in a gene-specific manner (Terakura et al., 2007). Recent structural analysis suggests that 6b displays an ADP-ribosyltransferase activity (Wang et al., 2011). Since 6b directly interacts with H3, it will be interesting to determine whether H3 can be modified by 6b and how potential 6b-mediated ribosylation of H3 may affect transcription.
Other Gram-Negative Bacteria
Gram-negative bacteria have evolved a sophisticated mechanism to deliver effector proteins into host cells. Indeed, the so-called type III secretion system (T3SS) provides a continuous channel for Type III effectors (T3Es) to travel from the bacterial cytoplasm directly into the cytoplasm of eukaryotic cells in a process that involves transport across (i) the two bacterial membranes separated by a peptidoglycan layer and (ii) the plasma membrane of the plant cell, which is surrounded by a thick cell wall (Buttner and He, 2009). In plant pathogenic bacteria, T3SSs are encoded by hrp (for HR and pathogenicity) genes that are required by bacteria to elicit the HR in resistant plants and to cause disease in susceptible plants (Lindgren et al., 1986)
In Ralstonia solanacearum, PopB is a small basic T3SS-secreted protein that carries a functional C-terminal bipartite NLS and a predicted helix forming a coiled-coil domain, suggesting that PopB might interact with other proteins (Gueneron et al., 2000). In addition to PopB and PopP2 (see below), at least three other R. solanacearum T3Es of unknown function (RSp0216, RSc1349, and RSc3272) appear to be nuclear localized when expressed in plant cells (Anne-Claire Cazalé, personal communication).
The T3E from Xanthomonas campestris pathovar vesicatoria (Xcv) HpaA (for Hrp-associated) is specifically required for disease development in pepper plants. hpaA mutants are affected in pathogenicity whereas they partially retain the ability to induce an HR (Huguet et al., 1998). HpaA presents two functional NLSs that are important for full HpaA-mediated disease development in the plant (Huguet et al., 1998).
Effector mining in Pseudomonas syringae pv. tomato DC3000, which infects Arabidopsis thaliana and tomato, has identified more than 30 candidate effector genes (Grant et al., 2006; Lindeberg et al., 2006; Cunnac et al., 2009). Systematic survey of P. syringae T3Es containing putative NLSs has identified AvrE, Hopl1, and HopY1, although their nuclear localization in plant cells remains to be demonstrated (Lionel Navarro, personal communication). The P. syringae T3E HopU1 displays mono-ADP-ribosyltransferase (ADP-RT) activity and mutation of HopU1 catalytic site abolishes HopU1-mediated suppression of plant innate immunity (Fu et al., 2007). HopU1 targets Arabidopsis RNA-binding proteins with RNA-recognition motifs (RRMs), including GRP7, a glycine-rich RNA-binding protein (GR-RBP). Arabidopsis grp7 mutant displays increased susceptibility to P. syringae inoculation as compared to wild-type plants. Both HopU1 and GRP7 present a dual nucleo-cytoplasmatic localization (Fu et al., 2007). The observation that ADP-ribosylation of GRP7 by HopU1 requires two arginine residues within the RRM indicates that this modification may affect the RNA-binding ability of GRP7. By disabling the function of GR-RBPs, the pathogen may modify the host RNA status, resulting in reduced amounts of immunity-related mRNAs available in the plant and suppression of host immunity.
The P. syringae T3E HopAI1 displays phospho-Thr lyase activity and disrupts defense signal transduction by directly inactivating MAPKs in plants (Zhang et al., 2007). A nuclear localization of HopAI1 has not been reported. However, based on the functional similarities between HopAI1 and the T3E OspF of the animal pathogen Shigella flexneri, which also displays phospho-Thr lyase activity that targets the phosphorylated MAPKs in the nucleus (Li et al., 2007), it is tempting to speculate that HopAI1 may be targeted to host cell nuclei. Interestingly, OspF remodels host chromatin by inducing dephosphorylation and deacetylation of H3, which leads to decreased expression of specific immunity-related genes (Arbibe et al., 2007). This function is accomplished through the interaction of OspF with host retinoblastoma protein, which has been linked to histone modification (Zurawski et al., 2009). Whether HopAI1 is also able to target nuclear MAPKs and/or modulate histone modifications in plant cells is an exciting perspective for future research.
Although in most cases the cellular activities that are manipulated by nuclear effectors remain to be determined, the action of the following nuclear T3Es has been relatively well characterized and illustrates varied pathogen strategies that lead to the establishment of a cellular environment that favors pathogen proliferation.
TAL Effectors from Xanthomonas and Ralstonia
Transcription activator-like (TAL) proteins, also called AvrBs3 family members, are T3Es only identified to date in plant pathogenic Xanthomonas spp. and R. solanacearum (for recent reviews, see Boch and Bonas, 2010; Bogdanove et al., 2010; Scholze and Boch, 2010). Historically, TAL effectors were the first plant pathogen T3Es shown to be specifically addressed to the nuclear compartment (Yang and Gabriel, 1995; Van den Ackerveken et al., 1996). Moreover, AvrBs3 is also a rare example of a plant pathogen T3E for which translocation into the plant nucleus was evidenced by immunodetection after bacterial infection (Szurek et al., 2002).
Transcription activator-like effectors act as transcriptional activators in the plant cell nucleus and provide a fascinating example of manipulation of the eukaryotic transcriptional machinery by directly promoting specific host gene reprogramming. More than a hundred TAL candidate sequences have been identified to date, mostly in Xanthomonas spp. genomes. Most TAL proteins contain NLSs and an acidic domain involved in transcriptional activation and localize to the nucleus through their interaction with importin-α (Szurek et al., 2001). However, the signature domain of TAL effectors is located in the central part of the proteins and consists of tandemly arranged nearly identical repeat units (Figure 3A). These repeats are 34- or more rarely 35-amino acid long and the number of repeats may vary from 1.5 to 33.5 repeats (Boch and Bonas, 2010) although a minimum of 6.5 repeats is required to detect a transcription activator function (Boch et al., 2009). These TAL repeats were shown to be a novel type of DNA-binding domain (Kay et al., 2007; Romer et al., 2007) in which the target DNA-recognition specificity results from a one-repeat-to-one-bp correlation with different repeat types exhibiting a different DNA base pair specificity (Boch et al., 2009; Moscou and Bogdanove, 2009). Each repeat domain of a TAL effector contains two hypervariable residues at positions 12 and 13 per repeat which have been termed RVDs (for repeat-variable di-residues) and that are directly involved in the pairing to one specific nucleotide of the target DNA sequence. Some RVDs are specific for a particular DNA bp whereas others recognize more than one bp (Figure 3A; Boch et al., 2009; Moscou and Bogdanove, 2009). The succession of RVDs in each repeat therefore determines the nature of the target promoter sequence that has been defined as a “TAL box” element (Scholze and Boch, 2011). Quite remarkably, this specific recognition property of TAL effectors can be used to create artificial TAL effectors with novel DNA-recognition specificities and opens the way to various biotechnological applications (Boch et al., 2009; Geissler et al., 2011).
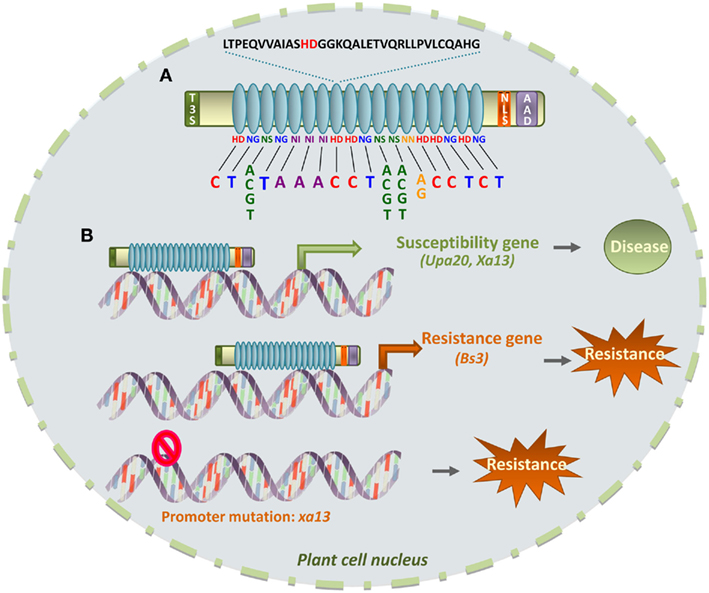
Figure 3. TAL effector-DNA specificity and implications in plant disease/resistance. (A) TAL effectors contain an N-terminal domain required for T3SS-dependent secretion (T3S), a tandem repeat domain (in blue), nuclear localization signals (NLS) and an acidic activation domain (AAD). The central repeat domain confers DNA-binding specificity. One 34-amino acid repeat is shown with the variable di-residue (in red) at positions 12 and 13. The type of di-residue confers specificity for one or several DNA bases, as indicated, and di-residues from each repeat define a specific DNA “TAL box.” (B) Upon binding to the TAL box in the promoter of a plant susceptibility gene, transcription is activated (green arrow), which contributes to disease development. Plant resistance may result from either a molecular decoy strategy where the TAL box drives the expression of a plant “executor” (Resistance) gene (orange arrow) to counter the pathogen, or from occurrence of a mutation in the TAL box DNA sequence which prevents binding of the TAL effector.
First structural data on TAL repeats revealed that a repeat folds into a helix-turn-helix structure reminiscent of a tetratrico peptide repeat (TPR; Murakami et al., 2010). TPRs are 34-amino acid long but are known to be involved in protein–protein interactions in prokaryotes and eukaryotes rather than protein–DNA interactions. The evolutionary origin of TAL effectors remains unknown but several types of TAL repeats exist in nature: whereas 34 aa repeats are prevalent in Xanthomonas spp., 35 aa repeats are found in R. solanacearum (Cunnac et al., 2004) or some Xanthomonas strains (Kay et al., 2005; Schornack et al., 2008). The biological implications of such differences are unclear but raise intriguing evolutionary questions since they suggest that TAL effectors containing 35-amino acid repeats independently arose by successive duplications of an initial variant repeat.
The pathogenic strategy of some Xanthomonas spp. appears to rely massively on host gene reprogramming by TAL effectors since some X. oryzae strains can harbor up to 28 TAL family members (including pseudogenes; Gonzalez et al., 2007), several of them being essential virulence factors for infection of rice (White et al., 2009). No TAL effector candidates were identified in the X. campestris pv. campestris (Xcc) strains B100, 8004, and ATCC 39913 for which a full sequence is available (da Silva et al., 2002; Vorhölter et al., 2003; Qian et al., 2005). However, recent work has identified between 1 and 4 TAL effector sequences in the genomes of 60% of a total of 55 surveyed Xcc strains (Boris Szurek and Laurent Noël, personal communication). The molecular functions of these newly identified TAL effector candidates are unknown but this finding suggests that TAL-based virulence strategies are widespread among xanthomonads.
The role of most TAL effectors in virulence is still unknown but there is some evidence of their role in pathogen proliferation and dispersal. For example, AvrBs3 from X. campestris pv. vesicatoria induces cell hypertrophy in leaves of susceptible pepper hosts, leading to epidermal rupture that is thought to be involved in bacterial dissemination (Marois et al., 2002). AvrBs3 binds to the UPA box, a TAL box found in the promoters of some pepper genes including UPA20 (Figure 3B). UPA20 encodes a TF that controls expression of auxin-induced genes and α-expansins and is crucial for the development of plant cell hypertrophy (Kay et al., 2007; Romer et al., 2007).
A second example illustrating how bacteria use a TAL effector to manipulate host transcription to its own benefit involves the rice gene Xa13. Xa13, which is transcriptionally activated by the TAL effector PthXo1 (Yang et al., 2006; Romer et al., 2010), is defined as a susceptibility gene since its expression facilitates X. oryzae pv. oryzae (Xoo) infection. It was recently shown that Xa13 corresponds to OsSWEET11 that belongs to a recently discovered family of sugar transporters mediating sugar efflux in plants (Chen et al., 2010). PthXo1-mediated induction of OsSweet11 transcription most probably causes glucose efflux from rice cells in order to feed bacteria. An additional TAL effector from Xoo, AvrXa7, triggers induction of OsSWEET14, and consequently allows overcoming xa13-mediated resistance by inducing sugar release through another SWEET transporter (Chen et al., 2010). A separate report showed that Xa13 is a plasma membrane protein interacting with two copper transporter proteins that contribute to decrease the copper content of xylem sap (Yuan et al., 2010). Since Xoo is a vascular pathogen that spreads through xylem vessels, PthXo1-dependent increased transcription of Xa13 might pave the way for Xoo successful infection by clearing its path of toxic amounts of copper (Yuan et al., 2010).
In order to circumvent TAL-mediated pathogenic strategies, resistant plants have developed defense strategies based on subverting the recognition specificities of TAL effectors to trap the pathogen. Indeed, point mutations in the promoter of a key susceptibility gene may alter TAL effector recognition, and subsequent gene activation, leading to plant resistance. For example, the resistance xa13 and susceptibility Xa13 alleles show sequence polymorphisms only in their promoter regions, resulting in induced expression of Xa13 but not recessive xa13 upon Xoo infection (Figure 3B; Chu et al., 2006). However, such a resistance strategy would be much limited (or presumably take longer to be effective) in some other cases where TAL effectors target multiple host susceptibility genes. The pepper Bs3 resistance gene illustrates a more subtle mechanism to promote resistance since, in this case, TAL effectors are lured into inducing the expression of a gene that promotes plant defense (Figure 3B; Boch and Bonas, 2010). Induction of Bs3 expression specifically triggers resistance and HR responses to Xanthomonas strains carrying the TAL T3E AvrBs3 (or related effectors displaying the same DNA-recognition specificity; Romer et al., 2007). Bs3 encodes a protein homologous to flavine-dependent mono-oxygenases, which play roles in auxin biosynthesis and glucosinolate metabolism, therefore contributing to plant defense against pathogens (Schlaich, 2007). Although it is not known whether Bs3-triggered cell death is due to toxin production or induction of defense signaling, resistant plants in this case have evolved a molecular trap to detect AvrBs3 activity. Indeed, by mimicking the upa20 promoter, BS3 subverts the virulence function of AvrBs3 and instead initiates plant immunity (Romer et al., 2007). Interestingly, this strategy should prove useful to engineer durable resistance to multiple pathogenic strains (or species). Along these lines, it has been already demonstrated that different TAL boxes combined into one promoter render this promoter responsive to several TAL effectors (Romer et al., 2009).
