 Emanuel Bojórquez-Quintal1Ana Velarde-Buendía2Ángela Ku-González1Mildred Carillo-Pech1Daniela Ortega-Camacho3Ileana Echevarría-Machado1
Emanuel Bojórquez-Quintal1Ana Velarde-Buendía2Ángela Ku-González1Mildred Carillo-Pech1Daniela Ortega-Camacho3Ileana Echevarría-Machado1 Igor Pottosin2
Igor Pottosin2 Manuel Martínez-Estévez1*
Manuel Martínez-Estévez1*- 1Unidad de Bioquímica y Biología Molecular de Plantas, Centro de Investigación Científica de Yucatán, Yucatán, México
- 2Centro Universitario de Investigaciones Biomédicas, Universidad de Colima, Colima, México
- 3Unidad de Ciencias del Agua, Centro de Investigación Científica de Yucatán, Yucatán, México
Despite its economic relevance, little is known about salt tolerance mechanisms in pepper plants. To address this question, we compared differences in responses to NaCl in two Capsicum chinense varieties: Rex (tolerant) and Chichen-Itza (sensitive). Under salt stress (150 mM NaCl over 7 days) roots of Rex variety accumulated 50 times more compatible solutes such as proline compared to Chichen-Itza. Mineral analysis indicated that Na+ is restricted to roots by preventing its transport to leaves. Fluorescence analysis suggested an efficient Na+ compartmentalization in vacuole-like structures and in small intracellular compartments in roots of Rex variety. At the same time, Na+ in Chichen-Itza plants was compartmentalized in the apoplast, suggesting substantial Na+ extrusion. Rex variety was found to retain more K+ in its roots under salt stress according to a mineral analysis and microelectrode ion flux estimation (MIFE). Vanadate-sensitive H+ efflux was higher in Chichen-Itza variety plants, suggesting a higher activity of the plasma membrane H+-ATPase, which fuels the extrusion of Na+, and, possibly, also the re-uptake of K+. Our results suggest a combination of stress tolerance mechanisms, in order to alleviate the salt-induced injury. Furthermore, Na+ extrusion to apoplast does not appear to be an efficient strategy for salt tolerance in pepper plants.
Introduction
The excess of soluble salts in soil, particularly NaCl, causes three types of stresses in plants: osmotic, ionic, and oxidative. These stresses reduce absorption and induce a massive efflux of water and ions (K+) in plant cells, resulting in water, and nutritional imbalances. The accumulation of Na+ to toxic concentrations and the production of reactive oxygen species (ROS) reduce the growth, yield, and production of economically important crops, such as cereals and vegetables (Munns and Tester, 2008; Bojórquez-Quintal et al., 2012). Plants in relation to salt can be classified into two groups: halophytes (growth stimulated at moderate and tolerant to high salinity) and glycophytes, which display a suppressed growth in a saline environment (Flowers and Colmer, 2008; Ruan et al., 2010). In halophytes, various adaptive mechanisms to tolerate high levels of salt have been identified and intensively studied (Ruan et al., 2010; Adolf et al., 2013; Shabala, 2013). Unfortunately, some of these mechanisms may not be directly transferred to crop plants, which are mostly glycophytes (Zhang and Shi, 2013). Yet, several crops are relatively salt resistant, and there are also substantial differences in the salt tolerance between nearly isogenic varieties within the same plant species. Salt tolerance is a complex trait controlled by many genes and involves various biochemical and physiological mechanisms. The fine tuning of these mechanisms is necessary to achieve a significant increase in tolerance to salt (Zhang and Shi, 2013; Adem et al., 2014).
Proline is the most common compatible osmolyte in plants and has therefore been extensively studied. The accumulation of this amino acid is an important regulatory mechanism under osmotic stress (Huang et al., 2013). Proline is a multifunctional amino acid (Szabados and Savouré, 2009). In many plant species, the accumulation of proline has been associated with tolerance to salt stress and has even been used as a marker to select tolerant genotypes (Ashraf and Harris, 2004). However, a negative correlation between the accumulation of proline and salt tolerance has also been reported, indicating discrepancies in its function (Lutts et al., 1999; Chen et al., 2007a). Proline accumulation is made possible by the increase in the expression and activity of the synthesis enzymes (Δ-pyrroline-5-carboxylate synthetase, P5CS; Δ-pyrroline-5-carboxylate reductase, P5CR) or by the decrease in the degradation enzymes, proline dehydrogenase or proline oxidase (PDH or POX), and P5C dehydrogenase (P5CDH) (Huang et al., 2013). Under salt stress, the P5CS1 and PDH genes are positively and negatively regulated, respectively (Kishor et al., 2005; Verslues and Sharma, 2010; Jaarsma et al., 2013). Similarly, the overexpression of the P5CS gene increases proline synthesis under salt stress and improves tolerance to salt (Kishore et al., 1995; Hmida-Sayari et al., 2005).
The roots are the first site of contact with high concentrations of Na+ in the soil and therefore of the uptake or absorption of salt. Na+ influx is mediated by non-selective cation channels (NSCC), high-affinity K+ transporters (HKTs), and low-affinity cation transporters (LCT) in the root epidermal cells (Apse and Blumwald, 2007; Plett and Moller, 2010; Maathuis, 2014). Na+ is then transported radially toward the root xylem via the apoplast and symplast. After being loaded into the xylem, Na+ is finally transported to the shoots by xylem flow (Adams and Shin, 2014). In contrast to halophytes, Na+ is not an essential element for most plants and becomes highly toxic at high concentrations, particularly in the aerial parts of the plant. Therefore, it is necessary to maintain efficient control of Na+ content and intracellular compartmentalization in plant tissues. The high-affinity potassium transporters (HKTs), the Na+/H+ SOS1 (salt overly sensitive) antiporters on the plasma membrane and the intracellular NHX antiporters (Na+/H+) are transporters involved in the Na+ homeostasis (Almeida et al., 2013; Adams and Shin, 2014).
The regulation of K+ homeostasis is essential for plant adaptation to biotic and abiotic stresses. This adaptation is associated with the wide range of functions in which K+ participates (Anschütz et al., 2014; Demidchik, 2014; Shabala and Pottosin, 2014). Recently, K+ retention in the cells of roots and leaves has been identified as an important trait for salt tolerance. A strong negative correlation between the magnitude of salt-induced K+ loss and salt tolerance, observed in various crop species, suggested K+ retention as a selection criterion between salt tolerant and sensitive varieties (Chen et al., 2005, 2007b,c; Smethurst et al., 2008; Lu et al., 2013; Wu et al., 2013; Bonales-Alatorre et al., 2013a). Furthermore, it has been observed that the exogenous administration of organic compounds and divalent cations prevents K+ efflux (Cuin and Shabala, 2005, 2007a,b; Shabala et al., 2006; Zhao et al., 2007; Chen et al., 2007a; Zepeda-Jazo et al., 2008). Efficient control of membrane potential due to the H+-ATPase activity was shown to be important for the salt tolerance in several species (Chen et al., 2007b; Cuin et al., 2008; Hariadi et al., 2011; Bose et al., 2013, 2014). A more negative membrane potential during salt stress reduces the driving force for the K+ loss and facilitates the K+ absorption, thus allowing plants to retain K+ in the cytosol (Chen et al., 2007b; Bose et al., 2013). Likewise, the H+-ATPase activity is essential to fuel Na+/H+ exchangers in the plasma membrane (SOS1). At the same time, a higher activity of the H+ pump consumes a large amount of ATP, hence has a higher energetic cost (Malagoli et al., 2008). Thus, keeping electrochemical gradients for physiologically important cations across the plasma membrane could present an energetic burden, so this tolerance mechanism cannot be considered permanent and may be used as a temporary solution at early times after the onset of the salt stress (Bose et al., 2013, 2014).
Peppers (Capsicum spp.) are an economically important genus of the Solanaceae family, which also includes tomatoes and potatoes. Among the 32 species native to America, C. annuum L., C. baccatum L., C. frutescens L., C. pubescens L., and C. chinense Jacq. are cultivated (Moscone et al., 2007; Perry et al., 2007). Overall, pepper plants are grown around the world because of their adaptation to different agro-climatic regions and their wide variety of shapes, sizes, colors, and pungencies of the fruit (Qin et al., 2014). However, these plants are sensitive to various biotic stresses, such as viruses and Oomycetes and abiotic factors such as drought and salinity. In fact, pepper plants are considered moderately sensitive, sensitive or highly susceptible to salt stress (Maas and Hoffman, 1977; Aktas et al., 2006). Nevertheless, despite their economic importance as a horticultural species, very little is known about the mechanisms of tolerance to high salt concentrations. To contribute to the understanding of salt stress in species of economic importance such as peppers, the difference in salt sensitivity of two varieties of the species C. chinense Jacq, commonly known as habanero pepper, was evaluated in this study. Furthermore, possible mechanisms of salt stress tolerance for the two varieties were addressed by electrophysiological studies using selective microelectrodes (MIFE) and by subcellular localization of Na+ using fluorescent indicators.
Materials and Methods
Plant Material and Growth Conditions
Habanero pepper (C. chinense Jacq.) seeds of the Chichen-Itza (Seminis®) and Rex (Mayan Chan obtained in CICY) varieties were used in this study. To disinfect the seeds, they were rinsed in 80% ethanol (v/v) for 5 min and washed continuously with sterile water. Seeds were then incubated with 30% (v/v) sodium hypochlorite (Cloralex 5% NaOCl) and Tween (1 drop) for 15 min. Washes were continuous, and the seeds were kept in sterile water for 48 h at 4°C in the dark. After stratification, seeds of both varieties were incubated (in the dark) in Petri dishes with filter paper moistened with sterile water until the emergence of the radicle.
For the hydroponic experiments, germinated seeds were transferred to plastic containers with vermiculite moistened with a Hoagland solution to a fifth of its ionic strength (H1/5). Seeds were incubated under photoperiods of 16/8 h light/dark at 25°C. The light intensity was 123 μ mol m−2 s−1. Seedlings were irrigated for 45 days with a sterile water solution and H1/5 with 7-day intervals. The modified Hoagland solution at one-fifth of its ionic strength (H1/5) contained the following: 1.2 mM KNO3, 0.8 mM Ca(NO3)2, 0.2 mM KH2PO4, 0.2 mM MgSO4, 50 μM CaCl2, 12.5 μM H3BO3, 1 μM MnSO4, 1 μM ZnSO4, 0.5 μM CuSO4, 0.1 μM (NH4)6Mo7O24, 0.1 μM NaCl, and 10 μM Fe-EDTA, pH 6.8.
