 William Oki Wong
William Oki Wong- 1State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing, China
- 2Department of Geology, Indiana University, Bloomington, IN, USA
- 3Department of Paleobiology, National Museum of Natural History, Washington, DC, USA
- 4Department of Entomology and BEES Program, University of Maryland, College Park, MD, USA
- 5College of Life Sciences, Capital Normal University, Beijing, China
- 6Paleontological Institute, Shenyang Normal University, Shenyang, China
- 7Botanische Staatssammlung München, Munich, Germany
Archaeamphora longicervia H. Q. Li was described as an herbaceous, Sarraceniaceae-like pitcher plant from the mid Early Cretaceous Yixian Formation of Liaoning Province, northeastern China. Here, a re-investigation of A. longicervia specimens from the Yixian Formation provides new insights into its identity and the morphology of pitcher plants claimed by Li. We demonstrate that putative pitchers of Archaeamphora are insect-induced leaf galls that consist of three components: (1) an innermost larval chamber; (2) an intermediate zone of nutritive tissue; and (3) an outermost wall of sclerenchyma. Archaeamphora is not a carnivorous, Sarraceniaceae-like angiosperm, but represents insect-galled leaves of the previously reported gymnosperm Liaoningocladus boii G. Sun et al. from the Yixian Formation.
Introduction
Li (2005) described Archaeamphora longicervia H. Q. Li as an “Early Cretaceous sarraceniacean-like pitcher plant” from the Yixian Formation of Liaoning Province in northeastern China. These fossils were interpreted by Li as an herbaceous plant having spirally arranged, ascidiate pitchers and phyllodia-like leaves with parallel veins, similar to modern Sarraceniaceae, a New World carnivorous plant family of Ericales represented by three extant genera: Sarracenia L., Darlingtonia Torr., and Heliamphora Benth. (Lloyd, 1942; Juniper et al., 1989; Schnell, 2002). However, doubts have been cast on whether Archaeamphora actually represents an early sarraceniacean plant, because its age contradicts the divergence time estimate, phylogeny, and biogeography of carnivorous Ericalean lineages (Fleischmann, 2010; Ellison et al., 2012; Brittnacher, 2013; Givnish, 2015; Sadowski et al., 2015). Here, we propose a new perspective on the morphology, identity, and affinity of A. longicervia in light of a re-investigation of newly collected Yixian specimens.
New Fossils of Archaeamphora longicervia
The two fossils presented here (Figure 1) were collected from the Jianshangou Bed of the Yixian Formation at Huangbanjigou Village near Beipiao City, Liaoning Province, northeastern China (Sun et al., 1998, 2001, 2011; Yang and Wang, 2013). The geological age of the Jianshangou Bed within the Yixian Formation can be established by multiple radiometric dates to ca. 122–125 Ma (Swisher et al., 2002; Zhou et al., 2003; Chang et al., 2009) or 125–127 Ma (Sun et al., 2011), which correlates to the Early Cretaceous late Barremian–early Aptian Stages. The fossils are preserved as compressions/impressions, with minimal carbonized organic material remaining. They were photographed with Nikon D700 and Panasonic DMC-FZ30 digital cameras, and fossil overlay drawings were drawn using the software Corel-Draw 10.0 (Corel Corporation, Ottawa, ON, Canada). Voucher specimens are deposited in the Indiana University Paleontology Type Collection, Department of Geology, Indiana University, Bloomington, IN, USA.
