 Sangram K. Lenka1‡
Sangram K. Lenka1‡ N. Ezekiel Nims1,2†‡
N. Ezekiel Nims1,2†‡ Kham Vongpaseuth2,3†
Kham Vongpaseuth2,3† Rosemary A. Boshar1
Rosemary A. Boshar1 Susan C. Roberts2,3
Susan C. Roberts2,3 Elsbeth L. Walker1,2*
Elsbeth L. Walker1,2*- 1Department of Biology, University of Massachusetts, Amherst, MA, USA
- 2Plant Biology Graduate Program, University of Massachusetts, Amherst, MA, USA
- 3Department of Chemical Engineering, University of Massachusetts, Amherst, MA, USA
Taxus cell suspension culture is a sustainable technology for the industrial production of paclitaxel (Taxol®), a highly modified diterpene anti-cancer agent. The methyl jasmonate (MJ)-mediated paclitaxel biosynthetic pathway is not fully characterized, making metabolic engineering efforts difficult. Here, promoters of seven genes (TASY, T5αH, DBAT, DBBT, PAM, BAPT, and DBTNBT), encoding enzymes of the paclitaxel biosynthetic pathway were isolated and used to drive MJ-inducible expression of a GUS reporter construct in transiently transformed Taxus cells, showing that elicitation of paclitaxel production by MJ is regulated at least in part at the level of transcription. The paclitaxel biosynthetic pathway promoters contained a large number of E-box sites (CANNTG), similar to the binding sites for the key MJ-inducible transcription factor AtMYC2 from Arabidopsis thaliana. Three MJ-inducible MYC transcription factors similar to AtMYC2 (TcJAMYC1, TcJAMYC2, and TcJAMYC4) were identified in Taxus. Transcriptional regulation of paclitaxel biosynthetic pathway promoters by transient over expression of TcJAMYC transcription factors indicated a negative rather than positive regulatory role of TcJAMYCs on paclitaxel biosynthetic gene expression.
Introduction
Paclitaxel (Taxol®; Figure 1) is a diterpene derived from plants in the genus Taxus. Taxol triggers anti-mitotic and cytotoxic activity by disrupting normal tubulin dynamics leading to dysfunction of microtubules (Schiff et al., 1979). Clinical application of Taxol® has been approved by the US Food and Drug Administration for several types of cancer treatment (www.fda.gov). Paclitaxel is also being used in arterial stents to inhibit scar tissue formation after implant (Bajaj and Garratt, 2010), thus the demand for this important compound is expected to increase. Paclitaxel constitutes only 0.01–0.03% of the dry weight of the bark of Taxus and total synthesis comprises several steps, and is therefore low yielding (Fu et al., 2009). Currently paclitaxel and its precursor are primarily derived from the needles of yew plants as well as Taxus suspension cell cultures (Frense, 2007; Vongpaseuth and Roberts, 2007; Kolewe et al., 2008; Ajikumar et al., 2010; Flores-Bustamante et al., 2010). The biosynthetic pathway leading to paclitaxel has been only partially elucidated (Croteau et al., 2006; Ketchum et al., 2007; Long et al., 2008) (Figure 1), and improved understanding of the four or five undefined pathway steps as well as the overall regulation of paclitaxel synthesis are needed in order to enable bioengineering approaches. This will allow enhanced production of paclitaxel and potentially may allow production of novel bioactive taxanes in plant cells.
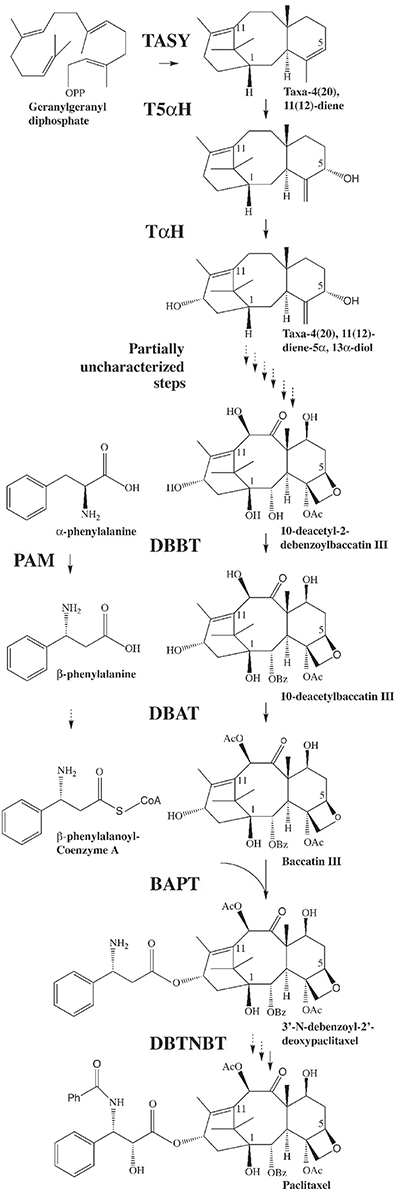
Figure 1. The biosynthetic pathway leading to paclitaxel. GGPP is synthesized by geranylgeranyl diphosphate synthase (GGPPS). GGPP is then converted to taxadiene by taxadiene synthase (TASY) (Wildung and Croteau, 1996) and then to taxadien-5α-ol by taxadien-5α-ol hydroxylase (T5α H) (Jennewein et al., 2004a). Several steps leading from the diol intermediate to functionalized taxanes, including the formation of the oxetane ring, are unknown. Taxane 2α-O-benzoyltransferase (DBBT) (Walker and Croteau, 2000b) produces 10-deacetylbaccatin III (10-DAB). 10-DAB is then converted to baccatin III by 10-deacetylbaccatin III-10-O-acetyltransferase (DBAT) (Walker and Croteau, 2000a). Baccatin III: 3-amino, 3-phenylpropanoyltransferase (BAPT) ligates the side chain [derived from phenylalanine via phenylalanine aminomutase (PAM) (Jennewein et al., 2004b)] to produce 3′-N-debenzoyl-2-deoxypaclitaxel (Walker et al., 2002a). An unknown P450-mediated hydroxylation of the side chain forms 3′-N-debenzoyl paclitaxel. 3′-N-debenzoyl-2-deoxypaclitaxel-N-benzoyltransferase (DBTNBT) then produces paclitaxel by benzoylation of the side chain (Walker et al., 2002b).
Taxus suspension cultures are commonly elicited using exogenous methyl jasmonate (MJ) to produce paclitaxel (Yukimune et al., 1996). The transcript levels of the known paclitaxel biosynthetic pathway genes in Taxus suspension cultures increase within 6 h after elicitation. Transcript levels peak at 12–18 h and then taper off to basal levels within 30 h. Accumulation of the taxane intermediates 10-deacetyl baccatin III and baccatin III (Figure 1) parallels transcript profiles, but with a 12–18 h lag. However, accumulation of paclitaxel occurs much later (3–6 days after elicitation), long after steady state transcript levels of the known pathway genes have returned to normal (Nims et al., 2006; Lenka et al., 2012). The mechanism(s) underlying the delayed correlation between steady state mRNA levels and accumulation of the desired metabolite paclitaxel are not understood. Still, the coordinated expression of the known paclitaxel pathway genes suggests that these genes could be under the control of a common regulatory regime. Generally, MJ application triggers profound transcriptional reprogramming in plant cells to modulate control machineries of a wide range of metabolite biosynthesis by interplay of both positive and negative regulators (Fonseca et al., 2009). Better understanding of the events between MJ application and metabolite accumulation will be useful for developing strategies to enhance production of taxanes in cultured cells.
Although MJ elicits a wide range of species-specific and structurally diverse secondary metabolic pathways of different biochemical origins, it appears that the basic MJ signaling machinery may be conserved in plants of different phylogenetic lineages (De Geyter et al., 2012). Key regulators belonging to the bHLH class of proteins regulating specialized Jasmonic acid (JA) mediated metabolite production have been identified in Catharanthus roseus, Arabidopsis thaliana, Nicotiana tabacum, and Nicotiana benthamiana (van der Fits and Memelink, 2000, 2001; van der Fits et al., 2000; Siberil et al., 2001; Chatel et al., 2003; Dombrecht et al., 2007; Gigolashvili et al., 2007a,b, 2008; Pre et al., 2008; Todd et al., 2010; Qi et al., 2011; Zhang et al., 2011, 2012). MYC2 can act as a positive regulator for several transcription factor (TF) genes and for genes encoding flavonoid biosynthesis pathway, but can also be a negative regulator, for example, of genes involved in indole glucosinolate synthesis (Dombrecht et al., 2007). In the absence of MJ, MYC2 action is repressed by JAZ proteins by forming repressor complexes with a group of other proteins including Novel Interactor of JAZ (NINJA) and TOPLESS (Browse, 2009; Chini et al., 2009). MJ elicitation leads to JAZ degradation by the SCFCOI1–ubiquitin–proteasome pathway releasing MYC2 from the repressor complex. The unbound MYC2 regulates target MJ-responsive genes by binding to E-boxes present in their promoters (Browse, 2009; Chini et al., 2009). The JA signaling mechanism also oscillates through a negative feedback loop involving MYC2 and JAZ proteins, in which JAZ blocks MYC2 activity at the protein level and MYC2 transcriptionally induces JAZ expression (Chini et al., 2007). Hence, upon MYC2 overexpression, MYC2 activity would be influenced by the relative abundance of the activator MYC2 and the JAZ repressor proteins.
