 Robert Backer1,2
Robert Backer1,2 Waheed Mahomed1,2Bianca J. Reeksting1,2
Waheed Mahomed1,2Bianca J. Reeksting1,2 Juanita Engelbrecht1,2
Juanita Engelbrecht1,2 Enrique Ibarra-Laclette3,4
Enrique Ibarra-Laclette3,4 Noëlani van den Berg1,2*
Noëlani van den Berg1,2*- 1Forestry and Agricultural Biotechnology Institute, University of Pretoria, Pretoria, South Africa
- 2Department of Genetics, Fruit Tree Biotechnology Program, University of Pretoria, Pretoria, South Africa
- 3Laboratorio Nacional de Genómica para la Biodiversidad-Langebio/Unidad de Genómica Avanzada, Centro de Investigación y Estudios Avanzados del – Instituto Politécnico Nacional, Irapuato, México
- 4Red de Estudios Moleculares Avanzados, Instituto de Ecología A.C., Xalapa, México
The NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 (NPR1) forms an integral part of the salicylic acid (SA) pathway in plants and is involved in cross-talk between the SA and jasmonic acid/ethylene (JA/ET) pathways. Therefore, NPR1 is essential to the effective response of plants to pathogens. Avocado (Persea americana) is a commercially important crop worldwide. Significant losses in production result from Phytophthora root rot, caused by the hemibiotroph, Phytophthora cinnamomi. This oomycete infects the feeder roots of avocado trees leading to an overall decline in health and eventual death. The interaction between avocado and P. cinnamomi is poorly understood and as such limited control strategies exist. Thus uncovering the role of NPR1 in avocado could provide novel insights into the avocado – P. cinnamomi interaction. A total of five NPR1-like sequences were identified. These sequences were annotated using FGENESH and a maximum-likelihood tree was constructed using 34 NPR1-like protein sequences from other plant species. The conserved protein domains and functional motifs of these sequences were predicted. Reverse transcription quantitative PCR was used to analyze the expression of the five NPR1-like sequences in the roots of avocado after treatment with salicylic and jasmonic acid, P. cinnamomi infection, across different tissues and in P. cinnamomi infected tolerant and susceptible rootstocks. Of the five NPR1-like sequences three have strong support for a defensive role while two are most likely involved in development. Significant differences in the expression profiles of these five NPR1-like genes were observed, assisting in functional classification. Understanding the interaction of avocado and P. cinnamomi is essential to developing new control strategies. This work enables further classification of these genes by means of functional annotation and is a crucial step in understanding the role of NPR1 during P. cinnamomi infection.
Introduction
Plants recognize and react to external threats much like any other living organism, eliciting a response to combat disease (Robert-Seilaniantz et al., 2011). Defense responses against biotrophic and hemibiotrophic pathogens are mainly dependent on the salicylic acid (SA) pathway (Glazebrook, 2005). Plants challenged by a biotrophic pathogen show a substantial increase in endogenous SA, a subsequent hypersensitive response (HR) at the site of infection and the onset of systemic acquired resistance (SAR; Malamy et al., 1990; Metraux et al., 1991; Rasmussen et al., 1991; Gaffney et al., 1993; Delaney et al., 1994). SAR is an important part of plant defense, providing long term, broad spectrum resistance which is effective against a wide variety of fungal, viral and bacterial pathogens at tissues distal to the initial site of infection (Frederich et al., 1996; Sticher et al., 1997; An and Mou, 2011; Robert-Seilaniantz et al., 2011). Increases in SA concentration have been conclusively linked to the establishment of SAR, for instance, exogenous application of SA or one of its biologically active analogs, 2,6-dichloroisonicotinic acid (INA) and benzo (1,2,3) thiadiazole-7-carbothioic acid S-methyl ester (BTH), is able to induce SAR (White, 1979; Ward et al., 1991; Lawton et al., 1995). Conversely, plants that express the transgene nahG which encodes for a salicylate hydroxylase, lack functionally active SA and are SAR compromised (Gaffney et al., 1993; Bi et al., 1995; Friedrich et al., 1995; Lawton et al., 1995).
The quest to discover the SA receptor led to the discovery of NONEXPRESSOR OF PATHOGENESIS-RELATED GENES1 (NPR1), a transcription co-factor protein encoded for by NPR1 (Cao et al., 1994). The majority of described NPR1 proteins contain ankyrin repeat and Broad Complex, Tramtrack and Bric a brac/Pox virus and Zinc finger (BTB/POZ) domains (Cao et al., 1997; Hepworth et al., 2005; Spoel et al., 2009). These domains are essential for protein–protein interactions and enable NPR1 to function as a co-activator (Cao et al., 1997; Rochon et al., 2006). In Arabidopsis NPR1 is found as an oligomer within the cytoplasm of uninduced cells and changes in SA concentration lead to an altered redox environment within the cell, supporting the nuclear localization of NPR1 in its monomeric form (Mou et al., 2003). It is worth noting that NPR1 is constitutively localized within the nucleus of several plant species, yet the perception of a SA signal is still required for the expression of pathogenesis-related (PR) genes (Kinkema et al., 2000; Le Henanff et al., 2009; Maier et al., 2011).
Multiple NPR1-like proteins seem to be present in most, if not all, plant species. Phylogenetic analysis of this family suggests the existence of three functionally distinct clades (Hepworth et al., 2005; Zhang et al., 2006; Peraza-Echeverria et al., 2012). Members of the first clade, AtNPR1 and AtNPR2, are mostly associated with positive SAR regulation (Cao et al., 1997, 1998). The second clade, AtNPR3, and AtNPR4, is associated with negative SAR regulation, yet is required for effective SAR induction (Liu et al., 2005; Zhang et al., 2006). The third clade, AtBOP1, and AtBOP2, is associated with the development of lateral organs (Hepworth et al., 2005). Phylogenetic analysis has since included NPR1-like proteins from multiple plant species (Le Henanff et al., 2009; Peraza-Echeverria et al., 2012; Shao et al., 2013), and although phylogenetic analysis alone is insufficient for functional annotation it may provide a basis for understanding functional variation (Liu et al., 2005; Zhang et al., 2006).
The most extensively studied member of the Arabidopsis NPR1-like family is AtNPR1. Mutants of this protein are more susceptible to virulent pathogens and display compromised expression of PR genes when compared to plants expressing wild-type NPR1 (Glazebrook et al., 1996; Cao et al., 1997). Complementation of these npr1 mutants using wild-type NPR1 restores the expression of PR genes as well as pathogen resistance and the induction of SAR (Cao et al., 1997). Various plants overexpressing NPR1 show increased PR gene expression and pathogen resistance (Cao et al., 1998; Friedrich et al., 2001; Malnoy et al., 2007; Le Henanff et al., 2011; Chen et al., 2012; Kumar et al., 2012). Overexpressing OsNPR1, the ortholog of AtNPR1 in rice, results in an increased resistance to bacterial blight, yet these transgenic plants show an increased susceptibility to herbivores (Yuan et al., 2007). Interestingly, herbivore hypersensitivity is alleviated when NPR1 is constitutively localized to the nucleus (Yuan et al., 2007). These results suggest that NPR1 is involved in the antagonistic cross-talk between the SA and jasmonic acid/ethylene (JA/ET) pathways, a theory supported by several other studies (Spoel et al., 2003; El Oirdi et al., 2011). Thus, NPR1 is considered the master regulator of defense responses in plants.
Additionally, NPR1 interacts with several members of the TGACG motif-binding factor (TGA) family of basic leucine zipper protein (bZIP) transcriptions factors (Zhang et al., 1999; Despres et al., 2000; Zhou et al., 2000). These transcription factors associate with the as-1-like (TGACG) promoter element within PR gene promoters and are responsible, at least in part, for their expression (Fan and Dong, 2002; Zhang et al., 2003). Moreover, the DNA binding affinity of TGA factors is increased when associated with NPR1 (Despres et al., 2000; Fan and Dong, 2002) and NPR1 may also deactivate the repression of PR genes by certain TGA factors (Rochon et al., 2006). This interaction describes the basis of NPR1-dependant gene expression, yet the realistically more complex mechanism involves several other factors (Li et al., 1999; Weigel et al., 2001; Chern et al., 2005; Zhang et al., 2006; Zwicker et al., 2007).
Evidence suggests that NPR3 and NPR4 are essential to establishing SAR even though they suppress NPR1-dependant gene expression (Liu et al., 2005; Zhang et al., 2006; Fu et al., 2012). Both NPR3 and NPR4 act as adaptors for Cullin 3 (CUL3) E3 ligase-facilitated ubiquitinylation and subsequent proteasome degradation of NPR1 (Fu et al., 2012). The degradation of NPR1 serves dual roles; turn-over of NPR1 as well as suppression of NPR1-dependant gene expression in SA naïve cells and cells undergoing HR (Spoel et al., 2009). The expression of NPR1-dependant genes during non-stress conditions and HR increases fitness costs and prevents the establishment of HR, respectively (Rate and Greenberg, 2001; Heidel et al., 2004). In SA naïve cells NPR4 strongly interacts with NPR1 thus preventing increases in expression of PR genes (Fu et al., 2012). Moreover, increased SA concentrations interrupt this interaction and increase the affinity of NPR3 for NPR1 (Fu et al., 2012). At the site of HR where SA concentrations are the highest NPR1 is rapidly degraded, while at distal tissues with intermediate SA concentrations NPR3 merely facilitates sufficient turn-over of NPR1 (Fu et al., 2012). The turn-over of transcription factors ensures optimal expression of target genes (Salghetti et al., 2000; Collins and Tansey, 2006), as seen for NPR1. Thus by responding to the concentration of SA, NPR3, and NPR4 prevent the untimely expression of PR genes, fine-tuning the defense response. It is therefore clear that understanding the role of the NPR1-like family is an important part of understanding defense responses in plants.
Avocado is an economically important fruit crop with an annual worldwide gross production value of US $ 3835 million1. The fruit are highly nutritious and contain high levels of monounsaturated fats making them popular for use in a wide variety of culinary products. The greatest threat to production is Phytophthora root rot (PRR), caused by the hemibiotrophic oomycete pathogen Phytophthora cinnamomi Rands (Hardham, 2005). Infection by P. cinnamomi results in decreased water and nutrient absorption due to necrosis of the avocado feeder roots, leading to a decline in tree health and eventual death ensuing economic losses (Zentmyer, 1984; Coffey, 1987). With a wide host range of more than 3000 plant species and the ability to persist in the environment (Weste, 1983; Hardham, 2005), effective control of P. cinnamomi is limited.
The use of phosphite trunk injections, tolerant rootstocks (e.g., Dusa®) and organic mulching practices are currently utilized by industry as methods for controlling PRR (Coffey, 1987; Giblin et al., 2005). Phosphite trunk injections have been a dependable method for over 30 years (Darvas et al., 1984; Pegg et al., 1985; Coffey, 1987; Kaiser et al., 1997; Giblin et al., 2005), yet evidence suggests that P. cinnamomi has the potential to develop decreased sensitivity against this fungicide (Duvenhage, 1994; Dobrowolski et al., 2008). Similar observations occur for metalaxyl, another decidedly effective fungicide (Darvas et al., 1984). Moreover, the lengthy selection process for PRR tolerant rootstocks (Gabor and Coffey, 1991; Menge, 1999; Kremer-Köhne and Mukhumo, 2003) limits the introduction of novel tolerant rootstocks, possibly providing the pathogen with an opportunity to overcome host tolerance.