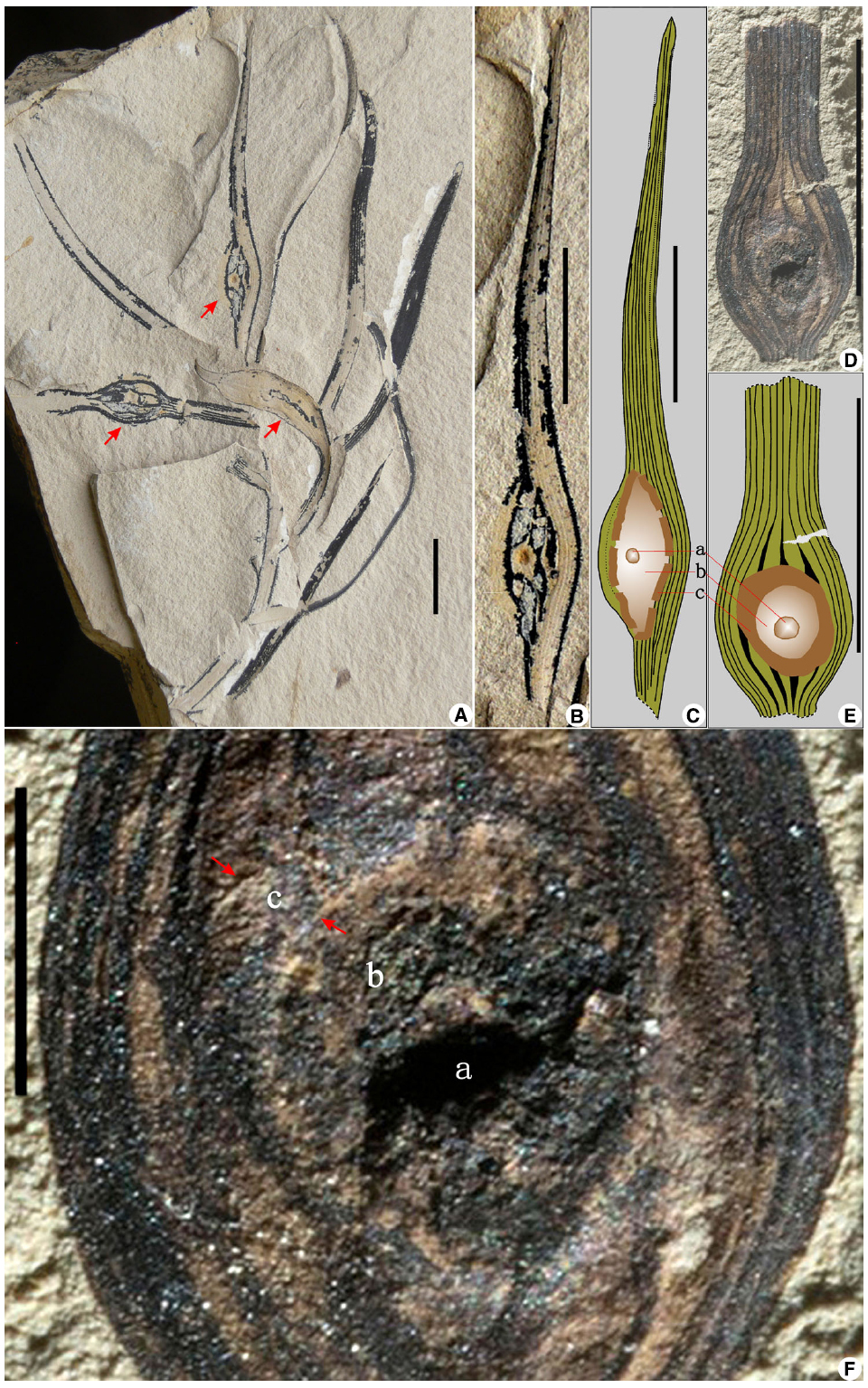
Figure 1. A leaf gall from the Early Cretaceous Yixian Formation of Liaoning, northeastern China. (A) A distal shoot with normal and galled leaves. Red arrows refer to three galls. (B) Partial enlargement of an upper gall in (A). (C) An overlay drawing of the gall in (B), showing gall features in cross-section. (D) A detached leaf fragment with a gall, showing tufts of nutritive tissue surrounding the innermost chamber and encapsulated by the outer sclerenchyma. (E) An overlay drawing of the gall in (D), with same features as in (C). (F) Partial enlargement of the gall in (D). Red arrows refer to the outermost sclerenchymatous layer of the gall. (a) Innermost larval chamber; (b) intermediate zone of gall nutritive tissue; (c) outermost sclerenchymatous layer. Scale bars = 1 cm, except in (F), which is 2 mm.
