 Roberta Acri-Nunes-Miranda
Roberta Acri-Nunes-Miranda Mariana Mondragón-Palomino
Mariana Mondragón-Palomino- Department of Cell Biology and Plant Biochemistry, University of Regensburg, Regensburg, Germany
The diverse flowers of Orchidaceae are the result of several major morphological transitions, among them the most studied is the differentiation of the inner median tepal into the labellum, a perianth organ key in pollinator attraction. Type A peloria lacking stamens and with ectopic labella in place of inner lateral tepals are useful for testing models on the genes specifying these organs by comparing their patterns of expression between wild-type and peloric flowers. Previous studies focused on DEFICIENS- and GLOBOSA-like MADS-box genes because of their conserved role in perianth and stamen development. The “orchid code” model summarizes this work and shows in Orchidaceae there are four paralogous lineages of DEFICIENS/AP3-like genes differentially expressed in each floral whorl. Experimental tests of this model showed the conserved, higher expression of genes from two specific DEF-like gene lineages is associated with labellum development. The present study tests whether eight MADS-box candidate SEP-, FUL-, AG-, and STK-like genes have been specifically duplicated in the Orchidaceae and are also differentially expressed in association with the distinct flower organs of Phalaenopsis hyb. “Athens.” The gene trees indicate orchid-specific duplications. In a way analogous to what is observed in labellum-specific DEF-like genes, a two-fold increase in the expression of SEP3-like gene PhaMADS7 was measured in the labellum-like inner lateral tepals of peloric flowers. The overlap between SEP3-like and DEF-like genes suggests both are associated with labellum specification and similar positional cues determine their domains of expression. In contrast, the uniform messenger levels of FUL-like genes suggest they are involved in the development of all organs and their expression in the ovary suggests cell differentiation starts before pollination. As previously reported AG-like and STK-like genes are exclusively expressed in gynostemium and ovary, however no evidence for transcriptional divergence was found in the stage investigated. Gene expression suggests a developmental regulatory system based on the combined activity of duplicate MADS-box genes. We discuss its feasibility based on documented protein interactions and patterns of expression.
Introduction
Over the last two decades comparative molecular genetic analysis of flower development and evolution have been fundamentally influenced by the ABC model of organ identity specification. This model resulted from the genetic analysis of floral homeotic mutants of Arabidopsis thaliana and Antirrhinum majus (Coen and Meyerowitz, 1991; Weigel and Meyerowitz, 1994).The original ABC and the extended ABCDE model associate the developmental determination of specific flower organs of Arabidopsis thaliana with the combinatorial activity of several classes of homeotic selector genes, most of which encode MADS domain developmental transcription factors: A- and E-class genes specify sepals; genes from classes A, B, and E determine petals; the combination of B-, C-, and E-class genes specify stamens; genes from class C and E determine carpels; and D- and E-class genes determine ovules (Reviewed in Theissen, 2001; Krizek and Fletcher, 2005).
Comparative studies demonstrate the conservation of homologs of the ABCDE genes across most Angiosperms (Becker and Theissen, 2003; Litt and Irish, 2003; Kramer et al., 2004; Zahn et al., 2005a,b, 2006), and suggest the regulatory principles of some of these homologs have been conserved during flower evolution (Whipple et al., 2004, 2007; Melzer et al., 2009; Cui et al., 2010).
The phylogenetic relationships of MADS-box genes have been investigated in depth and several studies consistently demonstrated AP1-/FUL-, DEF-/GLO-, AG-/STK-, and SEP-like genes form distinct phylogenetic groups (Purugganan et al., 1995; Theissen et al., 1996; Alvarez-Buylla et al., 2000; Becker and Theissen, 2003; Martinez-Castilla and Alvarez-Buylla, 2003; Nam et al., 2003). The finding that the genes in each clade or group of the MADS-box gene tree share distinct molecular sequences and have similar patterns of expression and functions suggests gene duplication and functional diversification of MADS-box genes played an important role in the evolution of flower morphology (Purugganan et al., 1995; Theissen et al., 1996). Furthermore, this relationship provides a solid comparative framework to generate and test hypotheses on the developmental program of non-model species.
It has been frequently observed that most non-grass monocot species like Tulipa gesneriana, Agapanthus praecox and Muscari armeniacum have several copies of DEF- and GLO-like genes that are expressed in the petaloid first whorl (Van Tunen et al., 1993; Kanno et al., 2003; Nakamura et al., 2005; Nakada et al., 2006). This shift in the pattern of expression respective to homologous genes from Arabidopsis thaliana and Antirrhinum majus is the basis of a proposed modification to the ABC model of flower identity specification for non-grass monocots. In this model DEF- and GLO-like genes determine tepal identity in the first whorl in addition to specifying inner tepal and stamen identity (Kanno et al., 2007). Nevertheless direct evidence supporting the modified ABC is still missing largely due to the technical hurdles of genetic analysis and stable transformation of non-grass monocots.
However, not all monocot species with petaloid flowers develop two whorls of identical perianth organs (tepals). Most flowers of Orchidaceae, the largest monocot family, have a highly differentiated zygomorphic perianth, including three types of organs: three outer tepals (sepals) in the first floral whorl, two inner lateral tepals (petals) as well as a frequently highly modified inner median tepal called labellum (or lip) in the second floral whorl (Rudall and Bateman, 2002) (Figure 1A). The labellum is positionally homologous to the adaxial tepal of other monocot flowers, but its position in the perianth is often the lowest because of a 180° torsion of the pedicel and/or ovary (resupination) that changes floral orientation (Arditti, 2002; Bateman and Rudall, 2006). The orientation of the labellum, its location in direct opposition to the fertile anther, its often distinct pattern of coloration as well as the presence of calli, spur, oil, or scent glands suggest its morphology is influenced by co-evolution with pollinators.
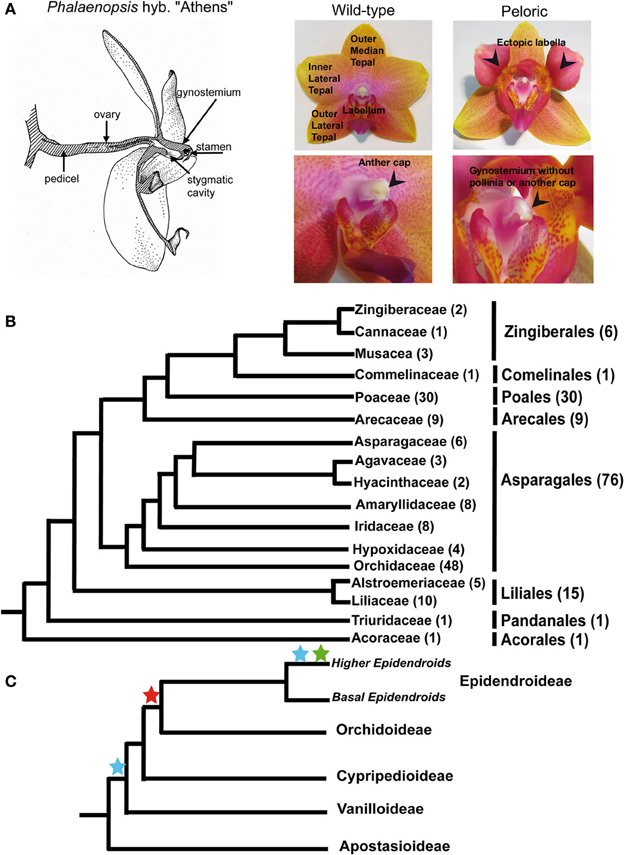
Figure 1. Phalaenopsis flower structure, distinctive floral features of wild-type and peloric hybrid “Athens.” Phylogenetic relationships the sequences studied. (A) Flower organs of wild-type and peloric mutant represented in the analysis. The stamen is located under the anther cap, a white laminar structure on top of the wild-type gynostemium (indicated with an arrow). The labella developing in place of inner lateral tepals as well as the organs missing in the gynostemium of peloric flowers are indicated with arrows. (B) Systematic relationships of the plant families represented in the phylogenetic analyses of MADS-box genes from monocots (based on Angiosperm phylogeny website version 12, www.mobot.org). The number sequences from every group in the dataset is indicated between brackets. (C) Systematic relationships of Orchidaceae subfamilies. The stars mark the points where the Orchidaceae subfamily composition of the gene trees suggests duplications might have occurred in SEP3-like genes (pale and dark blue), AG-like genes (red) and FUL-like II (green). The colors of the stars correspond to those employed to indicate these duplications in Figures 2–4.
The previously described features of the orchid perianth, together with the partial to complete suppression of three to five of the original six stamens are some of the major morphological transitions in the floral evolution of Orchidaceae (Bateman and Rudall, 2006). The developmental origins of these transitions, especially the ontogeny of the labellum and its role in orchid reproduction have been recurring topics in botany and evolutionary biology since the 19th century. The finding that MADS-box class B genes specify perianth and stamen identity in Arabidopsis put forward DEFICIENS- and GLOBOSA-like genes as candidates for experimentally addressing some of the key morphological transitions in orchid flower evolution (Mondragón-Palomino, 2013).
Several studies showed that in Orchidaceae there are four ancient lineages of DEF-like genes (Tsai et al., 2004; Mondragón-Palomino and Theißen, 2008, 2009). Their phylogeny served as a point of reference to compare the expression of DEF-like genes in wild-type representatives of most orchid subfamilies and flower terata. This analysis yielded the “orchid code,” a model where differential combinatorial expression of DEF-like genes from specific clades is associated with the development of each type of perianth organ: outer tepals, inner lateral tepals and labellum (Mondragón-Palomino and Theißen, 2008, 2011; Mondragón-Palomino, 2013). Namely, specification of labellum identity is associated with higher levels of mRNA from both clade three (PeMADS3-like) and clade four genes (PeMADS4-like) in this organ (Mondragón-Palomino and Theißen, 2011).
The ABCDE model already indicates several genes contribute to the specification of the identity of each flower organ. The involvement of several factors in flower development reflects the fact that MADS-domain transcription factors form higher order complexes (Egea-Cortines et al., 1999) that bind to CArG-box motifs in the regulatory regions of their targets. According to the floral quartet model (Honma and Goto, 2001; Theissen and Saedler, 2001), in Arabidopsis tetramers formed by AP1 and SEP3, determine sepal development, AP1, PI, AP3, and SEP are involved in the specification of petals while complexes of AP3, PI, SEP, and AG determine stamen identity and tetramers formed by AG and SEP dimers control carpel development.
Therefore, in order to realistically approach orchid flower development it is necessary to see beyond the “orchid code” and integrate information on the number and patterns of expression of additional candidate regulators of flower development. This information would enable approaching the development and evolution of gynostemium (colum) and ovary, organs that significantly contribute to the morphological diversity of the family. The orchid gynostemium is formed by the complete or partial union of male and female organs. This structure is often employed as diagnostic character in orchid systematics because of its highly complex species-specific combination of appendages as well as the position and characteristics of pollinia and anthers (Dressler, 1993; Rudall and Bateman, 2002) (Figure 1A).
The orchid ovary is inferior with respect to the bases of the perianth organs and formed by three carpels. In most orchids there are no divisions between carpels, but in genera from subfamilies Apostasioideae and Cypripedioideae the ovary has three locules (Dressler, 1993). Investigating MADS-box candidate SEP-, FUL-, AG-, and STK-like genes would also contribute to understanding the development of the carpel before and after pollination, an event that in Orchidaceae triggers ovary development.