For electrophysiological experiments and subcellular Na+ localization, seeds with radicles were transferred to Petri dishes containing modified Gamborg-B5 growth medium (Sales B5, Sigma) at half ionic strength (B5/2). B5/2 medium was supplemented with 0.5% sucrose (w/v) and 1% agar (w/v). The pH was adjusted to 5.8. Seedlings 10 days of age with a primary root 8–10 cm in length were used for this experiment.
NaCl Stress Treatment
Forty–five- to fifty-day-old seedlings (growing in vermiculite) of Rex and Chichen-Itza varieties were used. After pre-treatment with a solution of H1/5 for 7 days to avoid mechanical damage, seedlings homogeneous in size were selected. Three replicates of 10 seedlings of each variety were subjected to 7 days of treatment at concentrations of 0, 50, 100, and 150 mM NaCl in H1/5 solution. Treatments were performed in a culture room with photoperiods of 16/8 h light/dark at 25°C. At the end of the experiment, the seedlings were harvested and washed with sterile water to remove excess NaCl, and the fresh weight (FW), dry weight (DW), and the water content was determined by the formula (FW-DW)/FW. Each type of sample was dried in an electric oven at 60°C for 72 h. The leaves and roots were used for the determination of proline, Na+, and K+ content.
Proline Content
A modification of the method described by Bates et al. (1973) was used to determine proline content. Briefly, dry leaf and root tissues were macerated and homogenized in 10 mL of boiling water. For each reaction, 2 mL of the supernatant was mixed with 2 mL of acetic acid and 2 mL of ninhydrin. The reaction mixture was heated in a water bath at 100°C for 60 min, and the reaction was stopped in an ice bath. For extraction, 4 mL of toluene was added, and samples were mixed vigorously for 15–20 s. Samples were then set aside to allow separation of the organic and aqueous phases. The organic phase containing the chromophore was collected in a new tube, and absorbance was read at 520 nm using toluene as a blank. Proline concentration was determined from a standard curve, and concentrations were calculated based on DW.
Sodium and Potassium Quantification
Samples of dried leaves and roots were weighed, and HNO3:H2O2 was added at a 5:1 (v:v) ratio. Microwave digestion was performed at 1200 W using a ramp of 15 min to 200°C, 10 min to 200°C, and 5 min to 170°C. Subsequently, samples were adjusted to a volume of 25 mL with water (Milli Q), and Na+ and K+ content was quantified by inductively coupled plasma atomic emission spectroscopy (ICP-AES, Thermo IRIS Intrepid II XDL, New York, USA). Standard curves were used for each element.
MIFE Technique
The net flux of K+ and H+ on the surface of the roots of the two varieties of C. chinense was measured non-invasively by the microelectrode ion flux estimation (MIFE) technique (Newman, 2001). For MIFE studies, seedlings grown in vitro with roots of 8–10 cm in length were transferred and fixed horizontally to a measuring chamber. Subsequently, 25 mL of measurement solutions were added for K+ (0.5 mM KCl, 0.1 mM CaCl2, 5 mM MES, 2 mM Tris base, pH 6.0) and for H+ (0.5 mM KCl and 0.1 mM CaCl2, without pH-buffer) and incubated for 1 h to allow stabilization. Two electrodes selective for K+ and H+ were used in each experiment. For salt stress treatment, NaCl solution was added to the measuring chamber to a final concentration of 150 mM. Before the experiment, the microelectrodes were filled with 0.5 mM KCl for K+ or 0.15 mM NaCl and 0.04 mM KH2PO4 for H+, and the tip of each electrode was filled with the ion-selective resin (ion-liquid exchanger, LIX Fluka, Sigma-Aldrich) for the ion measured. Two electrodes were then mounted in a micromanipulator, and located perpendicular to the root axis 20–40 μm from the mature root zone, 1–2 cm from the root apex. Measurements were initiated by moving the electrodes 50 μm back and then forth and back in a cyclic manner every 8 s. The software CHART recorded potential differences between the two measurement positions and converted them into electrochemical potential differences using the Nernst slope. Net ion fluxes were calculated using the MIFEFLUX software for cylindrical diffusion geometry.
Localization and Subcellular Accumulation of Sodium
Sodium Green™ indicator (S-6901, Molecular Probes, Life Technologies) was used to evaluate the subcellular localization and accumulation of sodium in the roots of C. chinense. Seedlings grown in vitro were incubated in a measurement solution (0.5 mM KCl, 0.1 mM CaCl2, 5 mM MES, 2 mM Tris base, pH 6.0) supplemented or not with 150 mM NaCl. After 60 min of treatment, control, and NaCl treated seedlings were washed with distilled water and a solution of 0.5 mM CaCl2. Root segments (1–1.5 cm long) were cut from the mature zone to 1–2 cm from the root apex and were incubated for 60 min in Eppendorf tubes of 500 μL (measurement solution) with 10 μM of Sodium Green™ (sodium staining) and 20 μM of FM®4-64 (membrane staining, T-13320, Molecular Probes, Life Technologies). Excess dye was removed, and the primary root segments were placed on a slide. Approximately 10 μL of Vectashield (H-1000, Vector Laboratories, Inc.) with DAPI (nuclei staining, D3571, Molecular Probes, Life Technologies) was added. The fluorescence was observed using the confocal microscope FluoView™ FV1000 (Olympus, Japan). A UPLFLN 40X0 (oil, NA: 1.3) lens was used with a scanning speed of 10 μs/pixel. DAPI, Sodium Green™, and FM4-64 exhibit excitation and emission wavelengths of 358–461 nm, 507–532 nm, and 515–670 nm, respectively. Z images had a resolution of 512 × 512 pixels and were projected as a single image. Fluorophores were merged using the software FV10-ASW 3.01b.
Statistical Analysis
Data were analyzed using a One-Way analysis of variance (ANOVA) (Sigma Stat Version 3.1). Treatment averages were compared using Tukey's range test.
Results
Varieties of C. chinense Differ in Sensitivity to NaCl Stress
Different varieties of C. chinense such as Rex and Chichen-Itza differ in their morphologic characteristics (fruit color, size, and root system architecture), as shown in Figures 1A,C. These two varieties exhibit differing sensitivities to salt stress, Rex being more tolerant than Chichen-Itza (Figures 1B,C). A concentration of 150 mM of NaCl over 7 days of culture in hydroponic conditions had a strong impact on the growth of the two varieties. Loss of turgor, leaf abscission, and darkening of the root system were observed, especially in the variety Chichen-Itza (Figures 1B,C).

Figure 1. Morphologic and sensitivity differences to NaCl stress in two varieties of C. chinense. (A) Representative color of ripe habanero pepper fruits of the Rex (red) and Chichen-Itza (orange) varieties. (B) Forty-five-day-old seedlings of two varieties of C. chinense after 7 days of culture in H1/5 solution supplemented with 150 mM NaCl. (C) Appearance and sensitivity of Rex and Chichen-Itza seedlings after 7 days of treatment under control conditions (top) and 150 mM NaCl (bottom).
A significant reduction of fresh and DWs was also induced by NaCl in both genotypes (Figures 2A,B). Under salt stress, the FW reduction was greater in Chichen-Itza, (75%) than in the Rex variety (50%) (Figure 2A). The water content in the Rex variety was identical to the water content in control seedlings with no salt treatments. However, the water content in the Chichen-Itza variety was significantly lower than in the untreated controls (Figure 2C). It is noteworthy that although symptoms of stress (wilting and senescence) were observed at concentrations below 150 mM NaCl, the effect on growth parameters was not significant between varieties by the end of the measurement period (data not shown). For this reason, a dose of 150 mM NaCl was selected for subsequent studies. This concentration has been used in various studies with glycophytes such as A. thaliana, S. tuberosum, and S. lycopersicum (Apse et al., 1999; Rodríguez-Rosales et al., 2008; Jaarsma et al., 2013).
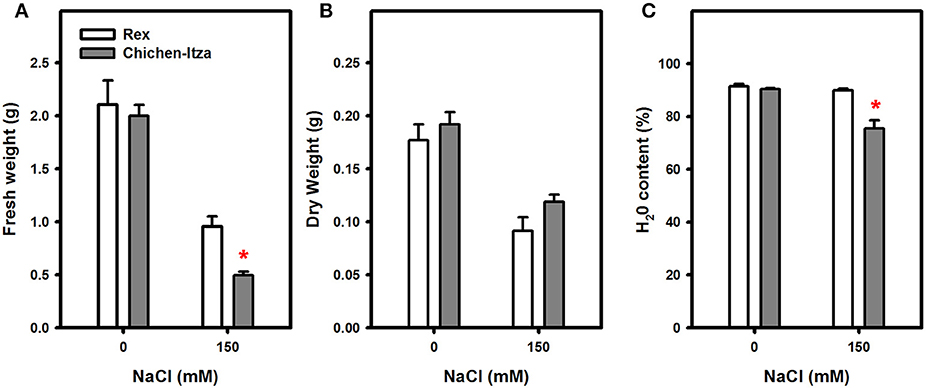
Figure 2. Effect of saline stress (150 mM NaCl) on fresh weight (A), dry weight (B), and water content (C) of Chichen-Itza and Rex varieties seedlings after 7 days of culture with 150 mM NaCl. The white bars (Rex) and gray bars (Chichen-Itza) represent the mean FW, DW, and WC for the different treatments, ME ± SD (n = 6 seedlings). The asterisk indicates statistically significant differences between varieties for each treatment (P < 0.050, Tukey's test).
Effect of NaCl on Proline Accumulation in Different Varieties of C. chinense
Many species of plants accumulate compatible solutes, such as proline, in response to abiotic stresses such as drought and salinity. In the varieties of C. chinense studied, proline accumulation was analyzed in the leaves and roots of seedlings grown in hydroponic cultures and subjected to 0 (control) or 150 mM NaCl for 7 days. Proline concentrations in the roots and leaves were similar in both varieties in the absence of salt. The proline concentration was higher in the leaves than in the roots (Figure 3). After 7 days of salt stress, the proline content in the Rex variety increased approximately 6.0-fold compared to control. However, in the Chichen-Itza variety, the values were similar to those in control seedlings growing without NaCl (Figure 3A). The same effect was observed in the roots as in the leaves. In the Rex variety, proline levels were 16-fold higher compared to control (Figure 3B). Surprisingly, the accumulation of proline in the roots was 50-fold higher in the Rex than in the Chichen-Itza variety after 7 days of treatment with NaCl (Figure 3B).