In this study we have investigated the role of three MJ-regulated bHLH TFs in Taxus cuspidata. These bHLH potential regulators are named JAMYC (TcJAMYC1, TcJAMYC2, and TcJAMYC4), based on their similarity to Arabidopsis MYC2, which has also been called JAMYC (Lorenzo et al., 2004). The similarity in sequence and expression between TcJAMYC(s) and the well-characterized AtMYC2 suggests a conserved response to MJ despite significant divergence between the gymnosperm and angiosperm lineages. The results presented here suggest that TcJAMYC(s) are regulators of MJ-mediated expression of taxane biosynthetic genes in Taxus cell cultures, but that they function as negative rather than positive regulators of these genes.
Materials and Methods
Cell Culture, MJ Elicitation, and Transient Transformation
Taxol producing Taxus cuspidata P991 suspension cell cultures were used for all experiments and are grown as described previously (Nims et al., 2006). For elicitation, 40 μl of 100% methyl jasmonate was added to 960 μl of 50% ethanol, the modified protocol used as described by (Patil et al., 2013). This solution was then filtered through a 0.2 μm PVDF filter. Cells were either elicited with MJ (dissolved in 50% EtOH) at a 100 μm final concentration or mock-elicited by adding 50% of equal volume EtOH. After 3 h, both batches of cells (~0.5 g) were transferred onto B5-agar plates, spread out in a circle in the center of the plate with a diameter of about 3 cm, and gently pressed into the agar to immobilize them. Within 2 h, the cells were transformed by bombardment using a PDS-1000 (Bio-RAD, Hercules, CA). Preparation of gold microcarriers was performed as described (Vongpaseuth et al., 2007). A minimum of three replicates of each experiment were performed. Forty-eight hours between bombardment and assay was provided to allow for optimal expression of the reporter genes.
GUS and LUC Assays
Transformed cells were ground in lysis buffer (1 ml; 100 mM KHPO4 pH 7.8 with 0.2% Triton X-100) using 3.2 mm chrome steel beads for 10 min at 30 Hz in a Retsch® MM 400 mixer mill (Retsch Inc., Irvine, California). The cell lysate was incubated on ice for 5 min, and the debris was removed by two rounds of centrifugation for 10 min at 16,000 × g. Luciferase and GUS activity of 10 μl of the cell lysate was measured by using Applied Biosystems' Tropix® Dual-Light® assay (Applied Biosystems, Foster City, CA) as per the manufacturer's instructions in a SpectraMax® M5 multi-mode microplate reader (Molecular Devices, Sunnyvale, CA).
Cloning of Paclitaxel Biosynthetic Gene Promoters
The upstream flanking regions of the biosynthetic pathway genes were cloned using inverse-PCR (Ochman et al., 1988) based on the cDNA sequences of DBBT (AF297618), DBAT (AF193765), PAM (AY582743), BAPT (AY563630), and DBTNBT (AF466397). TASY and T5αH promoters were cloned using PCR based on the sequence information of EF153471 and EF153470, respectively. PCR products were cloned into pDESTG221 to form translational fusions to GUS with 5–40 N-terminal amino acid residues of the pathway biosynthetic gene prior to the fusion point with GUS. cis-element analysis of these pathway promoters were performed using PLACE database (http://www.dna.affrc.go.jp/PLACE/) (Higo et al., 1999).
Vector Construction
A vector, pDESTG221, was constructed by modifying pPZP221 (Hajdukiewicz et al., 1994) to contain a Gateway recombination cassette B (Invitrogen, Carlsbad, CA), GUS reporter gene (Jefferson et al., 1987) and NOS terminator. The Taxus promoter DNA fragments were subcloned into pCR8/GW/TOPO vector and transferred to pDESTG221 using Gateway LR Clonase II (Invitrogen, Carlsbad, CA), creating translational fusions to the GUS reporter gene. The TcJAMYC TFs were amplified from the first-strand cDNA of MJ-elicited Taxus cuspidata suspension cell culture and ligated in between the CaMV35S promoter and a NOS 3′ sequence. The 35S:GUS construct was taken from pBI121:GUS and placed into the multiple cloning site of pPZP221 using EcoRI and BamHI restriction sites. The 35S:LUC construct was ligated into pTZ19u.
Degenerate Primer Amplification of TcJAMYC1
The conserved bHLH domain in the JAMYC proteins from Solanum tuberosum (AJ630505) (Boter et al., 2004) and Arabidopsis (At1g32640) (Abe et al., 1997) was used to design degenerate primers with the Consensus-Degenerate Hybrid Oligonucleotide Primers (CODE-HOP) program (Rose et al., 2003). Five primers were chosen, and two (Forward primer oDEGmyc.1 GAGAAGAACCTCTGAATCATGTTGARGCNGARMG; Reverse primer oDEGmyc.3 CAGCTTACATTTCAGTTCATTAATATAAGAAATNGCRTCNCC) produced a PCR product that was then used to screen a cDNA library by hybridization.
Cloning of TcJAMYC1, TcJAMYC2, and TcJAMYC4
Total RNA was extracted from two grams of Taxus cuspidata cell culture line P991, by guanidium isothiocyanate and cesium chloride gradient ultracentrifugation at 104,000 × g for 18 h, followed by phenol-chloroform extraction. Poly-A RNA (5 μg) was obtained from 1 mg total RNA using Poly-A Purist Mag-Kit (Ambion, Austin, TX). cDNA construction and cloning was performed using the ZAP Express cDNA Synthesis Kit (Agilent Technologies, Cedar Creek, TX). Plaques (1 × 106) from the primary library were screened using the Taxus bHLH fragment obtained by PCR. A 2.5 kb cDNA clone was isolated and sequenced, but the 5′-end was truncated. 5′-Rapid Amplification of cDNA Ends (RACE) was performed using RLM-RACE kit (Ambion, Austin, TX). The full-length cDNA of TcJAMYC1 (FJ608574) was cloned by PCR using the 5′-sequence obtained by RACE, and this product was cloned and sequenced. TcJAMYC2 (JX519289) and TcJAMYC4 (JX519290) were cloned by using sequence specific primers designed by mining an in-house 454 GS FLX sequencing dataset.
Phylogenetic Analysis
Multiple sequences alignments of the full length bHLH proteins were performed using ClustalW with default settings. Bootstrap method for estimating the standard error is used to plot the phylogram. The unrooted phylogenic tree was generated using MEGA5 (Tamura et al., 2011) by the Neighbor-Joining algorithm and values at branch-nodes are percentages of 1000 bootstrap repetitions. Also included in the analysis are the previously reported bHLH TFs recruited by JA signaling to induce secondary metabolite biosynthesis such as AtMYC2 (NM102998), CrMYC2 (AF283507), NtMYC2a (HM466974), NtMYC2b (HM466975), NbNbbHLH1 (GQ859152), NbNbbHLH2 (GQ859153), AtGL3 (NM148067), AtEGL3 (NM105042), AtTT8 (NM117050), AtbHLH28 (AT5G46830), AtMYC3 (At5G46760), and AtMYC4 (AT4G17880).
TcJAMYC1 Protein Purification
The TcJAMYC1 cDNA was recombined into pDEST17 (Invitrogen), an E. coli expression vector containing an N-terminal 6X His tag for affinity purification on a Nickel agarose column (Qiagen, Valencia, CA). The Rosetta 2(DE3) pLysS (EMD biosciences, Gibbstown, NJ) strain of E. coli containing this construct was grown to late log phase (OD600 = 0.8), induced with 1 mM IPTG, and then incubated with shaking for four more hours at 37°C. Cells were pelleted by centrifugation at 4400 × g, resuspended in 50 mM Tris-Cl pH 6.8, 20 mM β-mercaptoethanol, 2% SDS, 10% glycerol, and 10 mM imidazole, and 50 μl DNase1 (10 mg/ml), then incubated on ice for 30 min. Debris was removed by centrifugation at 17,000 × g for 20 min, and the TcJAMYC1 protein was bound to Ni-NTA resin (Qiagen, Valencia, CA). The resin was washed with buffer containing 250 mM NaCl, 50 mM Tris-Cl pH 6.8, 20 mM imidazole, and eluted with buffer containing 250 mM NaCl, 50 mM Tris-Cl pH 6.8, 300 mM imidazole. The eluted protein was further purified using a Centricon YM-3 centrifugal filter device (Millipore, Danvers, MA) and brought to a final protein concentration of 25 ng/μl in 50% glycerol.
Electrophoretic Mobility Shift Assays (EMSA)
Oligonucleotide probes (see Table 1) contained a six nucleotide E-box at the center of a 22 bp sequence. A four-nucleotide 5′-overhang was included in each double stranded probe to allow for 32P-labeling. Labeling reactions contained 100 mM Tris-HCl, 50 mM NaCl, 10 mM MgCl2, 0.025 % Triton X-100, pH 7.5, 2.2 mM of each dTTP, dATP, and dGTP and 22 μm double stranded oligo in a 23 μl total volume with 5 U of Klenow large fragment DNA polymerase (New England Biolabs, Ipswich, MA). Unincorporated nucleotides were removed using a 2 ml Sephadex G-25 (Sigma, St. Louis, MO) column. For each EMSA reaction, 175 fmol of double stranded oligonucleotide were used (labeling reaction diluted 1:125). EMSA buffer consisted of 20 mM HEPES-KOH pH 7.9, 20% glycerol, 0.2 mM EDTA, 100 mM KCl, 0.5 mM PMSF, and 1 mM DTT, 15 mM MgCl2, and 5 μg BSA. For standard reactions, 2 μl of TcJAMYC1 protein was used and the final volume of the reactions was 20 μl. Native polyacrylamide gel (7% acrylamide (29:1), 1% glycerol, 0.5X TBE) electrophoresis was used to separate the DNA probe that was bound by the protein and the free DNA probe. Gels were run at 4°C for 1.5 h at 82 V after being pre-run for 30 min.

