 Carl H. Mesarich
Carl H. Mesarich Joanna K. Bowen
Joanna K. Bowen Cyril Hamiaux
Cyril Hamiaux Matthew D. Templeton
Matthew D. Templeton- 1School of Biological Sciences, The University of Auckland, Auckland, New Zealand
- 2Host–Microbe Interactions, Bioprotection, The New Zealand Institute for Plant & Food Research Ltd, Auckland, New Zealand
- 3Human Responses, The New Zealand Institute for Plant & Food Research Limited, Auckland, New Zealand
Many plant-associated organisms, including microbes, nematodes, and insects, deliver effector proteins into the apoplast, vascular tissue, or cell cytoplasm of their prospective hosts. These effectors function to promote colonization, typically by altering host physiology or by modulating host immune responses. The same effectors however, can also trigger host immunity in the presence of cognate host immune receptor proteins, and thus prevent colonization. To circumvent effector-triggered immunity, or to further enhance host colonization, plant-associated organisms often rely on adaptive effector evolution. In recent years, it has become increasingly apparent that several effectors of plant-associated organisms are repeat-containing proteins (RCPs) that carry tandem or non-tandem arrays of an amino acid sequence or structural motif. In this review, we highlight the diverse roles that these repeat domains play in RCP effector function. We also draw attention to the potential role of these repeat domains in adaptive evolution with regards to RCP effector function and the evasion of effector-triggered immunity. The aim of this review is to increase the profile of RCP effectors from plant-associated organisms.
Effectors of Plant-associated Organisms
Diverse plant-associated organisms, including bacteria, fungi, oomycetes, nematodes, and insects, secrete or inject a suite of proteins, termed effectors, into the tissues of their prospective hosts (Bozkurt et al., 2012; Deslandes and Rivas, 2012; Mitchum et al., 2013; Jaouannet et al., 2014; Lo Presti et al., 2015). These effectors, which localize to the host apoplast, or are targeted to various plant cell compartments, function to promote colonization, typically by altering host physiology or by modulating host immune responses (Hogenhout et al., 2009; Win et al., 2012a). Certain host plants however, have evolved immune receptor proteins that are capable of directly or indirectly recognizing one or more of these effectors or their modulated host targets respectively, to trigger immune responses that prevent colonization (Böhm et al., 2014; Cui et al., 2015). To circumvent these recognition events, or to provide novel, altered, or extended effector functionalities that further enhance the colonization of susceptible hosts, plant-associated organisms often rely on effector modification through adaptive evolution, as driven by host-imposed selection pressure (e.g., Stergiopoulos et al., 2007; Win et al., 2007; Dong et al., 2014).
Several Effectors of Plant-associated Organisms are Repeat-containing Proteins
Proteins that make up the effector repertoires of plant-associated organisms possess a range of different features. For example, most carry a signal peptide for targeted secretion or delivery to the host environment. In addition, many effectors, particularly those of fungi, are small and/or cysteine-rich, while others may possess a nuclear localization signal (NLS) or, as shown for several effectors of filamentous plant-associated organisms, a conserved effector motif (Dou and Zhou, 2012). The secretomes, and thus effector repertoires, of plant-associated organisms also differ in their proportion of repeat-containing proteins (RCPs). This is best illustrated by the predicted secretomes of Melampsora larici-populina and Puccinia graminis f. sp. tritici, the fungal pathogens responsible for poplar leaf rust and wheat stem rust, respectively. In a study by Saunders et al. (2012), it was revealed that of the 1549 secreted proteins predicted from the proteome of M. larici-populina, 493 (~32%) were RCPs. In contrast, no RCPs could be identified among the 1852 secreted proteins predicted from the proteome of P. graminis f. sp. tritici (Saunders et al., 2012). As such, RCP effectors are expected to play an important role in promoting the colonization of some, but not all, plant-associated organisms. This is supported by the fact that several known effectors of plant-associated organisms are RCPs (Tables 1–3). For the purpose of this review, we define RCPs as those proteins that carry two or more copies of a tandemly or non-tandemly duplicated sequence or structural motif that is at least five amino acid residues in length.
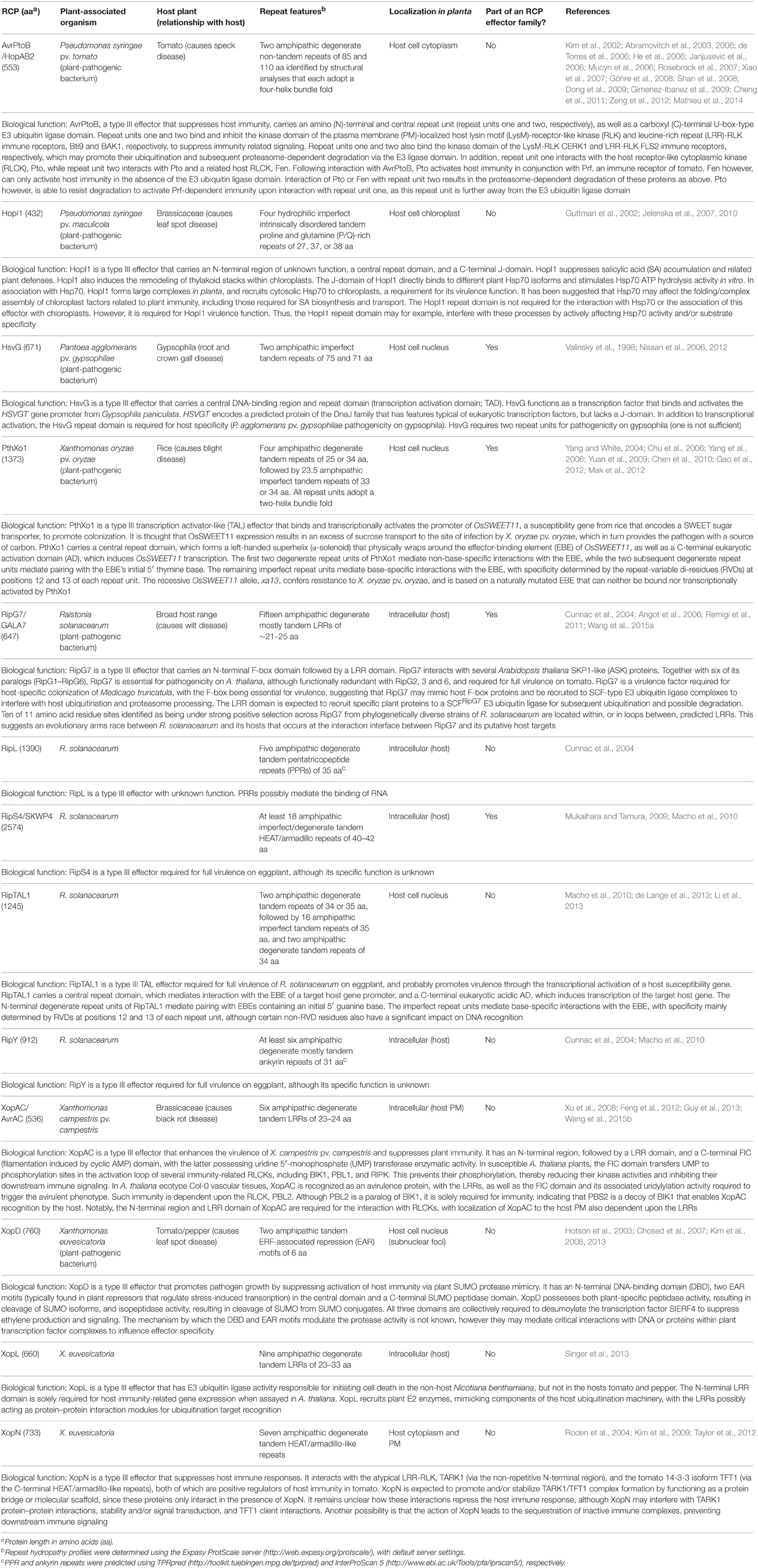
Table 1. Examples of repeat-containing protein (RCP) effectors from plant-associated bacteria.
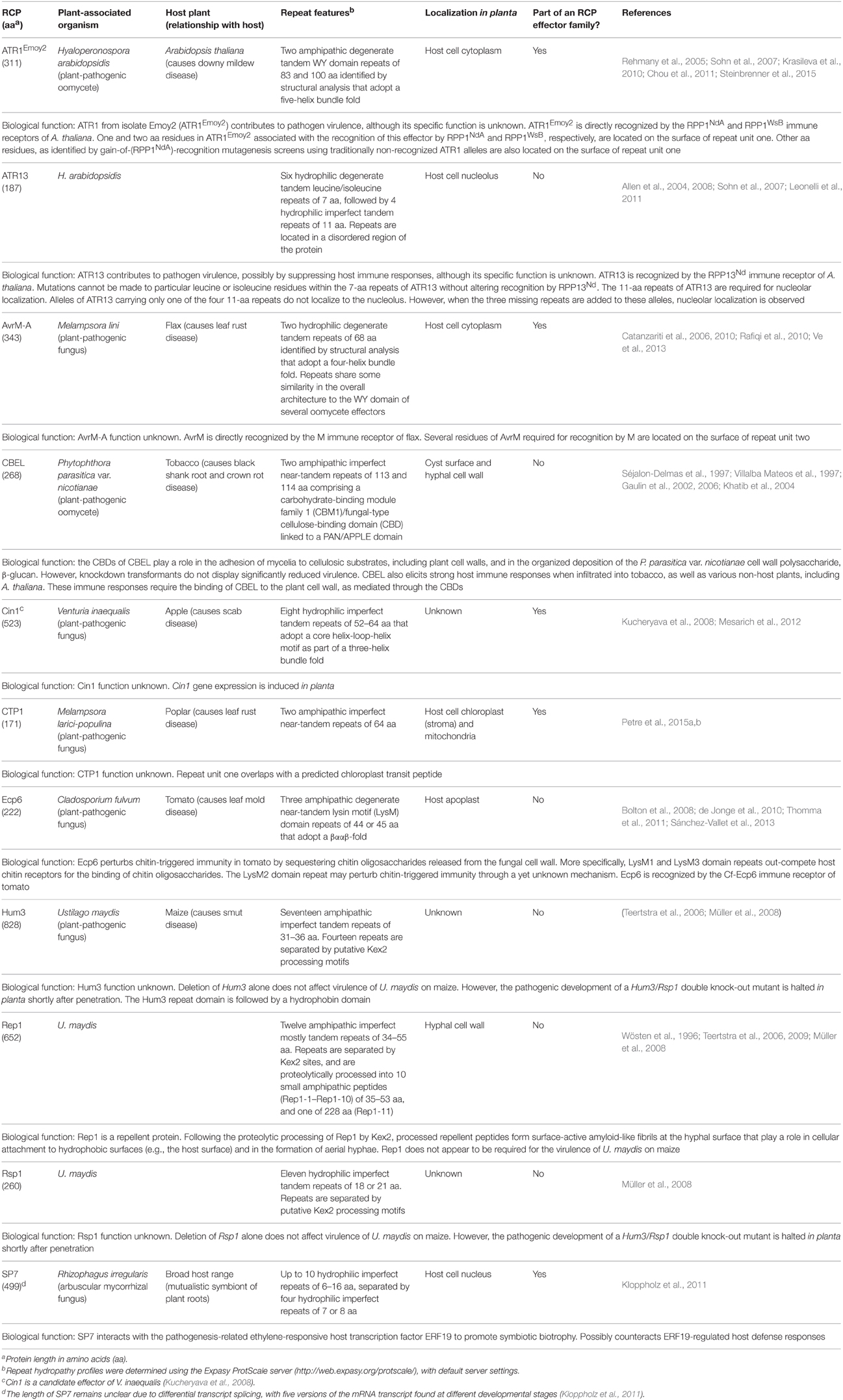
Table 2. Examples of repeat-containing protein (RCP) effectors and surface-associated RCPs from plant-associated fungi and oomycetes.
