 Iwona Żur
Iwona Żur Ewa Dubas
Ewa Dubas Monika Krzewska
Monika Krzewska Franciszek Janowiak
Franciszek Janowiak- The Franciszek Górski Institute of Plant Physiology, Polish Academy of Sciences, Kraków, Poland
Plant growth regulator (PGR) crosstalk and interaction with the plant’s genotype and environmental factors play a crucial role in microspore embryogenesis (ME), controlling microspore-derived embryo differentiation and development as well as haploid/doubled haploid plant regeneration. The complexity of the PGR network which could exist at the level of biosynthesis, distribution, gene expression or signaling pathways, renders the creation of an integrated model of ME-control crosstalk impossible at present. However, the analysis of the published data together with the results received recently with the use of modern analytical techniques brings new insights into hormonal regulation of this process. This review presents a short historical overview of the most important milestones in the recognition of hormonal requirements for effective ME in the most important crop plant species and complements it with new concepts that evolved over the last decade of ME studies.
Introduction
Plant growth regulators are known as key signaling molecules controlling plant growth and development, and initiating signal transduction pathways in response to environmental stimuli (Kohli et al., 2013). The role played by PGRs in ME has been examined widely but usually using the traditional ‘one-factor-at-a-time’ and ‘trial-and-error’ techniques. Hormonal requirements determined through such empirical methods were usually optimized for particular cultivars or genotypes. Once identified, positively acting combinations of PGRs were usually used standardly for years with small modifications introduced in the case of less responsive genotypes. Only in a few cases endogenous levels of PGRs were analyzed and taken into consideration in studies examining their influence on ME effectiveness (Dollmantel and Reinert, 1980; Delalonde and Coumans, 1998; Gorbunova et al., 2001; Lulsdorf et al., 2012). Moreover, usually only one or two groups of phytohormones were analyzed whereas there is growing evidence indicating that – as could be expected – it is the complex PGR crosstalk and its interaction with the plant’s genotype and environmental factors which controls the initiation and the course of the process. The complexity of the PGR network which could exist at the level of biosynthesis, distribution, gene expression, or signaling pathways, renders the creation of an integrated model of ME-control crosstalk impossible at present. However, the analysis of the published data together with the results received recently with the use of modern analytical techniques bring new insights into hormonal regulation of this process. New concepts that evolved over the last decade of ME studies together with a short historical review showing the most important milestones in the recognition of hormonal requirements for effective ME are presented below. The review concerns the most important crop plants, both model species, and species well-known for their recalcitrance to most in vitro approaches like oat, rye, grain legumes, and cassava (Supplementary Table S1).
Auxins and Cytokinins
Particularly important for in vitro cultures is the concerted action of auxins and cytokinins which control cell division and morphogenesis. These two hormone groups usually act antagonistically but their effects are modulated by plant genome and tissue specificity (Moubayidin et al., 2009). Various combinations of auxins and cytokinins have been used in media designated for in vitro anther culture, whereas in the majority of isolated microspore cultures exogenous PGRs were not required for ME initiation (for details see Supplementary Table S1). For several plant species instead of exogenous PGRs, co-culture with so-called immature ‘ovaries’ (accurately, pistils) is critical to sustain microspore-derived embryo development (Hul and Kasha, 1997; Li and Devaux, 2001; Zheng et al., 2002; Lantos et al., 2009). Similar or even better results could be received through the use of conditioned medium, prepared by culturing isolated ‘ovaries’ or microspores of responsive plant genotypes. In isolated microspore culture of recalcitrant wheat cultivars (Zheng et al., 2002) live ‘ovary’ co-culture alone was not effective, while the addition of medium preconditioned by ‘ovaries’ increased the yield of microspore-derived embryos more than 100-fold. Similarly, conditioned medium extracted from actively growing microspores of barley broke the recalcitrancy of isolated oat microspores and resulted in regeneration of fertile green plants (Sidhu and Davies, 2009). Despite many attempts, the effect of ‘ovary’ co-culture could not be successfully substituted by any treatment or any exogenously applied substance. It is supposed that the ‘ovaries’ are a sources of active signaling molecules that increase microspore-derived embryo yield and improve green plant regeneration. The involvement of auxin-like substances and/or arabinogalactans (Baldwin et al., 1993; Borderies et al., 2004; Letarte et al., 2006) has been postulated, but the mechanism of their influence remains unexplained.
Among auxins, the first and most widely used for ME initiation was IAA (Guha and Maheshwari, 1964). It is an essential phytohormone ubiquitous throughout the plant kingdom and involved in the regulation of a wide spectrum of physiological processes (Davies, 2010). Later on it was frequently replaced by more stable synthetic auxin analogs: 2,4-D, NAA, DIC or PIC and its combinations (for details see Supplementary Table S1; Table 1). 2,4-D is one of the most often used culture media supplements, applicable for both dicotyledonous and monocotyledonous plants (Raghavan, 2004). Its high effectiveness in the induction and maintenance of callus and suspension cultures from somatic tissues resulted in its application as ME stimulus. Other synthetic auxin analogs were also first tested in somatic tissue cultures as inducers of embryogenesis (PIC, DIC) or organogenesis (NAA; Gaspar et al., 1996). Natural auxins: IBA and PAA are used less frequently (for details see Supplementary Table S1). For decades, IBA has been used commercially for plant propagation, being more effective than IAA in stimulation of adventitious root formation. Its effectiveness can be at least partially explained by its higher stability and lower predisposition to the formation of inactive conjugates. Its possible direct involvement in ME initiation has been postulated recently (Dubas et al., 2013a). Similarly, PAA was applied mainly for stimulation of plant regeneration, its beneficial effects on androgenic plant production having been reported for wheat and barley (Ziauddin et al., 1992). In addition to acting as an active auxin, PAA may inhibit polar auxin transport (Morris and Johnson, 1987), regulating the level of free IAA. In plants, it is present at levels 10- to 100-fold lower in comparison with IAA. Due to its low activity (Normanly et al., 2010), it is usually supplemented in much higher concentrations to media used for ME (Table 1).
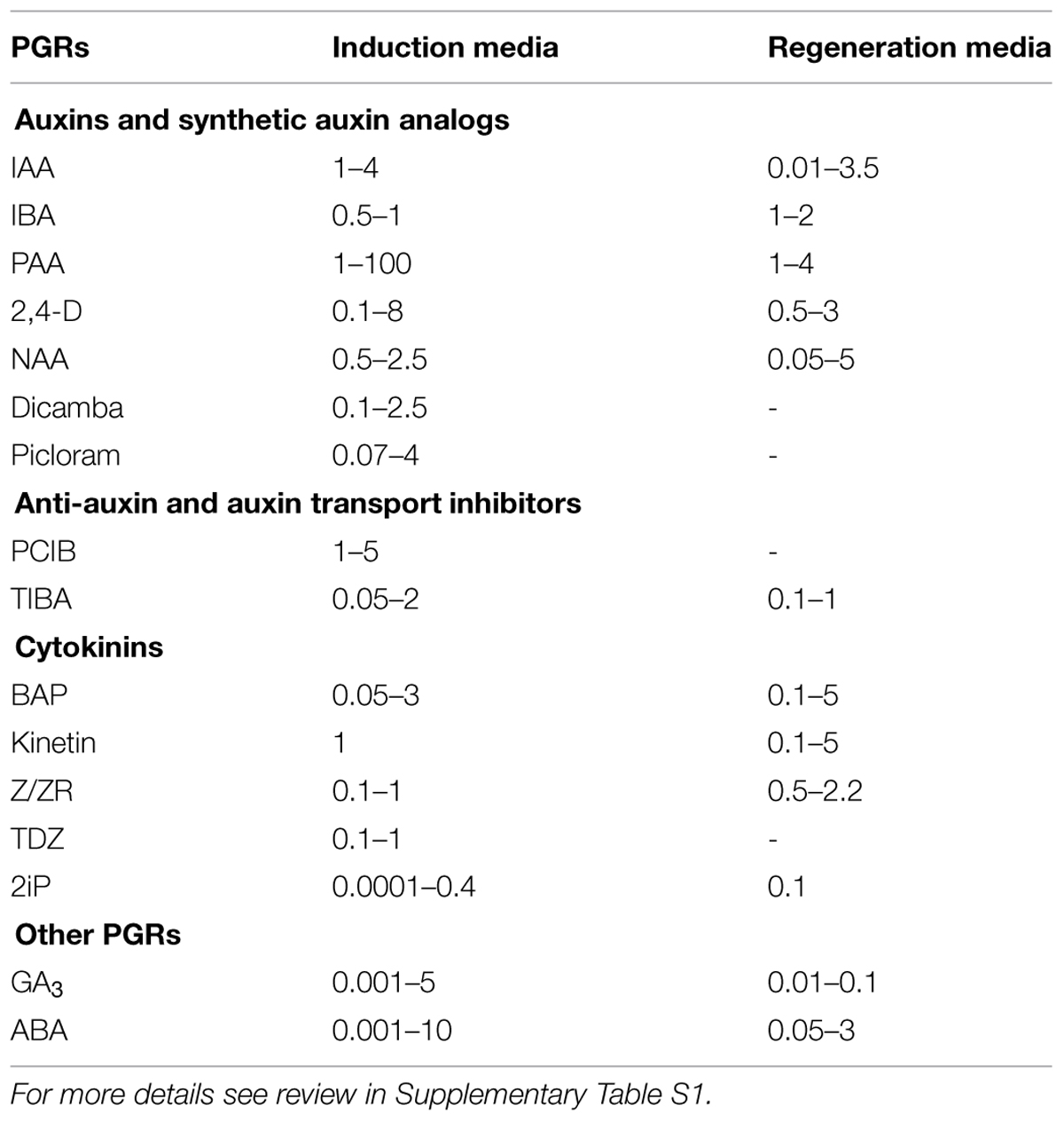
TABLE 1. The most popular PGRs and their concentration ranges [mg l-1] used standardly in media dedicated for ME induction and microspore-derived embryo regeneration.