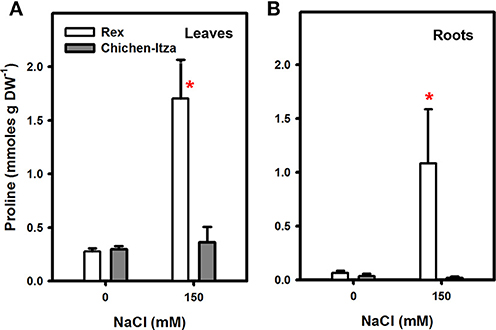
Figure 3. Effect of NaCl treatment on proline accumulation in leaves (A) and roots (B) of two varieties of C. chinense. The bars (white: Rex, gray: Chichen-Itza) represent the means of treatments with or without NaCl (150 mM), ME ± SD (n = 3). The asterisk indicates statistically significant differences between varieties for each treatment (P < 0.050, Tukey's test).
Potassium Retention and Sodium Accumulation in Roots Under Salt Stress
K+ and Na+ accumulation patterns in the two varieties of C. chinense are presented in Figure 4. In the absence of salt stress, K+ content in the leaves and roots did not significantly differ between these varieties (Figures 4A,D). A higher K+ concentration was observed in the leaves as compared to roots, due to the K+ accumulation on the top of transpiration stream (Conn and Gilliham, 2010). NaCl stress did not modify K+ content in the leaves in either variety of C. chinense (Figure 4A). However, a significant decrease in K+ was observed in the roots of seedlings from the Chichen-Itza variety under salt stress. No significant reduction of root K+ was observed in the Rex variety (Figure 4D). Furthermore, no differences in K+ were observed for concentrations lower than 150 mM of NaCl in either variety (Figure S1).
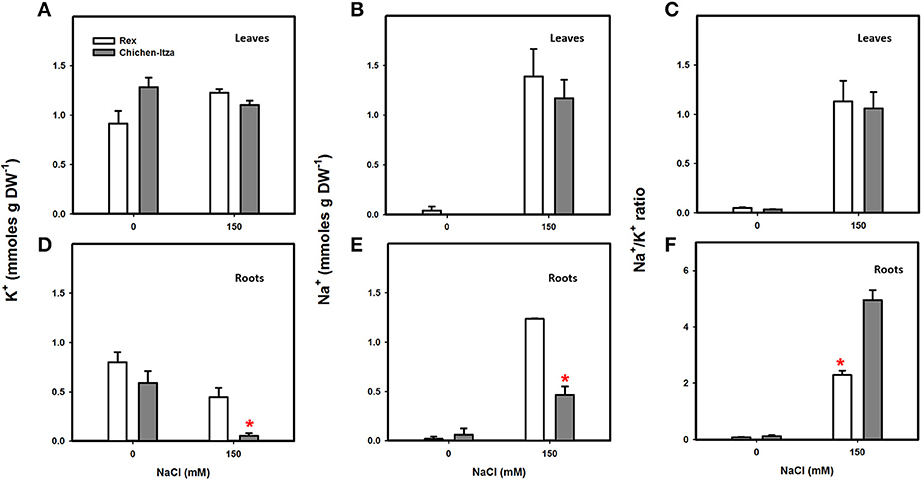
Figure 4. Potassium and sodium content in two varieties of C. chinense under NaCl stress conditions. Forty-five-day-old seedlings cultivated in hydroponic cultures (H1/5) for 7 days with 0 and 150 mM of NaCl. K+ content in the leaves (A) and roots (D) of Rex and Chichen-Itza varieties after treatment with salt. Na+ content in the leaves (B) and roots (E) of the two C. chinense varieties after 7 days of treatment. The Na+/K+ ratios in leaves (C) and roots (F) in control and NaCl treatments. Bars represent the average of the treatments with or without NaCl (150 mM), ME ± SD (n = 3). The asterisk indicates statistically significant differences between varieties for each treatment (P < 0.050, Tukey's test).
In absence of salt stress, Na+ content in the leaves and roots of both varieties was minimal, with values of 0.02 mmol and 0.04 mmol g DW−1, respectively. Treatment with 150 mM NaCl increased the Na+ content in the leaves; however, no differences were observed between the two varieties (Figure 4B). Na+ content increased in the roots treated with salt in both varieties and was surprisingly higher in Rex (twofold) compared to Chichen-Itza variety (Figure 4E). The Na+/K+ ratio in leaves and roots were very similar between pepper varieties when NaCl was not supplied (Figures 4C,F). As a consequence of increase in Na+ and decreases in K+ content by NaCl treatment, Chichen-Itza variety exhibited much higher Na+/K+ ratio in roots compared to the Rex variety (Figure 4F). In contrast, the Na+/K+ ratio in leaves at NaCl treatment did not differ between the genotypes (Figure 4C). Furthermore, at low and moderate concentrations of NaCl, the Rex variety exhibited higher Na+ content in the roots and much lower Na+ content in the leaves. The opposite effect was observed in the Chichen-Itza variety at 50 mM NaCl. Similar Na+ content was observed between shoots and roots at a concentration of 100 mM (Figure S2).
NaCl Induces K+ Efflux in Roots of C. chinense
As described so far, Chichen-Itza and Rex varieties differ in their sensitivity to salt stress, the latter being less affected (Figures 1, 2). The Rex variety retains 55% of K+ in salt-stress conditions, whereas Chichen-Itza looses about 90% of the root K+ (Figure 4). To deepen the study of this response, the K+ flux was measured using the non-invasive MIFE technique. The addition of 150 mM NaCl induced K+ efflux from the epidermal cells in the mature root zone of the two varieties of C. chinense (Figure 5). This efflux started immediately following NaCl treatment. A higher K+ efflux was observed in the roots of the Chichen-Itza variety compared to Rex variety (Figure 5A). The difference in K+ efflux in the 1 min was double and the relative difference even increased with time (Figure 5B). In the Rex variety K+ efflux was close zero after 35 min, whereas in Chichen-Itza a significant K+ efflux of about 50 nmol m−2 s−1 was observed at late times.
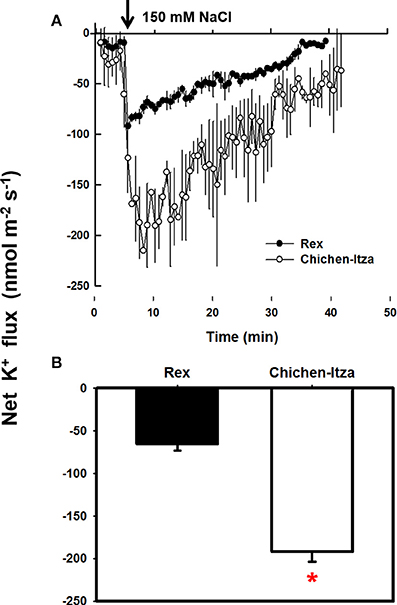
Figure 5. Measurement of the net flow of K+ in the roots of C. chinense in response to NaCl stress. (A) Kinetics of net K+ flux measured in the mature root zone of seedlings (10 days old) of the Rex (closed circles) and Chichen-Itza (open circles) varieties in response to 150 mM NaCl (the arrow indicates the time of addition of the treatment). (B) Mean net flow of K+ from the root of each variety in the first 10 min after application of 150 mM NaCl. ME ± SD (n = 4–5). The asterisk indicates statistically significant differences between varieties by treatment (P < 0.001, Tukey's test).
Effect of NaCl on H+ Efflux in Roots of C. chinense
In the roots of habanero pepper, NaCl stress caused significant changes in the net flux of H+ (Figure 6). Before starting the salt treatment (first 5 min), the net flux of H+ was zero in both varieties. Application of 150 mM NaCl induced a substantial H+ efflux (Figure 6A). In the roots of the Rex variety, NaCl induced H+ efflux was much lower as compared the Chichen-Itza variety (Figure 6B). Furthermore, a pre-treatment of seedlings with 1 mM sodium orthovanadate (an inhibitor of P-type H+-ATPases) strongly suppressed the H+ efflux from the mature root zone of both genotypes (Figure 7). Thus, the NaCl- induced H+ efflux was mediated by P-type H+-ATPases.
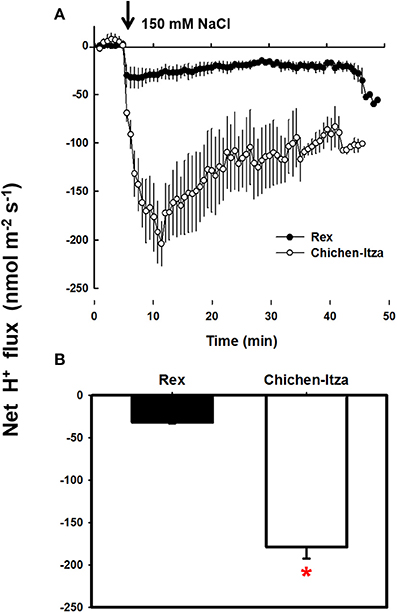
Figure 6. Effect of NaCl on the net H+ flux in the mature root area of C. chinense. (A) Kinetics of net flux of H+ measured in the roots of 10-day-old seedlings of Rex (closed circles) and Chichen-Itza (open circles) varieties after adding 150 mM NaCl (arrow indicates the time addition of the treatment). (B) Mean net flow of H+ from the root of each variety within the first 10 min of application of 150 mM NaCl. ME ± SD (n = 4–5). The asterisk indicates statistically significant differences between varieties by treatment (P < 0.001, Tukey's test).