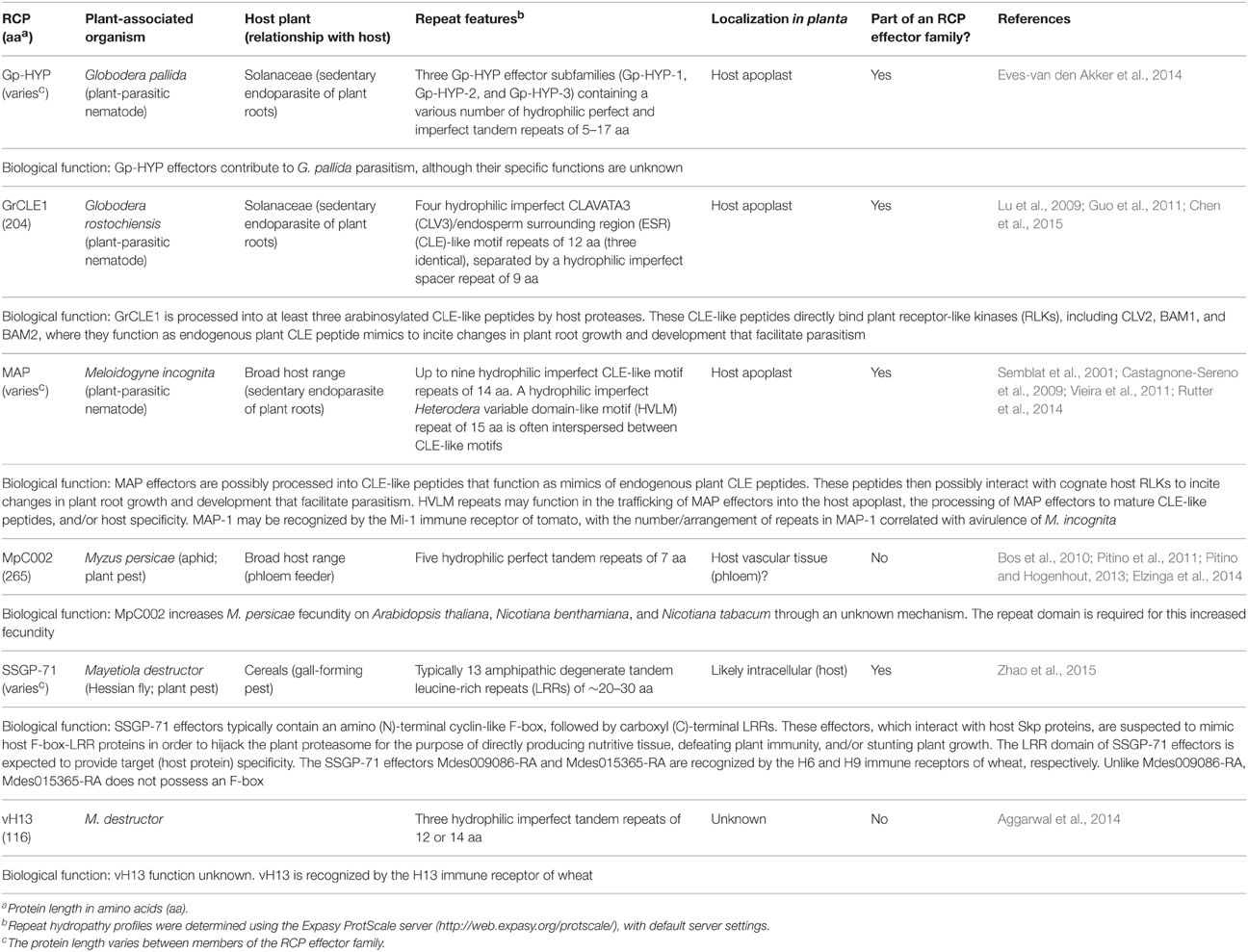
Table 3. Examples of repeat-containing protein (RCP) effectors from plant-associated nematodes and insects.
Various bioinformatic tools, databases, and servers are available for the detection of repeat domains in protein sequences (reviewed in Kajava, 2012; Luo and Nijveen, 2014). Typically, perfect (identical) or imperfect (near-identical) sequence repeats are easily detected, as are those repeats with homology to known functional domains. However, the detection of highly degenerate (divergent) sequence repeats, which carry amino acid substitutions, insertions, or deletions that have accumulated during evolution, is often more difficult. In some instances, degenerate sequence repeats may only be identified following an analysis of protein tertiary structure, for which servers are again available (see Kajava, 2012). Indeed, this has been the case for several effectors of plant-associated organisms. As an example of this, structural characterization of both the AvrM-A effector from Melampsora lini, a fungal rust pathogen of flax, as well as AvrPtoB, a type III effector from Pseudomonas syringae pv. tomato (Pst), the bacterial speck pathogen of tomato, revealed the presence of two four-helix bundle repeats (Figures 1A,B, 2B) (Dong et al., 2009; Cheng et al., 2011; Ve et al., 2013). Bioinformatic tools though, have been shown to play a key role in the identification of certain highly degenerate repeat domains. For example, Jiang et al. (2008) used the MEME algorithm (Bailey et al., 2015), together with hidden Markov model (HMM) searches, to identify RXLR effectors from two plant-associated oomycete species (Phytophthora sojae and Phytophthora ramorum) that carry conserved, but highly degenerate, C-terminal WYL motifs, or WY motifs, which often form tandem repeats. In oomycete plant pathogens, RXLR effectors represent one of the largest and most diverse effector families (Jiang et al., 2008). Jiang et al. (2008) demonstrated that approximately half of the abovementioned RXLR effectors possess WYL motifs, with 30% possessing between two and eight repeated WYL modules. A comparison of RXLR effector tertiary structures has since revealed that a three-helix bundle fold, termed the WY domain, is the basic structural unit adopted by the WY motifs (Boutemy et al., 2011; Win et al., 2012b). One of these structurally characterized RXLR effectors, ATR1, which is produced by Hyaloperonospora arabidopsidis, the oomycete downy mildew pathogen of Arabidopsis thaliana, carries two five-helix bundle WY domain repeats (Figure 2A) (Chou et al., 2011). Notably though, this tandem repeat was only identified upon structural characterization of ATR1, with a prior HMM-based bioinformatic screen identifying only one of the two WY domains present in this effector (Boutemy et al., 2011). This example therefore highlights the difficulties associated with identifying highly degenerate repeat domains. More recently though, Ye et al. (2015) have demonstrated that WYL motifs have highly conserved α-helical secondary structures. Furthermore, the few amino acid residues that are conserved between such WYL or WY motifs have been shown to be hydrophobic, occupying buried positions within these α-helices (Boutemy et al., 2011; Chou et al., 2011; Win et al., 2012b; Ye et al., 2015). Thus, an integrated approach, combining HMM screens, together with secondary structure predictions and surface accessibility profiles, can be employed to identify the degenerate, and often repeated, WYL or WY motifs present in oomycete RXLR effectors.
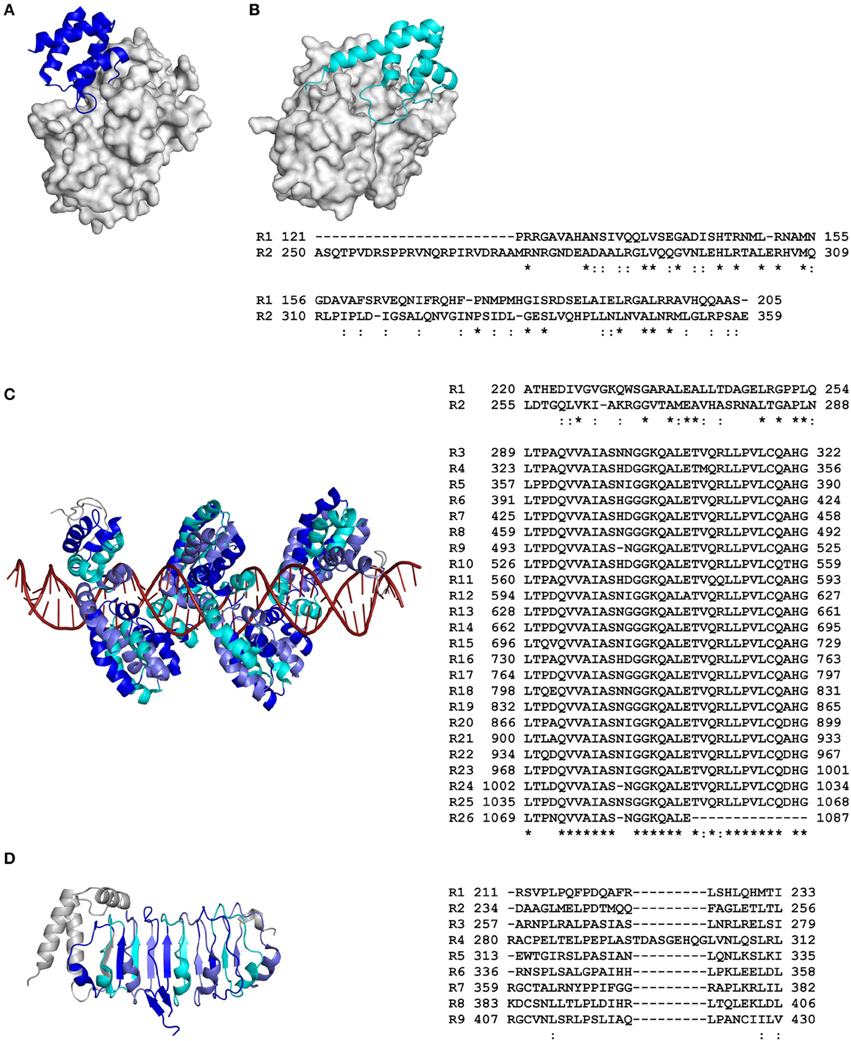
Figure 1. Primary and tertiary structures of repeat domains from RCP effectors of plant-associated bacteria. (A) Crystal structure of repeat unit one from the AvrPtoB effector of the tomato bacterial speck pathogen, Pseudomonas syringae pv. tomato (Pst), in complex with the tomato Pto kinase (Protein Data Bank [PDB] code 3HGK; Dong et al., 2009). (B) Nuclear magnetic resonance (NMR) structure of repeat unit two from AvrPtoB of Pst in complex with the BAK1 kinase domain from Arabidopsis thaliana (3TL8; Cheng et al., 2011). Note that in (A), AvrPtoB repeat unit one interacts with the Pto kinase in a different orientation to that of AvrPtoB repeat unit two with the BAK1 kinase domain in (B). (C) Crystal structure of the repeat domain from the PthXo1 transcription activator-like (TAL) effector of the bacterial rice pathogen, Xanthomonas oryzae pv. oryzae, bound to its natural DNA target (36 bp). The repeats pack together to form a left-handed superhelix (α-solenoid) that wraps around the DNA molecule (3UGM; Mak et al., 2012). (D) Crystal structure of the N-terminal leucine-rich repeat (LRR) domain from the XopL effector of the bacterial leaf spot pathogen of pepper and tomato, Xanthomonas euvesicatoria (4FCG; Singer et al., 2013). Structural coordinate files were downloaded from the Research Collaboratory for Structural Bioinformatics (RCSB) PDB (http://www.rcsb.org/pdb/home/home.do). Alternating repeat units are colored blue, slate, and cyan, respectively. Non-repetitive sequence is colored gray. The molecular surface of Pto kinase in (A) and BAK1 kinase domain in (B) are shown in gray, while the DNA molecule in (C) is colored red. An amino acid sequence alignment detailing the primary structure of each RCP effector repeat domain is shown to the right of each tertiary structure (as based on that presented in each tertiary structure). Repeat (R) units are numbered according to their position in the RCP effector. The start and end position of each repeat unit in the full-length RCP effector is shown. Conserved (*) and strongly similar (:) amino acid residues shared between repeat units are shown below the sequence alignment (based on full-length repeat units only). The figure was prepared using PyMol (https://www.pymol.org/) and Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/).