Although biochemical and histological studies have provided some insight into the avocado – P. cinnamomi interaction (Phillips et al., 1987; Botha and Kotze, 1989; Bekker et al., 2006; Sánchez-Pérez et al., 2009; García-Pineda et al., 2010), research on the molecular characteristics of this interaction have only recently gained attention (Mahomed and van den Berg, 2011; Reeksting et al., 2014). Our current understanding of the incompatible Arabidopsis thaliana – P. cinnamomi interaction provides limited information on compatible interactions. For example, in Arabidopsis the JA/ET pathway seems to be essential to P. cinnamomi resistance (Rookes et al., 2008), yet in avocado SA inhibits growth and colonization (García-Pineda et al., 2010). Such conspicuous differences between non-host and host interactions highlight the need to elucidate the host specific interaction between avocado and P. cinnamomi on a molecular level.
Thus defining the role of NPR1 in avocado could potentially provide novel insights into the avocado – P. cinnamomi interaction. This is the first study aimed at discovering and characterizing NPR1-like genes in Persea americana. We have discovered five NPR1-like genes from P. americana which harbor the ankyrin repeat and BTB/POZ domains and show sequence similarity to other known NPR1-like genes. Phylogenetic analysis reveals that the predicted protein sequences of these genes can be resolved into the three known phylogenetic clades of the NPR1-like family. We describe the expression of these genes in Dusa®, a PRR tolerant avocado rootstock, across five time points during treatment with SA, MeJA, and P. cinnamomi using RT–qPCR. Additionally, we measured the basal expression levels for each transcript across six different tissues. The findings of this study provide an invaluable resource for further study and functional characterization of the NPR1-like family in avocado. Future efforts could focus on intracellular interactions and localization as well as overexpression of defense related PaNPR1-like genes in wild-type and npr1 mutant Arabidopsis.
Materials and Methods
Sequence Annotation and Phylogenetic Analysis
Five NPR1-like gene sequences were obtained from the P. americana genome (Unpublished data). Sequences were arbitrarily assigned identifiers as follows: PaNPR1, PaNPR2, PaNPR3, PaNPR4, and PaNPR5. Open reading frames (ORF’s) were annotated using the online prediction tool FGENESH with the Vitis vinifera genome-specific parameters selected (Solovyev et al., 2006). Exon/intron positions of predicted CDSs were visualized using GSDS software v2.0 (Guo et al., 2007). Percentage amino acid similarity was calculated using SIAS2. Protein domains were predicted using PROSITE (Sigrist et al., 2010). Sequences were submitted to GenBank3: PaNPR1 [GenBank: KR056089], PaNPR2 [GenBank: KR056090], PaNPR3 [GenBank: KR056091], PaNPR4 [GenBank: KR056092], and PaNPR5 [GenBank: KR056093].
Phylogenetic Analysis
Additional NPR1-like protein sequences from other plant species were attained online at NCBI4 in order to perform alignments (Table 1). Sequences were aligned using Clustal W software v2.1 (Thompson et al., 1994). The best substitution model for the alignment was determined and subsequently used in construction of a maximum likelihood (ML) phylogenetic tree using the tools available in MEGA software v5.2 (Tamura et al., 2011). The initial tree was constructed using the neighbor-joining (NJ) method (Saitou and Nei, 1987) and bootstrapping (1000 replicates) was used to determine confidence.

TABLE 1. Additional NPR1-like protein sequences from other plant species.
Plant Material
One-year-old clonal PRR-tolerant Dusa® rootstock plantlets were provided by Westfalia Technological Services (Tzaneen, South Africa). Two phytohormone treatment groups were assigned and treated with 70 ml sodium salicylate (NaSA) solution (5 mM NaSA (Sigma–Aldrich, St. Louis, MO, USA), 0.1% Tween® 20 (v/v) (Sigma–Aldrich, St. Louis, MO, USA) or 70 ml methyl jasmonate (MeJA) solution (5 mM MeJA (Sigma–Aldrich, St. Louis, MO, USA), 0.1% ethanol (99.9%), 0.1% Tween® 20 (v/v)). A third treatment group was inoculated with 20 ml P. cinnamomi zoospore suspension (3 × 105 spores/ml) and 70 ml P. cinnamomi mycelial suspension. Each treatment contained three biological replicates with two plants per replicate. Control plants were either treated with 70 ml NaSA control solution (0.1% Tween®) or 70 ml MeJA control solution [0.1% Tween®, 0.1% ethanol (99.9%)]. Each control group contained three biological replicates with one plant per replicate. Plants were randomly assigned to either the treatment or control groups. All treatments and controls were applied directly to the soil at the base of the plant. Roots were harvested for all treatment and control groups at 6, 12, 18, 24, and 96 h. Samples were snap frozen in liquid nitrogen and stored at -80°C. Biological replicates were homogenized using the IKA® Tube Mill control (IKA®, Staufen, Germany) until a fine consistency was attained.
Mature grafted trees located at Westfalia (Tzaneen, South Africa) were used for the collection of tissue samples. Six tissue types were selected: feeder roots, mature green stems, mature green leaves, unripe fruit as well as stems and leaves from flush growth (young material). Samples were taken from a single orchard block which contained clonal Hass fruitstocks grafted onto clonal PRR-tolerant Duke 7 rootstocks. Fifteen trees were randomly selected from which two samples of each tissue were taken for each individual tree. Samples were snap frozen in liquid nitrogen and stored at -80°C. Tissue samples were randomly allocated to three groups of five trees, individual tissue samples from each group were then pooled and homogenized using the IKA® Tube Mill control (IKA®).
Phytophthora cinnamomi Infection
Zoospores were produced by placing P. cinnamomi colonized blocks of V8 agar (20% V8 juice (v/v), 0.25% CaCO3, agar 17g.l-1) into 90 mm Petri dishes containing 2% V8 broth for 3 days to allow sufficient mycelial growth. Cultures were then rinsed three times with dH2O and run-off stored for use as mycelial suspension. Filtered stream water was then added and cultures left under UV light for 2–3 days until sufficient sporangia formation was observed. Cultures were then cold-shocked at 4°C for 45 min and placed at room temperature for 1 h to allow zoospore release. Sufficient zoospore release and motility was monitored via microscopy. Inoculation was carried out immediately by pouring both the zoospore and mycelial suspension directly onto the soil at the base of the plants.
Nested PCR
Total genomic DNA was isolated from inoculated root samples. Nested LPV3 PCR was then performed in order to confirm successful infection of plant roots by P. cinnamomi as described by Engelbrecht et al. (2013). Results were visualized on 2% TAE agarose gel under non-denaturing conditions.
RNA Extraction
Total RNA was extracted from homogenized plant material using a modified version of the CTAB extraction method described by Chang et al. (1993). The chloroform: isoamyl alcohol step was repeated four to six times until the volume of the interphase diminished and the supernatant was clear. Samples were resuspended in diethylpyrocarbonate (DEPC) treated water containing 30 U/ml RiboLock RNase Inhibitor (Thermo Fisher Scientific Inc., Leicestershire, UK). RNA concentration and purity was assessed using the NanoDrop® ND-1000 spectrophotometer (Nanodrop Technologies Inc., Montchanin, DE, USA). RNA integrity was assessed on 2% TAE agarose gel under non-denaturing conditions.
Total RNA from P. cinnamomi infected tolerant (Dusa®) and susceptible (R0.12) avocado rootstocks at 0 h (uninfected control), 6, 12, and 24 h were obtained from Engelbrecht et al. (2013). RNA concentration and purity was assessed using the NanoDrop® ND-1000 spectrophotometer (Nanodrop Technologies, Inc., Montchanin, DE, USA). RNA integrity was assessed on 2% TAE agarose gel under non-denaturing conditions.
cDNA Synthesis
RNA was purified of any contaminants using the RNeasy MinElute Cleanup Kit (Qiagen Inc., Hilden, Germany) according to the manufacturer’s instructions followed by an on-column RNase-free DNase I (Thermo Fischer Scientific) treatment. cDNA was synthesized using the ImProm-IITM single strand cDNA synthesis kit (Promega Corporation, Madison, WI, USA) according to the manufacturer’s guidelines. First strand synthesis was primed using 0.5 μg random hexamers (Thermo Fisher Scientific). cDNA concentration and purity was assessed using the NanoDrop® ND-1000 spectrophotometer (Nanodrop Technologies). Genomic DNA (gDNA) contamination was assessed using the intron-spanning flavone-3-hydroxylase (F3H) primers as described in (Reeksting et al., 2014).
RT-qPCR
Primers for reverse transcription quantitative PCR were designed using CLC Genomics Workbench v5.1 (CLC Bio, Qiagen® Inc., Hilden, Germany) and quality assessed on NetPrimer (Premier Biosoft International, Palo Alto, CA, USA). Primers with annealing temperatures between 55 and 60°C, expected amplicons lengths of <150 bp and quality scores >95.0 (NetPrimer) were synthesized (Inqaba Biotec, Pretoria, South Africa; Table 2). Primer specificity was confirmed by performing conventional PCR and sequencing (African Center for Gene Technologies, Pretoria, South Africa) and by the presence of a single melting curve. A 1:3 serial dilution was derived from a comprehensive mix of treated and control cDNA samples. Calibration curves were then performed for each candidate and reference gene across multiple temperatures to ensure that efficiency (E) and correlation (R2) values were in accordance with MIQE guidelines (Bustin et al., 2009; Table 3). All reactions were performed using SensiMixTM SYBR® No-ROX kit (Bioline USA, Inc., Taunton, MA, USA) according to manufacturer’s instructions with a final reaction volume of 10 μl. Thermal cycling was performed on the Bio-Rad® CFX 96 instrument (Bio Rad laboratories., Hercules, CA, USA). Thermal cycling conditions were: 10 min at 95°C followed by 40 cycles of 10 s at 95°C, 15 s at Ta (Table 3) and 10 s at 72°C, a melting dissociation curve was constructed from 60 to 95°C at 0.5°C increments following the final cycle. Three endogenous control genes were used in order to normalize the data for each gene, namely Actin, 18S, and alpha-1 tubulin as reported by Reeksting et al. (2014). The stability of the reference genes were analyzed using Bio-Rad® CFX Manager software v1.5 (Bio Rad laboratories; Table 3). Normalized relative quantities (fold change) for genes were calculated using the method described by Pfaffl (2001). Expression values for each time-point in a specific treatment were calibrated against a control of the corresponding time-point. Cleanup of RT–qPCR products was performed using ZymocleanTM Gel DNA recovery kit (Zymo Research Corporation, Irvin, CA, USA) in preparation for sequencing. Sequencing reactions were performed using BigDye® Terminator v3.1 Cycle Sequencing Kit (Life Technologies, Thermo Fisher Scientific) according to the manufacturer’s guidelines. Reactions were precipitated using 3 M sodium acetate (pH 5.2) and submitted for sequencing (African Center for Gene Technologies, Pretoria, South Africa).

TABLE 2. Primer sequences used in reverse transcription quantitative PCR analysis.