Interestingly, besides auxins, several inhibitors of auxin biosynthesis (AZI) or auxin polar transport TIBA, NPA, or 1-NOA as well as anti-auxins OCPIB, PCIB have been used quite frequently (for details see Supplementary Table S1; Table 1). All these chemicals influence embryo development through disruption of auxin homeostasis (Lankova et al., 2010). However, depending on the plant species, type of substance and procedures used, the results were ambiguous or even contradictory. For example, preculture of Nicotiana tabacum anthers in the presence of the inhibitor of AZI and anti-auxin (OCPIB) resulted in enhanced plantlet formation (Dollmantel and Reinert, 1980). In contrast, IAA-oxidase activator, CPIBA, IAA transport inhibitor (quercetin), and IAA-oxidase inhibitor (dopamine) did not give positive results in maize anther cultures (Delalonde and Coumans, 1998). Due to structural similarity, anti-auxins (Jönsson, 1961) can compete with IAA at the binding site of its receptors (McRae and Bonner, 1953) and exhibit some antagonistic effects. Lower concentration of PCIB (20 μM) enhanced the development of microspore-derived embryos of Brassica juncea and B. napus, while higher doses were detrimental and resulted in a high frequency of morphologically abnormal embryo formation (Agarwal et al., 2006; Ahmadi et al., 2012). In B. rapa, critical ME-stimulating concentration of PCIB was twofold higher (40 μM), but similarly its overdose decreased microspore-derived embryo yield and increased the frequency of morphological abnormalities (Zhang et al., 2011). The effect of TIBA (1 μM), which conjugates specifically to the ingression site and inhibits polar transport of IAA, on barley ME was highly genotype-dependent (Cistué et al., 1999). Although it decreased the number of dividing microspores in some cultivars, a tendency to produce a higher percentage of embryos and to improve embryo quality was also observed. Higher doses (2–4 μM) of TIBA were beneficial for low responsive cultivar, increasing well developed embryo production and reducing albinism. TIBA could also affect later phases of microspore-derived embryo development. The treatment applied to B. napus cv. Topas at the preglobular/globular stages of embryo development resulted in altered shoot apical meristem development and in production of one fused cotyledon, which indicates a continuation of radial symmetry (Ramesar-Fortner and Yeung, 2006). However, tillers pre-treatment with 5 μM PCIB or 10 μM TIBA had no effect on ME induction in triticale anther culture (Żur et al., 2015). Similar to TIBA in the report of Cistué et al. (1999), PCIB stimulated plant regeneration but only in the highly recalcitrant triticale genotype. The supplementation of ME-induction medium with the same concentrations of TIBA or PCIB did not improve the efficiency of the process in the case of the recalcitrant genotype and significantly decreased the number of microspore-derived embryos produced by the responsive one (Żur et al., 2015).
The negative effect of NPA on embryogenesis was observed in microspore suspension of oak (Rodriguez-Sanz et al., 2014). This compound together with 1-NOA are potent synthetic auxin inhibitors (Lankova et al., 2010). It was proved that NPA strongly inhibits auxin efflux (Petrasek et al., 2003), whereas 1-NOA blocks both auxin influx and efflux. NPA interferes with actin dynamics being under the control of auxin itself, while 1-NOA action has been suggested to be related to the dynamics of membrane vesicle transporting auxin carriers (Titapiwatanakun and Murphy, 2009; Lankova et al., 2010).
Two major properties of cytokinins that predispose these adenine derivatives for in vitro cultures are their abilities to induce cell division and differentiation. Their effects result from co-action with auxins, but each of these PGR groups seems to control different phases of the cell cycle: auxins – DNA replication, whereas cytokinins – mitosis and cytokinesis (Gaspar et al., 1996). Among cytokinins, kinetin (N6-furfuryladenine; KN), 6-benzylaminopurine (BAP), and zeatin (Z) have been frequently tested both in ME induction and regeneration media (for details see Supplementary Table S1; Table 1). Other kinds of cytokinins, like thidiazuron (N-phenyl-N’-1,2,3-thiadiazol-5-ylurea; TDZ), N6-(2-isopentenyl)-adenine (2iP) or meta-topoline (6-3-hydroxybenzylaminopurine, mT) are less popular ingredients of culture media (Kumar et al., 2003; Grewal et al., 2009; Esteves et al., 2014). TDZ has been used successfully in vitro to induce adventitious shoot formation and to promote axillary shoot proliferation. It is particularly effective with recalcitrant woody species. However, prolonged exposure to this cytokinin may cause problems such as hyperhydricity and abnormal shoot or root development (Lu, 1993). 2iP was used for ME initiation in anther culture of Cicer arietium (Grewal et al., 2009), whereas regeneration medium supplemented with mT proved beneficial for green plants production in microspore culture of barley (Esteves et al., 2014).
Abscisic Acid
Besides auxins and cytokinins, ABA, known as a ubiquitous plant stress hormone, has been claimed to play a role in ME-inducing signal transduction system (Maraschin et al., 2005; Żur et al., 2012, 2015; Dubas et al., 2013b; Ahmadi et al., 2014). It is well documented that plant cells and tissues usually increase their ABA level in response to different biotic and abiotic stresses (Zeevaart and Creelman, 1988; Christmann et al., 2004; Cutler et al., 2010). During ME induction various stress conditions (e.g., starvation, cold, osmotic stresses) have been commonly used as a trigger of microspore switch toward sporophytic development pathway (Touraev et al., 1997; Zoriniants et al., 2005). ABA level increases in tissues and microspores exposed to these stresses and many reports have suggested a causal involvement of ABA in ME induction, describing the positive influence of ABA accumulation on the effectiveness of this process (Reynolds and Crawford, 1996; van Bergen et al., 1999; Wang et al., 1999; Żur et al., 2008, 2012). Furthermore, a positive relationship has been shown to exist between higher regeneration efficiency and increased endogenous ABA level during ME induction by osmotic stress in barley (Hoekstra et al., 1997; van Bergen et al., 1999). These positive ABA effects on ME have been confirmed in several manipulative experiments with a treatment with exogenous ABA or its inhibitor fluridone (Imamura and Harada, 1980; Reynolds and Crawford, 1996; Wang et al., 1999; Guzman and Arias, 2000). In Hordeum species the addition of 10-7 M ABA enhanced the regeneration of plants at sub-optimal anther pre-treatment conditions, while ABA-biosynthesis inhibitor fluridone strongly reduced regeneration efficiency, particularly green plant production (Hoekstra et al., 1997; van Bergen et al., 1999). In another study of rapeseed, ME efficiency was improved by exogenous ABA treatment – 0.5 mg ABA l-1 for 12 h enhanced ME threefold compared with untreated cultures and increased normal plantlet regeneration by 68% (Ahmadi et al., 2014). In turn Żur et al. (2008, 2012) observed a significant endogenous ABA increase during ME induction by low temperature stress in triticale. However, there was no linear relationship between the extent of ABA accumulation and ME efficiency in the population of 72 triticale DH lines (Żur et al., 2012). On the contrary, higher level of endogenous ABA significantly diminished green plant regeneration efficiency. Therefore, it seems that the induction of ME requires a certain genotype-specific threshold level of ABA, which initiates a signaling cascade switching the program of embryogenic development. Moreover, it seems that a specific PGRs homeostasis and auxins/cytokinins/ABA crosstalk is a more important prerequisite for effective ME than the level of individual PGRs (Żur et al., 2015). It has also been discovered that microspores’ membrane fluidity may indirectly affect the level of ABA accumulation within the cell (Dubas et al., 2013b). Those findings verified the hypothesis about the influence of ABA on ME induction in rapeseed and pointed out that increased ABA concentration (to about 2.1 μM) in heat-treated microspores enhanced ME. Altogether, the role of ABA in microspore reprogramming is complex – it acts as a common anti-stress factor increasing microspores’ viability during ME induction, and on the other hand, ABA-induced signaling cascade plays a vital role in the activation of many genes (mainly controlling the synthesis of LEA proteins), in the activity of enzymes and in the ox-redox status as well as interacts with other PGRs (Maraschin et al., 2005; Żur et al., 2012, 2014, 2015; Ahmadi et al., 2014).