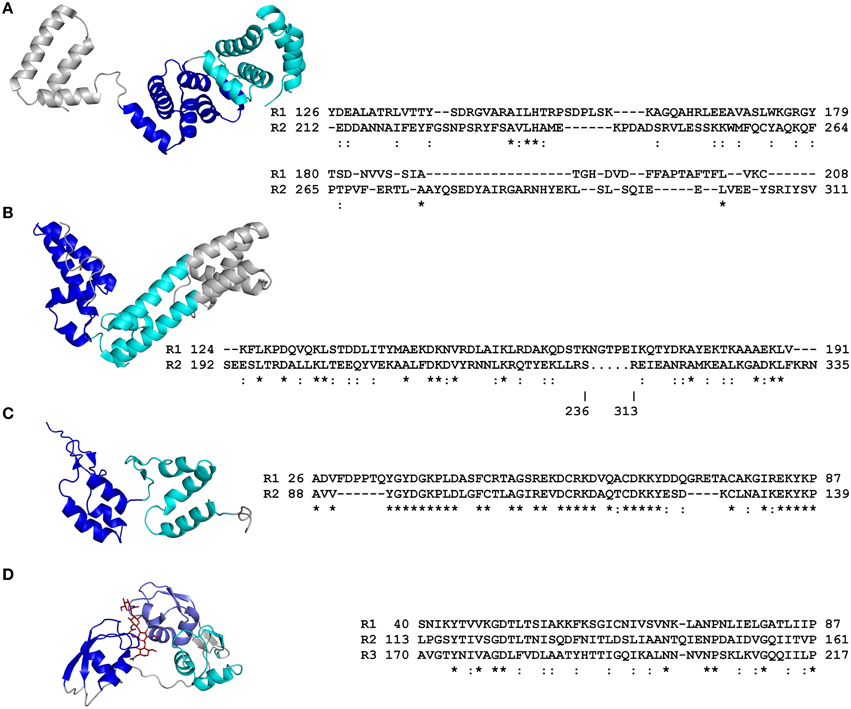
Figure 2. Primary and tertiary structures of repeat domains from RCP effectors of plant-associated fungi and an oomycete. (A) Crystal structure of the ATR1 effector from the Arabidopsis thaliana oomycete pathogen, Hyaloperonospora arabidopsidis (Protein Data Bank [PDB] code 3RMR; Chou et al., 2011). (B) Crystal structure of the AvrM-A effector from the flax rust fungus, Melampsora lini (4BJN; Ve et al., 2013). (C) Nuclear magnetic resonance (NMR) structure of repeat units 1 and 2 from the candidate effector Cin1 of the apple scab fungus, Venturia inaequalis (2LHT; Mesarich et al., 2012). (D) Crystal structure of the Ecp6 effector from the tomato leaf mold fungus, Cladosporium fulvum. The lysin motif (LysM) repeat units 1 and 3 coordinate the binding of a single chitin tetramer by means of an inter-repeat domain groove (4B8V; Sánchez-Vallet et al., 2013). Structural coordinate files were downloaded from the Research Collaboratory for Structural Bioinformatics (RCSB) PDB (http://www.rcsb.org/pdb/home/home.do). Alternating repeat units are colored blue, slate, and cyan, respectively. Non-repetitive sequence is colored gray. The chitin tetramer in (D) is colored red. An amino acid sequence alignment detailing the primary structure of each RCP effector repeat domain is shown to the right of each tertiary structure (as based on that presented in each tertiary structure). Repeat (R) units are numbered according to their position in the RCP effector. The start and end position of each repeat unit in the full-length RCP effector is shown. Conserved (*) and strongly similar (:) amino acid residues shared between repeat units are shown below the sequence alignment. The figure was prepared using PyMol (https://www.pymol.org/) and Clustal Omega (http://www.ebi.ac.uk/Tools/msa/clustalo/). Structure-based sequence alignments of repeat units from ATR1 and AvrM-A are adapted from Chou et al. (2011) and Ve et al. (2013), respectively.
Repeat Domains Play Diverse Roles in RCP Effector Function
Collectively, repeat domains play diverse roles in the biological function of RCP effectors from plant-associated organisms (Tables 1–3). In brief, these roles can range from directing effector localization, to mediating interaction with one or more specific RNA, DNA, protein, or carbohydrate targets, to providing effector stability. It is becoming increasingly clear that these roles are intimately linked to the composition or architecture of the repeat domains that perform them. For example, as shown in Figures 1, 2, the repeat domain of an RCP effector, like that of many other RCPs (Grove et al., 2008), frequently exhibits an extended modular, non-globular architecture. This in turn provides the effector with a larger surface area-to-volume ratio than that of a typical globular protein of equivalent amino acid length, a feature that is particularly well-suited to certain functional roles. This is elegantly illustrated by the transcription activator-like (TAL) effectors of the bacterial plant pathogens, Xanthomonas spp., which interact with host DNA in the plant cell nucleus to hijack host genes (by transcriptional activation) whose expression promotes bacterial growth and/or disease symptom formation (Boch and Bonas, 2010). TAL effectors carry a central repeat domain that possesses up to 33.5 near-identical tandem repeats of 30–42 amino acids in length, followed by a carboxyl (C)-terminal region that contains both NLSs and a eukaryotic acidic activation domain (Boch and Bonas, 2010). As shown for PthXo1, a TAL effector from the rice blight pathogen, Xanthomonas oryzae pv. oryzae, the central repeat domain forms an extended surface area of interaction with host DNA, in which the repeat domain adopts an α-solenoid structure that physically wraps around the DNA molecule (Figure 1C) (Deng et al., 2012; Mak et al., 2012). More specifically, the individual repeat units mediate the direct binding of single consecutive nucleotide bases within the promoter sequence (i.e., the effector-binding element; EBE) of a host gene. This specificity is governed by amino acid residues 12 and 13 of each repeat unit, termed the repeat-variable di-residues (RVDs), which make specific contact with the host DNA and play a stabilizing role, respectively (Boch et al., 2009; Moscou and Bogdanove, 2009). The functional relevance of this repeat structure was reinforced by artificial TAL effectors carrying a variable number of repeat units. Boch et al. (2009) were able to show that a minimum of 6.5 repeat units are necessary for EBE recognition and subsequent transcriptional activation, while 10.5 or more repeat units are required for strong target gene expression.
An extended modular, non-globular architecture, as adopted by the repeat domains of many RCPs, is also particularly well-suited to mediating various protein–protein interactions (Grove et al., 2008). Indeed, many classes of repeat domains serve as scaffolds or adaptors. When performing this role, different repeat units, or regions of a repeat unit, may organize multiple proteins into functional complexes. Alternatively, interactions between different proteins, or between proteins and other functional domains present in the RCP, may be facilitated (Grove et al., 2008). Importantly, these roles are supported by the inherent conformational flexibility of the repeat domain, as mediated through for instance, a flexible hydrophobic core (Kappel et al., 2010), or flexible inter-repeat hinges, loops, or linkers, similar to those found in Cin1, a candidate effector of unknown function from the apple scab fungus, Venturia inaequalis (Figure 2C) (Mesarich et al., 2012). Domains that may perform such a role include, for example, those comprising ankyrin or HEAT/armadillo repeats, which, like the repeat domains present in TAL effectors, adopt an α-solenoid-type architecture, as well as leucine-rich repeats (LRRs), which adopt an α/β-solenoid-like or horseshoe-type fold (Kajava, 2012). Notably, several effectors from plant-associated organisms carry such repeat domains. For example, effectors of the bacterial wilt pathogen, Ralstonia solanacearum, including RipAP, RipBB, RipBC, and RipY, carry ankyrin repeats (Peeters et al., 2013), while other effectors of R. solanacearum and Xanthomonas spp., including RipS1–RipS8, XopAD, and XopN, carry HEAT/armadillo repeats (White et al., 2009; Peeters et al., 2013). In addition, several effectors from R. solanacearum (RipG1–RipG7), Xanthomonas spp. (XopAC, XopAE, and XopL), and the gall-forming pest of cereals, Mayetiola destructor (SSGP-71 family), carry LRRs (Figure 1D) (Xu et al., 2008; White et al., 2009; Peeters et al., 2013; Zhao et al., 2015).
Of the effectors mentioned above, one of the best characterized to date is XopN, a type III effector widely conserved across Xanthomonas spp. that suppresses host immune responses (Roden et al., 2004; Kim et al., 2009; Taylor et al., 2012). XopN from the leaf spot pathogen of pepper and tomato, Xanthomonas euvesicatoria, carries seven tandem HEAT/armadillo-like repeats (Roden et al., 2004). This effector interacts with the atypical LRR-receptor-like kinase (RLK), TARK1 (via the XopN non-repetitive N-terminal region), and the 14-3-3 isoform, TFT1 (via the XopN C-terminal HEAT/armadillo-like repeats), two positive regulators of host immunity in tomato, near and at the plant cytoplasmic–plasma membrane (PM) interface, respectively (Kim et al., 2009; Taylor et al., 2012). In addition to these binary interactions, XopN also engages in tertiary interactions with TARK1 and TFT1 at the plant cytoplasmic–PM interface (Kim et al., 2009; Taylor et al., 2012). Here XopN is expected to promote and/or stabilize TARK1/TFT1 complex formation by functioning as a protein bridge or molecular scaffold (Taylor et al., 2012). Currently however, it remains unclear how these interactions suppress host immune responses. One possibility is that XopN interferes with TARK1 protein–protein interactions, stability and/or signal transduction, and in the case of TFT1, client interactions (Kim et al., 2009; Taylor et al., 2012). Another possibility, given that TARK1 and TFT1 do not interact in the absence of XopN, is that the binding of this effector to these proteins in either binary or tertiary complexes leads to the sequestration of inactive immune complexes at or near the plant cytoplasmic–PM interface, thereby preventing downstream immune signaling (Taylor et al., 2012).