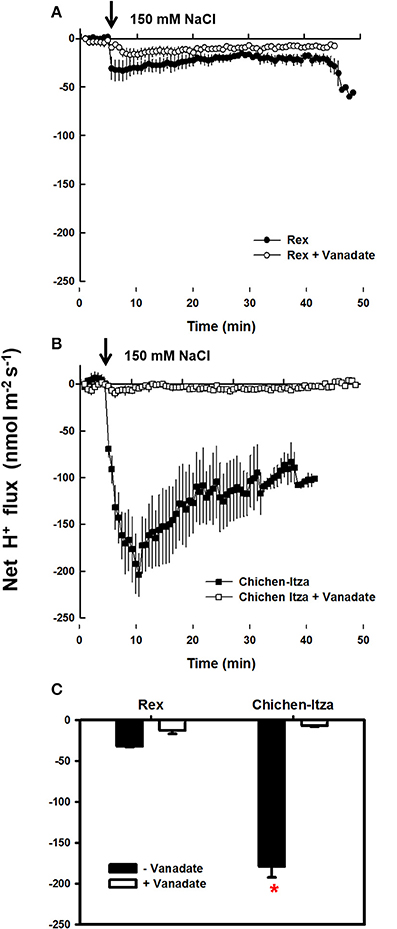
Figure 7. Effect of vanadate pre-treatment on NaCl-induced H+ efflux in roots of C. chinense. (A) Kinetics of net H+ flux measured in the mature root zone of the Rex variety and (B) of the Chichen-Itza variety. 10-day-old roots of the two strains were pre-treated with 1 mM vanadate for 45–60 min before adding of 150 mM NaCl (marked by an arrow). Open circles and squares indicate pre-treatment with vanadate. Closed circles and squares indicate no pre-treatment. (C) Average effect of vanadate pre-treatment on the net flow of H+ from the root of each variety in the first 10 min after the application of 150 mM NaCl, ME ± SD (n = 3–4). The asterisk indicates statistically significant differences between varieties by treatment (P < 0.001, Tukey's test).
Na+ Subcellular Localization in Roots
After 60 min of treatment with 150 mM NaCl, marked differences were observed with respect to Na+ localization in the mature root zone between two pepper genotypes (Figure 8). In the Rex variety, Sodium Green™ fluorescence was observed in vacuole-like structures of epidermal cells (red arrowheads, Figure 8A), suggesting an efficient mechanism for sodium compartmentalization in this variety. By contrast, in the Chichen-Itza variety, green fluorescence was observed around the epidermal cells in the mature root zone (white arrowheads, Figure 8B). Small endosomes stained with FM4-64 dye were observed in both pepper varieties under salt stress (yellow arrowheads, Figure 8). However, green fluorescence was not evident in these structures, indicating the absence of Na+ accumulation (merge, Figure 8). In epidermal cells of the Rex variety, a conglomeration of small structures with green fluorescence was observed around a larger structure comparable to a vacuole (red arrowheads, Figure 9A); similar structures were previously observed in Arabidopsis (Hamaji et al., 2009). In contrast to the Rex variety, in the Chichen-Itza a less pronounced and diffuse pattern of small compartments stained with Sodium Green™ was observed (red arrowheads, Figure 9A). Overall, the highest levels of fluorescence were observed outside the cells in Chichen-Itza (white arrowheads, Figure 9B), demonstrating that Na+ was mainly located in the apoplast. In control roots, Na+ indicator did not report any change of fluorescence for either variety (Figure S3).
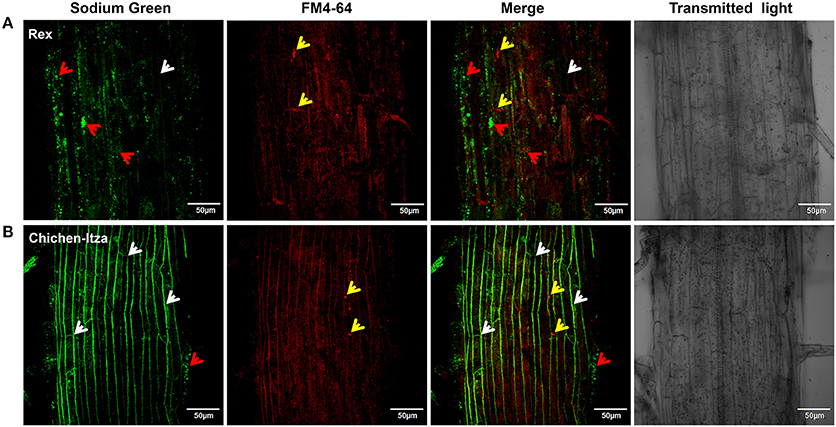
Figure 8. Sodium localization in epidermal cells of the primary root of habanero pepper C. chinense). (A) Localization of Na+ in the roots of the Rex and (B) Chichen-Itza varieties. 10-day-old habanero pepper roots were treated with 150 mM NaCl for 60 min and then stained with Sodium Green (Na+ detection) and FM4-64 (membrane staining) before the confocal images were taken. White arrowheads indicate the location of the Na+ around cells. Red arrowheads indicate the intracellular localization of Na+, and yellow arrowheads indicate the presence of endosomes. Fluorescence analysis was determined in the mature root zone. Images are representative of the analysis of three roots per treatment and variety.
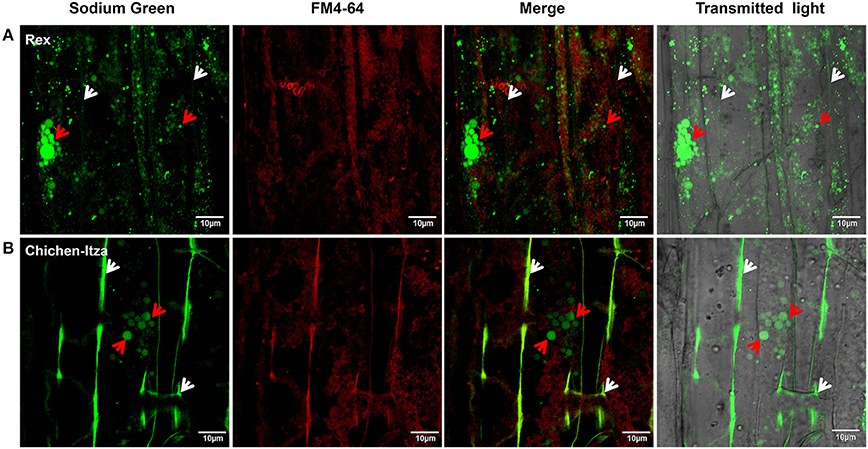
Figure 9. Sodium subcellular localization in the apoplast and subcellular compartments. Magnified images of the mature root area shown in Figure 8. (A) Localization of Na+ in the roots of the Rex and (B) Chichen-Itza varieties. The white arrowheads indicate the location of Na+ in the apoplast, and red arrowheads indicate the location of Na+ in subcellular structures. Images are representative of the analysis of three roots per treatment and variety.
Discussion
Sensitivity and Growth in C. chinense Seedlings Under NaCl Stress
Most crop plants that provide food for the world population are glycophytes and are very sensitive to high concentrations of salts in the soil, mainly NaCl. Salt stress is the main abiotic factor that affects growth, yield, and quality. Peppers (Capsicum spp.) are a major vegetable crop and are not exempt from the effect of salt throughout their ontogeny (Bojórquez-Quintal et al., 2012). Notably, pepper plants differ in their sensitivity to salt stress, including marked differences between varieties within the same species (Aktas et al., 2006). In this work, two varieties of C. chinense Jacq. were used as models. C. chinense Jacq. is a species in high demand in southeastern Mexico for its flavor and pungency and is commonly known as habanero pepper. These varieties exhibit different morphological characteristics and were shown to differ in their sensitivity to salt stress, with the Rex variety being more tolerant than the Chichen-Itza variety (Figure 1).
NaCl concentrations between 0 and 150 mM affect the growth of pepper plants, depending on the genotype, species, and condition of growth (Bojórquez-Quintal et al., 2012). In this study, the application of 150 mM NaCl had a dramatic impact on the growth of both varieties of C. chinense. Similar results were reported in C. annuum (Aktas et al., 2006). The FW, turgor, and water content of the Rex variety were less affected (Figure 2). In this variety, 70% of seedlings survived after treatment with NaCl, compared with 10% of the Chichen-Itza variety (data not shown). These results suggest the existence of intrinsic mechanisms of tolerance in the Rex variety to avoid the deleterious effect of NaCl.
Differences in Proline Accumulation Between Varieties of Habanero Peppers
Proline accumulation is one of the most common and important responses of plants to adverse environmental factors such as drought and salt stress. Proline is a multifunctional amino acid participating in a wide range of functions (Szabados and Savouré, 2009) and represents a potential biochemical marker for the salt tolerance in plants (Ashraf and Harris, 2004). In our study, different proline content was observed in two pepper varieties, contrasting in their salt sensitivity (Figure 3). In leaves of the Rex variety, proline content increased six times in the presence of NaCl, as compared to a non-significant increase in Chichen-Itza leaves. The leaves are the major site of proline synthesis (source organ). It has been suggested that proline accumulation in this organ occurs to maintain chlorophyll content and turgor to protect the photosynthetic activity under salt stress conditions (Yildiztugay et al., 2011; Huang et al., 2013). Furthermore, treatment with NaCl induced a dramatic (16-fold) increase in proline content in the roots, but only of the tolerant variety Rex. At low water potential, proline is thought to be transported from the leaf (source) to the roots (sink) for growth processes or other functions, such as osmotic adjustment depending on proline content (Sharma et al., 2011). Exogenous administration of proline reduced NaCl- induced K+ efflux in barley and Arabidopsis roots (Cuin and Shabala, 2005, 2007a,b). In Solanaceae such as the potato (S. tuberosum), increased proline content correlates with a higher expression of P5CS (synthesis gene) and decreased PDH gene expression (degradation) in tolerant but not in sensitive cultures (Jaarsma et al., 2013). Furthermore, overexpression of P5CS in N. tabacum and S. tuberosum stimulated proline accumulation under NaCl stress and improved the tolerance (Kishore et al., 1995; Hmida-Sayari et al., 2005). However, there is not always a good correlation between the accumulation of this osmolyte and tolerance to salt stress, and whether it is a symptom of damage or an indicator of tolerance is a matter of debates. For example, in rice (O. sativa), soybean (G. max), tomato (S. lycopersicum), and barley (H. vulgare), a negative correlation between proline accumulation and tolerance to stress has been reported. In these studies, sensitive genotypes accumulated more proline (Moftah and Michel, 1987; Aziz et al., 1998; Lutts et al., 1999; Chen et al., 2007a). In addition, proline synthesis is a metabolically expensive strategy (Shabala and Cuin, 2007; Shabala and Shabala, 2011). Nevertheless, proline content has been shown to be higher in many plant varieties tolerant to salt relative to their susceptible counterparts (Ashraf and Harris, 2004), as also demonstrated by the results of this work (Figure 3).