Finally, similar to Xanthomonas TAL proteins, HsvG and HsvB effectors of gall-forming Pantoea agglomerans act as transcriptional activators, although they are structurally distinct from TAL effectors (Nissan et al., 2006). HsvG and HsvB determine host specificity on gypsophila and beet, respectively. Both proteins present two functional NLSs required for their nuclear targeting and are able to bind DNA and activate transcription (Nissan et al., 2006; Weinthal et al., 2011). Although the mode of action of HsvG and HsvB remains unknown, it has been hypothesized that it involves modulation of host phytohormones associated with gall formation. A recent report showed that HsvG induces the transcriptional activation of a gene named HSVGT in Gypsophila paniculata (Nissan et al., 2011). HSVGT encodes a predicted acidic protein of the DnaJ family, which presents a bipartite NLS as well as zinc-finger and leucine zipper DNA-binding motifs, typical of TFs. HsvG binds to the HSVGT promoter indicating that HSVGT is a direct target of HsvG (Nissan et al., 2011). These data indicate that HsvG functions as a TF in gypsophila.
XopD from Xanthomonas campestris
XopD from the bacterial strain Xcv (XopDXcv) promotes bacterial growth and delays the onset of leaf chlorosis and necrosis in late infection stages of tomato, presumably to sustain bacterial populations in infected tissues (Kim et al., 2008). XopDXcv is a modular protein of 760 amino acids that shows different biochemical activities and contains (i) a recently identified N-terminal domain of 215 amino acids (Canonne et al., 2010), (ii) a helix-loop-helix domain (HLH), (iii) two tandemly repeated EAR (ERF-associated amphiphilic repression) motifs, previously described in plant transcriptional repressors during defense responses (Kazan, 2006), and (iv) a C-terminal cysteine protease domain with structural similarity with the yeast ubiquitin-like protease 1 (ULP1; Figure 4A). Consistent with its protein structure, XopDXcv displays small ubiquitin-like modifier (SUMO) protease (Canonne et al., 2010) and non-specific DNA-binding activities (Kim et al., 2008). XopDXcv has been additionally shown to repress transcription of defense- and senescence-associated plant genes in an EAR-dependent manner, suggesting that XopDXcv may target host TFs (Kim et al., 2008).
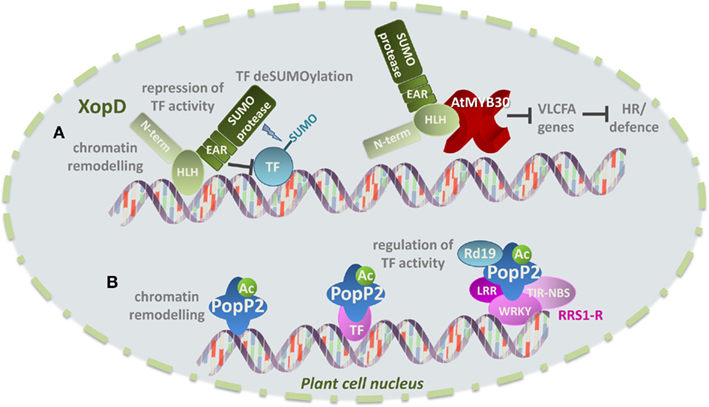
Figure 4. Examples of virulence strategies displayed by bacterial effectors in the plant cell nucleus. (A) XopD from the strain B100 of Xcc is a modular protein with an N-terminal domain of unknown function, a helix-loop-helix domain (HLH), three tandemly repeated transcriptional repressor domain of the EAR type and a C-terminal SUMO protease domain. It has been suggested that XopD DNA-binding activity through its HLH domain may provide access to chromatin and that XopD may thus modulate host transcription by altering chromatin remodeling. XopD may additionally interact with host TFs and repress their transcriptional activity directly via its EAR domains and/or by TF deSUMOylation. Recent work shows that the HLH domain of XopD targets the Arabidopsis TF AtMYB30 leading to repression of AtMYB30 trasncriptional activity and suppression of the plant HR and defense responses. (B) The R. solanacearum effector protein PopP2 displays acetyltransferase activity, suggesting that PopP2 may directly manipulate host transcription through chromating remodeling following acetylation of histone residues. PopP2 acetyltransferase activity may also promote the recruitment of TFs and enhance their DNA-binding affinity, resulting in modulation of host transcription. Finally, the interaction of PopP2 with host proteins, such as the cysteine protease RD19 and the resistance gene RRS1-R may additionally affect RRS1-R-meadited transcriptional reprogramming.
XopD is targeted to plant cell subnuclear structures named nuclear bodies (or nuclear foci; Hotson et al., 2003; Canonne et al., 2010). The molecular mechanism allowing XopD nuclear import remains unknown but a truncated XopD version containing only the HLH domain of XopD (amino acids 216–405), and not comprising its putative NLS, has been shown to be necessary and sufficient for XopD nuclear import and subnuclear targeting (Canonne et al., 2011). Intriguingly, expression of XopD appears to induce reorganization of the nuclear structure in host cells, leading to non-specific relocalization of all tested nuclear proteins into nuclear bodies. In addition, DAPI staining showed that DNA accumulation is weaker in nuclear bodies, where XopD is expressed, compared to the nucleoplasm, where DNA distribution seems to remain unaltered (Canonne et al., 2011). It is thus enticing to speculate that XopD-induced modification of the nuclear structure and protein distribution may be part of a general virulence strategy, which allows Xanthomonas to perturb plant cell responses to bacterial infection. Along these lines, it has been proposed that the presence of the HLH and EAR domains in XopD may provide access to chromatin and/or transcriptional units, leading to modulation of host transcription by affecting chromatin remodeling and/or TF activity (Kay and Bonas, 2009; Figure 4A).
In agreement with the idea that plant TFs and/or regulators might be direct targets of XopD, recent data show that XopD from the strain B100 of X. campestris pathovar campestris (XopDXccB100), which presents a similar protein structure to XopDXcv (Canonne et al., 2012), is able to target the Arabidopsis MYB TF AtMYB30. AtMYB30 is a positive regulator of defense and cell death associated responses through the activation of the lipid biosynthesis pathway that leads to the production of very long chain fatty acids (VLCFAs; Raffaele et al., 2008). In agreement with the finding that transcriptional activation of VLCFA-related genes by AtMYB30 is required to mount an efficient defense response during bacterial infection, it has been demonstrated that AtMYB30 transcriptional activity is tightly controlled by the plant cell. Indeed, AtMYB30 is able to induce partial nuclear relocalization of the secreted phospholipase AtsPLA2-α, which is otherwise localized intracellularly in Golgi-associated vesicles before being secreted to the extracellular space. The physical interaction between AtMYB30 and AtsPLA2-α leads to repression of the AtMYB30-mediated transcriptional activity and negative regulation of plant HR and defense responses (Froidure et al., 2010). These data highlight the importance of dynamic nucleocytoplasmic protein trafficking for the regulation of defense-related transcription. Interestingly, in addition to the control of AtMYB30 activity exerted by the plant cell, AtMYB30 transcriptional activation is additionally controlled by bacteria. Indeed, XopDXccB100 specifically interacts with AtMYB30. This protein association, which appears to be independent of nuclear foci formation, leads to inhibition of the transcriptional activation of AtMYB30 target genes and suppression of plant defense during Xanthomonas infection (Canonne et al., 2011; Figure 4A). The HLH domain of XopDXccB100 is necessary and sufficient to mediate interaction with AtMYB30 and repression of AtMYB30 transcriptional activation and plant resistance responses.
Considering XopD modular structure and varied biochemical activities, it is likely that XopD mediates multiple protein–DNA and protein–protein interactions to modulate host transcription. In addition, XopD-dependent bacterial strategies used to subvert plant resistance may vary depending on the Xanthomonas/host plant interaction. XopD SUMO protease, EAR transcription repressor, and DNA-binding activities are at least partially involved in promoting virulence in tomato during Xcv infection (Kim et al., 2008). As previously mentioned, the HLH domain of XopD is necessary and sufficient to suppress AtMYB30-mediated defense during Arabidopsis infection by Xcc, while the EAR motifs and Cys protease domains are not involved in this process. It is thus tempting to speculate that the EAR and the Cys protease domains in XopD are likely involved in targeting host defense-related components other than MYB30. Together, these data suggest that different XopD host targets are involved in the outcome of the interaction between Xcc and Arabidopsis. Future work should uncover additional XopD-related molecular interactions during Xanthomonas infection.
PopP2 from Ralstonia solanacearum
The YopJ/AvrRxv family of T3Es is present in both mammalian and plant pathogens, indicating that they play important roles in the interaction with the host (Orth, 2002; Roden et al., 2004). YopJ/AvrRxv-like effectors belong to the C55 peptidase family of the clan CE of cysteine proteases, which share a nucleophile cysteine and a predicted catalytic core composed of three conserved amino acid residues (H,D/E,C; Hotson and Mudgett, 2004; Rawlings et al., 2006). YopJ/AvrRxv family members differ in their subcellular localization, indicating that their host targets may be diverse. For example in Xcv, AvrRxv is a cytoplasmic protein (Bonshtien et al., 2005), XopJ is targeted to the plasma membrane (Thieme et al., 2007) and AvrBsT localizes to both cytoplasm and nucleus, although the biological significance of AvrBsT nuclear localization remains to be elucidated (Szczesny et al., 2010).
PopP2, a member of the YopJ/AvrRxv T3E family, is injected in plant cells by R. solanacearum, the causal agent of bacterial wilt in more than 200 plant species (Hayward, 1991). The RRS1-R resistance gene from Arabidopsis plants of the Nd-1 ecotype confers broad-spectrum resistance to several strains of R. solanacearum. RRS1-R encodes an R protein with original structure since it belongs to the Toll/interleukin1 receptor (TIR)-NBS-LRR subclass of R proteins and presents a C-terminal WRKY motif that is characteristic of the WKRY class of zinc-finger plant TFs (Deslandes et al., 2003; Figure 4B). RRS1-R presents a putative bipartite NLS and is localized in the nucleus. Intriguingly, PopP2, which presents an NLS that is required for its nuclear targeting (Deslandes et al., 2003), promotes nuclear accumulation of RRS1-R, possibly by preventing its proteasomal degradation (Tasset et al., 2010). PopP2 and RRS1-R physically interact in the nucleus of Arabidopsis cells but whether and how this protein interaction affects host transcription remains to be determined. The Arabidopsis cysteine protease RESPONSIVE TO DEHYDRATATION19 (RD19) was identified in a yeast two-hybrid screen as a PopP2-interacting protein (Figure 4B). In plant cells, PopP2 induces nuclear targeting of RD19, which is otherwise localized to mobile vacuole-associated vesicles and destined to the lytic vacuole (Bernoux et al., 2008). RD19 does not contain a predicted NLS and the molecular mechanism allowing its nuclear recruitment is still unknown. It has been proposed that a PopP2-induced membrane permeabilization process would lead to the release of RD19 from mobile vacuole-associated compartments into the cytoplasm, where it would become available for SUMOylation (Bernoux et al., 2008). Indeed, lysine residues with high probability of being SUMOylated are present in RD19. Therefore, it has been suggested that RD19 SUMOylation may generate the signal required for its nuclear translocation. Alternatively, PopP2 may intercept RD19 on its way to the vacuole through retrograde transport from the endomembrane system, which has some continuity with the nuclear envelope. RD19 interacts with PopP2, but not RRS1-R, in the plant cell nucleus. Since RD19, whose expression is induced by R. solanacearum infection, is required for Arabidopsis resistance to Ralstonia, it was proposed that RD19 associates with PopP2 to form a nuclear complex that is required for activation of the plant resistance response (Figure 4B). PopP2 interaction with RRS1-R may lead to regulation of defense-related gene expression either directly via RRS1-R WKRY domain or through the action of additional plant TFs (Deslandes et al., 2003; Tasset et al., 2010). In this context, it has been suggested that, once in the nucleus, RD19 may function as a transcriptional activator and/or compete with RRS1-R for similar or overlapping cis-elements in the promoters of defense-related genes (Bernoux et al., 2008).
The conserved cysteine residue in PopP2 catalytic triad is required to trigger RRS1-R-mediated resistance. Since PopP2 is able to display acetyltransferase activity, in addition to the proposed indirect modulation of transcription by PopP2 via its interaction with RRS1-R or RD19, it has been suggested that PopP2 may directly manipulate host transcription (Tasset et al., 2010; Figure 4B). Indeed, acetylation of lysine residues of histone tails facilitates access of TFs to DNA by disrupting higher-order packaging of the chromatin (Kornberg and Lorch, 1999). Addition of acetyl groups may also neutralize the positive charge of histones, thereby reducing their affinity for DNA (Hong et al., 1993). In addition, acetylation forms docking sites for recruitment of transcriptional co-activators and impairs the ability of the lysine side chain to form hydrogen bonds thereby enhancing specific or inhibiting non-specific DNA-binding activities of TFs (Mujtaba et al., 2004; Friedler et al., 2005). It has been therefore proposed that PopP2 autoacetylation and/or acetylation of its host targets may affect gene transcription in host cells (Tasset et al., 2010). Identification of host targets of PopP2 should provide insight into the molecular mechanisms developed by R. solanacearum to suppress plant resistance.
Phytoplasma
Phytoplasma are specialized bacteria that are intracellular obligate parasites of plant phloem tissue and their transmitting insect vectors. Phytoplasmas secrete effectors into cells of plants and insects to target host molecules that modulate plant development and increase phytoplasma fitness (Sugio et al., 2011b). As phytoplasma are located intracellularly for much of their life cycle, it is likely that their effectors are released into the cytoplasm of host cells via Sec-dependent translocation. Fifty-six secreted effector proteins were identified in AY-WB (Aster Yellows phytoplasma strain Witches’ Broom) and four contain NLSs. For one of these four effectors, named SAP11, it was further shown that the NLS sequence is required for SAP11 accumulation in Nicotiana benthamiana cell nuclei (Bai et al., 2009). Phytoplasmas are limited to the phloem sieve cells of their plant hosts, which have no nuclei, suggesting the possibility that SAP11 targets tissues beyond the phloem. Consistent with SAP11 nuclear targeting, SAP11 binds and destabilizes a class I and a subset of class II TCP TFs to manipulate plant development and jasmonic acid biosynthesis (Sugio et al., 2011a). As a result, fitness of the leafhopper insect vector is improved, which ensures efficient transmission of the phytoplasma to other plants (Sugio et al., 2011a).
Viruses
Several viral encoded proteins have been shown to translocate into the nuclei during infection. For example, nuclear targeting in infected cells has been observed for various proteins of the rhabdoviruses Sonchus yellow net virus (SYNV) and Maize fine streak virus (MFSV; Goodin et al., 2002; Tsai et al., 2005). An additional example of a viral protein addressed to nuclei is the 2b protein, a virulence determinant encoded by Cucumber mosaic virus (CMV) that suppresses initiation of post-transcriptional gene silencing (PTGS) in transgenic N. benthamiana. CMV 2b protein contains an NLS that is essential for translocation of 2b to the nuclei of tobacco cells. Furthermore, nuclear targeting of 2b protein is required for efficient suppression of PTGS (Lucy et al., 2000).