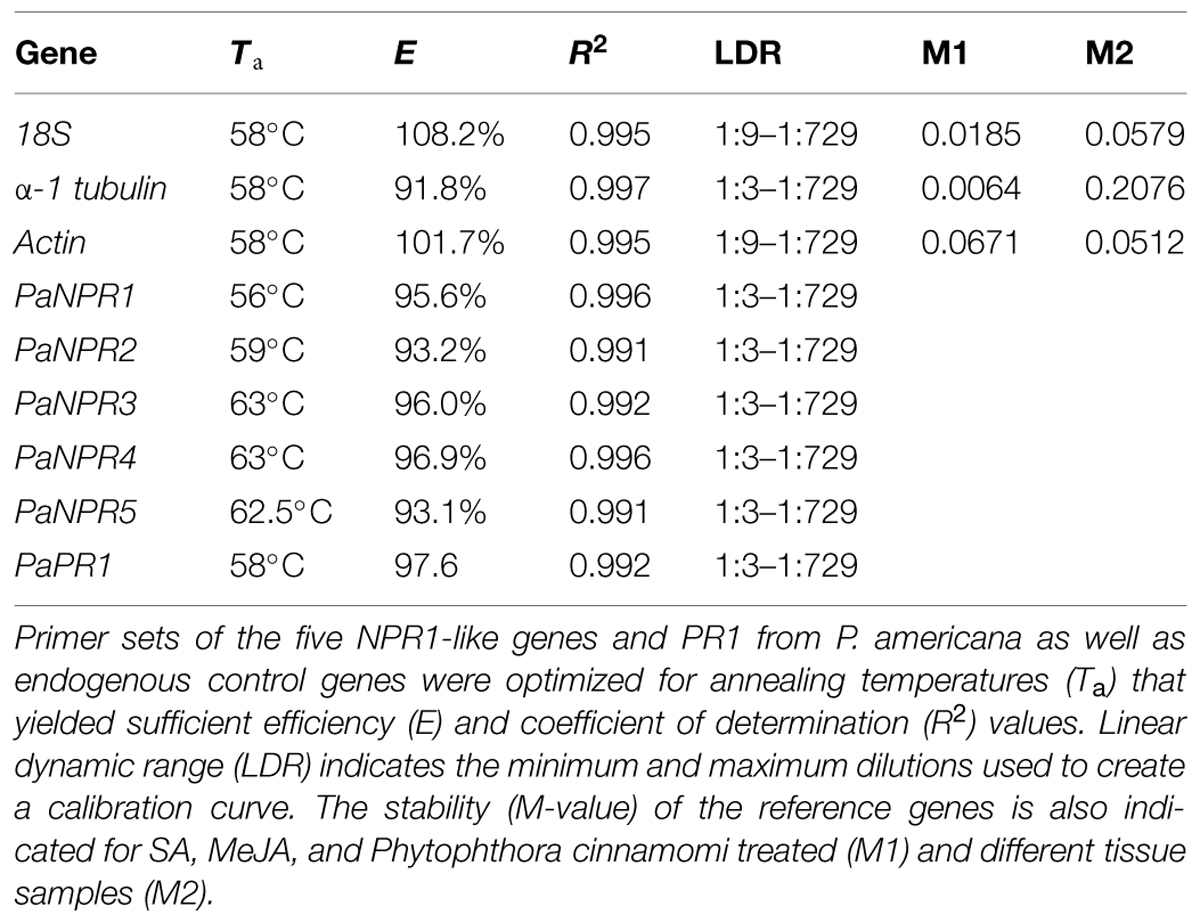
TABLE 3. Reverse transcription quantitative PCR optimization.
Statistical Analysis
A student’s t-test was performed to determine significance for quantitative gene expression analysis. SA, MeJA, and P. cinnamomi treated samples were compared to respective controls at each time point. Statistical analysis was performed using GraphPad Prism software v6.0.5 (GraphPad Software, Inc., La Jolla, CA, USA). Significance was assessed using a 95% confidence interval. Statistical analysis for various tissue samples was done using JMP 11 (SAS Institute Inc., Cary, NC, USA). Initial analysis was performed using one-way ANOVA followed by Tukey’s HSD test, adhoc. Significance was assessed using a 95% confidence interval. A Mann–Whitney unpaired t-test was performed to determine significance for quantitative gene expression analysis. Expression in tolerant plants was compared to that of susceptible plants at each respective time point. Statistical analysis was performed with the Statistics Online Computational Resource package5 Significance was assessed using a 95% confidence interval.
Results
In Silico Identification and Analysis
A total of five NPR1-like sequences were obtained from the unpublished P. americana genome. PaNPR1, PaNPR2, PaNPR3, PaNPR4, and PaNPR5, code for putative proteins of 601 aa, 590 aa, 476 aa, 642 aa, and 496 aa, respectively. Amino acid analysis revealed that PaNPR1 and PaNPR2 are most similar to AtNPR1 (52.46% identity, 64.08% similarity, and 55.94% identity, 68.35% similarity, respectively) while PaNPR4 is most similar to AtNPR3 (56.74% identity, 66.08% similarity). Furthermore, PaNPR3 and PaNPR5 are highly similar to AtBOP2 (78.90% identity, 83.71% similarity, and 81.57% and identity, 85.18% similarity, respectively). The predicted exon/intron structure further illustrates similarities between the avocado and Arabidopsis NPR1-like gene families (Figure 1A).
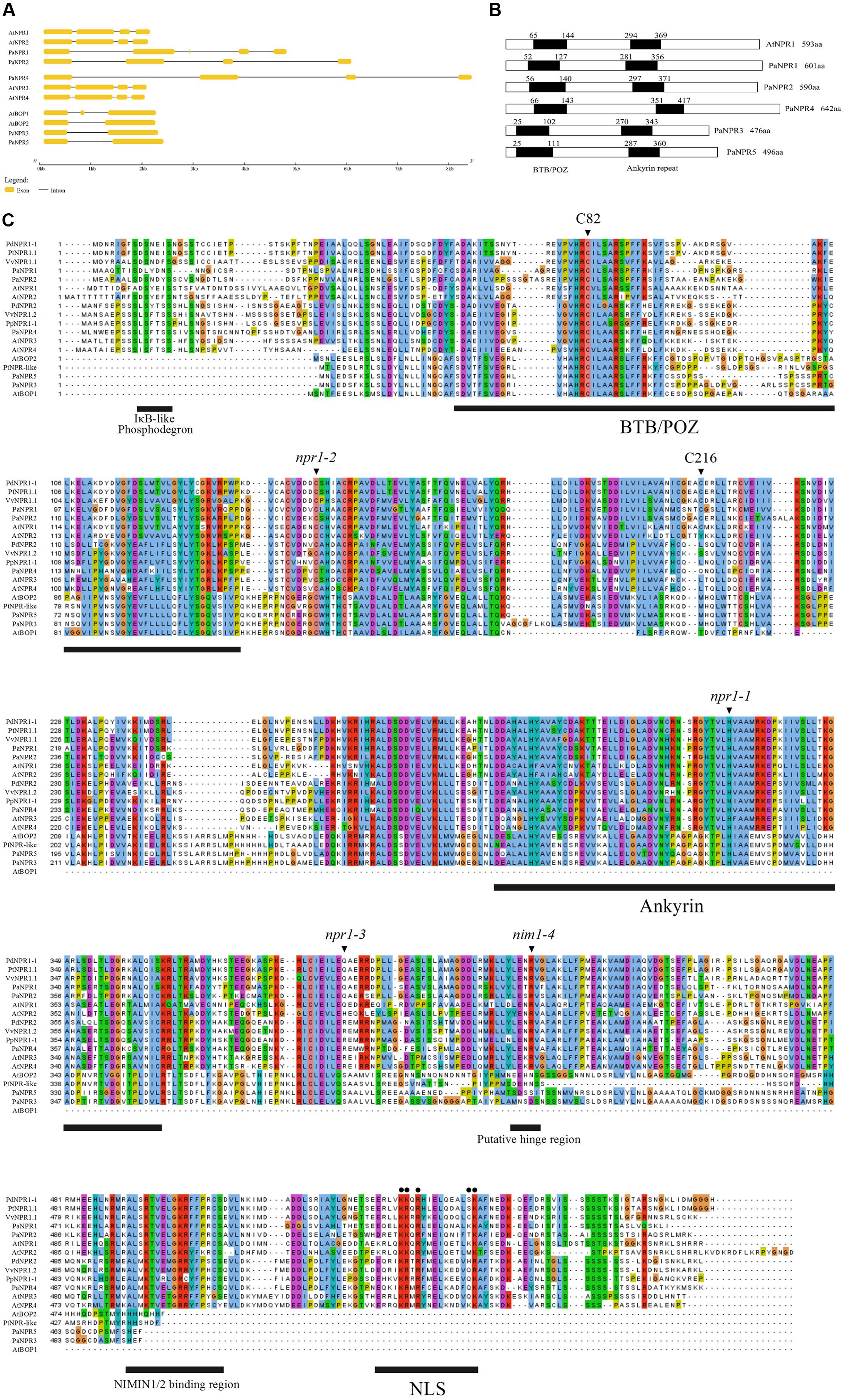
FIGURE 1. Exon/intron boundary and predicted coding sequence comparison of PaNPR1-like genes with other known NPR1-like sequences. (A) The predicted exon/intron structure of the NPR1-like family from Arabidopsis thaliana and Persea americana. Exons are denoted by yellow boxes while introns are represented by thin black lines. (B) A comparison of the positions of the BTB/POZ and ankyrin repeat domains between the PaNPR1-like and AtNPR1-like family of proteins. (C) A multiple alignment of PaNPR1-like proteins and several other known NPR1-like proteins from woody plants and Arabidopsis. The positions of amino acid changes causing the npr1-1(H), npr1-2 (C), npr1-3 (∗), and nim1-4 (R) mutants as well as the positions of the highly conserved cysteine residues at position 82 and 216 in Arabidopsis are indicated by black triangles above the alignment. The BTB/POZ and ankyrin repeat domains are indicated by black bars below the alignment. Several important motifs such as the IκB phosphodegron, LENRV hinge region, NIMIN1/2 binding site, and NLS1, are also indicated by black bars. The positions of important amino acids in the NLS1 of AtNPR1 are indicated by black dots above the alignment.
Analysis using PROSITE (Sigrist et al., 2010) reveals that all five PaNPR1-like proteins contain the BTB/POZ and ankyrin repeat domains at similar positions to AtNPR1 (Figure 1B). Conversely, only PaNPR1, PaNPR2, and PaNPR4 contain an NPR1-like C-terminal region which has been shown to be an essential component of NPR1 (Cao et al., 1997). The C-terminus contains the nuclear localization signal (NLS), a conserved penta-amino acid motif (LENRV) and a NIM INTERACTING (NIMIN) 1/2 protein binding site (Kinkema et al., 2000; Maier et al., 2011). PaNPR1 contains all five of the conserved basic amino acids that constitute the NLS1, PaNPR2 contains four of the five and PaNPR4 contains several conservative amino acid substitutions (Figure 1C). Similarly, the NIMIN1/2 binding region is completely conserved in PaNPR1 with one and three substitutions in PaNPR2 and PaNPR4, respectively, (Figure 1C). Furthermore, PaNPR4 contains the putative hinge region (LENRV motif) while PaNPR1 and PaNPR2, respectively, contain a conservative threonine and serine substitution at the third position of this motif (Figure 1C). The N-terminal of PaNPR2 contains an IκB-like phosphodegron motif (DSxxxS) which has been shown to be necessary for proteasome-mediated turnover of NPR1 (Spoel et al., 2009) while PaNPR1 contains a similar motif with a serine to lysine substitution at the second position (Figure 1C).
The PaNPR1-like protein sequences obtained in this study were subjected to phylogenetic analysis together with 34 full-length NPR1-like protein sequences from vascular and non-vascular plant species (Figure 2). This analysis reveals that PaNPR1 and PaNPR2 form a distinct group which is closely related to VvNPR1.1 from grapevine (70.83 and 76.33% similarity, respectively) as well as NPR1 from poplar (PtNPR1.1 and PdNPR1-1) and beet (BvNPR1), clustering within the clade containing AtNPR1 and AtNPR2 (Figure 2). On the other hand, PaNPR4 clusters within the clade containing AtNPR3 and AtNPR4 (Figure 2). Finally, PaNPR3 and PaNPR5 form a distinct group and are closely related to AtBOP2 from Arabidopsis (76.02 and 77.90% similarity, respectively; Figure 2).