The Tolerant Pepper Variety Accumulates More Sodium in the Roots than the Sensitive Variety
The hyperaccumulation of Na+, particularly in the leaves, inhibits protein synthesis, enzymatic activity, and photosynthesis. Therefore, plants have the ability to control the transport and distribution of Na+ to organs, tissues and cells where it causes less damage to protect against the accumulation of this cation. The most sensitive glycophytes (cereals or vegetables) are unable to control the transport of Na+; therefore, large amounts of this ion are translocated to the shoot (Maathuis, 2014), inducing senescence, growth inhibition, and eventually death of the plant (Roy et al., 2014). In contrast, most halophyte and some glycophytes plants tend to accumulate large amounts of Na+ in the leaves. It has been suggested that these plants use Na+ in addition to K+ to maintain turgor and growth (Hariadi et al., 2011; Adolf et al., 2013; Bonales-Alatorre et al., 2013a,b; Maathuis, 2014).
In our study (Figure 4), we observed that two varieties of habanero pepper (C. chinense) exhibit the same Na+ content in the leaves but exhibit differences in Na+ accumulation in the roots (after a minimum of 7 days of exposure to 150 mM NaCl). In plants of C. annuum treated with NaCl, a higher Na+ content has been reported in the shoots of sensitive genotypes compared to tolerant genotypes. In fact, a positive correlation was observed between the severity of the symptoms in the leaves and Na+ concentrations in the shoots (Aktas et al., 2006). This suggests an effect dependent on the accumulation of Na+ as proposed Roy et al. (2014). The senescence and the severity of the symptoms observed in the aerial part of Capsicum chinense in this study (Figure 1) seem to be due to an osmotic effect rather than an ionic effect due to the presence of Na+ in leaves (Figure 4B). In addition, this osmotic effect can be avoided by the accumulation of proline in the Rex variety (Figure 3A) as described in the previous section, as the effect of NaCl was less severe in that variety.
The root system is the first site of detection and the first line of defense against excess Na+ in cells (Ji et al., 2013). NSCC are the principal source of Na+ influx in the plant cell (Demidchik and Tester, 2002). As suggested for other species (Demidchik and Maathuis, 2007), NSCC can mediate Na+ influx of pepper plants (Rubio et al., 2003). In our study, the Rex variety, less affected by salt, exhibited a two–fold higher Na+ content in the root than in the Chichen-Itza variety (Figure 4E). Furthermore, the accumulation of Na+ in the root system of the tolerant variety was higher at low NaCl concentrations (50 mM) where reached its peak and was maintained at higher NaCl concentrations (150 mM). The levels of Na+ in the roots of the Chichen-Itza variety were lower and stable at all of the NaCl concentrations tested (Figure S2). Overall, the accumulation of Na+ is higher (in both roots and leaves) in the tolerant variety than in the sensitive variety (Figure S1 and Figure 4). At low and moderate salt concentration ranges, the Rex variety (tolerant) possesses more Na+ in the roots and much less in the leaves. However, the opposite was observed in the Chichen-Itza variety (sensitive) at 50 mM NaCl. Furthermore, at 100 mM NaCl, the same Na+ content was observed between the shoots and roots in the Chichen-Itza variety (Figure S2). The higher Na+ content in roots than in leaves suggests that exclusion mechanisms (SOS1, antiporters, and HKT1 transporters) and compartmentalization (NHX, antiporters) are present in roots and efficiently reduce the Na+ transport to leaves. These mechanisms have also been reported in other members of Solanaceae, such as tomatoes and potatoes (Queirós et al., 2009; Rodríguez-Rosales et al., 2009; Almeida et al., 2014). In particular, the HKTs (subfamily 1) exhibit an important role in the recovery of Na+ from the xylem to prevent its transport to the aerial part, and recirculate Na+ to the roots (Horie et al., 2009; Almeida et al., 2013; Adams and Shin, 2014).
Na+ Subcelular Localization in the Roots: Compartmentalization and Na+ Efflux in Different Varieties of C. chinense
After 60 min of treatment with 150 mM NaCl, Na+ was mainly located in vacuole-like structures in root epidermal cells in the Rex variety (Figure 9). This result suggests the existence of an efficient mechanism for Na+ compartmentalization in this genotype. Similarly, it has been reported that Na+ is confined in epidermal cells vacuoles and in the cortex in the roots of Arabidopsis and Citrus (Oh et al., 2009; Gonzalez et al., 2012). Furthermore, it is noteworthy that in the two varieties of habanero pepper, small compartments were stained with the fluorophore. These compartments were found in greater quantities in the roots of the Rex variety (Figure 9). Similar results were observed in the roots of A. thaliana under salt stress. Hamaji et al. (2009) reported that Na+ accumulates in the vacuoles as well as in small vesicular compartments around vacuoles. These authors suggest that the fusion of these vesicles with the main vacuole increases its size and the tolerance to excess salt. This explanation is logical, as we observed a conglomeration of structures stained with the fluorophore in the roots of the tolerant variety after a short period of NaCl stress (Figure 9). In tolerant salt includes such as mangroves and barley, a rapid increase in vacuolar volume in response to salt stress was observed. This phenomenon was not reported in sensitive species such as peas and tomatoes (Mimura et al., 2003).
Recently, in tobacco salt-acclimated BY2 cells accumulation of Na+ in vacuoles and small vesicles was reported. Interestingly, the putative VAMP711 (vesicle-associated membrane protein 711) and VPS46 (charged multivesicular body protein) proteins were highly induced in this BY2 cells, suggesting a Na+ transport mechanism for vesicle trafficking (Garcia de la Garma et al., 2014). Furthermore, the increased Na+ sequestration by vacuolar and small compartments in the Rex variety could be due to increased expression of NHX transporters as observed in tomato species with different sensitivity to salt (Galvez et al., 2012). Also, the up-regulation of V-ATPase (vacuolar-type H+-ATPase) and H+-PPase (vacuolar H+-pyrophosphatase) in Rex variety may result in a higher proton electrochemical gradient, which facilitates enhanced sequestering of ions into the vacuole and endosomes, reducing water potential and resulting in increased salt tolerance (Gaxiola et al., 2001; Bassil et al., 2012; Pittman, 2012). In different species of plants, overexpression of AVP1 (vacuolar H+-PPase) and co-overexpression with AtNHX1 enhances salt tolerance (Gaxiola et al., 2001; Pasapula et al., 2011; Shen et al., 2014). Finally, the reduction of Na+ loss via non-selective vacuolar channels could assist efficient vacuolar Na+ accumulation (Bonales-Alatorre et al., 2013b). In the Rex variety, all or some of these mechanisms may be involved in the Na+ compartmentalization.
On the contrary, in the Chichen-Itza variety, Na+ was mostly observed in the apoplastic area between cells of the root (Figures 8, 9). This result suggests an intensive Na+ extrusion toward the outside of the cell, likely via the Na+/H+ antiporters (SOS1) of the plasma membrane (Shi et al., 2002). This is consistent with a low content of Na+ (Figure 4) and with the large active H+ efflux, mediated by vanadate-sensitive H+-ATPase, in the roots of this variety. However, the high rate of Na+ extrusion in sensitive cultivars could have a high energetic cost (Malagoli et al., 2008). This opts for the use of intracellular Na+/H+ (NHX-type) antiporters as compared to SOS1-type ones; the latter may be more useful in early responses to acute salt stress. Candidate genes for the intracellular (NHX) and plasma membrane (SOS1) exchangers were revealed in the genome of Capsicum annuum (Qin et al., 2014).
Furthermore, we may not exclude at this moment that accumulation of the indicator at cell walls, which possess high esterase activity, could contribute to the observed fluorescence signal and the difference between Rex and Chichen-Itza.
K+ Retention in the Roots is a Tolerance Mechanism in Habanero Pepper Plants
Potassium is an essential nutrient throughout the life cycle of plants, including the adaptation to hostile environments. The regulation of K+ homeostasis plays a central role in tolerance to biotic and abiotic stresses in plants (Anschütz et al., 2014; Demidchik, 2014; Shabala and Pottosin, 2014). K+ efflux from the root is a common physiological reaction that occurs under a wide range of stress conditions (Demidchik, 2014). K+ retention in roots has been proved to confer salt tolerance in barley, wheat, lucerne, and poplar (Chen et al., 2007b,c; Cuin et al., 2008; Smethurst et al., 2008; Sun et al., 2009). In this study, the treatment with 150 mM NaCl significantly decreased the content of K+ in the roots of the sensitive variety but not in the tolerant variety of habanero pepper (Figure 4D). In contrast, K+ content in the leaves was not affected by treatment with NaCl in either genotype (Figure 4A). These data suggest that the ability to retain K+ by roots is one of the salt tolerance mechanisms of C. chinense.
Initial NaCl induced K+ efflux was higher in the Chichen-Itza as compared to the Rex variety and doubled after 10 min of exposure to NaCl. Similar differences in K+ efflux have been reported in barley (H. vulgare), wheat (T. aestivum) and alfalfa (M. sativa). This difference has been used as a selection criterion to distinguish salt-tolerant from salt-sensitive genotypes (Chen et al., 2005, 2007b,c; Smethurst et al., 2008), yet some plants, like rice, did not show such a correlation (Coskun et al., 2013). In our results, content and K+ efflux in roots (Figures 4, 5) were consistent with the observed differences in sensitivity between the varieties of habanero pepper (Figures 1, 2).
NaCl-induced K+ efflux may be through outward-rectifying K+ (KOR) channels activated by depolarization as demonstrated in barley (H. vulgare) and mangrove species (Chen et al., 2007b; Sun et al., 2009; Lu et al., 2013). The use of ion channels inhibitors in habanero pepper indicate that K+ efflux from the roots are likely mediated by KOR channels rather than by NSCC channels (Bojorquez-Quintal et al., in review). Furthermore, ROS and K+ deficiency have been associated with programmed cell-death (PCD) and necrosis (Anschütz et al., 2014; Demidchik, 2014; Shabala and Pottosin, 2014). Necrosis was observed in the roots of the Chichen-Itza variety (Figure 1), which have low ability to retain K+ (Figures 4, 5).
Differences in H+ Efflux in the Roots of Habanero Pepper Under NaCl Stress
H+-ATPases generate an electrochemical gradient that maintains membrane potential and transports ions between the cytosol and the external medium. Under salt stress, NaCl universally induces H+ efflux in the roots of cereals such as H. vulgare and T. aestivum, halophytes such as C. quinoa and even in the model plant A. thaliana (Chen et al., 2007b; Cuin et al., 2008; Hariadi et al., 2011; Bose et al., 2013, 2014). In C. chinense, NaCl also induced H+ efflux in the mature root zone. H+ flux differed significantly between the varieties (Figure 6). NaCl rapidly induced the H+ efflux in both varieties of habanero peppers. It was suppressed by vanadate, the inhibitor of P-type H+-ATPase (Figure 7). H+ pumping activity of the plasma membrane H+-ATPase is essential for salt tolerance (Palmgren and Nissen, 2010). It has been suggested that the ability to K+ retain is related to the increase in H+-ATPase activity, primarily through a membrane potential repolarization (Bose et al., 2013, 2014).
