 Rui Malhó
Rui Malhó- BioISI – Biosystems & Integrative Sciences Institute, Faculdade de Ciências, Universidade de Lisboa, Lisboa, Portugal
Apical cell growth seems to have independently evolved throughout the major lineages of life. To a certain extent, so does our body of knowledge on the mechanisms regulating this morphogenetic process. Studies on pollen tubes, root hairs, rhizoids, fungal hyphae, even nerve cells, have highlighted tissue and cell specificities but also common regulatory characteristics (e.g., ions, proteins, phospholipids) that our focused research sometimes failed to grasp. The working hypothesis to test how apical cell growth is established and maintained have thus been shaped by the model organism under study and the type of methods used to study them. The current picture is one of a dynamic and adaptative process, based on a spatial segregation of components that network to achieve growth and respond to environmental (extracellular) cues. Here, we explore some examples of our live imaging research, namely on cyclic nucleotide gated ion channels, lipid kinases and syntaxins involved in exocytosis. We discuss how their spatial distribution, activity and concentration suggest that the players regulating apical cell growth may display more mobility than previously thought. Furthermore, we speculate on the implications of such perspective in our understanding of the mechanisms regulating apical cell growth and their responses to extracellular cues.
Introduction
Apical tip growth is a form of cell extension common in all eukaryotes from rhizoids, pollen tubes, fungal hyphae to nerve cells. This growth form serves as a paradigm for cell polarity because cell extension is restricted to a narrow zone at the apex (Cheung and Wu, 2008). These cells are recognizably excellent models for cell research, particularly suitable for investigations on polarization, signal transduction, channel and ion flux activity, gene expression, cytoskeleton and wall structure, membrane dynamics and even cell-cell communication (Malhó et al., 2000; Onelli and Moscatelli, 2013; Sekereš et al., 2015).
As in any topic in science, advances in knowledge are dictated by current state of technology, the models under study and pre-existing ideas (typically represented in static diagrams such as the one represented in Figure 1). In the case of apical growth in plants, the outstanding technological advance registered in the past two decades, caused a perspective change from a morphological/structural approach to a dynamic/functional approach. When cell biology tools were predominantly used, researchers focused on larger cells that would grow straight and fast under in vitro conditions and would tolerate more harsh experimental conditions (e.g., microinjection, synthetic dyes, use of antibodies, and fixation methods). Lilium longiflorum was the paradigmatic example and underlying most experimental design was the hypothesis that if a molecule was important for tip growth, then its concentration should be higher in the apex (Malhó et al., 2000). The development of molecular tools in parallel with non-invasive methods to study live cells, made Arabidopsis thaliana a more accessible model and diversified tip growth studies. With the burst of genomics, a wider range of species (e.g., Physcomitrella patens) became also more prone to functional studies. Simple traits like fast growth rate or straight growth direction no longer hold as “universal” physiological controls and data interpretation must consider cellular and tissue variability.