Other Plant Growth Regulators
Despite many studies on microspore and anther culture in crop species, the effects of phytohormones such as gibberellins (GAs), brassinosteroids (BRs), jasmonic acid (JA), salicylic acid (SA), or ethylene on ME are not fully recognized.
GAs are involved in a wide range of developmental responses (Moshkov et al., 2008). They are required for normal pollen, anther and seed development, and are probably involved in a broad spectrum of responses to abiotic stress (Colebrook et al., 2014), but a complete understanding of their specific function remains elusive (Swain and Singh, 2005). Only scarce information is available on GAs in cultured cells. Plant tissue cultures can generally be induced to grow and differentiate without GAs. One of the most bioactive forms, GA3 is generally used in plant tissue to stimulate stem elongation. It was also supposed to be an essential ingredient of media for culturing cells at low densities (Stuart and Street, 1971). In microspore cultures of B. napus and Solanum tuberosum, GA3 improved plantlet regeneration (Supplementary Table S1), mainly via elongation of the embryo axis and acceleration of its maturation (Haddadi et al., 2008). Similarly Ahmadi et al. (2012) reported that the highest percentage of normal B. napus plantlet regeneration (40%) was received as a result of 0.05–0.1 mg l-1 GA3 treatment. More attention has been attracted by a wide range of synthetic substances, called ‘anti-gibberellins,’ which block GAs biosynthetic pathways. In the studies of Biddington et al. (1992), the addition of paclobutrazol into induction media inhibited embryo production in anther cultures of Brussels sprout. However, the authors suggested that this effect could be caused not only by inhibition of GA-biosynthesis but also by inhibition of sterol biosynthesis. Another inhibitor of GA-biosynthesis, uniconazole, applied to B. napus embryo at the globular stage of development significantly reduced axis elongation (Hays et al., 2002).
Brassinosteroids (BRs) are a class of plant steroidal hormones that regulate multiple developmental and physiological processes essential for plant growth and development. Their involvement in cell elongation and division, vascular differentiation, senescence, flowering time, male fertility, pollen development, seed size, photomorphogenesis, and resistance to biotic and abiotic stresses has been reported (Clouse et al., 1996; Li and Chory, 1999; Ye et al., 2010; Clouse, 2011). BRs, in particular 24-epibrassinolide (EBr), increased frequency of induction of both somatic (Azpeitia et al., 2003; Pullman et al., 2003) and ME (Ferrie et al., 2005; Malik et al., 2008). The role of EBr may be related to protection against abiotic stresses as its positive impact on the acquisition of thermotolerance was reported (Divi et al., 2010). Other BR, brassinolide (BL) also enhanced embryogenesis and the quality of microspore-derived embryos in B. napus and B. juncea (Ferrie et al., 2005; Belmonte et al., 2010). The addition of BRs did not affect plant regeneration but seems to influence chromosome doubling. Moreover, depletion of cellular BL decreases microspore-derived embryo production and disrupts the architecture of the apical meristems of B. napus (Belmonte et al., 2010).
As ME is induced by stress treatment it could be supposed that not only ABA, but also other stress hormones like jasmonic acid (JA), salicylic acid (SA), or ethylene can be involved in this process.
JA is widely distributed in the plant kingdom and regulates a wide range of processes from growth and photosynthesis to reproductive development. The most important is the role connected with plant defense reactions against biotic and abiotic stresses (Santino et al., 2013). In anther cultures of barley, ME-induction treatment resulted in higher expression of three genes encoding enzymes involved in JA biosynthesis (Jacquard et al., 2009). Ahmadi et al. (2014) claimed that the supplementation of induction medium with 1.0 mg l-1 JA for 24 h improved embryo yield in microspore cultures of B. napus. Moreover, the addition of 0.5 mg l-1 JA for 12 h resulted in better plantlet regeneration.
SA, a plant phenolic derivative, is now considered to be a hormone-like endogenous regulator and its role in the defense mechanisms against biotic and abiotic stress is well documented (Catinot et al., 2008). Being a mobile molecule, SA is capable of acting as a cell signal that senses, amplifies, and transmits information initiating the embryogenic program (Mulgund et al., 2012). There are several papers that describe the application of SA to culture media in order to improve ME efficiency. In the above-mentioned work, Ahmadi et al. (2014) reported a positive effect of short-term application (6 h) of 0.2 and 0.5 mg l-1 of SA on B. napus microspore-derived embryo yield. The mechanism of SA action could be connected with its ability to increase the activity of superoxide dismutase (H2O2-producing enzyme), and to inhibit ascorbate peroxidase and catalase activities (H2O2-decomposing enzymes), thus leading to endogenous H2O2 accumulation, which is supposed to initiate ME (Larqué-Saavedra, 1978, 1979; Leslie and Romani, 1988; Luo et al., 2001; Żur et al., 2014).
Ethylene is a gaseous plant hormone involved in many developmental processes – seed germination, root development, flower senescence, abscission, and fruit ripening (Kumar et al., 2009). Its biosynthesis is tightly regulated by internal signals and environmental stresses, like wounding, low temperature, hypoxia, or pathogen attack (Wang et al., 2002). Its role in in vitro callus growth, organo- and embryogenesis has been suggested (Kumar et al., 2009). It was reported that embryogenesis in barley can be stimulated by both promoters and antagonists of ethylene, depending on the genotype (Cho and Kasha, 1989). It suggests that the response depends upon how much ethylene is being produced and that an optimum level of ethylene is required for ME initiation. More often, positive effects induced by substances known as inhibitors of ethylene action – silver nitrate (Prem et al., 2005), activated charcoal (Prem et al., 2008), AVG or cobalt chloride (Leroux et al., 2009) were observed. On the other hand, there is also evidence reporting benefits from ethylene precursor ACC or promoter ETP. Their application increased ME initiation in anther culture of barley (Evans and Batty, 1994) and oat (Kiviharju et al., 2005).
New Concepts Describing PGRs Involvement in ME Regulation
New Kinds of PGRs Possibly Involved in ME Regulation
Although IBA is commonly considered to be only an IAA precursor and storage form (Woodward and Bartel, 2005; Korasick et al., 2013), some evidence suggests that it could act directly as an active auxin (Ludwig-Muller, 2000; Poupart and Waddell, 2000; Zolman et al., 2000). Recent results shown by Dubas et al. (2013a) suggest that the increased level of IBA in B. napus microspores under heat shock treatment might be used as a marker of cell embryogenic competence. However, IBA accumulation was not sufficient for ME initiation in the case of recalcitrant genotypes. Similar results were received in anther cultures of triticale (Żur et al., 2015), where higher concentration of IBA seems to be advantageous for effective ME induction. However, because IBA pool in triticale anthers comprises only about 1% of the total auxins content, it is questionable whether differences in its concentration are of any significance. In the same report, trans and cis isomers of tZ, cZ and tZR, cZR were detected in anthers of eight DH lines of triticale. Interestingly, cZ commonly regarded as cytokinin derivative without any or with low biological activity, prevailed significantly and positively correlated with ME induction. Similarly, in reports of Emery et al. (1998), Vyroubalová et al. (2009), and Kudo et al. (2012) cZ appeared to be the dominant form of cytokinins in specific plant organs and/or stages of development. In a recently published report, Gajdošová et al. (2011) concluded that cZ can be qualified as a regulator of cytokinin responses in plants under growth-limiting conditions. Another finding of Żur et al. (2015) was a relatively high concentration of KN-like compound and its negative correlation with ME efficiency. KN was the first compound identified as cytokinin, but for many years it was classified as a product of DNA rearrangement not produced by plant cells. This opinion started to change in the last decade, as sources of KN in biological samples were found in cellular DNA, plant tissues and extracts (Barciszewski et al., 1996, 1999, 2007; Ge et al., 2005). The role of endogenous KN and the molecular mechanisms of its action are not well understood, although some data indicate its strong antioxidant properties and some ABA- and JA-antagonistic effects (Barciszewski et al., 2000).