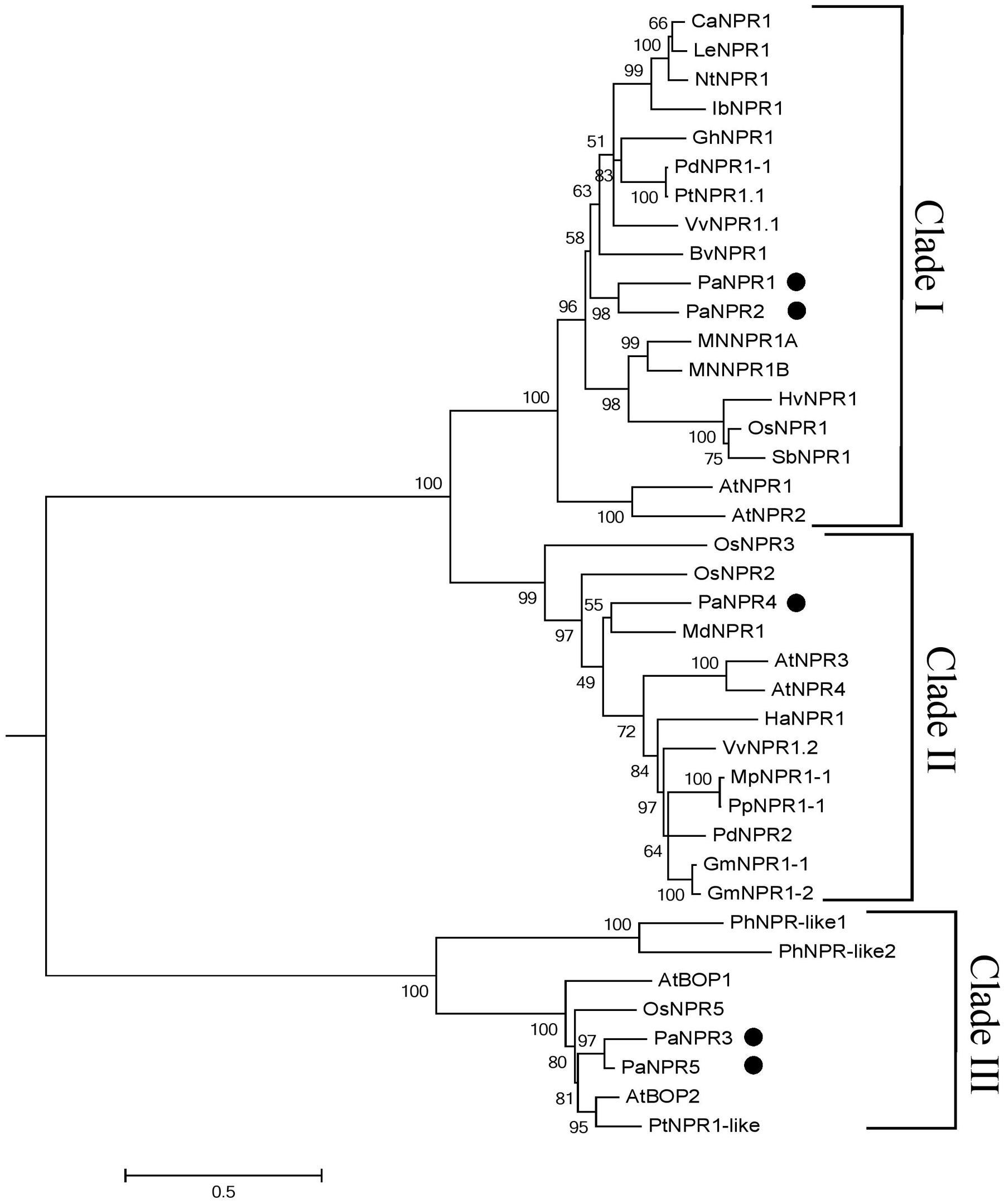
FIGURE 2. Phylogenetic analysis of the NPR1-like family of proteins from P. americana and several other species. A phylogenetic tree of five NPR1-like proteins from P. americana as well as NPR1-like proteins from other vascular and non-vascular plant species, including the NPR1-like family from Arabidopsis. The tree was generated in MEGA software v5.2 using the maximum likelihood (ML) method. A total of 1000 bootstrap replicates were performed and values are indicated above the branch points. The species of origin, identifiers and accession numbers are summarized in Table 1.
PaNPR1-like and PaPR1 Response to SA, MeJA, and P. cinnamomi
In order to evaluate all five PaNPR1-like genes, it is important to gage their expression in response to hormone treatment and pathogen challenge. It is well-known that SA application increases expression of AtNPR1 approximately twofold within 24 h, similar results are also obtained when Arabidopsis is inoculated with Hyaloperonospora parasitica (Ryals et al., 1997). Similarly, MeJA application has been shown to result in increased expression of NPR1 in rice and banana, although to a lesser extent than treatment with SA (Yuan et al., 2007; Endah et al., 2008). Thus, in order to investigate the response of all five PaNPR1-like genes, 1 year-old clonal PRR-tolerant Dusa® rootstock plantlets were treated with either SA, MeJA, or inoculated with P. cinnamomi and harvested at 6, 12, 18, 24, and 96 h. Furthermore, an ortholog of Arabidopsis PR1 from P. americana, PaPR1, was used as a SAR marker (Reeksting et al., 2014).
The expression of PaNPR1 was significantly down-regulated during SA treatment at 12 h (0.56-fold), returning to basal levels at 96 h (Figure 3A). Treatment with MeJA also decreased the expression of PaNPR1 but at a later time point, 24 h (0.61-fold), yet expression remained low at 96 h (Figure 3A). Infection with P. cinnamomi decreased the expression of PaNPR1 at 12 h (0.68-fold) followed by an increase at later time points, similar to treatment with SA, yet differences between treated and control samples were not significant (Figure 3A). However, significant down-regulation was seen at 96 h P. cinnamomi infection (0.55-fold), similar to treatment with MeJA (Figure 3A).
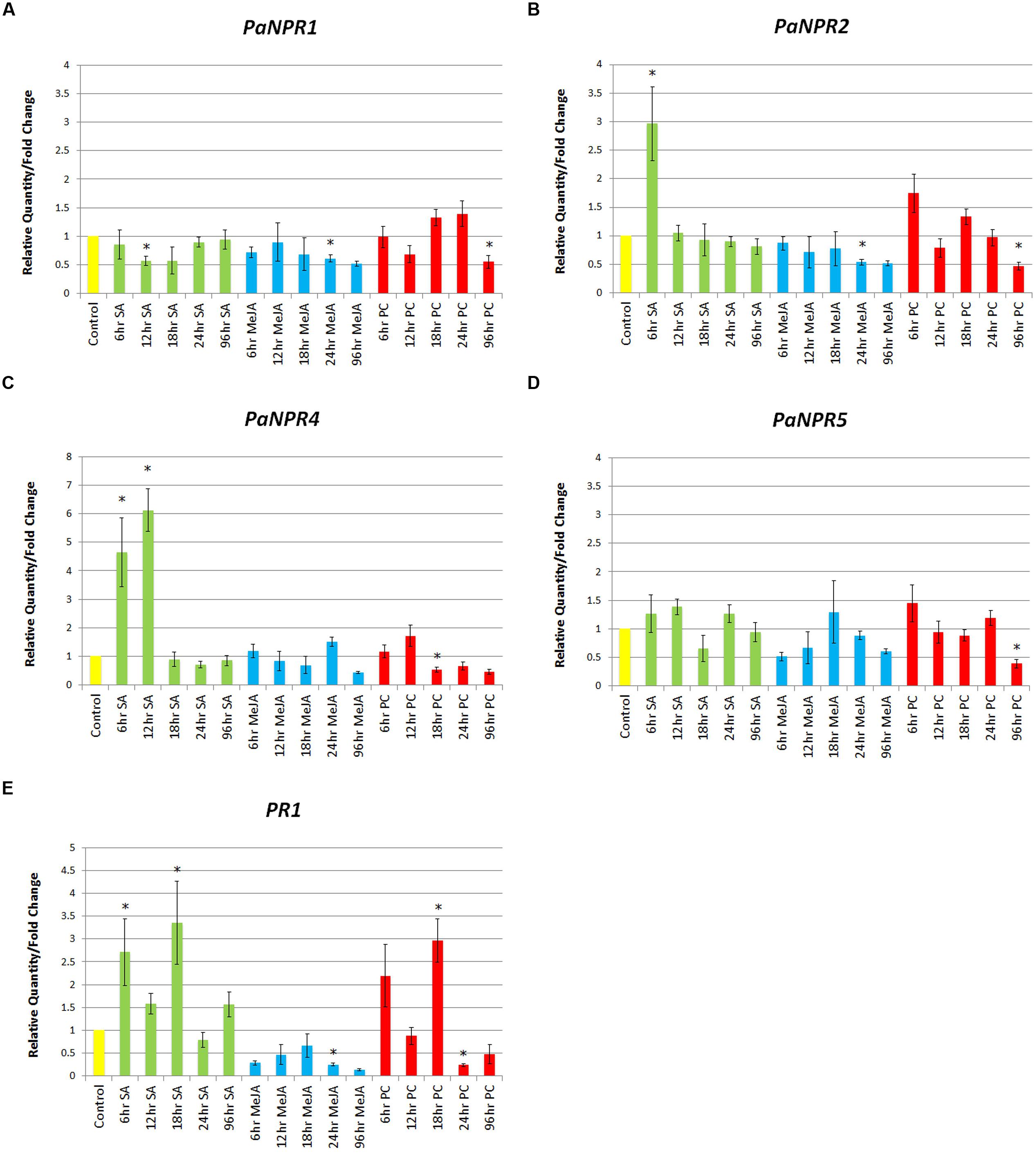
FIGURE 3. Expression of PaNPR1-like genes as well as PaPR1 in response to SA, JA, and Phytophthora cinnamomi treatment. Normalized relative quantities (fold change) for (A) PaNPR1, (B) PaNPR2, (C) PaNPR4, (D) PaNPR5, and (E) PaPR1 were calculated using the method described by Pfaffl (2001). The response is indicated by vertical bars across five time points for SA (green), MeJA (blue), and P. cinnamomi (red) treated samples, and is labeled on the horizontal axis. The samples were compared to untreated samples harvested at each time point, a representative bar for the controls (yellow) is also indicated. SE for each bar is shown. Significant differences (P < 0.05) between control and treated samples is denoted by an asterisk (∗) above the bar.
Treatment with SA significantly up-regulated PaNPR2 expression at 6 h (2.97-fold) when compared to control samples, returning to baseline levels at 12 h (Figure 3B). Plants treated with MeJA showed a progressive decline in the expression of PaNPR2 with a significant down-regulation at 24 h (0.53-fold) that remained low at 96 h (Figure 3B). Similarly, PaNPR2 was significantly down-regulated at 96 h after infection with P. cinnamomi (0.47-fold; Figure 3B). Unfortunately the presence of PaNPR3 could not be reliably detected in either treatment due to low transcript abundance and was therefore omitted from this part of the study.
Significant up-regulation of PaNPR4 was observed at 6 h (4.63-fold) and 12 h (6.13-fold) with a sharp drop to baseline levels at 18 h (Figure 3C). Treatment with MeJA yielded no significant changes in the expression of PaNPR4, yet expression seemed to be slightly lower than that of the controls, especially at 96 h (0.44-fold; Figure 3C). A significant down-regulation of PaNPR4 was observed at 18 h P. cinnamomi infection (0.53-fold) that remained low until the 96 h time point (Figure 3C). Conversely, PaNPR5 was not significantly altered by any of the hormone treatments, yet was significantly down-regulated by P. cinnamomi at 96 h (0.39-fold; Figure 3D).