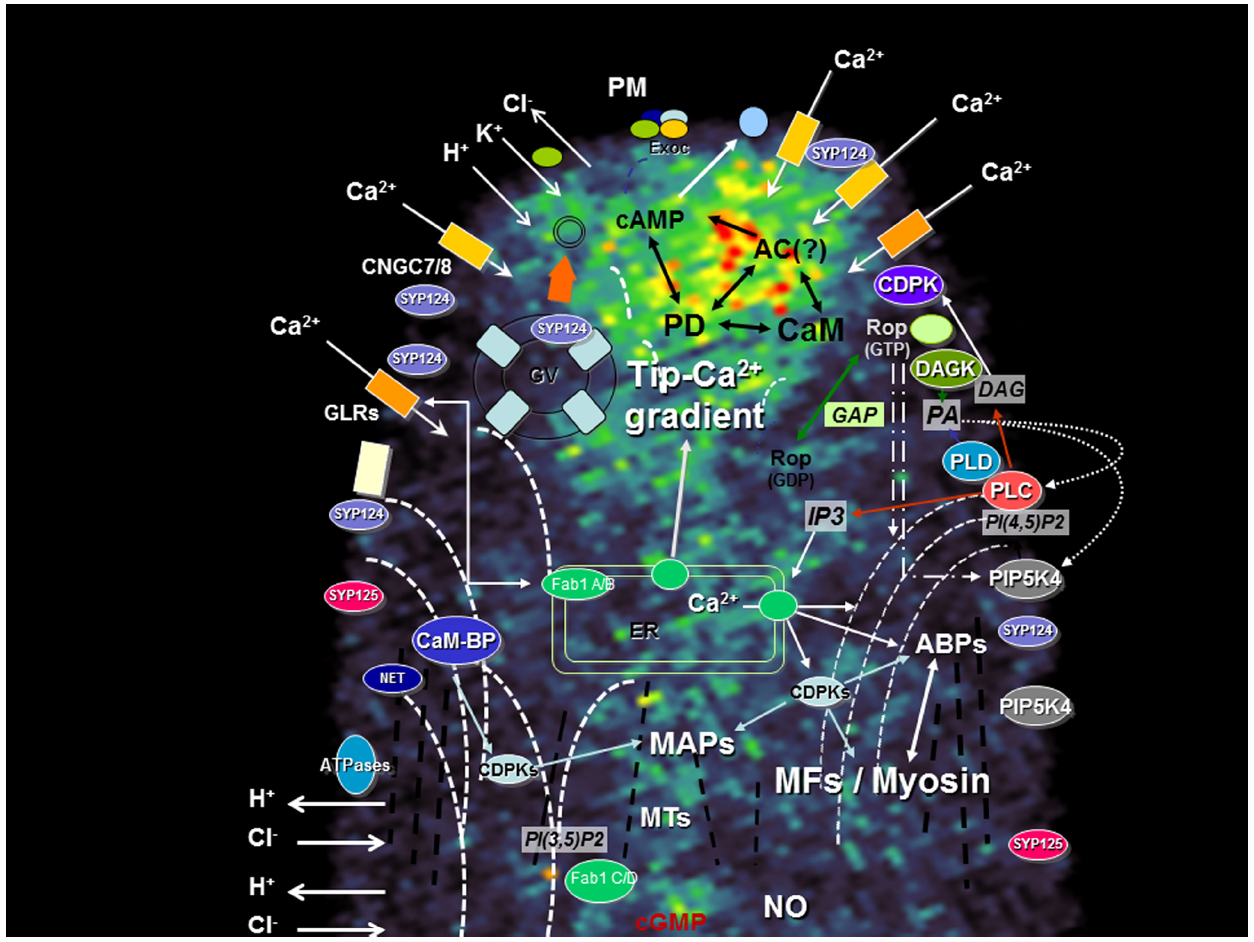
Figure 1. Apical region of a growing pollen tube depicting the main signaling transduction pathways and their components. A network between the different signaling pathways foresees the existence of a highly dynamic mechanism capable to interpret simultaneous extracellular cues and maintain polarity. The diagram is superimposed on a confocal image of a growing tobacco pollen tube loaded with FM1-43, a probe for endo-exocytosis [for methods see Camacho and Malhó (2003)]. ABPs, actin-binding proteins; AC, Adenylyl cyclase; CaM-BP, Calmodulin-binding protein; CDPK, Ca2+ dependent protein kinase; CNGC, cyclic nucleotide gated channel; DAG, diacylglycerol; DAGK, DAG kinase; Exoc, Exocyst; Fab1, PIKfyve/Fab1 kinase; GAP, Rop GTPase activating protein; GLR, Glutamate-like receptor; GV, Golgi vesicle; IP3, Inositol 1,4,5 triphosphate; MAPs, microtubule-associated protein; MFs, microfilaments (dashed white bars); MTs, microtubules (dashed black bars); NET, plant-specific Networked protein; NO, nitric oxide; PA, phosphatidic acid; PD, Phosphodiesterase; PI(3,5)P2, phosphatidylinositol-(3,5)-bis phosphate; PI(4,5)P2, phosphatidylinositol-(4,5)-bis phosphate; PIPK, phosphatidylinositol kinase; PLC, phospholipase C; PLD, phospholipase D; PM, plasma membrane; PMEs, pectin-methyl-esterases; R, IP3 receptor; SYP, syntaxin. Barbed arrows (↗) indicate direction of flux; Triangle arrows (△) indicate potential cross-regulatory effects. The curved MFs lines represent the actin fringe with larger cables extending to the sub-apex (and connecting to the plasma membrane) but not to the apical zone. The microtubules, where a fringe is not so visible, are represented as straight lines.
The diversity of species and characteristics of cells under study naturally generated a diversity of results (e.g., the natural growing environment of an Arabidopsis pollen tube is different from one of lily, from a root hair or from a moss rhizoid). Distinct asymmetric localization of components of the apical growth machinery (summarized in the diagram of Figure 1) were reported as part of new studies or reassessment of previous data. This, in turn, generated discrepancies, challenged old ideas and opened new perspectives. The architecture of the actin cytoskeleton (Vidali et al., 2009) and the secretory activity (Zonia and Munnik, 2008) are just two examples that challenged the “all-in-the-tip” concept and highlighted the importance of dynamics at the sub-apical region (Cheung and Wu, 2008; Sekereš et al., 2015). Here we explore the example of four different types of proteins recently studied by our group and which were found to be important for tip growth. In all four cases, we found that changes in growth pattern (e.g., redirectioning of growth axis, transient loss of polarity and recovery, oscillatory growth rates) were accompanied by protein delocalization. We discuss the implications of such findings and outline hypothesis to interpret the mechanisms underlying this complex machinery and their responses to extracellular stimuli.