High Concentrations of PGRs as a Stress Factor
Higher concentration (5–10 mg l-1) of 2,4-D stimulated ME initiation in some recalcitrant plant species, namely oat (Kiviharju and Tauriainen, 1999), Triticum turgidum (Jauhar, 2003), and cassava (Perera et al., 2014). It has been suggested that 2,4-D is not only an auxin analog but at higher concentrations acts as a stress factor effectively triggering embryogenic pathway of cell development (Gaj, 2004). The observed effect is probably the result of concerted PGRs action as evidence that 2,4-D regulates the activity of genes associated with auxin, ABA and ethylene biosynthesis has been reported (Raghavan et al., 2006). Short-term treatment with extremely high concentration of this substance (15–45 mg l-1 2,4-D for 15–45 min) has been recently used as an effective substitute of classical heat shock treatment (Ardebili et al., 2011) for B. napus ME initiation. It has been proposed as an alternative for plant species whose microspores are extremely sensitive to classical stresses.
Endogenous Level of PGRs and their Interaction with their Exogenously Applied Analogs
Inconsistent or even contradictory effects of various PGRs and their inhibitors suggest that the endogenous level of natural phytohormones and its balance with exogenously applied ones can be crucial both for yield and quality of microspore-derived embryos.
The first report pointing out that endogenous auxins level can determine anther culture responsiveness was published as early as by Dollmantel and Reinert (1980). Next, the results published by Gorbunova et al. (2001) indicated that wheat genotypes with high endogenous IAA content required lower concentration of 2,4-D in induction medium. Other data showing that anti-auxin (PCIB) and auxin transport inhibitor (TIBA) can stimulate microspore-derived embryo formation, probably due to overcoming the inhibitory effect of high auxin concentration, were published by Cistué et al. (1999) and Agarwal et al. (2006). Also in the case of triticale (Żur et al., 2015), for which ME-induction medium was supplemented with 1 mg l-1 DIC, 1 mg l-1 PIC, and 0.5 mg l-1 KN, anther cultures of responsive DH lines were characterized by significantly lower endogenous/exogenous auxins ratio in comparison to recalcitrant genotypes. In the same cultures, higher embryogenic potential was associated with significantly higher endogenous/exogenous cytokinins ratio.
Generally, exogenous PGRs are not required for ME in Brassica species. However, too low levels of endogenous auxins and/or cytokinins could disturb the proper course of the process, especially the transition from the radial to the bilateral microspore-derived embryo symmetry, as it was observed in B. napus (Ramesar-Fortner and Yeung, 2006). Similarly, the addition of 0.1–0.3 mg l-1 BAP significantly improved microspore-derived embryo yield in several B. rapa subspecies (Takahashi et al., 2012). Recently, Prem et al. (2012) and Dubas et al. (2014) showed how endogenous auxin distribution influenced embryo development in microspore suspension of B. napus. Precise endogenous auxin estimation in transgenic DR5rev::GFP or DR5::GUS microspores of highly embryogenic spring rape line (Dubas et al., 2014) revealed IAA concentration at 1.01 μM in microspores under ME-initiating heat treatment (1 day at 32°C). It could be supposed that such IAA concentration is optimal for further embryo development.
Crosstalk of Various PGRs and their Interaction with Stress Treatment and Plant Genotype in ME Initiation
A number of reviews highlighting phytohormone crosstalk in plant growth, development and response to abiotic and biotic stresses have been published recently (Depuydt and Hardtke, 2011; Hou et al., 2013; Kohli et al., 2013; Lyons et al., 2013; Wang et al., 2013). It is also well known that altered homeostasis of PGRs is one of the most dynamic changes in response to stress conditions (Kohli et al., 2013). As tissue/cell sensitivity to PGRs also changes during plant development in response to environmental or genetically coded changes (Davies, 2010), it could be supposed that interactions between PGRs, stress-induced responses and genotype-specific PGRs sensitivity coordinate microspore reprogramming and regulate the final efficiency of ME. The analysis of recent findings obtained with the use of modern analytical techniques brought some new insights into hormonal regulation of microspore reprogramming and the initiation of embryogenic development.
The results of extensive analysis of phytohormone content changes after exposure to various ME-inducing stress treatments in anthers of three highly recalcitrant legume species were presented by Lulsdorf et al. (2012). It was revealed that the most common response was increased level of IAA-asparagine, a putative IAA metabolite. Of the various cytokinins, only cZR increased after the application of stressors.
In B. napus, the level of various auxin forms depends significantly on the sample source (leaves, flower buds, isolated microspores) and temperature regimes during the growth of donor plants (10°C/18°C). For the first time Dubas et al. (2012, 2013a) showed that IBA prevailed in isolated microspores and its level could be reduced by low temperature. Interestingly, the combination of low temperature and heat shock reversed this effect. IAA level tends to change in a similar manner to IBA, both in responsive and recalcitrant genotypes. Based on these data, it could be concluded that noticeable changes in the level of both auxins forms caused by stress treatments are important for ME.
Similarly, low temperature ME-initiating treatment (3 weeks at 4°C) meaningfully changed PGRs homeostasis in several DH lines of triticale (Żur et al., 2015). Accumulation of IAA, IBA, cZ, cZR, and ABA together with a decrease in tZ content was observed in all studied genotypes. It was discovered that as result of cold treatment anthers of highly responsive triticale genotypes were characterized by higher concentrations of IBA, cZ, tZ, cZR, and lower amount of IAA and KN-like compound in comparison with recalcitrant ones. However, the effects of exogenously applied ABA, PCIB and TIBA suggest that none of the studied PGRs acts alone in the determination of embryogenic competency. An important prerequisite for effective ME seems to be a specific PGR homeostasis – lower auxin rate in comparison with cytokinins and ABA, and lower cytokinin/ABA ratio.
Genetically or/and environmentally determined changes in PGR sensitivity at least partially explain the importance of the timing of hormonal treatment/application. For example, Wassom et al. (2001) showed that modification of maize anther culture medium with various PGRs (ABA, GA3, ancymidol, or fluridone) was ineffective in comparison to donor plant treatments, where these substances were pipetted into whorls of field-grown plants 3 days before tassel harvest. Similarly, Liu et al. (2002a,b) and Zheng et al. (2003) demonstrated stimulation of ME in wheat by a combination of high temperature tillers pre-treatment with their ‘inducer chemical formulation.’ It seems that altered PGR homeostasis preceding microspore isolation and transfer to in vitro culture triggers changes important for effective ME initiation.
Summary
The data presented above indicate why standard culture medium optimization using the traditional ‘one-factor-at-a-time’ and ‘trial-and-error’ techniques, which require a considerable amount of time and effort, can sometimes be completely ineffective. As a large number of important crop species, cultivars or genotypes still remain highly recalcitrant to ME, only a much more precise recognition of molecular/physiological/metabolomical background that favors the initiation of embryogenic development could bring any substantial progress. As hormonal homeostasis seems to be one of the most important factors determining cell embryogenic competency, only a more comprehensive approach leading to the recognition of the mechanisms controlling the process could break the barrier of ME recalcitrancy.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fpls.2015.00424/abstract
Abbreviations
ABA, abscisic acid; ACC, 1-aminocyclopropane-l-carboxylic acid; AVG, aminoethoxyvinylglycine; AZI, 7-azaindole; BAP, 6-benzylaminopurine; BL, brassinolide; BR, brassinosteroid; CPIBA, chlorophenoxyisobutyric acid; DIC, 3,6-dichloro-2-methoxybenzoic acid (dicamba); 2,4-D, 2,4-dichlorophenoxyacetic acid; EBr, 4-epibrassinolide; ETP, Ethephon; GAs, gibberellins; GA3, gibberellic acid; IAA, indole-3-acetic acid; IBA, indole-3-butyric acid; 2iP, N6-(2-isopentenyl)-adenine; JA, jasmonic acid; ME, microspore embryogenesis; NAA, naphthalene-1-acetic acid; 1-NOA, 1-naphthoxyacetic acid; NPA, N-1-naphthylphthalamic acid; OCPIB, o-chlorophenoxy-isobutyric acid; PAA, phenylacetic acid; PCIB, p-chlorophenoxyisobutyric acid; PGR(s), plant growth regulator(s); PIC, 4-amino-3,5,6-trichloropicolinic acid (picloram); TDZ, N-phenyl-N′-1,2,3-thiadiazol-5-ylurea (thidiazuron); mT, 6-3-hydroxybenzylaminopurine (meta-topoline); KN, N6-furfuryladenine (kinetin); SA, salicylic acid; TIBA, 2,3,5-triiodobenzoic acid; Z, zeatin; ZR, zeatin riboside.