Other repeat domain architectures and compositions have been shown to play an important role in the function of RCP effectors from plant-associated organisms. One such example is provided by Ecp6, an effector of the tomato leaf mold fungus, Cladosporium fulvum, which carries three lysin motif (LysM) domains that each adopt a βααβ-fold as part of an overall globular structure (Figure 2D) (Bolton et al., 2008; Sánchez-Vallet et al., 2013). Ecp6 molecules sequester chitin oligosaccharides released from the cell wall of C. fulvum during infection. In doing so, Ecp6 prevents the recognition of these oligosaccharides by host chitin immune receptors, thereby perturbing chitin-triggered immunity (de Jonge et al., 2010). More specifically, two of the three LysM domains, LysM1, and LysM3, undergo chitin-induced dimerization, in which the domains cooperate to produce a deeply buried chitin-binding groove (Figure 2D). This groove binds a single chitin oligosaccharide with ultra-high affinity, and is sufficient to out-compete host chitin immune receptors for chitin binding (Sánchez-Vallet et al., 2013). Another example is provided by GrCLE1, an effector of the potato cyst nematode, Globodera rostochiensis (Lu et al., 2009). GrCLE1 possesses a variable domain, followed by a C-terminal region with four 12-amino acid repeats that have similarity to plant CLAVATA3 (CLV3)/endosperm surrounding region (ESR)-related (CLE) peptides (Lu et al., 2009). In plants, endogenous CLE protein precursors are post-translationally modified and proteolytically processed to give bioactive CLE peptides. These peptides then function as hormones that interact with various extracellular plant receptors to regulate many aspects of plant growth and development (Kucukoglu and Nilsson, 2015). Like plant CLE protein precursors, GrCLE1 is post-translationally modified and proteolytically processed by plant machinery to produce bioactive CLE-like peptides (Guo et al., 2011; Chen et al., 2015). These peptides then function as endogenous plant CLE peptide mimics, directly binding plant RLKs, including CLV2, BAM1, and BAM2, to alter plant root growth and development for the promotion of plant parasitism (Lu et al., 2009; Guo et al., 2011; Chen et al., 2015).
Several RCPs of Plant-associated Organisms are Surface-associated
An important point to stress is that several RCPs of plant-associated organisms are surface-associated. That is, they are attached to, or are integrated into, the cell wall and/or PM through various covalent/non-covalent linkages or transmembrane domains, and are at least partially exposed to the extracellular environment. Although not classified as typical secreted effectors, a number of these surface-associated RCPs, and more specifically their repeat domains, have been shown or are hypothesized to play a role in interactions between plant-associated organisms and their hosts (e.g., Görnhardt et al., 2000; Robold and Hardham, 2005; Lanver et al., 2010; Pradhan et al., 2012). An example is provided by CBEL, a cell wall glycoprotein from Phytophthora parasitica var. nicotianae (Ppn), the oomycete root pathogen responsible for black shank disease of tobacco (Nicotiana tabacum) (Séjalon-Delmas et al., 1997; Villalba Mateos et al., 1997). CBEL possesses two repeats, each comprising a carbohydrate-binding module family 1 (CBM1)/fungal-type cellulose-binding domain (CBD) attached to a PAN/APPLE domain (Séjalon-Delmas et al., 1997; Villalba Mateos et al., 1997). Functional analyses have determined that these CBDs play a role in the adhesion of Ppn mycelia to cellulosic substrates, including plant cell walls, and in the organized deposition of the Ppn cell wall polysaccharide, β-glucan (Villalba Mateos et al., 1997; Gaulin et al., 2002, 2006). Interestingly, CBEL also elicits strong host immune responses when infiltrated into tobacco (Villalba Mateos et al., 1997), as well as various non-host plants, including A. thaliana (Khatib et al., 2004; Gaulin et al., 2006). These responses are dependent upon the binding of CBEL to the plant cell wall, as mediated through the CBDs (Gaulin et al., 2006). A second example is provided by Rep1 of the corn smut fungus, Ustilago maydis, which carries 12 mostly tandem repeats of 34–55 amino acids in length (Wösten et al., 1996). These repeats, which carry Kex2 recognition sites, are processed in the secretory pathway to 11 repellent peptides that form rigid surface-active amyloid-like fibrils at the hyphal surface, and play a role in cellular attachment to hydrophobic surfaces (e.g., the plant surface) and in the formation of aerial hyphae (Wösten et al., 1996; Teertstra et al., 2006, 2009; Müller et al., 2008; Lanver et al., 2014).
Repeat Domains may Contribute to the Adaptive Evolution of RCP Effectors
Repeat domains can evolve in several different ways, including through changes in repeat unit number or order, as well as through amino acid substitutions or insertions/deletions (indels) in repeat units and/or associated interconnecting loop/linker regions. Changes in number or order, particularly for those repeat units encoded by long nucleotide sequences (≥10 nucleotides in length), likely evolve through intra- and inter-genic recombination events (Richard and Pâques, 2000). As shown in other systems, the mutation rates associated with these changes can be orders of magnitude greater than those associated with point mutations, accelerating the evolution of the coding sequence to which they belong (reviewed in Gemayel et al., 2010). Indeed, repeat unit number and/or order has commonly been shown to vary between RCP effectors and RCP effector candidates of individuals, strains, or isolates of the same species or pathovar of plant-associated organism (e.g., Allen et al., 2004; Heuer et al., 2007; Jelenska et al., 2007; Kucheryava et al., 2008; Aggarwal et al., 2014). Changes in repeat unit number have also been shown to accompany the evolutionary paths of certain effector families from plant-associated organisms (e.g., Goss et al., 2013). Furthermore, chimeric RCP effectors, resulting from a recombination event between homologous repeat domains, have been reported (e.g., Yang et al., 2005), a finding that is not surprising, given the high number of RCP effectors that belong to multi-protein families (Tables 1–3). Although generally not as quick to accumulate, amino acid substitutions, and indels also play an important role in generating sequence diversity within a repeat domain. However, these types of modification only occur following a duplication event. Again, such sequence variation has commonly been found to occur between the repeat units of RCP effectors or RCP effector candidates (see imperfect or degenerate repeat units listed in Tables 1–3), as well as between the repeat domains of RCP effectors and RCP effector candidates from individuals, strains, or isolates of the same species or pathovar of plant-associated organism (e.g., Kucheryava et al., 2008; Chou et al., 2011; Ve et al., 2013).
Of what relevance could this repeat domain variability be to plant-associated organisms? In industrial and animal-pathogenic yeasts, alterations to the repeat unit number, and/or order of surface-associated RCPs, termed adhesins, have been shown to impart changes in adhesion phenotype, which may permit the rapid adaptation of these organisms to different substrates and host tissues, respectively (reviewed in Verstrepen and Fink, 2009). Furthermore, variability in the repeat domains of RCPs has been linked to the evasion of host immune responses in animal systems (e.g., Madoff et al., 1996; Mendes et al., 2013). In plant-associated organisms, the first indication that repeat domain variability could confer RCP effectors with an adaptive advantage, by providing a source of functional diversity, flexibility, and/or a means of evading host recognition, was provided by the experimental manipulation of AvrBs3, a TAL effector from X. euvesicatoria (Herbers et al., 1992). Typically, in a compatible interaction with pepper plants, AvrBs3 transcriptionally activates UPA20, a host gene that encodes a basic helix-loop-helix transcription factor, to trigger plant cell hypertrophy (Marois et al., 2002; Kay et al., 2007). However, in an incompatible interaction, AvrBs3 transcriptionally activates Bs3, a pepper gene that encodes an executor resistance protein with homology to flavin monooxygenases, to trigger host immunity (Römer et al., 2007, 2009). To dissect the molecular basis of Bs3-dependent immunity, Herbers et al. (1992) generated random deletion derivatives of AvrBs3 that differed in their repeat unit number. While most AvrBs3 deletion derivatives lost their ability to trigger Bs3-dependent immunity, others gained a new host specificity, triggering immunity in pepper plants carrying Bs3-E, an allele of Bs3 (Herbers et al., 1992). This research, which was subsequently confirmed by repeat domain swaps between other TAL effectors (e.g., Yang et al., 2005), demonstrated that it is the order, and thus the sequence, of TAL repeat units that determines host specificity. In addition, this research raised the possibility that recombination within or between the repeat domains of TAL effectors could produce novel effectors capable of activating different host genes (and thus promoting different host interaction phenotypes) as a consequence of their altered DNA recognition specificities. Indeed, evidence for inter- and intra-genic recombination events between TAL effectors has since been provided (Yang and Gabriel, 1995; Yang et al., 2005).
Aside from those present in TAL effectors, other repeat domains have been implicated in the adaptive evolution of RCP effectors from plant-associated organisms. An example is provided by the hypervariable (Gp-HYP) effectors of the potato cyst nematode, Globodera pallida, which are targeted to the host apoplast throughout biotrophy, and are required for successful root colonization (Eves-van den Akker et al., 2014). Gp-HYP effectors, which possess several conserved regions and a central repeat domain, are encoded by a large and incredibly complex gene family. Based on repeat domain amino acid sequence, these effectors can be assigned to one of three subfamilies (Gp-HYP-1, -2, and -3), with members of Gp-HYP-1 and -3 demonstrating high variability in the number, sequence, and order of their tandem repeats (Eves-van den Akker et al., 2014). Notably, Gp-HYP genes exhibit unparalleled diversity between individuals of the same population, with no two nematodes possessing the same genetic complement of Gp-HYP-1 and -3 genes. While it remains unclear what functional role the Gp-HYP repeat domains play in the context of plant parasitism by G. pallida, it has been suggested that their variability may reflect functional diversity, possibly in specificity of ligand binding. It has also been suggested that this variability may reflect the need to evade host recognition, possibly providing an explanation as to why breeding broad-spectrum resistance against this nematode has been so difficult (Eves-van den Akker et al., 2014). In another example, it has been suggested that the duplication and subsequent sequence diversification of CLE-like repeats present in the GrCLE effectors of G. rostochiensis may represent an important mechanism for generating functional diversity required for host parasitism. This is based on the finding that the ectopic over-expression of different GrCLE RCP effectors in A. thaliana leads to a wide range of plant phenotypes (Lu et al., 2009).
For several RCP effectors, including ATR1 of H. arabidopsidis (and other RXLR effectors from plant-pathogenic oomycetes), as well as AvrM-A of M. lini, and AvrPtoB of Pst, sequence diversification has been shown to play a particularly important role in driving repeat domain evolution, with the repeat units present in these effectors lacking significant amino acid sequence homology (Jiang et al., 2008; Dong et al., 2009; Chou et al., 2011; Ve et al., 2013). Instead, typically only those amino acid residues required for maintenance or stabilization of the overall tertiary fold or structural core have remained conserved or physicochemically similar between repeat units (Cheng et al., 2011; Chou et al., 2011; Ve et al., 2013). This in turn has provided these effectors with a conserved structural framework for rapid diversification, a feature that may promote functional diversity, flexibility, and/or a means of evading host recognition. Certainly, the repeat units of AvrPtoB provide an excellent example of functional flexibility. As mentioned previously, the N terminus and central region of this effector each carry a single repeat unit that adopts a four-helix bundle fold (repeat units one and two, respectively; Figures 1A,B), while the C terminus carries a U-box-type E3 ubiquitin ligase domain (Abramovitch et al., 2006; Janjusevic et al., 2006; Dong et al., 2009; Cheng et al., 2011). Remarkably, both repeat units play distinct and multiple roles in modulating host immune responses. For example, repeat units one and two bind and inhibit the kinase domain of the PM-localized host LysM-RLK and LRR-RLK immune receptors, Bti9 and BAK1, respectively, to suppress immunity-related signaling (Göhre et al., 2008; Shan et al., 2008; Cheng et al., 2011; Zeng et al., 2012). Repeat units one and two also bind the kinase domain of the LysM-RLK CERK1 and LRR-RLK FLS2 immune receptors, respectively, which may promote their ubiquitination and subsequent proteasome-dependent degradation via the AvrPtoB E3 ligase domain (Göhre et al., 2008; Gimenez-Ibanez et al., 2009). In addition, repeat unit one interacts with the host receptor-like cytoplasmic kinase (RLCK) Pto, while repeat unit two interacts with Pto and a related host RLCK, Fen (Rosebrock et al., 2007; Dong et al., 2009; Mathieu et al., 2014). Of note, in line with the observed sequence diversity, structural analyses have determined that repeat unit one interacts with the Pto kinase in a different orientation to that of repeat unit two with the BAK1 kinase domain (Figures 1A,B) (Dong et al., 2009; Cheng et al., 2011). Interestingly, in conjunction with Prf, an immune receptor of tomato, Pto is able to activate host immunity following its interaction with AvrPtoB (Kim et al., 2002; Mucyn et al., 2006; Dong et al., 2009). Fen however, can only activate host immunity in the absence of the E3 ubiquitin ligase domain (Rosebrock et al., 2007). It has now been shown that interaction of either Pto or Fen with repeat unit two results in the proteasome-dependent degradation of these RLCKs as above (Rosebrock et al., 2007; Mathieu et al., 2014). Pto however, is able to resist AvrPtoB-mediated degradation and activate Prf-dependent immunity following its interaction with repeat unit one, as this repeat unit is further away from the E3 ubiquitin ligase domain (Mathieu et al., 2014). It has been suggested that Pto and Fen evolved as decoys of the aforementioned non-cytoplasmic kinases to provide immunity against Pst (Block and Alfano, 2011).
