 Petra Žádníková
Petra Žádníková Dajo Smet
Dajo Smet Qiang Zhu
Qiang Zhu Dominique Van Der Straeten
Dominique Van Der Straeten Eva Benková
Eva Benková- 1Department of Plant Systems Biology, Flanders Institute for Biotechnology, Ghent, Belgium
- 2Department of Plant Biotechnology and Bioinformatics, Ghent University, Ghent, Belgium
- 3Department of Physiology, Laboratory of Functional Plant Biology, Ghent University, Ghent, Belgium
- 4Institute of Science and Technology Austria, Klosterneuburg, Austria
Plants are sessile organisms that are permanently restricted to their site of germination. To compensate for their lack of mobility, plants evolved unique mechanisms enabling them to rapidly react to ever changing environmental conditions and flexibly adapt their postembryonic developmental program. A prominent demonstration of this developmental plasticity is their ability to bend organs in order to reach the position most optimal for growth and utilization of light, nutrients, and other resources. Shortly after germination, dicotyledonous seedlings form a bended structure, the so-called apical hook, to protect the delicate shoot meristem and cotyledons from damage when penetrating through the soil. Upon perception of a light stimulus, the apical hook rapidly opens and the photomorphogenic developmental program is activated. After germination, plant organs are able to align their growth with the light source and adopt the most favorable orientation through bending, in a process named phototropism. On the other hand, when roots and shoots are diverted from their upright orientation, they immediately detect a change in the gravity vector and bend to maintain a vertical growth direction. Noteworthy, despite the diversity of external stimuli perceived by different plant organs, all plant tropic movements share a common mechanistic basis: differential cell growth. In our review, we will discuss the molecular principles underlying various tropic responses with the focus on mechanisms mediating the perception of external signals, transduction cascades and downstream responses that regulate differential cell growth and consequently, organ bending. In particular, we highlight common and specific features of regulatory pathways in control of the bending of organs and a role for the plant hormone auxin as a key regulatory component.
Introduction
To compensate for their sessile lifestyle, plants developed unique mechanisms that provide them with an unusual level of developmental plasticity. The adaptive behavior of plants is noticeable throughout their life cycle. Already during early skotomorphogenesis (development in darkness), seedlings of dicotyledonous plants form a hook-like structure at the apical part of the hypocotyl to shield their tender shoot apical meristem and cotyledons from damage while penetrating the soil (Raz and Ecker, 1999). This apical hook is maintained in a closed stage while seedlings grow through the soil, and rapidly opens when exposed to light. Also during photomorphogenesis (development in light), shoot organs maintain the ability to coordinate their growth. Plants manage to direct growth in response to the perceived light; a process studied and described by Darwin and Darwin (1881) as phototropism. Equally important to light-directed growth is the ability of plants to sense and adjust growth in response to gravity. Gravitropism is of the utmost importance for plant growth and survival, in particular after underground germination, when gravity is the sole cue to direct both root and shoot growth. In higher plants, shoots are negatively gravitropic (growing opposite to the direction of the gravity vector), whereas roots are positively gravitropic (growing in the direction of the gravity vector; Knight, 1806). Changes in the direction of the gravity vector are continuously sensed and both the root and shoot are able to rapidly bend and realign their growth to maintain a vertical growth direction. Interestingly, despite the diversity of cues and nature of responding organs, all different types of organ bending, whether triggered by gravity or light, share the same mechanistic principle based on differential cell growth.
The first hints on the molecular basis of the differential growth underlying organ bending date to Darwin’s early studies on coleoptiles. Based on the bending of coleoptiles toward unilateral light, Darwin and Darwin (1881) predicted the existence of a messenger molecule which is transported from the site of light perception at the tip of coleoptile toward the site of response where bending occurs. Later, it was demonstrated that an asymmetrical accumulation of the plant hormone auxin at the non-versus light-stimulated side correlates with differential cell growth and organ bending (Boysen-Jensen, 1911) and a model implementing a role for auxin and its asymmetric distribution in the regulation of plant tropic responses was proposed (Cholodny, 1927, 1928; Went, 1928). Although the molecular basis underlying tropisms was unknown at that time, the Cholodny–Went model was an important conceptual step in the general understanding of the principles of plant movements. The model delineated the main processes mediating plant bending in response to environmental stimuli which encompass local signal sensing, and inter- and intracellular signal transition toward the site of response. Hence, the identification of the molecular components and mechanisms involved in the coordination of differential growth, underlying organ bending in response to environmental stimuli, became one of the main milestones of following research.
In our review we will discuss the current knowledge on the pathways engaged in the control of tropic responses and apical hook development, the developmental process which is similar to tropic responses largely based on coordinated differential cell growth. Similarities and specificities of mechanisms mediating organ bending in response to environmental stimuli will be highlighted.
Phototropism
Conversion of light into chemical energy during photosynthesis is essential for sustaining life on earth. Plants belong to the few species capable of such an energy transformation and by storing energy in carbohydrate molecules such as sugars, nutritional resources are generated for other organisms. To adjust their light absorption capacities, plants have evolved different light perception and signaling systems. Among them, phototropism or bending of an organ toward unidirectional blue light, is one of the most important adaptive responses. For the summary of the common and different factors involved in the regulation of phototropism see the Table 1.
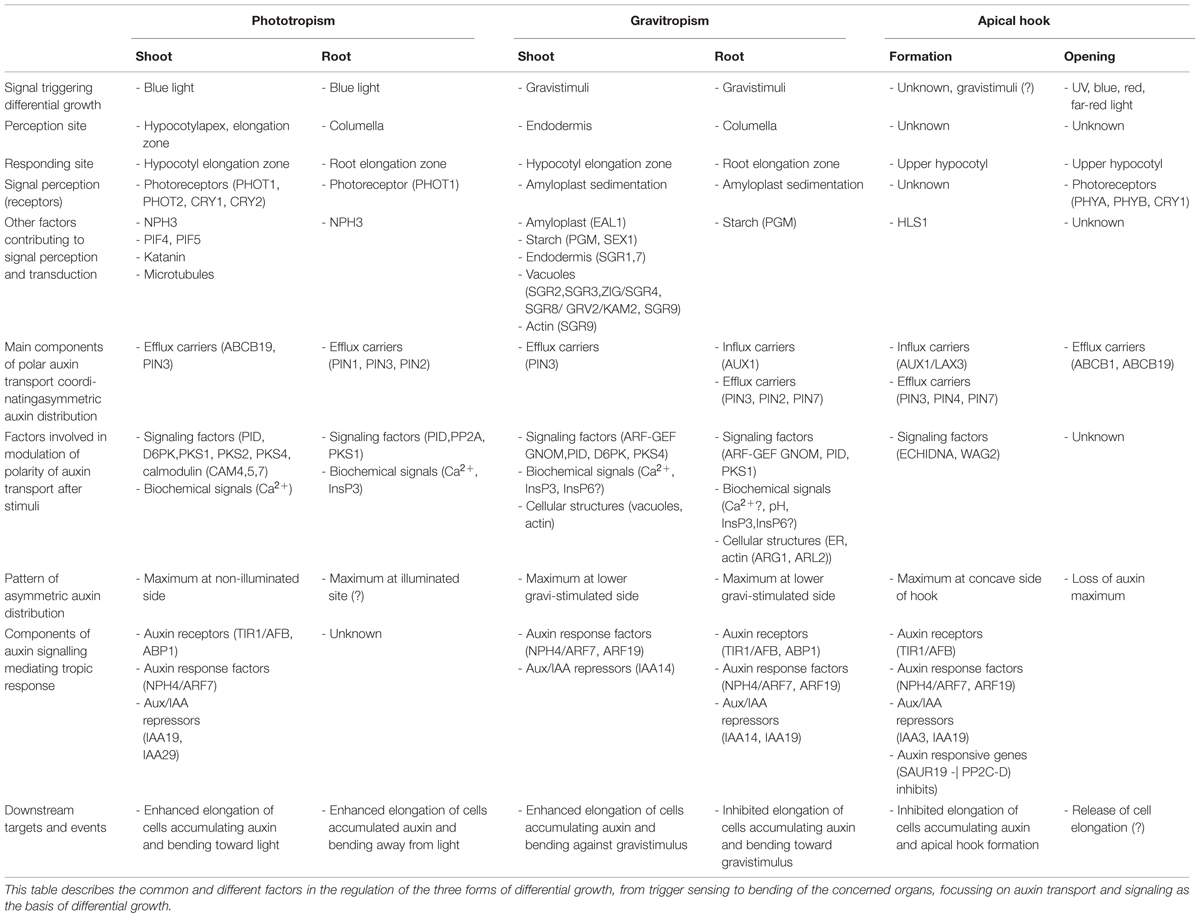
TABLE 1. Mechanism of differential growth in phototropism, gravitropism, and apical hook development.
Shoot Phototropism
Light Sensing
Although already in Darwin and Darwin (1881) the tip of coleoptiles was predicted to be the site of light perception, the identity of the light receptor(s) mediating the phototropic bending remained unknown for more than 100 years.
The isolation of Arabidopsis mutants defective in the response to unilateral light such as non-phototropic1 (nph1), nph2, nph3 and nph4 (Khurana and Poff, 1989; Liscum and Briggs, 1996) enabled the discovery of the receptors in the phototropic pathway. The NPH1 gene was found to encode a flavoprotein kinase and confirmed to act as a UV-A/blue receptor (Huala et al., 1997; Christie et al., 1998). Later, its homolog NPH1-LIKE1 (NPL1) was shown to contribute to the perception of blue light mainly at high fluence rates (Sakai et al., 2001). NPH1 and NPL1, later renamed to PHOTOTROPIN1 (PHOT1) and PHOTOTROPIN2 (PHOT2), are considered to be the major blue light receptors controlling phototropic responses (Briggs et al., 2001). Residual phototropic responses of phot1phot2 mutants to blue illumination suggested the involvement of other photoreceptors. The latter proved to be CRYPTOCHROME1 (CRY1) and CRYPTOCHROME2 (CRY2), because the mutation of CRY1 and CRY2 in the phot1phot2 mutant completely abolished the phototropic response (Ohgishi et al., 2004). The PHOT1 and PHOT2 function might also be partially replaced by phytochromes, the red/far-red sensitive light receptors, due to their ability to partially perceive blue light (Kami et al., 2010). Furthermore, activated phytochromes were shown to enhance the retention of PHOT1 and PHOT2 at the plasma membrane (Han et al., 2008), as well as to modulate the degree of phosphorylation of CRY1 and CRY2 (Ahmad et al., 1998), which indicates their indirect contribution to the regulation of phototropism.
Expression and protein localization of PHOT1 in coleoptile tips perfectly complied with the predicted perception site for phototropism (Knieb et al., 2004; Matsuda et al., 2011). In contrast, in dicotyledonous plants such as Arabidopsis, the expression pattern of PHOT1 and PHOT2 is much more extended and therefore the precise location of the perception site is less definite. Recent works indicated that the light perception site in Arabidopsis is indeed less strictly defined when compared to coleoptiles and might depend on the developmental status of the plant. As shown by Christie et al. (2011), light perception above and at the hypocotyl apex controls phototropic bending of de-etiolated Arabidopsis seedlings. In contrast, upon removal of the cotyledons and hypocotyl apex of etiolated Arabidopsis seedlings, the phototropic response remains unaffected. PHOT1 expression in the hypocotyl elongation zone is, however, essential for proper bending (Preuten et al., 2013). Furthermore, tissue-specific expression of PHOT1 in either the epidermis, cortex or endodermis suffices to trigger a full phototropic response, indicating that local activation of PHOT1 might lead to seedling-wide signaling, presumably mediated through a non-cell autonomous type of signal (Preuten et al., 2013).
Light Signal Transduction to Coordinate Shoot Differential Growth
Although phototropins PHOT1 and PHOT2, Ser/Thr kinases belonging to the AGC family (Bögre et al., 2003), have been recognized as principal light receptors mediating the phototropic response, the immediate downstream events triggered by light remain largely unknown. Mutant screens and the identification of photoreceptor-interacting factors provided important insights in the signal transduction cascade upon light sensing (Motchoulski and Liscum, 1999; Sullivan et al., 2009).
NON-PHOTOTROPIC HYPOCOTYL 3 (NPH3) was identified in early screens for mutants defective in phototropism, and despite its essential regulatory function, the mechanism of NPH3 action remained obscure for a long time (Liscum and Briggs, 1996). Recent results indicated that NPH3 acts as a substrate adapter in the CULLIN-based E3 ubiquitin ligase CRL3NPH3 complex (Roberts et al., 2011) and mediates mono- and multiubiquitination of PHOT1 under low-intensity blue light. This posttranslational modification of PHOT1 might promote clathrin-dependent internalization of the photoreceptor. In contrast, high-intensity blue light was found to trigger polyubiquitination which marks PHOT1 for proteasome-mediated degradation. The predicted function of NPH3 fits well with the previously demonstrated direct interaction between NPH3 and PHOT1 (Motchoulski and Liscum, 1999) and the observation that blue light irradiation might induce rapid internalization of PHOT1 (Kaiserli et al., 2009). In light of these results, NPH3 might primarily act in fine-tuning the photoreceptor activity through modulation of PHOT1 subcellular trafficking and desensitization by degradation.
Since Darwin’s experiments, evidence accumulated in support of a crucial role of an asymmetric auxin distribution, depending on functional polar auxin transport, for a proper phototropic response (Briggs, 1963; Epel et al., 1992; Friml et al., 2002). Yet, what do we know about the events coordinating the asymmetric auxin distribution and phototropic response downstream of PHOT1-mediated light perception? Among molecular components controlling polar auxin transport in plants (for review, see Petrášek and Friml, 2009), two auxin efflux carriers, ATP-BINDING CASSETTE B19 (ABCB19; Christie et al., 2011) and PIN-like 3 (PIN3; Friml et al., 2002), emerged to be involved in the control of the phototropic response. Loss of ABCB19 function results in an enhanced phototropic response and its expression pattern in the upper part of the hypocotyl of de-etiolated seedlings largely overlaps with that of PHOT1. Efforts to characterize a functional link between the photoreceptor and ABCB19 revealed that PHOT1 transiently interacts with ABCB19. Through this direct physical interaction PHOT1 promotes ABCB19 phosphorylation, which consequently leads to the attenuation of its auxin transport activity. Based on these observations it has been proposed that PHOT1 restrains ABCB19 activity, thereby increasing the accumulation of auxin in and above the hypocotyl apex to prime lateral auxin fluxes that are channeled to the elongation zone by the auxin transporter PIN3, hereby coordinating differential cell growth (Christie et al., 2011). The proposed model nicely complies with the previously presented phosphorylation gradient model which was based on the finding that unilateral irradiation induces a gradient of PHOT1 autophosphorylation across oat coleoptiles, with the highest level of phosphorylation occurring on the irradiated side (Salomon et al., 1997).
In contrast, lack of PIN3 activity causes reduced sensitivity to unilateral light, and localization of this transporter at lateral membranes of hypocotyl endodermal cells hinted at PIN3 involvement in the lateral redistribution of auxin in the lower elongation zone of the hypocotyl (Friml et al., 2002). However, whether and how PIN3 might coordinate auxin redistribution in endodermal cells to form local maxima in the cortex and epidermal cells at the non-illuminated side of the hypocotyl remained an open question. Recent work (Ding et al., 2011) provided a plausible mechanistic model postulating a light-induced local modulation of PIN3-dependent transport. The authors demonstrated that unilateral light polarizes PIN3 in the endodermal cells, which severely attenuates the PIN3 signal at the outer- and maintains a strong signal at the inner lateral membrane of illuminated endodermal cells. In contrast, no polarization of PIN3 was found in non-illuminated cells. The PIN3 polarization occurs downstream of PHOT1-dependent light perception, however, unlike ABCB19, PIN3 was not confirmed as a substrate of the PHOT1 kinase, suggesting an indirect PHOT1-mediated regulation of PIN3 polarization. Interestingly, transcription of PINOID (PID) kinase, previously implicated in the control of PIN polarization through phosphorylation (Friml et al., 2002; Ding et al., 2011), was found to decrease in response to light. Based on these observations, a model was proposed in which a light-dependent control of PID activity accounts for differential phosphorylation and polar targeting of PIN3, which subsequently results in an asymmetric auxin distribution during the phototropic response. Besides PID, D6 PROTEIN KINASE (D6PK), another member of the AGC kinase family, was shown to play a role in the phototropic response. However, unlike PID, D6PK-mediated phosphorylation of PIN3 seems to have a main impact on its auxin transport activity. Hence, D6PK might, through control of overall PIN3 transport activity rather than PIN3 polarity, contribute to the regulation of the phototropic response (Willige et al., 2013; Zourelidou et al., 2014). Haga and Sakai (2012) dissected the phototropic responses in three categories, pulse-induced phototropism, time-dependent second positive phototropism, and continuous-light-induced second positive phototropism. Interestingly, PID – PIN3 mediated auxin gradients were found to control primarily the pulse-induced but not the continuous-light-induced phototropism thus suggesting the existence of two functionally distinct mechanisms; a PIN-dependent mechanism in which transient stimulation is sufficient to induce phototropism, and a PIN-independent mechanism that requires continuous light stimulation (Haga et al., 2014).