Lastly, expression of PaPR1 was significantly up-regulated at 6 h (2.71-fold) and 18 h (3.35-fold), with an unexpected slump at 12 h (1.58-fold; Figure 3E). This decrease in expression coincided with the highest expression of PaNPR4 at 12 h and was relieved at 18 h when PaNPR4 expression returned to basal levels. The expression of PaPR1 was significantly down-regulated by MeJA at 24 h (0.25-fold) and remained low at 96 h (Figure 3E). A significant increase in PaPR1 expression was seen at 18 h (2.97-fold) followed by a significant decrease at 24 h (0.24-fold) following P. cinnamomi inoculation (Figure 3E). It was interesting to note that following inoculation with P. cinnamomi, the expression of PaPR1 is similar to that of SA at early time-points and MeJA at later time-points (Figure 3E).
PaNPR1-Like Expression in Various Avocado Tissues
AtBOP1 and AtBOP2 have been shown to be involved in the growth and development of lateral organs and to accumulate extensively in the proximal parts of these tissues (Hepworth et al., 2005). Thus determining whether any of the PaNPR1-like genes are overrepresented in specific tissues could assist in identifying possible AtNPR1 orthologs by eliminating possible ortholog of the Arabidopsis BOP genes in avocado. Consequently, feeder roots, mature stems, mature leaves, unripe fruit as well as stems and leaves from flush growth of mature avocado trees were sampled and basal expression of all five PaNPR1-like genes was determined.
The expression of PaNPR1, PaNPR2, and PaNPR4 was constitutive in all tissues yet higher basal levels were seen in more mature tissues than in younger tissues. The expression of PaNPR1 was highest in the roots and mature leaves, with the significantly less transcript detected in the young leaves (0.14-fold), young stems (0.11-fold) and unripe fruit (0.28-fold; Figure 4A). Similarly, PaNPR2 had significantly higher expression in roots and mature leaves when compared to young leaves (0.27-fold), young stems (0.24-fold) and unripe fruit (0.22-fold; Figure 4B). Expression of PaNPR4 was significantly higher in mature leaves (2.48-fold) and mature stems (2.61-fold), relative to the young leaves (0.74-fold), young stems (0.70-fold), and unripe fruit (0.44-fold; Figure 4D).
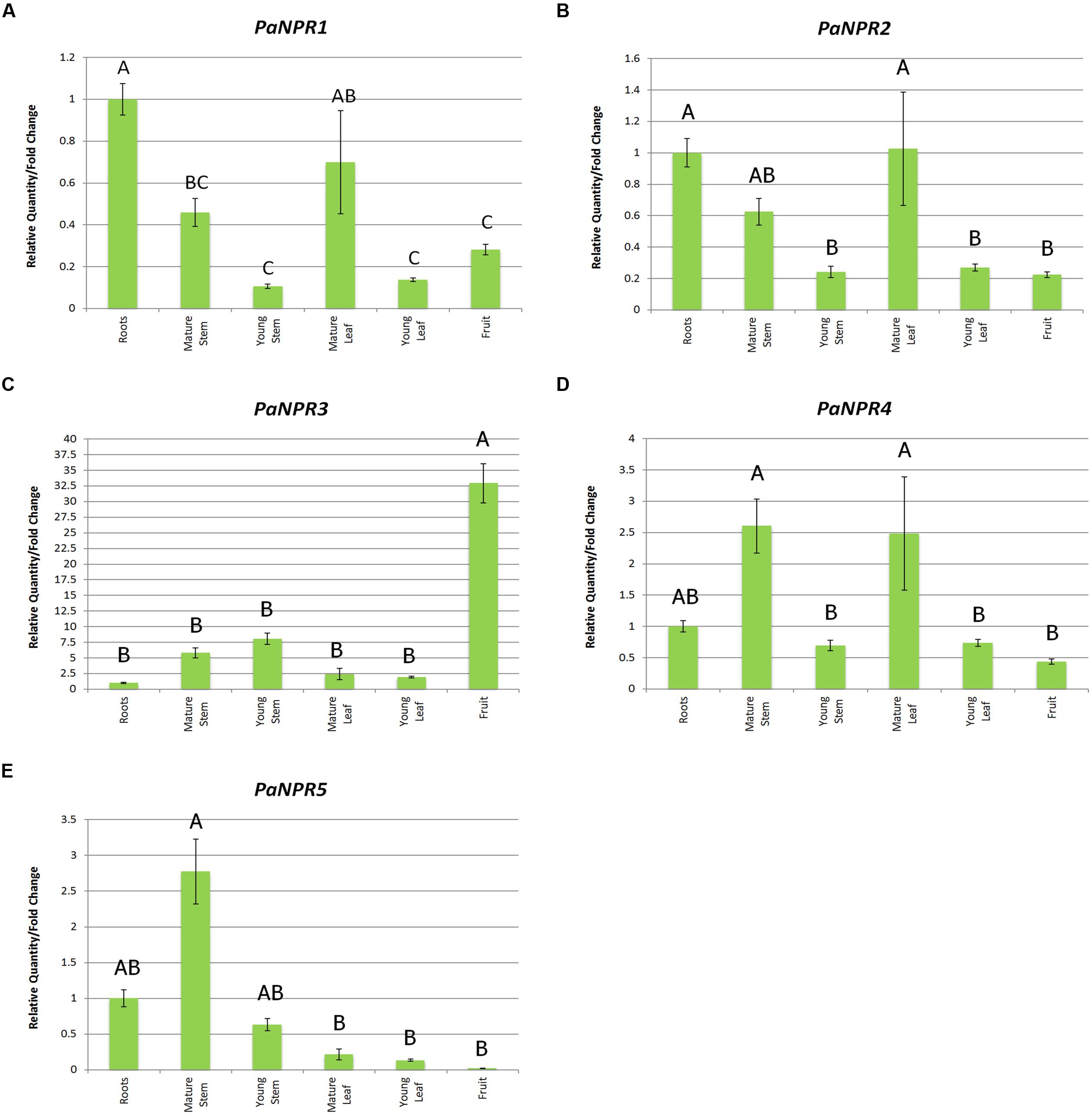
FIGURE 4. Expression of PaNPR1-like genes in various tissues. Normalized relative quantities (fold change) for (A) PaNPR1, (B) PaNPR2, (C) PaNPR3, (D) PaNPR4, and (E) PaNPR5 were calculated using the method described by Pfaffl (2001). The response is indicated by vertical bars across six tissues; feeder roots, mature green stems, mature green leaves, unripe fruit as well as stems and leaves from flush growth (young material), and is labeled on the horizontal axis. The expression in all tissues was calibrated using expression in the roots. SE for each bar is shown. Bars represented with the same letter are not significantly different at P < 0.05.
Conversely, PaNPR3 and PaNPR5 displayed patterns of expression unlike those of the aforementioned PaNPR1-like transcripts. Extremely high PaNPR3 transcript levels were found in the unripe fruit (32.94-fold) relative to the roots, mature stems (5.84-fold), young stems (8.08-fold) and young leaves (1.93-fold; Figure 4C). Inversely, expression of PaNPR5 was the lowest in unripe fruit (0.02-fold) and significantly higher in mature stems (2.77-fold; Figure 4E). The expression of this gene was also significantly less in mature leaves (0.21-fold) and young leaves (0.13-fold) when compared to mature stems (Figure 4E).
PaNPR1-Like Expression in Tolerant and Susceptible Avocado Rootstocks
The expression of NPR1-like genes has been shown to differ significantly between susceptible and tolerant banana cultivars challenged with Fusarium oxysporum Schlecht f. sp. cubense (Smith) Snyd (Foc). Thus determining whether such differences exist between tolerant and susceptible avocado rootstocks could provide insights into molecular differences which may affect P. cinnamomi tolerance. Consequently, RNA from both tolerant (Dusa®) and susceptible (R0.12) avocado rootstocks, infected with P. cinnamomi and harvested at 0 h (uninfected control), 6, 12, and 24 h, was obtained from Engelbrecht et al. (2013).
The expression of PaNPR2 was significantly lower in Dusa® (0.70-fold) as compared to R0.12 (1.87-fold) at 12 h (Figure 5B). PaNPR4 was expressed significantly lower in R0.12 (0.60-fold) when compared to Dusa® (0.94-fold) at 6 h (Figure 5C). However, PaNPR4 was expressed significantly lower in Dusa® (0.52-fold) when compared to R0.12 (0.95) at 12 h (Figure 5C). Expression of PaNPR1 and PaNPR5 was not significantly different when comparing Dusa and R0.12 at any of the time points (Figures 5A,D).
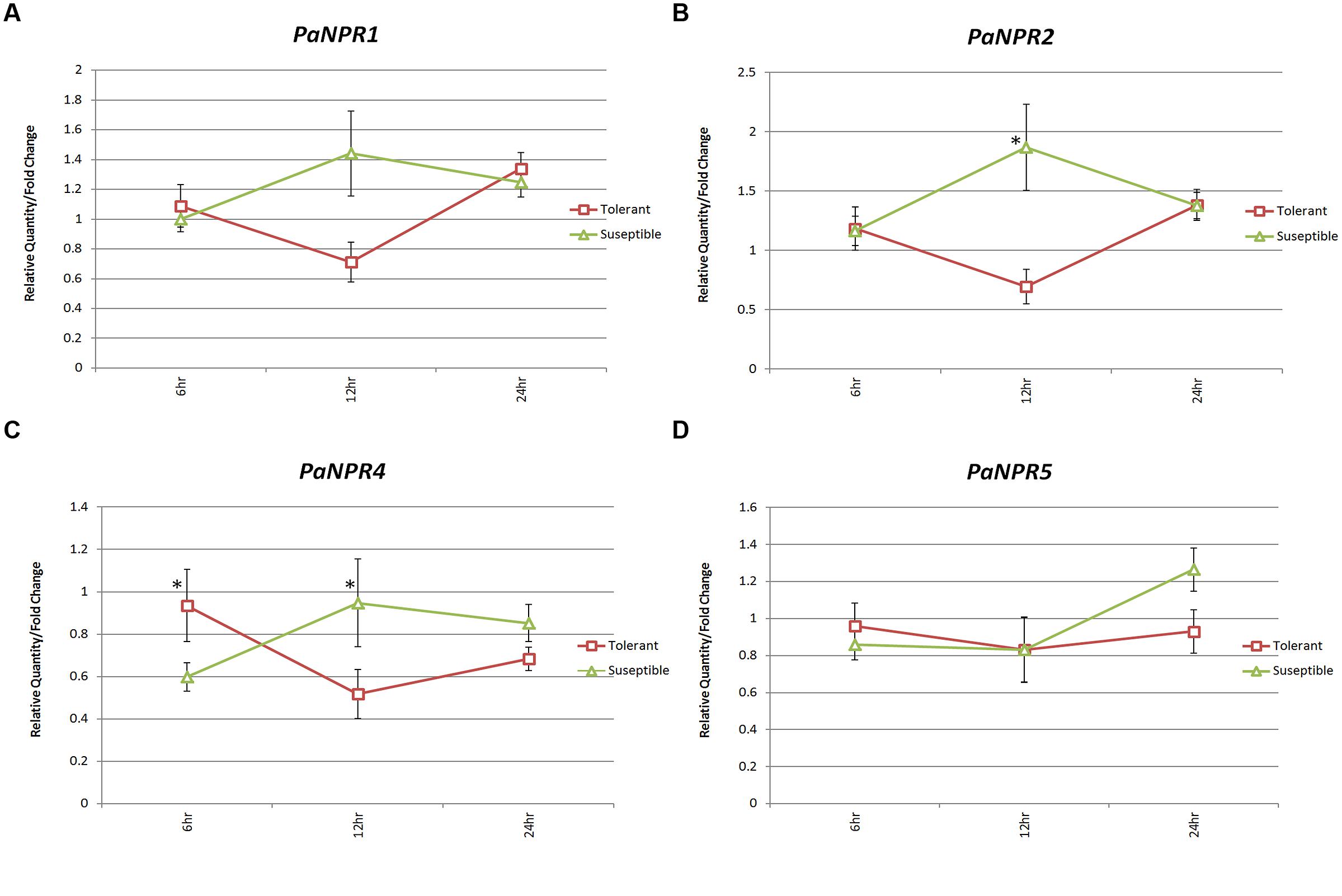
FIGURE 5. Expression of PaNPR1-like genes in tolerant (Dusa®) and susceptible (R0.12) avocado rootstocks infected with P. cinnamomi. Normalized relative quantities (fold change) for (A) PaNPR1, (B) PaNPR2, (C) PaNPR4, and (D) PaNPR5 were calculated using the method described by Pfaffl (2001). The response is indicated by horizontal lines (red – tolerant, green – susceptible) across three time points; 3, 6, and 24 h, labeled on the horizontal axis. The expression was calibrated using expression at 0 h (uninfected control) which was set to a normalized relative expression of 1. SE for each bar is shown. Significant differences (P < 0.05) between control and treated samples is denoted by an asterisk (*) above the data point.