Previous studies reported the expression of SEP-, FUL-, AG-, and STK-like genes in orchids (Reviewed in Mondragón-Palomino, 2013). According to this work the evolution of these genes is characterized by several instances of gene duplication as well as a conserved pattern of expression of each duplicate gene. Specifically, FRUITFULL-like genes are expressed mostly in the gynostemium and in some instances also in the perianth (Yu and Goh, 2000; Skipper et al., 2005; Chen et al., 2007; Chang et al., 2009) while AGAMOUS- and SEEDSTICK-like genes are expressed in gynostemium and ovary (Song et al., 2006; Xu et al., 2006; Hsu et al., 2010; Wang et al., 2011; Chen et al., 2012; Salemme et al., 2013). Most of the SEPALLATA-like genes isolated so far are expressed in all flower organs (Lu et al., 1993; Yu and Goh, 2000; Johansen and Frederiksen, 2002; Yu et al., 2002; Xu et al., 2006; Chang et al., 2009).
In the present study we isolated eight SEP-, FUL-, AG-, and STK-like genes from Phalaenopsis hyb. “Athens,” investigated their phylogenetic and orthology relationships and compared the patterns of expression in the perianth, column and developing organs of wild-type and peloric flowers with labella in place of inner lateral tepals and neither pollinia nor anther cap (Figure 1A). The ectopic labella are regarded as such because their shape, size, thickness, color and presence of calli are identical to those of the wild-type labellum.
The aims of this work are to investigate the association of additional MADS-box genes with the development of the labellum and pollinia, determine if paralogous SEP-, FUL-, AG-, and STK-like genes are also differentially expressed in the organs and stages investigated and in doing so contribute, in a way analogous to DEF-like genes, to the modularization of the orchid perianth. Although expression of some of the genes here investigated has been previously measured, not all studies include additional paralogs nor involve the simultaneous comparison of several MADS-box genes in homeotic organs or the developing ovary.
The results suggest a system of flower organ identity specification based on duplicate genes some of which are differentially expressed. We discuss the possible occurrence of dosage effects and their role in the preservation of these ancient duplicates. Because of the high prevalence of gene and genome duplication in plants, the origin and transcriptional divergence of orchid duplicate MADS-box genes reflect important processes shaping flower development, evolution and diversification in Angiosperms.
Materials and Methods
Plant Material
The origin and growth conditions of Phalaenopsis hybrid “Athens” (Epidendroideae) with wild-type or peloric flowers were described in Mondragón-Palomino and Theißen (2011). The organs of wild-type and peloric flower buds from of 0.9 to 1.0 cm in length were dissected, shock-frozen with liquid N2 and stored at −80°C.
RNA Isolation and cDNA Synthesis
Frozen flower organs and developing ovaries were individually ground with sterile steel beads. Total RNA was extracted with a buffer containing 8 M Guanidin-HCl, 20 mM MES, 20 mM EDTA pH 7,0. After a phenol, isoamyl alcohol, chloroform (25:24:1) extraction and centrifugation RNA was precipitated overnight with 1 N acetic acid and 100% ethanol. After centrifugation at 15,000 rpm for 10 min pellets were washed in 80% ethanol, dried by centrifugation and resuspended in 30–70 μ l RNase-free, sterile water. All steps were carried at 4°C.
Genomic DNA was removed with DNase I (RNase-free) from Fermentas (1 U/μ l, Fermentas, http://www.fermentas.com), following the manufacturer's protocol. Concentration, integrity and quality of total RNA were measured on an Agilent 2100 Bioanalyzer (Agilent Technologies, http://www.genomics.agilent.com).
Synthesis of cDNA was performed with 1 μ g of total DNAse-treated RNA from each sample with oligo (dT)15 AB05 as previously described (Mondragón-Palomino and Theißen, 2011). The quality of the resulting cDNA was verified by assessing the amplification of the internal reference genes Actin, Ubiquitin and EF1α of Phalaenopsis on an 1% agarose gel.
Primer Design
All primer pairs were designed with Primer3 v. 0.4.0 (http://frodo.wi.mit.edu/primer3). The sequences employed as target for primer design were identified in ML clade-specific alignments and phylogenies of orchid FUL-, AG-, STK-, and SEP-like genes previously described. The specificity of each primer pair was verified by the size of its amplicon in a 1% agarose gel. Annealing temperatures were obtained by gradient PCR with 1:5 diluted cDNA from flower buds or developing ovaries as template. The reactions were performed as described by the manufacturer (http://fermentas.com) in a Biometra TProfessional basic thermocycler gradient (http://www.biometra.de). Amplification efficiency (E) for each primer pair was calculated as previously described (Mondragón-Palomino and Theißen, 2011). Actin, Ubiquitin, and EF1α of Phalaenopsis were amplified with the corresponding primers from Mondragón-Palomino and Theißen (2011). All primers were ordered from biomers.net and their sequences are available in Supplementary Table I.
Quantitative Real-Time RT-PCR Assays
Assays were performed with Peqlab KAPA SYBR FAST qPCR Master Mix Universal (http://www.peqlab.com), in a Real-Time Thermo Cycler Realplex (Eppendorf, http://www.eppendorf.de). For each target gene and organ triplicate reactions with cDNA from each of two independent cDNA pools were performed. Samples were arranged in a 96-well plate according to the principle of maximization (Derveaux et al., 2010). In each plate were included a positive control (cDNA from flower buds), a non-template negative control (NTC) and two samples of DNase treated total RNA pooled from wild-type and peloric flowers to detect any genomic DNA contamination.
The quality of qPCR assay was assured by: sequencing the resulting products (Supplementary Figure 1), detection of a single amplicon of predicted size in a 1% agarose gel; a single specific peak in the melting curve of triplicate reactions, the cycle threshold (Cq) value of samples within a triplicate should not deviate by more than 0.50 cycle as well as on the validation of positive and negative controls. We employed the amplification conditions described in (Mondragón-Palomino and Theißen, 2011).
Normalization and Data Analysis
The Cq values of individual qPCR runs were exported to qBase plus v. 1.5 (Biogazelle, http://www.biogazelle.com) for further analysis. This program implements a modified ΔΔCt method (Hellemans et al., 2007) that takes the gene-specific amplification efficiencies calculated for each primer pair with standard curves. In this case, qBase plus employed the Cq values of EF1α, Ubiquitin, and Actin to generate a normalization factor. Normalization against three or more validated reference genes is considered most appropriate and universally applicable method (Vandesompele et al., 2002) and enables comparison with previous analysis of DEF- and GLO-like MADS-box gene expression in wild-type and peloric Phalaenopsis “Athens” (Mondragón-Palomino and Theißen, 2011).
The normalized quantities were rescaled relative to the sample with the lowest relative quantity, together with the corresponding standard errors they were exported to Excel v. 12.2.7 for Mac (Microsoft, http://www.microsoft.com) and to R v. 2.3.1 (R Foundation for Statistical Computing, http://www.r-projetic.org/foundation) to assess the correlation between samples from the same clade with Spearman's correlation test.
Sequences and Molecular Phylogenies
Keyword and BLAST searches in public databases (Supplementary Table I) with known MADS-box SEP-, FUL-, AG-, and STK-like genes from Zea mays and Oryza sativa returned 48 candidate orchid MADS-box sequences. To determine their evolutionary relationships and gene duplication history we performed computational alignments and inferred their relationships to other non-grass monocots. To support clade definition we included sequences of MADS-box genes from Oryza sativa and Zea mays (Supplementary Table II).
Sequences were obtained from across monocots, including Asparagales (76), Poales (30), and Liliales (15) (Figure 1B). The family best represented is Orchidaceae with 48 sequences, mostly from Phalaenopsis, Dendrobium, Oncidium, and Cymbidium, species of horticultural interest from the Epidendoideae, the largest and most derived orchid subfamily (Figure 1C).
Automated sequence alignment was carried out using the program Muscle v. 3.8.31 (Edgar, 2004) implemented in SeaView v. 4.3.5 (Gouy et al., 2010). All alignments were visually checked, manually improved and employed to obtain the corresponding nucleotide alignments.
Molecular phylogenies were inferred with Maximum Likelihood (ML) implemented in SeaView v. 4.3.5. ML analysis were performed with an optimized BioNJ tree as starting topology, a GTR model of substitution, aLRT (SH-like) as branch support, optimized invariables sites, optimized across-site rate variation. Tree searching operations took the best result from Nearest-Neighbor-Interchange and Subtree-Pruning-and-Regrafting.
The gene tree of FUL-like genes was rooted with a sample of basal angiosperm FUL-like genes which were previously employed for similar analyses (Pabón-Mora et al., 2013). SEP-like genes were rooted with a sample of SQUA-like genes from Angiosperms. In order to increase the resolution of both major clades SEP3- and SEP1, 2, and 4-like genes from basal Angiosperms were included. AG- and STK-like genes were rooted with Gymnosperms AG-like sequences following the example of (Kramer et al., 2004). All sequences employed for rooting and improving ingroup resolution are listed in Supplementary Table II.
Results
Orchidaceae-Specific Duplications Characterize the Phylogenies of SEP-, FUL-, AG-, and STK-Like Genes
Overall, the majority of the sequences from the same plant family form clades supported with probabilities larger than 0.95 (Figures 2–4). Although the phylogenetic relationships among these clades are topologically similar to those of their corresponding plant families (Figures 1B,C), they are often supported with probabilities smaller than 0.90.
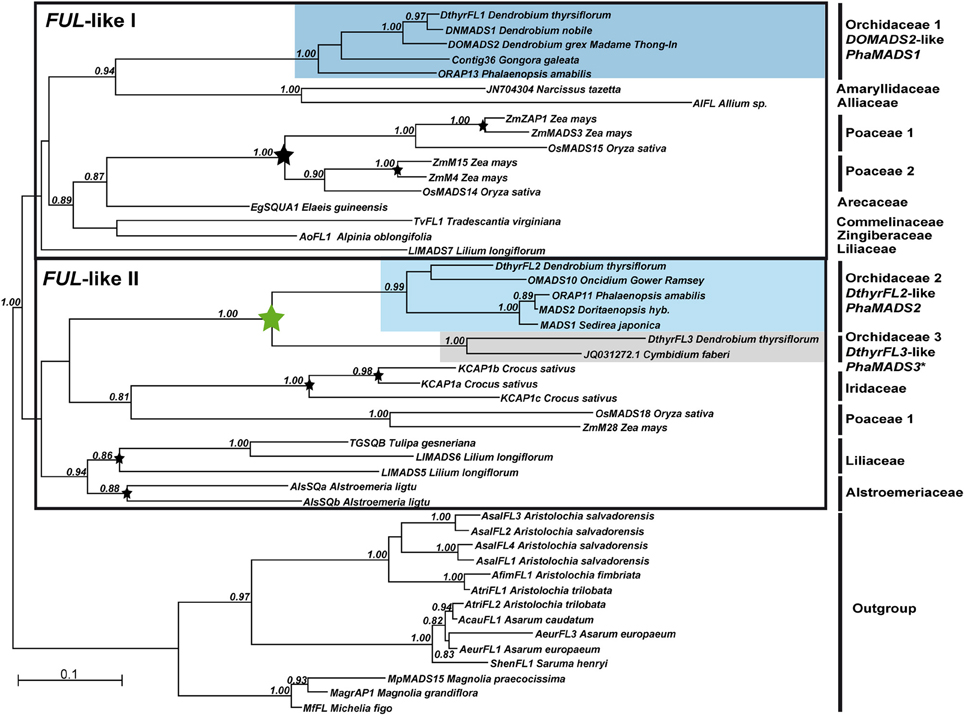
Figure 2. Maximum Likelihood phylogeny of monocot FUL-like genes. The two major monocot clades previously identified are indicated with a black frame (Litt and Irish, 2003). The clades containing sequences from Orchidaceae are highlighted if gene expression was characterized in Phalaenopsis hyb. “Athens.” Otherwise the non-isolated ortholog is marked with an asterisk. The numbers on every node indicate branch support above 0.80. The bars indicate clades of FUL-like genes from the plant families represented in the dataset as well as the outgroup. Black arrows point at the sequences employed as templates for qPCR primer design. Stars indicate gene duplications.