The NIa and NIb proteins of Tobacco etch potyvirus (TEV) contain two NLSs and have been shown to be translocated to the nucleus of infected cells (Restrepo et al., 1990; Carrington et al., 1991; Li et al., 1997). The NIa proteinase is required for proteolytic maturation of most TEV proteins (Carrington et al., 1988) whereas NIb functions as the RNA-dependent RNA polymerase (Allison et al., 1986). Mutations in the NLSs of NIb disrupt its nuclear translocation and RNA amplification of TEV (Li and Carrington, 1995; Li et al., 1997). It has been additionally demonstrated that NIa interacts with NIb and that this protein interaction is important during TEV genome replication (Li et al., 1997).
The protein p25 encoded by Beet necrotic yellow vein virus (BNYVV) is an important determinant of leaf symptom development and also governs BNYVV invasion of plant roots and induction of rootlet proliferation in sugar beet (Jupin et al., 1992; Tamada et al., 1999). p25 actively shuttles between the cytoplasm and the nucleus of infected cells (Haeberle and Stussi-Garaud, 1995; Vetter et al., 2004). An N-terminal monopartite NLS in p25 is responsible for nuclear accumulation of the protein via its interaction with importin-α, whereas a NES sequence in the C-terminus of p25 mediates its active nuclear export in a CRM1/exportin1-dependent manner. Importantly, modification of the wild-type distribution of p25 between the nuclear and cytoplasmic compartments is accompanied by alterations in p25-related symptoms during virus infection (Vetter et al., 2004). These data highlight the importance of nucleocytoplasmic protein trafficking on the production of necrotic symptoms during BNYVV infection.
Similarly, the regions involved in nucleocytoplasmic shuttling of the ORF3 protein of Groundnut rosette virus (GRV) have been mapped (Ryabov et al., 2004). ORF3 is required for viral RNA protection and movement through the phloem. In infected cells, ORF3 is localized in cytoplasmic granules and also in nuclei, preferentially targeting nucleoli, indicating that this protein can be transported between cytoplasm and nucleus during the course of virus infection (Ryabov et al., 1998, 2004). An arginine-rich NLS in ORF3 is responsible for its nuclear targeting whereas mutations in a leucine-rich NES disrupt nuclear import. The importance of nuclear import and export of GRV ORF3 protein is underlined by the high sequence conservation of the NLS and NES regions among different viruses (Ryabov et al., 2004).
The Cauliflower mosaic virus (CaMV) open reading frame VI product (P6) is a multifunctional protein essential for the viral infection cycle. P6 is a translational reinitiation factor that associates with the host translational machinery and thus permits translation of downstream ORFs (Park et al., 2001). The P6 protein is the major determinant of host specificity in CaMV and determines symptom severity (Daubert and Routh, 1990). P6 is an abundantly synthesized CaMV protein that in the cytoplasm of infected cells forms electron-dense inclusion bodies, also referred to as viroplasms, where virus replication and assembly occur (Mazzolini et al., 1989). Viroplasms have been additionally detected at the periphery of the nucleus and it has been shown that P6 is able to enter the nucleus during viral infection, suggesting that P6 is a nucleocytoplasmic shuttling protein (Haas et al., 2005). Nuclear localization of P6 is consistent with the finding that P6 interacts with nuclear-localized proteins (Park et al., 2001; Bureau et al., 2004). The N-terminal region of P6 presents an amphipatic α-helix containing a leucine zipper motif that is predicted to form a parallel coiled-coil structure. P6 N-terminal region is well conserved among CaMV strains and mediates the interaction between P6 molecules in vitro, making of this region an essential determinant for the formation of viroplasms (Haas et al., 2005). An NES has been identified in P6 Leu-rich sequence that bears some resemblance to NES sequences found in the BR1 protein of the geminivirus Squash leaf curl virus (Ward and Lazarowitz, 1999) and in several shuttling nuclear proteins, such as HIV Rev protein (Pollard and Malim, 1998). The NES in P6 determines its CRM1-dependent nuclear export and thus P6 localization in the cytoplasm (Haas et al., 2005). It has been suggested that P6 nuclear export probably occurs very rapidly in infected cells, so that only low amounts are present in the nucleus at any time. Therefore, activity of CRM1-dependent export pathway would limit the extent of P6 nuclear accumulation in the nucleus that could be deleterious for the CaMV infectious cycle. Different hypotheses regarding the role of P6 nuclear accumulation have been formulated. Since P6 is able to bind single- and double-stranded RNA (De Tapia et al., 1993; Cerritelli et al., 1998), it has been proposed that, similar to the Rev protein of HIV-1 (Pollard and Malim, 1998), P6 may control export of CaMV 35S RNA and its spliced versions. Additionally, P6 has been detected in the nucleolus where assembly of ribosomal subunits occurs, suggesting that P6 may interact directly with ribosomes before their export to render them competent for translation of the CaMV polycistronic mRNA. In this context, the P6-interacting ribosomal proteins L18 and L24 (Leh et al., 2000; Park et al., 2001) could be targets for P6 since they participate in the formation of the 60S ribosomal subunit in the nucleolus (Andersen et al., 2002). Finally, as a nucleocytoplasmic protein, P6 may play a role in inhibition of nonsense-mediated mRNA decay to prevent degradation of the 35S RNA and its spliced versions (Maquat and Carmichael, 2001). Indeed, P6 nuclear export is mediated by the CRM1 pathway (Kudo et al., 1998), which is known to be specifically used for export of the ribosomal subunits and of some cellular mRNAs (Weis, 2002).
The TIR-NBS-LRR immune receptor N is localized to the cytoplasm and the nucleus of uninfected tobacco cells. During Tobacco mosaic virus (TMV) infection, the viral replicase p50 is delivered into the plant cell cytoplasm, where it is recognized by the N protein, leading to the establishment of plant resistance (Burch-Smith et al., 2007). In TMV-infected cells, cytoplasmic p50 induces recruitment to the cytoplasm of NRIP1, a tobacco rhodanase sulfurtransferase that otherwise localizes to the stroma of chloroplasts. NRIP1 is able to interact with N but only in the presence of p50 and NRIP1 rhodanase sulfurtransferase activity is not necessary of its association with N or p50. In this context, a cytoplasmic pre-recognition complex is formed that contains NRIP1, p50, and possible additional host proteins (Caplan et al., 2008). Interaction of N with this pre-recognition complex would lead to its activation and, once activated, N would be either translocated into the nucleus or able to send a signal that activates the N nuclear pool, resulting in a successful defense response. Shuttling of p50-activated N from the cytoplasm to the nucleus appears to be required for an efficient defense response (Burch-Smith et al., 2007). Similarly, NRIP1 nuclear relocalization is necessary to provide full resistance to TMV infection (Caplan et al., 2008). Although the molecular mechanism behind p50-mediated NRIP1 nuclear relocalization remains to be determined, different hypothesis have been proposed to explain this finding. p50 might disrupt global chloroplast import by an unknown mechanism that would affect translocation of NRIP1. Otherwise, interaction with p50 might mask the chloroplast targeting signal in NRIP1 facilitating its nuclear import. Alternatively, NRIP1 may be released from chloroplasts into the cytoplasm and the nucleus following p50-induced permeabilization of the outer membrane. Finally, the close physical association between stromules and nuclei might enhance the nuclear import of chloroplastic factors, including NRIP1.
Filamentous Pathogens
Plant pathogenic fungi and oomycetes, collectively referred to as filamentous pathogens, are responsible for a variety of diseases in natural populations and agricultural crops. As sophisticated manipulators of plant cell functions, filamentous pathogens are intimately associated with host plants cells. As previously mentioned, haustoria are highly specialized structures that develop within plant cells and appear to be involved in nutrient acquisition (Hahn and Mendgen, 2001). In addition, haustoria contain specific membrane proteins required for pathogenicity (Avrova et al., 2008), are highly enriched in secreted effector proteins and most probably play a role in mediating effector translocation into host cells, although this hypothesis remains to be formally demonstrated (Panstruga and Dodds, 2009). Following secretion from the pathogen, effectors that carry host-translocation signals are transported into the plant cell. However, in order to reach the host cytoplasm, effectors need to travel across two membranes, one pathogen-derived and one host cell-derived membrane surrounding the haustorium, and the nature of the mechanism directing effector delivery is still unclear.
Catalogs of the complete set of secreted proteins for a number of filamentous pathogens have been recently generated thanks to extensive genome sequencing programs coupled with robust computational predictions of secretion signals and other sequence motifs characteristic of effectors (Dean et al., 2005; Kamper et al., 2006; Haas et al., 2009; Duplessis et al., 2011). From these studies, it is clear that oomycetes and phytopathogenic fungi secrete a suite of effector molecules that is considerably larger than the effector repertoires of phytopathogenic bacteria. These candidate effector proteins are predicted to modulate host innate immunity and enable parasitic infection (Kamoun, 2007; Hogenhout et al., 2009).
Oomycetes
Oomycota or oomycetes form a distinct phylogenetic lineage of fungus-like eukaryotic microorganisms. Species of the oomycete genus Phytophthora are devastating pathogens of dicotyledoneous plants. Sequencing and analysis of oomycete genomes has revealed that the predicted proteomes include a large repertoire of candidate host-translocated effector proteins that have been classified in two main classes, named RXLR and CRN (for crinkling and necrosis; Haas et al., 2009). Both RXLR and CRN effector proteins present a modular architecture and include a signal peptide, conserved N termini functioning in host delivery and highly diverse C-terminal domains directing the effector activity (Haas et al., 2009; Schornack et al., 2010). Indeed, RXLR effectors are defined by a conserved N-terminal RXLR motif, flanked by a high frequency of acidic (D/E) residues, that enables delivery of effector proteins inside plant cells (Whisson et al., 2007; Dou et al., 2008). It has been proposed that RxLR motifs enable effectors to bind to host cell surface phosphatidylinositol-3-phosphate (PI3P) and subsequently enter host cells through lipid raft-mediated endocytosis (Kale et al., 2010). However, this mechanism of effector translocation and the experiments that support it are still under debate (de Jonge et al., 2011; Stassen and Van den Ackerveken, 2011). Similar to RXLR effectors, N termini of CRN proteins present a conserved but not invariant LXLFLAK motif, which is required for effector targeting and translocation (Schornack et al., 2010). Aphanomyces euteiches CRN-like sequences carry a conserved N-terminal LQLYLALK motif similar to the Phytophthora LXLFLAK sequence (Gaulin et al., 2008). It has been demonstrated that the LQLYLALK motif in A. euteiches CRNs is able to mediate effector translocation (Schornack et al., 2010).
Five hundred sixty-three RXLR effectors are predicted for the strain T30-4 of Phytophthora infestans (Haas et al., 2009), whereas they appear to be absent in Pythium ultimum (Levesque et al., 2010) and A. euteiches (Gaulin et al., 2008), suggesting that these effectors have evolved only recently within the Peronosporales, coinciding with the appearance of haustoria (Levesque et al., 2010). In contrast, the CRN family is ubiquitous in plant pathogenic oomycetes (Gaulin et al., 2008; Levesque et al., 2010), suggesting that the CRNs belong to an ancient effector family that arose early in oomycete evolution before the emergence of haustoria. Recent evidence however indicates that a significant part of the oomycete secretome was probably acquired from fungi through horizontal gene transfer events, and this may have facilitated the spread of oomycetes to plant hosts (Richards et al., 2011). Conservation of the RXLR motif, which has not been described in fungal effectors, has enabled the computational development of genome-wide catalogs of candidate RXLR effectors from several oomycete pathogens (Tyler et al., 2006; Win et al., 2007; Jiang et al., 2008; Haas et al., 2009). In addition, putative RXLR effector genes have been used in high-throughput screens to predict novel functional activities (Vleeshouwers et al., 2008; Oh et al., 2009). Similarly, LXLFLAK-type motifs may be used in genome-wide searches to identify oomycete effector proteins.
Subcellular localization studies of diverse CRN C termini unrelated in sequence and derived from two divergent species (CRN2, CRNR8, CRN15, and CRN16 from P. infestans and AeCRN5 from A. euteiches) revealed that CRNs accumulate in plant cell nuclei (Schornack et al., 2010). NLS motifs were predicted in CRN8, CRN16, and AeCRN5, suggesting that CRN proteins recruit the host nuclear import machinery to achieve nuclear accumulation. Indeed, CRN2, CRNR8, CRN16, and AeCRN5 nuclear accumulation depends on the host nuclear import factor importin-α, and at least in the case of CRN8, on a functional NLS. Moreover, cell death induced by CRN8 requires its accumulation in host nuclei, suggesting that CRN proteins are targeted to the host nucleus during plant infection (Schornack et al., 2010). In P. sojae, CRN proteins PsCRN63 and PsCRN115, which are involved in virulence on soybean and suppression of host defense responses, contain an NLS motif (Liu et al., 2011). Despite their high sequence similarity (95.7% identity at the amino acid level), PsCRN63 triggers cell death in N. benthamiana whereas PsCRN115 is able to suppress cell death induced by the P. sojae necrosis-inducing protein PsojNIP (Qutob et al., 2002) and PsCRN63. A functional NLS is required for PsCRN63 induction of cell death and exclusion of PsCRN63 from the nucleus using a NES prevented cell death development, suggesting that PsCRN63 triggers cell death in the plant cell nucleus (Liu et al., 2011). In contrast, the NLS in PsCRN115 is not required for cell death suppression. Although the subcellular localization of PsCRN63 and PsCRN115 was not investigated, it was therefore proposed that PsCRN63 and PsCRN115 may share the same molecular host targets involved in cell death signaling and that their distinct activities are dependent or their nuclear localization (Liu et al., 2011).
Supressor of necrosis1 (SNE1) and Avr3a are nuclear-targeted effectors from P. infestans that suppress plant cell death responses. SNE1 is a highly hydrophilic secreted protein that presents NLSs and is able to translocate to the cell nucleus and suppress the action of secreted cell death-inducing effectors that are expressed during the necrotrophic growth phase of P. infestans, as well as programmed cell death mediated by a range of Avr–R protein interactions (Kelley et al., 2010). Avr3a has been shown to interact with and stabilize the U-box E3 ligase CMPG1 that is required for cell death trigerred by P. infestans elicitin INF1 (ICD) and is degraded by the 26S proteasome (Bos et al., 2010). This protein interaction, which leads to suppression of ICD, occurs in the nucleus and, more particularly, in the nucleolus of host cells (Gilroy et al., 2011). It has been therefore proposed that the nucleus is likely a major site of CMPG1 activity and 26S proteasome-mediated degradation. In an additional study, the authors searched for P. infestans proteins that contain a signal peptide and an NLS and identified four proteins called, Nuk6, Nuk7, Nuk10, and Nuk12. All four proteins were found in the nucleus of N. benthamiana cells in Agrobacterium-mediated transient expression assays. Nuclear localization of Nuk6, Nuk7, and Nuk10 is dependent on NbIMPα1 and NbIMPα2, whereas Nuk12 can target nuclei independently of these host α-importins (Kanneganti et al., 2007; Vleeshouwers et al., 2011). Interestingly, a mutation in the MOS6 gene, encoding AtImpα3 (one of the nine Arabidopsis α-importins), enhanced susceptibility to an additional oomycete plant pathogen Hyaloperonospora parasitica (Hpa; Palma et al., 2005). This finding confirms the importance of protein nucleocytoplasmic trafficking during defense responses against oomycete pathogens.