A New Perspective on Archaeamphora longicervia
The two fossils represent a distal shoot bearing spirally arranged leaves and a detached leaf fragment, respectively (Figures 1A,D). These leaves are lanceolate in form, about 50–80 × 2–6 mm in size, with nearly parallel veins, ca. 4–11 in number at the widest portions of each leaf, usually forking once near slightly constricted leaf bases, but unbranched and converging toward the acute or blunt apexes. The parallel veins on different parts of four leaves have been interrupted by a swollen, bulbous structure (ca. 4–10 × 4–5 mm in size), each of which is usually bordered by a grossly elliptical to orbicular ring zone, which often encloses a tiny, spheroidal lump or depression, about 0.8 mm in diameter (Figures 1B–F). Li (2005) interpreted such structures as pitchers of the carnivorous plant Archaeamphora longicervia. Instead, we explain these structures as foliar galls, which are abnormal swellings of plant tissues that contrast dramatically with normally developed leaves present on the shoot of the same specimens (Figure 1A). The plant host morphologically is more or less consistent with the Yixian conifer Liaoningocladus boii G. Sun et al., which bears helically arranged, narrowly lanceolate leaves (ca. 20–70 mm long by 2–5 mm wide) with a tapering to blunt apex, a somewhat contracted base, and about 4–11 in number of nearly parallel veins (Sun et al., 2000, 2001). These swollen, hemispheroidal to hemi-ellipsoidal leaf galls typically include an innermost larval chamber (Figures 1Ca,Ea,Fa) that would house the gall-inducing insect. A second, intermediately positioned zone (Figures 1Cb,Eb,Fb) is the parenchymatous nutritive tissue that is consumed by the gall-inducing insect and houses tissue with proliferative files of hyperplasic and hypertrophic cells (Labandeira and Phillips, 2002). A third major gall zone (Figures 1Cc,Ec,Fc) is the protective, hardened, sclerenchyma forming the outermost layer (Labandeira and Phillips, 2002).
Discussion
The age and place of origin of Archaeamphora are inconsistent with data retrieved from phylogenetic reconstructions of Sarraceniaceae and Ericales, respectively. Molecular clock dating, based on robust phylogenetic data, estimates an Eocene divergence time of 44–53 Ma for the Sarraceniaceae lineage (Ellison et al., 2012). This age estimate is further corroborated by fossil evidence from the sister group of Sarraceniaceae, Roridulaceae, that occur as leaves from Eocene Baltic amber dated at 35–47 Ma (Sadowski et al., 2015). Therefore, 122–127 million-year-old Cretaceous Archaeamphora is inordinately too old to be a member of the phylogenetically much younger Sarraceniaceae, or even Ericales. The Ericales is a lineage of core crown eudicots estimated at 92–102 Ma (Bell et al., 2010). By contrast, basal eudicots are considered to extend to the Barremian–Aptian boundary, corresponding to an age of 125 Ma (Sun et al., 2011; Wang et al., 2015). Also, the Clethraceae–Sarraceniaceae–Roridulaceae–Actinidiaceae clade is very likely to have originated in America (Ellison et al., 2012), rendering unlikely the assignment of Early Cretaceous Archaeamphora to the Sarraceniaceae of China.
All pitcher leaves of carnivorous plants are based upon epiascidiate leaves during ontogeny (Arber, 1941; Lloyd, 1942; Franck, 1975; Juniper et al., 1989; Fukushima et al., 2015) By contrast, the hollow, spheroidal to ellipsoidal cavities of Archaeamphora result from insect-induced galling. The “alae” of the pitcher, suggested by Li (2005), simply represent the foliar part of the uninfected gall. Li’s (2005) interpretation of different trapping “zones,” present in extant Sarraceniaceae pitchers, is untenable for the fossil leaves. A “fluid filled zone” at the bottom of the leaf interior, as suggested by Li (2005), is extremely unlikely due to the very narrow diameter (3–5 mm) of the cavity, where a constant water level would not be maintained due to capillary forces. The pitcher morphology of extant Sarraceniaceae is relatively constant at the species level. Li (2005) reported different “types of pitchers,” which he inferred to constitute different taxa that diversified before the Early Cretaceous. Instead, the variability in “pitcher” structure supports our explanation that these features result from more variable, tumor-like insect-induced growth reactions of the leaves, particularly the proliferation of nutritive tissue (Meyer, 1987). Because the position of the gall lacks constancy along the length of the leaf, the size of the “neck” part, as suggested by Li (2005), is similarly variable.
Insect-induced gall fossils provide unique opportunities for studying ecological links between first and second trophic levels in food chains, and the third trophic level of galls with parasitoid exit holes (Labandeira, 2002). The galling behavior of arthropods on plants has occurred in two mite lineages (Eriophyidae, Tetranychidae), and in the six major insect lineages Thysanoptera, Hemiptera, Coleoptera, Diptera, Lepidoptera, and Hymenoptera (Meyer, 1987; Shorthouse and Rohfritsch, 1992; Labandeira, 1998, 2002). The attacking insect is not only highly specific in selecting a particular host species, but often selects a certain host organ or tissue, and is associated with a distinctive structure, color, and physiology of the gall (Meyer, 1987; Shorthouse and Rohfritsch, 1992; Raman, 2011). The leaf galls presented here have been discovered from the host plant Liaoningocladus boii, which also hosts a beetle leaf miner (Ding et al., 2014), representing only two of the numerous associations occurring on this plant host elsewhere from the Yixian Formation (Sun et al., 2001).