Discussion
We identified five NPR1-like genes in avocado in an attempt to better understand avocado defense response signaling in general, and with regard to the response to PRR. Studies in numerous plant species have highlighted the fundamental involvement of NPR1 in many defense signaling pathways (Cao et al., 1997; Spoel et al., 2003; Le Henanff et al., 2011). This study is the first investigation of the NPR1-like gene family in P. americana, and sets the foundation for further functional characterization of the NPR1-like protein family in avocado.
The avocado genome contains five identifiable NPR1-like genes; these sequences share similar gene structures and protein sequence identities as well as conserved domains and motifs present in Arabidopsis NPR1-like sequences. The PaNPR1-like gene sequences have similar exon/intron structures compared to the members of the Arabidopsis NPR1-like gene family to which each is most similar. Specifically, the exon/intron structures of PaNPR1, PaNPR2, and PaNPR4 are comparable to that of AtNPR1-4. Interestingly, PaNPR1 contains an extra, short, exon (exon 3) which could either be an assembly error or a unique aspect of this gene. Furthermore, the exon/intron structures of PaNPR3 and PaNPR5 are similar to that of AtBOP2. The predicted coding sequences for all five PaNPR1-like genes harbor the BTB/POZ and ankyrin repeat domains, characteristic of NPR1-like proteins. These domains are crucial components of NPR1 and provide functions relating to NPR1-dependant co-activation of TGA transcription factors and protein–protein binding (Cao et al., 1997; Rochon et al., 2006). Only PaNPR2 contains the complete IκB phosphodegron motif (DSxxxS) found in AtNPR1, while PaNPR1 contains a lysine substitution at the first serine residue. Both serines of this motif are phosphorylated during SA treatment, leading to proteasome mediated turn-over and degradation of NPR1 (Spoel et al., 2009), suggesting that PaNPR2 may be regulated similarly. However, the effect of the serine to lysine substitution in PaNPR1 is unclear, particularly because basic amino acids such as lysine have been shown to undergo phosphorylation (Ciesla et al., 2011). Similar to AtNPR1, NLS1 sequences are also present in PaNPR1, PaNPR2, and PaNPR4 containing five, four, and three of the amino acids known to be essential to nuclear localization of AtNPR1 (Kinkema et al., 2000). Moreover, PaNPR1, PaNPR2, and PaNPR4 contain highly conserved NIMIN1/2 binding regions and LENRV motifs (Maier et al., 2011). Together these data suggest that PaNPR1, PaNPR2, and PaNPR4 are comparable to AtNPR1 and could possibly partake in the perception of SA and regulation of defense responses in avocado. Seemingly distinct, PaNPR3 and PaNPR5 do not contain these motifs and conserved regions, similar to the AtBOP proteins. The substantial difference in protein length and sequence composition of this subset of proteins suggests functionally diverse roles from PaNPR1, PaNPR2, and PaNPR4, and can be suggested that these proteins may be involved in certain aspects of tissue development as seen in AtBOP1 and AtBOP2 (Hepworth et al., 2005).
Phylogenetic analysis reveals that all five PaNPR1-like proteins group with other known NPR1-like sequences, clustering into three distinct clades (Peraza-Echeverria et al., 2012). PaNPR1 and PaNPR2 fall within the same clade as AtNPR1, which is a known positive regulator of SAR (Cao et al., 1997, 1998). PaNPR4 groups with AtNPR3 and AtNPR4 in the second clade. NPR1-like proteins within this group have been shown to negatively regulate SAR (Zhang et al., 2006), yet are able to perceive SA and are vital in mounting SAR (Fu et al., 2012). The last two, PaNPR3 and PaNPR5, fall within the third clade together with AtBOP1 and AtBOP2, which are known for their involvement in development of lateral organs (Hepworth et al., 2005). This phylogenetic analysis provides a second line of evidence suggesting possible functional distinctions between members of the PaNPR1-like protein family.
This study further describes the transcriptional response of the PaNPR1-like genes to SA, MeJA, and P. cinnamomi treatments as well as their expression levels in different tissues. Surprisingly, PaNPR1 was down regulated by SA at 12 h, contrasting with AtNPR1, which was up-regulated approximately twofold 24 h after SA application in Arabidopsis (Cao et al., 1998). This may point to an alternative function of PaNPR1 during defense responses and warrants further investigation. In banana cultivars which are resistant to Foc, NPR1 is up-regulated to a greater extent and at earlier time points after SA treatment than in susceptible cultivars (Endah et al., 2008). Similarly, PaNPR2 was up-regulated at the earliest time point after SA treatment in the PRR tolerant avocado rootstock Dusa®. Additionally, up-regulation of PaNPR2 corresponded to an increase in PaPR1 gene expression. Similarly, PaNPR4 was up-regulated soon after SA application and reached peak expression at 12 h. Interestingly, the highest expression of PaNPR4 corresponded to a substantial decrease in PaPR1 expression, suggesting that PaNPR4 may negatively regulate the expression of PaPR1. Treatment with MeJA led to decreased transcript abundance for PaNPR1, PaNPR2, and PaNPR4 for extended periods of time, opposite to that seen in rice and banana (Yuan et al., 2007; Endah et al., 2008). The regulation of NPR1 during antagonistic cross-talk between SA and JA mediated defense responses may thus differ between some monocot and dicot plants. Interestingly, the regulation of PaNPR1, PaNPR2, and PaNPR4 during P. cinnamomi treatment had expression patterns similar to that of SA treatment at earlier time points (6–18 h) and JA treatment at later time points (24–96 h). This could indicate the point at which P. cinnamomi switches from a biotrophic to a necrotrophic life stage, thus activating the SA and JA pathways, respectively.
Oddly, the induced expression of PaNPR2 and PaNPR4 during SA treatment was not observed during infection with P. cinnamomi. In our opinion three possible explanations exist: (1) PaNPR2 and PaNPR4 might be predominantly regulated at the protein level, (2) these proteins might not be involved in defense responses against P. cinnamomi or, (3) P. cinnamomi suppresses expression of these genes in order to promote successful host invasion. It has been noted that NPR1 is subject to extensive post-translational regulation (Mou et al., 2003; Spoel et al., 2009), thus changes in expression might not reflect the factual role of PaNPR1, PaNPR2, or PaNPR4 during defense response. Furthermore, Phytophthora species have been known to alter host gene expression in order to suppress host defense pathways and mediate infection (Oßwald et al., 2014).
A common trend in the expression of PaNPR1, PaNPR2, and PaNPR4 in various tissues was seen; transcript levels of these genes were significantly higher in mature tissues than immature tissues, an observation that may be explained by the establishment of SAR in mature tissues. Expression of PaNPR3 was undetectable in the roots in any of the treatments, yet this gene was expressed at much higher levels in aerial tissues, with the highest levels being detected in fruit. On the other hand, while PaNPR5 was readily detected in the roots, it was unresponsive to SA or MeJA treatments and significantly down-regulated by P. cinnamomi during later time points. These data support our initial hypothesis that PaNPR3 and PaNPR5 are unlikely to be involved in defense responses, and are instead more likely to be involved in development of certain tissues.
Finally, significant differences in the expression of PaNPR2 and PaNPR4 were observed when comparing tolerant (Dusa®) and susceptible (R0.12) avocado rootstock cultivars. The expression of PaNPR2 and PaNPR4 is significantly lower at 12 h after infection in Dusa® when compared to R0.12. In our opinion these observations could be explained when considering P. cinnamomi switching from a biotrophic to a necrotrophic life cycle. In this case, increased expression of PaNPR1-like defense related genes would likely suppress the JA/ET pathway and prevent effective control of P. cinnamomi. Thus it is conceivable that P. cinnamomi switches to a necrotrophic life cycle somewhere around 12 h after infection and that Dusa® reacts to this change more quickly than R0.12. This would explain, at least to some extent, tolerance in Dusa® and susceptibility in R0.12.
This study provides evidence assisting in the preliminary functional annotation of five newly discovered NPR1-like genes from avocado. Sequence structure and homology as well as phylogenetic analyze suggest that three PaNPR1-like proteins may be involved in defense responses, while the remaining two are most likely involved tissue development. Hormone and P. cinnamomi treatments, as well as expression in various tissues provide support for this and allow future research to focus on defense related PaNPR1-like proteins. Future efforts would be focused on intracellular interactions and localization of defense related PaNPR1-like proteins as well as the effect of overexpressing defense related PaNPR1-like genes in wild-type and npr1 mutant Arabidopsis. Information from this and future studies could aid in understanding PRR tolerance and lead to the development of more tolerant avocado rootstocks.
Author Contributions
RB drafted the manuscript, performed conceptual, and experimental design as well as performed the experimental work. WM assisted in expression analysis and experimental design. JE performed experimental work. BR provided general supervision and assisted in drafting the manuscript. EL sequenced the genome and provided the PaNPR1-like sequences. NvdB conceived the study, experimental design and assisted in drafting the manuscript. All authors contributed to and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to Dr. Luis Herrera-Estrella who provided us with access to the genome. We would also like to thank Dr. Sanushka Naidoo who advised on the structure and content of the study. This work was supported by the University of Pretoria and the Forestry and Agricultural Biotechnology Institute (FABI). Student funding was provided by the National Research Foundation and project funding was provided by the Hans Merensky Foundation.
Footnotes
- ^ http://faostat3.fao.org/faostat-gateway/go/to/home/E
- ^ http://imed.med.ucm.es/Tools/sias.html
- ^ http://www.ncbi.nlm.nih.gov/genbank/
- ^ http://www.ncbi.nlm.nih.gov/
- ^ http://www.socr.ucla.edu
References
An, C., and Mou, Z. (2011). Salicylic acid and its function in plant immunity. J. Integr. Plant Biol. 53, 412–428. doi: 10.1111/j.1744-7909.2011.01043.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Bekker, T. F., Kaiser, C., and Labuschagne, N. (2006). Efficacy of water soluble silicon against Phytophthora cinnamomi root rot of avocado: a progress report. S. Afr. Avocado Growers’ Assoc. Yearb. 29, 58–62.
Bi, Y. M., Kenton, P., Mur, L., Darby, R., and Draper, J. (1995). Hydrogen peroxide does not function downstream of salicylic acid in the induction of PR protein expression. Plant J. 8, 235–245. doi: 10.1046/j.1365-313X.1995.08020235.x
Botha, T., and Kotze, J. M. (1989). Exudates of avocado rootstocks and their possible role in resistance to Phytophthora cinnamomi. S. Afr. Avocado Growers’ Assoc. Yearb. 12, 64–65.