Other important components of the signaling cascade controlling the phototropic response are PHYTOCHROME KINASE SUBSTRATE1 (PKS1), PKS2, and PKS4. PKSs were identified in a screen for early blue light response genes (Lariguet et al., 2006) and genetic analyses confirmed their role in PHOT1- and PHOT2-dependent phototropism under weak intensities of blue light (Lariguet et al., 2006; Kami et al., 2014). PKSs localize to the plasma membrane and interact with PHOT1 and NPH3 (Lariguet et al., 2006). However, no indications for their regulation of the PHOT1 membrane localization and trafficking, resembling NPH3 function, could be found. Current analyses of auxin levels, rootward auxin transport and auxin signaling in pks1, pks2, and pks4 suggested that PKSs might contribute to the regulation of the phototropic response through modulation of local auxin signaling or lateral auxin transport, alternatively (Kami et al., 2014).
An important part of the response to blue light includes the rapid modulation of ion homeostasis such as calcium (Ca2+; Baum et al., 1999; Harada and Shimazaki, 2007). An earlier study revealed that a transient rise in cytosolic Ca2+ is important for PHOT1-mediated inhibition of hypocotyl growth, and the increase in Ca2+ attributed to PHOT2 function is required for blue light induced phototropism (Folta et al., 2003; Zhao et al., 2013). Detailed characterization of the PHOT role in Ca2+ homeostasis revealed that whereas PHOT1 seems to primarily control Ca2+ transport through plasma membranes, PHOT2 is mainly involved in the activation of intracellular Ca2+ resources and transport of Ca2+ from vacuoles and the endoplasmic reticulum (ER; Zhao et al., 2013).
Apart from blue light, also auxin might affect cytosolic Ca2+ levels. Inhibition of auxin transport upon 1-N-naphthylphthalamic acid (NPA) treatment inhibited a cytosolic Ca2+ increase in response to blue light, while auxin treatment led to a dramatic increase in cytosolic Ca2+ (Zhao et al., 2013), which support the role of the functional polar auxin transport and auxin in the blue light induced increase of Ca2+. However, functional links between light-controlled Ca2+ levels and auxin in the regulation of the phototropic response are still largely hypothetical. Considering recent findings on the involvement of cytosolic Ca2+ in PIN polarity establishment (Zhang et al., 2011), light- and auxin-stimulated modulation of Ca2+ levels might bring about a feedback regulation of PIN3 polarity establishment during the phototropic response. On the other hand, observations that PKSs interact with several members of the calmodulin family such as CAM4, CAM5, and CAM7, hint at a role of Ca2+ as a trigger of unknown downstream targets in the signaling cascade underlying the phototropic response (Zhao et al., 2013).
Auxin Coordinates Phototropic Organ Bending
Translation of the light-induced auxin gradient to an appropriate molecular response requires a functional hormone signaling cascade. Defects in auxin perception caused by the lack of the auxin receptors TIR1/AFB as well as ABP1 activity compromise the phototropic response (Möller et al., 2010; Effendi et al., 2011). This indicates that both TIR1- and ABP1-dependent auxin transduction cascades contribute to the regulation of phototropism. Although not much is known about events downstream of ABP1, it is presumably involved in control of the subcellular distribution of PIN auxin transporters and proper auxin distribution during phototropic response (Robert et al., 2010; Xu et al., 2010). In addition, ABP1, through negative feedback on auxin responses as demonstrated by Tromas et al. (2013), might fine-tune auxin signaling-dependent responses. Several components of the TIR1-controlled auxin signaling network, comprising the Auxin Response Factor (ARF) transcriptional regulators and their Aux/IAA repressors, including NPH4/ARF7, MSG2/IAA19, and IAA29, were recognized for their regulatory role in phototropism (Liscum and Briggs, 1996; Tatematsu et al., 2004; Okushima et al., 2005; Sun et al., 2013).
Aux/IAAs appear to be an important convergence point with the phytochrome-dependent light-regulated pathway. Transcription of IAA19 and IAA29 was found to be under control of PHYTOCHROME INTERACTING FACTOR4 (PIF4) and PIF5 (Sun et al., 2013). Originally, PIFs were identified as phytochrome interactors, which are phosphorylated and rapidly degraded to activate photomorphogenesis after light exposure (Castillon et al., 2007; Jeong and Choi, 2013). Recently published results indicated that these phytochrome-interacting partners might at the same time act in the regulatory pathway that mediates the attenuation of auxin signaling at the illuminated side of hypocotyls (Sun et al., 2013).
Activation of the auxin signaling pathway induces the expression of molecular targets, which, downstream of auxin, take part in the coordination of differential cell growth and thereby hypocotyl bending. Surprisingly, very few auxin-regulated target genes involved in phototropism are known so far.
Essential elements for the adaptive response of plant growth to the environment are cytoskeletal rearrangements. The microtubule cytoskeleton is an important component of the complex framework controlling cell growth. Microtubule organization directs the deposition of cellulose to the cell wall, thus contributing to the coordination of cell expansion and its directionality (Zandomeni and Schopfer, 1994). In elongating hypocotyl cells, cortical microtubules are arranged in parallel arrays mostly perpendicular to the hypocotyl axis. In response to blue light, microtubules undergo a rapid rearrangement into an orientation parallel to the hypocotyl axis (Laskowski, 1990; Nick et al., 1990). Recent work of Lindeboom et al. (2013) showed that this light-stimulated reorientation of cortical arrays involves the phototropin-dependent activation of the katanin-mediated severing of microtubules. The defective phototropic response in mutants lacking katanin demonstrate that the local light-induced microtubule rearrangement is an important mechanism for a proper phototropic response, however, molecular and mechanistic links between the rapid cytoskeletal response to light and other, in particular auxin-related events need to be explored.
Although the role of actin in the growth of cells is less evident, there is evidence supporting its involvement therein (Baskin and Bivens, 1995; Baluska et al., 2003). Experiments in etiolated rice coleoptiles clearly demonstrated that actin microfilaments are responsive to irradiation as well as auxin treatment. However, the developmental relevance of this actin dynamics modulation is unknown so far (Holweg et al., 2004).
As differential cell growth constitutes the mechanistic basis for organ bending during phototropism, genes controlling plant cell growth might be among expected auxin targets. Growth of a plant cell is driven by internal turgor pressure, and is dependent on the ability of the cell wall to extend under pressure. Turgor pressure results from the hydrostatic pressure exerted on the cell wall by the cytoplasm and its increase above a threshold leads to cell expansion (Boyer et al., 1985; Cosgrove, 1993; Peters and Tomos, 1996; Schopfer, 2006). On the other hand, the cell wall is a complex composite material consisting of stiff cellulose microfibrils, which are tethered by xyloglucans and embedded in a matrix of hydrosoluble pectins and structural proteins (Cosgrove, 2005; Burton et al., 2010). Hence, genes involved in the control of cell growth either through modulation of hydrostatic pressure, or by modifications of cell wall properties, such as relaxation through wall loosening, could be candidates. Indeed, several target genes of NPH4/ARF7 action, whose expression levels increase on the elongating side of phototropically stimulated Brassica oleracea hypocotyls, were revealed by a transcriptomic approach (Esmon et al., 2006). EXPA1 and EXPA8, two members of the α–expansin family, which mediate cell wall extension (Esmon et al., 2006), might be relevant for the establishment of phototropic curvature through control of cell wall expansion.
Negative Root Phototropism
While above-ground organs tend to bend toward blue light, underground organs such as roots typically turn away from the blue illumination. Interestingly, the same light receptor PHOT1 and downstream components such as NPH3, PKS, Ca2+, which control positive shoot phototropism as well as InsP3, are involved in negative root phototropism (Boccalandro et al., 2008; Salinas-Mondragon et al., 2010; Wan et al., 2012; Zhang et al., 2013). Similar to the shoot, unilateral blue light induces an asymmetric auxin distribution. However, while an auxin maximum forms at the shaded side of the shoot, auxin seems to accumulate in the meristem of epidermal cells at the illuminated side of the root (Zhang et al., 2013), although this conclusion in not fully supported by the work of Wan et al. (2012), indicating auxin accumulation at shaded side of root tip. Even though the pattern of auxin accumulation needs to be unambiguously resolved, theobserved asymmetry in auxin distribution hints at the involvement of a tightly controlled auxin transport in photo-stimulated roots. Indeed, auxin transport mutants such as pin2 and pin3, display defects in the root phototropic response (Wan et al., 2012; Zhang et al., 2013, 2014). Moreover, studies on the regulatory pathway acting downstream of light to modulate auxin transport activity, revealed striking similarities with the mechanism controlling shoot phototropism. As in the shoot, unilateral blue-light triggers the polarization of PIN3 in a PHOT1-, PINOID- , and PHOSPHATASE2A- (PP2A) dependent manner. In the root columella cells, PIN3 localizes to the outer lateral membranes at the illuminated side of the root and this blue light-induced PIN3 polarization might promote auxin redistribution toward the illuminated side of roots (Zhang et al., 2013). These results indicate that similar to the shoot, blue light-induced PIN3 polarization contributes to the control of an asymmetric auxin distribution during the negative root phototropic response. Besides PIN3, the activity of other root auxin transport components is also light-regulated. As recently shown, PIN1 and PIN2, major facilitators of rootward and shootward auxin streams, respectively, change their subcellular localization at the root apical meristem in response to illumination (Wan et al., 2012; Zhang et al., 2014). Hence, through common perception pathways light can effectively coordinate the activities of major auxin transporters and therefore the auxin distribution to control phototropic responses in different organs.
Gravitropism
As for their response to light, the correct recognition of the gravity vector by the root and shoot is of vital importance for plant growth and survival. In particular, during underground germination gravity is the key cue directing plant growth. Whereas root growth aligns with the gravity vector, the shoot is negatively gravitropic and grows against the gravity vector (Knight, 1806). For the summary of the common and different factors involved in the regulation of gravitropism see the Table 1.
Gravity Sensing
Specialized cells for the perception of gravity are called statocytes. In plants, these cells hold starch-containing plastids, referred to as amyloplasts, which sediment in the direction of gravity. Already back in Haberlandt (1900) and Němec (1900), amyloplasts were proposed to function as statoliths in gravity perception. This starch-statolith hypothesis is now widely accepted as a model for gravity perception in both roots and shoots of higher plants. However, different cell types function as statocytes in roots and shoots.
The root cap is essential for gravity perception in roots. Root cap removal through surgical ablation or genetic manipulation prevented gravitropic curvature of the root (Younis, 1954; Juniper et al., 1966; Tsugeki and Fedoroff, 1999) and high-precision laser ablation indicated the importance of the inner two cell layers of the root cap columella (Blancaflor et al., 1998). The perception of gravity outside the root cap was suggested by a continued gravitropic curvature of the root, while retaining a specific segment of the root elongation zone at a gravistimulated angle, even after a vertical orientation of the root cap (Mullen et al., 2000; Wolverton et al., 2002).However, gravity perception outside the root cap probably contributes only in a minor extent to the root gravitropic response.
In the shoot, starch-containing amyloplasts are found in the endodermis (Fukaki et al., 1998). The function of endodermal cells as statocytes is supported by several genetic studies. The Arabidopsis endodermal-amyloplastless (eal1) mutant, which lacks endodermal amyloplasts, exhibits hampered shoot gravitropism (Fujihira et al., 2000). Similarly, mutants in SCARECROW (SCR)/SHOOT GRAVITROPISM1 (SGR1) and SHORT-ROOT (SHR)/SGR7, transcription factors of the GRAS gene family, which are indispensable for normal shoot and root endodermis formation, are characterized by a deficient shoot gravitropic response. The deficient shoot gravitropism in the latter mutants supports the function of shoot endodermal cells as statocytes in gravity perception. On the other hand, the unaffected root gravitropism in endodermis defective mutants hints at a difference in the site of gravity perception between shoots and roots, supporting the role of columella cells as primary perception site in the root (Benfey et al., 1993; Fukaki et al., 1996, 1998; Yamauchi et al., 1997).
Upon gravistimulation, amyloplasts sediment to the gravity-associated bottom of the statocytes and trigger responses resulting in gravitropic bending (Sack et al., 1986; Saito et al., 2005; Leitz et al., 2009). The dense starch accumulation in amyloplasts suggests an important function of starch biosynthesis for gravitropism. In the Arabidopsis phosphoglucomutase (pgm) mutant, amyloplast sedimentation is dramatically affected due to a low starch content, which correlates with a reduced gravitropic response in roots as well as shoots (Caspar and Pickard, 1989; Kiss et al., 1989, 1997; Weise and Kiss, 1999). Restoration of the gravitropic response in the pgm mutant upon exposure to a hypergravity stimulus indicates that the amyloplast starch levels might be an important factor in the gravity sensing (Fitzelle and Kiss, 2001). This is further supported by the starch excess 1 (sex1) mutant phenotype. Due to a defective starch breakdown, amyloplasts are larger in the endodermal cells of light-grown sex1 shoots, while being unaffected in the root columella cells (Caspar et al., 1991). Consistently, gravitropic sensitivity increases dramatically in light-grown shoots, whilst remaining unaltered in the root (Vitha et al., 2007). Altogether, these results indicate that amyloplasts rather than the enclosed starch function as statoliths. However, starch promotes amyloplasts sedimentation and therefore represents an important factor in the control of the gravitropic response (Morita, 2010).
Amyloplast movements differ substantially between root and shoot statocytes. In roots, amyloplast sedimentation invariably occurs at the gravity-associated bottom of the statocytes (Yoder et al., 2001). In shoots, amyloplasts often exhibit dynamic saltatory movements rather than a static sedimentation, and are more unlikely to sediment to the gravity-associated statocytes’ bottom (Saito et al., 2005; Hashiguchi et al., 2013). Shoot statocytes contain large vacuoles and amyloplasts are enclosed by transvacuolar strands, i.e., cytoplasm enclosed by the tonoplast (Clifford et al., 1989). Amyloplast sedimentation through the vacuolar strands of shoot statocytes suggests vacuole involvement in gravity perception (Lawton et al., 1986; Clifford et al., 1989; Morita et al., 2002; Saito et al., 2005). Indeed, the phenotype of mutants in SGR2, SGR3, ZIG/SGR4, and GRV2/SGR8/KAM2, encoding proteins involved in membrane trafficking, vacuole formation and functioning, supports the importance of the central vacuole in shoot gravity sensing (Kato et al., 2002; Morita et al., 2002; Yano et al., 2003; Saito et al., 2005; Silady et al., 2008; Hashiguchi et al., 2013). The fact that vacuoles are Ca2+ and H+ reservoirs, might hint at an indirect role of vacuoles in gravity sensing and signal transduction (Perbal and Driss-Ecole, 2003). Root gravitropism remains unaffected in the sgr2, sgr3, zig/sgr4, and grv2/sgr8/kam2 mutants, excluding the importance of vacuoles in gravity perception in the root (Kato et al., 2002; Morita et al., 2002; Yano et al., 2003; Silady et al., 2008). Root columella cells exhibit a large ER, small vacuoles and abundant cytoplasm, and amyloplasts are not enclosed by the vacuolar membrane and thus sediment independently from the vacuole (Saito et al., 2005). The ER could fulfil the role of Ca2+ and H+ reservoir and amyloplast-ER interactions could mediate the signal transduction after gravity perception in the root (Perbal and Driss-Ecole, 2003). The role of Ca2+ and H+ as signals involved in the control of gravitropism will be discussed in detail below.