Cyclic Nucleotide Gated Channels—Mobile and Flexible Ion Influx
Ca2+ signaling plays a key role in all aspects of plant development including apical growth. Namely, a tip-high gradient of cytosolic free ([Ca2+]c) resulting from influx of extracellular Ca2+ seems crucial to establish polarity (Hepler et al., 2012). Evidence from pharmacological and genetic approaches indicate this influx occurs through at least two different types of Ca2+ -permeable channels—glutamate receptor-like proteins (GLRs) and cyclic nucleotide gated channels (CNGCs; Michard et al., 2011; Tunc-Ozdemir et al., 2013). CNGCs are cation channels with varying degrees of ion conduction selectivity harboring a cyclic nucleotide-binding domain and a calmodulin binding domain (Zelman et al., 2012). They can therefore integrate signals from distinct transduction pathways and could be functioning in a way that indirectly triggers a Ca2+ release from an internal store (Spalding and Harper, 2011). CNGCs were shown to be essential in tip-growing cells (Frietsch et al., 2007; Tunc-Ozdemir et al., 2013) and, in straight growing pollen tubes, GFP-CNGC7 was found to preferentially localize to the plasma membrane at the flanks of the growing tip (Figure 2A, top image). But at the onset of germination, perhaps the phase where apical Ca2+ influx is more relevant to establish a growth axis, the GFP-CNG7 signal was higher at the apex (Figure 2A, middle image). Similar observations were made in pollen tubes recovering and/or reorienting the growth axis (Figure 2A, lower image) suggesting that cells regulate the fine tuning of protein localization in response to extracellular cues. FRAP experiments of GFP fused to RLK (Receptor-Like-Kinase; Lee et al., 2008) showing apical fluorescence recovery support such hypothesis.
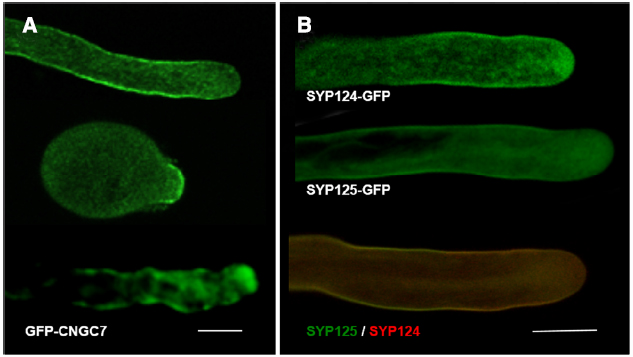
Figure 2. Confocal Imaging of GFP constructs of CNGC7, SYP124 and SYP125. (A) Arabidopsis pollen tubes expressing GFP-CNG7, in different growth phases—straight growth; upon germination; wiggling growth. For methods see Tunc-Ozdemir et al. (2013). Bar = 10 μm. (B) Tobacco pollen tubes expressing SYP124-GFP (upper image), SYP125-GFP (middle image), and co-expressing both constructs (lower image; SYP124 signal in red and SYP125 signal in green). For methods see Ul-Rehman et al. (2011). Bar = 10 μm.
Tip growing cells not only have to frequently adjust direction of growth axis but they also experience oscillations in growth rates. Several types of growth fluctuations (of asymmetric periodicity and intensity) have been observed and they seem to vary according to species and cell type. Whether these oscillations have any special physiological meaning or whether they are just a “built-in” characteristic is not clear. [Ca2+]c exhibits changes that correlated to such fluctuations of growth rates and reorientation of growth axis (Castanho Coelho and Malhó, 2006) so it is highly plausible that other components of the polarity mechanism (e.g., lipids and membrane-associated proteins) also exhibit changes in activity and/or localization. The timing of our observations (and the inherent physiological status of the cells) may thus influence our reports and help to explain some apparent discrepancies that exist in the literature.
PIP Kinases—Versatile and Key Transducers of “Signal to Form”
In the last years PtdIns(4,5)P2 and its synthesizing enzyme, phosphatidylinositol phosphate kinase (PIPK), have been intensively studied in plant cells, revealing a key role in the control of polar tip growth. Using fluorescence markers fused to the pleckstrin homology (PH) domain of the human PLCd1, PtdIns(4,5)P2 was found to accumulate at the tip of growing apical cells (Kost et al., 1999; Dowd et al., 2006; Helling et al., 2006; Ischebeck et al., 2008; Sousa et al., 2008). Analysis of the PIPK members from Arabidopsis thaliana, Oryza sativa, and Physcomitrella patens showed that they share some regulatory features with animal PIPKs but also exert plant-specific modes of regulation (Saavedra et al., 2012). Deletion or overexpression of these lipid kinases were found to cause distinct phenotypes and perturbations in cellular processes such as cell wall deposition, endocytosis, and actin bundling (Ischebeck et al., 2008, 2010; Kusano et al., 2008; Sousa et al., 2008; Stenzel et al., 2008; Zhao et al., 2010; Saavedra et al., 2011). Interestingly, in actively growing pollen tubes, all the six PIPK isoforms (AtPIP5K10, AtPIP5K11, AtPIP5K2, AtPIP5K4, AtPIP5K5, and AtPIP5K6) were found to localize preferentially at the flanks of the tube apex and not superimposed with the highest PtdIns(4,5)P2 concentration. However, it was also observed that the region displaying the highest protein fluorescent signal would change according to speed of growth and reorientation of the growth axis—slower growth resulting in delocalization from the flanks to the apex (e.g., Figure 6 of Ischebeck et al., 2008; Sousa et al., 2008). It has been suggested that distinct localization patterns of proteins may be the consequence of interactions with specific partners, which recruit them to different functional microdomains (Ischebeck et al., 2010). For example, PtdIns(4,5)P2 could be channeled to targets via specific interactions PIPK-downstream effectors (Heilmann and Heilmann, 2015) resulting in differential cellular responses and phenotypes as observed upon deletion of AtPIP5Ks (Ischebeck et al., 2008, 2010; Kusano et al., 2008; Sousa et al., 2008).