Bustin, S. A., Benes, V., Garson, J. A., Hellemans, J., Huggett, J., Kubista, M., et al. (2009). The MIQE guidelines: minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 55, 611–622. doi: 10.1373/clinchem.2008.112797
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cao, H., Bowling, S. A., Gordon, A. S., and Dong, X. (1994). Characterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 6, 1583–1592. doi: 10.1105/tpc.6.11.1583
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cao, H., Glazebrook, J., Clarke, J. D., Volko, S., and Dong, X. (1997). The Arabidopsis NPR1 gene that controls systemic acquired resistance encodes a novel protein containing ankyrin repeats. Cell 88, 57–63. doi: 10.1016/S0092-8674(00)81858-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Cao, H., Li, X., and Dong, X. (1998). Generation of broad-spectrum disease resistance by overexpression of an essential regulatory gene in systemic acquired resistance. Proc. Natl. Acad. Sci. U.S.A. 95, 6531–6536. doi: 10.1073/pnas.95.11.6531
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chang, S., Puryear, J., and Cairney, J. (1993). A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 11, 113–116. doi: 10.1007/BF02670468
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chen, X. K., Zhang, J. Y., Zhang, Z., Du, X. L., Du, B. B., and Qu, S. C. (2012). Overexpressing MhNPR1 in transgenic Fuji apples enhances resistance to apple powdery mildew. Mol. Biol. Rep. 39, 8083–8089. doi: 10.1007/s11033-012-1655-3
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Chern, M., Canlas, P. E., Fitzgerald, H. A., and Ronald, P. C. (2005). Rice NRR, a negative regulator of disease resistance, interacts with Arabidopsis NPR1 and rice NH1. Plant J. 43, 623–635. doi: 10.1111/j.1365-313X.2005.02485.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ciesla, J., Fraczyk, T., and Rode, W. (2011). Phosphorylation of basic amino acid residues in proteins: important but easily missed. Acta Biochim. Pol. 58, 137–148.
Coffey, D. M. (1987). Phytophthora root rot of avocado- an integrated approach to control in California. California Avocado Soc. Yearb. 71, 121–137.
Collins, G. A., and Tansey, W. P. (2006). The proteasome: a utility tool for transcription? Curr. Opin. Genet. Dev. 16, 197–202. doi: 10.1016/j.gde.2006.02.009
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Darvas, J. M., Toerien, J. C., and Milne, D. L. (1984). Control of avocado root rot by trunk injection with phosetyl-AI. Plant Dis. 68, 691–693. doi: 10.1094/PD-68-691
Delaney, T. P., Uknes, S., Vernooij, B., Friedrich, L., Weymann, K., Negrotto, D., et al. (1994). A central role of salicylic acid in plant disease resistance. Science 266, 1247–1250. doi: 10.1126/science.266.5188.1247
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Despres, C., Delong, C., Glaze, S., Liu, E., and Fobert, P. R. (2000). The Arabidopsis NPR1/NIM1 protein enhances the DNA binding activity of a subgroup of the TGA family of bZIP transcription factors. Plant Cell 12, 279–290. doi: 10.1105/tpc.12.2.279
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Dobrowolski, M. P., Shearer, B. L., Colquhoun, I. J., O’brien, P. A., and Hardy, G. E. S. (2008). Selection for decreased sensitivity to phosphite in Phytophthora cinnamomi with prolonged use of fungicide. Plant Pathol. 57, 928–936. doi: 10.1111/j.1365-3059.2008.01883.x
Duvenhage, J. (1994). Monitoring the resistance of Phytophthora cinnamomi to Fosetyl-Al and H3PO3. S. Afr. Avocado Growers’ Assoc. Yearb. 17, 35–37.
El Oirdi, M., El Rahman, T. A., Rigano, L., El Hadrami, A., Rodriguez, M. C., Daayf, F., et al. (2011). Botrytis cinerea manipulates the antagonistic effects between immune pathways to promote disease development in tomato. Plant Cell 23, 2405–2421. doi: 10.1105/tpc.111.083394
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Endah, R., Beyene, G., Kiggundu, A., van den Berg, N., Schluter, U., Kunert, K., et al. (2008). Elicitor and Fusarium-induced expression of NPR1-like genes in banana. Plant Physiol. Biochem. 46, 1007–1014. doi: 10.1016/j.plaphy.2008.06.007
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Engelbrecht, J., Duong, T. A., and van den Berg, N. (2013). Development of a nested quantitative real time PCR for detecting Phytophthora cinnamomi in Persea americana rootstocks. Plant Dis. 9, 1012–1017. doi: 10.1094/PDIS-11-12-1007-RE
Fan, W., and Dong, X. (2002). In vivo interaction between NPR1 and transcription factor TGA2 leads to salicylic acid-mediated gene activation in Arabidopsis. Plant Cell 14, 1377–1389. doi: 10.1105/tpc.001628
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Friedrich, L., Lawton, K., Dietrich, R., Willits, M., Cade, R., and Ryals, J. (2001). NIM1 overexpression in Arabidopsis potentiates plant disease resistance and results in enhanced effectiveness of fungicides. Mol. Plant-Microbe Interact. 14, 1114–1124. doi: 10.1094/MPMI.2001.14.9.1114
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Frederich, L., Lawton, K., Reuss, W., Masner, P., Specker, N., Gut Rella, M., et al. (1996). A benzothiadiazole induces systemic acquired resistance in tobacco. Plant J. 10, 61–70. doi: 10.1046/j.1365-313X.1996.10010061.x
Friedrich, L., Vernooij, E., Gaffney, T., Morse, A., and Ryals, J. (1995). Characterization of tobacco plants expressing a bacterial salicylate hydroxylase gene. Plant Mol. Biol. 29, 959–968. doi: 10.1007/BF00014969
Fu, Z. Q., Yan, S., Saleh, A., Wang, W., Ruble, J., Oka, N., et al. (2012). NPR3 and NPR4 are receptors for the immune signal salicylic acid in plants. Nature 486, 228–232. doi: 10.1038/nature11162
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Gabor, B. K., and Coffey, D. M. (1991). Comparison of rapid methods for evaluating resistance to Phytophthora cinnamomi in avocado rootstocks. Plant Dis. 75, 118–120. doi: 10.1094/PD-75-0118
Gaffney, T., Friedrich, L., Vernooij, B., Negrotto, D., Nye, G., Uknes, S., et al. (1993). Requirement of salicylic acid for the induction of systemic acquired resistance. Science 261, 754–756. doi: 10.1126/science.261.5122.754
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
García-Pineda, E., Benezer-Benezer, M., Gutiérrez-Segundo, A., Rangel-Sánchez, G., Arreola-Cortés, A., and Castro-Mercado, E. (2010). Regulation of defence responses in avocado roots infected with Phytophthora cinnamomi (Rands). Plant Soil 331, 45–56. doi: 10.1007/s11104-009-0225-5
Giblin, F., Pegg, K., Willingham, S., Anderson, J., Coates, L., Cooke, T., et al. (2005). “Phytophthora revisited,” in Proceedings of the New Zealand and Australia Avocado Grower’s Conference (Tauranga: New Zealand Avocado Growers Association).
Glazebrook, J. (2005). Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 43, 205–227. doi: 10.1146/annurev.phyto.43.040204.135923
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Glazebrook, J., Rogers, E. E., and Ausubel, F. M. (1996). Isolation of Arabidopsis mutants with enhanced disease susceptibility by direct screening. Genetics 143, 973–982.
Guo, A. Y., Zhu, Q. H., Chen, X., and Luo, J. C. (2007). [GSDS: a gene structure display server]. Yi Chuan 29, 1023–1026. doi: 10.1360/yc-007-1023
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hardham, A. R. (2005). Phytophthora cinnamomi. Mol. Plant Pathol. 6, 589–604. doi: 10.1111/j.1364-3703.2005.00308.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Heidel, A. J., Clarke, J. D., Antonovics, J., and Dong, X. (2004). Fitness costs of mutations affecting the systemic acquired resistance pathway in Arabidopsis thaliana. Genetics 168, 2197–2206. doi: 10.1534/genetics.104.032193
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Hepworth, S. R., Zhang, Y., Mckim, S., Li, X., and Haughn, G. W. (2005). BLADE-ON-PETIOLE-dependent signaling controls leaf and floral patterning in Arabidopsis. Plant Cell 17, 1434–1448. doi: 10.1105/tpc.104.030536
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kaiser, C., Whiley, A. W., Pegg, K. G., Hargreaves, P. A., and Weinhart, M. P. (1997). “Determination of critical root concentrations of phosphate to control Phytophthora root rot in avocado,” in Proceedings of the Joint Meeting of the Australian Avocado Grower’s Federation Inc and the New Zealand Avocado Grower’s Association, Tauranga, 155.
Kinkema, M., Fan, W., and Dong, X. (2000). Nuclear localization of NPR1 is required for activation of PR gene expression. Plant Cell 12, 2339–2350. doi: 10.1105/tpc.12.12.2339
Kremer-Köhne, S., and Mukhumo, M. L. (2003). “Breeding and field evaluation of new rootstocks for increased Hass yields and resistance to root rot in South Africa,” in Proceedings of V World Avocado Congress (Granada - Málaga: Actas V Congreso Mundial del Aguacate), 555–560. doi: 10.1007/s11248-012-9652-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Kumar, V., Joshi, S. G., Bell, A. A., and Rathore, K. S. (2012). Enhanced resistance against Thielaviopsis basicola in transgenic cotton plants expressing Arabidopsis NPR1 gene. Transgenic Res. 22, 359–368. doi: 10.1007/s11248-012-9652-9
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Lawton, K., Weymann, K., Friedrich, L., Vernooij, B., Uknes, S., and Ryals, J. (1995). Systemic acquired resistance in Arabidopsis requires salicylic acid but not ethylene. Mol. Plant-Microbe Interact. 8, 863–870. doi: 10.1094/MPMI-8-0863
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Le Henanff, G., Farine, S., Kieffer-Mazet, F., Miclot, A. S., Heitz, T., Mestre, P., et al. (2011). Vitis vinifera VvNPR1.1 is the functional ortholog of AtNPR1 and its overexpression in grapevine triggers constitutive activation of PR genes and enhanced resistance to powdery mildew. Planta 234, 405–417. doi: 10.1007/s00425-011-1412-1
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Le Henanff, G., Heitz, T., Mestre, P., Mutterer, J., Walter, B., and Chong, J. (2009). Characterization of Vitis vinifera NPR1 homologs involved in the regulation of pathogenesis-related gene expression. BMC Plant Biol. 9:54. doi: 10.1186/1471-2229-9-54
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Li, X., Zhang, Y., Clarke, J. D., Li, Y., and Dong, X. (1999). Identification and cloning of a negative regulator of systemic acquired resistance, SNI1, through a screen for suppressors of npr1-1. Cell 98, 329–339. doi: 10.1016/S0092-8674(00)81962-5
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Liu, G., Holub, E. B., Alonso, J. M., Ecker, J. R., and Fobert, P. R. (2005). An Arabidopsis NPR1-like gene, NPR4, is required for disease resistance. Plant J. 41, 304–318. doi: 10.1111/j.1365-313X.2004.02296.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Mahomed, W., and van den Berg, N. (2011). EST sequencing and gene expression profiling of defence-related genes from Persea americana infected with Phytophthora cinnamomi. BMC Plant Biol. 11:167. doi: 10.1186/1471-2229-11-167
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Maier, F., Zwicker, S., Huckelhoven, A., Meissner, M., Funk, J., Pfitzner, A. J., et al. (2011). NONEXPRESSOR OF PATHOGENESIS-RELATED PROTEINS1 (NPR1) and some NPR1-related proteins are sensitive to salicylic acid. Mol. Plant Pathol. 12, 73–91. doi: 10.1111/j.1364-3703.2010.00653.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Malamy, J., Carr, J. P., Klessig, D. F., and Raskin, I. (1990). Salicylic acid: a likely endogenous signal in the resistance response of tobacco to viral infection. Science 250, 1002–1004. doi: 10.1126/science.250.4983.1002
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Malnoy, M., Jin, Q., Borejsza-Wysocka, E. E., He, S. Y., and Aldwinckle, H. S. (2007). Overexpression of the apple MpNPR1 gene confers increased disease resistance in Malus x domestica. Mol. Plant-Microbe Interact. 20, 1568–1580. doi: 10.1094/MPMI-20-12-1568
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Menge, J. A. (1999). “Screening and evaluation of new rootstocks with resistance to Phytophthora cinnamomi,” in Proceedings of the California Avocado Research Symposium (San Juan Capistrano: California Avocado Society), 69–72.