References
Agarwal, P., Agarwal, P., Custers, J. M., Liu, C.-M., and Bhojwani, S. S. (2006). PCIB an antiauxin enhances microspore embryogenesis in microspore culture of Brassica juncea. Plant Cell Tissue Organ Cult. 86, 201–210. doi: 10.1007/s11240-006-9108-0
Ahmadi, B., Alizadeh, K., and Teixeira da Silva, J. A. (2012). Enhanced regeneration of haploid plantlets from microspores of Brassica napus L. using bleomycin, PCIB, and phytohormones. Plant Cell Tissue Organ Cult. 109, 525–533. doi: 10.1007/s11240-012-0119-8
Ahmadi, B., Shariatpanahi, M. E., and Teixeira da Silva, J. A. (2014). Efficient induction of microspore embryogenesis using abscisic acid, jasmonic acid and salicylic acid in Brassica napus L. Plant Cell Tissue Organ Cult. 116, 343–351. doi: 10.1007/s11240-013-0408-x
Ardebili, S. H., Shariatpanahi, M. E., Amiri, R., Emamifar, M., Oroojloo, M., Nematzadeh, G., et al. (2011). Effect of 2,4-D as a novel inducer of embryogenesis in microspores of Brassica napus L. Czech J. Genet. Plant Breed. 47, 114–122.
Azpeitia, A., Chan, J. L., Saenz, L., and Oropeza, C. (2003). Effect of 22(S),23(S)-homobrassinolide on somatic embryogenesis in plumule explants of Cocos nucifera (L.) cultured in vitro. J. Horticul. Sci. Biotechnol. 78, 591–596.
Baldwin, T. C., Mccann, M. C., and Roberts, K. (1993). A novel hydroxyproline-deficient arabinogalactan protein secreted by suspension-cultured cells of Daucus carota (purification and partial characterization). Plant Physiol. 103, 115–123.
Barciszewski, J., Massino, F., and Clark, B. F. C. (2007). Kinetin – A multiactive molecule. Int. J. Biol. Macromol. 40, 182–192. doi: 10.1016/j.ijbiomac.2006.06.024
Barciszewski, J., Rattan, S. I. S., Siboska, G., and Clark, B. F. C. (1999). Kinetin — 45 years on. Plant Sci. 148, 37–45. doi: 10.1016/S0168-9452(99)00116-118
Barciszewski, J., Siboska, G. E., Pedersen, B. O., Clark, B. F. C., and Rattan, S. I. S. (1996). Evidence for the presence of kinetin in DNA and cell extracts. FEBS Lett. 393, 197–200. doi: 10.1016/0014-5793(96)00884-8
Barciszewski, J., Siboska, G., Rattan, S. I. S., and Clark, B. F. C. (2000). Occurrence, biosynthesis and properties of kinetin (N6-furfuryladenine). Plant Growth Regul. 32, 257–265. doi: 10.1023/A:1010772421545
Belmonte, M., Elhiti, M., Waldner, B., and Stasolla, C. (2010). Depletion of cellular brassinolide decreases embryo production and disrupts the architecture of the apical meristems in Brassica napus microspore-derived embryos. J. Exp. Bot. 61, 2779–2794. doi: 10.1093/jxb/erq110
Biddington, N. L., Sutherland, R. A., and Robinson, H. T. (1992). The effects of gibberellic acid, fluridone, abscisic acid and paclobutrazol on anther culture of brussels sprouts. Plant Growth Regul. 11, 81–84. doi: 10.1007/bf00024438
Borderies, G., Le Béchec, M., Rossignol, M., Lafitte, C., Le Deunff, E., Beckert, M., et al. (2004). Characterization of proteins secreted during maize microspore culture: arabinogalactan proteins (AGPs) stimulate embryo development. Eur. J. Cell Biol. 83, 205–212. doi: 10.1078/0171-9335-00378
Catinot, J., Buchala, A., Abou-Mansour, E., and Metraux, J.-P. (2008). Salicylic acid production in response to biotic and abiotic stress depends on isochorismate in Nicotiana benthamiana. FEBS Lett. 582, 473–478. doi: 10.1016/j.febslet.2007.12.039
Cho, U. H., and Kasha, K. J. (1989). Ethylene production and embryogenesis from anther cultures of barley (Hordeum vulgare). Plant Cell Rep. 8, 415–417. doi: 10.1007/BF00270082
Christmann, A., Grill, E., and Meinhard, M. (2004). “Abscisic acid signalling,” in Plant Responses to Abiotic Stress. Topics in Current Genetics, eds H. Hirt and K. Shinozaki (Berlin: Springer), 39–71.
Cistué, L., Ramos, A., and Castillo, A. M. (1999). Influence of anther pretreatment and culture medium composition on the production of barley doubled haploids from model and low responding cultivars. Plant Cell Tissue and Organ Cult. 55, 159–166. doi: 10.1023/a:1006130028396
Clouse, S. D. (2011). Brassinosteroid signal transduction: from receptor kinase activation to transcriptional networks regulating plant development. Plant Cell 23, 1219–1230. doi: 10.1105/tpc.111.084475
Clouse, S. D., Langford, M., and Mcmorris, T. C. (1996). A brassinosteroid-insensitive mutant in Arabidopsis thaliana exhibits multiple defects in growth and development. Plant Physiol. 111, 671–678. doi: 10.1104/pp.111.3.671
Colebrook, E. H., Thomas, S. G., Phillips, A. L., and Hedden, P. (2014). The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 217, 67–75. doi: 10.1242/jeb.089938
Cutler, S. R., Rodriguez, P. L., Finkelstein, R. R., and Abrams, S. R. (2010). Abscisic acid: emergence of a core signaling network. Annu. Rev. Plant Biol. 61, 651–679. doi: 10.1146/annurev-arplant-042809-112122
Davies, P. (2010). “The plant hormones: their nature, occurrence, and functions,” in Plant Hormones, ed. P. Davies (Dordrecht: Springer), 1–15. doi: 10.1007/978-1-4020-2686-7_1
Delalonde, M., and Coumans, M. (1998). Effect of IAA content modulators on peroxidase activity and on endogenous IAA during cold pretreatment of maize anthers prior to androgenesis. Plant Growth Regul. 26, 123–130. doi: 10.1023/A:1006131620264
Depuydt, S., and Hardtke, C. S. (2011). Hormone signalling crosstalk in plant growth review regulation. Curr. Biol. 21, 365–373. doi: 10.1016/j.cub.2011.03.013
Divi, U. K., Rahman, T., and Krishna, P. (2010). Brassinosteroid-mediated stress tolerance in Arabidopsis shows interactions with abscisic acid, ethylene and salicylic acid pathways. BMC Plant Biol. 10:151. doi: 10.1186/1471-2229-10-151
Dollmantel, H. J., and Reinert, J. (1980). Auxin levels, antiauxin(s) and androgenic plantlet formation in isolated pollen cultures of Nicotiana tabacum. Protoplasma 103, 155–162. doi: 10.1007/BF01276672
Dubas, E., Benkova, E., Janowiak, F., Waligórski, P., Dziurka, M., Krzewska, M., et al. (2013a). “Endogenous auxin and ABA in microspore embryogenesis of oilseed rape (Brassica napus L.),” in Proceedings of Conference European Frontiers of Plant Reproduction Research, 2-4 October 2013, Oslo, p.79.
Dubas, E., Janowiak, F., Krzewska, M., Hura, T., and Żur, I. (2013b). Endogenous ABA concentration and cytoplasmic membrane fluidity in microspores of oilseed rape (Brassica napus L.) genotypes differing in responsiveness to androgenesis induction. Plant Cell Rep. 32, 1465–1475. doi: 10.1007/s00299-013-1458-6
Dubas, E., Benkova, E., Waligorski, P., Dziurka, M., and Żur, I. (2012). “The involvement of endogenous auxins in androgenesis of Brassica napus,” in Proceedings of XXX Conference on Embryology “Plants, Animals, Humans”, Jurata.