Actin filaments have been identified as important mediators of gravitropism (reviewed by Blancaflor, 2002). They occur within the transvacuolar strands in shoot statocytes and surround amyloplasts in root statocytes (Traas et al., 1987; Tominaga et al., 2000; Saito et al., 2005). The enhanced root and shoot gravitropism upon disruption of actin microfilaments indicates an inhibitory function (Yamamoto and Kiss, 2002; Hou et al., 2004; Palmieri and Kiss, 2005; Saito et al., 2005). The negative role of actin in gravisensing is supported by the observation that saltatory movements of shoot endodermal amyloplasts are increased in the Arabidopsis mutant sgr9 and that amyloplasts do not sediment but rather move around, or cluster together, entangled in actin filaments (Nakamura et al., 2011). Disruption of actin in sgr9 by either Latrunculin B or mutation of FRIZZYSHOOT1 (FIZ1), involved in actin microfilament formation (Kato et al., 2010), restores normal amyloplast sedimentation and thus normal shoot gravitropism. SGR9, a RING-type E3 ligase, presumably regulates the dissociation of amyloplasts from the actin microfilaments and thereby control the balance between amyloplast sedimentation and saltatory movements in the shoot endodermal cells (Nakamura et al., 2011). Hence, actin microfilaments probably regulate gravity perception by fine-tuning the dynamics of amyloplast sedimentation and saltatory movements.
Signal Transduction
Amyloplast sedimentation in response to gravity is considered to excite a biochemical signal that converts gravity perception into gravitropic curvature. Rapid modulation of the cytoplasmic pH, Ca2+ and inositol 1,4,5-triphosphate (InsP3) levels in response to a gravistimulus qualified them as early signaling molecules. How these signaling molecules might coordinate gravitropic bending and possibly the asymmetric redistribution of auxin, the hormonal signal coordinating differential cell growth, will be discussed in the following paragraph.
Coordinated Auxin Redistribution During Gravitropic Responses
Similar to phototropism, gravitropic curvature results from a differential auxin distribution (Cholodny, 1927; Went, 1928; Evans, 1991). In gravistimulated roots, auxin accumulates in the epidermal cells at the lower side of the root apex and coordinates bending of the root through the inhibition of cell elongation (Friml et al., 2002; Ottenschläger et al., 2003). Monitoring of the auxin dynamics using the auxin sensitive reporter (DII)-VENUS demonstrated that auxin rapidly relocalizes to the lower side of roots within minutes after gravistimulation (Band et al., 2012; Brunoud et al., 2012). Research on the mechanism controlling this rapid auxin redistribution upon gravistimulation, provided a detailed picture on the directionality of the auxin flow, and demonstrated that the auxin transport machinery acting at the root tip was instrumental (Bennett et al., 1996; Gälweiler et al., 1998; Luschnig et al., 1998; Müller et al., 1998; Friml et al., 2002; Swarup et al., 2005; Harrison and Masson, 2008). Auxin is transported acropetally through the central vasculature toward the root tip. Auxin transporters including the AUX1 influx carrier and the PIN1, PIN4, PIN7 as well as ABCB1 and ABCB19 efflux carriers procure the auxin movement in this direction. In the columella, AUX1, PIN3 and PIN2 regulate the basipetal auxin transport through the lateral root cap and epidermis toward the root elongation zone where the auxin distribution controls the dynamics of cell growth. However, what are the mechanisms triggering the differential auxin transport resulting in local auxin maxima at the lower side of the root elongation zone upon gravistimulation? The agravitropic phenotype of the aux1, pin2, pin3 and pin3, pin7 mutants delineated AUX1, PIN2, PIN3 and PIN7 as key transporters coordinating the auxin redistribution upon gravistimulation. Subsequent studies on the polar membrane localization and subcellular dynamics of the auxin transporters, revealed that within a few minutes after gravistimulation, the PIN3 and PIN7localization changed from apolar to polar with maxima at the bottom of root columella cells. This rapid PINs relocation might increase the efficiency of the auxin flow toward the lower side of the gravistimulated root (Friml et al., 2002; Kleine-Vehn et al., 2010). Beyond the root columella, AUX1 and PIN2 procure auxin transport through the lateral root cap and epidermis toward the root elongation zone, where the accumulation of auxin limits cell expansion (Luschnig et al., 1998; Müller et al., 1998; Swarup et al., 2005).
Similar to the root, auxin redistribution upon shoot gravistimulation depends on the activity of the polar auxin transport machinery (Rakusová et al., 2011). In the pin3 mutant, the asymmetrical auxin distribution in gravistimulated hypocotyls is significantly reduced and gravitropic responses are limited. Consistently, upon gravistimulation PIN3 relocalizes to the lower side of the endodermal cells where it mediates an asymmetrical auxin distribution, resulting in auxin accumulation to the lower side of the gravistimulated shoot (Friml et al., 2002; Rakusová et al., 2011).
Redirection of the auxin stream after gravistimulation requires a rapid adaptation of the auxin transport machinery. PIN3 and PIN7 polarization in both roots and PIN3 in shoots presumably occurs via ARF-GEF GNOM-dependent protein transcytosis (Geldner et al., 2003; Kleine-Vehn et al., 2010; Rakusová et al., 2011). Upon gravistimulation PIN3 polarization was demonstrated to rest also on PID kinase-dependent regulation. PID overexpression impeded gravity-induced PIN3 polarization and consequently caused a defective shoot gravitropism, whereas hypocotyl growth remained unaltered. On the other hand, in pid loss-of-function seedlings gravity still induced an asymmetrical PIN3 localization (Rakusová et al., 2011). Apart from PID, D6PK, also a member of the AGC kinase family, was shown to regulate shoot gravitropism. Shoot gravitropism is defective in d6pk mutants. The reduced phosphorylation of PIN3, suggest D6PK involvement in shoot gravitropism through regulation of PIN3 phosphorylation and therefore auxin transport (Zourelidou et al., 2009; Willige et al., 2013). D6PK levels at the plasma membrane correlate with PIN3 phosphorylation and consequently PIN3-dependent auxin efflux. Plasma membrane D6PK abundance depends on ARF-GEF GNOM activity (Barbosa et al., 2014), which is in line with the dependency of PIN3 polarization on ARF-GEF GNOM in shoot gravitropism (Geldner et al., 2003; Kleine-Vehn et al., 2010; Rakusová et al., 2011). D6PK polarity is independent from PID (Barbosa et al., 2014), excluding D6PK function directly down-stream from PID in the regulation of auxin transport in shoot gravitropism. Similarly, PIN2-mediated auxin transport is rapidly altered to promote auxin redistribution upon gravistimulation of the root. Whereas the PIN2 level increases in the epidermal cells at the lower side of the root apex, at the upper side PIN2 undergoes endocytosis and lytic degradation (Abas et al., 2006). Again, PID kinase was shown to regulate auxin transport and gravitropism by targeting the PIN2 auxin efflux carrier in the root. The pid-9 mutant is characterized by reduced PIN2-mediated basipetal auxin transport and gravitropism. However, in contrast with PID overexpression, the asymmetrical auxin distribution of PIN2 upon gravistimulation remained unaltered in the pid-9 mutant, suggesting a PIN2 polarity-independent regulation of auxin transport during gravitropism (Sukumar et al., 2009). Similar to negative phototropism in the root, PHYTOCHROME KINASE SUBSTRATE1 (PKS1) seems also involved in root gravitropism. However, whilst PKS1 positively regulates root phototropism, root gravitropism is counteracted by PKS1. The gravitropic response of etiolated roots is enhanced in the pks1 mutant, whereas it is slightly reduced upon overexpression of PKS1 (Boccalandro et al., 2008). On the other hand, shoot gravitropism is normal in etiolated pks1pks2pks4 mutants (Lariguet et al., 2006). Overexpression of PKS4, however, inhibited shoot gravitropism whilst upon mutation of PKS4 the gravitropic response of etiolated seedlings was not substantially affected (Schepens et al., 2008), suggesting redundancy among the PKS kinases. Hence, the PKS family plays important functions during light-regulated tropic growth responses, whereas in darkness their involvement in gravitropism seems minor and requires more research.
Biochemical Signals Involved in Signal Transduction Upon Gravity Sensing
Although full understanding of the molecular mechanisms controlling gravitropic bending and auxin transport modulation after a gravistimulus is still lacking, there are several indications for the involvement of signaling molecules such as Ca2+ and InsP3 (inositol1,4,5-trisphosphate) in these regulations.
Calcium has been implicated as a signal in the transduction of gravity sensing to differential growth (Sinclair and Trewavas, 1997; Fasano et al., 2002). Gravistimulation induces an increase in the cytoplasmic Ca2+ in the shoot within seconds (Legué et al., 1997; Plieth and Trewavas, 2002; Toyota et al., 2008, 2013). In roots, however, it remains obscure whether an increase in cytoplasmic Ca2+ occurs upon gravistimulation (Legué et al., 1997; Toyota et al., 2008). A recent study of Monshausen et al. (2011) reports asymmetrical transient Ca2+ changes, with Ca2+ levels decreasing and increasing at the upper and lower site of gravistimulated roots respectively. These changes in the Ca2+ concentration are, however, much slower compared to shoot gravitropism (minutes compared to seconds). Furthermore, the dependency of both root and shoot gravitropism on Ca2+ signaling is supported by the defective gravitropism upon administration of Ca2+chelators (Lee et al., 1983; Daye et al., 1984; Millet and Pickard, 1988; Toyota et al., 2008). Curvature of the root in response to unilateral administration of Ca2+ also suggests the involvement in differential growth (Hasenstein and Evans, 1988; Ishikawa and Evans, 1992).
Another signaling molecule implicated in gravity sensing and gravitropic curvature is InsP3 (Perera et al., 1999, 2001). Upon gravistimulation, InsP3 levels temporarily increase in Arabidopsis and oat inflorescence stems before returning to basal levels (Perera et al., 2001, 2006). Furthermore, the inhibition of InsP3 production substantially attenuates the graviresponse in Arabidopsis roots and oat shoots (Perera et al., 2001; Andreeva et al., 2010; Salinas-Mondragon et al., 2010). Consistently, inhibition of InsP3-mediated signaling postpones and reduces the gravitropic response of Arabidopsis roots and shoots (Perera et al., 2006; Smith et al., 2013). Nevertheless, no InsP3 receptor was yet identified in plants (Wheeler and Brownlee, 2008; Munnik and Testerink, 2009). InsP3 is an important intermediate in the biosynthesis of InsP6 (inositol hexakisphosphate; Testerink and Munnik, 2011). Furthermore, both InsP3 and InsP6 are able to induce the release of Ca2+ (Krinke et al., 2007; Im et al., 2010; Gillaspy, 2011). Hence, the question remains whether InsP6 rather than InsP3 functions as signaling molecule in gravitropism. The root of the mips1 (myo-Inositol-1-phosphate synthase) mutant, which is characterized by reduced levels of InsP6 (Raboy, 2003), is agravitropic (Chen and Xiong, 2010). The absence of a significant difference in gravitropic root curvature between pin2 and the pin2mips1 double mutant, suggests that the defective root gravitropism in mips1 is due to dysfunction of PIN2, which plays a key role in auxin redistribution upon gravistimulation. This is supported by the slower PIN2 aggregation in mips1 upon Brefeldin A treatment, indicative of an altered PIN2 trafficking (Chen and Xiong, 2010). Further research is required to elucidate whether InsP6 rather than InsP3 or both, are more important in gravitropism.
The function of InsP3 in gravitropic signaling could be linked to that of Ca2+. Gravistimulation could induce IsnP3 production, which in turn could inducea Ca2+ release (Ghosh et al., 1988; Gilroy et al., 1990; Perera et al., 2006). This is supported by the corresponding changes in InsP3 and Ca2+ levels upon gravistimulation (Toyota et al., 2008, 2013). Hence, through Ca2+- and/or InsP3-induced Ca2+ release, a sustained Ca2+ increase could be elicited mediating gravitropic signaling (Toyota et al., 2013). The opening of stretch-activated Ca2+ channels by the mechanical strain exerted by sedimenting amyloplasts on actin microfilaments, could be a mechanism for gravistimulation-induced Ca2+ release (Perbal and Driss-Ecole, 2003). In root statocytes the large ER could function as an intracellular Ca2+ reservoir, which upon contact with sedimenting amyloplasts could release Ca2+. In shoot statocytes on the other hand, the large central vacuole could function as an intracellular Ca2+ reservoir, releasing Ca2+, possibly upon contact with the sedimenting amyloplasts or through actin filament-activated mechanosensitive Ca2+ channels (Perbal and Driss-Ecole, 2003).
Recent works indicated that Ca2+ and InsP3 might control gravitropic responses through the modulation of PIN polar targeting and auxin distribution (Lee and Evans, 1985; Plieth and Trewavas, 2002; Benjamins et al., 2003; Zhang et al., 2011). Firstly, mutation in INOSITOL POLYPHOSPHATE 5-PHOSPHATASE 13 (5PTase13), encoding an enzyme involved in phosphatidylinositol metabolism, was found to enhance the root gravitropic response. The reduced sensitivity of the 5PTase13 mutant to the polar auxin transport inhibitor NPA, denoted the involvement of enhanced polar auxin transport and consistently, an altered PIN2 distribution, with an expression extended to the root elongation zone in the 5PTase13 mutant, has been observed (Wang et al., 2009). Another indication for the role of InsP3 and Ca2+ in the control of the PIN-dependent auxin distribution during gravisignaling emerged from a screen for suppressors of the PIN1 gain-of-function phenotype (Zhang et al., 2011). The ectopic presence of basally localized PIN1 in the epidermal cells resulted in root agravitropism, which was suppressed after mutation of suppressor of PIN1 overexpression (SUPO1). SUPO1 (SAL1/FRY1/HOS2/ALX8/RON1) was shown to code for a protein with inositol polyphosphate 1-phosphatase and 3′(2′),5′-bisphosphate nucleotidase activities (Zhang et al., 2011). Elevated cytosolic InsP3 and Ca2+ levels, either due to the lack of SUPO1 activity or as result of pharmacological treatment, were found to modify the PIN polar membrane localization and consequently the root gravity response (Zhang et al., 2011). Altogether, these findings hint at the role of InsP3-mediated Ca2+ release in the control of PIN-mediated auxin distribution and subsequently auxin-dependent gravitropism.
Apart from an increase in both Ca2+ and InsP3, gravistimulation also induces a rapid increase in the cytoplasmic and cell wall pH of root cap columella cells (Scott and Allen, 1999; Fasano et al., 2001; Johannes et al., 2001). In the Arabidopsis pgm mutant, cytoplasmic alkalization upon gravistimulation is, however, inhibited, indicating the dependency on amyloplast sedimentation. Furthermore, the Arabidopsis mutant arg1 is defective in the gravity-induced cytoplasmic pH changes within the root columella cells, partly explaining the altered root gravitropic response of the mutant (Boonsirichai et al., 2003). Although the increase in cytoplasmic pH in the root cap columella cells is probably important for root gravitropism, the spatial and temporal uniformity of the pH changes in the root cap columella cells indicate that pH changes do not transduce directional gravity information (Fasano et al., 2001).
The function of actin microfilaments in gravitropism is not confined to gravisensing (discussed above). Modulation of the polar membrane localization of auxin transporters after a gravistimulus is largely dependent on the actin cytoskeleton (Geldner et al., 2001). Stabilization of actin microfilaments limits the subcellular dynamics of auxin transporters and consequently inhibits the asymmetrical auxin accumulation and the root gravitropic response (Dhonukshe et al., 2008). The Arabidopsis ALTERED RESPONSE TO GRAVITY 1 (ARG1) and ARG1-LIKE 2 (ARL2) genes further strengthen the role of actin in the regulation of PIN dynamics. ARG1 interacts with the cytoskeleton, supposedly with actin microfilaments, and arg1 and arg1-like 2 mutants exhibit a reduced shoot and root gravitropism, as well as defects in the asymmetrical auxin redistribution (Fukaki et al., 1997; Sedbrook et al., 1999; Boonsirichai et al., 2003; Guan et al., 2003; Harrison and Masson, 2008). Furthermore, rearrangements of the actin cytoskeleton during cell expansion after gravistimulation also suggest its involvement as a regulator of cell elongation (Pozhvanov et al., 2013).