Finally, sequencing of the Hpa genome predicted 134 high-confidence effector gene candidates of the RxLR type (HaRxLs), which is significantly fewer than in Phytophthora genomes (Baxter et al., 2010). Notably, a recent study showed association of the plant cell nucleus with developing haustoria in Arabidopsis mesophyll cells, possibly by moving through the actin cytoskeleton (Caillaud et al., 2011). In this context, it is tempting to speculate that haustoria directly influence the position of the nucleus in the cell in order to enhance delivery of effectors that compromise defense-related nuclear processes. Indeed, 66% of 49 tested HaRxLs target the nucleus, either as strictly nuclear or nuclear–cytoplasmic. However, only 37.5% of the 16 strictly nuclear HaRxLs carried a canonical NLS (Caillaud et al., 2011), suggesting that endogenous host proteins may facilitate nuclear delivery of these effectors. Interestingly, 21% of the HaRxLs tested localized to the nucleolus, indicating that Hpa hijacks the plant cell transcriptional machinery perhaps by acting on RNA biogenesis, transport, or splicing, on ribosome biogenesis and thereby on protein translation to prevent de novo induction of defense responses.
Fungi
Similarly to oomycetes, pathogenic fungi have elaborated intricate parasitic relationships to feed from their host plants and the effector-dependent transcriptional reprogramming of host cells is much plausible. The production of apoplastic effectors by filamentous fungi is well documented (de Jonge et al., 2011; Stassen and Van den Ackerveken, 2011). Similar to what has been described for oomycete effectors, the N termini of various fungal effectors have been reported to carry degenerate RxLR motifs that bind to PI3P and mediate effector translocation, although this may not be a universal means of effector uptake (Kale et al., 2010). Indeed, powdery mildew and rust fungi encode small secreted proteins that share an N-terminal Y/F/WxC motif that is situated downstream of signal peptide and not found in effectors from non-haustorial fungi or oomycetes. It has been suggested that this motif mediates translocation of fungal haustorial effectors into plant cells (Godfrey et al., 2010).
The Uf-RTP1 protein from the rust fungus Uromyces fabae is the only effector from a pathogenic fungus for which a nuclear localization has been described to date. Indeed, Uf-RTP1 is a haustorial secreted effector that presents a signal peptide and a bipartite NLS that mediates Uf-RTP1 accumulation in plant cell nuclei (Kemen et al., 2005). Homologs of Uf-RTP1 have been found in other pathogenic rust fungi such as Puccinia graminis, Melampsora spp., Uromyces appendiculatus, or Hemileia vastatrix (Puthoff et al., 2008; Duplessis et al., 2011). However, the absence of obvious structural features within their protein sequences renders prediction of their putative function(s) a difficult task. Mining of effectors in the genome of Ustilago maydis identified among the secreted effector candidates 14 proteins that contain a putative NLS (Mueller et al., 2008), suggesting that virulence of this pathogen may also rely on nuclear targeting of effector proteins.
Nematodes
Research aimed at the identification and characterization of pathogenicity effectors of plant nematodes has been intensively developed since genome sequences of these pathogenic organisms were first reported. Most nematode species are restricted to the roots in which they form specialized feeding sites to become sedentary endoparasites (Gheysen and Mitchum, 2011). Two groups of pathogens have been particularly studied: the root knot nematodes (Meloidogyne spp.) and the cyst nematodes (Heterodera spp. and Globodera spp.).
First suspicions about the existence of nuclear-localized proteins involved in nematode parasitism of plants came from analysis of first lists of genes encoding secretory proteins and identified as candidate effectors. These proteins are expressed and produced in pharyngeal glands and subsequently delivered into plant cells by the stylet, a protrusible hollow mouth spear (Bellafiore and Briggs, 2010). In Heterodera glycines, the identification of specific secretory gland-expressed genes revealed 51 candidate effectors, 15 of which displayed canonical NLSs (Gao et al., 2003), whereas inspection of the Meloidogyne incognita secretome yielded a total of 66 effectors putatively addressed to the plant nucleus (Bellafiore et al., 2008). These 66 candidates included 26 proteins with an NLS motif and 40 additional proteins with putative nucleotide binding activity such as DNA or chromatin interaction motifs. Similarly, three esophageal gland-specific gene products from Meloidogyne chitwoodi are predicted to be nuclear localized in host cells following cleavage of the leader peptide for protein secretion (Roze et al., 2008). These lists of potential NLS-effectors include many proteins of unknown function (Gao et al., 2003; Roze et al., 2008) but also helicases, histones, DNA-binding domain proteins, and the Nucleosome Assembly Protein, NAP-1 (Bellafiore et al., 2008).
Functional characterization of nematode effectors provided direct evidence of their localization to the plant nucleus. A GFP fusion to the Heterodera schachtii secreted protein Hs-UBI1, which encodes a protein with a mono-ubiquitin domain, was shown to be targeted to the nucleolus of tobacco cells (Tytgat et al., 2004). Interestingly, other nematode effectors with some similarity to components of E3-ubiquitin ligase complexes, such as a SKP1-homolog protein, have been identified in other species (Gao et al., 2003; Bellafiore et al., 2008) but their role during parasitic infection remains elusive.
A functional analysis of identified NLS-containing effectors in H. glycines demonstrated that two out of the eight tested effector proteins were imported into the nuclei of both onion epidermal cells and Arabidopsis protoplasts (Elling et al., 2007). One of these protein fusions was further transported into the nucleolus. Mutation analysis of the NLS domains confirmed their role in effector nuclear uptake. The authors also showed that GFP fusions with three additional effectors also accumulated in the plant nucleus, but only with effector truncated versions (containing the NLS domain) and not with the full-length proteins (Elling et al., 2007). Whether this differential behavior is due to artifactual mislocalization or reflects the possible existence of an effector processing mechanism within the plant cell remains to be determined.
Another report also showed that some effectors encoded by genes belonging to the large SPRYSEC family in Globodera pallida are addressed to the host cell nucleus. These stylet-secreted effectors harbor a SPRY domain of 120 amino acids and unknown function that was first identified in some receptors from Dictyostelium. Two G. pallida SPRYSEC family proteins harbor NLSs and present nucleolar localization in both tobacco leaf and root cells whereas some other SPRYSEC proteins are cytoplasmic (Jones et al., 2009). It was hypothesized that SPRYSEC effectors suppress host defenses through interaction with a range of host targets in different cell compartments (Jones et al., 2009). Interestingly, similar to viral and bacterial effectors, two SPRYSEC proteins were also shown to interact with resistance proteins, including the potato R protein Gpa2 (Rehman et al., 2009; Sacco et al., 2009).
Since some stylet-secreted proteins have features of DNA-binding proteins (Bellafiore et al., 2008), it is tempting to speculate that some nuclear-localized nematode effectors might act in reprogramming host transcription. Although expression of some TFs involved in plant development have been shown to be induced in nematode feeding sites (Grunewald et al., 2008; Barcala et al., 2010), there is however no evidence supporting a direct role of nematode effectors in modifying plant gene expression. It has been reported that a 13-amino acid peptide, 16D10, secreted from the esophageal glands of M. incognita, stimulates root growth and the generation of extensive lateral roots in tobacco hairy roots. 16D10 has been shown to interact in planta with two putative plant SCARECROW-like TFs (Huang et al., 2006). However, 16D10 has not been visualized in the plant cell nucleus and its role in plant cell developmental processes, as well as the role of its interacting TFs, remains to be determined.
Concluding Remarks
Pathogenic microorganisms use an extremely diverse panoply of effector proteins to counteract plant defense and ensure successful colonization of their hosts. Although different subcellular compartments are targeted by effector proteins following their translocation into plant cells, it is now apparent that a significant number of these effector molecules are specifically addressed to the host plant cell nucleus. Nuclear targeting of effectors appears to be a general microbial strategy since examples have been documented in each main class of pathogenic organisms from viruses to nematodes, although no nuclear effectors from bacterial Gram-positive plant pathogens have been described to date.
As mentioned above, the search for NLSs in the available repertoires of predicted effector proteins from different microbes may help identify nuclear translocated effectors. However, it is worth mentioning that up to 45% of yeast proteins present a predicted bipartite or monopartite NLS (and therefore the potential to enter the nucleus via the classical nuclear import pathway), although only 25.8% of proteins localize to the nucleus at steady state when chromosomally tagged with GFP (Huh et al., 2003). Moreover, only 25.8 and 30.9% of the yeast proteins that have been localized to the nucleus in the global GFP screen contain a putative bipartite or monopartite NLS, respectively (Lange et al., 2007). Therefore, about 43% of steady-state nuclear proteins in yeast may not use the classical nuclear import pathway to enter the nucleus. Similarly, in silico prediction of NLSs is not systematically associated to nuclear localization of effector proteins (Caillaud et al., 2011) and, in some cases, NLSs are dispensable for effector nuclear targeting (Canonne et al., 2011). Therefore, caution should be taken when interpreting data from in silico effector analysis and functional studies of predicted effector proteins are absolutely required to validate their subcellular localization.
Interestingly, nuclear translocation of effector proteins does not seem to be restricted to pathogenic microorganisms since targeting of effector proteins to the plant cell nucleus has been also recently discovered in the context of plant–microbe symbiotic interactions. Indeed, the secreted effector MiSSP7 from the ectomycorrhizal fungus Laccaria bicolor is detected in host cell nuclei where it induces reprogramming of the plant cell transcriptome to favor mutualism (Plett et al., 2011). Another report established that the arbuscular mycorrhizal fungus Glomus intraradices secretes a protein, SP7, which interacts with the pathogenesis-related TF ERF19 in the plant nucleus to promote symbiotic biotrophy (Kloppholz et al., 2011). These findings highlight the importance of the host nucleus in determining the fate of parasitic or mutualist interactions and the almost ubiquitous microbial strategy that involves direct interactions with host DNA and/or nuclear proteins. As illustrated above for pathogens, outputs resulting from these molecular interactions in the nucleus encompass various pathogenicity processes and multiple stages of host infection, such as the suppression of plant defense responses, the metabolic priming of host cells, or pathogen dispersal.
It is clear that, despite significant recent advances in our field describing nuclear targeting of effector proteins, a lot remains to be done to obtain a global view on how transcriptional reprogramming of host cells or specific targeting of nuclear proteins promote pathogen infection and plant disease. Oligonucleotide microarrays or mRNA sequencing approaches should shed light on the large-scale transcriptomic changes undergone by host cells during nuclear targeting of effector proteins. Finally, a whole cellular dynamics outlook, integrating the contribution to virulence of nuclear effectors and that of effectors targeted to additional subcellular compartments, represents an exciting perspective for future research.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Our work is supported by funds from French ANR (ANR JC08_324792) and is part of the Laboratoire d’Excellence (LABEX) entitled TULIP (ANR-10-LABX-41). We thank Anne-Claire Cazalé, Lionel Navarro, Laurent Noël, and Boris Szurek for communicating unpublished results and apologize to all colleagues whose work could not be discussed because of space limitations.
References
Alfano, J. R., and Collmer, A. (2004). Type III secretion system effector proteins: double agents in bacterial disease and plant defense. Annu. Rev. Phytopathol. 42, 385–414.
Allison, R., Johnston, R. E., and Dougherty, W. G. (1986). The nucleotide sequence of the coding region of tobacco etch virus genomic RNA: evidence for the synthesis of a single polyprotein. Virology 154, 9–20.
Anand, A., Krichevsky, A., Schornack, S., Lahaye, T., Tzfira, T., Tang, Y., Citovsky, V., and Mysore, K. S. (2007). Arabidopsis VIRE2 INTERACTING PROTEIN2 is required for Agrobacterium T-DNA integration in plants. Plant Cell 19, 1695–1708.
Andersen, J. S., Lyon, C. E., Fox, A. H., Leung, A. K., Lam, Y. W., Steen, H., Mann, M., and Lamond, A. I. (2002). Directed proteomic analysis of the human nucleolus. Curr. Biol. 12, 1–11.
Arbibe, L., Kim, D. W., Batsche, E., Pedron, T., Mateescu, B., Muchardt, C., Parsot, C., and Sansonetti, P. J. (2007). An injected bacterial effector targets chromatin access for transcription factor NF-kappaB to alter transcription of host genes involved in immune responses. Nat. Immunol. 8, 47–56.
Asai, T., Tena, G., Plotnikova, J., Willmann, M. R., Chiu, W. L., Gomez-Gomez, L., Boller, T., Ausubel, F. M., and Sheen, J. (2002). MAP kinase signalling cascade in Arabidopsis innate immunity. Nature 415, 977–983.
Avrova, A. O., Boevink, P. C., Young, V., Grenville-Briggs, L. J., van West, P., Birch, P. R., and Whisson, S. C. (2008). A novel Phytophthora infestans haustorium-specific membrane protein is required for infection of potato. Cell. Microbiol. 10, 2271–2284.
Bai, X., Correa, V. R., Toruno, T. Y., Ammar el, D., Kamoun, S., and Hogenhout, S. A. (2009). AY-WB phytoplasma secretes a protein that targets plant cell nuclei. Mol. Plant Microbe Interact. 22, 18–30.
Bako, L., Umeda, M., Tiburcio, A. F., Schell, J., and Koncz, C. (2003). The VirD2 pilot protein of Agrobacterium-transferred DNA interacts with the TATA box-binding protein and a nuclear protein kinase in plants. Proc. Natl. Acad. Sci. U.S.A. 100, 10108–10113.
Ballas, N., and Citovsky, V. (1997). Nuclear localization signal binding protein from Arabidopsis mediates nuclear import of Agrobacterium VirD2 protein. Proc. Natl. Acad. Sci. U.S.A. 94, 10723–10728.
Barcala, M., Garcia, A., Cabrera, J., Casson, S., Lindsey, K., Favery, B., Garcia-Casado, G., Solano, R., Fenoll, C., and Escobar, C. (2010). Early transcriptomic events in microdissected Arabidopsis nematode-induced giant cells. Plant J. 61, 698–712.
Baxter, L., Tripathy, S., Ishaque, N., Boot, N., Cabral, A., Kemen, E., Thines, M., Ah-Fong, A., Anderson, R., Badejoko, W., Bittner-Eddy, P., Boore, J. L., Chibucos, M. C., Coates, M., Dehal, P., Delehaunty, K., Dong, S., Downton, P., Dumas, B., Fabro, G., Fronick, C., Fuerstenberg, S. I., Fulton, L., Gaulin, E., Govers, F., Hughes, L., Humphray, S., Jiang, R. H., Judelson, H., Kamoun, S., Kyung, K., Meijer, H., Minx, P., Morris, P., Nelson, J., Phuntumart, V., Qutob, D., Rehmany, A., Rougon-Cardoso, A., Ryden, P., Torto-Alalibo, T., Studholme, D., Wang, Y., Win, J., Wood, J., Clifton, S. W., Rogers, J., Van den Ackerveken, G., Jones, J. D., McDowell, J. M., Beynon, J., and Tyler, B. M. (2010). Signatures of adaptation to obligate biotrophy in the Hyaloperonospora arabidopsidis genome. Science 330, 1549–1551.