Table 1. Oligonucleotide probes used for EMSA.
Sub-Cellular Localization of TcJAMYC1
A C-terminal translational fusion was made by cloning in frame TcJAMYC1 cDNA and soluble modified GFP (smGFP) into the psmGFP vector (CD3-326; Arabidopsis Biological Resource Center) under the control of a CaMV35S promoter. The 35S::TcJAMYC1::GFP fusion construct was then transiently expressed in intact Arabidopsis mesophyll protoplasts as per the tape-Arabidopsis sandwich method (Wu et al., 2009). Protoplasts were imaged by a Zeiss 510 Meta laser scanning confocal microscope (Nikon) with excitation at 488 nm, and the fluorescence emission signal was recovered between 520 and 550 nm.
Results
MJ Induces Paclitaxel Pathway Gene Promoters
Transcript profiling studies in Taxus cultured cells previously demonstrated the MJ-induced mRNA accumulation of the paclitaxel biosynthetic pathway genes (hereafter referred to as “pathway genes”) (Nims et al., 2006; Lenka et al., 2012). To investigate MJ-responsive promoter activity of pathway genes, promoters of seven known pathway genes viz. taxadiene synthase (TASY; JF338876), taxadien-5α-ol hydroxylase (T5αH; JF338877), taxane 2α-O-benzoyltransferase (DBBT; JF338880), 10-deacetylbaccatin III-10-O-acetyltransferase (DBAT; FJ603641), phenylalanine aminomutase (PAM; FJ603642), baccatin III: 3-amino, 3-phenylpropanoyltransferase (BAPT; JF338879) and 3′-N-debenzoyl-2-deoxypaclitaxel-N-benzoyltransferase (DBTNBT; FJ603644) were cloned. Constructs containing a β-glucuronidase (GUS) reporter gene driven by each of these promoters were made (Figure 2A), and were delivered into the Taxus cuspidata P991 culture cells for transient reporter gene expression by particle bombardment. The cells were either mock-elicited or elicited with MJ at a final concentration of 100 μm. 35S:LUC was co-bombarded in all experiments as control for variable transformation efficiency (Figure 2B). Reporter gene activity is presented as a GUS/LUC ratio to control for transformation efficiency between samples. For each pathway gene promoter construct, GUS enzyme activity was at least 1.5-fold higher in MJ-elicited cells compare to the control (Figure 3). Thus, we concluded that the promoters of the pathway genes are activated by MJ. This result, in combination with increased amount of steady state mRNA for the endogenous genes after MJ elicitation (Nims et al., 2006; Lenka et al., 2012; Li et al., 2012b), implies that MJ treatment causes increased transcription of pathway genes.
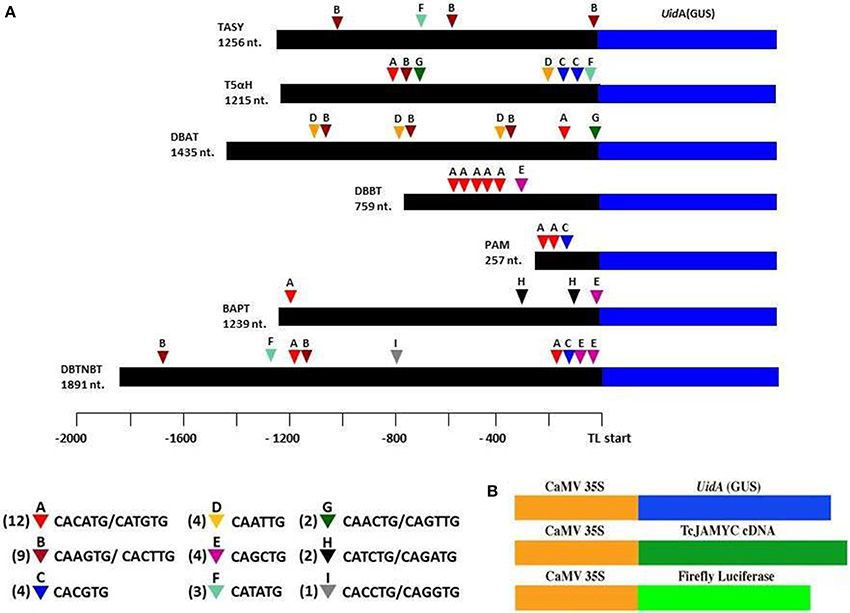
Figure 2. The pathway promoters cloned using inverse-PCR and the locations of the various E-box elements found in all seven gene promoter sequences. (A) Numbers next to the triangles indicate the frequency of that E-box element. Promoter regions were translationally fused to GUS so that 5–40 codons after the ATG of the paclitaxel pathway gene are encoded as an N-terminal extension on the GUS protein. (B) The GUS TcJAMYC(s) full-length cDNA, and firefly luciferase (LUC) genes, all driven by the Cauliflower Mosaic Virus (CaMV) 35S promoter, that were used in bombardment assays.
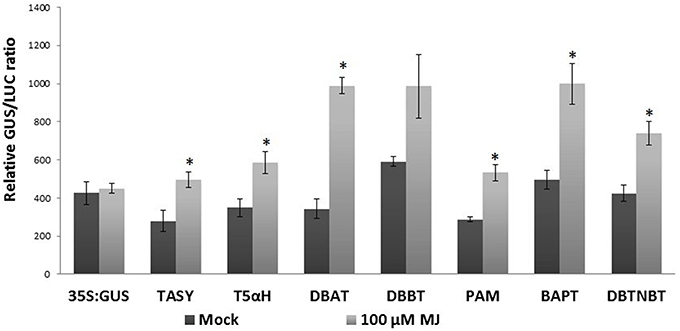
Figure 3. Pathway promoter activation by MJ. Taxus suspension cultures were plated onto B5 agar plates and bombarded with the promoter:GUS reporter constructs. 35S:GUS was also bombarded as control. Forty-eight hours was allowed between bombardment and assaying the GUS activity. Results are presented as a GUS/LUC ratio. n = 3 in all samples, error bars are SE (*p < 0.05, t-test).
Promoter Analysis
Since the pathway gene promoters were activated upon MJ-elicitation and since all the pathway genes respond in a uniform time course (Nims et al., 2006; Li et al., 2012b), there could be an MJ-responsive transcriptional regulatory mechanism that influences these promoters. Considering this, an in silico analysis of the pathway promoters was performed using PLACE (http://www.dna.affrc.go.jp/PLACE/) (Higo et al., 1999). This analysis revealed that there are 37 E-boxes (CANNTG) found in the 8052 nt of the collective pathway gene promoter sequence that we cloned (Figure 2A). A disproportionately large number of potential E-box sites are found in the promoters of the pathway genes, since only 31 E-boxes would be expected to occur at random in a sequence of this length. E-boxes have been found in defense gene promoters in other plants (Kim et al., 1992). bHLH proteins typically bind to these E-box nucleotide motifs (Qian et al., 2007). For example, the Arabidopsis bHLH MYC2 protein preferentially binds to the E-box sequence CACGTG. Furthermore, E-boxes are commonly found on the promoters of genes that respond to MJ (Dombrecht et al., 2007; Montiel et al., 2011). The Solanum JAMYC2 binds a related, “T/G-box” sequence (AACGTG) on the proteinase inhibitor promoter (Boter et al., 2004). The T/G-box was found in only four of the Taxus gene promoters (DBBT (twice), TASY, DBAT, and DBTNBT). Hence, the over-representation of E-box sites on the cloned promoters suggests the MJ-mediated transcriptional regulation of pathway promoters could be imparted by jasmonate-responsive bHLH MYC proteins.
Cloning TcJAMYC1, TcJAMYC2, and TcJAMYC4
The well-conserved bHLH sequences from Arabidopsis MYC2 (At1g32640) and Solanum JAMYC10 (AJ630506) were used to design degenerate primers with the CODE-HOP program (Rose et al., 2003). These primers were used to amplify a 172 nt fragment from Taxus cDNA derived from suspension cultures that had been elicited with MJ for 6 h (Figure 4A). The amplified sequence displayed 94% identity at the amino acid level to the Solanum JAMYC bHLH region. Only one gene was cloned using this set of degenerate primers. The Taxus fragment was used to obtain a full-length cDNA by screening a λ cDNA library derived from MJ-elicited cells (see Materials and methods). The protein encoded by the full-length cDNA possesses 43% overall identity with the Solanum JAMYC, and 59% overall similarity (Figure 4A).