Auxin Coordinates Gravitropic Bending
Similar to phototropism, the accumulation of auxin in gravistimulated organs triggers downstream responses through a transduction cascade consisting of the TIR1/AFB auxin receptor family, ARFs and the Aux/IAA repressor family. Several components of the auxin signaling pathway were demonstrated to contribute to the regulation of the gravitropic response in both shoot and root (reviewed by Masson et al., 2002). Auxin regulates its own efflux during root gravitropism by affecting the abundance of PIN proteins. PIN2 vacuolar targeting and degradation depends on a (SCFTIR1/AFB)-mediated signaling (Baster et al., 2013), whereas PIN2 endocytosis depends on ABP1-mediated signaling (Paciorek et al., 2005). The PIN2 turnover is the result from biosynthesis, endocytosis and vacuolar targeting and degradation. Upon gravistimulation, this PIN2 turnover is differentially regulated by auxin at the upper and lower side of the root to promote its efflux at the lower side, and to inhibit its efflux at the upper side of the gravistimulated root (Paciorek et al., 2005; Baster et al., 2013). Mutation of the MASSUGU 2 (MSG2)/IAA19 gene results in a reduced hypocotyl gravitropic response, whereas growth and developmental processes other than differential growth are unaffected (Watahiki and Yamamoto, 1997; Tatematsu et al., 2004). Likewise, mutation of SOLITARY ROOT (SLR)/IAA14, results in agravitropic roots and shoots (Fukaki et al., 2002). The severely abolished auxin-dependent up-regulation of PIN gene expression in the slr1/iaa14 mutant (Vieten et al., 2005), suggests an Aux/IAA-dependent regulation of auxin transport upon gravitropism. Furthermore, an asymmetrical auxin redistribution in the root in response to gravistimulation triggers the expression of the auxin response factor ARF19 (Band et al., 2012). After mutation of ARF7/NPH4 and ARF19, both hypocotyl and root are characterized by an agravitropic phenotype (Okushima et al., 2005), reminiscent of the phenotype of the slr/iaa14 mutant (Fukaki et al., 2002). However, in contrast to the arf19 single mutant, the arf7/nph4 single mutant is characterized by a reduced shoot gravitropism. Furthermore, neither of these mutations conferred an agravitropic root phenotype (Liscum and Briggs, 1996; Ruegger et al., 1997; Harper et al., 2000; Okushima et al., 2005; Li et al., 2006). ARF7/NPH4 and ARF19 are expressed in both the root and the shoot. ARF19 expression is stronger in the root, whereas ARF7/NPH4 expression is stronger in the shoot (Okushima et al., 2005; Wilmoth et al., 2005; Li et al., 2006). The normal root gravitropic response of the single mutants indicates that these ARFs act largely redundant in the root. In contrast, the stronger expression of ARF7/NPH4 in the shoot, is consistent with the stronger effect of arf7/nph4 on shoot gravitropism, indicating that these ARFs function less redundantly compared to the root. The induced expression of common auxin responsive genes by ARF7/NPH4 and ARF19 supports their redundant functions in gravitropism (Okushima et al., 2005; Wilmoth et al., 2005). Furthermore, whereas the nph4/arf 7 single mutant exhibited no obvious reduction, the auxin-dependent relocation of PIN1 and PIN2 was reduced in the arf7 arf19 double mutant (Sauer et al., 2006), which supports an ARF-dependent auxin transport in root gravitropism. Altogether these results support an Aux/IAA-ARF-dependent regulation of gravitropism, presumably partly through the regulation of auxin transport. Despite evidence for the involvement of several components of the auxin signaling cascade in the control of gravitropic bending, clarification of downstream targets and mechanisms that regulate differential cell growth requires deeper investigation.
Apical Hook Development
Another remarkable example of tightly coordinated differential growth in plants is the development of the apical hook. Typically, dicotyledonous plants form a hook-like structure, which protects the delicate apical meristem and cotyledons from damage when penetrating through the soil (Raz and Ecker, 1999). The apical hook is an exceptionally dynamic structure formed by gradual bending of the apical part of the hypocotyl soon after germination, it is maintained in a closed stage while the hypocotyl penetrates through the soil, and rapidly opens upon light exposure (Raz and Ecker, 1999; Vandenbussche et al., 2010; Žádníková et al., 2010). All phases of the hook development depend on the phase-specific coordination of differential growth and cell division (Raz and Koornneef, 2001). For the summary of the common and different factors involved in the regulation of hook asymmetrical growth see the Table 1.
Apical Hook Formation
Apical hook formation is orchestrated by differential growth along the apical-basal axis of the hypocotyl, with distinct contributions of cell elongation and cell division on the opposite sides of the structure. A few, but significant, cell division events are localized at the apical concave (inner) side of the hook, while cell elongation is reduced and the arc length is shorter. In contrast, cell elongation is an active process at the convex (outer) side of the hook that permits the arc length to increase. At the basal side, the cell elongation rate is equal between the outer and inner sides and, therefore, without curvature. Thus, coordinated differential cell elongation and cell division mechanisms are essential for proper hook development (Silk and Erickson, 1978; Raz and Koornneef, 2001). Hence, bending of the apical part of the hypocotyl during hook formation is reminiscent of root and shoot tropic responses. Similar to tropic responses, an asymmetric distribution of auxin was found to be indispensable for apical hook formation. During apical hook formation, auxin has been shown to define the concave side of the apical curvature, based on polar auxin transport (Raz and Ecker, 1999). Treatment with the auxin efflux inhibitor NPA or the auxin influx inhibitor 1-naphtoxyacetic acid (NOA) abolishes the asymmetric auxin distribution and hook formation (Vandenbussche et al., 2010; Žádníková et al., 2010). Detailed analyses of the components of the polar auxin transport machinery based on mutant phenotype characterizations and expression studies helped to elucidate the key auxin influx and efflux carriers involved in the regulation of auxin transport during apical hook formation. Expression patterns, and membrane polarities of auxin transporters in particular, helped to outline the directions of auxin transport resulting in the formation of the auxin maximum at the concave side of the hook. According to the model, auxin synthesized in the cotyledons, the shoot apical meristem and the apical portion of the hypocotyl is transported basipetally toward the root. The latter basipetal transport is enhanced by the auxin influx carriers of the AUX/LAX family, which control the auxin flow from the cotyledons and shoot apical meristem, and by the efflux carriers PIN1 and PIN3, which act in the central cylinder. Lateral redistribution of auxin through the endodermis is promoted by PIN3 and further through the cortex and the epidermis by AUX1, PIN3 and PIN4. In the cortex, auxin transport is basipetally directed by PIN3 and PIN4 and in the epidermis by PIN3, PIN7, AUX1 and LAX3. Importantly, the accumulation of auxin at the concave side of the apical hook was proposed to result from a PIN asymmetry at the convex versus the concave side. The higher abundance of PIN3 and PIN4 at the cell membranes on the convex compared to concave side of the apical hook presumably enhances the draining of auxin from the outer cortex and epidermal layers and the formation of an auxin maximum at the concave side (Vandenbussche et al., 2010; Žádníková et al., 2010). Other important players in the control of hook formation are upstream regulators of auxin carrier activity. AUX1 exocytosis to the plasma membrane depends on the activity of the trans-Golgi network-localized ECHIDNA protein complex (Boutté et al., 2013). WAG2, a member of the AGC kinase family was found to phosphorylate PIN3 and to regulate establishment of the auxin maxima in the apical hook. Hence, it has been proposed that WAG2 might contribute to the regulation of hook formation through either control of PIN3 activity or polarity; although other PIN-independent mechanism cannot be excluded (Willige et al., 2012).
Although the polarity and expression patterns of auxin transporters provide indications for the direction of the auxin flow resulting in auxin maxima during apical hook formation, primary cues triggering this asymmetric pattern of auxin distribution and hook formation remain obscure. For tropic responses such as gravi-, or phototropism, both triggering stimuli and perception mechanisms are well defined. However, the trigger(s) of apical hook formation remain(s) unknown. Hypothetically, several candidate mechanisms controlling the onset of hook formation are plausible. To fit within a tightly enclosed seed coat, maturing embryos form a typical U-shaped structure with two cotyledons bent toward the embryonic root. Could this embryonic body shape predetermine incipient apical hook formation, after germination? Another factor that might contribute to the control of early apical hook formation is gravity. It has been shown that Helianthus annuus L. seedlings treated in the clinostat display a defective apical hook development, hence, supporting the role of gravisensing in hook formation (Macdonald et al., 1983). On the other hand, pea seedlings grown in a space station were able to form a partially closed apical hook, suggesting additional intrinsic mechanisms (Miyamoto et al., 2014). Thus, it seems that the formation of the apical hook might result from several mutually complementary mechanisms, and their identification and characterization are a challenge for future research.
To control hook formation, auxin locally accumulating at the concave side of the hook requires a functional transduction pathway to activate relevant downstream responses. Attenuation of auxin signaling by the expression of the auxin signaling repressor axr3 at the concave side of the hook prevents hook formation (Vandenbussche et al., 2010). Similarly, auxin perception and signaling mutants, including transport inhibitor response (tir1; Dharmasiri et al., 2005), and auxin resistant1 (axr1), short hypocotyl2-2 (shy2-2), massugu (msg2/iaa19) and non-phototropic hypocotyl4 (nph4/msg1/arf7), auxin response factor19 (arf19; Lehman et al., 1996; Žádníková et al., 2010), respectively, do not form an apical hook, indicating an indispensable role for auxin signaling (Boerjan et al., 1995; Zhao et al., 2001). An important regulator of apical hook development is HOOKLESS1 (HLS1), an ethylene-responsive gene that encodes an N-acetyltransferase. HLS1 was found to inhibit the AUXIN RESPONSE FACTOR2 (ARF2) repressor and, thus, to control the auxin response (Lehman et al., 1996). Apical hook formation is completely prevented in the loss-of-function mutant hls1 (Li et al., 2004), whereas overexpression of HLS1 causes apical hook exaggeration (Lehman et al., 1996). Among the auxin-responsive genes, several members of the SMALL AUXIN UPREGULATED RNA (SAUR) gene family are highly expressed in tissues undergoing differential cell expansion (McClure and Guilfoyle, 1989; Esmon et al., 2006). Arabidopsis seedlings expressing stabilized, auxin-inducible SAUR19-24 proteins are characterized by increased cell expansion, and impaired apical hook development, indicating that SAUR19–24 proteins function as positive effectors of cell expansion and therefore might contribute to the regulation of apical hook development (Spartz et al., 2012). Recent work of Spartz et al. (2014) provided new insights into the molecular mechanisms underlying SAUR19 action and demonstrated that SAUR19, through inhibition of PP2C-D subfamily type 2C protein phosphatases, activates plasma membrane H+ -ATPases to promote cell expansion.
Light Triggered Apical Hook Opening
To ensure a timely start of photosynthesis, the apical hook opens shortly upon seedling exposure to light after emergence from the soil. For most dicotyledonous species, including Arabidopsis, light is the key trigger for hook opening (Liscum and Hangarter, 1993b). However, in some species light might have an opposite effect. Red or far-red light-induced hook exaggeration has been demonstrated to occur in a wide variety of species including tomato (Solanum lycopersicum), carrot (Daucus carota), and okra (Abelmoschus esculentus; Shichijo and Hashimoto, 2013). Those observations indicate that the effect of light on apical hook opening is controlled differently among species.
Light quality and intensity are two important factors that influence apical hook opening. In Arabidopsis, UV, blue, red and far-red light can effectively stimulate apical hook opening, while infrared light has no impact on apical hook development (Liscum and Hangarter, 1993a,b). Low-fluence red and blue light effectively trigger apical hook opening, and this effect is reversible by far-red light (Liscum and Hangarter, 1993b). Low UV-B doses are sufficient to inhibit hypocotyl growth and apical hook opening in etiolated seedlings, independently of phytochromes and cryptochromes (Ballaré et al., 1991; Kim et al., 1998; Li et al., 2013). A recent study demonstrated that a short light pulse is sufficient to trigger apical hook opening. However, this pulsed light effect results in an incompletely opened hook, which may reflect a self-protection mechanism of premature seedlings (Smet et al., 2014).
Apical hook opening is part of a plants’ photoresponse, which is achieved through multiple photoreceptors (Chen et al., 2004; Jiao et al., 2007). In plants, various photoreceptors absorb different light spectra: red and far-red light is mainly absorbed by phytochromes (phyA-phyE; Quail et al., 1995; Rockwell et al., 2006); UV-A and blue light are mainly perceived by cryptochromes (CRY1 and CRY2; Li and Yang, 2007) and phototropins (PHOT1 and PHOT2; Iino, 2006; Kimura and Kagawa, 2006); the UV RESISTANCE LOCUS 8 (UV8) is activated by UV-B light (Rizzini et al., 2011; Tilbrook et al., 2013). Currently, only PHYA, PHYB and CRY1 have been reported to act in the regulation of apical hook development (Reed et al., 1994; Fox et al., 2012; Smet et al., 2014). Phytochromes A and B were proposed to mediate apical hook opening in response to red and far-red light (Liscum and Hangarter, 1993b; Reed et al., 1994; Smet et al., 2014). Loss of CRY1 activity interferes with hook formation and the opening process is faster when compared to the wild type (Fox et al., 2012). Although several studies proposed the role of light signaling in the modulation of auxin transport (Wu et al., 2010) and auxin signaling (Stowe-Evans et al., 2001; Tepperman et al., 2001; DeBlasio et al., 2003; Nagashima et al., 2008; Wu et al., 2010; Kami et al., 2012), the mechanism of how phytochromes and cryptomes coordinate apical hook opening is largely unknown.
Besides phytochromes and cryptochromes, the function of other photoreceptors in apical hook development needs to be investigated. UV-B could possibly trigger apical hook opening (Ballaré et al., 1991). However, whether UVR8 or other yet-to-be identified UV-B receptor(s) control apical hook opening is unknown. Phototropins, receptors mediating phototropic responses, were so far not reported for their role in apical hook opening. In summary, although light-induced apical hook opening is well-established, the molecular mechanism of this process remains largely unknown.
Conclusion
The ability of plant organs to bend in response to different environmental stimuli represents one of the most crucial adaptive mechanisms compensating for the lack of plant locomotion. Differential growth is the common mechanistic basis underlying all forms of tropic movements in plants. Deciphering the molecular components and mechanisms underlying differential growth in response to various environmental stimuli revealed that multiple common elements from perception to signal transduction, as well as specific elements are involved (Table 1; Figure 1). The perception sites for light and gravity differ for the shoot and the root. In addition, the response to these two stimuli rests on different mechanisms. In phototropism, light is perceived through photoreceptors, whereas in gravitropism, gravity is sensed through the sedimentation of starch-containing amyloplasts in statocytes. The triggers and perception site for apical hook formation remain, however, in the dark. To transduce the perception of both light and gravity in organ bending, common, as well as different, biochemical signals and factors are involved. Calcium ions and InsP3 appear as important common biochemical signals in both tropic responses. The increase in cytoplasmic pH in the root cap columella cells in response to gravity is another important biochemical signal, important more specifically for root gravitropism.
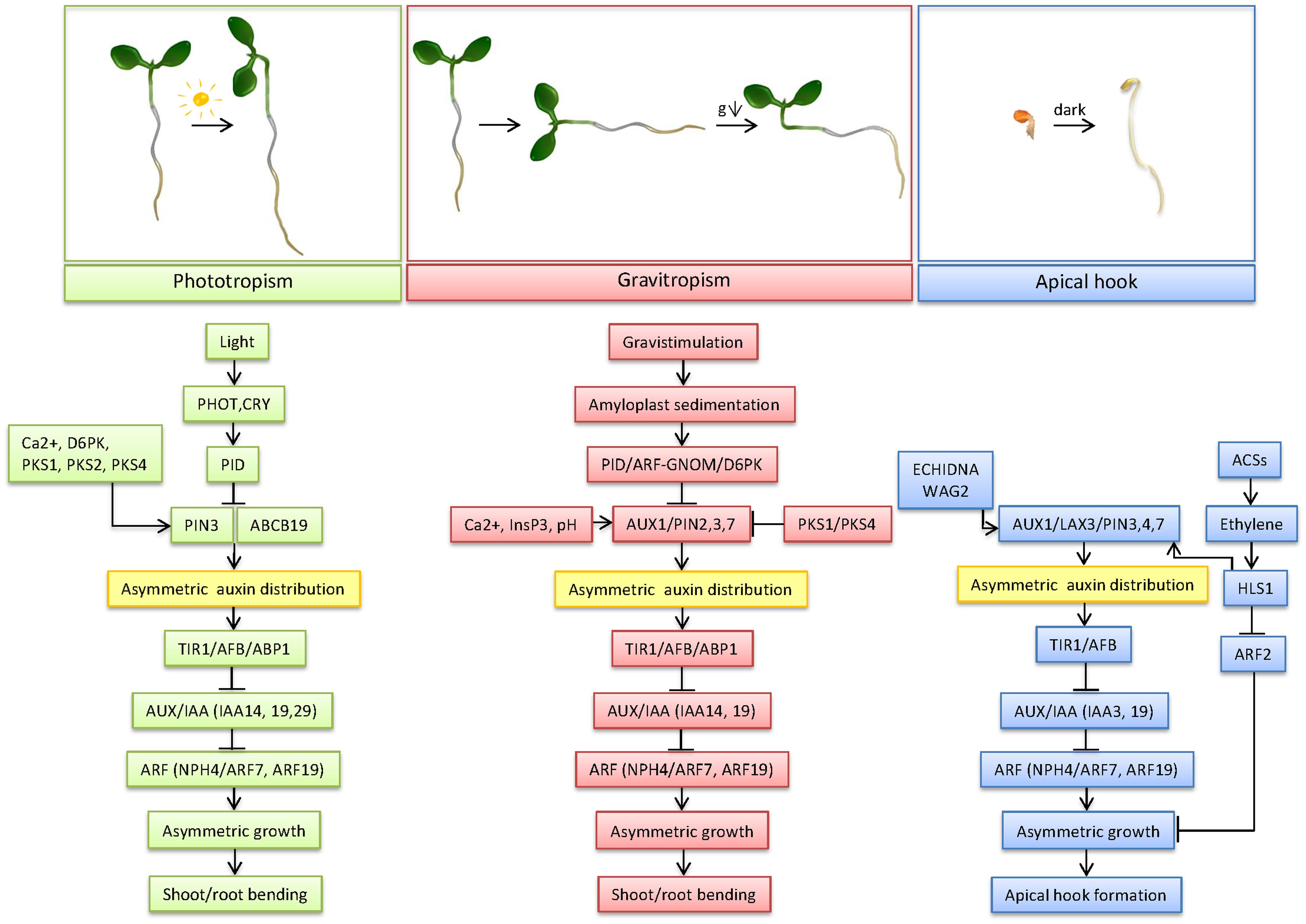
FIGURE 1. Mechanism of differential growth in phototropism, gravitropism and apical hook development. This model depicts the most important factors in the regulation of the three forms of differential growth, from trigger sensing to bending of the concerned organs, centered around auxin transport and signaling as key mechanisms underlying differential growth.