Dubas, E., Moravčíková, J., Libantová, J., Matušíková, I., Benková, E., Żur, I., et al. (2014). The influence of heat stress on auxin distribution in transgenic B. napus microspores and microspore-derived embryos. Protoplasma 251, 1077–1087. doi: 10.1007/s00709-014-0616-1
Emery, R. J. N., Leport, L., Barton, J. E., Turner, N. C., and Atkins, C. A. (1998). cis-Isomers of cytokinins predominate in chickpea seeds throughout their development. Plant Physiol. 117, 1515–1523. doi: 10.1104/pp.117.4.1515
Esteves, P., Clermont, I., Marchand, S., and Belzile, F. (2014). Improving the efficiency of isolated microspore culture in six-row spring barley: II-exploring novel growth regulators to maximize embryogenesis and reduce albinism. Plant Cell Rep. 33, 871–879. doi: 10.1007/s00299-014-1563-1
Evans, J., and Batty, N. (1994). Ethylene precursors and antagonists increase embryogenesis of Hordeum vulgare L. anther culture. Plant Cell Rep. 13, 676–678. doi: 10.1007/BF00231622
Ferrie, A. M. R., Dirpaul, J., Krishna, P., Krochko, J., and Keller, W. A. (2005). Effects of brassinosteroids on microspore embryogenesis in Brassica species. In Vitro Cell. Dev. Biol. Plant 41, 742–745. doi: 10.1079/ivp2005690
Gaj, M. (2004). Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 43, 27–47. doi: 10.1023/B:GROW.0000038275.29262.fb
GajdoŠová, S., Motyka, V., Hoyerova, K., Dobrev, P. I., and Kaminek, M. (2011). cis-zeatin type cytokinins and their function under growth limiting conditions. Febs J. 278, 313.
Gaspar, T., Kevers, C., Penel, C., Greppin, H., Reid, D. M., and Thorpe, T. A. (1996). Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell. Dev. Biol. Plant 32, 272–289. doi: 10.1007/BF02822700
Ge, L., Yong, J. W. H., Goh, N. K., Chia, L. S., Tan, S. N., and Ong, E. S. (2005). Identification of kinetin and kinetin riboside in coconut (Cocos nucifera L.) water using a combined approach of liquid chromatography-tandem mass spectrometry, high performance liquid chromatography and capillary electrophoresis. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 829, 26–34. doi: 10.1016/j.jchromb.2005.09.026
Gorbunova, V. Y., Kruglova, N. N., and Abramov, S. N. (2001). The induction of androgenesis in vitro in spring soft wheat. balance of exogenous and endogenous phytohormones. Biol. Bull. 28, 25–30. doi: 10.1023/A:1026602603527
Grewal, R., Lulsdorf, M., Croser, J., Ochatt, S., Vandenberg, A., and Warkentin, T. (2009). Doubled-haploid production in chickpea (Cicer arietinum L.): role of stress treatments. Plant Cell Rep. 28, 1289–1299. doi: 10.1007/s00299-009-0731-1
Guha, S., and Maheshwari, S. C. (1964). In vitro production of embryos from anthers of Datura. Nature 204, 497. doi: 10.1038/204497a0
Guzman, M., and Arias, F. J. Z. (2000). Increasing anther culture efficiency in rice (Oryza sativa L.) using anthers from ratooned plants. Plant Sci. 151, 107–114. doi: 10.1016/s0168-9452(99)00204-6
Haddadi, P., Moieni, A., Karimadeh, G., and Abdollahi, M. R. (2008). Effects of gibberellin, abscisic acid and embryo desiccation on normal plantlet regeneration, secondary embryogenesis and callogenesis in microspore culture of Brassica napus L. cv. PF(704). Int. J. Plant Prod. 2, 153–162.
Hays, D. B., Yeung, E. C., and Pharis, R. P. (2002). The role of gibberellins in embryo axis development. J. Exp. Bot. 53, 1747–1751. doi: 10.1093/jxb/erf017
Hoekstra, S., Vanbergen, S., Vanbrouwershaven, I. R., Schilperoort, R. A., and Wang, M. (1997). Androgenesis in Hordeum vulgare L: effects of mannitol, calcium and abscisic acid on anther pretreatment. Plant Sci. 126, 211–218. doi: 10.1016/s0168-9452(97)00096-4
Hou, X., Ding, L., and Yu, H. (2013). Crosstalk between GA and JA signaling mediates plant growth and defense. Plant Cell Rep. 32, 1067–1074. doi: 10.1007/s00299-013-1423-4
Hul, T., and Kasha, K. J. (1997). Improvement of isolated microspore culture of wheat (Triticum aestivum L.) through ovary co-culture. Plant Cell Rep. 16, 520–525. doi: 10.1007/BF01142316
Imamura, J., and Harada, H. (1980). Effects of abscisic acid and water-stress on the embryo and plantlet formation in anther culture of Nicotiana tabacum cv Samsun. Z. Pflanzenphysiol. 100, 285–289. doi: 10.1016/S0044-328X(80)80232-7
Jacquard, C., Mazeyrat-Gourbeyre, F., Devaux, P., Boutilier, K., Baillieul, F., and Clement, C. (2009). Microspore embryogenesis in barley: anther pre-treatment stimulates plant defence gene expression. Planta 229, 393–402. doi: 10.1007/s00425-008-0838-6
Jauhar, P. P. (2003). Formation of 2n gametes in durum wheat haploids: sexual polyploidization. Euphytica 133, 81–94. doi: 10.1023/a:1025692422665
Jönsson, A. (1961). “Chemical structure and growth activity of auxin and antiauxins,” in Encyclopedia of Plant Physiology, ed. W. Ruhland (Berlin: Springer), 959–1006.
Kiviharju, E., Moisander, S., and Laurila, J. (2005). Improved green plant regeneration rates from oat anther culture and the agronomic performance of some DH lines. Plant Cell Tissue Organ Cult. 81, 1–9. doi: 10.1007/s11240-004-1560-0
Kiviharju, E. M., and Tauriainen, A. A. (1999). 2,4-Dichlorophenoxyacetic acid and kinetin in anther culture of cultivated and wild oats and their interspecific crosses: plant regeneration from A. sativa L. Plant Cell Rep. 18, 582–588. doi: 10.1007/s002990050626
Kohli, A., Sreenivasulu, N., Lakshmanan, P., and Kumar, P. P. (2013). The phytohormone crosstalk paradigm takes center stage in understanding how plants respond to abiotic stresses. Plant Cell Rep. 32, 945–957. doi: 10.1007/s00299-013-1461-y
Korasick, D. A., Enders, T. A., and Strader, L. C. (2013). Auxin biosynthesis and storage forms. J. Exp. Bot. 64, 2541–2555. doi: 10.1093/jxb/ert080
Kudo, T., Makita, N., Kojima, M., Tokunaga, H., and Sakakibara, H. (2012). Cytokinin activity of cis-zeatin and phenotypic alterations induced by overexpression of putative cis-zeatin-o-glucosyltransferase in rice. Plant Physiol. 160, 319–331. doi: 10.1104/pp.112.196733
Kumar, H. G. A., Murthy, H. N., and Paek, K. Y. (2003). Embryogenesis and plant regeneration from anther cultures of Cucumis sativus L. Sci. Hortic. 98, 213–222. doi: 10.1016/S0304-4238(03)00003-7
Kumar, V., Parvatam, G., and Ravishankar, G. A. (2009). AgNO3 – a potential regulator of ethylene activity and plant growth modulator. Electronic J. Biotechnol. 12, 1–16. doi: 10.2225/vol12-issue2-fulltext-1
Lankova, M., Smith, R. S., Pesek, B., Kubes, M., Zazimalova, E., Petrasek, J., et al. (2010). Auxin influx inhibitors 1-NOA, 2-NOA, and CHPAA interfere with membrane dynamics in tobacco cells. J. Exp. Bot. 61, 3589–3598. doi: 10.1093/jxb/erq172
Lantos, C., Juhász, A., Somogyi, G., Ötvös, K., Vági, P., Mihály, R., et al. (2009). Improvement of isolated microspore culture of pepper (Capsicum annuum L.) via co-culture with ovary tissues of pepper or wheat. Plant Cell Tissue Organ Cult. 97, 285–293. doi: 10.1007/s11240-009-9527-9
Larqué-Saavedra, A. (1978). The antiranspirant effect of acetylsalcylic acid on Phaseolus vulgaris. Physiol. Plant. 43, 126–128. doi: 10.1111/j.1399-3054.1978.tb01579.x