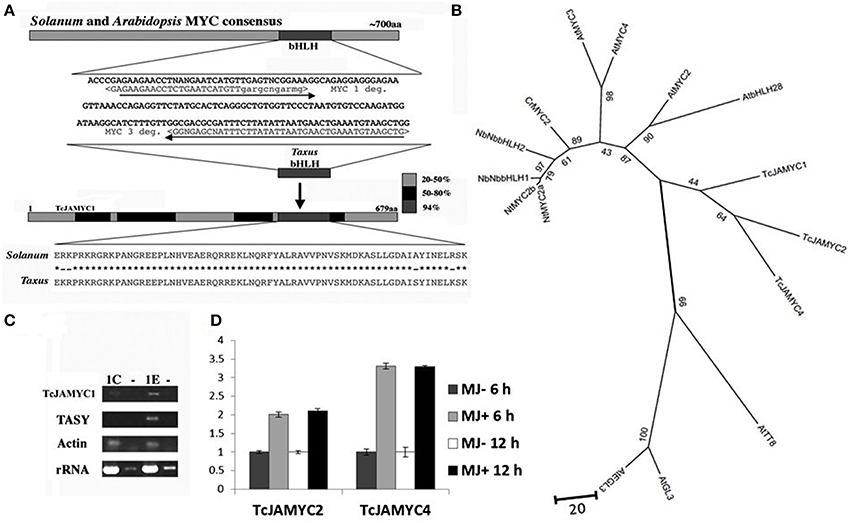
Figure 4. Cloning of TcJAMYC1. (A) Degenerate primers were designed from the conserved bHLH region in order to amplify a Taxus DNA fragment. This fragment was used to obtain a full-length cDNA. (B) The relationship between the three TcJAMYCs and 12 other known bHLH protein sequences. The tree was created by using 12 known bHLH proteins recruited by JA signaling for enhanced production of secondary metabolite biosynthesis in other plants along with the TcJAMYCs protein. This is an unrooted Neighbor-Joining phylogram and values at branch-nodes are percentages of 1000 bootstrap repetitions. (C) RT-PCR analysis of TcJAMYC1, Taxadiene synthase (TASY, AY424738) rRNA (AF259290), and actin (derived from P. contorta actin; Genbank M36171.1) after 1 h of MJ elicitation. Both rRNA and actin fragments were amplified as internal controls. The control (mock elicited) cells are labeled 1C and the MJ-elicited cells are labeled 1E. The lanes labeled (–) represent the results of amplification of RNA without reverse transcription, as a control for contaminating genomic DNA in our RNA preparations. (D) qRT-PCR analysis of TcJAMYC2 and TcJAMYC4 after 6 and 12 h of MJ elicitation. Induction values for target genes normalized to the endogenous controls Taxus actin (JF735995). Error bars are SD.
To clarify the relationship of these Taxus genes to the bHLH genes induced by JA signaling for enhanced production of secondary metabolite biosynthesis in plants (De Geyter et al., 2012), an unrooted phylogram was generated from the Taxus MYC and 12 other known bHLH protein sequences using the Neighbor-Joining method (Figure 4B). The Taxus bHLH falls into the same clade as AtMYC2, confirming the similarity between these genes. Based on the similarity to the other well-characterized JAMYC bHLH transcription factors involved in JA mediated production of secondary metabolite, we refer to this Taxus gene as TcJAMYC1 for Taxus cuspidata jasmonate MYC1 (TcJAMYC1).
Previously characterized JA-MYC genes from tomato and Arabidopsis are positively regulated by MJ with increased mRNA levels upon MJ addition (Boter et al., 2004; Dombrecht et al., 2007). To determine whether TcJAMYC1 responds to MJ application, semi-quantitative RT-PCR was performed to compare relative transcript abundance at 1 h after MJ elicitation (Figure 4C). This result indicates that there is an increase in TcJAMYC1 transcripts after MJ elicitation, again emphasizing the resemblance between TcJAMYC1 and the Arabidopsis JAMYC, AtMYC2. To find other members of JAMYC family in Taxus, we searched for similar cDNA sequences in a recently generated 454 GS FLX Taxus transcriptome sequence dataset (Lenka and Walker, unpublished). Similar to the TcJAMYC1 mRNA, TcJAMYC2 and TcJAMYC4 also showed MJ-induced transcript accumulation in Taxus culture cells as measured by quantitative PCR both at 6 and 12 h (Figure 4D). Two other partial MYC cDNAs were identified from the Taxus transcriptome sequencing data (TcJAMYC3 and TcJAMYC5), but, these two MYC transcripts were not inducible by MJ (data not shown). Cloning of TcJAMYC2 and TcJAMYC4 was carried out using sequence specific primers designed from cDNA contigs.
TcJAMYC1 Binds to Pathway Promoters in vitro and is Localized to the Nucleus
To determine whether the TcJAMYC protein physically interacts with the E-box elements found in the pathway promoters, electrophoretic mobility shift assays (EMSA) were performed using one of the TF proteins TcJAMYC1. EMSAs are used to determine whether a protein can bind to a specific DNA probe sequence in vitro. This binding is visualized as a retarded migration rate through a native polyacrylamide gel. 6X-HIS tagged TcJAMYC1 was expressed in E. coli and purified on Ni-NTA agarose columns. The molecular weight for the tagged TcJAMYC1 protein was 73 kD, as expected (Figure 5A). To determine whether TcJAMYC1 binds to E-box elements, the most abundant E-box (CATGTG) in the pathway promoters was used initially as a probe sequence (Table 1). The TcJAMYC1 protein bound the CATGTG sequence (Figure 5B). To determine if the binding was specific to TcJAMYC1, the GUS protein, expressed in and purified from E. coli (Figure 5A), was used in the EMSA. This assay demonstrated that the binding of the CATGTG sequence is specific to the TcJAMYC1 protein, as GUS did not bind to the DNA probe (Figure 5B). Finally, a mutated DNA probe that does not contain an E-box (Table 1) was used in the EMSA (Figure 5C). The TcJAMYC1 protein did not bind to this DNA sequence, demonstrating that the TcJAMYC1 protein specifically binds to the E-box element CATGTG.
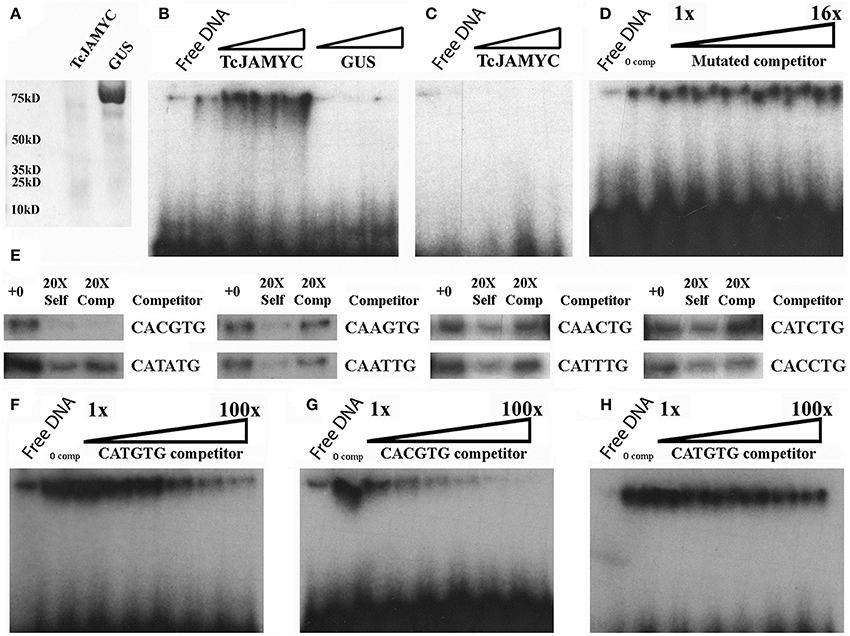
Figure 5. Electrophoretic mobility shift binding assays for TcJAMYC. (A) The TcJAMYC1 and the GUS proteins on a Coomassie blue-stained SDS page gel after nickel column purification. (B) EMSA using the TcJAMYC1 (left to right: ~25, 50, 75, 100 ng) and GUS (left to right: ~25, 50, and 75 ng) proteins at increasing amounts using the most common E-box element found on pathway promoters (CATGTG) as probe. (C) Binding assay using TcJAMYC1 at increasing amounts (left to right: ~25, 50, and 75 ng) and a mutated probe (Table 1). (D) Competition assay with the TcJAMYC1 protein and the CATGTG probe against increasing amounts of mutated cold competitor at 0X, 1X, 2X, 4X, 8X, and 16X excess. (E) Competition assays using TcJAMYC1 protein and the CATGTG radio labeled probe against all other E-box elements found in the pathway promoters. The cold competitor is listed to the right of the panels. +0: no competitor, 20X Self: the CATGTG cold competitor at 20X excess, 20X Comp: the cold competitor at 20X excess. (F) TcJAMYC1 self-competition assay (to determine binding affinity to CATGTG). The 1–100X range of cold competitor is 1X, 5X, 10X, 20X, 50X, and 100X excess, left to right in each gel. (G) A competition assay between radio labeled CATGTG against the CACGTG cold competitor. (H) A competition assay between radio labeled CACGTG against the CATGTG cold competitor.