Maintaining a more negative membrane potential during salt stress prevents the loss of K+ in the cytosol. In the salt-tolerant Arabidopsis relative species, T. halophila, a more negative membrane potential, correlated with a better K+ retention, was observed during salt stress (Volkov and Amtmann, 2006). In transgenic A. thaliana (HO, heme oxygenase), K+ retention was regulated by the increased H+-ATPase activity (Bose et al., 2013). A higher H+-ATPase activity maintained a more negative membrane potential and improved K+ retention in tolerant genotypes of H. vulgare (Chen et al., 2007b). In contrast, higher H+ transport activity was observed in the sensitive variety Chichen-Itza of C. chinense (Figure 6). Kinetics and magnitude of the H+ efflux coincides with that of the K+ efflux (Figure 5). Thus, it may be hypothesized that the descending phase of the H+ efflux reflects the condition of membrane potential repolarization, so that the leakage of K+ through KOR channels would be also reduced.
Furthermore, the plasma membrane H+-ATPase activity provides the driving force the Na+ extrusion via the Na+/H+ (SOS1) exchanger. In this work, we observed a massive accumulation of Na+ in the apoplast of the Chichen-Itza variety roots (Figure 9). These data suggest that SOS1 antiporter may be very active in the Chichen-Itza variety. However, Na+ efflux has a high energetic cost for the cell, especially keeping in mind a futile Na+ cycling between cytosol and apoplast. It may recruit the ATP available for other metabolic processes, thus, being detrimental to growth and yield. For this reason, the activation of H+-ATPase cannot be considered a permanent solution and might only be a temporal mechanism, as described by other studies (Ramani et al., 2006; Bose et al., 2013).
Possible Tolerance Mechanism in Peppers
Salt tolerance is a complex multigenic trait that involves many biochemical and physiological processes to achieve salt tolerance. In this paper, we demonstrate differences in salt sensitivity between two varieties of C. chinense Jacq, one of the five domesticated pepper species. We have analyzed several parameters of salt stress responses in both genotypes and their differences that may underlie their differential salt tolerance (Figure 10). One of salt tolerance mechanisms is the osmotic adjustment through the accumulation of compatible solutes (in this case, proline) in roots and leaves to maintain the absorption and prevent the loss of water (1). A second tolerance mechanism is the efficient control of Na+ transport by confining this ion to the roots, possibly through the recovery of Na+ from the xylem by HKT1 transporters (at low, moderate and high concentration of NaCl) to avoid transport to photosynthetic tissues (2). Furthermore, if the Na+ content in roots is high, this ion needs to be excluded from the cytosol to avoid toxicity. A third tolerance mechanism was observed in the tolerant variety (Rex), Na+ was efficiently compartmentalized into vacuole-like structures and small compartments which can act as osmolytes. This mechanism is possibly mediated by vacuolar and endosomal NHX antiporters (3). Additional mechanism appears to be involved in salt-sensitive variety (Chichen-Itza), which extrudes large amounts of Na+ into the apoplast (4). However, this mechanism appears to be less efficient due to its large energy cost. As in many other plant species, the regulation of K+ homeostasis through its retention in roots is crucial in habanero peppers (5), demonstrating the universality of this mechanism to the salt stress tolerance of crops.
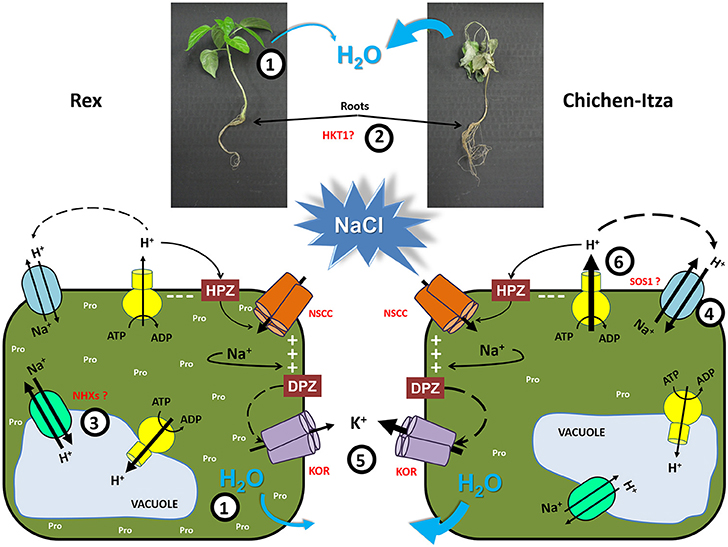
Figure 10. Schematic model of habanero pepper plant tolerance mechanisms. (1) Osmotic adjustment through the accumulation of compatible solutes as proline in leaves and roots to maintain the absorption and prevent the loss of water. (2) Efficient control of Na+ transport by confining this ion to roots, possibly through the recovery of Na+ from the xylem by HKT1 transporters to avoid transport to photosynthetic tissues. (3) Na+ compartmentalization in vacuole-like structures and small subcellular compartments, where it can act as an osmolyte, suggesting the involvement of vacuolar and endosomal NHX transporters. (4) Na+ extrusion or efflux from the cytosol to the apoplast by SOS1 exchangers. (5) Regulation of K+ homeostasis through K+ retention in roots is crucial in habanero peppers. (6) Increase in the H+-ATPase activity to generate the proton force for cell membrane repolarization or Na+ extrusion. (KOR), outward-rectifying K+ channels. HPZ is hyperpolarization and DPZ is depolarization.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
For help with the confocal microscopy analysis we thank Luis Alberto Cruz Silva (Instituto de Ecología A.C.). We thank Lucila A. Sanchez Cach for their excellent technical assistance (Centro de Investigación Científica de Yucatán, A.C.). This work was supported by CONACYT project # 166621.
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fpls.2014.00605/abstract
Figure S1. K+ content of two varieties of C. chinense at different NaCl concentrations. K+ content in the leaves (A) and roots (B) after treatment with salt. Forty-five-day-old seedlings cultivated in H1/5 for 7 days with 0, 50, 100 or 150 mM of NaCl. Bars represent averages for treatments with or without NaCl, ME ± SD (n = 3). The asterisk indicates statistically significant differences between varieties by treatment (P < 0.050, Tukey's test).
Figure S2. Na+ content in Rex and Chichen-Itza varieties at different NaCl concentrations. Na+ content in the leaves (A) and roots (B) after 7 days under salt stress conditions. Forty-five-day-old seedlings were cultivated in H1/5 with 0, 50, 100, and 150 mM of NaCl. Bars represent the average effect of the treatments with or without NaCl, ME ± SD (n = 3). The asterisk indicates statistically significant differences between varieties by treatment (P < 0.050, Tukey's test).
Figure S3. Roots of control seedlings exhibit an absence of Sodium Green™ fluorescence. Roots of untreated Rex (A) and Chichen-Itza (B) varieties stained with Sodium Green (SG), FM4-64, and DAPI. Images are representative of the analysis of three roots per treatment and variety.
References
Adams, E., and Shin, S. (2014). Transport, signalling, and homeostasis of potassium and sodium in plants. J. Integr. Plant Biol. 56, 231–249. doi: 10.1111/jipb.12159
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Adem, G. D., Roy, S. J., Zhou, M., Bowman, J., and Shabala, S. (2014). Evaluating contribution of ionic, osmotic and oxidative stress components towards salinity tolerance in barley. BMC Plant Biol. 14:113. doi: 10.1186/1471-2229-14-113
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Adolf, V. I., Jacobsen, S. E., and Shabala, S. (2013). Salt tolerance mechanisms in quinoa (Chenopodium quinoa Willd.). Environ. Exp. Bot. 92, 43–54. doi: 10.1016/j.envexpbot.2012.07.004
Aktas, H., Abak, K., and Cakmak, I. (2006). Genotypic variation in the response of pepper to salinity. Sci. Hortic. 110, 260–266. doi: 10.1016/j.scienta.2006.07.017
Almeida, P., de Boer, G.-J., and de Boer, A. H. (2014). Differences in shoot Na+ accumulation between two tomato species are due to differences in ion affinity of HKT1;2. J. Plant Physiol. 171, 438–447. doi: 10.1016/j.jplph.2013.12.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Almeida, P., Katsching, D., and de Boer, A. H. (2013). HKT transporters-State of the art. Int. J. Mol. Sci. 14, 20359–20385. doi: 10.3390/ijms141020359
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Anschütz, U., Becker, D., and Shabala, S. (2014). Going beyond nutrition: regulation of potassium homoeostasis as a common denominator of plant adaptive responses to environment. J. Plant Physiol. 171, 670–687. doi: 10.1016/j.jplph.2014.01.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Apse, M. P., Aharon, G. S., Snedden, W. A., and Blumwald, E. (1999). Salt tolerance conferred by overexpression of a vacuolar Na+/H+ antiport in Arabidopsis. Science 285, 1256–1258. doi: 10.1126/science.285.5431.1256
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Apse, M. P., and Blumwald, E. (2007). Na+ transport in plants. FEBS Lett. 581, 2247–2254. doi: 10.1016/j.febslet.2007.04.014
Ashraf, M., and Harris, P. J. C. (2004). Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 166, 3–16. doi: 10.1016/j.plantsci.2003.10.024
Aziz, A., Martin-Tanguy, J., and Larher, F. (1998). Stress-induced changes in polyamine and tyramine levels can regulate proline accumulation in tomato leaf discs treated with sodium chloride. Physiol. Plant. 104, 195–202. doi: 10.1034/j.1399-3054.1998.1040207.x
Bassil, E., Coku, A., and Blumwald, E. (2012). Cellular ion homeostasis: emerging roles of intracellular NHX Na+/H+ antiporters in plant growth and development. J. Exp. Bot. 63, 5727–5740. doi: 10.1093/jxb/ers250
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bates, L. S., Waldren, R. P., and Teare, D. (1973). Rapid determination of free proline for water-stress studies. Plant Soil 39, 2005–2007. doi: 10.1007/BF00018060
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bojórquez-Quintal, J. E., Echevarría-Machado, I., Medina-Lara, F., and Martínez-Estévez, M. (2012). Plants challenges in a salinized world: the case of Capsicum. Afr. J. Biotechnol. 11, 13614–13626. doi: 10.5897/AJB12.2145