Phosphatidylinositol phosphate kinases have membrane occupation and recognition nexus (MORN) motifs which are thought to be the plasma membrane localizing module (Kusano et al., 2008) but other modules were shown to be important for correct subcellular localization (Mikami et al., 2010; Stenzel et al., 2012). E.g., AtPIP5K5 and NtPIP5K6-1 require non-conserved linker (LIM) domain for correct localization in pollen tubes, as the deletion of the N-terminal and MORN domain did not affect their apical plasma membrane localization (Stenzel et al., 2012). It is thus possible that protein modules responsible for plasma membrane localization are distinct in each PIPK allowing a fine tuning that depends on differences in physiological and/or developmental status of cells, such as polarized and non-polarized. Full comprehension of the localization mechanisms will probably involve comparison of the function of the MORN, LIM, and kinase domains of every PIPKs in cells exhibiting the same stage in development.
Syntaxins—Spatial Targeting of Secretion and Membrane Recycling
Apical growth occurs by continuous vesicle secretion and delivery of new wall material. Therefore, the exact sub-cellular location of endocytic and exocytic domains is essential to determine cellular responses and reshape form. In pollen tubes, it was previously assumed that exocytosis events occur mostly at the extreme apex, where [Ca2+]c is higher, while membrane recycling (endocytosis) would take place further back from the tip, at the flanks of the apex and/or at sub-apical regions (Picton and Steer, 1981; Castanho Coelho and Malhó, 2006). Recent data questioned this paradigm suggesting that the preferential location for fusion is on a limited membrane domain at the sub-apical flanks and not at the extreme apex (Bove et al., 2008; Zonia and Munnik, 2008). Similar observations have been made in root hairs (Ovecka et al., 2005). The hypothesis of two endocytic modes co-existing according to the growth conditions—a clathrin-dependent and an independent one—was raised (Camacho and Malhó, 2003; Monteiro et al., 2005) and experimental evidence to support it was obtained by Moscatelli et al. (2007) and McKenna et al. (2009). We have also obtained evidence that two pollen-specific SNAREs (for soluble N-ethylmaleimide sensitive factor attachment protein receptor), syntaxins SYP125 and SYP124 (Silva et al., 2010; Ul-Rehman et al., 2011) have a complementary, not fully overlapping, dynamic distribution (Figure 2B). Similarly to PIPK proteins, the observed changes in protein accumulation at the plasma membrane might reflect specific interactions with unidentified targets (e.g., PIP2 and phosphatidic acid) which, under natural growth conditions of pollen tubes, could translate into discrete asymmetric secretory events and relate to the two endocytic modes. Indirect support for such hypothesis derives from our observations that in pollen tubes, [Ca2+]c and membrane fusion exhibit frequent non-linear changes correlated to growth rates and reorientation of growth axis (Castanho Coelho and Malhó, 2006). Over-expression of PIP5K4 or Rab GTPases was also found to perturb SYP124 localization (Silva et al., 2010). The changes in SYP124 and SYP125 distribution observed upon growth modulation are thus likely to reflect membrane dynamics and a repositioning of the vesicle’s docking machinery upon intra- and extracellular cues. In nerve cells, syntaxins were reported to be involved in rapid and slow endocytosis (Xu et al., 2013) and could thus act as regulators of both endo- and exocytosis. VAMP726, another member of the SNARES family, was also shown to mediate fusion of endo- and exocytic compartments in pollen tube tip growth (Guo and McCubbin, 2012). A similar role has just been proposed for the exocyst (Jose et al., 2015), a complex known to be present in tip growing cells (Zárský et al., 2013).
Similarly to our CNGC observations, these findings highlight the importance of syntaxins in secretion and tip growth but must be interpreted considering that the localization reported reflects protein accumulation and not necessarily activity.
FAB Kinases- Dynamics in the Secretory Pathway
Most of the data now available relating apical polarity and growth focused on the role of plasma membrane (integral or associated) events. The plasma membrane and its links to the cell wall and the cytoskeleton are the obvious main targets for perceiving and transducing both intra- and extracellular cues. Notwithstanding, we have recently obtained data suggesting that PIKfyve/Fab1 kinases localized to the endomembrane compartment are also involved in the regulation of plasma membrane recycling events and thus in the maintenance of polarity (Serrazina et al., 2014).