To test the specificity of binding further, a competition assay was performed using the CATGTG probe against increasing amounts of non-specific DNA competitor (the mutated DNA described above). The binding efficiency to the CATGTG probe did not diminish (Figure 5D), again demonstrating that the TcJAMYC1 protein specifically binds to the CATGTG sequence. Eight additional types of E-box sequences were found in the pathway promoters (Figure 2A). To determine whether TcJAMYC1 also binds these E-boxes, we tested oligonucleotides containing seven of these sequences in competition assays against the CATGTG probe. The binding efficiency of TcJAMYC1 toward the CAGCTG E-box element was not examined. As shown in Figure 5E, the only E-box element that competed well-against the CATGTG probe was the CACGTG element. This demonstrates that the TcJAMYC1 protein preferentially binds to two specific E-box sequences found in the pathway gene promoters: CACGTG and the CATGTG.
To test which of these two sequences, CATGTG or CACGTG, is most efficiently bound by TcJAMYC1, competition assays between these two DNA elements were performed. The CATGTG probe competed against itself, as previously shown in Figure 5E, and this was used as reference for binding specificity (Figure 5F). A competition assay was performed using the CATGTG probe against increasing amounts of CACGTG competitor (Figure 5G). The CACGTG fragment competed for binding of the CATGTG probe more effectively than CATGTG self-competitor (compare Figures 5F,G). This demonstrates that TcJAMYC1 preferentially binds to the CACGTG as compared to the CATGTG sequence. To further characterize this binding selectivity, a competition assay was performed with the CACGTG probe against the unlabeled CATGTG competitor (Figure 5H). Increasing amounts of CATGTG competitor did not efficiently compete for binding, confirming the preference for binding of CACGTG. Thus, the TcJAMYC1 protein specifically binds to CACGTG and secondarily the CATGTG E-box elements.
Nuclear localization of TcJAMYC1 was confirmed by co-localization of TcJAMYC1:GFP fusion protein in the transformed Arabidopsis protoplast nucleus (Supplementary Figures S1 A–J). This evidence suggests that TcJAMYC1 recruited to nuclear compartment might regulate pathway genes by specific interaction through the E-box elements.
Regulation of Pathway Promoters by TcJAMYCs
To investigate whether TcJAMYCs influence transcription of the paclitaxel pathway genes, co-bombardment experiments were performed using each of the three full-length TcJAMYC cDNA under the control of the CaMV35S promoter in combination with the pathway gene promoter:GUS reporter constructs (Figures 2A, 6A–F, Table 2). Mock-elicited cells were bombarded with each pathway gene promoter:GUS construct either alone or in combination with separate 35S:TcJAMYC constructs. TcJAMYC1 negatively regulated DBBT, BAPT, and DBTNBT promoters by at least 3-fold, while all the other promoters were unaffected by co-bombardment with TcJAMYC1 (Table 2, Figure 6A). This demonstrates that TcJAMYC1 negatively regulates the promoters of the last three late pathway genes and does not affect any of the early or intermediate pathway gene promoters. On the other hand TcJAMYC2 induces the early pathway gene promoter T5αH, by more than 1.5-fold. TcJAMYC2 also induces PAM promoter by more than 2.5-fold. Expression of BAPT is slightly repressed by the TcJAMYC2 action and regulation of TASY, DBAT, DBBT, and DBTNBT promoters were not significantly influenced by transient over-expression of TcJAMYC2 in Taxus cultured cells (Table 2, Figure 6C). TcJAMYC4 has not much regulatory impact on the T5αH gene promoter; while all other pathway promoters were down regulated by TcJAMYC4 transient over-expression except TASY, which was weakly induced (Table 2, Figure 6E).
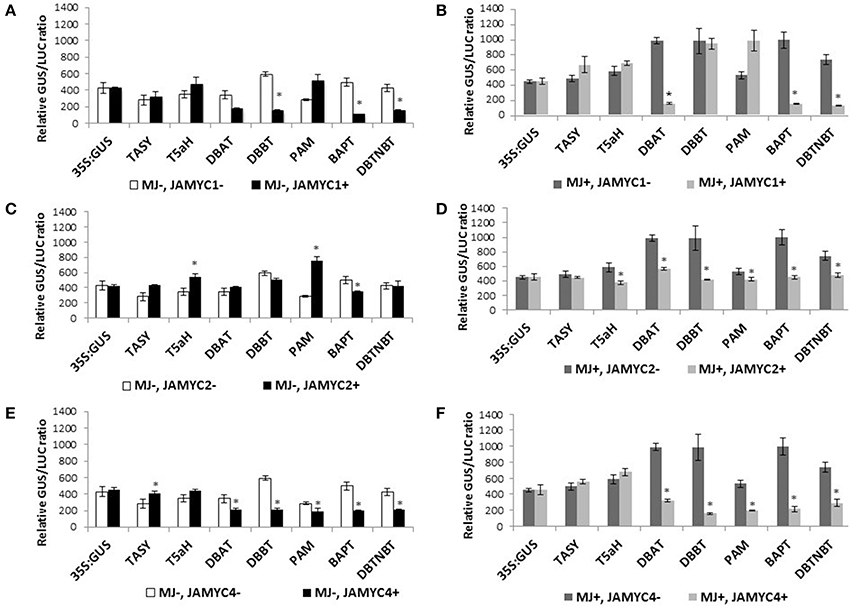
Figure 6. (A–F) Promoter activation by TcJAMYC(s). Mock-elicited Taxus cultures were plated onto B5 agar plates and bombarded with the promoter:GUS fusions alone (−MJ, −MYC) or combination with the CaMV35S:TcJAMYCs effector plasmid (−MJ, +MYC) separately. MJ-elicited Taxus cultures were plated onto B5 agar plates and bombarded with the promoter:GUS fusions alone (+MJ, −MYC) or in combination with the CaMV35S:TcJAMYC effector plasmid (+MJ, +MYC). Results are presented as a GUS/LUC ratio. n = 3 in all samples, error bars are SE (*p < 0.05, t-test).
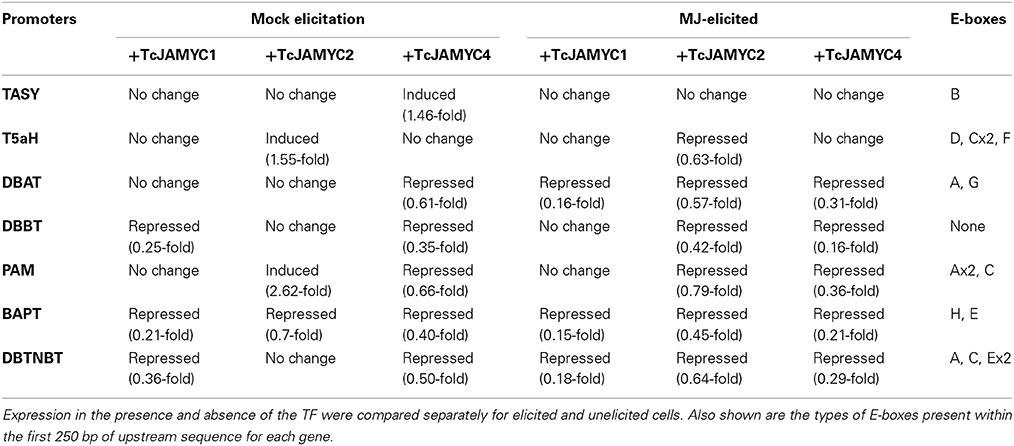
Table 2. Changes in promoter activity caused by co-expression of TcJAMYC TF.
To determine the effect of MJ elicitation on the promoter-TcJAMYC interaction, co-bombardment experiments were performed using cells that had been elicited with 100 μm MJ 6 h prior to bombardment. The elicited cells were bombarded with each promoter:GUS construct alone or in combination with the separate 35S:TcJAMYCs. The DBAT, BAPT, and DBTNBT promoters were highly repressed following co-bombardment with the 35S:TcJAMYC1 effector despite MJ elicitation (Table 2, Figure 6B). The initial two pathway genes, TASY, T5αH, and PAM along with the DBBT promoter were not significantly regulated with MJ elicitation when co-bombarded with the 35S:TcJAMYC1 effector (Table 2, Figure 6B). However, transient over-expression of TcJAMYC2 led to a decrease in activity of all the pathway promoters when MJ elicitation was applied prior to bombardment, except TASY, which was unaffected (Table 2, Figure 6D). Similarly, TcJAMYC4 also negatively regulated all the pathway promoters except the first two early pathway gene promoters TASY and T5αH, whose activity remain unchanged (Table 2, Figure 6F). Thus, expression of single TcJAMYCs in elicited cells most often repressed pathway gene expression.
Discussion
Transcriptional Activation of Pathway Promoters
MJ has been shown to induce terpene production in conifers (Hudgins et al., 2004), glucosinolates in Arabidopsis (Mikkelsen et al., 2003; Mewis et al., 2006), alkaloids in Papaver (Facchini and Park, 2003), Catharanthus (Menke et al., 1999), and Nicotiana (Shoji et al., 2000), as well as proteinase inhibitors in many plant species including important agricultural crops (Farmer and Ryan, 1990; Bolter, 1993; Ton et al., 2007). MJ elicitation has also been implicated in the activation of the defense related leucine aminopeptidase promoter in Nicotiana, the strictosidine synthase promoter in Catharanthus, and a sesquiterpene synthase promoter in Nicotiana are activated with MJ elicitation (Yin et al., 1997; Menke et al., 1999; Boter et al., 2004). Exogenously added MJ also induces the production of paclitaxel and baccatin III in Taxus cell suspension culture (Yukimune et al., 1996). In this investigation, we have shown that seven paclitaxel biosynthetic pathway gene promoters are likewise activated by MJ elicitation in Taxus cuspidata P991 suspension cell cultures. This result explains, at least in part, the previously observed increase in mRNA abundance for these genes that follows MJ elicitation (Nims et al., 2006; Li et al., 2012b), and indicates that the transcriptional response to MJ is conserved across land plant phylogeny from gymnosperms to angiosperms. The coordinated MJ inducible regulation of all the pathway promoters may indicate the presence of a common transcriptional regulatory mechanism for defense-related biosynthetic pathway genes.