The difference between the perception and the response sites in both gravitropism and phototropism indicates that a signal must be transported between both sites. The polar transport of the plant hormone auxin is indeed indispensable for the differential growth underlying gravitropism, phototropism, as well as apical hook development. In all forms of differential growth, an asymmetrical auxin distribution is found in the bend of the root and the shoot. Despite the firmly established strong correlation between an asymmetric auxin distribution and differential growth, there is a longstanding enigma with respect to auxin accumulation and cell growth patterns (Table 1; Figure 1). Whereas during root gravitropism and apical hook formation auxin accumulates at the inner side of the curvature to restrict cell expansion, shoot gravi- and phototropism and presumably also root phototropism are driven by auxin maxima at the outer side of the curvature, where auxin stimulates cell growth. This apparent discrepancy in auxin accumulation patterns and consequences for local cell growth indicate that auxin might coordinate differential growth through specific mechanisms, which might depend on the developmental context. In addition, auxin is known to regulate cell growth in a concentration-dependent way. At optimal concentrations, auxin stimulates, whereas at supraoptimal concentrations it inhibits cell growth, respectively (Taiz and Zeiger, 2006). Hence, depending on the concentration of auxin, different cell growth responses might be triggered. The molecular basis of the concentration-dependent effects of auxin on cell growth are so far unknown. It might be envisioned that specific circuits within the TIR1/Aux-IAA/ARF network, which in Arabidopsis consists of 6 TIR1/AFBs, 29 Aux/IAAs and 23 ARFs homologues are activated in dependence on the auxin concentrations to mediate certain developmental responses. Such a scenario is strongly supported by the recent study of Calderón Villalobos et al. (2012) demonstrating that the formation of specific TIR1/AFB–Aux/IAA receptor pairs is auxin concentration dependent.
The dependence of all forms of differential growth on an asymmetrical auxin distribution indicates that auxin carriers fulfill an indispensable role. PIN3 efflux carriers appear to be one of the key components of the polar auxin transport machinery involved in gravitropism, phototropism and apical hook formation (Friml et al., 2002; Žádníková et al., 2010; Ding et al., 2011; Rakusová et al., 2011; Haga and Sakai, 2012; Zhang et al., 2013). In the shoot endodermal cells, light, as well as gravity, stimulate pathways targeting the PIN3 auxin efflux carriers and modulating their activity. Similarly, in root columella cells, the PIN3 polarity is changed in response to both light and gravity. Hence, PIN3 proteins are utmost susceptible to changes in light and gravity and promote the redirection of the auxin flow. This implies that pathways mediating qualitatively highly different external inputs might partially overlap and crosstalk to coordinate and/fine-tune responses of organs to different stimuli. Apart from PIN3, other, both different and common, auxin carriers are also involved in the three forms of differential growth. In root gravitropism, PIN2 is, together with AUX1, involved in the basipetal transport of auxin from the lateral root cap and epidermis to the root elongation zone, whereupon auxin is asymmetrically distributed by PIN3. In addition, the auxin efflux carriers PIN2 and PIN1 are also important for negative root phototropism (Wan et al., 2012; Zhang et al., 2014). Apart from root gravitropism, AUX1 is also important for apical hook development, where it regulates the lateral auxin distribution through the cortex and epidermis and basipetal auxin transport in the epidermis together with PIN3, PIN7 and LAX3 (Vandenbussche et al., 2010). PIN7 is also active in root gravitropism. Apical hook opening after light exposure of the shoot relies on ABCB transporters, among which importantly ABCB19 plays a prime role (Wu et al., 2010). This involvement of ABCB transporters also applies consistently to light-induced shoot phototropism (Christie et al., 2011).
Interestingly, several common components of the auxin signaling pathway, such as ARF7/NPH4 and ARF19 as well as IAA14, and IAA19, were found to control the bending of different organs regardless of the nature of the stimulus. This further strengthens the assumption that the developmental context might play a crucial role in defining the specificity of the response. The current knowledge on downstream molecular components and mechanisms which direct differential cell growth during specific tropic responses downstream of auxin is surprisingly scarce. Hence, the identification of factors and detailed understanding of networks coordinating differential growth remain a challenge for future research.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Annick Bleys for her assistance with preparing the manuscript. This work was supported by the European Research Council with a Starting Independent Research grant (ERC-2007-Stg-207362-HCPO) and the and the Grant Agency of the Czech Republic (GACR, GA13-39982S) to EB, and by research grants from the Research Foundation Flanders (G.0656.13N) and Ghent University to DS. DS holds a PhD fellowship from the Research Foundation Flanders. We apologize that, because of space restrictions, the scientific contributions of only a limited number of original articles could be cited and discussed.
References
Abas, L., Benjamins, R., Malenica, N., Paciorek, T., Wiśniewska, J., Wirniewska, J., et al. (2006). Intracellular trafficking and proteolysis of the Arabidopsis auxin-efflux facilitator PIN2 are involved in root gravitropism. Nat. Cell Biol. 8, 249–256. doi: 10.1038/ncb1369
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ahmad, M., Jarillo, J. A., Smirnova, O., and Cashmore, A. R. (1998). The CRY1 blue light photoreceptor of Arabidopsis interacts with phytochrome A in vitro. Mol. Cell 1, 939–948. doi: 10.1016/S1097-2765(00)80094-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Andreeva, Z., Barton, D., Armour, W. J., Li, M. Y., Liao, L.-F., McKellar, H. L., et al. (2010). Inhibition of phospholipase C disrupts cytoskeletal organization and gravitropic growth in Arabidopsis roots. Planta 232, 1263–1279. doi: 10.1007/s00425-010-1256-0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ballaré, C. L., Barnes, P. W., and Kendrick, R. E. (1991). Photomorphogenic effects of UV-B radiation on hypocotyl elongation in wild type and stable-phytochrome-deficient mutant seedlings of cucumber. Physiol. Plant. 83, 652–658. doi: 10.1111/j.1399-3054.1991.tb02483.x
Baluska, F., Wojtaszek, P., Volkmann, D., and Barlow, P. (2003). The architecture of polarized cell growth: the unique status of elongating plant cells. Bioessays 25, 569–576. doi: 10.1002/bies.10282
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Band, L. R., Wells, D. M., Larrieu, A., Sun, J., Middleton, A. M., French, A. P., et al. (2012). Root gravitropism is regulated by a transient lateral auxin gradient controlled by a tipping-point mechanism. Proc. Natl. Acad. Sci. U.S.A. 109, 4668–4673. doi: 10.1073/pnas.1201498109
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Barbosa, I. C. R., Zourelidou, M., Willige, B. C., Weller, B., and Schwechheimer, C. (2014). D6 PROTEIN KINASE activates auxin transport-dependent growth and PIN-FORMED phosphorylation at the plasma membrane. Dev. Cell 29, 674–685. doi: 10.1016/j.devcel.2014.05.006
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Baskin, T. I., and Bivens, N. J. (1995). Stimulation of radial expansion in Arabidopsis roots by inhibitors of actomyosin and vesicle secretion but not by various inhibitors of metabolism. Planta 197, 514–521. doi: 10.1007/BF00196673
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Baster, P., Robert, S., Kleine-Vehn, J., Vanneste, S., Kania, U., Grunewald, W., et al. (2013). SCF(TIR1/AFB)-auxin signalling regulates PIN vacuolar trafficking and auxin fluxes during root gravitropism. EMBO J. 32, 260–274. doi: 10.1038/emboj.2012.310
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Baum, G., Long, J. C., Jenkins, G. I., and Trewavas, A. J. (1999). Stimulation of the blue light phototropic receptor NPH1 causes a transient increase in cytosolic Ca2+. Proc. Natl. Acad. Sci. U.S.A. 96, 13554–13559. doi: 10.1073/pnas.96.23.13554
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Benfey, P. N., Linstead, P. J., Roberts, K., Schiefelbein, J. W., Hauser, M. T., and Aeschbacher, R. A. (1993). Root development in Arabidopsis: four mutants with dramatically altered root morphogenesis. Development 119, 57–70.
Benjamins, R., Ampudia, C. S. G., Hooykaas, P. J. J., and Offringa, R. (2003). PINOID-mediated signaling involves calcium-binding proteins. Plant Physiol. 132, 1623–1630. doi: 10.1104/pp.103.019943
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bennett, M. J., Marchant, A., Green, H. G., May, S. T., Ward, S. P., Millner, P. A., et al. (1996). Arabidopsis AUX1 gene: a permease-like regulator of root gravitropism. Science 273, 948–950. doi: 10.1126/science.273.5277.948
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Blancaflor, E. B. (2002). The cytoskeleton and gravitropism in higher plants. J. Plant Growth Regul. 21, 120–136. doi: 10.1007/s003440010041
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Blancaflor, E. B., Fasano, J. M., and Gilroy, S. (1998). Mapping the functional roles of cap cells in the response of Arabidopsis primary roots to gravity. Plant Physiol. 116, 213–222. doi: 10.1104/pp.116.1.213
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boccalandro, H. E., De Simone, S. N., Bergmann-Honsberger, A., Schepens, I., Fankhauser, C., and Casal, J. J. (2008). PHYTOCHROME KINASE SUBSTRATE1 regulates root phototropism and gravitropism. Plant Physiol. 146, 108–115. doi: 10.1104/pp.107.106468
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boerjan, W., Cervera, M. T., Delarue, M., Beeckman, T., Dewitte, W., Bellini, C., et al. (1995). Superroot, a recessive mutation in Arabidopsis, confers auxin overproduction. Plant Cell 7, 1405–1419. doi: 10.1105/tpc.7.9.1405
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bögre, L., Okrész, L., Henriques, R., and Anthony, R. G. (2003). Growth signalling pathways in Arabidopsis and the AGC protein kinases. Trends Plant Sci. 8, 424–431. doi: 10.1016/S1360-1385(03)00188-2
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boonsirichai, K., Sedbrook, J. C., Chen, R., Gilroy, S., and Masson, P. H. (2003). ALTERED RESPONSE TO GRAVITY is a peripheral membrane protein that modulates gravity-induced cytoplasmic alkalinization and lateral auxin transport in plant statocytes. Plant Cell 15, 2612–2625. doi: 10.1105/tpc.015560
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boutté, Y., Jonsson, K., McFarlane, H. E., Johnson, E., Gendre, D., Swarup, R., et al. (2013). ECHIDNA-mediated post-Golgi trafficking of auxin carriers for differential cell elongation. Proc. Natl. Acad. Sci. U.S.A. 110, 16259–16264. doi: 10.1073/pnas.1309057110
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boyer, J. S., Cavalieri, A. J., and Schulze, E. -D. (1985). Control of the rate of cell enlargement: excision, wall relaxation, and growth-induced water potentials. Planta 163, 527–543. doi: 10.1007/BF00392710
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Boysen-Jensen, P. (1911). La transmission de l’irritation phototropique dans l’avena. Bull. Acad. R. Des Sci. Lett. 3, 1–24.
Briggs, W. R. (1963). Mediation of phototropic responses of corn coleoptiles by lateral transport of auxin. Plant Physiol. 38, 237–247. doi: 10.1104/pp.38.3.237
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Briggs, W. R., Beck, C. F., Cashmore, A. R., Christie, J. M., Hughes, J., Jarillo, J. A., et al. (2001). The phototropin family of photoreceptors. Plant Cell 13, 993–997. doi: 10.1105/tpc.13.5.993
Brunoud, G., Wells, D. M., Oliva, M., Larrieu, A., Mirabet, V., Burrow, A. H., et al. (2012). A novel sensor to map auxin response and distribution at high spatio-temporal resolution. Nature 482, 103–106. doi: 10.1038/nature10791
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Burton, R. A., Gidley, M. J., and Fincher, G. B. (2010). Heterogeneity in the chemistry, structure and function of plant cell walls. Nat. Chem. Biol. 6, 724–732. doi: 10.1038/nchembio.439
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Calderón Villalobos, L. I. A., Lee, S., De Oliveira, C., Ivetac, A., Brandt, W., Armitage, L., et al. (2012). A combinatorial TIR1/AFB-Aux/IAA co-receptor system for differential sensing of auxin. Nat. Chem. Biol. 8, 477–485. doi: 10.1038/nchembio.926
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Caspar, T., Lin, T. P., Kakefuda, G., Benbow, L., Preiss, J., and Somerville, C. (1991). Mutants of Arabidopsis with altered regulation of starch degradation. Plant Physiol. 95, 1181–1188. doi: 10.1104/pp.95.4.1181
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Caspar, T., and Pickard, B. G. (1989). Gravitropism in a starchless mutant of Arabidopsis: implications for the starch-statolith theory of gravity sensing. Planta 177, 185–197. doi: 10.1007/BF00392807
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Castillon, A., Shen, H., and Huq, E. (2007). Phytochrome interacting factors: central players in phytochrome-mediated light signaling networks. Trends Plant Sci. 12, 514–521. doi: 10.1016/j.tplants.2007.10.001
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chen, H., and Xiong, L. (2010). myo-inositol-1-phosphate synthase is required for polar auxin transport and organ development. J. Biol. Chem. 285, 24238–24247. doi: 10.1074/jbc.M110.123661
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chen, M., Chory, J., and Fankhauser, C. (2004). Light signal transduction in higher plants. Annu. Rev. Genet. 38, 87–117. doi: 10.1146/annurev.genet.38.072902.092259
Cholodny, N. (1928). Beiträge zur hormonalen Theorie von Tropismen. Planta 6, 118–134. doi: 10.1007/BF01915869
Christie, J. M., Reymond, P., Powell, G. K., Bernasconi, P., Raibekas, A. A., Liscum, E., et al. (1998). Arabidopsis NPH1: a flavoprotein with the properties of a photoreceptor for phototropism. Science 282, 1698–1701. doi: 10.1126/science.282.5394.1698
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Christie, J. M., Yang, H., Richter, G. L., Sullivan, S., Thomson, C. E., Lin, J., et al. (2011). phot1 inhibition of ABCB19 primes lateral auxin fluxes in the shoot apex required for phototropism. PLoS Biol. 9:e1001076. doi: 10.1371/journal.pbio.1001076
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Clifford, P. E., Douglas, S., and Mccartney, G. W. (1989). Amyloplast sedimentation in shoot statocytes having a large, central vacuole: further interpretation from electron microscopy. J. Exp. Bot. 40, 1341–1346. doi: 10.1093/jxb/40.12.1341
Cosgrove, D. J. (1993). How do plant cell walls extend? Plant Physiol. 102, 1–6. doi: 10.1104/pp.102.1.1
Cosgrove, D. J. (2005). Growth of the plant cell wall. Nat. Rev. Mol. Cell Biol. 6, 850–861. doi: 10.1038/nrm1746
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Darwin, C., and Darwin, F. (1881). The Power of Movement in Plants. New York, NY: D. Appleton and Co.