Bellafiore, S., and Briggs, S. P. (2010). Nematode effectors and plant responses to infection. Curr. Opin. Plant Biol. 13, 442–448.
Bellafiore, S., Shen, Z., Rosso, M. N., Abad, P., Shih, P., and Briggs, S. P. (2008). Direct identification of the Meloidogyne incognita secretome reveals proteins with host cell reprogramming potential. PLoS Pathog. 4, e1000192. doi:10.1371/journal.ppat.1000192
Bernoux, M., Timmers, T., Jauneau, A., Briere, C., de Wit, P. J., Marco, Y., and Deslandes, L. (2008). RD19, an Arabidopsis cysteine protease required for RRS1-R-mediated resistance, is relocalized to the nucleus by the Ralstonia solanacearum PopP2 effector. Plant Cell 20, 2252–2264.
Block, A., Li, G., Fu, Z. Q., and Alfano, J. R. (2008). Phytopathogen type III effector weaponry and their plant targets. Curr. Opin. Plant Biol. 11, 396–403.
Boch, J., and Bonas, U. (2010). Xanthomonas AvrBs3 family-type III effectors: discovery and function. Annu. Rev. Phytopathol. 48, 419–436.
Boch, J., Scholze, H., Schornack, S., Landgraf, A., Hahn, S., Kay, S., Lahaye, T., Nickstadt, A., and Bonas, U. (2009). Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509–1512.
Bogdanove, A. J., Schornack, S., and Lahaye, T. (2010). TAL effectors: finding plant genes for disease and defense. Curr. Opin. Plant Biol. 13, 394–401.
Bonshtien, A., Lev, A., Gibly, A., Debbie, P., Avni, A., and Sessa, G. (2005). Molecular properties of the Xanthomonas AvrRxv effector and global transcriptional changes determined by its expression in resistant tomato plants. Mol. Plant Microbe Interact. 18, 300–310.
Bos, J. I., Armstrong, M. R., Gilroy, E. M., Boevink, P. C., Hein, I., Taylor, R. M., Zhendong, T., Engelhardt, S., Vetukuri, R. R., Harrower, B., Dixelius, C., Bryan, G., Sadanandom, A., Whisson, S. C., Kamoun, S., and Birch, P. R. (2010). Phytophthora infestans effector AVR3a is essential for virulence and manipulates plant immunity by stabilizing host E3 ligase CMPG1. Proc. Natl. Acad. Sci. U.S.A. 107, 9909–9914.
Burch-Smith, T. M., and Dinesh-Kumar, S. P. (2007). The functions of plant TIR domains. Sci. STKE 2007, pe46.
Burch-Smith, T. M., Schiff, M., Caplan, J. L., Tsao, J., Czymmek, K., and Dinesh-Kumar, S. P. (2007). A novel role for the TIR domain in association with pathogen-derived elicitors. PLoS Biol. 5, e68. doi:10.1371/journal.pbio.0050068
Bureau, M., Leh, V., Haas, M., Geldreich, A., Ryabova, L., Yot, P., and Keller, M. (2004). P6 protein of Cauliflower mosaic virus, a translation reinitiator, interacts with ribosomal protein L13 from Arabidopsis thaliana. J. Gen. Virol. 85, 3765–3775.
Buttner, D., and He, S. Y. (2009). Type III protein secretion in plant pathogenic bacteria. Plant Physiol. 150, 1656–1664.
Caillaud, M. C., Piquerez, S. J., Fabro, G., Steinbrenner, J., Ishaque, N., Beynon, J., and Jones, J. D. (2011). Subcellular localization of the Hpa RxLR effector repertoire identifies a tonoplast-associated protein HaRxL17 that confers enhanced plant susceptibility. Plant J.doi: 10.1111/j.1365-313X.2011.04787.x
Canonne, J., Marino, D., Jauneau, A., Pouzet, C., Briere, C., Roby, D., and Rivas, S. (2011). The Xanthomonas type III effector XopD targets the Arabidopsis transcription factor AtMYB30 to suppress plant defence. Plant Cell 23, 3498–3511.
Canonne, J., Marino, D., Noël, L. D., Arechaga, I., Pichereaux, C., Rossignol, M., Roby, D., and Rivas, S. (2010). Detection and functional characterization of a 215 amino acid N-terminal extension in the Xanthomonas type III effector XopD. PLoS ONE 5, e15773. doi:10.1371/journal.pone.0015773
Canonne, J., Pichereaux, C., Marino, D., Roby, D., Rossignol, M., and Rivas, S. (2012). Identification of the protein sequence of the type III effector XopD from the B100 strain of Xanthomonas campestris pv. campestris. Plant Signal. Behav. (in press).
Caplan, J. L., Mamillapalli, P., Burch-Smith, T. M., Czymmek, K., and Dinesh-Kumar, S. P. (2008). Chloroplastic protein NRIP1 mediates innate immune receptor recognition of a viral effector. Cell 132, 449–462.
Carrington, J. C., Cary, S. M., and Dougherty, W. G. (1988). Mutational analysis of Tobacco etch virus polyprotein processing: cis and trans proteolytic activities of polyproteins containing the 49-kilodalton proteinase. J. Virol. 62, 2313–2320.
Carrington, J. C., Freed, D. D., and Leinicke, A. J. (1991). Bipartite signal sequence mediates nuclear translocation of the plant potyviral NIa protein. Plant Cell 3, 953–962.
Cerritelli, S. M., Fedoroff, O. Y., Reid, B. R., and Crouch, R. J. (1998). A common 40 amino acid motif in eukaryotic RNases H1 and caulimovirus ORF VI proteins binds to duplex RNAs. Nucleic Acids Res. 26, 1834–1840.
Chen, L. Q., Hou, B. H., Lalonde, S., Takanaga, H., Hartung, M. L., Qu, X. Q., Guo, W. J., Kim, J. G., Underwood, W., Chaudhuri, B., Chermak, D., Antony, G., White, F. F., Somerville, S. C., Mudgett, M. B., and Frommer, W. B. (2010). Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 468, 527–532.
Chisholm, S. T., Coaker, G., Day, B., and Staskawicz, B. J. (2006). Host-microbe interactions: shaping the evolution of the plant immune response. Cell 124, 803–814.
Christie, P. J. (2004). Type IV secretion: the Agrobacterium VirB/D4 and related conjugation systems. Biochim. Biophys. Acta 1694, 219–234.
Christie, P. J., Atmakuri, K., Krishnamoorthy, V., Jakubowski, S., and Cascales, E. (2005). Biogenesis, architecture, and function of bacterial type IV secretion systems. Annu. Rev. Microbiol. 59, 451–485.
Chu, Z., Fu, B., Yang, H., Xu, C., Li, Z., Sanchez, A., Park, Y. J., Bennetzen, J. L., Zhang, Q., and Wang, S. (2006). Targeting xa13, a recessive gene for bacterial blight resistance in rice. Theor. Appl. Genet. 112, 455–461.
Citovsky, V., Kapelnikov, A., Oliel, S., Zakai, N., Rojas, M. R., Gilbertson, R. L., Tzfira, T., and Loyter, A. (2004). Protein interactions involved in nuclear import of the Agrobacterium VirE2 protein in vivo and in vitro. J. Biol. Chem. 279, 29528–29533.
Citovsky, V., Kozlovsky, S. V., Lacroix, B., Zaltsman, A., Dafny-Yelin, M., Vyas, S., Tovkach, A., and Tzfira, T. (2007). Biological systems of the host cell involved in Agrobacterium infection. Cell. Microbiol. 9, 9–20.
Clapier, C. R., and Cairns, B. R. (2009). The biology of chromatin remodeling complexes. Annu. Rev. Biochem. 78, 273–304.
Cunnac, S., Lindeberg, M., and Collmer, A. (2009). Pseudomonas syringae type III secretion system effectors: repertoires in search of functions. Curr. Opin. Microbiol. 12, 53–60.
Cunnac, S., Occhialini, A., Barberis, P., Boucher, C., and Genin, S. (2004). Inventory and functional analysis of the large Hrp regulon in Ralstonia solanacearum: identification of novel effector proteins translocated to plant host cells through the type III secretion system. Mol. Microbiol. 53, 115–128.
da Silva, A. C., Ferro, J. A., Reinach, F. C., Farah, C. S., Furlan, L. R., Quaggio, R. B., Monteiro-Vitorello, C. B., Van Sluys, M. A., Almeida, N. F., Alves, L. M., do Amaral, A. M., Bertolini, M. C., Camargo, L. E., Camarotte, G., Cannavan, F., Cardozo, J., Chambergo, F., Ciapina, L. P., Cicarelli, R. M., Coutinho, L. L., Cursino-Santos, J. R., El-Dorry, H., Faria, J. B., Ferreira, A. J., Ferreira, R. C., Ferro, M. I., Formighieri, E. F., Franco, M. C., Greggio, C. C., Gruber, A., Katsuyama, A. M., Kishi, L. T., Leite, R. P., Lemos, E. G., Lemos, M. V., Locali, E. C., Machado, M. A., Madeira, A. M., Martinez-Rossi, N. M., Martins, E. C., Meidanis, J., Menck, C. F., Miyaki, C. Y., Moon, D. H., Moreira, L. M., Novo, M. T., Okura, V. K., Oliveira, M. C., Oliveira, V. R., Pereira, H. A., Rossi, A., Sena, J. A., Silva, C., de Souza, R. F., Spinola, L. A., Takita, M. A., Tamura, R. E., Teixeira, E. C., Tezza, R. I., Trindade dos Santos, M., Truffi, D., Tsai, S. M., White, F. F., Setubal, J. C., and Kitajima, J. P. (2002). Comparison of the genomes of two Xanthomonas pathogens with differing host specificities. Nature 417, 459–463.
Daubert, S., and Routh, G. (1990). Point mutations in Cauliflower mosaic virus gene VI confer host-specific symptom changes. Mol. Plant Microbe Interact. 3, 341–345.
Davis, E. L., Hussey, R. S., Mitchum, M. G., and Baum, T. J. (2008). Parasitism proteins in nematode-plant interactions. Curr. Opin. Plant Biol. 11, 360–366.
de Jonge, R., Bolton, M. D., and Thomma, B. P. (2011). How filamentous pathogens co-opt plants: the ins and outs of fungal effectors. Curr. Opin. Plant Biol. 14, 400–406.
De Tapia, M., Himmelbach, A., and Hohn, T. (1993). Molecular dissection of the Cauliflower mosaic virus translation transactivator. EMBO J. 12, 3305–3314.
Dean, R. A., Talbot, N. J., Ebbole, D. J., Farman, M. L., Mitchell, T. K., Orbach, M. J., Thon, M., Kulkarni, R., Xu, J. R., Pan, H., Read, N. D., Lee, Y. H., Carbone, I., Brown, D., Oh, Y. Y., Donofrio, N., Jeong, J. S., Soanes, D. M., Djonovic, S., Kolomiets, E., Rehmeyer, C., Li, W., Harding, M., Kim, S., Lebrun, M. H., Bohnert, H., Coughlan, S., Butler, J., Calvo, S., Ma, L. J., Nicol, R., Purcell, S., Nusbaum, C., Galagan, J. E., and Birren, B. W. (2005). The genome sequence of the rice blast fungus Magnaporthe grisea. Nature 434, 980–986.
Dehesh, K., Hung, H., Tepperman, J. M., and Quail, P. H. (1992). A transcription factor with twin autonomous DNA-binding domains of closely related but different target sequence specificity. EMBO J. 11, 4131–4144.
Deslandes, L., Olivier, J., Peeters, N., Feng, D. X., Khounlotham, M., Boucher, C., Somssich, I., Genin, S., and Marco, Y. (2003). Physical interaction between RRS1-R, a protein conferring resistance to bacterial wilt, and PopP2, a type III effector targeted to the plant nucleus. Proc. Natl. Acad. Sci. U.S.A. 100, 8024–8029.
Deslandes, L., and Rivas, S. (2011). The plant cell nucleus: a true arena for the fight between plants and pathogens. Plant Signal. Behav. 6, 42–48.
Dingwall, C., Sharnick, S. V., and Laskey, R. A. (1982). A polypeptide domain that specifies migration of nucleoplasmin into the nucleus. Cell 30, 449–458.
Djamei, A., Pitzschke, A., Nakagami, H., Rajh, I., and Hirt, H. (2007). Trojan horse strategy in Agrobacterium transformation: abusing MAPK defense signaling. Science 318, 453–456.
Dou, D., Kale, S. D., Wang, X., Jiang, R. H., Bruce, N. A., Arredondo, F. D., Zhang, X., and Tyler, B. M. (2008). RXLR-mediated entry of Phytophthora sojae effector Avr1b into soybean cells does not require pathogen-encoded machinery. Plant Cell 20, 1930–1947.
Duplessis, S., Cuomo, C. A., Lin, Y. C., Aerts, A., Tisserant, E., Veneault-Fourrey, C., Joly, D. L., Hacquard, S., Amselem, J., Cantarel, B. L., Chiu, R., Coutinho, P. M., Feau, N., Field, M., Frey, P., Gelhaye, E., Goldberg, J., Grabherr, M. G., Kodira, C. D., Kohler, A., Kues, U., Lindquist, E. A., Lucas, S. M., Mago, R., Mauceli, E., Morin, E., Murat, C., Pangilinan, J. L., Park, R., Pearson, M., Quesneville, H., Rouhier, N., Sakthikumar, S., Salamov, A. A., Schmutz, J., Selles, B., Shapiro, H., Tanguay, P., Tuskan, G. A., Henrissat, B., Van de Peer, Y., Rouze, P., Ellis, J. G., Dodds, P. N., Schein, J. E., Zhong, S., Hamelin, R. C., Grigoriev, I. V., Szabo, L. J., and Martin, F. (2011). Obligate biotrophy features unraveled by the genomic analysis of rust fungi. Proc. Natl. Acad. Sci. U.S.A. 108, 9166–9171.
Durrant, W. E., and Dong, X. (2004). Systemic acquired resistance. Annu. Rev. Phytopathol. 42, 185–209.
Elling, A. A., Davis, E. L., Hussey, R. S., and Baum, T. J. (2007). Active uptake of cyst nematode parasitism proteins into the plant cell nucleus. Int. J. Parasitol. 37, 1269–1279.
Friedler, A., Veprintsev, D. B., Freund, S. M., von Glos, K. I., and Fersht, A. R. (2005). Modulation of binding of DNA to the C-terminal domain of p53 by acetylation. Structure 13, 629–636.
Froidure, S., Canonne, J., Daniel, X., Jauneau, A., Briere, C., Roby, D., and Rivas, S. (2010). AtsPLA2-alpha nuclear relocalization by the Arabidopsis transcription factor AtMYB30 leads to repression of the plant defense response. Proc. Natl. Acad. Sci. U.S.A. 107, 15281–15286.