Conclusion and Perspective
Analyses of protein sequence and tertiary structure have revealed that several effectors of plant-associated organisms are RCPs. As reviewed here, repeat domains play diverse roles in RCP effector function. Furthermore, repeat domains may contribute to the rapid adaptive evolution of RCP effectors, providing a source of functional diversity, flexibility, and/or a means of evading host recognition. With these points in mind, it is perhaps not surprising that increased attention has been given to the identification of RCP effectors from plant-associated organisms (e.g., Mueller et al., 2008; Raffaele et al., 2010; Rudd et al., 2010; Saunders et al., 2012; Rafiqi et al., 2013). Undoubtedly, as (1) more genomes of plant-associated organisms are sequenced; (2) the tools of repeat identification become more powerful; and (3) additional effectors are structurally characterized, many more RCP effectors will be identified. The ongoing challenge will be to understand the precise roles that repeat domains play in the function and adaptive evolution of these effectors. Curiously, many of the repeat domain classes discussed in this review are also co-opted by plants to mediate ligand recognition and/or signaling associated with symbiosis, immunity, as well as physiology and development (Palma et al., 2005; Wang et al., 2006; Laluk et al., 2011; Gust et al., 2012; Böhm et al., 2014; Cui et al., 2015; Kucukoglu and Nilsson, 2015). Thus, as shown for the CLE-like repeats of GrCLE1 from G. rostochiensis (Lu et al., 2009; Guo et al., 2011), it is likely that many RCP effector repeat domains mimic host components associated with these processes to facilitate colonization.
Although not discussed in this review, we acknowledge that repeat domains can be intrinsically disordered (ID); a feature characterized by conformational flexibility and a lack of secondary or tertiary structure under physiological conditions (Dyson and Wright, 2005). In fact, repetitive sequence, along with a preponderance of charged and hydrophilic amino acid residues, is often a hallmark of ID (Dyson and Wright, 2005). Like the ordered (structured) repeat domains described above, ID regions carry out diverse roles in protein function, ranging from providing a flexible linker between structured domains, to mediating protein–protein interactions (Dyson and Wright, 2005). To date, examples of RCP effectors with such a repeat domain architecture remain limited, although ID has been predicted for the P/Q-rich repeats of HopI1, a type III effector from the Brassicaceae leaf spot pathogen, P. syringae pv. maculicola (Table 1; Jelenska et al., 2010; Marín and Ott, 2014). Of relevance, many ID regions are known to undergo induced folding upon interaction with their physiological targets, a process that gives rise to the unusual combination of low affinity and high specificity, which may allow these interactions to be readily reversible or may confer flexibility and promiscuity to target binding (Dyson and Wright, 2005). Furthermore, likely owing to a lack of structural constraints, ID protein sequences often evolve at a faster rate than ordered protein sequences, acquiring a greater number of single amino acid substitutions, insertions, deletions, and repeat unit expansions (Brown et al., 2011; Nilsson et al., 2011). Consequently, ID repeat domains are also of great interest to understanding how RCP effectors circumvent host recognition, or acquire novel, altered, and extended effector functionalities that further enhance the colonization of susceptible hosts (Marín et al., 2013; Marín and Ott, 2014).
Author Contributions
CM, JB, and MT conceived the review. CM wrote the manuscript. CM and CH prepared Figures 1, 2. CM and JB constructed Tables 1–3. CM, JB, CH, and MT critically revised the manuscript. All authors approved the final version of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Erik Rikkerink and Xiaolin Sun (The New Zealand Institute for Plant & Food Research) for critically reviewing the manuscript. CM acknowledges financial support provided by The New Zealand Bio-Protection Research Centre (BPRC), Lincoln University.
References
Abramovitch, R. B., Janjusevic, R., Stebbins, C. E., and Martin, G. B. (2006). Type III effector AvrPtoB requires intrinsic E3 ubiquitin ligase activity to suppress plant cell death and immunity. Proc. Natl. Acad. Sci. U.S.A. 103, 2851–2856. doi: 10.1073/pnas.0507892103
Abramovitch, R. B., Kim, Y.-J., Chen, S., Dickman, M. B., and Martin, G. B. (2003). Pseudomonas type III effector AvrPtoB induces plant disease susceptibility by inhibition of host programmed cell death. EMBO J. 22, 60–69. doi: 10.1093/emboj/cdg006
Aggarwal, R., Subramanyam, S., Zhao, C., Chen, M. S., Harris, M. O., and Stuart, J. J. (2014). Avirulence effector discovery in a plant galling and plant parasitic arthropod, the Hessian fly (Mayetiola destructor). PLoS ONE 9:e100958. doi: 10.1371/journal.pone.0100958
Allen, R. L., Bittner-Eddy, P. D., Grenville-Briggs, L. J., Meitz, J. C., Rehmany, A. P., Rose, L. E., et al. (2004). Host-parasite coevolutionary conflict between Arabidopsis and downy mildew. Science 306, 1957–1960. doi: 10.1126/science.1104022
Allen, R. L., Meitz, J. C., Baumber, R. E., Hall, S. A., Lee, S. C., Rose, L. E., et al. (2008). Natural variation reveals key amino acids in a downy mildew effector that alters recognition specificity by an Arabidopsis resistance gene. Mol. Plant Pathol. 9, 511–523. doi: 10.1111/j.1364-3703.2008.00481.x
Angot, A., Peeters, N., Lechner, E., Vailleau, F., Baud, C., Gentzbittel, L., et al. (2006). Ralstonia solanacearum requires F-box-like domain-containing type III effectors to promote disease on several host plants. Proc. Natl. Acad. Sci. U.S.A. 103, 14620–14625. doi: 10.1073/pnas.0509393103
Bailey, T. L., Johnson, J., Grant, C. E., and Noble, W. S. (2015). The MEME suite. Nucleic Acids Res. 43, W39–W49. doi: 10.1093/nar/gkv416
Block, A., and Alfano, J. R. (2011). Plant targets for Pseudomonas syringae type III effectors: virulence targets or guarded decoys? Curr. Opin. Microbiol. 14, 39–46. doi: 10.1016/j.mib.2010.12.011
Boch, J., and Bonas, U. (2010). Xanthomonas AvrBs3 family-type III effectors: discovery and function. Ann. Rev. Phytopathol. 48, 419–436. doi: 10.1146/annurev-phyto-080508-081936
Boch, J., Scholze, H., Schornack, S., Landgraf, A., Hahn, S., Kay, S., et al. (2009). Breaking the code of DNA binding specificity of TAL-type III effectors. Science 326, 1509–1512. doi: 10.1126/science.1178811
Böhm, H., Albert, I., Fan, L., Reinhard, A., and Nürnberger, T. (2014). Immune receptor complexes at the plant cell surface. Curr. Opin. Plant Biol. 20, 47–54. doi: 10.1016/j.pbi.2014.04.007
Bolton, M. D., van Esse, H. P., Vossen, J. H., de Jonge, R., Stergiopoulos, I., Stulemeijer, I. J., et al. (2008). The novel Cladosporium fulvum lysin motif effector Ecp6 is a virulence factor with orthologues in other fungal species. Mol. Microbiol. 69, 119–136. doi: 10.1111/j.1365-2958.2008.06270.x
Bos, J. I., Prince, D., Pitino, M., Maffei, M. E., Win, J., and Hogenhout, S. A. (2010). A functional genomics approach identifies candidate effectors from the aphid species Myzus persicae (green peach aphid). PLoS Genet. 6:e1001216. doi: 10.1371/journal.pgen.1001216
Boutemy, L. S., King, S. R. F., Win, J., Hughes, R. K., Clarke, T. A., Blumenschein, T. M. A., et al. (2011). Structures of Phytophthora RXLR effector proteins. J. Biol. Chem. 286, 35834–35842. doi: 10.1074/jbc.M111.262303
Bozkurt, T. O., Schornack, S., Banfield, M. J., and Kamoun, S. (2012). Oomycetes, effectors, and all that jazz. Curr. Opin. Plant Biol. 15, 483–492. doi: 10.1016/j.pbi.2012.03.008
Brown, C. J., Johnson, A. K., Dunker, A. K., and Daughdrill, G. W. (2011). Evolution and disorder. Curr. Opin. Struct. Biol. 21, 441–446. doi: 10.1016/j.sbi.2011.02.005
Castagnone-Sereno, P., Semblat, J.-P., and Castagnone, C. (2009). Modular architecture and evolution of the map-1 gene family in the root-knot nematode Meloidogyne incognita. Mol. Genet. Genomics 282, 547–554. doi: 10.1007/s00438-009-0487-x
Catanzariti, A.-M., Dodds, P. N., Lawrence, G. J., Ayliffe, M. A., and Ellis, J. G. (2006). Haustorially expressed secreted proteins from flax rust are highly enriched for avirulence elicitors. Plant Cell 18, 243–256. doi: 10.1105/tpc.105.035980
Catanzariti, A.-M., Dodds, P. N., Ve, T., Kobe, B., Ellis, J. G., and Staskawicz, B. J. (2010). The AvrM effector from flax rust has a structured C-terminal domain and interacts directly with the M resistance protein. Mol. Plant Microbe Interact. 23, 49–57. doi: 10.1094/MPMI-23-1-0049
Chen, L. Q., Hou, B. H., Lalonde, S., Takanaga, H., Hartung, M. L., Qu, X. Q., et al. (2010). Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 468, 527–532. doi: 10.1038/nature09606
Chen, S., Lang, P., Chronis, D., Zhang, S., De Jong, W. S., Mitchum, M. G., et al. (2015). In planta processing and glycosylation of a nematode CLAVATA3/ENDOSPERM SURROUNDING REGION-like effector and its interaction with a host CLAVATA2-like receptor to promote parasitism. Plant Physiol. 167, 262–272. doi: 10.1104/pp.114.251637
Cheng, W., Munkvold, K. R., Gao, H., Mathieu, J., Schwizer, S., Wang, S., et al. (2011). Structural analysis of Pseudomonas syringae AvrPtoB bound to host BAK1 reveals two similar kinase-interacting domains in a type III effector. Cell Host Microbe 10, 616–626. doi: 10.1016/j.chom.2011.10.013