Bonales-Alatorre, E., Pottosin, I., Shabala, L., Chen, Z.-H., Zeng, F., Jacobsen, S. E., et al. (2013a). Differential activity of plasma and vacuolar membrane transporters contributes to genotypic differences in salinity tolerance in a halophyte species, Chenopodium quinoa. Int. J. Mol. Sci. 14, 9267–9285. doi: 10.3390/ijms14059267
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bonales-Alatorre, E., Shabala, S., Chen, Z. H., and Pottosin, I. (2013b). Reduced tonoplast fast-activating and slow-activating channel activity is essential for conferring salinity tolerance in a facultative halophyte, quinoa. Plant Physiol. 162, 940–952. doi: 10.1104/pp.113.216572
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bose, J., Shabala, L., Pottosin, I., Zeng, F., Velarde-Buendía, A. M., Massart, A., et al. (2014). Kinetics of xylem loading, membrane potential maintenance, and sensitivity of K+-permeable channels to reactive oxygen species: physiological traits that differentiate salinity tolerance between pea and barley. Plant Cell Environ. 37, 589–600. doi: 10.1111/pce.12180
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bose, J., Xie, Y., Shen, W., and Shabala, S. (2013). Haem oxygenase modifies salinity tolerance in Arabidopsis by controlling K+ retention via regulation of the plasma membrane H+-ATPase and by altering SOS1 transcript levels in roots. J. Exp. Bot. 64, 471–481. doi: 10.1093/jxb/ers343
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, Z., Cuin, T. A., Zhou, M., Twomey, A., Naidu, B. P., and Shabala, S. (2007a). Compatible solute accumulation and stress-mitigating effects in barley genotypes contrasting in their salt tolerance. J. Exp. Bot. 58, 4245–4255. doi: 10.1093/jxb/erm284
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, Z., Newman, I., Zhou, M., Mendham, N., Zhang, G., and Shabala, S. (2005). Screening plants for salt tolerance by measuring K+ flux: a case study for barley. Plant Cell Environ. 28, 1230–1246. doi: 10.1111/j.1365-3040.2005.01364.x
Chen, Z., Pottosin, I. I., Cuin, T. A., Fuglsang, A. T., Tester, M., Jha, D., et al. (2007b). Root plasma membrane transporters controlling K+/Na+ homeostasis in salt-stressed barley. Plant Physiol. 145, 1714–1725. doi: 10.1104/pp.107.110262
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chen, Z., Zhou, M., Newman, I. A., Mendham, N. J., Zhang, G., and Shabala, S. (2007c). Potassium and sodium relations in salinised barley tissues as a basis of differential salt tolerance. Funct. Plant Biol. 34, 150–162. doi: 10.1071/FP06237
Conn, S., and Gilliham, M. (2010). Comparative physiology of elemental distributions in plants. Ann. Bot. 105, 1081–1102. doi: 10.1093/aob/mcq027
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Coskun, D., Britto, D. T., Jean, Y.-K., Kabir, I., Tolay, I., Torum, A. A., et al. (2013). K+ efflux and retention in response to NaCl stress do not predict salt tolerance in contrasting genotypes of rice (Oryza sativa L.). PLoS ONE 8:e57767. doi: 10.1371/journal.pone.0057767
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cuin, T. A., Betts, S. A., Chalmandrier, R., and Shabala, S. (2008). A root's ability to retain K+ correlates with salt tolerance in wheat. J. Exp. Bot. 59, 2697–2706. doi: 10.1093/jxb/ern128
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cuin, T. A., and Shabala, S. (2005). Exogenously supplied compatible solutes rapidly ameliorate NaCl-induced potassium efflux from barley roots. Plant Cell Physiol. 46, 1924–1933. doi: 10.1093/pcp/pci205
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cuin, T. A., and Shabala, S. (2007a). Amino acids regulate salinity-induced potassium efflux in barley root epidermis. Planta 225, 753–761. doi: 10.1007/s00425-006-0386-x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cuin, T. A., and Shabala, S. (2007b). Compatible solutes reduce ROS-induced potassium efflux in Arabidopsis roots. Plant Cell Environ. 30, 875–885. doi: 10.1111/j.1365-3040.2007.01674.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Demidchik, V. (2014). Mechanism and physiological roles of K+ efflux from root cells. J. Plant Physiol. 171, 696–707. doi: 10.1016/j.jplph.2014.01.015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Demidchik, V., and Maathuis, F. J. M. (2007). Physiological roles of nonselective cation channels in plants: from salt stress to signaling and development. New Phytol. 175, 387–404. doi: 10.1111/j.1469-8137.2007.02128.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Demidchik, V., and Tester, M. (2002). Sodium fluxes through nonselective cation channels in the plasma membrane of protoplasts from Arabidopsis roots. Plant Physiol. 128, 379–387. doi: 10.1104/pp.010524
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Flowers, T. J., and Colmer, T. D. (2008). Salinity tolerance in halophytes. New Phytol. 179, 945–963. doi: 10.1111/j.1469-8137.2008.02531.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Galvez, F. J., Baghour, M., Hao, G., Cagnac, O., Rodriguez-Rosales, M. P., and Venema, K. (2012). Expression of LeNHX isoforms in response to salt stress in salt sensitive and salt tolerant tomato species. Plant Physiol. Biochem. 51, 109–115. doi: 10.1016/j.plaphy.2011.10.012
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Garcia de la Garma, J., Fernandez-Garcia, N., Bardisi, E., Pallol, B., Asencio-Rubio, J. S., Bru, R., et al. (2014). New insights into plant salt acclimation: the roles of vesicle trafficking and reactive oxygen species signalling in mitochondria and the endomembrane system. New Phytol. doi: 10.1111/nph.12997. [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gaxiola, R. A., Li, J., Undurraga, S., Dang, L. M., Allen, G. J., Alper, S. L., et al. (2001). Drought- and salt-tolerant plants result from overexpression of the AVP1 H+pump. Proc. Natl. Acad. Sci. U.S.A. 98, 11444–11449. doi: 10.1073/pnas.191389398
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gonzalez, P., Syvertsen, J. P., and Etxeberria, E. (2012). Sodium distribution in salt-stressed Citrus Rootstock seedlings. HortScience 47, 1504–1511.
Hamaji, K., Nagira, M., Yoshida, K., Ohnishi, M., Oda, Y., Uemura, T., et al. (2009). Dynamic aspects of ion accumulation by vesicle traffic under salt stress in Arabidopsis. Plant Cell Physiol. 50, 2023–2033. doi: 10.1093/pcp/pcp143
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hariadi, Y., Marandon, K., Tian, Y., Jacobsen, S. E., and Shabala, S. (2011). Ionic and osmotic relations in quinoa (Chenopodium quinoa Willd.) plants grown at various salinity levels. J. Exp. Bot. 62, 185–193. doi: 10.1093/jxb/erq257
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hmida-Sayari, A., Gargouri-Bouzid, R., Bidani, A., Jaoua, L., Savouré, A., and Jaoua, S. (2005). Overexpression of Δ1-pyrroline-5-carboxylate synthetase increases proline production and confers salt tolerance in transgenic potato plants. Plant Sci. 169, 746–752. doi: 10.1016/j.plantsci.2005.05.025
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Horie, T., Hauser, F., and Schroeder, J. I. (2009). HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci. 14, 660–668. doi: 10.1016/j.tplants.2009.08.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Huang, Z., Zhao, L., Chen, D., Liang, M., Liu, Z., Shao, H., et al. (2013). Salt stress encourages proline accumulation by regulating proline biosynthesis and degradation in Jerusalem Artichoke plantlets. PLoS ONE 8:e62085. doi: 10.1371/journal.pone.0062085
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jaarsma, R., de Vries, R. S. M., and de Boer, A. H. (2013). Effect of salt stress on growth, Na+ accumulation and proline metabolism in potato (Solanum tuberosum) cultivars. PLoS ONE 8:e60183. doi: 10.1371/journal.pone.0060183
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ji, H., Pardo, J. M., Batelli, G., Van Oosten, M. J., Bressan, R. A., and Li, X. (2013). The salt overly sensitive (SOS) pathway: established and emerging roles. Mol. Plant 6, 275–286. doi: 10.1093/mp/sst017
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kishor, P. B. K., Sangam, S., Amrutha, R. N., Laxmi, P. S., Naidu, K. R., Rao, K. R. S. S., et al. (2005). Regulation of proline biosynthesis, degradation, uptake and transport in higher plants: its implications in plant growth and abiotic stress tolerance. Curr. Sci. 88, 424–438.
Kishore, B., Hong, Z., Miao, G., Hu, C., and Verma, D. (1995). Overexpression of delta-pyrroline-5-carboxylatesynthetase increase proline production and confers osmotolerance in transgenic plants. Plant Physiol. 108, 1387–1394.
Lu, Y., Li, N., Sun, J., Hou, P., Jing, X., Zhu, H., et al. (2013). Exogenous hydrogen peroxide, nitric oxide and calcium mediate root ion fluxes in two non-secretor mangrove species subjected to NaCl stress. Tree Physiol. 33, 81–95. doi: 10.1093/treephys/tps119
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lutts, S., Majerus, V., and Kinet, J. M. (1999). NaCl effects on proline metabolism in rice (Oryza sativa L.) seedlings. Physiol. Plant. 105, 450–458. doi: 10.1034/j.1399-3054.1999.105309.x
Maas, E. V., and Hoffman, G. J. (1977). Crop salt tolerance, current assessment. ASCE J. Irrig. Drain. Div. 103, 115–134.