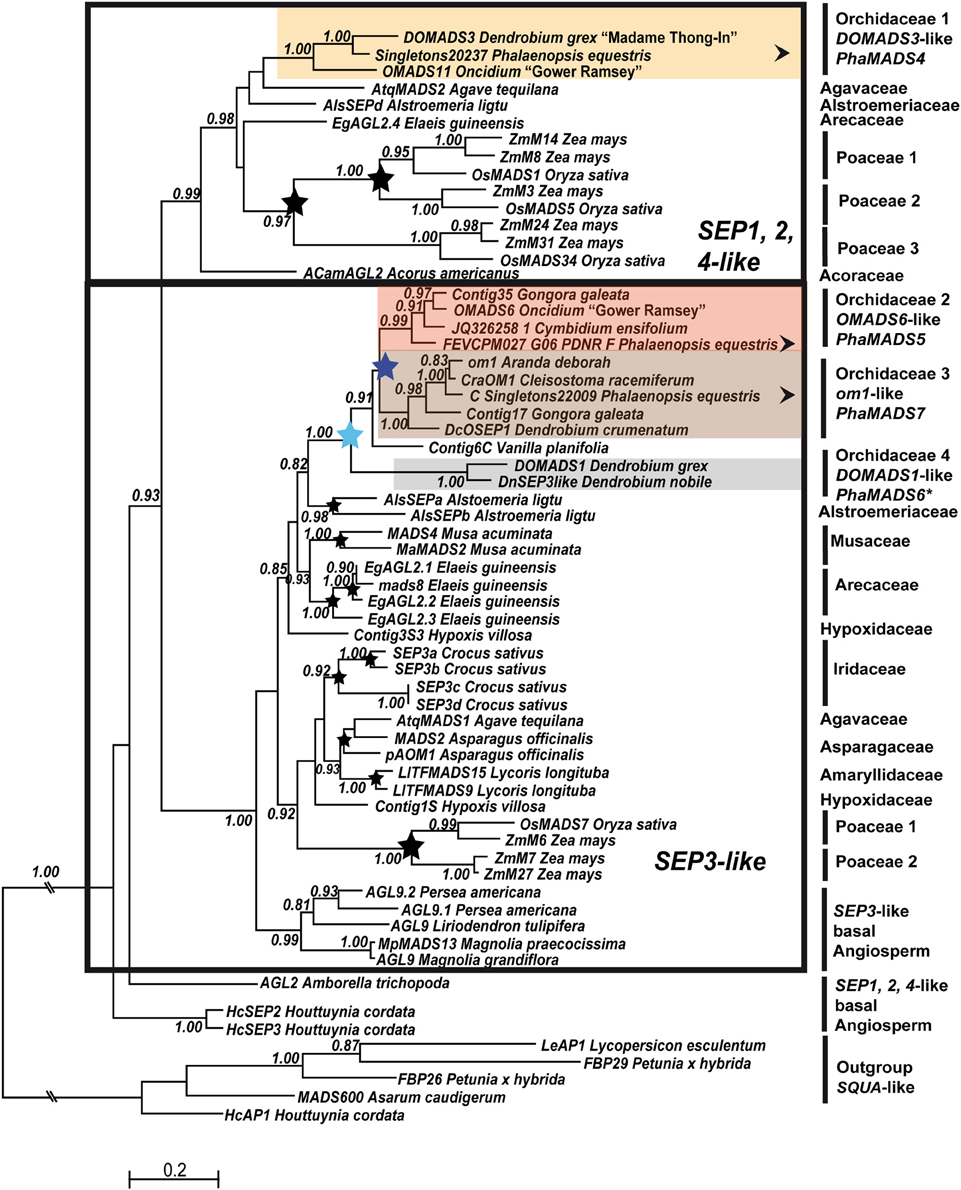
Figure 3. Maximum Likelihood phylogeny of monocot SEP-like genes. The two major monocot clades previously identified are indicated with a black frame (Zahn et al., 2005a). Orchidaceae-specific clades and other annotations are as described in Figure 2.
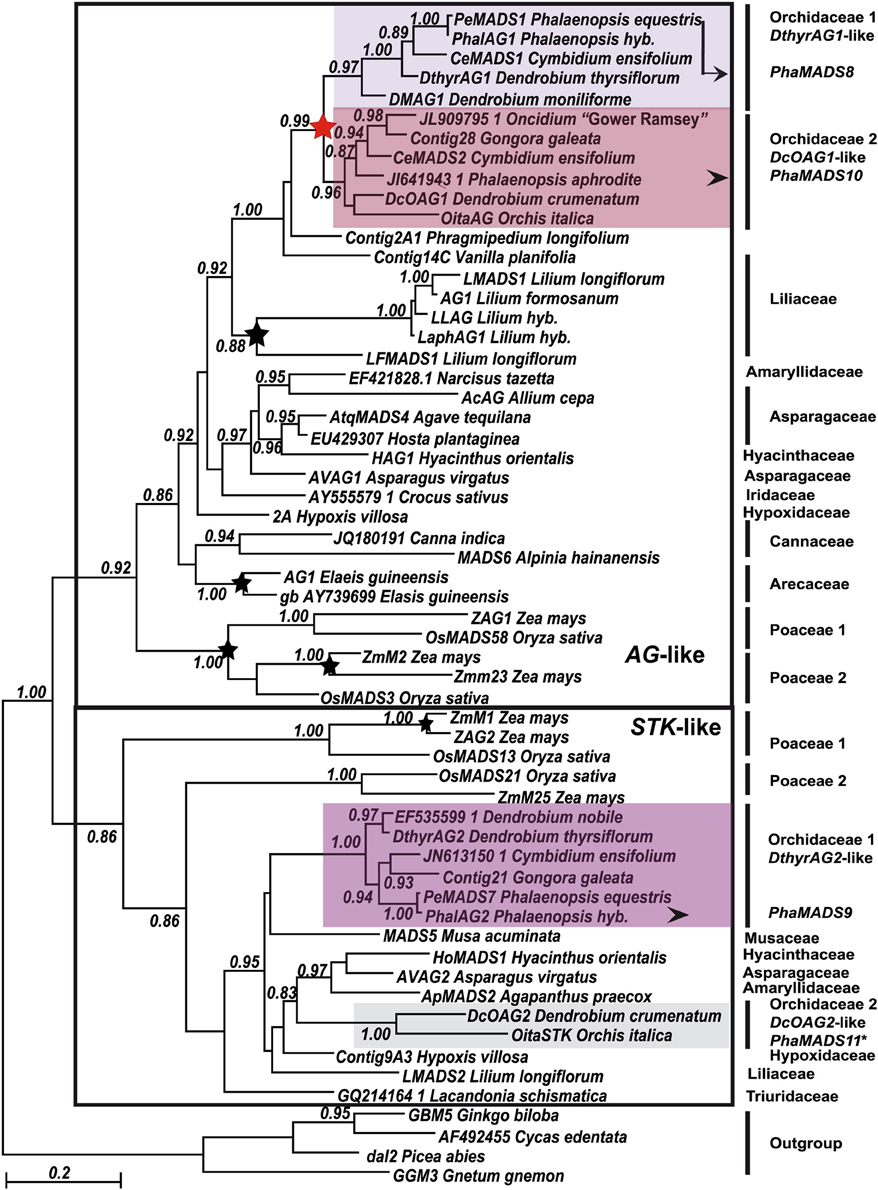
Figure 4. Maximum Likelihood phylogeny of monocot AG- and STK-like genes. Major monocot clades previously identified are indicated with a black frame (Kramer et al., 2004). Orchidaceae-specific clades and other annotations are as described in Figure 2.
Our phylogenetic analysis of monocot APETALA1/FRUITFULL genes identified the two major groups FUL-like I and FUL-like II previously inferred by Litt and Irish (2003). Despite employing a broader sample of monocot sequences, these groups are still unresolved as their low statistical support suggests. Notwithstanding, in both groups our analysis confirmed several grass- and species-specific gene duplications in Crocus, Lilium, and Alstroemeria. Furthermore in the FUL-like I subgroup there is a single, clade of orchid-specific DOMADS2-like genes clearly grouped with other genes belonging to species in Asparagales. In the FUL-like II genes there are two well-supported clades of DthyrFL2- and DthyrFL3-like sequences. However their relationship to other genes from species in Asparagales is not well-resolved (Figure 2).
While the C-terminal domain encoded by DthyrFL1 and DthyrFL2-like genes has the conserved FUL-like motif LPPWML, the sequences encoding DthyrFL3-like proteins have an early stop codon that eliminates the motif (Supplementary Figure 2A). As suggested by the alignment, the loss of this motif has already taken place in the common ancestor of Dendrobium and Cymbidium (Epidendroideae).
Similar changes in the region encoding the C-terminal domain caused by early stop codons and major deletions leading to loss of regions potentially involved in protein-protein interactions have been documented in OMADS3-like DEF-like genes form Orchidaceae (Mondragón-Palomino et al., 2009). The divergence of the C-terminal domains of both OMADS3- and DthyrFL3-like genes might imply their proteins adopted novel patterns of higher order molecular interaction (Supplementary Figure 2A) (Mondragón-Palomino et al., 2009).
In comparison with proteins like ZAP1 and ZmMADS3, which have C-terminal domains rich in glutamine and serine, the corresponding orchid FUL-like II proteins do not have similar long homopolymeric stretches. Because some of these repetitive sequences have been involved in transcriptional activation and associated to morphological evolution (Gerber et al., 1994; Lindqvist et al., 2007) their absence in orchid FUL-like proteins suggests divergence of their functional properties.
The analysis of SEP-like genes supports a division in two major clades: SEP1, 2, 4-like genes and SEP3-like genes. In monocots SEP1, 2, 4-like genes are characterized by four subclades, three of them [OsMADS1, OsMADS5, und RMADS217-like genes (OsMADS34)] are grass-specific (Zahn et al., 2005a) (Figure 3). In the single, non-resolved group containing non-grass species we identified a well-supported group of DOMADS3-like genes (Figure 3). Like other monocot lineages, the C-terminal domain of DOMADS3-like genes has a conserved SEP I motif, but also a rather divergent or missing SEP II motif (Supplementary Figure 2C).