Fu, Z. Q., Guo, M., Jeong, B. R., Tian, F., Elthon, T. E., Cerny, R. L., Staiger, D., and Alfano, J. R. (2007). A type III effector ADP-ribosylates RNA-binding proteins and quells plant immunity. Nature 447, 284–288.
Galan, J. E., and Wolf-Watz, H. (2006). Protein delivery into eukaryotic cells by type III secretion machines. Nature 444, 567–573.
Gao, B., Allen, R., Maier, T., Davis, E. L., Baum, T. J., and Hussey, R. S. (2003). The parasitome of the phytonematode Heterodera glycines. Mol. Plant Microbe Interact. 16, 720–726.
Garcia-Rodriguez, F. M., Schrammeijer, B., and Hooykaas, P. J. (2006). The Agrobacterium VirE3 effector protein: a potential plant transcriptional activator. Nucleic Acids Res. 34, 6496–6504.
Gaulin, E., Madoui, M. A., Bottin, A., Jacquet, C., Mathe, C., Couloux, A., Wincker, P., and Dumas, B. (2008). Transcriptome of Aphanomyces euteiches: new oomycete putative pathogenicity factors and metabolic pathways. PLoS ONE 3, e1723. doi:10.1371/journal.pone.0001723
Geissler, R., Scholze, H., Hahn, S., Streubel, J., Bonas, U., Behrens, S. E., and Boch, J. (2011). Transcriptional activators of human genes with programmable DNA-specificity. PLoS ONE 6, e19509. doi:10.1371/journal.pone.0019509
Gelvin, S. B. (1998). The introduction and expression of transgenes in plants. Curr. Biol. 9, 227–232.
Gheysen, G., and Mitchum, M. G. (2011). How nematodes manipulate plant development pathways for infection. Curr. Opin. Plant Biol. 14, 415–421.
Gilroy, E. M., Taylor, R. M., Hein, I., Boevink, P., Sadanandom, A., and Birch, P. R. (2011). CMPG1-dependent cell death follows perception of diverse pathogen elicitors at the host plasma membrane and is suppressed by Phytophthora infestans RXLR effector AVR3a. New Phytol. 190, 653–666.
Godfrey, D., Bohlenius, H., Pedersen, C., Zhang, Z., Emmersen, J., and Thordal-Christensen, H. (2010). Powdery mildew fungal effector candidates share N-terminal Y/F/WxC-motif. BMC Genomics 11, 317. doi:10.1186/1471-2164-11-317
Gonzalez, C., Szurek, B., Manceau, C., Mathieu, T., Sere, Y., and Verdier, V. (2007). Molecular and pathotypic characterization of new Xanthomonas oryzae strains from West Africa. Mol. Plant Microbe Interact. 20, 534–546.
Goodin, M. M., Dietzgen, R. G., Schichnes, D., Ruzin, S., and Jackson, A. O. (2002). pGD vectors: versatile tools for the expression of green and red fluorescent protein fusions in agroinfiltrated plant leaves. Plant J. 31, 375–383.
Gordeeva, A. V., Zvyagilskaya, R. A., and Labas, Y. A. (2003). Cross-talk between reactive oxygen species and calcium in living cells. Biochemistry Mosc. 68, 1077–1080.
Grant, S. R., Fisher, E. J., Chang, J. H., Mole, B. M., and Dangl, J. L. (2006). Subterfuge and manipulation: type III effector proteins of phytopathogenic bacteria. Annu. Rev. Microbiol. 60, 425–449.
Grunewald, W., Karimi, M., Wieczorek, K., Van de Cappelle, E., Wischnitzki, E., Grundler, F., Inze, D., Beeckman, T., and Gheysen, G. (2008). A role for AtWRKY23 in feeding site establishment of plant-parasitic nematodes. Plant Physiol. 148, 358–368.
Gueneron, M., Timmers, A. C., Boucher, C., and Arlat, M. (2000). Two novel proteins, PopB, which has functional nuclear localization signals, and PopC, which has a large leucine-rich repeat domain, are secreted through the hrp-secretion apparatus of Ralstonia solanacearum. Mol. Microbiol. 36, 261–277.
Haas, B. J., Kamoun, S., Zody, M. C., Jiang, R. H., Handsaker, R. E., Cano, L. M., Grabherr, M., Kodira, C. D., Raffaele, S., Torto-Alalibo, T., Bozkurt, T. O., Ah-Fong, A. M., Alvarado, L., Anderson, V. L., Armstrong, M. R., Avrova, A., Baxter, L., Beynon, J., Boevink, P. C., Bollmann, S. R., Bos, J. I., Bulone, V., Cai, G., Cakir, C., Carrington, J. C., Chawner, M., Conti, L., Costanzo, S., Ewan, R., Fahlgren, N., Fischbach, M. A., Fugelstad, J., Gilroy, E. M., Gnerre, S., Green, P. J., Grenville-Briggs, L. J., Griffith, J., Grunwald, N. J., Horn, K., Horner, N. R., Hu, C. H., Huitema, E., Jeong, D. H., Jones, A. M., Jones, J. D., Jones, R. W., Karlsson, E. K., Kunjeti, S. G., Lamour, K., Liu, Z., Ma, L., Maclean, D., Chibucos, M. C., McDonald, H., McWalters, J., Meijer, H. J., Morgan, W., Morris, P. F., Munro, C. A., O’Neill, K., Ospina-Giraldo, M., Pinzon, A., Pritchard, L., Ramsahoye, B., Ren, Q., Restrepo, S., Roy, S., Sadanandom, A., Savidor, A., Schornack, S., Schwartz, D. C., Schumann, U. D., Schwessinger, B., Seyer, L., Sharpe, T., Silvar, C., Song, J., Studholme, D. J., Sykes, S., Thines, M., van de Vondervoort, P. J., Phuntumart, V., Wawra, S., Weide, R., Win, J., Young, C., Zhou, S., Fry, W., Meyers, B. C., van West, P., Ristaino, J., Govers, F., Birch, P. R., Whisson, S. C., Judelson, H. S., and Nusbaum, C. (2009). Genome sequence and analysis of the Irish potato famine pathogen Phytophthora infestans. Nature 461, 393–398.
Haas, M., Geldreich, A., Bureau, M., Dupuis, L., Leh, V., Vetter, G., Kobayashi, K., Hohn, T., Ryabova, L., Yot, P., and Keller, M. (2005). The open reading frame VI product of Cauliflower mosaic virus is a nucleocytoplasmic protein: its N terminus mediates its nuclear export and formation of electron-dense viroplasms. Plant Cell 17, 927–943.
Haeberle, A. M., and Stussi-Garaud, C. (1995). In situ localization of the non-structural protein P25 encoded by beet necrotic yellow vein virus RNA 3. J. Gen. Virol. 76(Pt 3), 643–650.
Hahn, M., and Mendgen, K. (2001). Signal and nutrient exchange at biotrophic plant-fungus interfaces. Curr. Opin. Plant Biol. 4, 322–327.
Hauck, P., Thilmony, R., and He, S. Y. (2003). A Pseudomonas syringae type III effector suppresses cell wall-based extracellular defense in susceptible Arabidopsis plants. Proc. Natl. Acad. Sci. U.S.A. 100, 8577–8582.
Hayward, A. C. (1991). Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol. 29, 65–87.
Hirooka, T., Rogowsky, P. M., and Kado, C. I. (1987). Characterization of the virE locus of Agrobacterium tumefaciens plasmid pTiC58. J. Bacteriol. 169, 1529–1536.
Hogenhout, S. A., Van der Hoorn, R. A., Terauchi, R., and Kamoun, S. (2009). Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 22, 115–122.
Hong, L., Schroth, G. P., Matthews, H. R., Yau, P., and Bradbury, E. M. (1993). Studies of the DNA binding properties of histone H4 amino terminus. Thermal denaturation studies reveal that acetylation markedly reduces the binding constant of the H4 “tail” to DNA. J. Biol. Chem. 268, 305–314.
Hotson, A., Chosed, R., Shu, H., Orth, K., and Mudgett, M. B. (2003). Xanthomonas type III effector XopD targets SUMO-conjugated proteins in planta. Mol. Microbiol. 50, 377–389.
Hotson, A., and Mudgett, M. B. (2004). Cysteine proteases in phytopathogenic bacteria: identification of plant targets and activation of innate immunity. Curr. Opin. Plant Biol. 7, 384–390.
Huang, G., Dong, R., Allen, R., Davis, E. L., Baum, T. J., and Hussey, R. S. (2006). A root-knot nematode secretory peptide functions as a ligand for a plant transcription factor. Mol. Plant Microbe Interact. 19, 463–470.
Huguet, E., Hahn, K., Wengelnik, K., and Bonas, U. (1998). hpaA mutants of Xanthomonas campestris pv. vesicatoria are affected in pathogenicity but retain the ability to induce host-specific hypersensitive reaction. Mol. Microbiol. 29, 1379–1390.
Huh, W. K., Falvo, J. V., Gerke, L. C., Carroll, A. S., Howson, R. W., Weissman, J. S., and O’Shea, E. K. (2003). Global analysis of protein localization in budding yeast. Nature 425, 686–691.
Jiang, R. H., Tripathy, S., Govers, F., and Tyler, B. M. (2008). RXLR effector reservoir in two Phytophthora species is dominated by a single rapidly evolving superfamily with more than 700 members. Proc. Natl. Acad. Sci. U.S.A. 105, 4874–4879.
Jones, J. T., Kumar, A., Pylypenko, L. A., Thirugnanasambandam, A., Castelli, L., Chapman, S., Cock, P. J., Grenier, E., Lilley, C. J., Phillips, M. S., and Blok, V. C. (2009). Identification and functional characterization of effectors in expressed sequence tags from various life cycle stages of the potato cyst nematode Globodera pallida. Mol. Plant Pathol. 10, 815–828.
Jupin, I., Guilley, H., Richards, K. E., and Jonard, G. (1992). Two proteins encoded by Beet necrotic yellow vein virus RNA 3 influence symptom phenotype on leaves. EMBO J. 11, 479–488.
Kalderon, D., Richardson, W. D., Markham, A. F., and Smith, A. E. (1984a). Sequence requirements for nuclear location of simian virus 40 large-T antigen. Nature 311, 33–38.
Kalderon, D., Roberts, B. L., Richardson, W. D., and Smith, A. E. (1984b). A short amino acid sequence able to specify nuclear location. Cell 39, 499–509.
Kale, S. D., Gu, B., Capelluto, D. G., Dou, D., Feldman, E., Rumore, A., Arredondo, F. D., Hanlon, R., Fudal, I., Rouxel, T., Lawrence, C. B., Shan, W., and Tyler, B. M. (2010). External lipid PI3P mediates entry of eukaryotic pathogen effectors into plant and animal host cells. Cell 142, 284–295.
Kale, S. D., and Tyler, B. M. (2011). Entry of oomycete and fungal effectors into plant and animal host cells. Cell. Microbiol. 13, 1839–1848.
Kamoun, S. (2007). Groovy times: filamentous pathogen effectors revealed. Curr. Opin. Plant Biol. 10, 358–365.
Kamper, J., Kahmann, R., Bolker, M., Ma, L. J., Brefort, T., Saville, B. J., Banuett, F., Kronstad, J. W., Gold, S. E., Muller, O., Perlin, M. H., Wosten, H. A., de Vries, R., Ruiz-Herrera, J., Reynaga-Pena, C. G., Snetselaar, K., McCann, M., Perez-Martin, J., Feldbrugge, M., Basse, C. W., Steinberg, G., Ibeas, J. I., Holloman, W., Guzman, P., Farman, M., Stajich, J. E., Sentandreu, R., Gonzalez-Prieto, J. M., Kennell, J. C., Molina, L., Schirawski, J., Mendoza-Mendoza, A., Greilinger, D., Munch, K., Rossel, N., Scherer, M., Vranes, M., Ladendorf, O., Vincon, V., Fuchs, U., Sandrock, B., Meng, S., Ho, E. C., Cahill, M. J., Boyce, K. J., Klose, J., Klosterman, S. J., Deelstra, H. J., Ortiz-Castellanos, L., Li, W., Sanchez-Alonso, P., Schreier, P. H., Hauser-Hahn, I., Vaupel, M., Koopmann, E., Friedrich, G., Voss, H., Schluter, T., Margolis, J., Platt, D., Swimmer, C., Gnirke, A., Chen, F., Vysotskaia, V., Mannhaupt, G., Guldener, U., Munsterkotter, M., Haase, D., Oesterheld, M., Mewes, H. W., Mauceli, E. W., DeCaprio, D., Wade, C. M., Butler, J., Young, S., Jaffe, D. B., Calvo, S., Nusbaum, C., Galagan, J., and Birren, B. W. (2006). Insights from the genome of the biotrophic fungal plant pathogen Ustilago maydis. Nature 444, 97–101.
Kanneganti, T. D., Bai, X., Tsai, C. W., Win, J., Meulia, T., Goodin, M., Kamoun, S., and Hogenhout, S. A. (2007). A functional genetic assay for nuclear trafficking in plants. Plant J. 50, 149–158.
Kay, S., Boch, J., and Bonas, U. (2005). Characterization of AvrBs3-like effectors from a Brassicaceae pathogen reveals virulence and avirulence activities and a protein with a novel repeat architecture. Mol. Plant Microbe Interact. 18, 838–848.
Kay, S., and Bonas, U. (2009). How Xanthomonas type III effectors manipulate the host plant. Curr. Opin. Microbiol. 12, 37–43.
Kay, S., Hahn, S., Marois, E., Hause, G., and Bonas, U. (2007). A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 318, 648–651.
Kazan, K. (2006). Negative regulation of defence and stress genes by EAR-motif-containing repressors. Trends Plant Sci. 11, 109–112.
Kelley, B. S., Lee, S. J., Damasceno, C. M., Chakravarthy, S., Kim, B. D., Martin, G. B., and Rose, J. K. (2010). A secreted effector protein (SNE1) from Phytophthora infestans is a broadly acting suppressor of programmed cell death. Plant J. 62, 357–366.
Kemen, E., Kemen, A. C., Rafiqi, M., Hempel, U., Mendgen, K., Hahn, M., and Voegele, R. T. (2005). Identification of a protein from rust fungi transferred from haustoria into infected plant cells. Mol. Plant Microbe Interact. 18, 1130–1139.
Kim, J. G., Taylor, K. W., Hotson, A., Keegan, M., Schmelz, E. A., and Mudgett, M. B. (2008). XopD SUMO protease affects host transcription, promotes pathogen growth, and delays symptom development in Xanthomonas-infected tomato leaves. Plant Cell 20, 1915–1929.
Kinkema, M., Fan, W., and Dong, X. (2000). Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 12, 2339–2350.
Kitakura, S., Fujita, T., Ueno, Y., Terakura, S., Wabiko, H., and Machida, Y. (2002). The protein encoded by oncogene 6b from Agrobacterium tumefaciens interacts with a nuclear protein of tobacco. Plant Cell 14, 451–463.