In plants, the Fab1 phosphatidylinositol-3-monophosphate 5-kinases produce phosphatidylinositol (3,5)-bisphosphate [PtdIns(3,5)P2], a phosphoinositide implicated in endomembrane trafficking and pH control in the vacuole (Dove et al., 2009; Bak et al., 2013). In pollen tubes, we found that AtFAB1B-GFP had a highly mobile reticulate-like distribution, distinct from γ-TIP (Serrazina et al., 2014) and decorating the sub-apical region in a manner similar to the actin cytoskeleton—a sort of V-shaped collar that adjusts and “accompanies” the reorientation of the growth axis (Vidali et al., 2009). This suggests that the protein is localized in protein storage vacuoles, endoplasmic reticulum and possibly trans-Golgi network mediating transport to and from the plasma membrane (Gary et al., 1998; Wang et al., 2011). Additionally, we found that FAB1 deletion resulted in lower internalization rates and perturbed secretion and deposition of new wall material (Serrazina et al., 2014). Analogous observations were made in root hairs (Hirano et al., 2011). FAB1 deletion was also found to impair acidification of the endomembrane compartment and to cause abnormal pollen tube diameter (Serrazina et al., 2014). The perturbation of proton gradients across membranes was reported to affect membrane curvature and vesicularization (Hope et al., 1989), protein sorting (Hurtado-Lorenzo et al., 2006), and ion fluxes, all of which are processes essential for tip growth.
These results confirm that apical polarity is not solely dependent on a positive feedback mechanism based on a single protein or ion influx localization but rather on an orchestrated network of signals. Gui et al. (2014) have recently found that overexpression of LePRK1, a pollen-specific and plasma membrane-localized receptor-like kinase, dramatically affects tube morphology in a process that can be counterbalanced by an actin bundling protein (PLIM2a) in a Ca2+-responsive manner.
Conclusion and Perspectives
The establishment and maintenance of apical polarity relies on a dynamic, mobile network of signaling mechanisms. Currently, our conceptual models focus mostly on apical and sub-apical localization (of ions, proteins, lipids), particularly at the plasma membrane (or associated with). Here we provided examples of four classes of proteins related to ion and lipid signaling which indicates that a careful analysis of localization, activity and mobility is required in order to fully assign their role in apical growth. This rationale can probably be extended to other equally relevant signaling components identified in these cells (e.g., Rop GTPases, Ca2+-dependent protein kinases, actin and actin-binding proteins, etc; Figure 1; for a review see Onelli and Moscatelli, 2013). The results obtained with FAB kinases further suggest the importance of plasma membrane—endomembrane signaling raising new perspectives in the study of apical growth mechanisms. Signaling to other organelles and compartments is also likely to play key roles in the establishment of polarity.
We have discussed the hypothesis that, in nature, and upon a myriad of environmental (extracellular) cues, cellular responses involve flexible positioning of proteins, lipids and ion fluxes. Such dynamics may go partly unnoticed in the set-ups we devise for our experimental planning which are conditioned by the species under study, the technical approach, the pre-existing knowledge and, most importantly, by the requirements to test individual stimuli. Thus, transient gradients or peaks of activity/localization may or not be recorded (even dismissed) depending on the experimental set-up, cellular conditions and our biased previous background. Large single cell analysis and settings mimicking (or bearing in mind) the natural environment where cells develop, are required but may be difficult to implement. To compare and interpret results obtained with different experimental as well as plant systems will be a challenge, but one that must be tackled.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by Fundação Ciência e Tecnologia (FCT/MCTES/PIDDAC, Portugal) with post-doc fellowship to LS (SFRH/BPD/63619/2009), FD (SFRH/BPD/81635/2011), and research funds to RM (PEst-OE/BIA/UI4046/2014; UID/MULTI/04046/2013).