Conclusion
Early Cretaceous Archaeamphora longicervia is not a carnivorous, Sarraceniaceae-like angiosperm, but represents insect-galled leaves of the fossil gymnosperm Liaoningocladus boii from the Yixian Formation of northeastern China.
Author Contributions
WW and DD designed the research plan. WW, CL, and AF wrote the manuscript. WW and DD photographed specimens. CL identified the gall and its tissues. WW drew line-drawings and arranged the figure. All authors performed analyses, commented the text and approved the final manuscript.
Conflict of Interest Statement
The reviewer Chunlin Sun declares that, despite having collaborated with the author David Leonard Dilcher, the review process was handled objectively. The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We would like to thank Dr. Zhengyuan Wang, PetroChina Refining & Chemicals R&D Center, Beijing for providing the fossil specimen shown in Figure 1A. We also thank Prof. Zhenyu Li and Mrs. Si Shen, Institute of Botany, Chinese Academy of Sciences, Beijing for helpful discussions. We are grateful to the three reviewers Profs. David Smyth, Chunlin Sun, and Tom Givnish for their helpful and constructive comments and suggestions. This work was supported by the National Natural Science Foundation of China (grant no. 41372029), and the State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences (grant no. 123106). This is contribution 309 of the Evolution of Terrestrial Ecosystems consortium at the National Museum of Natural History.
References
Arber, A. (1941). On the morphology of the pitcher-leaves in Heliamphora, Sarracenia, Darlingtonia, Cephalotus, and Nepenthes. Ann. Bot. 5, 563–578.
Bell, C. D., Soltis, D. E., and Soltis, P. S. (2010). The age and diversification of the angiosperms re-visited. Am. J. Bot. 97, 1296–1303. doi: 10.3732/ajb.0900346
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Brittnacher, J. (2013). Phylogeny and biogeography of the Sarraceniaceae. Carniv. Pl. Newslett. 42, 99–106.
Chang, S. C., Zhang, H. C., Renne, P. R., and Fang, Y. (2009). High-precision 40Ar/39Ar age for the Jehol Biota. Palaeogeogr. Palaeoclimatol. Palaeoecol. 280, 94–104. doi: 10.1016/j.palaeo.2009.06.021
Ding, Q. L., Labandeira, C. C., and Ren, D. (2014). Biology of a leaf miner (Coleoptera) on Liaoningocladus boii (Coniferales) from the Early Cretaceous of northeastern China and the leaf-mining biology of possible insect clades. Arthropod Syst. Phylo. 72, 281–308.
Ellison, A. M., Butler, E. D., Hicks, E. J., Naczi, R. F. C., Calie, P. J., Bell, C. D., et al. (2012). Phylogeny and biogeography of the carnivorous plant family Sarraceniaceae. PLoS ONE 7:e39291. doi: 10.1371/journal.pone.0039291
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Fleischmann, A. (2010). “Evolution of carnivorous plants,” in Carnivorous Plants and Their Habitats, ed. S. McPherson (Redfern: Dorset), 68–123.
Franck, D. H. (1975). Early histogenesis of the adult leaves of Darlingtonia californica (Sarraceniaceae) and its bearing on the nature of epiascidiate foliar appendages. Am. J. Bot. 62, 116–132. doi: 10.2307/2441587
Fukushima, K., Fujita, H., Yamaguchi, T., Kawaguchi, M., Tsukaya, H., and Hasebe, M. (2015). Oriented cell division shapes carnivorous pitcher leaves of Sarracenia purpurea. Nat. Commun. 6, 6450. doi: 10.1038/ncomms7450
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Givnish, T. J. (2015). New evidence on the origin of carnivorous plants. Proc. Natl. Acad. Sci. U.S.A. 112, 10–11. doi: 10.1073/pnas.1422278112
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Juniper, B. E., Robins, R. J., and Joel, D. M. (1989). The Carnivorous Plants. London: Academic Press.
Labandeira, C. C. (1998). Early history of arthropod and vascular plant associations. Annu. Rev. Earth Planet. Sci. 26, 329–377. doi: 10.1146/annurev.earth.26.1.329
Labandeira, C. C. (2002). “The history of associations between plants and animals,” in Plant–Animal Interactions: An Evolutionary Approach, eds C. M. Herrera and O. Pellmyr (London: Blackwell Science), 26–74, 248–261.