Kloppholz, S., Kuhn, H., and Requena, N. (2011). A secreted fungal effector of Glomus intraradices promotes symbiotic biotrophy. Curr. Biol. 21, 1204–1209.
Kornberg, R. D., and Lorch, Y. (1999). Chromatin-modifying and -remodeling complexes. Curr. Opin. Genet. Dev. 9, 148–151.
Kudo, N., Wolff, B., Sekimoto, T., Schreiner, E. P., Yoneda, Y., Yanagida, M., Horinouchi, S., and Yoshida, M. (1998). Leptomycin B inhibition of signal-mediated nuclear export by direct binding to CRM1. Exp. Cell Res. 242, 540–547.
Lacroix, B., and Citovsky, V. (2009). Agrobacterium aiming for the host chromatin: Host and bacterial proteins involved in interactions between T-DNA and plant nucleosomes. Commun. Integr. Biol. 2, 42–45.
Lacroix, B., Loyter, A., and Citovsky, V. (2008). Association of the Agrobacterium T-DNA-protein complex with plant nucleosomes. Proc. Natl. Acad. Sci. U.S.A. 105, 15429–15434.
Lange, A., Mills, R. E., Lange, C. J., Stewart, M., Devine, S. E., and Corbett, A. H. (2007). Classical nuclear localization signals: definition, function, and interaction with importin alpha. J. Biol. Chem. 282, 5101–5105.
Leh, V., Yot, P., and Keller, M. (2000). The Cauliflower mosaic virus translational transactivator interacts with the 60S ribosomal subunit protein L18 of Arabidopsis thaliana. Virology 266, 1–7.
Levesque, C. A., Brouwer, H., Cano, L., Hamilton, J. P., Holt, C., Huitema, E., Raffaele, S., Robideau, G. P., Thines, M., Win, J., Zerillo, M. M., Beakes, G. W., Boore, J. L., Busam, D., Dumas, B., Ferriera, S., Fuerstenberg, S. I., Gachon, C. M., Gaulin, E., Govers, F., Grenville-Briggs, L., Horner, N., Hostetler, J., Jiang, R. H., Johnson, J., Krajaejun, T., Lin, H., Meijer, H. J., Moore, B., Morris, P., Phuntmart, V., Puiu, D., Shetty, J., Stajich, J. E., Tripathy, S., Wawra, S., van West, P., Whitty, B. R., Coutinho, P. M., Henrissat, B., Martin, F., Thomas, P. D., Tyler, B. M., De Vries, R. P., Kamoun, S., Yandell, M., Tisserat, N., and Buell, C. R. (2010). Genome sequence of the necrotrophic plant pathogen Pythium ultimum reveals original pathogenicity mechanisms and effector repertoire. Genome Biol. 11, R73.
Li, H., Xu, H., Zhou, Y., Zhang, J., Long, C., Li, S., Chen, S., Zhou, J. M., and Shao, F. (2007). The phosphothreonine lyase activity of a bacterial type III effector family. Science 315, 1000–1003.
Li, J., Krichevsky, A., Vaidya, M., Tzfira, T., and Citovsky, V. (2005). Uncoupling of the functions of the Arabidopsis VIP1 protein in transient and stable plant genetic transformation by Agrobacterium. Proc. Natl. Acad. Sci. U.S.A. 102, 5733–5738.
Li, X. H., and Carrington, J. C. (1995). Complementation of Tobacco etch potyvirus mutants by active RNA polymerase expressed in transgenic cells. Proc. Natl. Acad. Sci. U.S.A. 92, 457–461.
Li, X. H., Valdez, P., Olvera, R. E., and Carrington, J. C. (1997). Functions of the Tobacco etch virus RNA polymerase (NIb): subcellular transport and protein-protein interaction with VPg/proteinase (NIa). J. Virol. 71, 1598–1607.
Lindeberg, M., Cartinhour, S., Myers, C. R., Schechter, L. M., Schneider, D. J., and Collmer, A. (2006). Closing the circle on the discovery of genes encoding Hrp regulon members and type III secretion system effectors in the genomes of three model Pseudomonas syringae strains. Mol. Plant Microbe Interact. 19, 1151–1158.
Lindgren, P. B., Peet, R. C., and Panopoulos, N. J. (1986). Gene cluster of Pseudomonas syringae pv. “phaseolicola” controls pathogenicity of bean plants and hypersensitivity of nonhost plants. J. Bacteriol. 168, 512–522.
Liu, J., and Coaker, G. (2008). Nuclear trafficking during plant innate immunity. Mol. Plant 1, 411–422.
Liu, T., Ye, W., Ru, Y., Yang, X., Gu, B., Tao, K., Lu, S., Dong, S., Zheng, X., Shan, W., Wang, Y., and Dou, D. (2011). Two host cytoplasmic effectors are required for pathogenesis of Phytophthora sojae by suppression of host defenses. Plant Physiol. 155, 490–501.
Loyter, A., Rosenbluh, J., Zakai, N., Li, J., Kozlovsky, S. V., Tzfira, T., and Citovsky, V. (2005). The plant VirE2 interacting protein 1. a molecular link between the Agrobacterium T-complex and the host cell chromatin? Plant Physiol. 138, 1318–1321.
Lucy, A. P., Guo, H. S., Li, W. X., and Ding, S. W. (2000). Suppression of post-transcriptional gene silencing by a plant viral protein localized in the nucleus. EMBO J. 19, 1672–1680.
Ma, K. W., Flores, C., and Ma, W. (2011). Chromatin configuration as a battlefield in plant-bacteria interactions. Plant Physiol. 157, 535–543.
Marois, E., Van den Ackerveken, G., and Bonas, U. (2002). The Xanthomonas type III effector protein AvrBs3 modulates plant gene expression and induces cell hypertrophy in the susceptible host. Mol. Plant Microbe Interact. 15, 637–646.
Mazzolini, L., Dabos, P., Constantin, S., and Yot, P. (1989). Further evidence that viroplasms are the site of Cauliflower mosaic virus genome replication by reverse transcription during viral infection. J. Gen. Virol. 70, 3439–3449.
McCann, H. C., and Guttman, D. S. (2008). Evolution of the type III secretion system and its effectors in plant-microbe interactions. New Phytol. 177, 33–47.
Meier, I. (2007). Composition of the plant nuclear envelope: theme and variations. J. Exp. Bot. 58, 27–34.
Merkle, T. (2001). Nuclear import and export of proteins in plants: a tool for the regulation of signalling. Planta 213, 499–517.
Moscou, M. J., and Bogdanove, A. J. (2009). A simple cipher governs DNA recognition by TAL effectors. Science 326, 1501.
Mou, Z., Fan, W., and Dong, X. (2003). Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 113, 935–944.
Mueller, O., Kahmann, R., Aguilar, G., Trejo-Aguilar, B., Wu, A., and de Vries, R. P. (2008). The secretome of the maize pathogen Ustilago maydis. Fungal Genet. Biol. 45(Suppl. 1), S63–S70.
Mujtaba, S., He, Y., Zeng, L., Yan, S., Plotnikova, O., Sachchidanand, Sanchez, R., Zeleznik-Le, N. J., Ronai, Z., and Zhou, M. M. (2004). Structural mechanism of the bromodomain of the coactivator CBP in p53 transcriptional activation. Mol. Cell 13, 251–263.
Mur, L. A., Kenton, P., Lloyd, A. J., Ougham, H., and Prats, E. (2008). The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 59, 501–520.
Murakami, M. T., Sforca, M. L., Neves, J. L., Paiva, J. H., Domingues, M. N., Pereira, A. L., Zeri, A. C., and Benedetti, C. E. (2010). The repeat domain of the type III effector protein PthA shows a TPR-like structure and undergoes conformational changes upon DNA interaction. Proteins 78, 3386–3395.
Nissan, G., Manulis-Sasson, S., Chalupowicz, L., Teper, D., Yeheskel, A., Pasmanik-Chor, M., Sessa, G., and Barash, I. (2011). The type III effector HsvG of the gall-forming Pantoea agglomerans mediates expression of the host gene HSVGT. Mol. Plant Microbe Interact.doi: 10.1094/MPMI-06-11-0173
Nissan, G., Manulis-Sasson, S., Weinthal, D., Mor, H., Sessa, G., and Barash, I. (2006). The type III effectors HsvG and HsvB of gall-forming Pantoea agglomerans determine host specificity and function as transcriptional activators. Mol. Microbiol. 61, 1118–1131.
Oh, S. K., Young, C., Lee, M., Oliva, R., Bozkurt, T. O., Cano, L. M., Win, J., Bos, J. I., Liu, H. Y., van Damme, M., Morgan, W., Choi, D., Van der Vossen, E. A., Vleeshouwers, V. G., and Kamoun, S. (2009). In planta expression screens of Phytophthora infestans RXLR effectors reveal diverse phenotypes, including activation of the Solanum bulbocastanum disease resistance protein Rpi-blb2. Plant Cell 21, 2928–2947.
Ossareh-Nazari, B., Gwizdek, C., and Dargemont, C. (2001). Protein export from the nucleus. Traffic 2, 684–689.
Palma, K., Zhang, Y., and Li, X. (2005). An importin alpha homolog, MOS6, plays an important role in plant innate immunity. Curr. Biol. 15, 1129–1135.
Panstruga, R., and Dodds, P. N. (2009). Terrific protein traffic: the mystery of effector protein delivery by filamentous plant pathogens. Science 324, 748–750.
Park, H. S., Himmelbach, A., Browning, K. S., Hohn, T., and Ryabova, L. A. (2001). A plant viral “reinitiation” factor interacts with the host translational machinery. Cell 106, 723–733.
Patel, S. S., Belmont, B. J., Sante, J. M., and Rexach, M. F. (2007). Natively unfolded nucleoporins gate protein diffusion across the nuclear pore complex. Cell 129, 83–96.
Plett, J. M., Kemppainen, M., Kale, S. D., Kohler, A., Legue, V., Brun, A., Tyler, B. M., Pardo, A. G., and Martin, F. (2011). A secreted effector protein of Laccaria bicolor is required for symbiosis development. Curr. Biol. 21, 1197–1203.
Puthoff, D. P., Neelam, A., Ehrenfried, M. L., Scheffler, B. E., Ballard, L., Song, Q., Campbell, K. B., Cooper, B., and Tucker, M. L. (2008). Analysis of expressed sequence tags from Uromyces appendiculatus hyphae and haustoria and their comparison to sequences from other rust fungi. Phytopathology 98, 1126–1135.
Qian, W., Jia, Y., Ren, S. X., He, Y. Q., Feng, J. X., Lu, L. F., Sun, Q., Ying, G., Tang, D. J., Tang, H., Wu, W., Hao, P., Wang, L., Jiang, B. L., Zeng, S., Gu, W. Y., Lu, G., Rong, L., Tian, Y., Yao, Z., Fu, G., Chen, B., Fang, R., Qiang, B., Chen, Z., Zhao, G. P., Tang, J. L., and He, C. (2005). Comparative and functional genomic analyses of the pathogenicity of phytopathogen Xanthomonas campestris pv. campestris. Genome Res. 15, 757–767.
Qutob, D., Kamoun, S., and Gijzen, M. (2002). Expression of a Phytophthora sojae necrosis-inducing protein occurs during transition from biotrophy to necrotrophy. Plant J. 32, 361–373.
Raffaele, S., Vailleau, F., Leger, A., Joubes, J., Miersch, O., Huard, C., Blee, E., Mongrand, S., Domergue, F., and Roby, D. (2008). A MYB transcription factor regulates very-long-chain fatty acid biosynthesis for activation of the hypersensitive cell death response in Arabidopsis. Plant Cell 20, 752–767.
Rawlings, N. D., Morton, F. R., and Barrett, A. J. (2006). MEROPS: the peptidase database. Nucleic Acids Res. 34, D270–D272.
Rehman, S., Postma, W., Tytgat, T., Prins, P., Qin, L., Overmars, H., Vossen, J., Spiridon, L. N., Petrescu, A. J., Goverse, A., Bakker, J., and Smant, G. (2009). A secreted SPRY domain-containing protein (SPRYSEC) from the plant-parasitic nematode Globodera rostochiensis interacts with a CC-NB-LRR protein from a susceptible tomato. Mol. Plant Microbe Interact. 22, 330–340.
Restrepo, M. A., Freed, D. D., and Carrington, J. C. (1990). Nuclear transport of plant potyviral proteins. Plant Cell 2, 987–998.
Richards, T. A., Soanes, D. M., Jones, M. D., Vasieva, O., Leonard, G., Paszkiewicz, K., Foster, P. G., Hall, N., and Talbot, N. J. (2011). Horizontal gene transfer facilitated the evolution of plant parasitic mechanisms in the oomycetes. Proc. Natl. Acad. Sci. U.S.A. 108, 15258–15263.
Rivas, S. (2011). Nuclear dynamics during plant innate immunity. Plant Physiol. doi: 10.1104/pp.111.186163
Robbins, J., Dilworth, S. M., Laskey, R. A., and Dingwall, C. (1991). Two interdependent basic domains in nucleoplasmin nuclear targeting sequence: identification of a class of bipartite nuclear targeting sequence. Cell 64, 615–623.
Roden, J., Eardley, L., Hotson, A., Cao, Y., and Mudgett, M. B. (2004). Characterization of the Xanthomonas AvrXv4 effector, a SUMO protease translocated into plant cells. Mol. Plant Microbe Interact. 17, 633–643.
Romer, P., Hahn, S., Jordan, T., Strauss, T., Bonas, U., and Lahaye, T. (2007). Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 318, 645–648.
Romer, P., Recht, S., and Lahaye, T. (2009). A single plant resistance gene promoter engineered to recognize multiple TAL effectors from disparate pathogens. Proc. Natl. Acad. Sci. U.S.A. 106, 20526–20531.
Romer, P., Recht, S., Strauss, T., Elsaesser, J., Schornack, S., Boch, J., Wang, S., and Lahaye, T. (2010). Promoter elements of rice susceptibility genes are bound and activated by specific TAL effectors from the bacterial blight pathogen, Xanthomonas oryzae pv. oryzae. New Phytol. 187, 1048–1057.
Roze, E., Hanse, B., Mitreva, M., Vanholme, B., Bakker, J., and Smant, G. (2008). Mining the secretome of the root-knot nematode Meloidogyne chitwoodi for candidate parasitism genes. Mol. Plant Pathol. 9, 1–10.
Ryabov, E. V., Kim, S. H., and Taliansky, M. (2004). Identification of a nuclear localization signal and nuclear export signal of the umbraviral long-distance RNA movement protein. J. Gen. Virol. 85, 1329–1333.
Ryabov, E. V., Oparka, K. J., Santa Cruz, S., Robinson, D. J., and Taliansky, M. E. (1998). Intracellular location of two groundnut rosette umbravirus proteins delivered by PVX and TMV vectors. Virology 242, 303–313.