In the analyses of (Zahn et al., 2005a), monocot SEP3-like genes are divided in three major groups, two of them being grass-specific. In the non-grass sequences we reproducibly identified several well-supported species-specific duplications in Alstroemeria, Musa, Eleais, Crocus, Asparagus, and Lycoris (Figure 3). Nonetheless, the node at the base of non-grass monocot group is supported with a probability lower than 0.8.
In Orchidaceae we identified two successive family-specific duplications (Figure 3). The earliest one involves the clade of DOMADS1-like genes and a group containing Vanilla planifolia Contig6C and the ancestor of OMADS6- and om1-like genes, which subsequently resulted from a second duplication event. The sequences in OMADS6- and om1-like genes have well-conserved SEP I and SEP II motifs while both DOMADS1-like genes have a divergent K-domain and a truncated C-domain (Supplementary Figure 2C). However, sequences from a broader sample of species are needed to precisely date the origin of these clades and the process behind their diversification.
In the case of monocot AG-like MADS-box genes, previous analyses reported three clades, two of them exclusively involving Poaceae-specific sequences and duplications (Kramer et al., 2004). The present analysis shows that in addition to the grass subclades there is a group of non-grass genes involving several species-specific duplications in Liliaceae and Arecaceae. Although the relationships among most non-grass sequences are statistically well supported, they do not always reproduce those of the plant groups where they belong.
Our analysis identified two clades of Orchidaceae-specific AG-like genes: DthyrAG1-like and DcOAG1-like (Figure 4). The relationship of both clades to sequences from Phragmipedium longifolium (Cypripedioideae) and Vanilla planifolia (Vanilloideae), two relatively basal orchid species (Figure 1B), suggests these clades might be the result of a relatively recent duplication in Epidendroideae or already in Orchidoideae (Figure 4). Furthermore, the analysis supports the relationship between the Orchidaceae clade and that of Liliaceae AG-like genes.
Previous analyses of monocot STK-like genes identified two sister clades of Poaceae-specific genes (Kramer et al., 2004). Outside of these groups the present analysis identified a poorly resolved group of non-grass genes. Among them, there are two clades of DthyrAG2- and DcOAG2-like genes from Orchidaceae (Figure 3). Although the relationship between both clades with each other and the rest of the sequences is not clear, they share conserved AGI and AGII motifs (Supplementary Figure 2B) and several Orchidaceae-specific substitutions, indels and in the case of DcOAG2, an early stop codon that eliminates the last seven amino acids of its C-terminal domain.
Patterns of Expression
FUL-like genes PhaMADS1 and PhaMADS2 are highly expressed in the ovary before pollination
Both FUL-like I PhaMADS1 and FUL-like II PhaMADS2 are transcribed at a relatively uniform and low level in perianth and gynostemium and are expressed at their highest level in the ovary before pollination.
The similarity in the patterns of expression of these paralogs in wild-type and peloric flowers is reflected by a correlation of 0.88 (Spearman's test) (Figure 5). At the level of individual organs a noteworthy difference is the 60% increase on the expression of PhaMAD2 in the gynostemium of peloric flowers (Figure 5). In addition, a low level of expression of PhaMADS1, but especially of PhaMADS2 was detected in leaves (Supplementary Figure 3).
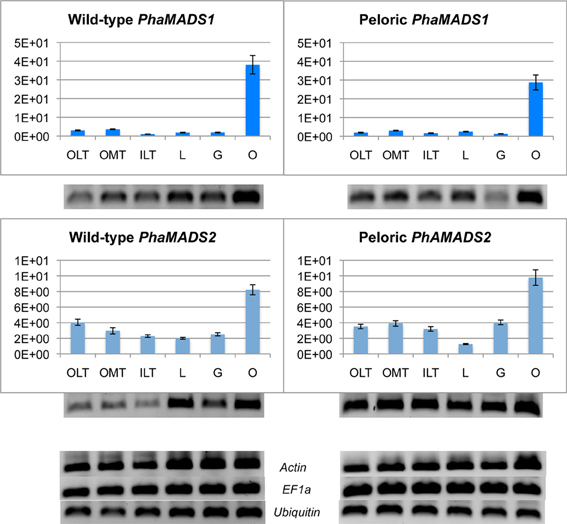
Figure 5. Normalized expression of orchid FUL-like genes. Expression of PhaMADS1 and PhaMADS2 in six flower organs of wild-type and peloric of Phalaenopsis hybrid “Athens.” OLT, outer lateral tepals; OMT, outer median tepals; ILT, inner lateral tepals; L, labellum; G, gynostemium; O, ovary. Expression of the target genes was normalized to the geometric average expression of three internal control genes: Actin, EF1α and Ubiquitin. Each column represents the expression obtained from six samples (three replicates from each of two different cDNA pools). The error bars represent the standard errors of the replicates. The y-axis is in arbitrary fluorescence units. The qPCR products from each sample series are presented below their corresponding columns in the graph.
In this study it was not possible to isolate the Phalaenopsis ortholog of DthyrFL3, representative of the second clade of FUL-like II genes (Figure 2). Because previous studies have not documented the expression of genes from this lineage in individual flower organs, it is not clear whether in addition to being expressed in developing ovules they are also associated with perianth formation (Skipper et al., 2005).
SEPALLATA-like gene PhaMADS7 is differentially expressed in inner lateral tepals and labellum
SEP-like genes PhaMADS4, PhaMADS5, and PhaMADS7 are expressed in all flower organs. Among the eight genes here measured the SEP3-like gene PhaMADS7 showed the largest expression differences between wild-type and peloric flowers: in the labellum-like inner lateral tepals its expression increased 196% and in the labellum 235% (Figure 6). In contrast, in the outer lateral tepals its expression increased 81%. The expression of PhaMADS5, the second SEP3-like gene measured, also increased in the outer median tepal, inner lateral tepals and labellum of peloric flowers, albeit at a lower level (65, 80, and 88% respectively) and decreased by 65% in the peloric gynostemium (Figure 6). The expression of SEP1-, 2, 4-like gene PhaMADS4 also showed an interesting increased of 130% in the peloric labellum (Figure 6).
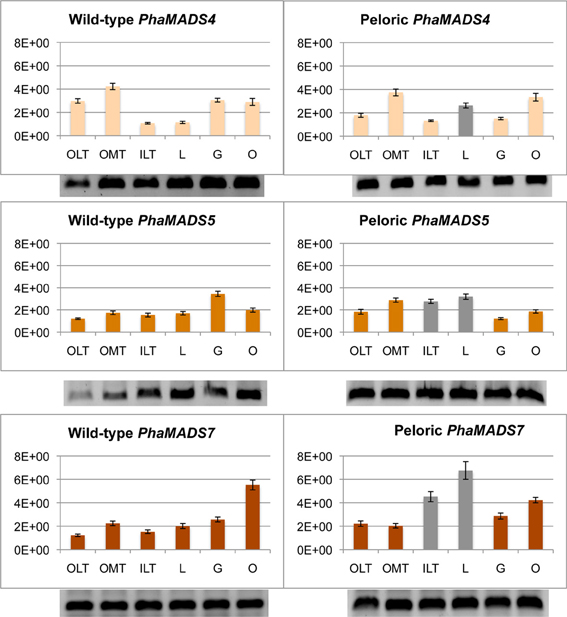
Figure 6. Normalized expression of orchid SEPALLATA-like genes. Expression of PhaMADS4 and PhaMADS5 and PhaMADS7 in six flower organs of wild-type and peloric of Phalaenopsis hybrid “Athens.” Replicates, normalization, graphics and abbreviations are as in Figure 5.
Furthermore, messengers of PhaMADS5 were detected in wild-type leaves and roots as well as in leaves of plants producing peloric flowers, while expression of PheMADS7 was detected in wild-type leaves (Supplementary Figure 3).
AGAMOUS-like genes PhaMADS8 and PhaMADS10 and STK-like gene PhaMADS9 are only expressed in gynostemium and ovary
In gynostemium and ovary there are relevant differences on the relative level of messengers from PhaMADS8 and PhaMADS9 (both AG-like genes), and PhaMADS10 (STK-like gene). While PhaMADS8 and PhaMADS10 are expressed twice as high in the gynostemium as in the ovary there is no significant difference in the expression of PhaMADS9 in those organs (Figure 7).
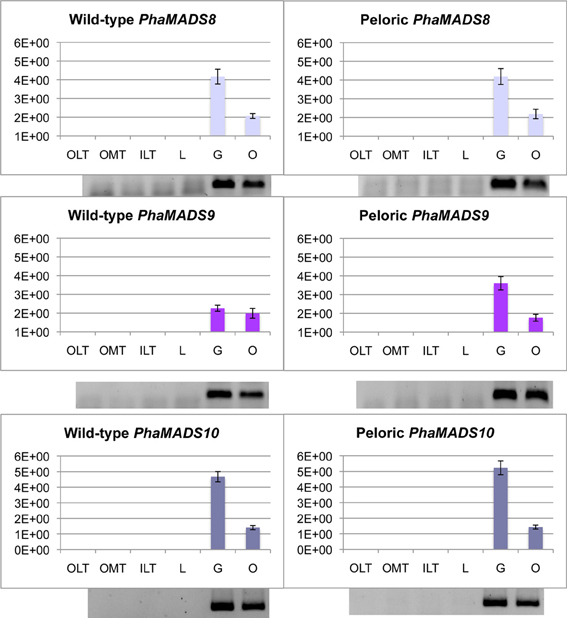
Figure 7. Normalized expression of orchid AGAMOUS- and SEEDSTICK-like genes. Expression of PhaMADS8, PhaMADS9 and PhaMADS10 in six flower organs of wild-type and peloric Phalaenopsis hybrid “Athens.” Replicates, normalization, graphics and abbreviations are as described in Figure 5.
PhaMADS9 is expressed 60% more in the peloric gynostemium as in the wild-type (Figure 7). In contrast, there are no significant changes in the levels of expression of PhaMADS8 and PhaMADS10 in the organs of wild-type and peloric flowers. Messengers for none of these three genes were detected in leaves and roots (Supplementary Figure 3).
Differential activity of FUL-, AG-, STK-, and SEP-like genes during ovary development in phalaenopsis
Expression of the eight SEP-, FUL-, AG-, and STK-like genes previously described was also measured in the developing ovary of Phalaenopsis hyb. 56 days after pollination (DAP) (Supplementary Figure 4), when ovule differentiation takes place after a phase of ovary growth and proliferation of ovule primordia (Zhang and O'neill, 1993). The significant differences we observed mostly involve the STK-like gene PhaMADS9, which expressed 5–27 times higher than the rest of the transcripts measured (Figure 8). The levels of expression in the ovary before and after pollination cannot be compared because they were measured in tissues of a different Phalaenopsis variety due to ovary abortion after self- or cross-pollination of Phalaenopsis hyb. “Athens.”