Three Jasmonate-Responsive bHLHs in Taxus
In the model plant Arabidopsis thaliana, MYC2 acts as a versatile regulator that is capable of both positive and negative regulation of particular pathways, including the transcriptional orchestration of other TFs (Dombrecht et al., 2007). By leveraging information about MYC2 obtained in model systems (Arabidopsis and Solanum) we have cloned TcJAMYC1, TcJAMYC2, and TcJAMYC4 from T. cuspidata, which produces the important pharmaceutical paclitaxel. Over-expression of particular TcJAMYCs caused positive regulation of several paclitaxel pathway gene promoters. However, most promoters (DBAT, DBBT, BAPT, and DBTNBT) were down regulated or not influenced by the individual action of the three TcJAMYC TFs (Table 2, Figures 6A–F). No strong correlation between the presence of specific E-box binding site types and the activation of particular promoters was noted. By contrast, CAGCTG was found only in those promoters (DBBT, BAPT, and DBTNBT) that were negatively regulated when co-expressed with TcJAMYC1 and TcJAMYC4 (Table 2, Figures 2A, 6A–F). Possibly, the TcJAMYCs act as heterodimers, and the effect of overexpressing a single TcJAMYC could be to disrupt the normal formation of MYC dimers, to produce a negative effect on gene expression. Alternatively, the role of TcJAMYCs in the regulation of particular paclitaxel synthesis steps may be negative. A limitation of the Taxus system is that efficient stable transformation is not yet standardized for most of the commonly used, paclitaxel-producing cell lines, such as the P991 cell line used in this study. This limits our ability to test combinations of potentially heterodimeric TcJAMYCs and to determine the effect of TcJAMYCs on the native gene activation in stably transformed cultures.
Negative Regulation of Pathway Promoters Following MJ Elicitation
Arabidopsis thaliana MYC2 negatively regulates some JA responsive genes and it negatively regulates its own expression (Dombrecht et al., 2007; Chico et al., 2008; Gangappa et al., 2010). In the studies presented here, we uncovered evidence for a negative regulation of the promoters of the paclitaxel biosynthetic genes by TcJAMYC proteins. In previous studies, we demonstrated that steady state mRNA levels for pathway genes increase by 6 h after MJ elicitation, remain high for about 24–30 h, and return to original low levels by about 48 h (Nims et al., 2006). This rapid up- then down-regulation of the endogenous genes indicates positive regulation for the first 24 h following elicitation, followed by negative regulation after the 24 h time point. In the studies presented here, the level of reporter gene expression was always measured 48 h after bombardment, which is ~54 h after MJ elicitation. Thus, when MJ elicitation was used, measurements were taken at a time when endogenous negative regulatory mechanisms are likely to be operative. This may help to explain the relatively weak levels of promoter activation (1.6- to 2.8-fold) that we observed from all the reporter constructs as opposed to the very strong enhancement of steady state RNA observed in RNA gel blot (Nims et al., 2006) and RNA-seq experiments (3.3- to 6.8-fold) (Li et al., 2012b). The measurements were made at this relatively late time because, following transformation by bombardment, sufficient time must be provided to allow transcription and translation of both the effector construct (TcJAMYCs) and the reporter constructs. Furthermore, reporter activity was not consistent at time points prior to 48 h post-bombardment, preventing use of earlier time points for measurements.
In no case did expression of a TcJAMYC cause enhanced reporter expression in MJ-elicited cells. Indeed, in most cases, reporter gene expression was negatively impacted by co-expression of a TcJAMYC. The most straightforward interpretation of this result is that TcJAMYCs are involved in the negative regulation of pathway genes that occurs 24–48 h after MJ elicitation. Possibly, when TcJAMYCs are overexpressed in MJ-elicited cells, negative regulation is enhanced, and lower reporter gene expression is observed. Although this is not the desired result from a metabolic engineering perspective, it could be an indication that TcJAMYCs directly regulate pathway promoters, presumably through binding to E-box elements they contain. Further efforts to introduce silencing of TcJAMYC1, 2, and 4 by MJ-inducible RNAi expression may knock down its negative regulation on late pathway genes and thus, may enable increased late taxane accumulation. An alternative explanation could be that TcJAMYCs positively regulate TFs that in turn negatively regulate pathway promoters, in much the same way that Arabidopsis MYC2 regulates expression of ERF11, At1g33760, and WRKY26 to impart negative regulation on several promoters (Dombrecht et al., 2007). In this scenario, overexpression of TcJAMYCs stimulates further the natural down-regulation of pathway genes that follows their initial activation by MJ, and leads to the observed decrease in reporter gene expression. Despite the seemingly conflicting or minimal effects from individual MYC proteins, it is evident that the MYC transcription factors directly bind to and affect, albeit negatively, the activation of promoters driving expression of this important medicinal plant biosynthetic pathway.
Other Transcription Factors Potentially Regulating MJ-Responsive Taxus Genes
MJ-mediated transcriptional regulation of entire secondary pathways is not likely to be orchestrated by the action of a single TF. Combinatorial action of AP2–ERF and bHLH factors has already been shown in the JA mediated elicitation of nicotine and alkaloid biosynthesis (Zhang et al., 2011; De Geyter et al., 2012). Several other classes of TFs such as WRKYs and MYBs also have been shown to regulate JA-induced responses (Fonseca et al., 2009). Recently, an MJ-inducible WRKY transcription factor involved in promoter activation of the paclitaxel pathway gene encoding DBAT was described (Li et al., 2012a,b). The biosynthetic gene promoters described here contain putative WRKY binding sites (TGAC) on both the plus and minus strands, although not a frequencies higher than expected by chance. Thus, these promoters may be regulated by this transcription factor. Discovering the key TFs uncoupled from negative regulators and capable of partially mimicking MJ activity by activating expression of specific sets of genes is a key challenge for biologically sustainable metabolite production.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Dr. Donna Gibson at the U.S. Plant Soil and Nutrition laboratory, USDA (Ithaca, NY) for providing Taxus cuspidata suspension cultures and Dr. Jennifer Normanly for her support and advice. This work was funded by NIH grant #S11100007200000 to ELW, JN, and SCR and by two UMass Plant Biology Graduate Program Gilgut Fellowships to EN and KV.
Supplementary Material
The Supplementary Material for this article can be found online at: http://www.frontiersin.org/journal/10.3389/fpls.2015.00115/abstract
Supplementary Figure S1. Sub-cellular localization of TcJAMYC1. 35S::TcJAMYC1::GFP fusion construct was transiently expressed in intact Arabidopsis mesophyll protoplasts and were imaged by a Zeiss 510 Meta laser scanning confocal microscope.
References
Abe, H., Yamaguchi-Shinozaki, K., Urao, T., Iwasaki, T., Hosokawa, D., and Shinozaki, K. (1997). Role of arabidopsis MYC and MYB homologs in drought- and abscisic acid-regulated gene expression. Plant Cell 9, 1859–1868.
Ajikumar, P. K., Xiao, W. H., Tyo, K. E., Wang, Y., Simeon, F., Leonard, E., et al. (2010). Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 330, 70–74. doi: 10.1126/science.1191652
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bajaj, N. R., and Garratt, K. N. (2010). Clinical studies with paclitaxel – eluting stent systems. Curr. Pharm. Des. 16, 4025–4036. doi: 10.2174/138161210794454860
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bolter, C. J. (1993). Methyl jasmonate induces papain inhibitor(s) in tomato leaves. Plant Physiol. 103, 1347–1353.