References
Bak, G., Lee, E.-J., Lee, Y., Kato, M., Segami, S., Sze, H., et al. (2013). Rapid structural changes and acidification of guard cell vacuoles during stomatal closure require phosphatidylinositol 3,5-bisphosphate. Plant Cell 25, 2202–2216. doi: 10.1105/tpc.113.110411
Bove, J., Vaillancourt, B., Kroeger, J., Hepler, P. K., Wiseman, P. W., and Geitmann, A. (2008). Magnitude and direction of vesicle dynamics in growing pollen tubes using spatiotemporal image correlation spectroscopy and fluorescence recovery after photobleaching. Plant Physiol. 147, 1646–1658. doi: 10.1104/pp.108.120212
Camacho, L., and Malhó, R. (2003). Endo-exocytosis in the pollen tube apex is differentially regulated by Ca2+ and GTPases. J. Exp. Bot. 54, 83–92. doi: 10.1093/jxb/erg043
Castanho Coelho, P., and Malhó, R. (2006). Correlative analysis of apical secretion and [Ca2+]c in pollen tube growth and reorientation. Plant Signal. Behav. 1, 152–157. doi: 10.4161/psb.1.3.2999
Cheung, A., and Wu, H. M. (2008). Structural and signaling networks for the polar cell growth machinery in pollen tubes. Annu. Rev. Plant Biol. 59, 547–572. doi: 10.1146/annurev.arplant.59.032607.092921
Dove, S. K., Dong, K., Kobayashi, T., Williams, F. K., and Michell, R. H. (2009). Phosphatidylinositol 3,5-bisphosphate and Fab1p/PIKfyve under PPIn endo-lysosome function. Biochem. J. 419, 1–13. doi: 10.1042/BJ20081950
Dowd, P. E., Courso, S., Skirpan, A. L., Kao, T.-H., and Gilroy, S. (2006). Petunia phospholipase C1 is involved in pollen tube growth. Plant Cell 18, 1438–1453. doi: 10.1105/tpc.106.041582
Frietsch, S., Wang, Y.-F., Sladek, C., Poulsen, L. R., Romanowsky, S. M., Schroeder, J. I., et al. (2007). A cyclic nucleotide-gated channel is essential for polarized tip growth of pollen. Proc. Natl. Acad. Sci. U.S.A. 104, 14531–14536. doi: 10.1073/pnas.0701781104
Gary, J. D., Wurmser, A. E., Bonangelino, C. J., Weisman, L. S., and Emr, S. D. (1998). Fab1p is essential for PtdIns(3)P 5-kinase activity and the maintenance of vacuolar size and membrane homeostasis. J. Cell Biol. 143, 65–79. doi: 10.1083/jcb.143.1.65
Gui, C. P., Dong, X., Liu, H. K., Huang, W. J., Zhang, D., Wang, S. J., et al. (2014). Overexpression of the tomato pollen receptor kinase LePRK1 rewires pollen tube growth to a blebbing mode. Plant Cell 26, 3538–3555. doi: 10.1105/tpc.114.127381
Guo, F., and McCubbin, A. (2012). The pollen-specific R-SNARE/longin PiVAMP726 mediates fusion of endo- and exocytic compartments in pollen tube tip growth. J. Exp. Bot. 63, 3083–3095. doi: 10.1093/jxb/ers023
Heilmann, M., and Heilmann, I. (2015). Plant phosphoinositides-complex networks controlling growth and adaptation. Biochim. Biophys. Acta 1851, 759–769. doi: 10.1016/j.bbalip.2014.09.018
Helling, D., Possart, A., Cottier, S., Klahre, U., and Kost, B. (2006). Pollen tube tip growth depends on plasma membrane polarization mediated by tobacco PLC3 activity and endocytic membrane recycling. Plant Cell 18, 3519–3534. doi: 10.1105/tpc.106.047373
Hepler, P. K., Kunkel, J. G., Rounds, C. M., and Winship, L. J. (2012). Calcium entry into pollen tubes. Trends Plant Sci. 17, 32–38. doi: 10.1016/j.tplants.2011.10.007
Hirano, T., Matsuzawa, T., Takegawa, K., and Sato, M. H. (2011). Loss-of-function and gain-of-function mutations in FAB1A/B impair endomembrane homeostasis, conferring pleiotropic developmental abnormalities in Arabidopsis. Plant Physiol. 155, 797–807. doi: 10.1104/pp.110.167981
Hope, M. J., Redelmeier, T. E., Wong, K. F., Rodrigueza, W., and Cullis, P. R. (1989). Phospholipid asymmetry in large unilamellar vesicles induced by transmembrane pH gradients. Biochemistry 28, 4181–4187. doi: 10.1021/bi00436a009