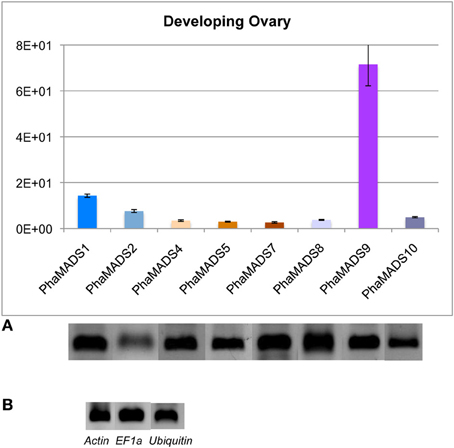
Figure 8. Normalized expression of orchid FUL-, SEP-, AG-, and STK-like MADS-box genes in developing ovary. (A) Normalized expression of MADS-box genes PhaMADS1, PhaMADS2 (FUL-like), PhaMADS4, PhaMADS5, PhaMADS7 (SEP-like), PhaMADS8, PhaMADS10 (AG-like) and PhaMADS9 (STK-like) in developing ovary of Phalaenopsis hybrid “Athens.” (B) Expression of three internal control genes (Actin, EF1α and Ubiquitin) in the developing ovaries. Replicates, normalization, graphics and abbreviations are as described in Figure 5.
Discussion
Orchidaceae Duplicate MADS-Box Genes have been Retained for Millions of Years
Because of the high representation of Orchidaceae genes in the phylogenies, we identified several well-supported, family-specific duplications which broaden the previously identified monocot MADS-box groups FRUITFUL- (FUL-), AGAMOUS- (AG-), SEEDSTICK- (STK-), and SEPALLATA-like (SEP-) genes, (Litt and Irish, 2003; Kramer et al., 2004; Zahn et al., 2005a, 2006). Because the sample of MADS-box genes from Orchidaceae species is rather biased toward subfamily Epidendroideae the conclusions drawn from this dataset regarding number of paralogs and date of duplication might turn out to be different as a broader dataset becomes available.
The phylogenetic relationships identified within the clades of FUL-like DOMADS2-like genes and DtyhrFL2-like genes are consistent with previous phylogenetic analyses (Chen et al., 2007). Because genes from Orchidaceae are present in both FUL-like I and FUL-like II monocot clades our analysis suggests these groups originated earlier in Angiosperm evolution than initially dated (Litt and Irish, 2003). The phylogenetic analysis here presented reproduces recent studies involving Orchidaceae AG-/STK- (Salemme et al., 2013) and describes the occurrence of two duplications in Orchidaceae SEP3-like genes.
The groups of duplicate Orchidaceae SEP-, FUL-, AG-, and STK-like genes are ancient as suggested by their origin at different points in the history of Orchidaceae (Figure 1C). The molecular phylogenies suggest the duplication generating OMADS6- and om1-like SEP3 genes (Figure 3) probably took place after the divergence of family Vanilloideae, at least 62 million years ago (MYA) (Ramirez et al., 2007). In contrast, the duplication of DthyrAG1- and DcOAG1-like AG genes is relatively more recent, probably involving the ancestor of subfamily Orchidoideae, about 56 MYA (Figure 4). Genes in all other clades are already present in several species from the so-called “higher” Epidendroids, whose origin dates back to 54 MYA (Ramirez et al., 2007). As more data becomes available, it will become clearer whether these apparently recent paralogs are Epidendroideae-specific or more ancient (Figure 1C).
The mechanisms behind the retention of these paralogs are possibly associated with dosage effects. Assuming the different levels of messengers result in distinct amounts of protein products, it is foreseeable their concentration plays a role in the formation of regulatory protein complexes and eventually the activation of target genes. In this context, the biased retention of Phalaenopsis duplicate MADS-box genes is feasibly explained by the gene balance hypothesis, which states duplicated gene retention following a Whole Genome Duplication (WGD) would avoid the harmful consequences of dosage imbalance among interacting proteins. Dosage effects are already implied by the association of different levels of DEF-like genes PeMADS3 and PeMADS4 and the development of inner lateral tepals and labellum (Mondragón-Palomino and Theißen, 2011).
Expression of SEP-, FUL-, AG-, and STK-Like Genes in Phalaenopsis
The two-fold increase of PhaMADS7 (SEP3-like) expression in the inner lateral tepal and the labellum of peloric flowers suggests it might have a distinct role in the development of the inner perianth. These aspects will be discussed in “A transcriptional model for Phalaenopsis flower development.”
Our results agree with previous work reporting SEP-like genes from orchids are expressed in all flower organs during development, in a way analogous to SEP1, SEP2, and SEP3 from Arabidopsis thaliana (Lu et al., 1993; Yu and Goh, 2000; Johansen and Frederiksen, 2002; Yu et al., 2002; Xu et al., 2006; Chang et al., 2009) (Summarized in Supplementary Table III). Based on these studies it has been argued that in orchids SEP-like genes are involved in floral transition and flower organ identity specification.
Our findings in Phalaenopsis also agree with the fact in other non-Poales monocots like Asparagus officinalis (dioecious, Asparagaceae), Eleais guineensis (monoecious Arecaceae) and Musa acuminata (dioecious, Musaceae) SEP-like genes are expressed in the inflorescence, in all flower organs and flower meristems and thus might be involved in their development (Caporali et al., 2000; Tzeng et al., 2003; Adam et al., 2007; Tsaftaris et al., 2011) as well as in the differences between male and female flowers of Musa acuminata (Elitzur et al., 2010) and Agave tequilana (Agavaceae) (Delgado Sandoval et al., 2011).
The domains of expression of FUL-like genes PhaMADS1 and PhaMADS2 were generally similar to those of their orthologs in other orchid species (Yu and Goh, 2000; Skipper et al., 2005; Chen et al., 2007; Chang et al., 2009) (summarized in Supplementary Table IIIA). However they differ with previous studies in that both genes are also expressed in the perianth organs (Figure 5). This might be due to the fact gene expression was measured in developing tepals where distinctive features like shape, color and appendages are not yet defined while DOMADS2 and OMADS10, ORAP11, and ORAP13 were measured in fully-developed tepals (Yu and Goh, 2000; Chen et al., 2007; Chang et al., 2009) (Summarized in Supplementary Table III).
The relatively high level of expression of PhaMADS1 and PhaMADS2 in the ovary agrees with previous studies suggesting that monocot FUL genes might have a role fruit development in a way analogous to Arabidopsis' FRUITFULL rather than to AP1 (Litt and Irish, 2003). In accordance to this, the sequences of PhaMADS1 and PhaMADS2 like all members of the AP1/FUL, AGL6, and SEP gene subfamilies share a conserved, C-terminal, hydrophobic FUL-like and the M/LPPGWLA SEPII motives (Supplementary Figures 2A,B respectively) (Litt and Irish, 2003; Zahn et al., 2005a).
In Arabidopsis FRUITFULL is key to fruit morphogenesis after fertilization by mediating elongation and cell differentiation within fruit valve layers (Gu et al., 1998). Because orchid ovary development starts after fertilization, it is intriguing that PhaMADS1 and PhaMADS2 are highly expressed before this event, thus implying unknown aspects of ovary cell differentiation might have an earlier starting point (Figure 5).
Although sequence similarity with FUL does not explain the expression of both genes in all four whorls, it is a characteristic shared with other non-grass monocots like: Crocus (Iridaceae), Tradescantia (Commelinaceae), Lilium (Liliaceae), Agapanthus (Agapanthaceae), and Elaeis (Arecaceae) (Tsaftaris et al., 2004; Preston and Kellogg, 2006; Adam et al., 2007). In our view the patterns observed support the notion that in non-grass monocots expression of FUL-like genes in all four floral whorls is the ancestral state (Preston and Kellogg, 2006) and suggests in these species FUL-like genes might also play a role in the development of all flower organs. Nonetheless a genetic definition of class A function in non-grass monocots would require analysis of mutants or specific silencing of each paralog.
The expression of PhaMADS8, PhaMADS10 (both AG-like) and PhaMADS9 (STK-like) in gynostemium and ovary agrees with most previous transcriptional characterizations of AG- and STK-like genes in orchids (Skipper et al., 2006; Song et al., 2006; Xu et al., 2006; Hsu et al., 2010; Wang et al., 2011; Chen et al., 2012; Salemme et al., 2013) (Summarized in Supplementary Table III). Exceptionally, and probably due to differences in the developmental stages analyzed, messengers for AG-like genes DcOAG1, CeMADS2, and OitaAG as well as STK-like gene OitaSTK are also detected in perianth organs (Xu et al., 2006; Wang et al., 2011), particularly in stages closer to anthesis (Salemme et al., 2013).
The highly similar patterns of expression of duplicate AG- and STK-like genes suggest they are redundantly involved in the development of gynostemium and ovary in the stages investigated. Nevertheless recent studies on multitepal (Cymbidium ensifolium) and glyp (Phalaenopsis) might suggest otherwise. In multitepal the gynostemium is replaced by an ectopic flower which produces outer and inner tepal-like structures centripetally (Wang et al., 2011), in a way analogous to agamous from A. thaliana. In the wild-type, AG-like genes CeMADS1 (ortholog of PhaMADS8) and CeMADS2 (ortholog of PhaMADS10) are strongly expressed in the gynostemium while CeMADS2 is weakly expressed in the perianth organs. However, CeMADS1 is not expressed in the gynostemiumless buds of the multitepal mutant and CeMADS2 remains weakly expressed in the perianth (Wang et al., 2011). Further analysis is needed to determine whether the level of expression reported supports a role in perianth development.
In agreement with our results PeMADS1 (ortholog of PhaMADS8) and PeMADS7 (ortholog of PhaMADS9) are expressed in ovary and gynostemium of Phalaenopsis equestris (Chen et al., 2012). However they might play different roles in the development of these structures as suggested by the differential expression of PeMADS1 in the gynostemium-like inner lateral tepals of the glyp mutant of Phalaenopsis hyb. “CD1” (Chen et al., 2012).
Further analysis of gynostemium and ovary mutants is required to discern the specific roles in column development of paralogous orchid AG- and STK-like genes.
In agreement previously described work in orchids, AG-like genes in other non-grass monocots, like LLAG1, HAG1, AcAG, CsAG1a, and AtqMADS4 from Lilium longiflorum, Hyacinthus orientalis, Allium cepa, Crocus sativus, and Agave tequilana respectively, are expressed in stamens and carpels, suggesting a role in the development of these organs by analogy to the domains of expression of AGAMOUS in the ABC model (Li et al., 2002; Benedito et al., 2004; Tsaftaris et al., 2005; Hsu et al., 2010; Delgado Sandoval et al., 2011; Li et al., 2013).
In contrast to the expression of orchid STK-like genes in gynostemium and ovaries (Skipper et al., 2006; Song et al., 2006; Xu et al., 2006; Hsu et al., 2010; Chen et al., 2012; Salemme et al., 2013) (Supplementary Table III), non-grass monocots genes LMADS2 (Lilium longiflorum), HoMADS1 (Hyacinthus orientalis), and LsSTK (Lacandonia schismatica) are expressed exclusively in the carpel, mainly in the ovules (Tzeng, 2002; Xu et al., 2004; Alvarez-Buylla et al., 2010). In Orchidaceae expression in the gynostemium is probably due to adnation of stamens and style.
A Transcriptional Model for Phalaenopsis Flower Development
The classic ABC model of flower organ identity specification was based on genetic analysis of single and multiple mutants affecting distinct groups of flower organs in Antirhinum majus and Arabidopsis thaliana. Although initially the genes behind the phenotypes observed were not known, it was possible to employ the tools of classical genetics to understand how combinations of different functions affected the development of specific organs in particular whorls (Bowman et al., 1991).