Boter, M., Ruiz-Rivero, O., Abdeen, A., and Prat, S. (2004). Conserved MYC transcription factors play a key role in jasmonate signaling both in tomato and Arabidopsis. Genes Dev. 18, 1577–1591. doi: 10.1101/gad.297704
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Browse, J. (2009). Jasmonate passes muster: a receptor and targets for the defense hormone. Annu. Rev. Plant Biol. 60, 183–205. doi: 10.1146/annurev.arplant.043008.092007
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chatel, G., Montiel, G., Pre, M., Memelink, J., Thiersault, M., Saint-Pierre, B., et al. (2003). CrMYC1, a Catharanthus roseus elicitor- and jasmonate-responsive bHLH transcription factor that binds the G-box element of the strictosidine synthase gene promoter. J. Exp. Bot. 54, 2587–2588. doi: 10.1093/jxb/erg275
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chico, J. M., Chini, A., Fonseca, S., and Solano, R. (2008). JAZ repressors set the rhythm in jasmonate signaling. Curr. Opin. Plant Biol. 11, 486–494. doi: 10.1016/j.pbi.2008.06.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chini, A., Boter, M., and Solano, R. (2009). Plant oxylipins: COI1/JAZs/MYC2 as the core jasmonic acid-signalling module. FEBS J. 276, 4682–4692. doi: 10.1111/j.1742-4658.2009.07194.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chini, A., Fonseca, S., Fernandez, G., Adie, B., Chico, J. M., Lorenzo, O., et al. (2007). The JAZ family of repressors is the missing link in jasmonate signalling. Nature 448, 666–671. doi: 10.1038/nature06006
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Croteau, R., Ketchum, R. E. B., Long, R. M., Kaspera, R., and Wildung, M. R. (2006). Taxol biosynthesis and molecular genetics. Phytochem. Rev. 5, 75–97. doi: 10.1007/s11101-005-3748-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Geyter, N., Gholami, A., Goormachtig, S., and Goossens, A. (2012). Transcriptional machineries in jasmonate-elicited plant secondary metabolism. Trends Plant Sci. 17, 349–359. doi: 10.1016/j.tplants.2012.03.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Dombrecht, B., Xue, G. P., Sprague, S. J., Kirkegaard, J. A., Ross, J. J., Reid, J. B., et al. (2007). MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell 19, 2225–2245. doi: 10.1105/tpc.106.048017
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Facchini, P. J., and Park, S. U. (2003). Developmental and inducible accumulation of gene transcripts involved in alkaloid biosynthesis in opium poppy. Phytochemistry 64, 177–186. doi: 10.1016/S0031-9422(03)00292-9
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Farmer, E. E., and Ryan, C. A. (1990). Interplant communication: airborne methyl jasmonate induces synthesis of proteinase inhibitors in plant leaves. Proc. Natl. Acad. Sci. U.S.A. 87, 7713–7716. doi: 10.1073/pnas.87.19.7713
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Flores-Bustamante, Z. R., Rivera-Orduna, F. N., Martinez-Cardenas, A., and Flores-Cotera, L. B. (2010). Microbial paclitaxel: advances and perspectives. J. Antibiot. 63, 460–467. doi: 10.1038/ja.2010.83
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fonseca, S., Chico, J. M., and Solano, R. (2009). The jasmonate pathway: the ligand, the receptor and the core signalling module. Curr. Opin. Plant Biol. 12, 539–547. doi: 10.1016/j.pbi.2009.07.013
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frense, D. (2007). Taxanes: perspectives for biotechnological production. Appl. Microbiol. Biotechnol. 73, 1233–1240. doi: 10.1007/s00253-006-0711-0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Fu, Y., Li, S., Zu, Y., Yang, G., Yang, Z., Luo, M., et al. (2009). Medicinal chemistry of paclitaxel and its analogues. Curr. Med. Chem. 16, 3966–3985. doi: 10.2174/092986709789352277
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gangappa, S. N., Prasad, V. B., and Chattopadhyay, S. (2010). Functional interconnection of MYC2 and SPA1 in the photomorphogenic seedling development of Arabidopsis. Plant Physiol. 154, 1210–1219. doi: 10.1104/pp.110.163717
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gigolashvili, T., Berger, B., Mock, H. P., Muller, C., Weisshaar, B., and Flugge, U. I. (2007a). The transcription factor HIG1/MYB51 regulates indolic glucosinolate biosynthesis in Arabidopsis thaliana. Plant J. 50, 886–901. doi: 10.1111/j.1365-313X.2007.03099.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gigolashvili, T., Engqvist, M., Yatusevich, R., Muller, C., and Flugge, U. I. (2008). HAG2/MYB76 and HAG3/MYB29 exert a specific and coordinated control on the regulation of aliphatic glucosinolate biosynthesis in Arabidopsis thaliana. New Phytol. 177, 627–642. doi: 10.1111/j.1469-8137.2007.02295.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gigolashvili, T., Yatusevich, R., Berger, B., Muller, C., and Flugge, U. I. (2007b). The R2R3-MYB transcription factor HAG1/MYB28 is a regulator of methionine-derived glucosinolate biosynthesis in Arabidopsis thaliana. Plant J. 51, 247–261. doi: 10.1111/j.1365-313X.2007.03133.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hajdukiewicz, P., Svab, Z., and Maliga, P. (1994). The small, versatile pPZP family of Agrobacterium binary vectors for plant transformation. Plant Mol. Biol. 25, 989–994. doi: 10.1007/BF00014672
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Higo, K., Ugawa, Y., Iwamoto, M., and Korenaga, T. (1999). Plant cis-acting regulatory DNA elements (PLACE) database: 1999. Nucleic Acids Res. 27, 297–300. doi: 10.1093/nar/27.1.297
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Hudgins, J. W., Christiansen, E., and Franceschi, V. R. (2004). Induction of anatomically based defense responses in stems of diverse conifers by methyl jasmonate: a phylogenetic perspective. Tree Physiol. 24, 251–264. doi: 10.1093/treephys/24.3.251
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jefferson, R. A., Kavanagh, T. A., and Bevan, M. W. (1987). GUS fusions: beta-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 6, 3901–3907.
Jennewein, S., Long, R. M., Williams, R. M., and Croteau, R. (2004a). Cytochrome p450 taxadiene 5alpha-hydroxylase, a mechanistically unusual monooxygenase catalyzing the first oxygenation step of taxol biosynthesis. Chem. Biol. 11, 379–387. doi: 10.1016/j.chembiol.2004.02.022
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Jennewein, S., Wildung, M. R., Chau, M., Walker, K., and Croteau, R. (2004b). Random sequencing of an induced Taxus cell cDNA library for identification of clones involved in Taxol biosynthesis. Proc. Natl. Acad. Sci. U.S.A. 101, 9149–9154. doi: 10.1073/pnas.0403009101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ketchum, R. E., Horiguchi, T., Qiu, D., Williams, R. M., and Croteau, R. B. (2007). Administering cultured Taxus cells with early precursors reveals bifurcations in the taxoid biosynthetic pathway. Phytochemistry 68, 335–341. doi: 10.1016/j.phytochem.2006.10.019
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kim, S. R., Choi, J. L., Costa, M. A., and An, G. (1992). Identification of G-box sequence as an essential element for methyl jasmonate response of potato proteinase inhibitor II promoter. Plant Physiol. 99, 627–631. doi: 10.1104/pp.99.2.627
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Kolewe, M. E., Gaurav, V., and Roberts, S. C. (2008). Pharmaceutically active natural product synthesis and supply via plant cell culture technology. Mol. Pharm. 5, 243–256. doi: 10.1021/mp7001494
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lenka, S. K., Boutaoui, N., Paulose, B., Vongpaseuth, K., Normanly, J., Roberts, S. C., et al. (2012). Identification and expression analysis of methyl jasmonate responsive ESTs in paclitaxel producing Taxus cuspidata suspension culture cells. BMC Genomics 13:148. doi: 10.1186/1471-2164-13-148
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Li, S., Zhang, P., Zhang, M., Fu, C., and Yu, L. (2012a). Functional analysis of a WRKY transcription factor involved in transcriptional activation of the DBAT gene in Taxus chinensis. Plant Biol (Stuttg). 15, 19–26. doi: 10.1111/j.1438-8677.2012.00611.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Li, S. T., Zhang, Z. P., Zhang, M., Fu, C. H., Zhao, C. F., Dong, Y. S., et al. (2012b). Transcriptional profile of Taxus chinensis cells in response to methyl jasmonate. BMC Genomics 13:295. doi: 10.1186/1471-2164-13-295
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Long, R. M., Lagisetti, C., Coates, R. M., and Croteau, R. B. (2008). Specificity of the N-benzoyl transferase responsible for the last step of Taxol biosynthesis. Arch. Biochem. Biophys. 477, 384–389. doi: 10.1016/j.abb.2008.06.021
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lorenzo, O., Chico, J. M., Sanchez-Serrano, J. J., and Solano, R. (2004). JASMONATE-INSENSITIVE1 encodes a MYC transcription factor essential to discriminate between different jasmonate-regulated defense responses in Arabidopsis. Plant Cell 16, 1938–1950. doi: 10.1105/tpc.022319
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Menke, F. L., Champion, A., Kijne, J. W., and Memelink, J. (1999). A novel jasmonate- and elicitor-responsive element in the periwinkle secondary metabolite biosynthetic gene Str interacts with a jasmonate- and elicitor-inducible AP2-domain transcription factor, ORCA2. EMBO J. 18, 4455–4463. doi: 10.1093/emboj/18.16.4455
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mewis, I., Tokuhisa, J. G., Schultz, J. C., Appel, H. M., Ulrichs, C., and Gershenzon, J. (2006). Gene expression and glucosinolate accumulation in Arabidopsis thaliana in response to generalist and specialist herbivores of different feeding guilds and the role of defense signaling pathways. Phytochemistry 67, 2450–2462. doi: 10.1016/j.phytochem.2006.09.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mikkelsen, M. D., Petersen, B. L., Glawischnig, E., Jensen, A. B., Andreasson, E., and Halkier, B. A. (2003). Modulation of CYP79 genes and glucosinolate profiles in Arabidopsis by defense signaling pathways. Plant Physiol. 131, 298–308. doi: 10.1104/pp.011015
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Montiel, G., Zarei, A., Korbes, A. P., and Memelink, J. (2011). The Jasmonate-responsive element from the ORCA3 promoter from Catharanthus roseus is active in Arabidopsis and is controlled by the transcription factor AtMYC2. Plant Cell Physiol. 52, 578–587. doi: 10.1093/pcp/pcr016
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Nims, E., Dubois, C. P., Roberts, S. C., and Walker, E. L. (2006). Expression profiling of genes involved in paclitaxel biosynthesis for targeted metabolic engineering. Metab. Eng. 8, 385–394. doi: 10.1016/j.ymben.2006.04.001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ochman, H., Gerber, A. S., and Hartl, D. L. (1988). Genetic applications of an inverse polymerase chain reaction. Genetics 120, 621–623.