Hurtado-Lorenzo, A., Skinner, M., El Annan, J., Futai, M., Sun-Wada, G. H., Bourgoin, S., et al. (2006). V-ATPase interacts with ARNO and Arf6 in early endosomes and regulates the protein degradative pathway. Nat. Cell Biol. 8, 124–136. doi: 10.1038/ncb1348
Ischebeck, T., Stenzel, I., and Heilmann, I. (2008). Type B phosphatidylinositol-4-phosphate 5-kinases mediate Arabidopsis and Nicotiana tabacum pollen tube growth by regulating apical pectin secretion. Plant Cell 20, 3312–3330. doi: 10.1105/tpc.108.059568
Ischebeck, T., Stenzel, I., Hempel, F., Jin, X., Mosblech, A., and Heilmann, I. (2010). Phosphatidylinositol-4,5-bisphosphate influences Nt-Rac5-mediated cell expansion in pollen tubes of Nicotiana tabacum. Plant J. 65, 453–468. doi: 10.1111/j.1365-313X.2010.04435.x
Jose, M., Tollis, S., Nair, D., Mitteau, R., Velours, C., Massoni-Laporte, A., et al. (2015). A quantitative imaging-based screen reveals the exocyst as a network hub connecting endocytosis and exocytosis. Mol. Biol. Cell 26, 2519–2534. doi: 10.1091/mbc.E14-11-1527
Kost, B., Lemichez, E., Spielhofer, P., Hong, Y., Tolias, K., Carpenter, C., et al. (1999). Rac homologues and compartmentalized phosphatidylinositol 4, 5-bisphosphate act in a common pathway to regulate polar pollen tube growth. J. Cell Biol. 145, 317–330. doi: 10.1083/jcb.145.2.317
Kusano, H., Testerink, C., Vermeer, J. E., Tsuge, T., Shimada, H., Oka, A., et al. (2008). The Arabidopsis phosphatidylinositol phosphate 5-kinase PIP5K3 is a key regulator of root hair tip growth. Plant Cell 20, 367–380. doi: 10.1105/tpc.107.056119
Lee, Y. J., Szumlanski, A., Nielsen, E., and Yang, Z. (2008). Rho-GTPase-dependent filamentous actin dynamics coordinate vesicle targeting and exocytosis during tip growth. J. Cell Biol. 181, 1155–1168. doi: 10.1083/jcb.200801086
Malhó, R., Camacho, L., and Moutinho, A. (2000). Signaling pathways in pollen tube growth and reorientation. Ann. Bot. 85(Suppl. A), 59–68. doi: 10.1006/anbo.1999.0991
McKenna, S. T., Kunkel, J. G., Bosch, M., Rounds, C. M., Vidali, L., Winship, L. J., et al. (2009). Exocytosis precedes and predicts the increase in growth in oscillating pollen tubes. Plant Cell 21, 3026–3040. doi: 10.1105/tpc.109.069260
Michard, E., Lima, P. T., Borges, F., Silva, A. C., Portes, M. T., Carvalho, J. E., et al. (2011). Glutamate receptor-like genes form Ca2+ channels in pollen tubes and are regulated by pistil D-serine. Science 332, 434–437. doi: 10.1126/science.1201101
Mikami, K., Saavedra, L., Hiwatashi, Y., Uji, T., Hasebe, M., and Sommarin, M. (2010). A dibasic amino acid pair conserved in the activation loop directs plasma membrane localization and is necessary for activity of plant type I/II phosphatidylinositol phosphate kinase. Plant Physiol. 153, 1004–1015. doi: 10.1104/pp.109.152686
Monteiro, D., Liu, Q., Lisboa, S., Scherer, G. E. F., Quader, H., and Malhó, R. (2005). Phosphoinositides and phosphatidic acid regulate pollen tube growth and reorientation through modulation of [Ca2+]c and membrane secretion. J. Exp. Bot. 56, 1665–1674. doi: 10.1093/jxb/eri163
Moscatelli, A., Ciampolini, F., Rodighiero, S., Onelli, E., Cresti, M., Santo, N., et al. (2007). Distinct endocytic pathways identified in tobacco pollen tubes using charged nanogold. J. Cell Sci. 120, 3804–3819. doi: 10.1242/jcs.012138
Onelli, E., and Moscatelli, A. (2013). Endocytic pathways and recycling in growing pollen tubes. Plants 2, 211–229. doi: 10.3390/plants2020211
Ovecka, M., Lang, I., Baluška, F., Ismail, A., Illes, P., and Lichtscheidl, I. (2005). Endocytosis and vesicle trafficking during tip growth of root hairs. Protoplasma 226, 39–54. doi: 10.1007/s00709-005-0103-9
Picton, J. M., and Steer, M. W. (1981). Determination of secretory vesicle production rates by dictyossomes in pollen tubes of Tradescantia using cytochalasin D. J. Cell Sci. 49, 261–272.