Labandeira, C. C., and Phillips, T. L. (2002). Stem borings and petiole galls from Pennsylvanian tree ferns of Illinois, USA: implications for the origin of the borer and galler functional-feeding-groups and holometabolous insects. Palaeontographica Abt. B 264, 1–84.
Li, H. Q. (2005). Early Cretaceous sarraceniacean-like pitcher plants from China. Acta Bot. Gallica 152, 227–234. doi: 10.1080/12538078.2005.10515473
Raman, A. (2011). Morphogenesis of insect-induced plant galls: facts and questions. Flora 206, 517–533. doi: 10.1016/j.flora.2010.08.004
Sadowski, E. M., Seyfullah, L. J., Sadowski, F., Fleischmann, A., Behling, H., and Schmidt, A. R. (2015). Carnivorous leaves from Baltic amber. Proc. Natl. Acad. Sci. U.S.A. 112, 190–195. doi: 10.1073/pnas.1414777111
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Schnell, D. E. (2002). Carnivorous Plants of the United States and Canada, 2nd Edn. Portland: Timber Press.
Shorthouse, J. D., and Rohfritsch, O. (1992). Biology of Insect-induced Galls. Oxford: Oxford University Press.
Sun, G., Dilcher, D. L., Wang, H. S., and Chen, Z. D. (2011). A eudicot from the Early Cretaceous of China. Nature 471, 625–628. doi: 10.1038/nature09811
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sun, G., Dilcher, D. L., Zheng, S. L., and Zhou, Z. K. (1998). In search of the first flower: a Jurassic angiosperm, Archaefructus, from Northeast China. Science 282, 1692–1695. doi: 10.1126/science.282.5394.1692
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Sun, G., Zheng, S. L., Dilcher, D. L., Wang, Y. D., and Mei, S. W. (2001). Early Angiosperms and Their Associated Plants from Western Liaoning, China. Shanghai: Shanghai Scientific and Technological Education Publishing House.
Sun, G., Zheng, S. L., and Mei, S. W. (2000). Discovery of Liaoningocladus gen. nov. from the lower part of Yixian Formation (Upper Jurassic) in western Liaoning, China. Acta Palaeontol. Sin. 39(Suppl.), 200–208.
Swisher, C. C. III, Wang, X. L., Zhou, Z. H., Wang, Y. Q., Jin F., Zhang, J. Y., et al. (2002). Further support for a Cretaceous age for the feathered dinosaur beds of Liaoning, China: new 40Ar/39Ar dating of the Yixian and Tuchengzi formations. Chin. Sci. Bull. 47, 135–138.
Wang, W., Dilcher, D. L., Sun, G., Wang, H. S., and Chen, Z. D. (2015). Accelerated evolution of angiosperms: evidence from Ranunculales phylogeny by integrating living and fossil data. J. Syst. Evol. 53, 1–7. doi: 10.1111/jse.12090
Yang, Y., and Wang, Q. (2013). The earliest fleshy cone of Ephedra from the Early Cretaceous Yixian Formation of Northeast China. PLoS ONE 8:e53652. doi: 10.1371/journal.pone.0053652
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Zhou, Z. H., Barrett, P. M., and Hilton, J. (2003). An exceptionally preserved Lower Cretaceous ecosystem. Nature 421, 807–814. doi: 10.1038/nature01420
PubMed Abstract | Full Text | CrossRef Full Text | Google Scholar
Keywords: angiosperm, Archaeamphora longicervia, carnivorous plant, Early Cretaceous, leaf gall, Liaoningocladus boii, Sarraceniaceae, Yixian Formation
Citation: Wong WO, Dilcher DL, Labandeira CC, Sun G and Fleischmann A (2015) Early Cretaceous Archaeamphora is not a carnivorous angiosperm. Front. Plant Sci. 6:326. doi: 10.3389/fpls.2015.00326
Received: 25 January 2015; Accepted: 26 April 2015;
Published: 07 May 2015.
Edited by:
Stefan De Folter, Centro de Investigación y de Estudios Avanzados del Instituto Politécnico Nacional, MexicoReviewed by:
David Smyth, Monash University, AustraliaChunlin Sun, Jilin University, China
Tom Givnish, University of Wisconsin-Madison, USA
Copyright © 2015 Wong, Dilcher, Labandeira, Sun and Fleischmann. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: William Oki Wong (Qi Wang), State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, 20 Nanxincun, Xiangshan, Haidian District, Beijing 100093, China,aGFwcHlraW5nQGliY2FzLmFjLmNu