Daye, S., Biro, R. L., and Roux, S. J. (1984). Inhibition of gravitropism in oat coleoptiles by the calcium chelator, ethyleneglycol-bis-(beta-aminoethyl ether)-N,N’-tetraacetic acid. Physiol. Plant. 61, 449–454. doi: 10.1111/j.1399-3054.1984.tb06354.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
DeBlasio, S. L., Mullen, J. L., Luesse, D. R., and Hangarter, R. P. (2003). Phytochrome modulation of blue light-induced chloroplast movements in Arabidopsis. Plant Physiol. 133, 1471–1479. doi: 10.1104/pp.103.029116
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dharmasiri, N., Dharmasiri, S., Weijers, D., Lechner, E., Yamada, M., Hobbie, L., et al. (2005). Plant development is regulated by a family of auxin receptor F box proteins. Dev. Cell 9, 109–119. doi: 10.1016/j.devcel.2005.05.014
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dhonukshe, P., Grigoriev, I., Fischer, R., Tominaga, M., Robinson, D. G., Hasek, J., et al. (2008). Auxin transport inhibitors impair vesicle motility and actin cytoskeleton dynamics in diverse eukaryotes. Proc. Natl. Acad. Sci. U.S.A. 105, 4489–4494. doi: 10.1073/pnas.0711414105
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ding, Z., Galván-Ampudia, C. S., Demarsy, E., Łangowski,Ł., Kleine-Vehn, J., Fan, Y., et al. (2011). Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis. Nat. Cell Biol. 13, 447–452. doi: 10.1038/ncb2208
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Effendi, Y., Rietz, S., Fischer, U., and Scherer, G. F. E. (2011). The heterozygous abp1/ABP1 insertional mutant has defects in functions requiring polar auxin transport and in regulation of early auxin-regulated genes. Plant J. 65, 282–294. doi: 10.1111/j.1365-313X.2010.04420.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Epel, B. L., Warmbrodt, R. P., and Bandurski, R. S. (1992). Studies on the longitudinal and lateral transport of IAA in the shoots of etiolated corn seedlings. J. Plant Physiol. 140, 310–318. doi: 10.1016/S0176-1617(11)81084-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Esmon, C. A., Tinsley, A. G., Ljung, K., Sandberg, G., Hearne, L. B., and Liscum, E. (2006). A gradient of auxin and auxin-dependent transcription precedes tropic growth responses. Proc. Natl. Acad. Sci. U.S.A. 103, 236–241. doi: 10.1073/pnas.0507127103
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Evans, M. L. (1991). Gravitropism: interaction of sensitivity modulation and effector redistribution. Plant Physiol. 95, 1–5. doi: 10.1104/pp.95.1.1
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fasano, J. M., Massa, G. D., and Gilroy, S. (2002). Ionic signaling in plant responses to gravity and touch. J. Plant Growth Regul. 21, 71–88. doi: 10.1007/s003440010049
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fasano, J. M., Swanson, S. J., Blancaflor, E. B., Dowd, P. E., Kao, T. H., and Gilroy, S. (2001). Changes in root cap pH are required for the gravity response of the Arabidopsis root. Plant Cell 13, 907–921. doi: 10.1105/tpc.13.4.907
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fitzelle, K. J., and Kiss, J. Z. (2001). Restoration of gravitropic sensitivity in starch-deficient mutants of Arabidopsis by hypergravity. J. Exp. Bot. 52, 265–275. doi: 10.1093/jexbot/52.355.265
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Folta, K. M., Lieg, E. J., Durham, T., and Spalding, E. P. (2003). Primary inhibition of hypocotyl growth and phototropism depend differently on phototropin-mediated increases in cytoplasmic calcium induced by blue light. Plant Physiol. 133, 1464–1470. doi: 10.1104/pp.103.024372
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fox, A. R., Soto, G. C., Jones, A. M., Casal, J. J., Muschietti, J. P., and Mazzella, M. A. (2012). cry1 and GPA1 signaling genetically interact in hook opening and anthocyanin synthesis in Arabidopsis. Plant Mol. Biol. 80, 315–324. doi: 10.1007/s11103-012-9950-x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Friml, J., Wiśniewska, J., Benková, E., Mendgen, K., and Palme, K. (2002). Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 415, 806–809. doi: 10.1038/415806a
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fujihira, K., Kurata, T., Watahiki, M. K., Karahara, I., and Yamamoto, K. T. (2000). An agravitropic mutant of Arabidopsis, endodermal-amyloplast less 1, that lacks amyloplasts in hypocotyl endodermal cell layer. Plant Cell Physiol. 41, 1193–1199. doi: 10.1093/pcp/pcd046
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fukaki, H., Fujisawa, H., and Tasaka, M. (1996). How do plant shoots bend up? the initial step to elucidate the molecular mechanisms of shoot gravitropism using Arabidopsis thaliana. J. Plant Res. 109, 129–137. doi: 10.1007/BF02344538
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fukaki, H., Fujisawa, H., and Tasaka, M. (1997). The RHG gene is involved in root and hypocotyl gravitropism in Arabidopsis thaliana. Plant Cell Physiol. 38, 804–810. doi: 10.1093/oxfordjournals.pcp.a029238
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fukaki, H., Tameda, S., Masuda, H., and Tasaka, M. (2002). Lateral root formation is blocked by a gain-of-function mutation in the SOLITARY-ROOT/IAA14 gene of Arabidopsis. Plant J. 29, 153–168. doi: 10.1046/j.0960-7412.2001.01201.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fukaki, H., Wysocka-Diller, J., Kato, T., Fujisawa, H., Benfey, P. N., and Tasaka, M. (1998). Genetic evidence that the endodermis is essential for shoot gravitropism in Arabidopsis thaliana. Plant J. 14, 425–430. doi: 10.1046/j.1365-313X.1998.00137.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gälweiler, L., Guan, C., Müller, A., Wisman, E., Mendgen, K., Yephremov, A., et al. (1998). Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 282, 2226–2230. doi: 10.1126/science.282.5397.2226
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Geldner, N., Anders, N., Wolters, H., Keicher, J., Kornberger, W., Muller, P., et al. (2003). The Arabidopsis GNOM ARF-GEF mediates endosomal recycling, auxin transport, and auxin-dependent plant growth. Cell 112, 219–230. doi: 10.1016/S0092-8674(03)00003-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Geldner, N., Friml, J., Stierhof, Y. D., Jürgens, G., and Palme, K. (2001). Auxin transport inhibitors block PIN1 cycling and vesicle trafficking. Nature 413, 425–428. doi: 10.1038/35096571
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ghosh, T. K., Eis, P. S., Mullaney, J. M., Ebert, C. L., and Gill, D. L. (1988). Competitive, reversible, and potent antagonism of inositol 1,4,5-trisphosphate-activated calcium release by heparin. J. Biol. Chem. 263, 11075–11079.
Gillaspy, G. E. (2011). The cellular language of myo-inositol signaling. New Phytol. 192, 823–839. doi: 10.1111/j.1469-8137.2011.03939.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gilroy, S., Read, N. D., and Trewavas, A. J. (1990). Elevation of cytoplasmic calcium by caged calcium or caged inositol triphosphate initiates stomatal closure. Nature 346, 769–771. doi: 10.1038/346769a0
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Guan, C., Rosen, E. S., Boonsirichai, K., Poff, K. L., and Masson, P. H. (2003). The ARG1-LIKE2 gene of Arabidopsis functions in a gravity signal transduction pathway that is genetically distinct from the PGM pathway. Plant Physiol. 133, 100–112. doi: 10.1104/pp.103.023358
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Haberlandt, G. (1900). Über die perception des geotropischen reizes. Ber. Dtsch. Bot. Ges. 18, 261–272.
Haga, K., Hayashi, K.-I., and Sakai, T. (2014). PINOID AGC kinases are necessary for phytochrome-mediated enhancement of hypocotyl phototropism in Arabidopsis. Plant Physiol. 166, 1535–1545. doi: 10.1104/pp.114.244434
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Haga, K., and Sakai, T. (2012). PIN auxin efflux carriers are necessary for pulse-induced but not continuous light-induced phototropism in Arabidopsis. Plant Physiol. 160, 763–776. doi: 10.1104/pp.112.202432
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Han, I.-S., Tseng, T.-S., Eisinger, W., and Briggs, W. R. (2008). Phytochrome A regulates the intracellular distribution of phototropin 1-green fluorescent protein in Arabidopsis thaliana. Plant Cell 20, 2835–2847. doi: 10.1105/tpc.108.059915
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Harada, A., and Shimazaki, K. (2007). Phototropins and blue light-dependent calcium signaling in higher plants. Photochem. Photobiol. 83, 102–111. doi: 10.1562/2006-03-08-IR-837
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Harper, R. M., Stowe-Evans, E. L., Luesse, D. R., Muto, H., Tatematsu, K., Watahiki, M. K., et al. (2000). The NPH4 locus encodes the auxin response factor ARF7, a conditional regulator of differential growth in aerial Arabidopsis tissue. Plant Cell 12, 757–770. doi: 10.1105/tpc.12.5.757
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Harrison, B. R., and Masson, P. H. (2008). ARL2, ARG1 and PIN3 define a gravity signal transduction pathway in root statocytes. Plant J. 53, 380–392. doi: 10.1111/j.1365-313X.2007.03351.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hasenstein, K. H., and Evans, M. L. (1988). Effects of cations on hormone transport in primary roots of Zea mays1. Plant Physiol. 86, 890–894. doi: 10.1104/pp.86.3.890
Hashiguchi, Y., Tasaka, M., and Morita, M. T. (2013). Mechanism of higher plant gravity sensing. Am. J. Bot. 100, 91–100. doi: 10.3732/ajb.1200315
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Holweg, C., Süsslin, C., and Nick, P. (2004). Capturing in vivo dynamics of the actin cytoskeleton stimulated by auxin or light. Plant Cell Physiol. 45, 855–863. doi: 10.1093/pcp/pch102
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hou, G., Kramer, V. L., Wang, Y.-S., Chen, R., Perbal, G., Gilroy, S., et al. (2004). The promotion of gravitropism in Arabidopsis roots upon actin disruption is coupled with the extended alkalinization of the columella cytoplasm and a persistent lateral auxin gradient. Plant J. 39, 113–125. doi: 10.1111/j.1365-313X.2004.02114.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Huala, E., Oeller, P. W., Liscum, E., Han, I. S., Larsen, E., and Briggs, W. R. (1997). Arabidopsis NPH1: a protein kinase with a putative redox-sensing domain. Science 278, 2120–2123. doi: 10.1126/science.278.5346.2120
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Iino, M. (2006). Toward understanding the ecological functions of tropisms: interactions among and effects of light on tropisms. Curr. Opin. Plant Biol. 9, 89–93. doi: 10.1016/j.pbi.2005.11.012
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Im, Y. J., Phillippy, B. Q., and Perera, I. Y. (2010). “InsP3 in plant cells,” in Lipid Signaling in Plants, ed. T. Munnik (Berlin: Springer), 145–160. doi: 10.1007/978-3-642-03873-0_10
Ishikawa, H., and Evans, M. L. (1992). Induction of curvature in maize roots by calcium or by thigmostimulation: role of the postmitotic isodiametric growth zone. Plant Physiol. 100, 762–768. doi: 10.1104/pp.100.2.762
Jeong, J., and Choi, G. (2013). Phytochrome-interacting factors have both shared and distinct biological roles. Mol. Cells 35, 371–380. doi: 10.1007/s10059-013-0135-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Jiao, Y., Lau, O. S., and Deng, X. W. (2007). Light-regulated transcriptional networks in higher plants. Nat. Rev. Genet. 8, 217–230. doi: 10.1038/nrg2049
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Johannes, E., Collings, D. A., Rink, J. C., and Allen, N. S. (2001). Cytoplasmic pH dynamics in maize pulvinal cells induced by gravity vector changes. Plant Physiol. 127, 119–130. doi: 10.1104/pp.127.1.119
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Juniper, B. E., Gorves, S., Landau-Schachar, B., and Audus, L. J. (1966). Root cap and the perception of gravity. Nature 209, 93–94. doi: 10.1038/209093a0
Kaiserli, E., Sullivan, S., Jones, M. A., Feeney, K. A., and Christie, J. M. (2009). Domain swapping to assess the mechanistic basis of Arabidopsis phototropin 1 receptor kinase activation and endocytosis by blue light. Plant Cell 21, 3226–3244. doi: 10.1105/tpc.109.067876
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kami, C., Allenbach, L., Zourelidou, M., Ljung, K., Schütz, F., Isono, E., et al. (2014). Reduced phototropism in pks mutants may be due to altered auxin-regulated gene expression or reduced lateral auxin transport. Plant J. 77, 393–403. doi: 10.1111/tpj.12395
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kami, C., Hersch, M., Trevisan, M., Genoud, T., Hiltbrunner, A., Bergmann, S., et al. (2012). Nuclear phytochrome A signaling promotes phototropism in Arabidopsis. Plant Cell 24, 566–576. doi: 10.1105/tpc.111.095083
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kami, C., Lorrain, S., Hornitschek, P., and Fankhauser, C. (2010). Light-regulated plant growth and development. Curr. Top. Dev. Biol. 91, 29–66. doi: 10.1016/S0070-2153(10)91002-8
Kato, T., Morita, M. T., Fukaki, H., Yamauchi, Y., Uehara, M., Niihama, M., et al. (2002). SGR2, a phospholipase-like protein, and ZIG/SGR4, a SNARE, are involved in the shoot gravitropism of Arabidopsis. Plant Cell 14, 33–46. doi: 10.1105/tpc.010215
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kato, T., Morita, M. T., and Tasaka, M. (2010). Defects in dynamics and functions of actin filament in Arabidopsis caused by the dominant-negative actin fiz1-induced fragmentation of actin filament. Plant Cell Physiol. 51, 333–338. doi: 10.1093/pcp/pcp189
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Khurana, J. P., and Poff, K. L. (1989). Mutants of Arabidopsis thaliana with altered phototropism. Planta 178, 400–406. doi: 10.1007/BF00391868
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kim, B. C., Tennessen, D. J., and Last, R. L. (1998). UV-B-induced photomorphogenesis in Arabidopsis thaliana. Plant J. 15, 667–674. doi: 10.1046/j.1365-313x.1998.00246.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kimura, M., and Kagawa, T. (2006). Phototropin and light-signaling in phototropism. Curr. Opin. Plant Biol. 9, 503–508. doi: 10.1016/j.pbi.2006.07.003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kiss, J. Z., Guisinger, M. M., Miller, A. J., and Stackhouse, K. S. (1997). Reduced gravitropism in hypocotyls of starch-deficient mutants of Arabidopsis. Plant Cell Physiol. 38, 518–525. doi: 10.1093/oxfordjournals.pcp.a029199
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kiss, J. Z., Hertel, R., and Sack, F. D. (1989). Amyloplasts are necessary for full gravitropic sensitivity in roots of Arabidopsis thaliana. Planta 177, 198–206. doi: 10.1007/BF00392808
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kleine-Vehn, J., Ding, Z., Jones, A. R., Tasaka, M., Morita, M. T., and Friml, J. (2010). Gravity-induced PIN transcytosis for polarization of auxin fluxes in gravity-sensing root cells. Proc. Natl. Acad. Sci. U.S.A. 107, 22344–22349. doi: 10.1073/pnas.1013145107
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Knieb, E., Salomon, M., and Rüdiger, W. (2004). Tissue-specific and subcellular localization of phototropin determined by immuno-blotting. Planta 218, 843–851. doi: 10.1007/s00425-003-1164-7
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Knight, T. (1806). On the direction of the radicle and germen during the vegetation of seeds. Philos. Trans. R. Soc. Lond. Biol. Sci. 99, 108–120. doi: 10.1098/rstl.1806.0006
Krinke, O., Ruelland, E., Valentová, O., Vergnolle, C., Renou, J.-P., Taconnat, L., et al. (2007). Phosphatidylinositol 4-kinase activation is an early response to salicylic acid in Arabidopsis suspension cells. Plant Physiol. 144, 1347–1359. doi: 10.1104/pp.107.100842
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lariguet, P., Schepens, I., Hodgson, D., Pedmale, U. V., Trevisan, M., Kami, C., et al. (2006). PHYTOCHROME KINASE SUBSTRATE 1 is a phototropin 1 binding protein required for phototropism. Proc. Natl. Acad. Sci. U.S.A. 103, 10134–10139. doi: 10.1073/pnas.0603799103
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Laskowski, M. J. (1990). Microtubule orientation in pea stem cells: a change in orientation follows the initiation of growth rate decline. Planta 181, 44–52. doi: 10.1007/BF00202323
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lawton, J. R., Juniper, B. E., and Hawes, C. R. (1986). Ultrastructural changes in statocytes in grass nodes after short periods of gravistimulation: do statoliths fall through the vacuole? Evidence from thick section high voltage EM. J. Exp. Bot. 37, 693–704. doi: 10.1093/jxb/37.5.693
Lee, J. S., and Evans, M. L. (1985). Polar transport of auxin across gravistimulated roots of maize and its enhancement by calcium. Plant Physiol. 77, 824–827. doi: 10.1104/pp.77.4.824
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lee, J. S., Mulkey, T. J., and Evans, M. L. (1983). Gravity-induced polar transport of calcium across root tips of maize. Plant Physiol. 73, 874–876. doi: 10.1104/pp.73.4.874
Legué, V., Blancaflor, E., Wymer, C., Perbal, G., Fantin, D., and Gilroy, S. (1997). Cytoplasmic free Ca2+ in Arabidopsis roots changes in response to touch but not gravity. Plant Physiol. 114, 789–800. doi: 10.1104/pp.114.3.789
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lehman, A., Black, R., and Ecker, J. R. (1996). HOOKLESS1, an ethylene response gene, is required for differential cell elongation in the Arabidopsis hypocotyl. Cell 85, 183–194. doi: 10.1016/S0092-8674(00)81095-8
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Leitz, G., Kang, B. -H., Schoenwaelder, M. E. A., and Staehelin, L. A. (2009). Statolith sedimentation kinetics and force transduction to the cortical endoplasmic reticulum in gravity-sensing Arabidopsis columella cells. Plant Cell 21, 843–860. doi: 10.1105/tpc.108.065052
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, H., Johnson, P., Stepanova, A., Alonso, J. M., and Ecker, J. R. (2004). Convergence of signaling pathways in the control of differential cell growth in Arabidopsis. Dev. Cell 7, 193–204. doi: 10.1016/j.devcel.2004.07.002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, J., Dai, X., and Zhao, Y. (2006). A role for auxin response factor 19 in auxin and ethylene signaling in Arabidopsis. Plant Physiol. 140, 899–908. doi: 10.1104/pp.105.070987
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, J., Yang, L., Jin, D., Nezames, C. D., Terzaghi, W., and Deng, X. W. (2013). UV-B-induced photomorphogenesis in Arabidopsis. Protein Cell 4, 485–492. doi: 10.1007/s13238-013-3036-7
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, Q.-H., and Yang, H.-Q. (2007). Cryptochrome signaling in plants. Photochem. Photobiol. 83, 94–101. doi: 10.1562/2006-02-28-IR-826
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lindeboom, J. J., Nakamura, M., Hibbel, A., Shundyak, K., Gutierrez, R., Ketelaar, T., et al. (2013). A mechanism for reorientation of cortical microtubule arrays driven by microtubule severing. Science 342, 1245533. doi: 10.1126/science.1245533
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Liscum, E., and Briggs, W. R. (1996). Mutations of Arabidopsis in potential transduction and response components of the phototropic signaling pathway. Plant Physiol. 112, 291–296. doi: 10.1104/pp.112.1.291
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Liscum, E., and Hangarter, R. P. (1993a). Light-stimulated apical hook opening in wild-type Arabidopsis thaliana seedlings. Plant Physiol. 101, 567–572.