Although a similar approach is not yet feasible in orchids, an initial, informative approach is associating the patterns of expression of DEF-, GLO- SEP-, FUL-, AG-, and STK-like MADS-box genes in wild-type and mutant flowers with the development of particular organs. The resulting patterns are the basis of models on evolution and development that later on can be genetically evaluated.
In the following paragraphs we propose how the genes profiled would be involved in the specification of distinct flower organ identities in Phalaenopsis (Summarized in Figure 9).
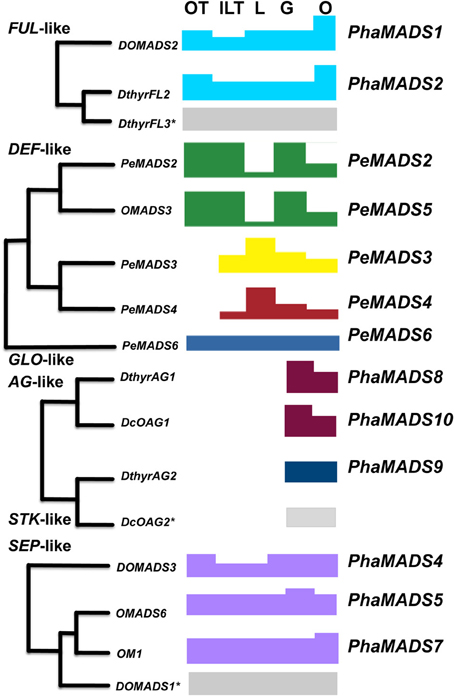
Figure 9. A transcriptional model for Phalaenopsis flower development. Summary of expression patterns of orchid paralogous FUL-, DEF-, GLO-, AG-, STK-, and SEP-like genes in Phalaenopsis perianth and reproductive organs. The combined differential activity and levels of expression of genes from different clades in each organ are represented with blocks of different colors and mapped on their domains of expression in the perianth, column and ovary. Genes corresponding to a clade with an asterisk were isolated and measured in other studies and their pattern of expression is included here for completion.
Outer tepal identity: DEF-, GLO-, SEP-, and FUL-like genes
In this system of organ identity specification outer tepals would be defined by the differential expression of DEF-like genes PeMADS2 (clade 1) and PeMADS5 (clade 2) together with PeMADS6 (GLO-like gene) (Mondragón-Palomino and Theißen, 2011) (Figure 9). SEP-like genes would contribute to the specification of this whorl as they do in the case of Arabidopsis thaliana and rice (Pelaz et al., 2000; Cui et al., 2010). Because of its higher level of expression we assume PhaMADS4 (SEP1, 2, 4-like) makes a larger contribution to organ identity determination in this whorl (Figures 6, 9).
The activity of the three FUL-like genes identified might be relevant for the development of the outer perianth. However, a floral mutant affected on the first whorl must first be analyzed to determine their specific role.
Inner lateral tepals and labellum identity is associated with differential activity of specific DEF- and SEP3-like genes
The increase in expression of SEP3-like gene PhaMADS7 in the peloric inner perianth suggests an association between its differential expression (Figure 6) and the development of the inner lateral tepals and labellum. Most importantly, the highest levels of expression of this gene in peloric flowers overlap with those of PeMADS3 and PeMADS4, two DEF-like genes, which in a similar way are highly expressed in peloric inner-lateral tepals and wild-type labellum and therefore have been associated to the development of this organ (Mondragón-Palomino and Theißen, 2011) (Figure 9). Because the pattern of differential expression of SEP3-like gene PhaMADS7 increased in the peloric inner perianth in a way analogous to that of PeMADS3 and PeMADS4 it is feasible their domains of expression are determined by a common upstream regulatory gene whose activity reflects adaxial-abaxial positional cues on the flower meristem.
The feasibility of regulatory protein-protein interactions between the transcription factors encoded by PhaMADS7 (SEP3-like) and both DEF- and GLO-like genes has been experimentally documented in orchid Dendrobium crumenatum between DcOAP3B, DcOPI, and DcOSEP1, product of the ortholog of PhaMADS7. These complexes are analogous to those formed in Arabidopsis thaliana by class B proteins AP3 and PI with SEP3, their most abundant interaction partner (Smaczniak et al., 2012). These proteins together with AP1 or AG are key in the specification of petal and stamen identity, respectively (Honma and Goto, 2001; Theissen and Saedler, 2001).
Stamen development in the gynostemium
In Arabidopsis thaliana MADS-box gene classes B, C, and E determine the development of stamens. Our results suggest the abortion of stamen development might be associated to the important change of expression of PeMADS2 (DEF-like, clade 2) in the peloric gynostemium (Mondragón-Palomino and Theißen, 2011) as well as the 65% decrease in the expression of SEP3-like gene PhaMADS5 (Figure 6), which encodes a potential interaction partner for PeMADS2. In contrast, there are not changes in the expression of AG-like genes PhaMADS8 and PhaMADS10 (Figure 7).
Ovary and ovule development: SEP-, FUL-, AG-, and STK-like genes
The functional conservation of the genes controlling ovary and ovule development in eudicots and monocots (Favaro et al., 2002, 2003; Dreni et al., 2007; Cui et al., 2010), suggests PhaMADS9 (STK-like), encoded by the gene most highly expressed in the ovary after pollination, might play a role in ovule identity determination, by forming complexes with the products of co-expressed PhaMADS8 and PhaMADS10 (both AG-like) as well as PhaMADS4, PhaMADS5 and PhaMADS7 (SEP-like) genes (Figure 9).
Conclusions and Perspectives
Clearly the dataset presently available for phylogenetic reconstruction of MADS-box gene evolution in non-grass monocots must still be broadened beyond the species represented in this study to enable detailed inference of the number and age of paralogs in each group of genes as well as comparative analysis of molecular orchid evo-devo.
In this context, our study in Phalaenopsis hyb. “Athens” suggests, albeit a few exceptions, that the MADS-box genes investigated generally share their patterns of expression with other non-grass monocots. A key difference however, is that for a fraction of Orchidaceae, and especially for Phalaenopsis it is already clear each group of genes is represented by two or more paralogs (Figures 2–4, 9). The joint expression of these paralogs hints at a system of regulatory activities determined by the differential expression of some of them in specific domains (Figure 9).
Concretely, the profiles obtained suggests flower organ identity results from the activity of multiple groups of duplicate FUL-, DEF-, AG-, STK-, and SEP-like genes. Orchid flower morphology might then be to a great extent the result of an extended developmental “toolkit” that in the course of evolution enabled a complex network of regulatory interactions with a broader, organ-specific group of downstream targets. Previous analysis of the patterns of expression in orchid DEF-like MADS-box genes suggested the subfunctionalization of developmental paralogs might increase the genotypic modularity of organisms by generating novel domains of expression (Mondragón-Palomino and Theißen, 2008). Once duplicate genes acquired different domains and levels of expression, they also might attain particular groups of interaction partners and downstream targets. The resulting developmental modules (e.g., the labellum or the gynostemium) might then respond independently to natural selection (e.g., pollinators) eventually giving rise to morphologically distinct flower organs.
The prevalent occurrence of gene duplication has major consequences for the regulation of orchid flower development and requires addressing the following topics:
(a) Positional cues defining gene expression
While FUL, SEP-, and GLO-like genes seem to be expressed in all flower organs, DEF-, AG-, and STK-like genes have domains of expression in specific perianth or reproductive organs (Figure 9). This difference suggests the expression of certain genes is sensitive to specific positional cues. Understanding their nature and mechanisms of action is key to learning how duplicates diverged transcriptionally and became associated to the development of particular organs.
(b) Functional redundancy
In order to clarify whether AG-, FUL-, and SEP3-like paralogs are transcriptionally redundant it would be necessary to investigate a broader array of developmental stages and tissues with approaches like RNA-seq on microdissected tissues. On the other hand, classic genetic analysis or genetic transformation (transient or stable) of at least one from the recently advanced model species (reviewed in Mondragón-Palomino, 2013) would offer a more direct approach to their actual function or functions.
(c) Role of C-terminal domain variation in protein-protein interactions
In Orchidaceae FUL-, DEF-, AG-, STK-, and SEP-like genes there is one clade where the sequence of the C-terminal domain substantially diverges from all others investigated (Supplementary Figure 2 and (Mondragón-Palomino et al., 2009)). It remains to be elucidated whether the proteins encoded by the members of these distinct lineages modify the formation of regulatory higher order interactions.
Author Contributions
Roberta Acri-Nunes-Miranda collected plant material, performed experiments, analyzed data, prepared illustrations and drafted the manuscript. Mariana Mondragón-Palomino designed the study, analyzed data, prepared illustrations and wrote manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank all reviewers for their valuable comments which helped us improve the manuscript. Mariana Mondragón-Palomino is funded by a stipend from the “Programm zur Realisierung der Chancengleichheit für Frauen in Forschung und Lehre.”
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fpls.2014.00076/abstract
Supplementary Figure S1 | Sequences of qPCR amplicons. Amino acid translation of the sequences used for qPCR primer design and the corresponding sequences of the products amplified with such primers. Due to sequencing the amplicon size is not identical to the one given in Supplementary Table II.
Supplementary Figure S2 | C-terminal domains of FUL-, AG/STK and SEP-like genes from Orchidaceae. Sequences are grouped according to the clades in the corresponding phylogenies. Framed in black are conserved sequence motifs mentioned in the text. Color-coding indicates the biochemical properties of each amino acid.
Supplementary Figure S3 | Expression of orchid FUL-, SEP-, AG-, and STK-like MADS-box genes in leaves and roots. (A) Expression of seven MADS-box genes in leaves (L) and roots (R) of Phalaenopsis hybrid “Athens” wild-type and peloric. C stands for control amplification with wild-type flower bud cDNA. (B) Expression of three internal control genes (Actin, Ubiquitin, and EF1α) in the cDNA pools from leaves and roots.
Supplementary Figure S4 |Transverse sections of developing ovary from Phalaenospsis hyb. 56 DAP. (A) Distal section of monocarpellate ovary with parietal placentation. (B) Proximal section (near to the pedicel). P and O indicate placenta and ovules, respectively.
Supplementary Table I | Characteristics and sequences of the primers pairs employed for qPCR.
Supplementary Table II | List of sequences employed in alignments, phylogenetic analysis and primer design.
Supplementary Table III | Expression of FUL-, AG/STK and SEP-like MADS-box genes from Orchidaceae.
References
Adam, H., Jouannic, S., Orieux, Y., Morcillo, F., Richaud, F., Duval, Y., et al. (2007). Functional characterization of MADS box genes involved in the determination of oil palm flower structure. J. Exp. Bot. 58, 1245–1259. doi: 10.1093/jxb/erl263
Alvarez-Buylla, E. R., Ambrose, B. A., Flores-Sandoval, E., Englund, M., Garay-Arroyo, A., Garcia-Ponce, B., et al. (2010). B-Function expression in the flower center underlies the homeotic phenotype of Lacandonia schismatica (Triuridaceae). Plant Cell 22, 3543–3559. doi: 10.1105/tpc.109.069153
Alvarez-Buylla, E. R., Pelaz, S., Liljegren, S. J., Gold, S. E., Burgeff, C., Ditta, G. S., et al. (2000). An ancestral MADS-box gene duplication occurred before the divergence of plants and animals. Proc. Natl. Acad. Sci. U.S.A. 97, 5328–5333. doi: 10.1073/pnas.97.10.5328
Arditti, J. (2002). “Resupination,” in Proceedings of the 17th World Orchid Conference, eds J. Arditti and H. Nair (Borneo: Natural History Publications), 111–121.