Saavedra, L., Balbi, V., Lerche, J., Mikami, K., Heilmann, I., and Sommarin, M. (2011). PIPKs are essential for rhizoid elongation and caulonemal cell development in the moss Physcomitrella patens. Plant J. 67, 635–647. doi: 10.1111/j.1365-313X.2011.04623.x
Saavedra, L., Mikami, K., Malhó, R., and Sommarin, M. (2012). PIP kinases and their role in plant tip growing cells. Plant Signal. Behav. 7, 1302–1305. doi: 10.4161/psb.21547
Sekereš, J., Pleskot, R., Pejchar, P., Žárský, V., and Potocký, M. (2015). The song of lipids and proteins: dynamic lipid–protein interfaces in the regulation of plant cell polarity at different scales. J. Exp. Bot. 66, 1587–1598. doi: 10.1093/jxb/erv052
Serrazina, S., Vaz Dias, F., and Malhó, R. (2014). Characterization of FAB1 phosphatidylinositol kinases in Arabidopsis pollen tube growth and fertilization. New Phytol. 203, 784–793. doi: 10.1111/nph.12836
Silva, P., Rehman, R., Rato, C., Di Sansebastiano, G.-P., and Malhó, R. (2010). Asymmetric localization of Arabidopsis syntaxins at the pollen tube apical and sub-apical zones is involved in tip growth. BMC Plant Biol. 10:179. doi: 10.1186/1471-2229-10-179
Sousa, E., Kost, B., and Malhó, R. (2008). Arabidopsis phosphatidylinositol-4-monophosphate 5-kinase 4 regulates pollen tube growth and polarity by modulating membrane recycling. Plant Cell 20, 3050–3064. doi: 10.1105/tpc.108.058826
Spalding, E. P., and Harper, J. F. (2011). The ins and outs of cellular Ca2+ transport. Curr. Opin. Plant Biol. 14, 715–720. doi: 10.1016/j.pbi.2011.08.001
Stenzel, I., Ischebeck, T., Konig, S., Holubowska, A., Sporysz, M., Hause, B., et al. (2008). The type B phosphatidylinositol-4-phosphate 5-kinase 3 is essential for root hair formation in Arabidopsis thaliana. Plant Cell 20, 124–141. doi: 10.1105/tpc.107.052852
Stenzel, I., Ischebeck, T., Quint, M., and Heilmann, I. (2012). Variable regions of PI4P 5-kinases direct PtdIns(4,5)P2 towards alternative regulatory functions in tobacco pollen tubes. Front. Plant Sci. 2:114. doi: 10.3389/fpls.2011.00114
Tunc-Ozdemir, M., Rato, C., Brown, E., Rogers, S., Mooneyham, A., Frietsch, S., et al. (2013). Cyclic nucleotide gated channels 7 and 8 are essential for male reproductive fertility. PLoS ONE 8:e55277. doi: 10.1371/journal.pone.0055277
Ul-Rehman, R., Silva, P., and Malhó, R. (2011). Localization of Arabidopsis SYP125 syntaxin in the plasma membrane sub-apical and distal zones of growing pollen tubes. Plant Signal. Behav. 6, 665–670. doi: 10.4161/psb.6.5.14423
Vidali, L., Rounds, C. M., Hepler, P. K., and Bezanilla, M. (2009). Lifeact-mEGFP reveals a dynamic apical F-Actin network in tip growing plant cells. PLoS ONE 4:e5744. doi: 10.1371/journal.pone.0005744
Wang, H., Zhuang, X. H., Hillmer, S., Robinson, D. G., and Jiang, L. W. (2011). Vacuolar sorting receptor (VSR) proteins reach the plasma membrane in germinating pollen tubes. Mol. Plant 4, 845–853. doi: 10.1093/mp/ssr011
Xu, J., Luo, F., Zhang, Z., Xue, L., Wu, X.-S., Chiang, H.-C., et al. (2013). SNARE proteins synaptobrevin, SNAP-25, and syntaxin are involved in rapid and slow endocytosis at synapses. Cell Rep. 3, 1414–1421. doi: 10.1016/j.celrep.2013.03.010
Zárský, V., Kulich, I., Fendrych, M., and Pečenková, T. (2013). Exocyst complexes multiple functions in plant cells secretory pathways. Curr Opin. Plant Biol. 16, 726–733. doi: 10.1016/j.pbi.2013.10.013
Zelman, A. K., Dawe, A., Gehring, C., and Berkowitz, G. A. (2012). Evolutionary and structural perspectives of plant cyclic nucleotide-gated cation channel. Front. Plant Sci. 3:95. doi: 10.3389/fpls.2012.00095
Zhao, Y., Yan, A., Feijó, J. A., Furutani, M., Takenawa, T., Hwang, I., et al. (2010). Phosphoinositides regulate clathrin-dependent endocytosis at the tip of pollen tubes in Arabidopsis and tobacco. Plant Cell 22, 4031–4044. doi: 10.1105/tpc.110.076760
Keywords: Ca2+, cyclic nucleotides, syntaxins, phosphoinositides, signaling, tip growth
Citation: Malhó R, Serrazina S, Saavedra L, Dias FV and Rehman RU (2015) Ion and lipid signaling in apical growth—a dynamic machinery responding to extracellular cues. Front. Plant Sci. 6:816. doi: 10.3389/fpls.2015.00816
Received: 30 June 2015; Accepted: 18 September 2015;
Published: 06 October 2015.
Edited by:
Eric Ruelland, Centre National de la Recherche Scientifique, FranceReviewed by:
Imara Y. Perera, North Carolina State University, USAHaitao Shi, Hainan University, China
Copyright © 2015 Malhó, Serrazina, Saavedra, Dias and Rehman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rui Malhó and Reiaz Ul-Rehman, BioISI – Biosystems & Integrative Sciences Institute, Faculdade de Ciências, Universidade de Lisboa, Campo Grande, 1749-016 Lisboa, Portugal,ci5tYWxob0BmYy51bC5wdCw=cmVpYXpyZWhtYW5AeWFob28uY28uaW4=
†Present address: Laura Saavedra, Cátedra de Fisiología Vegetal, Facultad de Ciencias Exactas, Físicas y Naturales, Universidad Nacional de Córdoba, Córdoba, Argentina; Reiaz Ul-Rehman, Department of Bioresources, University of Kashmir, Hazratbal, Srinagar 190006, India