Liscum, E., and Hangarter, R. P. (1993b). Genetic evidence that the red-absorbing form of phytochrome B modulates gravitropism in Arabidopsis thaliana. Plant Physiol. 103, 15–19.
Luschnig, C., Gaxiola, R. A., Grisafi, P., and Fink, G. R. (1998). EIR1, a root-specific protein involved in auxin transport, is required for gravitropism in Arabidopsis thaliana. Genes Dev. 12, 2175–2187. doi: 10.1101/gad.12.14.2175
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Macdonald, I. R., Gordon, D. C., Hart, J. W., and Maher, E. P. (1983). The positive hook: the role of gravity in the formation and opening of the apical hook. Planta 158, 76–81. doi: 10.1007/BF00395406
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Masson, P. H., Tasaka, M., Morita, M. T., Guan, C., Chen, R., and Boonsirichai, K. (2002). Arabidopsis thaliana: a model for the study of root and shoot gravitropism. Arabidopsis Book 1, e0043. doi: 10.1199/tab.0043
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Matsuda, S., Kajizuka, T., Kadota, A., Nishimura, T., and Koshiba, T. (2011). NPH3- and PGP-like genes are exclusively expressed in the apical tip region essential for blue-light perception and lateral auxin transport in maize coleoptiles. J. Exp. Bot. 62, 3459–3466. doi: 10.1093/jxb/err019
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
McClure, B. A., and Guilfoyle, T. (1989). Rapid redistribution of auxin-regulated RNAs during gravitropism. Science 243, 91–93. doi: 10.1126/science.11540631
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Millet, B., and Pickard, B. G. (1988). Early wrong-way response occurs in orthogravitropism of maize roots treated with lithium. Physiol. Plant. 72, 555–559. doi: 10.1111/j.1399-3054.1988.tb09165.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Miyamoto, K., Yamasaki, T., Uheda, E., and Ueda, J. (2014). Analysis of apical hook formation in Alaska pea with a 3-D clinostat and agravitropic mutant ageotropum. Front. Plant Sci. 5:137. doi: 10.3389/fpls.2014.00137
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Möller, B., Schenck, D., and Lüthen, H. (2010). Exploring the link between auxin receptors, rapid cell elongation and organ tropisms. Plant Signal. Behav. 5, 601–603. doi: 10.4161/psb.11501
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Monshausen, G. B., Miller, N. D., Murphy, A. S., and Gilroy, S. (2011). Dynamics of auxin-dependent Ca2+ and pH signaling in root growth revealed by integrating high-resolution imaging with automated computer vision-based analysis. Plant J. 65, 309–318. doi: 10.1111/j.1365-313X.2010.04423.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Morita, M. T. (2010). Directional gravity sensing in gravitropism. Annu. Rev. Plant Biol. 61, 705–720. doi: 10.1146/annurev.arplant.043008.092042
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Morita, M. T., Kato, T., Nagafusa, K., Saito, C., Ueda, T., Nakano, A., et al. (2002). Involvement of the vacuoles of the endodermis in the early process of shoot gravitropism in Arabidopsis. Plant Cell 14, 47–56. doi: 10.1105/tpc.010216
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Motchoulski, A., and Liscum, E. (1999). Arabidopsis NPH3: a NPH1 photoreceptor-interacting protein essential for phototropism. Science 286, 961–964. doi: 10.1126/science.286.5441.961
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mullen, J. L., Wolverton, C., Ishikawa, H., and Evans, M. L. (2000). Kinetics of constant gravitropic stimulus responses in Arabidopsis roots using a feedback system. Plant Physiol. 123, 665–670. doi: 10.1104/pp.123.2.665
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Müller, A., Guan, C., Gälweiler, L., Tänzler, P., Huijser, P., Marchant, A., et al. (1998). AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J. 17, 6903–6911. doi: 10.1093/emboj/17.23.6903
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Munnik, T., and Testerink, C. (2009). Plant phospholipid signaling: “in a nutshell.” J. Lipid Res. 50(Suppl.), S260–S265. doi: 10.1194/jlr.R800098-JLR200
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nagashima, A., Suzuki, G., Uehara, Y., Saji, K., Furukawa, T., Koshiba, T., et al. (2008). Phytochromes and cryptochromes regulate the differential growth of Arabidopsis hypocotyls in both a PGP19-dependent and a PGP19-independent manner. Plant J. 53, 516–529. doi: 10.1111/j.1365-313X.2007.03358.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Nakamura, M., Toyota, M., Tasaka, M., and Morita, M. T. (2011). An Arabidopsis E3 ligase, SHOOT GRAVITROPISM9, modulates the interaction between statoliths and F-actin in gravity sensing. Plant Cell 23, 1830–1848. doi: 10.1105/tpc.110.079442
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Němec, B. (1900). Über die art der wahrnehmung des schwerkraftes bei den pflanzen. Ber. Dtsch. Bot. Ges. 18, 241–245.
Nick, P., Bergfeld, R., Schafer, E., and Schopfer, P. (1990). Unilateral reorientation of microtubules at the outer epidermal wall during photo- and gravitropic curvature of maize coleoptiles and sunflower hypocotyls. Planta 181, 162–168. doi: 10.1007/BF02411533
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ohgishi, M., Saji, K., Okada, K., and Sakai, T. (2004). Functional analysis of each blue light receptor, cry1, cry2, phot1, and phot2, by using combinatorial multiple mutants in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 101, 2223–2228. doi: 10.1073/pnas.0305984101
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Okushima, Y., Overvoorde, P. J., Arima, K., Alonso, J. M., Chan, A., Chang, C., et al. (2005). Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in Arabidopsis thaliana: unique and overlapping functions of ARF7 and ARF19. Plant Cell 17, 444–463. doi: 10.1105/tpc.104.028316
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ottenschläger, I., Wolff, P., Wolverton, C., Bhalerao, R. P., Sandberg, G., Ishikawa, H., et al. (2003). Gravity-regulated differential auxin transport from columella to lateral root cap cells. Proc. Natl. Acad. Sci. U.S.A. 100, 2987–2991. doi: 10.1073/pnas.0437936100
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Paciorek, T., Zazímalová, E., Ruthardt, N., Petrásek, J., Stierhof, Y.-D., Kleine-Vehn, J., et al. (2005). Auxin inhibits endocytosis and promotes its own efflux from cells. Nature 435, 1251–1256. doi: 10.1038/nature03633
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Palmieri, M., and Kiss, J. Z. (2005). Disruption of the F-actin cytoskeleton limits statolith movement in Arabidopsis hypocotyls. J. Exp. Bot. 56, 2539–2550. doi: 10.1093/jxb/eri248
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Perbal, G., and Driss-Ecole, D. (2003). Mechanotransduction in gravisensing cells. Trends Plant Sci. 8, 498–504. doi: 10.1016/j.tplants.2003.09.005
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Perera, I. Y., Heilmann, I., and Boss, W. F. (1999). Transient and sustained increases in inositol 1,4,5-trisphosphate precede the differential growth response in gravistimulated maize pulvini. Proc. Natl. Acad. Sci. U.S.A. 96, 5838–5843. doi: 10.1073/pnas.96.10.5838
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Perera, I. Y., Heilmann, I., Chang, S. C., Boss, W. F., and Kaufman, P. B. (2001). A role for inositol 1,4,5-trisphosphate in gravitropic signaling and the retention of cold-perceived gravistimulation of oat shoot pulvini. Plant Physiol. 125, 1499–1507. doi: 10.1104/pp.125.3.1499
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Perera, I. Y., Hung, C.-Y., Brady, S., Muday, G. K., and Boss, W. F. (2006). A universal role for inositol 1,4,5-trisphosphate-mediated signaling in plant gravitropism. Plant Physiol. 140, 746–760. doi: 10.1104/pp.105.075119
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Peters, W. S., and Tomos, A. D. (1996). The history of tissue tension. Ann. Bot. 77, 657–665. doi: 10.1093/aob/77.6.657
Petrášek, J., and Friml, J. (2009). Auxin transport routes in plant. Development 136, 2675–2688. doi: 10.1242/dev.030353
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Plieth, C., and Trewavas, A. J. (2002). Reorientation of seedlings in the earth’s gravitational field induces cytosolic calcium transients. Plant Physiol. 129, 786–796. doi: 10.1104/pp.011007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Pozhvanov, G. A., Suslov, D. V., and Medvedev, S. S. (2013). Actin cytoskeleton rearrangements during the gravitropic response of Arabidopsis roots. Tsitologiia 55, 28–35. doi: 10.1134/S1990519X13020120
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Preuten, T.,Hohm, T., Bergmann, S., and Fankhauser, C. (2013). Defining the site of light perception and initiation of phototropism in Arabidopsis. Curr. Biol. 23, 1934–1938. doi: 10.1016/j.cub.2013.07.079
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Quail, P. H., Boylan, M. T., Parks, B. M., Short, T. W., Xu, Y., and Wagner, D. (1995). Phytochromes: photosensory perception and signal transduction. Science 268, 675–680. doi: 10.1126/science.7732376
Raboy, V. (2003). myo-Inositol-1,2,3,4,5,6-hexakisphosphate. Phytochemistry 64, 1033–1043. doi: 10.1016/S0031-9422(03)00446-1
Rakusová, H., Gallego-Bartolomé, J., Vanstraelen, M., Robert, H. S., Alabadí, D., Blázquez, M. A., et al. (2011). Polarization of PIN3-dependent auxin transport for hypocotyl gravitropic response in Arabidopsis thaliana. Plant J. 67, 817–826. doi: 10.1111/j.1365-313X.2011.04636.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Raz, V., and Ecker, J. R. (1999). Regulation of differential growth in the apical hook of Arabidopsis. Development 126, 3661–3668.
Raz, V., and Koornneef, M. (2001). Cell division activity during apical hook development. Plant Physiol. 125, 219–226. doi: 10.1104/pp.125.1.219
Reed, J. W., Nagatani, A., Elich, T. D., Fagan, M., and Chory, J. (1994). Phytochrome A and phytochrome B have overlapping but distinct functions in Arabidopsis development. Plant Physiol. 104, 1139–1149.
Rizzini, L., Favory, J.-J., Cloix, C., Faggionato, D., O’Hara, A., Kaiserli, E., et al. (2011). Perception of UV-B by the Arabidopsis UVR8 protein. Science 332, 103–106. doi: 10.1126/science.1200660
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Robert, S., Kleine-Vehn, J., Barbez, E., Sauer, M., Paciorek, T., Baster, P., et al. (2010). ABP1 mediates auxin inhibition of clathrin-dependent endocytosis in Arabidopsis. Cell 143, 111–121. doi: 10.1016/j.cell.2010.09.027
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Roberts, D., Pedmale, U. V., Morrow, J., Sachdev, S., Lechner, E., Tang, X., et al. (2011). Modulation of phototropic responsiveness in Arabidopsis through ubiquitination of phototropin 1 by the CUL3-Ring E3 ubiquitin ligase CRL3(NPH3). Plant Cell 23, 3627–3640. doi: 10.1105/tpc.111.087999
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rockwell, N. C., Su, Y.-S., and Lagarias, J. C. (2006). Phytochrome structure and signaling mechanisms. Annu. Rev. Plant Biol. 57, 837–858. doi: 10.1146/annurev.arplant.56.032604.144208
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ruegger, M., Dewey, E., Hobbie, L., Brown, D., Bernasconi, P., Turner, J., et al. (1997). Reduced naphthylphthalamic acid binding in the tir3 mutant of Arabidopsis is associated with a reduction in polar auxin transport and diverse morphological defects. Plant Cell 9, 745–757. doi: 10.1105/tpc.9.5.745
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sack, F. D., Suyemoto, M. M., and Leopold, A. C. (1986). Amyloplast sedimentation and organelle saltation in living corn columella cells. Am. J. Bot. 73, 1692–1698. doi: 10.2307/2444235
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Saito, C., Morita, M. T., Kato, T., and Tasaka, M. (2005). Amyloplasts and vacuolar membrane dynamics in the living graviperceptive cell of the Arabidopsis inflorescence stem. Plant Cell 17, 548–558. doi: 10.1105/tpc.104.026138
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sakai, T., Kagawa, T., Kasahara, M., Swartz, T. E., Christie, J. M., Briggs, W. R., et al. (2001). Arabidopsis nph1 and npl1: blue light receptors that mediate both phototropism and chloroplast relocation. Proc. Natl. Acad. Sci. U.S.A. 98, 6969–6974. doi: 10.1073/pnas.101137598
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Salinas-Mondragon, R. E., Kajla, J. D., Perera, I. Y., Brown, C. S., and Sederoff, H. W. (2010). Role of inositol 1,4,5-triphosphate signalling in gravitropic and phototropic gene expression. Plant Cell Environ. 33, 2041–2055. doi: 10.1111/j.1365-3040.2010.02204.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Salomon, M., Zacherl, M., and Rudiger, W. (1997). Asymmetric, blue light-dependent phosphorylation of a 116-kilodalton plasma membrane protein can be correlated with the first- and second-positive phototropic curvature of oat coleoptiles. Plant Physiol. 115, 485–491.