Chosed, R., Tomchick, D. R., Brautigam, C. A., Mukherjee, S., Negi, V. S., Machius, M., et al. (2007). Structural analysis of Xanthomonas XopD provides insights into substrate specificity of ubiquitin-like protein proteases. J. Biol. Chem. 282, 6773–6782. doi: 10.1074/jbc.M608730200
Chou, S., Krasileva, K. V., Holton, J. M., Steinbrenner, A. D., Alber, T., and Staskawicz, B. J. (2011). Hyaloperonospora arabidopsidis ATR1 effector is a repeat protein with distributed recognition surfaces. Proc. Natl. Acad. Sci. U.S.A. 108, 13323–13328. doi: 10.1073/pnas.1109791108
Chu, Z., Yuan, M., Yao, J., Ge, X., Yuan, B., Xu, C., et al. (2006). Promoter mutations of an essential gene for pollen development result in disease resistance in rice. Genes Dev. 20, 1250–1255. doi: 10.1101/gad.1416306
Cui, H., Tsuda, K., and Parker, J. E. (2015). Effector-triggered immunity: from pathogen perception to robust defense. Ann. Rev. Plant Biol. 66, 487–511. doi: 10.1146/annurev-arplant-050213-040012
Cunnac, S., Occhialini, A., Barberis, P., Boucher, C., and Genin, S. (2004). Inventory and functional analysis of the large Hrp regulon in Ralstonia solanacearum: identification of novel effector proteins translocated to plant host cells through the type III secretion system. Mol. Microbiol. 53, 115–128. doi: 10.1111/j.1365-2958.2004.04118.x
de Jonge, R., van Esse, H. P., Kombrink, A., Shinya, T., Desaki, Y., Bours, R., et al. (2010). Conserved fungal LysM effector Ecp6 prevents chitin-triggered immunity in plants. Science 329, 953–955. doi: 10.1126/science.1190859
de Lange, O., Schreiber, T., Schandry, N., Radeck, J., Braun, K. H., Koszinowski, J., et al. (2013). Breaking the DNA-binding code of Ralstonia solanacearum TAL effectors provides new possibilities to generate plant resistance genes against bacterial wilt disease. New Phytol. 199, 773–786. doi: 10.1111/nph.12324
Deng, D., Yan, C., Pan, X., Mahfouz, M., Wang, J., Zhu, J. K., et al. (2012). Structural basis for sequence-specific recognition of DNA by TAL effectors. Science 335, 720–723. doi: 10.1126/science.1215670
Deslandes, L., and Rivas, S. (2012). Catch me if you can: bacterial effectors and plant targets. Trends Plant Sci. 17, 644–655. doi: 10.1016/j.tplants.2012.06.011
de Torres, M., Mansfield, J. W., Grabov, N., Brown, I. R., Ammouneh, H., Tsiamis, G., et al. (2006). Pseudomonas syringae effector AvrPtoB suppresses basal defence in Arabidopsis. Plant J. 47, 368–382. doi: 10.1111/j.1365-313X.2006.02798.x
Dong, J., Xiao, F., Fan, F., Gu, L., Cang, H., Martin, G. B., et al. (2009). Crystal structure of the complex between Pseudomonas effector AvrPtoB and the tomato Pto kinase reveals both a shared and a unique interface compared with AvrPto-Pto. Plant Cell 21, 1846–1859. doi: 10.1105/tpc.109.066878
Dong, S., Stam, R., Cano, L. M., Song, J., Sklenar, J., Yoshida, K., et al. (2014). Effector specialization in a lineage of the Irish potato famine pathogen. Science 343, 552–555. doi: 10.1126/science.1246300
Dou, D., and Zhou, J.-M. (2012). Phytopathogen effectors subverting host immunity: different foes, similar battleground. Cell Host Microbe 12, 484–495. doi: 10.1016/j.chom.2012.09.003
Dyson, H. J., and Wright, P. E. (2005). Intrinsically unstructured proteins and their functions. Nat. Rev. Mol. Cell Biol. 6, 197–208. doi: 10.1038/nrm1589
Elzinga, D. A., De Vos, M., and Jander, G. (2014). Suppression of plant defenses by a Myzus persicae (green peach aphid) salivary effector protein. Mol. Plant Microbe Interact. 27, 747–756. doi: 10.1094/MPMI-01-14-0018-R
Eves-van den Akker, S., Lilley, C. J., Jones, J. T., and Urwin, P. E. (2014). Identification and characterisation of a hyper-variable apoplastic effector gene family of the potato cyst nematodes. PLoS Pathog. 10:e1004391. doi: 10.1371/journal.ppat.1004391
Feng, F., Yang, F., Rong, W., Wu, X., Zhang, J., Chen, S., et al. (2012). A Xanthomonas uridine 5′-monophosphate transferase inhibits plant immune kinases. Nature 485, 114–118. doi: 10.1038/nature10962
Gao, H., Wu, X., Chai, J., and Han, Z. (2012). Crystal structure of a TALE protein reveals an extended N-terminal DNA binding region. Cell Res. 22, 1716–1720. doi: 10.1038/cr.2012.156
Gaulin, E., Dramé, N., Lafitte, C., Torto-Alalibo, T., Martinez, Y., Ameline-Torregrosa, C., et al. (2006). Cellulose binding domains of a Phytophthora cell wall protein are novel pathogen-associated molecular patterns. Plant Cell 18, 1766–1777. doi: 10.1105/tpc.105.038687
Gaulin, E., Jauneau, A., Villalba, F., Rickauer, M., Esquerré-Tugayé, M. T., and Bottin, A. (2002). The CBEL glycoprotein of Phytophthora parasitica var-nicotianae is involved in cell wall deposition and adhesion to cellulosic substrates. J. Cell Sci. 115, 4565–4575. doi: 10.1242/jcs.00138
Gemayel, R., Vinces, M. D., Legendre, M., and Verstrepen, K. J. (2010). Variable tandem repeats accelerate evolution of coding and regulatory sequences. Ann. Rev. Genet. 44, 445–477. doi: 10.1146/annurev-genet-072610-155046
Göhre, V., Spallek, T., Häweker, H., Mersmann, S., Mentzel, T., Boller, T., et al. (2008). Plant pattern-recognition receptor FLS2 is directed for degradation by the bacterial ubiquitin ligase AvrPtoB. Curr. Biol. 18, 1824–1832. doi: 10.1016/j.cub.2008.10.063
Gimenez-Ibanez, S., Hann, D. R., Ntoukakis, V., Petutschnig, E., Lipka, V., and Rathjen, J. P. (2009). AvrPtoB targets the LysM receptor kinase CERK1 to promote bacterial virulence on plants. Curr. Biol. 19, 423–429. doi: 10.1016/j.cub.2009.01.054
Goss, E. M., Press, C. M., and Grünwald, N. J. (2013). Evolution of RXLR-class effectors in the oomycete plant pathogen Phytophthora ramorum. PLoS ONE 8:e79347. doi: 10.1371/journal.pone.0079347
Görnhardt, B., Rouhara, I., and Schmelzer, E. (2000). Cyst germination proteins of the potato pathogen Phytophthora infestans share homology with human mucins. Mol. Plant Microbe Interact. 13, 32–42. doi: 10.1094/MPMI.2000.13.1.32
Grove, T. Z., Cortajarena, A. L., and Regan, L. (2008). Ligand binding by repeat proteins: natural and designed. Curr. Opin. Struct. Biol. 18, 507–515. doi: 10.1016/j.sbi.2008.05.008
Guo, Y., Ni, J., Denver, R., Wang, X., and Clark, S. E. (2011). Mechanisms of molecular mimicry of plant CLE peptide ligands by the parasitic nematode Globodera rostochiensis. Plant Phys. 157, 476–484. doi: 10.1104/pp.111.180554
Gust, A. A., Willmann, R., Desaki, Y., Grabherr, H. M., and Nürnberger, T. (2012). Plant LysM proteins: modules mediating symbiosis and immunity. Trends Plant Sci. 17, 495–502. doi: 10.1016/j.tplants.2012.04.003
Guttman, D. S., Vinatzer, B. A., Sarkar, S. F., Ranall, M. V., Kettler, G., and Greenberg, J. T. (2002). A functional screen for the type III (Hrp) secretome of the plant pathogen Pseudomonas syringae. Science 295, 1722–1726. doi: 10.1126/science.295.5560.1722
Guy, E., Lautier, M., Chabannes, M., Roux, B., Lauber, E., Arlat, M., et al. (2013). xopAC-triggered immunity against Xanthomonas depends on Arabidopsis receptor-like cytoplasmic kinase genes PBL2 and RIPK. PLoS ONE 8:e73469. doi: 10.1371/journal.pone.0073469
He, P., Shan, L., Lin, N. C., Martin, G. B., Kemmerling, B., Nürnberger, T., et al. (2006). Specific bacterial suppressors of MAMP signaling upstream of MAPKKK in Arabidopsis innate immunity. Cell 125, 563–575. doi: 10.1016/j.cell.2006.02.047
Herbers, K., Conrads-Strauch, J., and Bonas, U. (1992). Race-specificity of plant resistance to bacterial spot disease determined by repetitive motifs in a bacterial avirulence protein. Nature 356, 172–174. doi: 10.1038/356172a0
Heuer, H., Yin, Y. N., Xue, Q. Y., Smalla, K., and Guo, J. H. (2007). Repeat domain diversity of avrBs3-like genes in Ralstonia solanacearum strains and association with host preferences in the field. Appl. Environ. Microbiol. 73, 4379–4384. doi: 10.1128/AEM.00367-07
Hogenhout, S. A., van der Hoorn, R. A. L., Terauchi, R., and Kamoun, S. (2009). Emerging concepts in effector biology of plant-associated organisms. Mol. Plant Microbe Interact. 22, 115–122. doi: 10.1094/MPMI-22-2-0115
Hotson, A., Chosed, R., Shu, H., Orth, K., and Mudgett, M. B. (2003). Xanthomonas type III effector XopD targets SUMO-conjugated proteins in planta. Mol. Microbiol. 50, 377–389. doi: 10.1046/j.1365-2958.2003.03730.x
Janjusevic, R., Abramovitch, R. B., Martin, G. B., and Stebbins, C. E. (2006). A bacterial inhibitor of host programmed cell death defenses is an E3 ubiquitin ligase. Science 311, 222–226. doi: 10.1126/science.1120131
Jaouannet, M., Rodriguez, P. A., Thorpe, P., Lenoir, C. J., MacLeod, R., Escudero-Martinez, C., et al. (2014). Plant immunity in plant-aphid interactions. Front. Plant Sci. 5:663. doi: 10.3389/fpls.2014.00663
Jelenska, J., van Hal, J. A., and Greenberg, J. T. (2010). Pseudomonas syringae hijacks plant stress chaperone machinery for virulence. Proc. Natl. Acad. Sci. U.S.A. 107, 13177–13182. doi: 10.1073/pnas.0910943107
Jelenska, J., Yao, N., Vinatzer, B. A., Wright, C. M., Brodsky, J. L., and Greenberg, J. T. (2007). A J domain virulence effector of Pseudomonas syringae remodels host chloroplasts and suppresses defenses. Curr. Biol. 17, 499–508. doi: 10.1016/j.cub.2007.02.028
Jiang, R. H., Tripathy, S., Govers, F., and Tyler, B. M. (2008). RXLR effector reservoir in two Phytophthora species is dominated by a single rapidly evolving superfamily with more than 700 members. Proc. Natl. Acad. Sci. U.S.A. 105, 4874–4879. doi: 10.1073/pnas.0709303105
Kajava, A. V. (2012). Tandem repeats in proteins: from sequence to structure. J. Struct. Biol. 179, 279–288. doi: 10.1016/j.jsb.2011.08.009