Sacco, M. A., Koropacka, K., Grenier, E., Jaubert, M. J., Blanchard, A., Goverse, A., Smant, G., and Moffett, P. (2009). The cyst nematode SPRYSEC protein RBP-1 elicits Gpa2- and RanGAP2-dependent plant cell death. PLoS Pathog. 5, e1000564. doi:10.1371/journal.ppat.1000564
Schlaich, N. L. (2007). Flavin-containing monooxygenases in plants: looking beyond detox. Trends Plant Sci. 12, 412–418.
Scholze, H., and Boch, J. (2011). TAL effectors are remote controls for gene activation. Curr. Opin. Microbiol. 14, 47–53.
Schornack, S., Minsavage, G. V., Stall, R. E., Jones, J. B., and Lahaye, T. (2008). Characterization of AvrHah1, a novel AvrBs3-like effector from Xanthomonas gardneri with virulence and avirulence activity. New Phytol. 179, 546–556.
Schornack, S., van Damme, M., Bozkurt, T. O., Cano, L. M., Smoker, M., Thines, M., Gaulin, E., Kamoun, S., and Huitema, E. (2010). Ancient class of translocated oomycete effectors targets the host nucleus. Proc. Natl. Acad. Sci. U.S.A. 107, 17421–17426.
Schrammeijer, B., Risseeuw, E., Pansegrau, W., Regensburg-Tuink, T. J., Crosby, W. L., and Hooykaas, P. J. (2001). Interaction of the virulence protein VirF of Agrobacterium tumefaciens with plant homologs of the yeast Skp1 protein. Curr. Biol. 11, 258–262.
Shen, Q. H., and Schulze-Lefert, P. (2007). Rumble in the nuclear jungle: compartmentalization, trafficking, and nuclear action of plant immune receptors. EMBO J. 26, 4293–4301.
Stassen, J. H., and Van den Ackerveken, G. (2011). How do oomycete effectors interfere with plant life? Curr. Opin. Plant Biol. 14, 407–414.
Sugio, A., Kingdom, H. N., Maclean, A. M., Grieve, V. M., and Hogenhout, S. A. (2011a). Phytoplasma protein effector SAP11 enhances insect vector reproduction by manipulating plant development and defense hormone biosynthesis. Proc. Natl. Acad. Sci. U.S.A. 108, E1254–E1263.
Sugio, A., MacLean, A. M., Kingdom, H. N., Grieve, V. M., Manimekalai, R., and Hogenhout, S. A. (2011b). Diverse targets of phytoplasma effectors: from plant development to defense against insects. Annu. Rev. Phytopathol. 49, 175–195.
Szczesny, R., Buttner, D., Escolar, L., Schulze, S., Seiferth, A., and Bonas, U. (2010). Suppression of the AvrBs1-specific hypersensitive response by the YopJ effector homolog AvrBsT from Xanthomonas depends on a SNF1-related kinase. New Phytol. 187, 1058–1074.
Szurek, B., Marois, E., Bonas, U., and Van den Ackerveken, G. (2001). Eukaryotic features of the Xanthomonas type III effector AvrBs3: protein domains involved in transcriptional activation and the interaction with nuclear import receptors from pepper. Plant J. 26, 523–534.
Szurek, B., Rossier, O., Hause, G., and Bonas, U. (2002). Type III-dependent translocation of the Xanthomonas AvrBs3 protein into the plant cell. Mol. Microbiol. 46, 13–23.
Tamada, T., Uchino, H., Kusume, T., and Saito, M. (1999). RNA 3 deletion mutants of Beet necrotic yellow vein virus do not cause rhizomania disease in sugar beets. Phytopathology 89, 1000–1006.
Tao, Y., Xie, Z., Chen, W., Glazebrook, J., Chang, H. S., Han, B., Zhu, T., Zou, G., and Katagiri, F. (2003). Quantitative nature of Arabidopsis responses during compatible and incompatible interactions with the bacterial pathogen Pseudomonas syringae. Plant Cell 15, 317–330.
Tasset, C., Bernoux, M., Jauneau, A., Pouzet, C., Brière, C., Kieffer-Jacquinod, S., Rivas, S., Marco, Y., and Deslandes, L. (2010). Autoacetylation of the Ralstonia solanacearum effector PopP2 targets a lysine residue essential for RRS1-R mediated immunity in Arabidopsis. PLoS Pathog. 6, e1001202. doi:10.1371/journal.ppat.1001202
Terakura, S., Ueno, Y., Tagami, H., Kitakura, S., Machida, C., Wabiko, H., Aiba, H., Otten, L., Tsukagoshi, H., Nakamura, K., and Machida, Y. (2007). An oncoprotein from the plant pathogen Agrobacterium has histone chaperone-like activity. Plant Cell 19, 2855–2865.
Thieme, F., Szczesny, R., Urban, A., Kirchner, O., Hause, G., and Bonas, U. (2007). New type III effectors from Xanthomonas campestris pv. vesicatoria trigger plant reactions dependent on a conserved N-myristoylation motif. Mol. Plant Microbe Interact. 20, 1250–1261.
Tinland, B., Koukolikova-Nicola, Z., Hall, M. N., and Hohn, B. (1992). The T-DNA-linked VirD2 protein contains two distinct functional nuclear localization signals. Proc. Natl. Acad. Sci. U.S.A. 89, 7442–7446.
Tinland, B., Rohfritsch, O., Michler, P., and Otten, L. (1990). Agrobacterium tumefaciens T-DNA gene 6b stimulates rol-induced root formation, permits growth at high auxin concentrations and increases root size. Mol. Gen. Genet. 223, 1–10.
Torres, M. A., Dangl, J. L., and Jones, J. D. (2002). Arabidopsis gp91phox homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. U.S.A. 99, 517–522.
Tsai, C. W., Redinbaugh, M. G., Willie, K. J., Reed, S., Goodin, M., and Hogenhout, S. A. (2005). Complete genome sequence and in planta subcellular localization of maize fine streak virus proteins. J. Virol. 79, 5304–5314.
Tyler, B. M., Tripathy, S., Zhang, X., Dehal, P., Jiang, R. H., Aerts, A., Arredondo, F. D., Baxter, L., Bensasson, D., Beynon, J. L., Chapman, J., Damasceno, C. M., Dorrance, A. E., Dou, D., Dickerman, A. W., Dubchak, I. L., Garbelotto, M., Gijzen, M., Gordon, S. G., Govers, F., Grunwald, N. J., Huang, W., Ivors, K. L., Jones, R. W., Kamoun, S., Krampis, K., Lamour, K. H., Lee, M. K., McDonald, W. H., Medina, M., Meijer, H. J., Nordberg, E. K., Maclean, D. J., Ospina-Giraldo, M. D., Morris, P. F., Phuntumart, V., Putnam, N. H., Rash, S., Rose, J. K., Sakihama, Y., Salamov, A. A., Savidor, A., Scheuring, C. F., Smith, B. M., Sobral, B. W., Terry, A., Torto-Alalibo, T. A., Win, J., Xu, Z., Zhang, H., Grigoriev, I. V., Rokhsar, D. S., and Boore, J. L. (2006). Phytophthora genome sequences uncover evolutionary origins and mechanisms of pathogenesis. Science 313, 1261–1266.
Tytgat, T., Vanholme, B., De Meutter, J., Claeys, M., Couvreur, M., Vanhoutte, I., Gheysen, G., Van Criekinge, W., Borgonie, G., and Coomans, A. (2004). A new class of ubiquitin extension proteins secreted by the dorsal pharyngeal gland in plant parasitic cyst nematodes. Mol. Plant Microbe Interact. 17, 846–852.
Tzfira, T., and Citovsky, V. (2002). Partners-in-infection: host proteins involved in the transformation of plant cells by Agrobacterium. Trends Cell Biol. 12, 121–129.
Tzfira, T., Vaidya, M., and Citovsky, V. (2001). VIP1, an Arabidopsis protein that interacts with Agrobacterium VirE2, is involved in VirE2 nuclear import and Agrobacterium infectivity. EMBO J. 20, 3596–3607.
Tzfira, T., Vaidya, M., and Citovsky, V. (2004). Involvement of targeted proteolysis in plant genetic transformation by Agrobacterium. Nature 431, 87–92.
Van den Ackerveken, G., Marois, E., and Bonas, U. (1996). Recognition of the bacterial avirulence protein AvrBs3 occurs inside the host plant cell. Cell 87, 1307–1316.
van der Biezen, E. A., and Jones, J. D. G. (1998). Plant disease resistance proteins and the gene for gene concept. Trends Biochem. Sci. 23, 454–456.
Veena, Jiang, H., Doerge, R. W., and Gelvin, S. B. (2003). Transfer of T-DNA and Vir proteins to plant cells by Agrobacterium tumefaciens induces expression of host genes involved in mediating transformation and suppresses host defense gene expression. Plant J. 35, 219–236.
Vergunst, A. C., Schrammeijer, B., den Dulk-Ras, A., de Vlaam, C. M., Regensburg-Tuink, T. J., and Hooykaas, P. J. (2000). VirB/D4-dependent protein translocation from Agrobacterium into plant cells. Science 290, 979–982.
Vetter, G., Hily, J. M., Klein, E., Schmidlin, L., Haas, M., Merkle, T., and Gilmer, D. (2004). Nucleo-cytoplasmic shuttling of the Beet necrotic yellow vein virus RNA-3-encoded p25 protein. J. Gen. Virol. 85, 2459–2469.
Vleeshouwers, V. G., Raffaele, S., Vossen, J. H., Champouret, N., Oliva, R., Segretin, M. E., Rietman, H., Cano, L. M., Lokossou, A., Kessel, G., Pel, M. A., and Kamoun, S. (2011). Understanding and exploiting late blight resistance in the age of effectors. Annu. Rev. Phytopathol. 49, 507–531.
Vleeshouwers, V. G., Rietman, H., Krenek, P., Champouret, N., Young, C., Oh, S. K., Wang, M., Bouwmeester, K., Vosman, B., Visser, R. G., Jacobsen, E., Govers, F., Kamoun, S., and Van der Vossen, E. A. (2008). Effector genomics accelerates discovery and functional profiling of potato disease resistance and Phytophthora infestans avirulence genes. PLoS ONE 3, e2875. doi:10.1371/journal.pone.0002875
Vorhölter, F. J., Thias, T., Meyer, F., Bekel, T., Kaiser, O., Puhler, A., and Niehaus, K. (2003). Comparison of two Xanthomonas campestris pathovar campestris genomes revealed differences in their gene composition. J. Biotechnol. 106, 193–202.
Wang, M., Soyano, T., Machida, S., Yang, J. Y., Jung, C., Chua, N. H., and Yuan, Y. A. (2011). Molecular insights into plant cell proliferation disturbance by Agrobacterium protein 6b. Genes Dev. 25, 64–76.
Ward, B. M., and Lazarowitz, S. G. (1999). Nuclear export in plants. Use of geminivirus movement proteins for a cell-based export assay. Plant Cell 11, 1267–1276.
Weinthal, D. M., Barash, I., Tzfira, T., Gaba, V., Teper, D., Sessa, G., and Manulis-Sasson, S. (2011). Characterization of nuclear localization signals in the type III effectors HsvG and HsvB of the gall-forming bacterium Pantoea agglomerans. Microbiology 157, 1500–1508.
Weis, K. (2002). Nucleocytoplasmic transport: cargo trafficking across the border. Curr. Opin. Cell Biol. 14, 328–335.
Whisson, S. C., Boevink, P. C., Moleleki, L., Avrova, A. O., Morales, J. G., Gilroy, E. M., Armstrong, M. R., Grouffaud, S., van West, P., Chapman, S., Hein, I., Toth, I. K., Pritchard, L., and Birch, P. R. (2007). A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature 450, 115–118.
White, F. F., Potnis, N., Jones, J. B., and Koebnik, R. (2009). The type III effectors of Xanthomonas. Mol. Plant Pathol. 10, 749–766.
Wiermer, M., Palma, K., Zhang, Y., and Li, X. (2007). Should i stay or should i go? Nucleocytoplasmic trafficking in plant innate immunity. Cell. Microbiol. 9, 1880–1890.
Win, J., Morgan, W., Bos, J., Krasileva, K. V., Cano, L. M., Chaparro-Garcia, A., Ammar, R., Staskawicz, B. J., and Kamoun, S. (2007). Adaptive evolution has targeted the C-terminal domain of the RXLR effectors of plant pathogenic oomycetes. Plant Cell 19, 2349–2369.
Yang, B., Sugio, A., and White, F. F. (2006). Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. U.S.A. 103, 10503–10508.
Yang, Y., and Gabriel, D. W. (1995). Xanthomonas avirulence/pathogenicity gene family encodes functional plant nuclear targeting signals. Mol. Plant Microbe Interact. 8, 627–631.
Yuan, M., Chu, Z., Li, X., Xu, C., and Wang, S. (2010). The bacterial pathogen Xanthomonas oryzae overcomes rice defenses by regulating host copper redistribution. Plant Cell 22, 3164–3176.
Zaltsman, A., Krichevsky, A., Loyter, A., and Citovsky, V. (2010). Agrobacterium induces expression of a host F-box protein required for tumorigenicity. Cell Host Microbe 7, 197–209.
Zhang, J., Shao, F., Cui, H., Chen, L. J., Li, H. T., Zou, Y., Long, C. Z., Lan, L. F., Chai, J. J., Chen, S., Tang, X. Y., and Zhou, J. M. (2007). A Pseudomonas syringae effector inactivates MAPKs to suppress PAMP-Induced immunity in plants. Cell Host Microbe 1, 175–185.
Zhang, Y., and Li, X. (2005). A putative nucleoporin 96 Is required for both basal defense and constitutive resistance responses mediated by suppressor of npr1-1, constitutive 1. Plant Cell 17, 1306–1316.
Zurawski, D. V., Mumy, K. L., Faherty, C. S., McCormick, B. A., and Maurelli, A. T. (2009). Shigella flexneri type III secretion system effectors OspB and OspF target the nucleus to downregulate the host inflammatory response via interactions with retinoblastoma protein. Mol. Microbiol. 71, 350–368.
Keywords: microbial effector, NES, NLS, nuclear pore complex, plant immunity, nucleocytoplasmic protein translocation, resistance protein, transcription factor
Citation: Rivas S and Genin S (2011) A plethora of virulence strategies hidden behind nuclear targeting of microbial effectors. Front. Plant Sci. 2:104. doi: 10.3389/fpls.2011.00104
Received: 18 October 2011;
Accepted: 09 December 2011;
Published online: 27 December 2011.
Edited by:
Vitaly Citovsky, State University of New York at Stony Brook, USAReviewed by:
Thierry Rouxel, Institut National de la Recherche Agronomique, FranceJens Staal, Ghent University, Belgium
Copyright: © 2011 Rivas and Genin. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Susana Rivas, Laboratoire des Interactions Plantes-Microorganismes, UMR CNRS/INRA 2594/441, Chemin de Borde Rouge, BP 52627, 31326 Castanet-Tolosan Cedex, France. e-mail:c3VzYW5hLnJpdmFzQHRvdWxvdXNlLmlucmEuZnI=