Larqué-Saavedra, A. (1979). Stomatal closure in response to acetylsalicylic acid treatment. Z. Pflanzenphysiol. 93, 371–375. doi: 10.1016/S0044-328X(79)80271-8
Leroux, B., Carmoy, N., Giraudet, D., Potin, P., Larher, F., and Bodin, M. (2009). Inhibition of ethylene biosynthesis enhances embryogenesis of cultured microspores of Brassica napus. Plant Biotechnol. Rep. 3, 347–353. doi: 10.1007/s11816-009-0109-4
Leslie, C. A., and Romani, R. J. (1988). Inhibition of ethylene biosynthesis by salicylic acid. Plant Physiol. 88, 833–837. doi: 10.1104/pp.88.3.833
Letarte, J., Simion, E., Miner, M., and Kasha, K. J. (2006). Arabinogalactans and arabinogalactan-proteins induce embryogenesis in wheat (Triticum aestivum L.) microspore culture. Plant Cell Rep. 24, 691–698. doi: 10.1007/s00299-005-0013-5
Li, H., and Devaux, P. (2001). Enhancement of microspore culture efficiency of recalcitrant barley genotypes. Plant Cell Rep. 20, 475–481. doi: 10.1007/s002990100368
Li, J. M., and Chory, J. (1999). Brassinosteroid actions in plants. J. Exp. Bot. 50, 275–282. doi: 10.1093/jexbot/50.332.275
Liu, W., Zheng, M. Y., and Konzak, C. F. (2002a). Improving green plant production via isolated microspore culture in bread wheat (Triticum aestivum L.). Plant Cell Rep. 20, 821–824. doi: 10.1007/s00299-001-0408-x
Liu, W. G., Zheng, M. Y., Polle, E. A., and Konzak, C. F. (2002b). Highly efficient doubled-haploid production in wheat (Triticum aestivum L.) via induced microspore embryogenesis. Crop Sci. 42, 686–692. doi: 10.2135/cropsci2002.0686
Lu, C.-Y. (1993). The use of thidiazuron in tissue culture. In Vitro Cell. Dev. Biol. Plant 29, 92–96. doi: 10.1007/BF02632259
Ludwig-Muller, J. (2000). Indole-3-butyric acid in plant growth and development. Plant Growth Regul. 32, 219–230. doi: 10.1023/a:1010746806891
Lulsdorf, M., Yuan, H., Slater, S., Vandenberg, A., Han, X., and Zaharia, L. I. (2012). Androgenesis-inducing stress treatments change phytohormone levels in anthers of three legume species (Fabaceae). Plant Cell Rep. 31, 1255–1267. doi: 10.1007/s00299-012-1246-8
Luo, J. P., Jiang, S. T., and Pan, L. J. (2001). Enhanced somatic embryogenesis by salicylic acid of Astragalus adsurgens Pall.: relationship with H2O2 production and H2O2-metabolizing enzyme activities. Plant Sci. 161, 125–132. doi: 10.1016/s0168-9452(01)00401-0
Lyons, R., Manners, J. M., and Kazan, K. (2013). Jasmonate biosynthesis and signaling in monocots: a comparative overview. Plant Cell Rep. 32, 815–827. doi: 10.1007/s00299-013-1400-y
Malik, M. R., Wang, F., Dirpaul, J. M., Zhou, N., Hammerlindl, J., Keller, W., et al. (2008). Isolation of an embryogenic line from non-embryogenic Brassica napus cv. Westar through microspore embryogenesis. J. Exp. Bot. 59, 2857–2873. doi: 10.1093/jxb/ern149
Maraschin, S. F., De Priester, W., Spaink, H. P., and Wang, M. (2005). Androgenic switch: an example of plant embryogenesis from the male gametophyte perspective. J. Exp. Bot. 56, 1711–1726. doi: 10.1093/jxb/eri190
McRae, D. H., and Bonner, J. (1953). Chemical structure and antiauxin activity. Physiol. Plant. 6, 485–510. doi: 10.1111/j.1399-3054.1953.tb08406.x
Morris, D. A., and Johnson, C. F. (1987). Regulation of auxin transport in pea (Pisum sativum) by phenylacetic acid – inhibition of polar auxin transport in intact plants and stem segments. Planta 172, 408–416. doi: 10.1007/bf00398671
Moshkov, I. E., Novikova, G. V., Hall, M. A., and George, E. F. (2008). “Plant growth regulators III: gibberellins, ethylene, abscisic acid, their analogues and inhibitors; miscellaneous compounds,” in Plant Propagation by Tissue Culture, eds E. F. George, M. Hall, and G.-J. Klerk (Dordrecht: Springer), 227–281.
Moubayidin, L., Di Mambro, R., and Sabatini, S. (2009). Cytokinin-auxin crosstalk. Trends Plant Sci. 14, 557–562. doi: 10.1016/j.tplants.2009.06.010
Mulgund, G. S., Meti, N. T., Malabadi, R. B., Nataraja, K., and Kumar, S. V. (2012). Role of salicyclic acid on conifer somatic embryogenesis. Res. Biotechnol. 3, 57–61.
Normanly, J., Slovin, J., and Cohen, J. (2010). “auxin biosynthesis and metabolism,” in Plant Hormones, ed. P. Davies (Dordrecht: Springer), 36–62. doi: 10.1007/978-1-4020-2686-7_3
Perera, P. I. P., Ordonez, C. A., Becerra Lopez-Lavalle, L. A., and Dedicova, B. (2014). A milestone in the doubled haploid pathway of cassava. Protoplasma 251, 233–246. doi: 10.1007/s00709-013-0543-546
Petrasek, J., Cerna, A., Schwarzerova, K., Elckner, M., Morris, D. A., and Zazimalova, E. (2003). Do phytotropins inhibit auxin efflux by impairing vesicle traffic? Plant Physiol. 131, 254–263. doi: 10.1104/pp.012740
Poupart, J., and Waddell, C. S. (2000). The rib1 mutant is resistant to indole-3-butyric acid, an endogenous auxin in Arabidopsis. Plant Physiol. 124, 1739–1751. doi: 10.1104/pp.124.4.1739
Prem, D., Gupta, K., and Agnihotri, A. (2005). Effect of various exogenous and endogenous factors on microspore embryogenesis in Indian mustard (Brassica juncea (L.) Czern and Coss). In Vitro Cell. Dev. Biol. Plant 41, 266–273. doi: 10.1079/IVP2005636
Prem, D., Gupta, K., Sarkar, G., and Agnihotri, A. (2008). Activated charcoal induced high frequency microspore embryogenesis and efficient doubled haploid production in Brassica juncea. Plant Cell Tissue Organ Cult. 93, 269–282. doi: 10.1007/s11240-008-9373-1
Prem, D., Solis, M. T., Barany, I., Rodriguez-Sanz, H., Risueno, M. C., and Testillano, P. S. (2012). A new microspore embryogenesis system under low temperature which mimics zygotic embryogenesis initials, expresses auxin and efficiently regenerates doubled-haploid plants in Brassica napus. BMC Plant Biol. 12:127. doi: 10.1186/1471-2229-12-127
Pullman, G. S., Zhang, Y., and Phan, B. H. (2003). Brassinolide improves embryogenic tissue initiation in conifers and rice. Plant Cell Rep. 22, 96–104. doi: 10.1007/s00299-003-0674-x
Raghavan, C., Ong, E., Dalling, M., and Stevenson, T. (2006). Regulation of genes associated with auxin, ethylene and ABA pathways by 2,4-dichlorophenoxyacetic acid in Arabidopsis. Funct. Integr. Genomics 6, 60–70. doi: 10.1007/s10142-005-0012-1
Raghavan, V. (2004). Role of 2,4-dichlorophenoxyacetic acid (2,4-D) in somatic embryogenesis on cultured zygotic embryos of Arabidopsis: cell expansion, cell cycling, and morphogenesis during continuous exposure of embryos to 2,4-D. Am. J. Bot. 91, 1743–1756. doi: 10.3732/ajb.91.11.1743
Ramesar-Fortner, N. S., and Yeung, E. C. (2006). Physiological influences in the development and function of the shoot apical meristem of microspore-derived embryos of Brassica napus ‘Topas’. Can. J. Bot. 84, 371–383. doi: 10.1139/b06-007
Reynolds, T. L., and Crawford, R. L. (1996). Changes in abundance of an abscisic acid-responsive, early cysteine-labeled metallothionein transcript during pollen embryogenesis in bread wheat (Triticum aestivum). Plant Mol. Biol. 32, 823–829. doi: 10.1007/bf00020480
Rodriguez-Sanz, H., Manzanera, J.-A., Solis, M.-T., Gomez-Garay, A., Pintos, B., Risueno, M. C., et al. (2014). Early markers are present in both embryogenesis pathways from microspores and immature zygotic embryos in cork oak, Quercus suber L. BMC Plant Biol. 14:224. doi: 10.1186/s12870-014-0224-4