Bateman, R. M., and Rudall, P. J. (2006). “The good, the bad and the ugly: using naturally occurring terata to distinguish the possible from the impossible in orchid floral evolution,” in Monocots: Comparative Biology and Evolution. Excluding Poales, eds J. T. Columbus, E. A. Friar, J. M. Porter, L. M. Prince, and M. G. Simpson (Claremont, CA: Rancho Santa Ana Botanical Garden), 481–496.
Becker, A., and Theissen, G. (2003). The major clades of MADS-box genes and their role in the development and evolution of flowering plants. Mol. Phyl. Evol. 29, 464–489. doi: 10.1016/S1055-7903(03)00207-0
Benedito, V. A., Visser, P. B., Van Tuyl, J. M., Angenent, G. C., De Vries, S. C., and Krens, F. A. (2004). Ectopic expression of LLAG1, an AGAMOUS homologue from lily (Lilium longiflorum Thunb.) causes floral homeotic modifications in Arabidopsis. J. Exp. Bot. 55, 1391–1399. doi: 10.1093/jxb/erh156
Bowman, J. L., Smyth, D. R., and Meyerowitz, E. M. (1991). Genetic interactions among floral homeotic genes of Arabidopsis. Development 112, 1–20.
Caporali, E., Spada, A., Losa, A., and Marziani, G. (2000). The MADS box gene AOM1 is expressed in reproductive meristems and flowers of the dioecious species Asparagus officinalis. Sex. Plant Rep. 13, 151–156. doi: 10.1007/s004970000049
Chang, Y. Y., Chiu, Y. F., Wu, J. W., and Yang, C. H. (2009). Four orchid (Oncidium ‘Gower Ramsey’) AP1/AGL9-like MADS box genes show novel expression patterns and cause different effects on floral transition and formation in Arabidopsis thaliana. Plant Cell Physiol. 50, 1425–1438. doi: 10.1093/pcp/pcp087
Chen, D., Guo, B., Hexige, S., Zhang, T., Shen, D., and Ming, F. (2007). SQUA-like genes in the orchid Phalaenopsis are expressed in both vegetative and reproductive tissues. Planta 226, 369–380. doi: 10.1007/s00425-007-0488-0
Chen, Y. Y., Lee, P. F., Hsiao, Y. Y., Wu, W. L., Pan, Z. J., Lee, Y. I., et al. (2012). C- and D-class MADS-box genes from Phalaenopsis equestris (Orchidaceae) display functions in gynostemium and ovule development. Plant Cell Physiol. 53, 1053–1067. doi: 10.1093/pcp/pcs048
Coen, E. S., and Meyerowitz, E. M. (1991). The war of the whorls: genetic interactions controlling flower development. Nature 353, 31–37. doi: 10.1038/353031a0
Cui, R., Han, J., Zhao, S., Su, K., Wu, F., Du, X., et al. (2010). Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). Plant J. 61, 767–781. doi: 10.1111/j.1365-313X.2009.04101.x
Delgado Sandoval, S. D. C., Abraham Juárez, M. J., and Simpson, J. (2011). Agave tequilana MADS genes show novel expression patterns in meristems, developing bulbils and floral organs. Sex. Plant Rep. 25, 11–26. doi: 10.1007/s00497-011-0176-x
Derveaux, S., Vandesompele, J., and Hellemans, J. (2010). How to do successful gene expression analysis using real-time PCR. Methods 50, 227–230. doi: 10.1016/j.ymeth.2009.11.001
Dreni, L., Jacchia, S., Fornara, F., Fornari, M., Ouwerkerk, P. B. F., An, G., et al. (2007). The D-lineage MADS-box gene OsMADS13 controls ovule identity in rice. Plant J. 52, 690–699. doi: 10.1111/j.1365-313X.2007.03272.x
Dressler, R. L. (1993). Phylogeny and Classification of the Orchid Family. Portland, OR: Discorides Press.
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucl. Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Egea-Cortines, M., Saedler, H., and Sommer, H. (1999). Ternary complex formation between the MADS-box proteins SQUAMOSA, DEFICIENS and GLOBOSA is involved in the control of floral architecture in Antirrhinum majus. EMBO J. 18, 5370–5379. doi: 10.1093/emboj/18.19.5370
Elitzur, T., Vrebalov, J., Giovannoni, J. J., Goldschmidt, E. E., and Friedman, H. (2010). The regulation of MADS-box gene expression during ripening of banana and their regulatory interaction with ethylene. J. Exp. Bot. 61, 1523–1535. doi: 10.1093/jxb/erq017
Favaro, R., Immink, R., Ferioli, V., Bernasconi, B., Byzova, M., Angenent, G., et al. (2002). Ovule-specific MADS-box proteins have conserved protein-protein interactions in monocot and dicot plants. Mol. Gen. Genomics 268, 152–159. doi: 10.1007/s00438-002-0746-6
Favaro, R., Pinyopich, A., Battaglia, R., Kooiker, M., Borghi, L., Ditta, G., et al. (2003). MADS-box protein complexes control carpel and ovule development in Arabidopsis. Plant Cell 15, 2603–2611. doi: 10.1105/tpc.015123
Gerber, H. P., Seipel, K., Georgiev, O., Höfferer, M., Hug, M., Rusconi, S., et al. (1994). Transcriptional activation modulated by homopolymeric glutamine and proline stretches. Science 263, 808–811. doi: 10.1126/science.8303297
Gouy, M., Guindon, S., and Gascuel, O. (2010). SeaView version 4: a multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 27, 221–224. doi: 10.1093/molbev/msp259
Gu, Q., Ferrándiz, C., Yanofsky, M. F., and Martienssen, R. (1998). The FRUITFULL MADS-box gene mediates cell differentiation during Arabidopsis fruit development. Development 125, 1509–1517.
Hellemans, J., Mortier, G., De Paepe, A., Speleman, F., and Vandesompele, J. (2007). qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 8:R19. doi: 10.1186/gb-2007-8-2-r19
Honma, T., and Goto, K. (2001). Complexes of MADS-box proteins are sufficient to convert leaves into floral organs. Nature 409, 525–529. doi: 10.1038/35054083
Hsu, H. F., Hsieh, W. P., Chen, M. K., Chang, Y. Y., and Yang, C. H. (2010). C/D class MADS box genes from two monocots, orchid (Oncidium ‘Gower Ramsey’) and lily (Lilium longiflorum), exhibit different effects on floral transition and formation in Arabidopsis thaliana. Plant Cell Physiol. 51, 1029–1045. doi: 10.1093/pcp/pcq052
Johansen, B., and Frederiksen, S. (2002). “Orchid flowers: evolution and molecular development,” in Developmental Genetics and Plant Evolution, eds Q. C. B. Cronk, R. M. Bateman, and J. A. Hawkins (London: Taylor & Francis), 206–219. doi: 10.1201/9781420024982.ch10
Kanno, A., Nakada, M., Akita, Y., and Hirai, M. (2007). Class B gene expression and the modified ABC model in nongrass monocots. Sci. World J. 7, 268–279. doi: 10.1100/tsw.2007.86
Kanno, A., Saeki, H., Kameya, T., Saedler, H., and Theissen, G. (2003). Heterotopic expression of class B floral homeotic gene supports a modified ABC model for tulip (Tulipa gesneriana). Plant Mol. Biol. 52, 831–841. doi: 10.1023/A:1025070827979
Kramer, E. M., Jaramillo, M. A., and Di Stilio, V. S. (2004). Patterns of gene duplication and functional evolution during the diversification of the AGAMOUS subfamily of MADS box genes in angiosperms. Genetics 166, 1011–1023. doi: 10.1534/genetics.166.2.1011
Krizek, B. A., and Fletcher, J. C. (2005). Molecular mechanisms of flower development: an armchair guide. Nat. Rev. Genet. 6, 688–698. doi: 10.1038/nrg1675
Li, H.-Y., Zhao, R., Wang, C., Zhang, L.-Y., Zhao, H., and Wang, Y.-Q. (2013). Molecular cloning and transcriptional analysis of the putative AGAMOUS homolog AcAG in Onion (Allium cepa). Plant Mol. Biol. Rep. 31, 1346–1357. doi: 10.1007/s11105-013-0607-y
Li, Q., Li, X., Bai, S., Lu, W., and Zhang, X. (2002). Isolation of HAG1 and its regulation by plant hormones during in vitro floral organogenesis in Hyacinthus orientalis L. Planta 215, 533–540. doi: 10.1007/s00425-002-0796-3
Lindqvist, C., Laakkonen, L., and Albert, V. A. (2007). Polyglutamine variation in a flowering time protein correlates with island age in a Hawaiian plant radiation. BMC Evol. Biol. 7:105. doi: 10.1186/1471-2148-7-105
Litt, A., and Irish, V. (2003). Duplication and diversification in the APETALA1/FRUITFULL floral homeotic gene lineage: implications for the evolution of floral development. Genetics 165, 821–833. Available online at: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1462802/
Lu, Z. X., Wu, M., Loh, C. S., Yeong, C. Y., and Goh, C. J. (1993). Nucleotide sequence of a flower-specific MADS box cDNA clone from orchid. Plant Mol. Biol. 23, 901–904. doi: 10.1007/BF00021545
Martinez-Castilla, L. P., and Alvarez-Buylla, E. R. (2003). Adaptive evolution in the Arabidopsis MADS-box gene family inferred from its complete resolved phylogeny. Proc. Natl. Acad. Sci. U.S.A. 100, 13407–13412. doi: 10.1073/pnas.1835864100
Melzer, R., Wang, Y.-Q., and Theiãÿen, G. N. (2009). The naked and the dead: the ABCs of gymnosperm reproduction and the origin of the angiosperm flower. Semin. Cell Dev. Biol. 21, 118–128. doi: 10.1016/j.semcdb.2009.11.015
Mondragón-Palomino, M. (2013). Perspectives on MADS-box expression during orchid flower evolution and development. Front. Plant Sci. 4, 1–9. doi: 10.3389/fpls.2013.00377
Mondragón-Palomino, M., Hiese, L., Härter, A., Koch, M. A., and Theissen, G. (2009). Positive selection and ancient duplications in the evolution of class B floral homeotic genes of orchids and grasses. BMC Evol. Biol. 9:81. doi: 10.1186/1471-2148-9-81
Mondragón-Palomino, M., and Theißen, G. (2008). MADS about the evolution of orchid flowers. Trends. Plant Sci. 13, 51–59. doi: 10.1016/j.tplants.2007.11.007
Mondragón-Palomino, M., and Theißen, G. (2009). Why are orchid flowers so diverse? Reduction of evolutionary constraints by paralogues of class B floral homeotic genes. Ann. Bot. 104, 583. doi: 10.1093/aob/mcn258
Mondragón-Palomino, M., and Theißen, G. (2011). Conserved differential expression of paralogous DEFICIENS- and GLOBOSA-like MADS-box genes in the flowers of Orchidaceae: refining the ‘orchid code’ Plant J. 66, 1008–1019. doi: 10.1111/j.1365-313X.2011.04560.x
Nakada, M., Komatsu, M., Ochiai, T., Ohtsu, K., Nakazono, M., Nishizawa, N. K., et al. (2006). Isolation of MaDEF from Muscari armeniacum and analysis of its expression using laser microdissection. Plant Sci. 170, 143–150. doi: 10.1016/j.plantsci.2005.08.021
Nakamura, T., Fukuda, T., Nakano, M., Hasebe, M., Kameya, T., and Kanno, A. (2005). The modified ABC model explains the development of the petaloid perianth of Agapanthus praecox ssp. orientalis (Agapanthaceae) flowers. Plant Mol. Biol. 58, 435–445. doi: 10.1007/s11103-005-5218-z
Nam, J., Depamphilis, C. W., Ma, H., and Nei, M. (2003). Antiquity and evolution of the MADS-box gene family controlling flower development in plants. Mol. Biol. Evol. 20, 1435–1447. doi: 10.1093/molbev/msg152
Pabón-Mora, N., Hidalgo, O., Gleissberg, S., and Litt, A. (2013). Assessing duplication and loss of APETALA1/FRUITFULL homologs in Ranunculales. Front. Plant. Sci. 4:358. doi: 10.3389/fpls.2013.00358
Pelaz, S., Ditta, G. S., Baumann, E., Wisman, E., and Yanofsky, M. F. (2000). B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature 405, 200–203. doi: 10.1038/35012103
Preston, J. C., and Kellogg, E. A. (2006). Reconstructing the evolutionary history of paralogous APETALA1/FRUITFULL-like genes in grasses (Poaceae). Genetics 174, 421–437. doi: 10.1534/genetics.106.057125
Purugganan, M. D., Rounsley, S.D., Schmidt, R. J., and Yanfosky, M. F. (1995). Molecular evolution of flower development: Diversification of the plant MADS-box regulatory gene family. Genetics 140, 345–356.