Kappel, C., Zachariae, U., Dölker, N., and Grubmüller, H. (2010). An unusual hydrophobic core confers extreme flexibility to HEAT repeat proteins. Biophys. J. 99, 1596–1603. doi: 10.1016/j.bpj.2010.06.032
Kay, S., Hahn, S., Marois, E., Hause, G., and Bonas, U. (2007). A bacterial effector acts as a plant transcription factor and induces a cell size regulator. Science 318, 648–651. doi: 10.1126/science.1144956
Khatib, M., Lafitte, C., Esquerré-Tugayé, M.-T., Bottin, A., and Rickauer, M. (2004). The CBEL elicitor of Phytophthora parasitica var. nicotianae activates defence in Arabidopsis thaliana via three different signalling pathways. New Phytol. 162, 501–510. doi: 10.1111/j.1469-8137.2004.01043.x
Kim, J. G., Li, X., Roden, J. A., Taylor, K. W., Aakre, C. D., Su, B., et al. (2009). Xanthomonas T3S effector XopN suppresses PAMP-triggered immunity and interacts with a tomato atypical receptor-like kinase and TFT1. Plant Cell 21, 1305–1323. doi: 10.1105/tpc.108.063123
Kim, J.-G., Stork, W., and Mudgett, M. B. (2013). Xanthomonas type III effector XopD desumoylates tomato transcription factor SlERF4 to suppress ethylene responses and promote pathogen growth. Cell Host Microbe 13, 143–154. doi: 10.1016/j.chom.2013.01.006
Kim, J.-G., Taylor, K. W., Hotson, A., Keegan, M., Schmelz, E. A., and Mudgett, M. B. (2008). XopD SUMO protease affects host transcription, promotes pathogen growth, and delays symptom development in Xanthomonas-infected tomato leaves. Plant Cell 20, 1915–1929. doi: 10.1105/tpc.108.058529
Kim, Y. J., Lin, N.-C., and Martin, G. B. (2002). Two distinct Pseudomonas effector proteins interact with the Pto kinase and activate plant immunity. Cell 109, 589–598. doi: 10.1016/S0092-8674(02)00743-2
Kloppholz, S., Kuhn, H., and Requena, N. (2011). A secreted fungal effector of Glomus intraradices promotes symbiotic biotrophy. Curr. Biol. 21, 1204–1209. doi: 10.1016/j.cub.2011.06.044
Krasileva, K. V., Dahlbeck, D., and Staskawicz, B. J. (2010). Activation of an Arabidopsis resistance protein is specified by the in planta association of its leucine-rich repeat domain with the cognate oomycete effector. Plant Cell 22, 2444–2458. doi: 10.1105/tpc.110.075358
Kucheryava, N., Bowen, J. K., Sutherland, P. W., Conolly, J. J., Mesarich, C. H., Rikkerink, E. H., et al. (2008). Two novel Venturia inaequalis genes induced upon morphogenetic differentiation during infection and in vitro growth on cellophane. Fungal Genet. Biol. 45, 1329–1339. doi: 10.1016/j.fgb.2008.07.010
Kucukoglu, M., and Nilsson, O. (2015). CLE peptide signaling in plants – the power of moving around. Physiol. Plant. 155, 74–87. doi: 10.1111/ppl.12358
Laluk, K., Abuqamar, S., and Mengiste, T. (2011). The Arabidopsis mitochondria-localized pentatricopeptide repeat protein PGN functions in defense against necrotrophic fungi and abiotic stress tolerance. Plant Phys. 156, 2053–2068. doi: 10.1104/pp.111.177501
Lanver, D., Berndt, P., Tollot, M., Naik, V., Vranes, M., Warmann, T., et al. (2014). Plant surface cues prime Ustilago maydis for biotrophic development. PLoS Pathog. 10:e1004272. doi: 10.1371/journal.ppat.1004272
Lanver, D., Mendoza-Mendoza, A., Brachmann, A., and Kahmann, R. (2010). Sho1 and Msb2-related proteins regulate appressorium development in the smut fungus Ustilago maydis. Plant Cell 22, 2085–2101. doi: 10.1105/tpc.109.073734
Leonelli, L., Pelton, J., Schoeffler, A., Dahlbeck, D., Berger, J., Wemmer, D. E., et al. (2011). Structural elucidation and functional characterization of the Hyaloperonospora arabidopsidis effector protein ATR13. PLoS Pathog. 7:e1002428. doi: 10.1371/journal.ppat.1002428
Li, L., Atef, A., Piatek, A., Ali, Z., Piatek, M., Aouida, M., et al. (2013). Characterization and DNA-binding specificities of Ralstonia TAL-like effectors. Mol. Plant 6, 1318–1330. doi: 10.1093/mp/sst006
Lo Presti, L., Lanver, D., Schweizer, G., Tanaka, S., Liang, L., Tollot, M., et al. (2015). Fungal effectors and plant susceptibility. Ann. Rev. Plant Biol. 66, 513–545. doi: 10.1146/annurev-arplant-043014-114623
Lu, S. W., Chen, S., Wang, J., Yu, H., Chronis, D., Mitchum, M. G., et al. (2009). Structural and functional diversity of CLAVATA3/ESR (CLE)-like genes from the potato cyst nematode Globodera rostochiensis. Mol. Plant Microbe Interact. 22, 1128–1142. doi: 10.1094/MPMI-22-9-1128
Luo, H., and Nijveen, H. (2014). Understanding and identifying amino acid repeats. Brief. Bioinformatics. 15, 582–591. doi: 10.1093/bib/bbt003
Macho, A. P., Guidot, A., Barberis, P., Beuzón, C. R., and Genin, S. (2010). A competitive index assay identifies several Ralstonia solanacearum type III effector mutant strains with reduced fitness in host plants. Mol. Plant Microbe Interact. 23, 1197–1205. doi: 10.1094/MPMI-23-9-1197
Madoff, L. C., Michel, J. L., Gong, E. W., Kling, D. E., and Kasper, D. L. (1996). Group B streptococci escape host immunity by deletion of tandem repeat elements of the alpha C protein. Proc. Natl. Acad. Sci. U.S.A. 93, 4131–4136. doi: 10.1073/pnas.93.9.4131
Mak, A. N., Bradley, P., Cernadas, R. A., Bogdanove, A. J., and Stoddard, B. L. (2012). The crystal structure of TAL effector PthXo1 bound to its DNA target. Science 335, 716–719. doi: 10.1126/science.1216211
Marín, M., and Ott, T. (2014). Intrinsic disorder in plant proteins and phytopathogenic bacterial effectors. Chem. Rev. 114, 6912–6932. doi: 10.1021/cr400488d
Marín, M., Uversky, V. N., and Ott, T. (2013). Intrinsic disorder in pathogen effectors: protein flexibility as an evolutionary hallmark in a molecular arms race. Plant Cell 25, 3153–3157. doi: 10.1105/tpc.113.116319
Marois, E., Van den Ackerveken, G., and Bonas, U. (2002). The Xanthomonas type III effector protein AvrBs3 modulates plant gene expression and induces cell hypertrophy in the susceptible host. Mol. Plant Microbe Interact. 15, 637–646. doi: 10.1094/MPMI.2002.15.7.637
Mathieu, J., Schwizer, S., and Martin, G. B. (2014). Pto kinase binds two domains of AvrPtoB and its proximity to the effector E3 ligase determines if it evades degradation and activates plant immunity. PLoS Pathog. 10:e1004227. doi: 10.1371/journal.ppat.1004227
Mendes, T. A., Lobo, F. P., Rodrigues, T. S., Rodrigues-Luiz, G. F., daRocha, W. D., Fujiwara, R. T., et al. (2013). Repeat-enriched proteins are related to host cell invasion and immune evasion in parasitic protozoa. Mol. Biol. Evol. 30, 951–963. doi: 10.1093/molbev/mst001
Mesarich, C. H., Schmitz, M., Tremouilhac, P., McGillivray, D. J., Templeton, M. D., and Dingley, A. J. (2012). Structure, dynamics and domain organization of the repeat protein Cin1 from the apple scab fungus. BBA Proteins Proteom. 1824, 1118–1128. doi: 10.1016/j.bbapap.2012.06.015
Mitchum, M. G., Hussey, R. S., Baum, T. J., Wang, X., Elling, A. A., Wubben, M., et al. (2013). Nematode effector proteins: an emerging paradigm of parasitism. New Phytol. 199, 879–894. doi: 10.1111/nph.12323
Moscou, M. J., and Bogdanove, A. J. (2009). A simple cipher governs DNA recognition by TAL effectors. Science 326, 1501. doi: 10.1126/science.1178817
Mucyn, T. S., Clemente, A., Andriotis, V. M., Balmuth, A. L., Oldroyd, G. E., Staskawicz, B. J., et al. (2006). The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate specific plant immunity. Plant Cell 18, 2792–2806. doi: 10.1105/tpc.106.044016
Mueller, O., Kahmann, R., Aguilar, G., Trejo-Aguilar, B., Wu, A., and de Vries, R. P. (2008). The secretome of the maize pathogen Ustilago maydis. Fungal Genet. Biol. 45(Suppl. 1), S63–S70. doi: 10.1016/j.fgb.2008.03.012
Mukaihara, T., and Tamura, N. (2009). Identification of novel Ralstonia solanacearum type III effector proteins through translocation analysis of hrpB-regulated gene products. Microbiology 155, 2235–2244. doi: 10.1099/mic.0.027763-0
Müller, O., Schreier, P. H., and Uhrig, J. F. (2008). Identification and characterization of secreted and pathogenesis-related proteins in Ustilago maydis. Mol. Genet. Genomics 279, 27–39. doi: 10.1007/s00438-007-0291-4
Nilsson, J., Grahn, M., and Wright, A. P. H. (2011). Proteome-wide evidence for enhanced positive Darwinian selection within intrinsically disordered regions in proteins. Genome Biol. 12:R65. doi: 10.1186/gb-2011-12-7-r65
Nissan, G., Manulis-Sasson, S., Chalupowicz, L., Teper, D., Yeheskel, A., Pasmanik-Chor, M., et al. (2012). The type III effector HsvG of the gall-forming Pantoea agglomerans mediates expression of the host gene HSVGT. Mol. Plant Microbe Interact. 25, 231–240. doi: 10.1094/MPMI-06-11-0173
Nissan, G., Manulis-Sasson, S., Weinthal, D., Mor, H., Sessa, G., and Barash, I. (2006). The type III effectors HsvG and HsvB of gall-forming Pantoea agglomerans determine host specificity and function as transcriptional activators. Mol. Microbiol. 61, 1118–1131. doi: 10.1111/j.1365-2958.2006.05301.x
Palma, K., Zhang, Y., and Li, X. (2005). An importin α homolog, MOS6, plays an important role in plant innate immunity. Curr. Biol. 15, 1129–1135. doi: 10.1016/j.cub.2005.05.022
Peeters, N., Carrère, S., Anisimova, M., Plener, L., Cazalé, A. C., and Genin, S. (2013). Repertoire, unified nomenclature and evolution of the Type III effector gene set in the Ralstonia solanacearum species complex. BMC Genomics 14:859. doi: 10.1186/1471-2164-14-859
Petre, B., Lorrain, C., Saunders, D. G. O., Win, J., Sklenar, J., Duplessis, S., et al. (2015a). Rust fungal effectors mimic host transit peptides to translocate into chloroplasts. Cell. Microbiol. doi: 10.1111/cmi.12530. [Epub ahead of print].