Metraux, J. P., Ahl-Goy, P., Staub, T., Speich, J., Steinemann, A., Ryals, J., et al. (1991). “Induced resistance in cucumber in response to 2,6-dichloroisonicotinic acid and pathogens,” in Advances in Molecular Genetics of Plant-Microbe Interactions, Vol. 1. eds H. Hennecke and D. S. Verma (Berlin: Springer), 432–439.
Mou, Z., Fan, W., and Dong, X. (2003). Inducers of plant systemic acquired resistance regulate NPR1 function through redox changes. Cell 113, 935–944. doi: 10.1016/S0092-8674(03)00429-X
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Oßwald, W., Fleischmann, F., Rigling, D., Coelho, A. C., Cravador, A., Diez, J., et al. (2014). Strategies of attack and defence in woody plant–Phytophthora interactions. For. Pathol. 44, 169–190. doi: 10.1111/efp.12096
Pegg, K. G., Whiley, A. W., Saranah, J. B., and Glass, R. J. (1985). Control of Phytophthora root rot of avocado with phosphorous acid. Aust. Plant Pathol. 14, 25–29. doi: 10.1071/APP9850025
Peraza-Echeverria, S., Santamaria, J. M., Fuentes, G., Menendez-Ceron, M. A., Vallejo-Reyna, M. A., and Herrera-Valencia, V. A. (2012). The NPR1 family of transcription cofactors in papaya: insights into it’s structure, phylogeny and expression. Genes Genom. 34, 379–390. doi: 10.1007/s13258-011-0218-7
Pfaffl, M. W. (2001). A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 29, e45. doi: 10.1093/nar/29.9.e45
Phillips, D., Grant, B. R., and Weste, G. (1987). Histological changes in the roots of an avocado cultivar, Duke 7, infected with Phytophthora cinnamomi. Phytopathology 77, 691–698. doi: 10.1094/Phyto-77-691
Rasmussen, J. B., Hammerschmidt, R., and Zook, M. N. (1991). Systemic induction of salicylic acid accumulation in cucumber after inoculation with Pseudomonas syringae pv syringae. Plant Physiol. 97, 1342–1347. doi: 10.1104/pp.97.4.1342
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rate, D. N., and Greenberg, J. T. (2001). The Arabidopsis aberrant growth and death2 mutant shows resistance to Pseudomonas syringae and reveals a role for NPR1 in suppressing hypersensitive cell death. Plant J. 27, 203–211. doi: 10.1046/j.0960-7412.2001.1075umedoc.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Reeksting, B. J., Coetzer, N., Mahomed, W., Engelbrecht, J., and van den Berg, N. (2014). De novo sequencing, assembly, and analysis of the root transcriptome of Persea americana (Mill.) in response to Phytophthora cinnamomi and flooding. PLoS ONE 9:e86399. doi: 10.1371/journal.pone.0086399
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Robert-Seilaniantz, A., Grant, M., and Jones, J. D. (2011). Hormone crosstalk in plant disease and defense: more than just jasmonate-salicylate antagonism. Annu. Rev. Phytopathol. 49, 317–343. doi: 10.1146/annurev-phyto-073009-114447
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rochon, A., Boyle, P., Wignes, T., Fobert, P. R., and Despres, C. (2006). The coactivator function of Arabidopsis NPR1 requires the core of its BTB/POZ domain and the oxidation of C-terminal cysteines. Plant Cell 18, 3670–3685. doi: 10.1105/tpc.106.046953
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Rookes, J. E., Wright, M. L., and Cahill, D. M. (2008). Elucidation of defence responses and signalling pathways induced in Arabidopsis thaliana following challenge with Phytophthora cinnamomi. Physiol. Mol. Plant Pathol. 72, 151–161. doi: 10.1016/j.pmpp.2008.08.005
Ryals, J., Weymann, K., Lawton, K., Friedrich, L., Ellis, D., Steiner, H. Y., et al. (1997). The Arabidopsis NIM1 protein shows homology to the mammalian transcription factor inhibitor I kappa B. Plant Cell 9, 425–439. doi: 10.1105/tpc.9.3.425
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Saitou, N., and Nei, M. (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 4, 406–425.
Salghetti, S. E., Muratani, M., Wijnen, H., Futcher, B., and Tansey, W. P. (2000). Functional overlap of sequences that activate transcription and signal ubiquitin-mediated proteolysis. Proc. Natl. Acad. Sci. U.S.A. 97, 3118–3123. doi: 10.1073/pnas.97.7.3118
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sánchez-Pérez, J., Jaimes-Lara, M., Salgado-Garciglia, R., and López-Meza, J. (2009). Root extracts from Mexican avocado Persea americana var. drymifolia inhibit the mycelial growth of the oomycete Phytophthora cinnamomi. Eur. J. Plant Pathol. 124, 595–601. doi: 10.1007/s10658-009-9446-y
Shao, Y., Zhang, H., He, H., Cheng, B., and Xiang, Y. (2013). Molecular cloning and characterization of orthologues of NPR1 gene from poplar. J. Phytopathol. 161, 35–42. doi: 10.1111/jph.12002
Sigrist, C. J., Cerutti, L., De Castro, E., Langendijk-Genevaux, P. S., Bulliard, V., Bairoch, A., et al. (2010). PROSITE, a protein domain database for functional characterization and annotation. Nucleic Acids Res. 38, D161–D166. doi: 10.1093/nar/gkp885
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Solovyev, V., Kosarev, P., Seledsov, I., and Vorobyev, D. (2006). Automatic annotation of eukaryotic genes, pseudogenes and promoters. Genome Biol. 7, 10.11–10.12. doi: 10.1186/gb-2006-7-s1-s10
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Spoel, S. H., Koornneef, A., Claessens, S. M., Korzelius, J. P., Van Pelt, J. A., Mueller, M. J., et al. (2003). NPR1 modulates cross-talk between salicylate- and jasmonate-dependent defense pathways through a novel function in the cytosol. Plant Cell 15, 760–770. doi: 10.1105/tpc.009159
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Spoel, S. H., Mou, Z., Tada, Y., Spivey, N. W., Genschik, P., and Dong, X. (2009). Proteasome-mediated turnover of the transcription coactivator NPR1 plays dual roles in regulating plant immunity. Cell 137, 860–872. doi: 10.1016/j.cell.2009.03.038
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sticher, L., Mauch-Mani, B., and Metraux, J. P. (1997). Systemic acquired resistance. Annu. Rev. Phytopathol. 35, 235–270. doi: 10.1146/annurev.phyto.35.1.235
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Thompson, J. D., Higgins, D. G., and Gibson, T. J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22, 4673–4680. doi: 10.1093/nar/22.22.4673
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Ward, E. R., Uknes, S. J., Williams, S. C., Dincher, S. S., Wiederhold, D. L., Alexander, D. C., et al. (1991). Coordinate gene activity in response to agents that induce systemic acquired resistance. Plant Cell 3, 1085–1094. doi: 10.1105/tpc.3.10.1085
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Weigel, R. R., Bauscher, C., Pfitzner, A. J., and Pfitzner, U. M. (2001). NIMIN-1, NIMIN-2 and NIMIN-3, members of a novel family of proteins from Arabidopsis that interact with NPR1/NIM1, a key regulator of systemic acquired resistance in plants. Plant Mol. Biol. 46, 143–160. doi: 10.1023/A:1010652620115
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Weste, G. (1983). Phytophthora cinnamomi: the dynamics of chlamydospore formation and survival. J. Phytopathol. 106, 163–176. doi: 10.1111/j.1439-0434.1983.tb00039.x
White, R. F. (1979). Acetylsalicylic acid (aspirin) induces resistance to tobacco mosaic virus in tobacco. Virology 99, 410–412. doi: 10.1016/0042-6822(79)90019-9
Yuan, Y., Zhong, S., Li, Q., Zhu, Z., Lou, Y., Wang, L., et al. (2007). Functional analysis of rice NPR1-like genes reveals that OsNPR1/NH1 is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotechnol. J. 5, 313–324. doi: 10.1111/j.1467-7652.2007.00243.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zentmyer, G. A. (1984). Avocado diseases. Int. J. Pest Manage. 30, 388–400. doi: 10.1080/09670878409370915
Zhang, Y., Cheng, Y. T., Qu, N., Zhao, Q., Bi, D., and Li, X. (2006). Negative regulation of defense responses in Arabidopsis by two NPR1 paralogs. Plant J. 48, 647–656. doi: 10.1111/j.1365-313X.2006.02903.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhang, Y., Fan, W., Kinkema, M., Li, X., and Dong, X. (1999). Interaction of NPR1 with basic leucine zipper protein transcription factors that bind sequences required for salicylic acid induction of the PR-1 gene. Proc. Natl. Acad. Sci. U.S.A. 96, 6523–6528. doi: 10.1073/pnas.96.11.6523
Zhang, Y., Tessaro, M. J., Lassner, M., and Li, X. (2003). Knockout analysis of Arabidopsis transcription factors TGA2, TGA5, and TGA6 reveals their redundant and essential roles in systemic acquired resistance. Plant Cell 15, 2647–2653. doi: 10.1105/tpc.014894
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhou, J. M., Trifa, Y., Silva, H., Pontier, D., Lam, E., Shah, J., et al. (2000). NPR1 differentially interacts with members of the TGA/OBF family of transcription factors that bind an element of the PR-1 gene required for induction by salicylic acid. Mol. Plant-Microbe Interact. 13, 191–202. doi: 10.1094/MPMI.2000.13.2.191
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zwicker, S., Mast, S., Stos, V., Pfitzner, A. J., and Pfitzner, U. M. (2007). Tobacco NIMIN2 proteins control PR gene induction through transient repression early in systemic acquired resistance. Mol. Plant Pathol. 8, 385–400. doi: 10.1111/j.1364-3703.2007.00399.x
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: avocado, Phytophthora cinnamomi, NPR1, expression analysis, salicylic acid, jasmonic acid, pathogenesis-related
Citation: Backer R, Mahomed W, Reeksting BJ, Engelbrecht J, Ibarra-Laclette E and van den Berg N (2015) Phylogenetic and expression analysis of the NPR1-like gene family from Persea americana (Mill.). Front. Plant Sci. 6:300. doi: 10.3389/fpls.2015.00300
Received: 22 January 2015; Accepted: 14 April 2015;
Published online: 29 April 2015
Edited by:
Georgina Hernandez, Centro de Ciencias Genomicas – Universidad Nacional Autonoma de Mexico, MexicoReviewed by:
Dacheng Tian, Nanjing University, ChinaDaniel Pinero, Universidad Nacional Autonoma de Mexico, Mexico
Adriana Garay, Universidad Nacional Autónoma de México, Mexico
Copyright © 2015 Backer, Mahomed, Reeksting, Engelbrecht, Ibarra-Laclette and van den Berg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Noëlani van den Berg, Department of Genetics, Fruit Tree Biotechnology Program, University of Pretoria, Plant Sciences Complex Room 5-17, Corner Lynwood and Roper Street, Pretoria 0002, South Africa,bm9lbGFuaS52ZGJlcmdAZmFiaS51cC5hYy56YQ==