Patil, R. A., Kolewe, M. E., and Roberts, S. C. (2013). Cellular aggregation is a key parameter associated with long term variability in paclitaxel accumulation in Taxus suspension cultures. Plant Cell Tissue Organ Cult. 112, 303–310. doi: 10.1007/s11240-012-0237-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Pre, M., Atallah, M., Champion, A., De Vos, M., Pieterse, C. M., and Memelink, J. (2008). The AP2/ERF domain transcription factor ORA59 integrates jasmonic acid and ethylene signals in plant defense. Plant Physiol. 147, 1347–1357. doi: 10.1104/pp.108.117523
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Qi, T., Song, S., Ren, Q., Wu, D., Huang, H., Chen, Y., et al. (2011). The Jasmonate-ZIM-domain proteins interact with the WD-Repeat/bHLH/MYB complexes to regulate Jasmonate-mediated anthocyanin accumulation and trichome initiation in Arabidopsis thaliana. Plant Cell 23, 1795–1814. doi: 10.1105/tpc.111.083261
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Qian, W., Tan, G., Liu, H., He, S., Gao, Y., and An, C. (2007). Identification of a bHLH-type G-box binding factor and its regulation activity with G-box and Box I elements of the PsCHS1 promoter. Plant Cell Rep. 26, 85–93. doi: 10.1007/s00299-006-0202-x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rose, T. M., Henikoff, J. G., and Henikoff, S. (2003). CODEHOP (COnsensus-DEgenerate Hybrid Oligonucleotide Primer) PCR primer design. Nucleic Acids Res. 31, 3763–3766. doi: 10.1093/nar/gkg524
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schiff, P. B., Fant, J., and Horwitz, S. B. (1979). Promotion of microtubule assembly in vitro by taxol. Nature 277, 665–667. doi: 10.1038/277665a0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Shoji, T., Yamada, Y., and Hashimoto, T. (2000). Jasmonate induction of putrescine N-methyltransferase genes in the root of Nicotiana sylvestris. Plant Cell Physiol. 41, 831–839. doi: 10.1093/pcp/pcd001
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Siberil, Y., Benhamron, S., Memelink, J., Giglioli-Guivarc'h, N., Thiersault, M., Boisson, B., et al. (2001). Catharanthus roseus G-box binding factors 1 and 2 act as repressors of strictosidine synthase gene expression in cell cultures. Plant Mol. Biol. 45, 477–488. doi: 10.1023/A:1010650906695
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Todd, A. T., Liu, E., Polvi, S. L., Pammett, R. T., and Page, J. E. (2010). A functional genomics screen identifies diverse transcription factors that regulate alkaloid biosynthesis in Nicotiana benthamiana. Plant J. 62, 589–600. doi: 10.1111/j.1365-313X.2010.04186.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ton, J., D'alessandro, M., Jourdie, V., Jakab, G., Karlen, D., Held, M., et al. (2007). Priming by airborne signals boosts direct and indirect resistance in maize. Plant J. 49, 16–26. doi: 10.1111/j.1365-313X.2006.02935.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
van der Fits, L., and Memelink, J. (2000). ORCA3, a jasmonate-responsive transcriptional regulator of plant primary and secondary metabolism. Science 289, 295–297. doi: 10.1126/science.289.5477.295
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
van der Fits, L., and Memelink, J. (2001). The jasmonate-inducible AP2/ERF-domain transcription factor ORCA3 activates gene expression via interaction with a jasmonate-responsive promoter element. Plant J. 25, 43–53. doi: 10.1046/j.1365-313x.2001.00932.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
van der Fits, L., Zhang, H., Menke, F. L., Deneka, M., and Memelink, J. (2000). A Catharanthus roseus BPF-1 homologue interacts with an elicitor-responsive region of the secondary metabolite biosynthetic gene Str and is induced by elicitor via a JA-independent signal transduction pathway. Plant Mol. Biol. 44, 675–685. doi: 10.1023/A:1026526522555
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vongpaseuth, K., Nims, E., St Amand, M., Walker, E. L., and Roberts, S. C. (2007). Development of a particle bombardment-mediated transient transformation system for Taxus spp. cells in culture. Biotechnol. Prog. 23, 1180–1185. doi: 10.1021/bp0700307
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Vongpaseuth, K., and Roberts, S. C. (2007). Advancements in the understanding of Paclitaxel metabolism in tissue culture. Curr. Pharm. Biotechnol. 8, 219–236. doi: 10.2174/138920107781387393
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Walker, K., and Croteau, R. (2000a). Molecular cloning of a 10-deacetylbaccatin III-10-O-acetyl transferase cDNA from Taxus and functional expression in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 97, 583–587. doi: 10.1073/pnas.97.2.583
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Walker, K., and Croteau, R. (2000b). Taxol biosynthesis: molecular cloning of a benzoyl-CoA:taxane 2a-O-benzoyltransferase cDNA from Taxus and functional expression in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 97, 13591–13596. doi: 10.1073/pnas.250491997
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Walker, K., Fujisaki, S., Long, R., and Croteau, R. (2002a). Molecular cloning and heterologous expression of the C-13 phenylpropanoid side chain CoA acyltransferase that functions in Taxol synthesis. Proc. Natl. Acad. Sci. U.S.A. 99, 12715–12720. doi: 10.1073/pnas.192463699
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Walker, K., Long, R., and Croteau, R. (2002b). The final acylation step in Taxol biosynthesis: cloning of the taxoid C13-side chain N-benzoyltransferase from Taxus. Proc. Natl. Acad. Sci. U.S.A. 99, 9166–9171. doi: 10.1073/pnas.082115799
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wildung, M. R., and Croteau, R. (1996). A cDNA clone for taxadiene synthase, the diterpene cyclase that catalyzes the committed step of Taxol biosynthesis. J. Biol. Chem. 271, 9201–9204. doi: 10.1074/jbc.271.16.9201
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Wu, F. H., Shen, S. C., Lee, L. Y., Lee, S. H., Chan, M. T., and Lin, C. S. (2009). Tape-Arabidopsis sandwich – a simpler Arabidopsis protoplast isolation method. Plant Methods 5; 16. doi: 10.1186/1746-4811-5-16
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yin, S., Mei, L., Newman, J., Back, K., and Chappell, J. (1997). Regulation of sesquiterpene cyclase gene expression. Characterization of an elicitor- and pathogen-inducible promoter. Plant Physiol 115, 437–451. doi: 10.1104/pp.115.2.437
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Yukimune, Y., Tabata, H., Higashi, Y., and Hara, Y. (1996). Methyl jasmonate-induced overproduction of paclitaxel and baccatin III in Taxus cell suspension cultures. Nat. Biotechnol. 14, 1129–1132. doi: 10.1038/nbt0996-1129
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Zhang, H. B., Bokowiec, M. T., Rushton, P. J., Han, S. C., and Timko, M. P. (2012). Tobacco transcription factors NtMYC2a and NtMYC2b form nuclear complexes with the NtJAZ1 repressor and regulate multiple jasmonate-inducible steps in nicotine biosynthesis. Mol. Plant 5, 73–84. doi: 10.1093/mp/ssr056
Zhang, H., Hedhili, S., Montiel, G., Zhang, Y., Chatel, G., Pre, M., et al. (2011). The basic helix-loop-helix transcription factor CrMYC2 controls the jasmonate-responsive expression of the ORCA genes that regulate alkaloid biosynthesis in Catharanthus roseus. Plant J. 67, 61–71. doi: 10.1111/j.1365-313X.2011.04575.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: Taxus cuspidata, paclitaxel, methyl jasmonate, E-box, JA-MYC, MYC2
Citation: Lenka SK, Nims NE, Vongpaseuth K, Boshar RA, Roberts SC and Walker EL (2015) Jasmonate-responsive expression of paclitaxel biosynthesis genes in Taxus cuspidata cultured cells is negatively regulated by the bHLH transcription factors TcJAMYC1, TcJAMYC2, and TcJAMYC4. Front. Plant Sci. 6:115. doi: 10.3389/fpls.2015.00115
Received: 08 January 2015; Accepted: 11 February 2015;
Published online: 26 February 2015.
Edited by:
Daniel Anthony Dias, The University of Melbourne, AustraliaReviewed by:
Mingshu Cao, AgResearch, Grasslands Research Center, New ZealandQing Liu, Commonwealth Scientific and Industrial Research Organisation, Australia
Copyright © 2015 Lenka, Nims, Vongpaseuth, Boshar, Roberts and Walker. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elsbeth L. Walker, Department of Biology, University of Massachusetts, 611 North Pleasant St., Amherst, MA 01003, USA e-mail:ZXdhbGtlckBiaW8udW1hc3MuZWR1
†Present address: N. Ezekiel Nims, Ra Pharmaceuticals, Inc., Cambridge, MA, USA;
Kham Vongpaseuth, U.S. Department of Agriculture, Animal and Plant Health Inspection Service, Biotechnology Regulatory Services, Riverdale, MD, USA
‡These authors have contributed equally to this work.