Ramirez, S. R., Gravendeel, B., Singer, R. B., Marshall, C. R., and Pierce, N. E. (2007). Dating the origin of the Orchidaceae from a fossil orchid with its pollinator. Nature 448, 1042–1045. doi: 10.1038/nature06039
Rudall, P. J., and Bateman, R. M. (2002). Roles of synorganisation, zygomorphy and heterotopy in floral evolution: The gynostemium and labellum of orchids and other lilioid monocots. Biol. Rev. 77, 403–441. doi: 10.1017/S1464793102005936
Salemme, M., Sica, M., Gaudio, L., and Aceto, S. (2013). The OitaAG and OitaSTK genes of the orchid Orchis italica: a comparative analysis with other C- and D-class MADS-box genes. Mol. Biol. Rep. 40, 3523–3535. doi: 10.1007/s11033-012-2426-x
Skipper, M., Johansen, L. B., Pedersen, K. B., Frederiksen, S., and Johansen, B. (2006). Cloning and transcription analysisi of an AGAMOUS- and SEEDSTICK orthologo in the orchid Dendrobium thyrsiflorum (Reichb. f.). Gene 366, 266–274. doi: 10.1016/j.gene.2005.08.014
Skipper, M., Pedersen, K. B., Johansen, L. B., Frederiksen, S., Irish, V. F., and Johansen, B. (2005). Identification and quantification of expression levels of three FRUITFULL-like MADS-box genes from the orchid Dendrobium thyrsiflorum (Reich. f.). Plant Sci. 169, 579–586. doi: 10.1016/j.plantsci.2005.04.011
Smaczniak, C., Immink, R. G. H., Muiño, J. M., Blanvillain, R., Busscher, M., Busscher-Lange, J., et al. (2012). Characterization of MADS-domain transcription factor complexes in Arabidopsis flower development. Proc. Natl. Acad. Sci. U.S.A. 109, 1560–1565. doi: 10.1073/pnas.1112871109
Song, I.-J., Nakamura, T., Fukuda, T., Yokoyama, J., Ito, T., Ichikawa, H., et al. (2006). Spatiotemporal expression of duplicate AGAMOUS orthologues during floral development in Phalaenopsis. Dev. Genes. Evol. 216, 301–313. doi: 10.1007/s00427-005-0057-0
Theissen, G. (2001). Development of floral organ identity: stories from the MADS house. Curr. Opin. Plant. Biol. 4, 75–85. doi: 10.1016/S1369-5266(00)00139-4
Theissen, G., Kim, J. T., and Saedler, H. (1996). Classification and phylogeny of the MADS-box multigene family suggest defined roles of MADS-box gene subfamilies in the morphological evolution of eukaryotes. J. Mol. Evol. 43, 484–516. doi: 10.1007/BF02337521
Tsaftaris, A., Pasentsis, K., Makris, A., Darzentas, N., Polidoros, A., Kalivas, A., et al. (2011). The study of the E-class SEPALLATA3-like MADS-box genes in wild-type and mutant flowers of cultivated saffron crocus (Crocus sativus L.) and its putative progenitors. J. Plant Physiol. 168, 1675–1684. doi: 10.1016/j.jplph.2011.03.015
Tsaftaris, A. S., Pasentis, K., Iliopoulos, I., and Polidoros, A. N. (2004). Isolation of three homologous AP1-like MADS-box genes in crocus (Crocus sativus L) and characterization of their expression. Plant Sci. 166, 1235–1243. doi: 10.1016/j.plantsci.2003.12.037
Tsaftaris, A. S., Pasentsis, K., and Polidoros, A. N. (2005). Isolation of a differentially spliced C-type flower specific AG-like MADS-box gene from Crocus sativus and characterization of its expression. Biol. Plant. 49, 499–504. doi: 10.1007/s10535-005-0041-6
Tsai, W. C., Kuoh, C. S., Chuang, M. H., Chen, W. H., and Chen, H. H. (2004). Four DEF-like MADS box genes displayed distinct floral morphogenetic roles in Phalaenopsis orchid. Plant Cell Physiol. 46, 831–844. doi: 10.1093/pcp/pch095
Tzeng, T. Y. (2002). Ectopic expression of carpel-specific MADS box genes from Lily and Lisianthus causes similar homeotic conversion of sepal and petal in Arabidopsis. Plant Physiol. 130, 1827–1836. doi: 10.1104/pp.007948
Tzeng, T. Y., Hsiao, C. C., Chi, P. J., and Yang, C. H. (2003). Two lily SEPALLATA-like genes cause different effects on floral formation and floral transition in Arabidopsis. Plant Physiol. 133, 1091–1101. doi: 10.1104/pp.103.026997
Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N., De Paepe, A., et al. (2002). Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3:RESEARCH0034. doi: 10.1186/gb-2002-3-7-research0034
Van Tunen, A. J., Eikelboom, W., and Angenent, G. (1993). Floral organogenesis in Tulipa. Flow. Newsl. 16, 33–38.
Wang, S. Y., Lee, P. F., Lee, Y. I., Hsiao, Y. Y., Chen, Y. Y., Pan, Z. J., et al. (2011). Duplicated C-class MADS-Box genes reveal distinct roles in gynostemium development in Cymbidium ensifolium (Orchidaceae). Plant Cell Physiol. 52, 563–577. doi: 10.1093/pcp/pcr015
Weigel, D., and Meyerowitz, E. M. (1994). The ABCs of floral homeotic genes. Cell 78, 203–209. doi: 10.1016/0092-8674(94)90291-7
Whipple, C. J., Ciceri, P., Padilla, C. M., Ambrose, B. A., Bandong, S. L., and Schmidt, R. (2004). Conservation of B-class floral homeotic gene function between maize and Arabidopsis. Development 131, 6083–6091. doi: 10.1242/dev.01523
Whipple, C. J., Zanis, M. J., Kellogg, E. A., and Schmidt, R. J. (2007). Conservation of B class gene expression in the second whorl of a basal grass and outgroups links the origin of loducules and petals. Proc. Natl. Acad. Sci. U.S.A. 104, 1081–1086. doi: 10.1073/pnas.0606434104
Xu, H. Y., Li, X. G., Li, Q. Z., Bai, S. N., Lu, W. L., and Zhang, X. S. (2004). Characterization of HoMADS 1 and its induction by plant hormones during in vitro ovule development in Hyacinthus orientalis L. Plant Mol. Biol. 55, 209–220. doi: 10.1007/s11103-004-0181-7
Xu, Y., Teo, L. L., Zhou, J., Kumar, P. P., and Yu, H. (2006). Floral organ identity genes in the orchid Dendrobium crumenatum. Plant J. 46, 54–68. doi: 10.1111/j.1365-313X.2006.02669.x
Yu, H., and Goh, C. J. (2000). Identification and characterization of three orchid MADS-box genes of the AP1/AGL9 subfamily during floral transition. Plant Physiol. 123, 1325–1336. doi: 10.1104/pp.123.4.1325
Yu, H., Hua, S., and Goh, C. J. (2002). Spatial and temporal expression of the orchid floral homeotic gene DOMADS1 is mediated by its upstream regulatory regions. Plant Mol. Biol. 49, 225–237. doi: 10.1023/A:1014958118852
Zahn, L. M., Kong, H., Leebens-Mack, J. H., Kim, S., Soltis, P. S., Landherr, L. L., et al. (2005a). The evolution of the SEPALLATA subfamily of MADS-box genes: a pre-angiosperm origin with multiple duplications throughout angiosperm history. Genetics 169, 2209–2223. doi: 10.1534/genetics.104.037770
Zahn, L. M., Leebens-Mack, J., Depamphilis, C. W., Ma, H., and Theissen, G. (2005b). To B or not to B a flower: the role of DEFICIENS and GLOBOSA orthologs in the evolution of the angiosperms. J. Hered. 96, 225–240. doi: 10.1093/jhered/esi033
Zahn, L. M., Leebens-Mack, J. H., Arrington, J. M., Hu, Y., Landherr, L. L., Depamphilis, C. W., et al. (2006). Conservation and divergence in the AGAMOUS subfamily of MADS-box genes: evidence of independent sub- and neofunctionalization events. Evol. Dev. 8, 30–45. doi: 10.1111/j.1525-142X.2006.05073.x
Keywords: Orchidaceae, flower evolution, evo-devo, qPCR, gene duplication, labellum
Citation: Acri-Nunes-Miranda R and Mondragón-Palomino M (2014) Expression of paralogous SEP-, FUL-, AG- and STK-like MADS-box genes in wild-type and peloric Phalaenopsis flowers. Front. Plant Sci. 5:76. doi: 10.3389/fpls.2014.00076
Received: 15 November 2013; Accepted: 17 February 2014;
Published online: 12 March 2014.
Edited by:
Michael J. Scanlon, Cornell University, USACopyright © 2014 Acri-Nunes-Miranda and Mondragón-Palomino. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mariana Mondragón-Palomino, Department of Cell Biology and Plant Biochemistry, University of Regensburg, Universitätsstraße 31, 93053 Regensburg, Germany e-mail:bWFyaWFuYS5tb25kcmFnb25AYmlvbG9naWUudW5pLXJlZ2Vuc2J1cmcuZGU=