Maathuis, F. J. M. (2014). Sodium in plants: perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 65, 849–858. doi: 10.1093/jxb/ert326
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Malagoli, P., Britto, D. T., Schulze, L. M., and Kronzucker, H. J. (2008). Futile Na+ cycling at the root plasma membrane in rice (Oryza sativa L.): kinetics, energetic, and relationship to salinity tolerance. J. Exp. Bot. 59, 4109–4117. doi: 10.1093/jxb/ern249
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mimura, T., Kura-Hotta, M., Tsujimura, T., Ohnishi, M., Miura, M., Okazaki, Y., et al. (2003). Rapid increase of vacuolar volumen in response to salt stress. Planta 216, 397–402. doi: 10.1007/s00425-002-0878-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Moftah, A. E., and Michel, B. E. (1987). The effect of sodium chloride on solute potential and proline accumulation in soybean leaves. Plant Physiol. 83, 238–240. doi: 10.1104/pp.83.2.238
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Moscone, E. A., Scaldaferro, M. A., Grabiele, M., Cecchini, N. M., Sánchez-García, Y., Jarret, R., et al. (2007). The evolution of chili peppers (Capsicum-Solanaceae): a cytogenetic perspective. Acta Hortic. 745, 137–170. Available online at: http://www.actahort.org/books/745/745_5.htm
Munns, R., and Tester, M. (2008). Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 59, 651–681. doi: 10.1146/annurev.arplant.59.032607.092911
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Newman, I. A. (2001). Ion transport in roots: measurement of fluxes using ion-selective microelectrodes to characterize transporter function. Plant Cell Environ. 24, 1–14. doi: 10.1046/j.1365-3040.2001.00661.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Oh, D.-H., Leidi, E., Zhang, Q., Hwang, S.-M., Li, Y., Quintero, F. J., et al. (2009). Loss of halophytism by interference with SOS1 expression. Plant Physiol. 151, 210–222. doi: 10.1104/pp.109.137802
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Palmgren, M., and Nissen, P. (2010). P-type ATPases. Annu. Rev. Biophys. 40, 243–266. doi: 10.1146/annurev.biophys.093008.131331
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pasapula, V., Shen, G., Kuppu, S., Paez-Valencia, J., Mendoza, M., Hou, P., et al. (2011). Expression of an Arabidopsis vacuolar H+-pyrophosphatase gene (AVP1) in cotton improves drought and salt tolerance and increase fibre yield in the field conditions. Plant Biotechnol. J. 9, 88–99. doi: 10.1111/j.1467-7652.2010.00535.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Perry, L., Dickau, R., Zarrillo, S., Holst, I., Pearsall, D. M., Piperno, D. R., et al. (2007). Starch fossils and the domestication and dispersal of chili peppers (Capsicum spp. L.) in the Americas. Science 325, 986–988. doi: 10.1126/science.1136914
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pittman, J. K. (2012). Multiple transport pathways for medianting intracellular pH homeostasis: the contribution of H+/ion exchangers. Front. Plant Sci. 3:11. doi: 10.3389/fpls.2012.00011
Plett, D. C., and Moller, I. S. (2010). Na+ transport in glycophytic plants: what we know and would like to know. Plant Cell Environ. 33, 612–626. doi: 10.1111/j.1365-3040.2009.02086.x
Qin, C., Yu, C., Shen, Y., Fang, X., Chen, L., Min, J., et al. (2014). Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. U.S.A. 111, 5135–5140. doi: 10.1073/pnas.1400975111
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Queirós, F., Fontes, N., Silva, P., Almeida, D., Maeshima, M., Gerós, H., et al. (2009). Activity of tonoplast proton pumps and Na+/H+ exchange in potato cell cultures is modulated by salt. J. Exp. Bot. 60, 1363–1374. doi: 10.1093/jxb/erp011
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ramani, B., Reeck, T., Debez, A., Stelzer, R., Huchzermeyer, B., Schmidt, A., et al. (2006). Aster tripolium L. and Sesuvium portulacastrum L.: two halophytes, two strategies to survive in saline habitats. Plant Physiol. Biochem. 44, 395–408. doi: 10.1016/j.plaphy.2006.06.007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rodríguez-Rosales, M. P., Gálvez, F. J., Huertas, R., Aranda, M. N., Baghour, M., Cagnac, O., et al. (2009). Plant NHX catión/proton antiporters. Plant Signal. Behav. 4, 265–276. doi: 10.4161/psb.4.4.7919
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rodríguez-Rosales, M. P., Jiang, X., Gálvez, F. J., Aranda, M. N., Cubero, B., and Venema, K. (2008). Overexpression of the tomato K+/H+ antiporter LeNHX2 confers salt tolerance by improving potassium compartmentalization. New Phytol. 179, 366–377. doi: 10.1111/j.1469-8137.2008.02461.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Roy, S. J., Negrão, S., and Tester, M. (2014). Salt resistant crop plants. Curr. Opin. Biotechnol. 26, 115–124. doi: 10.1016/j.copbio.2013.12.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ruan, C.-J., Teixeira da Silva, J. A., Mopper, S., Qin, P., and Lutts, S. (2010). Halophyte improvement for a salinized world. CRC. Crit. Rev. Plant Sci. 29, 329–359. doi: 10.1080/07352689.2010.524517
Rubio, F., Flores, P., Navarro, J. M., and Martínez, V. (2003). Effects of Ca2+, K+ and cGMP on Na+ uptake in pepper plants. Plant Sci. 165, 1043–1049. doi: 10.1016/S0168-9452(03)00297-8
Shabala, S. (2013). Learning from halophytes: physiological basis and strategies to improve abiotic stress tolerance in crops. Ann. Bot. 112, 1209–1221. doi: 10.1093/aob/mct205
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shabala, S., and Cuin, T. A. (2007). Potassium transport and plant salt tolerance. Physiol. Plant. 133, 651–669. doi: 10.1111/j.1399-3054.2007.01008.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shabala, S., Demidchik, V., Shabala, L., Cuin, T. A., Smith, S. J., Miller, A. J., et al. (2006). Extracellular Ca2+ ameriorates NaCl-induced K+ loss from Arabidopsis root and leaf cells by controlling plasma membrane K+-permeable channels. Plant Physiol. 141, 1653–1665. doi: 10.1104/pp.106.082388
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shabala, S., and Pottosin, I. (2014). Regulation of potassium transport in plants under hostile conditions: implications for abiotic and biotic stress tolerance. Physiol. Plant. 151, 257–279. doi: 10.1111/ppl.12165
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shabala, S., and Shabala, L. (2011). Ion transport and osmotic adjustment in plants and bacteria. Biomol. Concepts 2, 407–419. doi: 10.1515/BMC.2011.032
Sharma, S., Villamor, J. G., and Verslues, P. E. (2011). Essential role of tissue-specific proline synthesis and catabolism in growth and redox balance at low water potential. Plant Physiol. 157, 292–304. doi: 10.1104/pp.111.183210
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shen, G., Wei, J., Qiu, X., Hu, R., Kuppu, S., Auld, D., et al. (2014). Co-overexpression of AVP1 and AtNHX1 in cotton further improves drought and salt tolerance in transgenic cotton plants. Plant Mol. Biol. Rep. doi: 10.1007/s11105-014-0739-8. [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shi, H., Quintero, F. J., Pardo, J. M., and Zhu, J.-K. (2002). The putative plasma membrane Na+/H+ antiporter SOS1 controls long-distance Na+ transport in plants. Plant Cell 14, 465–477. doi: 10.1105/tpc.010371
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Smethurst, C. F., Rix, K., Garnett, T., Auricht, G., Bayart, A., Lane, P., et al. (2008). Multiple traits associated with salt tolerance in lucerne: revealing the underlying cellular mechanisms. Funct. Plant Biol. 35, 640–650. doi: 10.1071/FP08030
Sun, J., Dai, S., Wang, R., Chen, S., Li, N., Zhou, X., et al. (2009). Calcium mediates root K+/Na+ homeostasis in poplar species differing in salt tolerance. Tree Physiol. 29, 1175–1186. doi: 10.1093/treephys/tpp048
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Szabados, L., and Savouré, A. (2009). Proline: a multifunctional amino acid. Trends Plant Sci. 15, 89–97. doi: 10.1016/j.tplants.2009.11.009
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Verslues, P. E., and Sharma, S. (2010). Proline metabolism and its implications for plant-environmental interaction. Arabidopsis Book 8:e0140. doi: 10.1199/tab.0140
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Volkov, V., and Amtmann, A. (2006). Thellungiella halophila, a salt-tolerant relative of Arabidopsis thaliana, has specific root ion-channel features supporting K+/Na+ homeostasis under salinity stress. Plant J. 48, 342–353. doi: 10.1111/j.1365-313X.2006.02876.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wu, H., Shabala, L., Barry, K., Zhou, M., and Shabala, S. (2013). Ability of leaf mesophyll to retain potassium correlates with salinity tolerance in wheat and barley. Physiol. Plant. 149, 515–527. doi: 10.1111/ppl.12056
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yildiztugay, E., Sekmen, A. H., Turkan, I., and Kucukoduk, M. (2011). Elucidation of physiological and biochemical mechanisms of an endemic halophyte Centaurea tuzgoluensis under salt stress. Plant Physiol. Biochem. 49, 816–824. doi: 10.1016/j.plaphy.2011.01.021
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zepeda-Jazo, I., Shabala, S., Chen, Z., and Pottosin, I. I. (2008). Na+-K+ transport in roots under salt stress. Plant Signal. Behav. 3, 401–403. doi: 10.4161/psb.3.6.5429
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhang, J.-L., and Shi, H. (2013). Physiological and molecular mechanisms of plant salt tolerance. Photosyn. Res. 115, 1–22. doi: 10.1007/s11120-013-9813-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhao, F., Song, C. P., He, J., and Zhu, H. (2007). Polyamines improve K+/Na+ homeostasis in barley seedlings by regulating root ion channel activities. Plant Physiol. 145, 1061–1072. doi: 10.1104/pp.107.105882
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: salt tolerance, pepper, roots, proline accumulation, sodium compartmentalization, potassium retention, ion fluxes, H+-ATPase
Citation: Bojórquez-Quintal E, Velarde-Buendía A, Ku-González Á, Carillo-Pech M, Ortega-Camacho D, Echevarría-Machado I, Pottosin I and Martínez-Estévez M (2014) Mechanisms of salt tolerance in habanero pepper plants (Capsicum chinense Jacq.): Proline accumulation, ions dynamics and sodium root-shoot partition and compartmentation. Front. Plant Sci. 5:605. doi: 10.3389/fpls.2014.00605
Received: 13 August 2014; Accepted: 17 October 2014;
Published online: 12 November 2014.
Edited by:
Vadim Volkov, London Metropolitan University, UKReviewed by:
Sergey Shabala, University of Tasmania, AustraliaVadim Volkov, London Metropolitan University, UK
Soledad Francisca Undurraga, Universidad Mayor, Chile
Copyright © 2014 Bojórquez-Quintal, Velarde-Buendía, Ku-González, Carillo-Pech, Ortega-Camacho, Echevarría-Machado, Pottosin and Martínez-Estévez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuel Martínez-Estévez, Unidad de Bioquímica y Biología Molecular de Plantas, Centro de Investigación Científica de Yucatán (CICY), Calle 30 # 130, Col. Chuburná de Hidalgo, Mérida 97200, México e-mail:bHVpc21hbmhAY2ljeS5teA==