Sauer, M., Balla, J., Luschnig, C., Wisniewska, J., Reinöhl, V., Friml, J., et al. (2006). Canalization of auxin flow by Aux/IAA-ARF-dependent feedback regulation of PIN polarity. Genes Dev. 20, 2902–2911. doi: 10.1101/gad.390806
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schepens, I., Boccalandro, H. E., Kami, C., Casal, J. J., and Fankhauser, C. (2008). PHYTOCHROME KINASE SUBSTRATE4 modulates phytochrome-mediated control of hypocotyl growth orientation. Plant Physiol. 147, 661–671. doi: 10.1104/pp.108.118166
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schopfer, P. (2006). Biomechanics of plant growth. Am. J. Bot. 93, 1415–1425. doi: 10.3732/ajb.93.10.1415
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Scott, A. C., and Allen, N. S. (1999). Changes in cytosolic pH within Arabidopsis root columella cells play a key role in the early signaling pathway for root gravitropism. Plant Physiol. 121, 1291–1298. doi: 10.1104/pp.121.4.1291
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sedbrook, J. C., Chen, R., and Masson, P. H. (1999). ARG1 (altered response to gravity) encodes a DnaJ-like protein that potentially interacts with the cytoskeleton. Proc. Natl. Acad. Sci. U.S.A. 96, 1140–1145. doi: 10.1073/pnas.96.3.1140
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Shichijo, C., and Hashimoto, T. (2013). “Light-induced exaggeration of the hypocotyl hook: its developmental basis and significance,” in From Seed Germination to Young Plants, ed. C. A. Busso (New York, NY: Nova Sci. Pub. Inc.), 39–59.
Silady, R. A., Ehrhardt, D. W., Jackson, K., Faulkner, C., Oparka, K., and Somerville, C. R. (2008). The GRV2/RME-8 protein of Arabidopsis functions in the late endocytic pathway and is required for vacuolar membrane flow. Plant J. 53, 29–41. doi: 10.1111/j.1365-313X.2007.03314.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Silk, W. K., and Erickson, R. O. (1978). Kinematics of hypocotyl curvature. Am. J. Bot. 65, 310–319. doi: 10.2307/2442271
Sinclair, W., and Trewavas, A. J. (1997). Calcium in gravitropism. A re-examination. Planta 203, S85–S90. doi: 10.1007/PL00008120
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Smet, D., Žádníková, P., Vandenbussche, F., Benková, E., and Van Der Straeten, D. (2014). Dynamic infrared imaging analysis of apical hook development in Arabidopsis: the case of brassinosteroids. New Phytol. 202, 1398–1411. doi: 10.1111/nph.12751
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Smith, C. M., Desai, M., Land, E. S., and Perera, I. Y. (2013). A role for lipid-mediated signaling in plant gravitropism. Am. J. Bot. 100, 153–160. doi: 10.3732/ajb.1200355
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Spartz, A. K., Lee, S. H., Wenger, J. P., Gonzalez, N., Itoh, H., Inzé, D., et al. (2012). The SAUR19 subfamily of SMALL AUXIN UP RNA genes promote cell expansion. Plant J. 70, 978–990. doi: 10.1111/j.1365-313X.2012.04946.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Spartz, A. K., Ren, H., Park, M. Y., Grandt, K. N., Lee, S. H., Murphy, A. S., et al. (2014). SAUR inhibition of PP2C-D phosphatases activates plasma membrane H+-ATPases to promote cell expansion in Arabidopsis. Plant Cell 26, 2129–2142. doi: 10.1105/tpc.114.126037
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Stowe-Evans, E. L., Luesse, D. R., and Liscum, E. (2001). The enhancement of phototropin-induced phototropic curvature in Arabidopsis occurs via a photoreversible phytochrome A-dependent modulation of auxin responsiveness. Plant Physiol. 126, 826–834. doi: 10.1104/pp.126.2.826
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sukumar, P., Edwards, K. S., Rahman, A., Delong, A., and Muday, G. K. (2009). PINOID kinase regulates root gravitropism through modulation of PIN2-dependent basipetal auxin transport in Arabidopsis. Plant Physiol. 150, 722–735. doi: 10.1104/pp.108.131607
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sullivan, S., Thomson, C. E., Kaiserli, E., and Christie, J. M. (2009). Interaction specificity of Arabidopsis 14-3-3 proteins with phototropin receptor kinases. FEBS Lett. 583, 2187–2193. doi: 10.1016/j.febslet.2009.06.011
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sun, J., Qi, L., Li, Y., Zhai, Q., and Li, C. (2013). PIF4 and PIF5 transcription factors link blue light and auxin to regulate the phototropic response in Arabidopsis. Plant Cell 25, 2102–2114. doi: 10.1105/tpc.113.112417
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Swarup, R., Kramer, E. M., Perry, P., Knox, K., Leyser, H. M. O., Haseloff, J., et al. (2005). Root gravitropism requires lateral root cap and epidermal cells for transport and response to a mobile auxin signal. Nat. Cell Biol. 7, 1057–1065. doi: 10.1038/ncb1316
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tatematsu, K., Kumagai, S., Muto, H., Sato, A., Watahiki, M. K., Harper, R. M., et al. (2004). MASSUGU2 Encodes Aux/IAA19, an auxin-regulated protein that functions together with the transcriptional activator NPH4/ARF7 to regulate differential growth responses of hypocotyl and formation of lateral roots in Arabidopsis thaliana. Plant Cell 16, 379–393. doi: 10.1105/tpc.018630
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tepperman, J. M., Zhu, T., Chang, H. S., Wang, X., and Quail, P. H. (2001). Multiple transcription-factor genes are early targets of phytochrome A signaling. Proc. Natl. Acad. Sci. U.S.A. 98, 9437–9442. doi: 10.1073/pnas.161300998
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Testerink, C., and Munnik, T. (2011). Molecular, cellular, and physiological responses to phosphatidic acid formation in plants. J. Exp. Bot. 62, 2349–2361. doi: 10.1093/jxb/err079
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tilbrook, K., Arongaus, A. B., Binkert, M., Heijde, M., Yin, R., and Ulm, R. (2013). The UVR8 UV-B photoreceptor: perception, signaling and response. Arabidopsis Book 11, e0164. doi: 10.1199/tab.0164
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tominaga, M., Yokota, E., Vidali, L., Sonobe, S., Hepler, P. K., and Shimmen, T. (2000). The role of plant villin in the organization of the actin cytoskeleton, cytoplasmic streaming and the architecture of the transvacuolar strand in root hair cells of Hydrocharis. Planta 210, 836–843. doi: 10.1007/s004250050687
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Toyota, M., Furuichi, T., Sokabe, M., and Tatsumi, H. (2013). Analyses of a gravistimulation-specific Ca2+ signature in Arabidopsis using parabolic flights. Plant Physiol. 163, 543–554. doi: 10.1104/pp.113.223313
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Toyota, M., Furuichi, T., Tatsumi, H., and Sokabe, M. (2008). Cytoplasmic calcium increases in response to changes in the gravity vector in hypocotyls and petioles of Arabidopsis seedlings. Plant Physiol. 146, 505–514. doi: 10.1104/pp.107.106450
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Traas, J. A., Doonan, J. H., Rawlins, D. J., Shaw, P. J., Watts, J., and Lloyd, C. W. (1987). An actin network is present in the cytoplasm throughout the cell cycle of carrot cells and associates with the dividing nucleus. J. Cell Biol. 105, 387–395. doi: 10.1083/jcb.105.1.387
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tromas, A., Paque, S., Stierlé, V., Quettier, A.-L., Muller, P., Lechner, E., et al. (2013). Auxin-binding protein 1 is a negative regulator of the SCF(TIR1/AFB) pathway. Nat. Commun. 4, 2496. doi: 10.1038/ncomms3496
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tsugeki, R., and Fedoroff, N. V. (1999). Genetic ablation of root cap cells in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 96, 12941–12946. doi: 10.1073/pnas.96.22.12941
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vandenbussche, F., Petrásek, J., Žádníková, P., Hoyerová, K., Pesek, B., Raz, V., et al. (2010). The auxin influx carriers AUX1 and LAX3 are involved in auxin-ethylene interactions during apical hook development in Arabidopsis thaliana seedlings. Development 137, 597–606. doi: 10.1242/dev.040790
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vieten, A., Vanneste, S., Wisniewska, J., Benková, E., Benjamins, R., Beeckman, T., et al. (2005). Functional redundancy of PIN proteins is accompanied by auxin-dependent cross-regulation of PIN expression. Development 132, 4521–4531. doi: 10.1242/dev.02027
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Vitha, S., Yang, M., Sack, F. D., and Kiss, J. Z. (2007). Gravitropism in the starch excess mutant of Arabidopsis thaliana. Am. J. Bot. 94, 590–598. doi: 10.3732/ajb.94.4.590
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wan, Y., Jasik, J., Wang, L., Hao, H., Volkmann, D., Menzel, D., et al. (2012). The signal transducer NPH3 integrates the phototropin1 photosensor with PIN2-based polar auxin transport in Arabidopsis root phototropism. Plant Cell 24, 551–565. doi: 10.1105/tpc.111.094284
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wang, Y., Lin, W.-H., Chen, X., and Xue, H.-W. (2009). The role of Arabidopsis 5PTase13 in root gravitropism through modulation of vesicle trafficking. Cell Res. 19, 1191–1204. doi: 10.1038/cr.2009.105
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Watahiki, M. K., and Yamamoto, K. T. (1997). The massugu1 mutation of Arabidopsis identified with failure of auxin-induced growth curvature of hypocotyl confers auxin insensitivity to hypocotyl and leaf. Plant Physiol. 115, 419–426. doi: 10.1104/pp.115.2.419
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Weise, S. E., and Kiss, J. Z. (1999). Gravitropism of inflorescence stems in starch-deficient mutants of Arabidopsis. Int. J. Plant Sci. 160, 521–527. doi: 10.1086/314142
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wheeler, G. L., and Brownlee, C. (2008). Ca2+ signalling in plants and green algae–changing channels. Trends Plant Sci. 13, 506–514. doi: 10.1016/j.tplants.2008.06.004
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Willige, B. C., Ahlers, S., Zourelidou, M., Barbosa, I. C. R., Demarsy, E., Trevisan, M., et al. (2013). D6PK AGCVIII kinases are required for auxin transport and phototropic hypocotyl bending in Arabidopsis. Plant Cell 25, 1674–1688. doi: 10.1105/tpc.113.111484
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Willige, B. C., Ogiso-Tanaka, E., Zourelidou, M., and Schwechheimer, C. (2012). WAG2 represses apical hook opening downstream from gibberellin and PHYTOCHROME INTERACTING FACTOR 5. Development 139, 4020–4028. doi: 10.1242/dev.081240
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wilmoth, J. C., Wang, S., Tiwari, S. B., Joshi, A. D., Hagen, G., Guilfoyle, T. J., et al. (2005). NPH4/ARF7 and ARF19 promote leaf expansion and auxin-induced lateral root formation. Plant J. 43, 118–130. doi: 10.1111/j.1365-313X.2005.02432.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wolverton, C., Mullen, J. L., Ishikawa, H., and Evans, M. L. (2002). Root gravitropism in response to a signal originating outside of the cap. Planta 215, 153–157. doi: 10.1007/s00425-001-0726-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Wu, G., Cameron, J. N., Ljung, K., and Spalding, E. P. (2010). A role for ABCB19-mediated polar auxin transport in seedling photomorphogenesis mediated by cryptochrome 1 and phytochrome B. Plant J. 62, 179–191. doi: 10.1111/j.1365-313X.2010.04137.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Xu, T., Wen, M., Nagawa, S., Fu, Y., Chen, J.-G., Wu, M.-J., et al. (2010). Cell surface- and rho GTPase-based auxin signaling controls cellular interdigitation in Arabidopsis. Cell 143, 99–110. doi: 10.1016/j.cell.2010.09.003
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yamamoto, K., and Kiss, J. Z. (2002). Disruption of the actin cytoskeleton results in the promotion of gravitropism in inflorescence stems and hypocotyls of Arabidopsis. Plant Physiol. 128, 669–681. doi: 10.1104/pp.010804
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yamauchi, Y., Fukaki, H., Fujisawa, H., and Tasaka, M. (1997). Mutations in the SGR4, SGR5 and SGR6 loci of Arabidopsis thaliana alter the shoot gravitropism. Plant Cell Physiol. 38, 530–535. doi: 10.1093/oxfordjournals.pcp.a029201
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yano, D., Sato, M., Saito, C., Sato, M. H., Morita, M. T., and Tasaka, M. (2003). A SNARE complex containing SGR3/AtVAM3 and ZIG/VTI11 in gravity-sensing cells is important for Arabidopsis shoot gravitropism. Proc. Natl. Acad. Sci. U.S.A. 100, 8589–8594. doi: 10.1073/pnas.1430749100
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Yoder, T. L., Zheng, H. Q., Todd, P., and Staehelin, L. A. (2001). Amyloplast sedimentation dynamics in maize columella cells support a new model for the gravity-sensing apparatus of roots. Plant Physiol. 125, 1045–1060. doi: 10.1104/pp.125.2.1045
Žádníková, P., Petrášek, J., Marhavȳ, P., Raz, V., Vandenbussche, F., Ding, Z., et al. (2010). Role of PIN-mediated auxin efflux in apical hook development of Arabidopsis thaliana. Development 137, 607–617. doi: 10.1093/jxb/5.3.357
Zandomeni, K., and Schopfer, P. (1994). Mechanosensory microtubule reorientation in the epidermis of maize coleoptiles subjected to bending stress. Protoplasma 182, 96–101. doi: 10.1242/dev.041277
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhang, J., Vanneste, S., Brewer, P. B., Michniewicz, M., Grones, P., Kleine-Vehn, J., et al. (2011). Inositol trisphosphate-induced Ca2+ signaling modulates auxin transport and PIN polarity. Dev. Cell 20, 855–866. doi: 10.1007/BF01403471
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhang, K.-X., Xu, H.-H., Gong, W., Jin, Y., Shi, Y.-Y., Yuan, T.-T., et al. (2014). Proper PIN1 distribution is needed for root negative phototropism in Arabidopsis. PLoS ONE 9:e85720. doi: 10.1371/journal.pone.0085720
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhang, K.-X., Xu, H.-H., Yuan, T.-T., Zhang, L., and Lu, Y.-T. (2013). Blue-light-induced PIN3 polarization for root negative phototropic response in Arabidopsis. Plant J. 76, 308–321. doi: 10.1111/tpj.12298
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhao, X., Wang, Y.-L., Qiao, X.-R., Wang, J., Wang, L.-D., Xu, C.-S., et al. (2013). Phototropins function in high-intensity blue light-induced hypocotyl phototropism in Arabidopsis by altering cytosolic calcium. Plant Physiol. 162, 1539–1551.
Zhao, Y., Christensen, S. K., Fankhauser, C., Cashman, J. R., Cohen, J. D., Weigel, D., et al. (2001). A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 291, 306–309. doi: 10.1104/pp.113.216556
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zourelidou, M., Absmanner, B., Weller, B., Barbosa, I. C. R., Willige, B. C., Fastner, A., et al. (2014). Auxin efflux by PIN-FORMED proteins is activated by two different protein kinases, D6 PROTEIN KINASE and PINOID. elife 3:e02860. doi: 10.1126/science.291.5502.306
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: differential growth, auxin, apical hook, phototropism, gravitropism, hormonal crosstalk
Citation: Žádníková P, Smet D, Zhu Q, Van Der Straeten D and Benková E (2015) Strategies of seedlings to overcome their sessile nature: auxin in mobility control. Front. Plant Sci. 6:218. doi: 10.3389/fpls.2015.00218
Received: 09 October 2014; Accepted: 19 March 2015;
Published online: 14 April 2015.
Edited by:
Tohru Hashimoto, Uozaki Life Science Laboratory, JapanReviewed by:
Günther F. E. Scherer, Leibniz Universität Hannover, GermanyBjörn C. Willige, Salk Institute, USA
Copyright © 2015 Žádníková, Smet, Zhu, Van Der Straeten and Benková. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Dominique Van Der Straeten, Department of Physiology, Laboratory of Functional Plant Biology, Ghent University, K.L. Ledeganckstraat 35, B-9000 Ghent, BelgiumZG9taW5pcXVlLnZhbmRlcnN0cmFldGVuQHVnZW50LmJl; Eva Benková, Institute of Science and Technology Austria (IST Austria), Am Campus 1, 3400 Klosterneuburg, AustriaZXZhLmJlbmtvdmFAaXN0LmFjLmF0
†Present address: Petra Žádníková, Institute of Developmental Genetics, Cluster of Excellence on Plant Sciences, Heinrich Heine University, Düsseldorf, Germany
‡These authors have contributed equally to this work.