Santino, A., Taurino, M., De Domenico, S., Bonsegna, S., Poltronieri, P., Pastor, V., et al. (2013). Jasmonate signaling in plant development and defense response to multiple (a)biotic stresses. Plant Cell Rep. 32, 1085–1098. doi: 10.1007/s00299-013-1441-2
Sidhu, P., and Davies, P. (2009). Regeneration of fertile green plants from oat isolated microspore culture. Plant Cell Rep. 28, 571–577. doi: 10.1007/s00299-009-0684-4
Stuart, R., and Street, H. E. (1971). Studies on growth in culture of plant cells.10. Further studies on conditioning of culture media by suspensions of Acer pseudoplatanus L cells. J. Exp. Bot. 22, 96. doi: 10.1093/jxb/22.1.96
Swain, S. M., and Singh, D. P. (2005). Tall tales from sly dwarves: novel functions of gibberellins in plant development. Trends Plant Sci. 10, 123–129. doi: 10.1016/j.tplants.2005.01.007
Takahashi, Y., Yokoi, S., and Takahata, Y. (2012). Effects of genotypes and culture conditions on microspore embryogenesis and plant regeneration in several subspecies of Brassica rapa L. Plant Biotechnol. Rep. 6, 297–304. doi: 10.1007/s11816-012-0224-5
Titapiwatanakun, B., and Murphy, A. S. (2009). Post-transcriptional regulation of auxin transport proteins: cellular trafficking, protein phosphorylation, protein maturation, ubiquitination, and membrane composition. J. Exp. Bot. 60, 1093–1107. doi: 10.1093/jxb/ern240
Touraev, A., Vicente, O., and Heberlebors, E. (1997). Initiation of microspore embryogenesis by stress. Trends Plant Sci. 2, 297–302. doi: 10.1016/s1360-1385(97)89951-7
van Bergen, S., Kottenhagen, M. J., Van Der Meulen, R. M., and Wang, M. (1999). The role of abscisic acid in induction of androgenesis: a comparative study between Hordeum vulgare L. cvs. Igri and Digger. J. Plant Growth Regul. 18, 135–143. doi: 10.1007/pl00007061
Vyroubalová, S., Vaclavikova, K., Tureckova, V., Novak, O., Smehilova, M., Hluska, T., et al. (2009). Characterization of new maize genes putatively involved in cytokinin metabolism and their expression during osmotic stress in relation to cytokinin levels. Plant Physiol. 151, 433–447. doi: 10.1104/pp.109.142489
Wang, F., Cui, X., Sun, Y., and Dong, C. -H. (2013). Ethylene signaling and regulation in plant growth and stress responses. Plant Cell Rep. 32, 1099–1109. doi: 10.1007/s00299-013-1421-6
Wang, K. L. C., Li, H., and Ecker, J. R. (2002). Ethylene biosynthesis and signaling networks. Plant Cell 14, S131–S151. doi: 10.1105/tpc.001768
Wang, M., Hoekstra, S., Van Bergen, S., Lamers, G. E. M., Oppedijk, B. J., Van Der Heijden, M. W., et al. (1999). Apoptosis in developing anthers and the role of ABA in this process during androgenesis in Hordeum vulgare L. Plant Mol. Biol. 39, 489–501. doi: 10.1023/a:1006198431596
Wassom, J. J., Mei, C., Rocheford, T. R., and Widholm, J. M. (2001). Interaction of environment and ABA and GA treatments on the maize anther culture response. Plant Cell Tissue Organ Cult. 64, 69–72. doi: 10.1023/a:1010671613695
Woodward, A. W., and Bartel, B. (2005). Auxin: regulation, action, and interaction. Ann. Bot. 95, 707–735. doi: 10.1093/aob/mci083
Ye, Q., Zhu, W., Li, L., Zhang, S., Yin, Y., Ma, H., et al. (2010). Brassinosteroids control male fertility by regulating the expression of key genes involved in Arabidopsis anther and pollen development. Proc. Natl. Acad. Sci. U.S.A. 107, 6100–6105. doi: 10.1073/pnas.0912333107
Zeevaart, J. A. D., and Creelman, R. A. (1988). Metabolism and physiology of abscisic acid. Annu. Rev. Plant Physiol. Plant Mol. Biol. 39, 439–473. doi: 10.1146/annurev.arplant.39.1.439
Zhang, Y., Wang, A. J., Liu, Y., Wang, Y. S., and Feng, H. (2011). Effects of the antiauxin PCIB on microspore embryogenesis and plant regeneration in Brassica rapa. Sci. Hortic. 130, 32–37. doi: 10.1016/j.scienta.2011.06.047
Zheng, M. Y., Liu, W., Weng, Y., Polle, E., and Konzak, C. F. (2003). “Production of doubled haploids in wheat (Triticum aestivum L.) through microspore embryogenesis triggered by inducer chemicals,” in Doubled Haploid Production in Crop Plants, eds M. Maluszynski, K. J. Kasha, B. P. Forster, and I. Szarejko (Dordrecht: Springer), 83–94.
Zheng, M., Weng, Y., Liu, W., and Konzak, C. (2002). The effect of ovary-conditioned medium on microspore embryogenesis in common wheat (Triticum aestivum L.). Plant Cell Rep. 20, 802–807. doi: 10.1007/s00299-001-0411-2
Ziauddin, A., Marsolais, A., Simion, E., and Kasha, K. J. (1992). Improved plant regeneration from wheat anther and barley microspore culture using phenylacetic acid (PAA). Plant Cell Rep. 11, 489–498. doi: 10.1007/BF00236263
Zolman, B. K., Yoder, A., and Bartel, B. (2000). Genetic analysis of indole-3-butyric acid responses in Arabidopsis thaliana reveals four mutant classes. Genetics 156, 1323–1337.
Zoriniants, S., Tashpulatov, A. S., Heberle-Bors, E., and Touraev, A. (2005). “The role of stress in the induction of haploid microspore embryogenesis,” in Haploids in Crop Improvement II, eds D. Palmer, W. Keller, and K. Kasha (Berlin: Springer), 35–52.
Żur, I., Dubas, E., Golemiec, E., Szechynska-Hebda, M., Janowiak, F., and Wedzony, M. (2008). Stress-induced changes important for effective androgenic induction in isolated microspore culture of triticale (×Triticosecale Wittm.). Plant Cell Tissue Organ Cult. 94, 319–328.
Żur, I., Dubas, E., Krzewska, M., Janowiak, F., Hura, K., Pociecha, E., et al. (2014). Antioxidant activity and ROS tolerance in triticale (×Triticosecale Wittm.) anthers affect the efficiency of microspore embryogenesis. Plant Cell Tissue Organ Cult. 119, 79–94. doi: 10.1007/s11240-014-0515-3
Żur, I., Dubas, E., Krzewska, M., Waligórski, P., Dziurka, M., and Janowiak, F. (2015). Hormonal requirements for effective induction of microspore embryogenesis in triticale (×Triticosecale Wittm.) anther cultures. Plant Cell Rep. 34, 47–62. doi: 10.1007/s00299-014-1686-4
Żur, I., Krzewska, M., Dubas, E., Gołebiowska-Pikania, G., Janowiak, F., and Stojałowski, S. (2012). Molecular mapping of loci associated with abscisic acid accumulation in triticale (×Triticosecale Wittm.) anthers in response to low temperature stress inducing androgenic development. Plant Growth Regul. 68, 483–492.
Keywords: crop species, hormonal regulation, microspore embryogenesis, plant growth regulators, phytohormone crosstalk
Citation: Żur I, Dubas E, Krzewska M and Janowiak F (2015) Current insights into hormonal regulation of microspore embryogenesis. Front. Plant Sci. 6:424. doi: 10.3389/fpls.2015.00424
Received: 27 February 2015; Accepted: 26 May 2015;
Published online: 10 June 2015.
Edited by:
Jose M. Segui-Simarro, Universitat Politècnica de València, SpainReviewed by:
Jochen Kumlehn, Leibniz Institute of Plant Genetics and Crop Plant Research (IPK), GermanyAlison Ferrie, National Research Council of Canada, Canada
Copyright © 2015 Żur, Dubas, Krzewska and Janowiak. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Iwona Żur, The Franciszek Górski Institute of Plant Physiology, Polish Academy of Sciences, Niezapominajek 21, 30-239 Kraków, Poland,aS56dXJAaWZyLXBhbi5lZHUucGw=