Petre, B., Saunders, D. G., Sklenar, J., Lorrain, C., Win, J., Duplessis, S., et al. (2015b). Candidate effector proteins of the rust pathogen Melampsora larici-populina target diverse plant cell compartments. Mol. Plant Microbe Interact. 28, 689–700. doi: 10.1094/MPMI-01-15-0003-R
Pitino, M., Coleman, A. D., Maffei, M. E., Ridout, C. J., and Hogenhout, S. A. (2011). Silencing of aphid genes by dsRNA feeding from plants. PLoS ONE 6:e25709. doi: 10.1371/journal.pone.0025709
Pitino, M., and Hogenhout, S. A. (2013). Aphid protein effectors promote aphid colonization in a plant species-specific manner. Mol. Plant Microbe Interact. 26, 130–139. doi: 10.1094/MPMI-07-12-0172-FI
Pradhan, B. B., Ranjan, M., and Chatterjee, S. (2012). XadM, a novel adhesin of Xanthomonas oryzae pv. oryzae, exhibits similarity to Rhs family proteins and is required for optimum attachment, biofilm formation, and virulence. Mol. Plant Microbe Interact. 25, 1157–1170. doi: 10.1094/MPMI-02-12-0049-R
Raffaele, S., Win, J., Cano, L. M., and Kamoun, S. (2010). Analyses of genome architecture and gene expression reveal novel candidate virulence factors in the secretome of Phytophthora infestans. BMC Genomics 11:637. doi: 10.1186/1471-2164-11-637
Rafiqi, M., Gan, P. H. P., Ravensdale, M., Lawrence, G. J., Ellis, J. G., Jones, D. A., et al. (2010). Internalization of flax rust avirulence proteins into flax and tobacco cells can occur in the absence of the pathogen. Plant Cell 22, 2017–2032. doi: 10.1105/tpc.109.072983
Rafiqi, M., Jelonek, L., Akum, N. F., Zhang, F., and Kogel, K.-H. (2013). Effector candidates in the secretome of Piriformospora indica, a ubiquitous plant-associated fungus. Front. Plant Sci. 4:228. doi: 10.3389/fpls.2013.00228
Rehmany, A. P., Gordon, A., Rose, L. E., Allen, R. L., Armstrong, M. R., Whisson, S. C., et al. (2005). Differential recognition of highly divergent downy mildew avirulence gene alleles by RPP1 resistance genes from two Arabidopsis lines. Plant Cell 17, 1839–1850. doi: 10.1105/tpc.105.031807
Remigi, P., Anisimova, M., Guidot, A., Genin, S., and Peeters, N. (2011). Functional diversification of the GALA type III effector family contributes to Ralstonia solanacearum adaptation on different plant hosts. New Phytol. 192, 976–987. doi: 10.1111/j.1469-8137.2011.03854.x
Richard, G.-F., and Pâques, F. (2000). Mini- and microsatellite expansions: the recombination connection. EMBO Rep. 1, 122–126. doi: 10.1093/embo-reports/kvd031
Römer, P., Hahn, S., Jordan, T., Strauss, T., Bonas, U., and Lahaye, T. (2007). Plant pathogen recognition mediated by promoter activation of the pepper Bs3 resistance gene. Science 318, 645–648. doi: 10.1126/science.1144958
Römer, P., Strauss, T., Hahn, S., Scholze, H., Morbitzer, R., Grau, J., et al. (2009). Recognition of AvrBs3-like proteins is mediated by specific binding to promoters of matching pepper Bs3 alleles. Plant Physiol. 150, 1697–1712. doi: 10.1104/pp.109.139931
Robold, A. V., and Hardham, A. R. (2005). During attachment Phytophthora spores secrete proteins containing thrombospondin type 1 repeats. Curr. Genet. 47, 307–315. doi: 10.1007/s00294-004-0559-8
Roden, J. A., Belt, B., Ross, J. B., Tachibana, T., Vargas, J., and Mudgett, M. B. (2004). A genetic screen to isolate type III effectors translocated into pepper cells during Xanthomonas infection. Proc. Natl. Acad. Sci. U.S.A. 101, 16624–16629. doi: 10.1073/pnas.0407383101
Rosebrock, T. R., Zeng, L., Brady, J. J., Abramovitch, R. B., Xiao, F., and Martin, G. B. (2007). A bacterial E3 ubiquitin ligase targets a host protein kinase to disrupt plant immunity. Nature 448, 370–374. doi: 10.1038/nature05966
Rudd, J. J., Antoniw, J., Marshall, R., Motteram, J., Fraaije, B., and Hammond-Kosack, K. (2010). Identification and characterisation of Mycosphaerella graminicola secreted or surface-associated proteins with variable intragenic coding repeats. Fungal Genet. Biol. 47, 19–32. doi: 10.1016/j.fgb.2009.10.009
Rutter, W. B., Hewezi, T., Maier, T. R., Mitchum, M. G., Davis, E. L., Hussey, R. S., et al. (2014). Members of the Meloidogyne avirulence protein family contain multiple plant ligand-like motifs. Phytopathology 104, 879–885. doi: 10.1094/PHYTO-11-13-0326-R
Sánchez-Vallet, A., Saleem-Batcha, R., Kombrink, A., Hansen, G., Valkenburg, D. J., Thomma, B. P., et al. (2013). Fungal effector Ecp6 outcompetes host immune receptor for chitin binding through intrachain LysM dimerization. Elife 2:e00790. doi: 10.7554/eLife.00790
Saunders, D. G., Win, J., Cano, L. M., Szabo, L. J., Kamoun, S., and Raffaele, S. (2012). Using hierarchical clustering of secreted protein families to classify and rank candidate effectors of rust fungi. PLoS ONE 7:e29847. doi: 10.1371/journal.pone.0029847
Séjalon-Delmas, N., Mateos, F. V., Bottin, A., Rickauer, M., Dargent, R., and Esquerre-Tugaye, M. T. (1997). Purification, elicitor activity, and cell wall localization of a glycoprotein from Phytophthora parasitica var. nicotianae, a fungal pathogen of tobacco. Phytopathology 87, 899–909. doi: 10.1094/PHYTO.1997.87.9.899
Semblat, J. P., Rosso, M. N., Hussey, R. S., Abad, P., and Castagnone-Sereno, P. (2001). Molecular cloning of a cDNA encoding an amphid-secreted putative avirulence protein from the root-knot nematode Meloidogyne incognita. Mol. Plant Microbe Interact. 14, 72–79. doi: 10.1094/MPMI.2001.14.1.72
Shan, L., He, P., Li, J., Heese, A., Peck, S. C., Nürnberger, T., et al. (2008). Bacterial effectors target the common signaling partner BAK1 to disrupt multiple MAMP receptor-signaling complexes and impede plant immunity. Cell Host Microbe 4, 17–27. doi: 10.1016/j.chom.2008.05.017
Singer, A. U., Schulze, S., Skarina, T., Xu, X., Cui, H., Eschen-Lippold, L., et al. (2013). A pathogen type III effector with a novel E3 ubiquitin ligase architecture. PLoS Pathog. 9:e1003121. doi: 10.1371/journal.ppat.1003121
Sohn, K. H., Lei, R., Nemri, A., and Jones, J. D. (2007). The downy mildew effector proteins ATR1 and ATR13 promote disease susceptibility in Arabidopsis thaliana. Plant Cell 19, 4077–4090. doi: 10.1105/tpc.107.054262
Steinbrenner, A. D., Goritschnig, S., and Staskawicz, B. J. (2015). Recognition and activation domains contribute to allele-specific responses of an Arabidopsis NLR receptor to an oomycete effector protein. PLoS Pathog. 11:e1004665. doi: 10.1371/journal.ppat.1004665
Stergiopoulos, I., De Kock, M. J. D., Lindhout, P., and De Wit, P. J. (2007). Allelic variation in the effector genes of the tomato pathogen Cladosporium fulvum reveals different modes of adaptive evolution. Mol. Plant Microbe Interact. 20, 1271–1283. doi: 10.1094/MPMI-20-10-1271
Taylor, K. W., Kim, J.-G., Su, X. B., Aakre, C. D., Roden, J. A., Adams, C. M., et al. (2012). Tomato TFT1 is required for PAMP-triggered immunity and mutations that prevent T3S effector XopN from binding to TFT1 attenuate Xanthomonas virulence. PLoS Pathog. 8:e1002768. doi: 10.1371/journal.ppat.1002768
Teertstra, W. R., Deelstra, H. J., Vranes, M., Bohlmann, R., Kahmann, R., Kämper, J., et al. (2006). Repellents have functionally replaced hydrophobins in mediating attachment to a hydrophobic surface and in formation of hydrophobic aerial hyphae in Ustilago maydis. Microbiology 152, 3607–3612. doi: 10.1099/mic.0.29034-0
Teertstra, W. R., van der Velden, G. J., de Jong, J. F., Kruijtzer, J. A., Liskamp, R. M., Kroon-Batenburg, L. M., et al. (2009). The filament-specific Rep1-1 repellent of the phytopathogen Ustilago maydis forms functional surface-active amyloid-like fibrils. J. Biol. Chem. 284, 9153–9159. doi: 10.1074/jbc.M900095200
Thomma, B. P. H. J., Nürnberger, T., and Joosten, M. H. A. J. (2011). Of PAMPs and effectors: the blurred PTI-ETI dichotomy. Plant Cell 23, 4–15. doi: 10.1105/tpc.110.082602
Valinsky, L., Manulis, S., Nizan, R., Ezra, D., and Barash, I. (1998). A pathogenicity gene isolated from the pPATH plasmid of Erwinia herbicola pv. gypsophilae determines host specificity. Mol. Plant Microbe Interact. 11, 753–762. doi: 10.1094/MPMI.1998.11.8.753
Ve, T., Williams, S. J., Catanzariti, A.-M., Rafiqi, M., Rahman, M., Ellis, J. G., et al. (2013). Structures of the flax-rust effector AvrM reveal insights into the molecular basis of plant-cell entry and effector-triggered immunity. Proc. Natl. Acad. Sci. U.S.A. 110, 17594–17599. doi: 10.1073/pnas.1307614110
Verstrepen, K. J., and Fink, G. R. (2009). Genetic and epigenetic mechanisms underlying cell-surface variability in protozoa and fungi. Ann. Rev. Genet. 43, 1–24. doi: 10.1146/annurev-genet-102108-134156
Vieira, P., Danchin, E. G. J., Neveu, C., Crozat, C., Jaubert, S., Hussey, R. S., et al. (2011). The plant apoplasm is an important recipient compartment for nematode secreted proteins. J. Exp. Bot. 62, 1241–1253. doi: 10.1093/jxb/erq352
Villalba Mateos, F., Rickauer, M., and Esquerré-Tugayé, M. T. (1997). Cloning and characterization of a cDNA encoding an elicitor of Phytophthora parasitica var. nicotianae that shows cellulose-binding and lectin-like activities. Mol. Plant Microbe Interact. 10, 1045–1053. doi: 10.1094/MPMI.1997.10.9.1045
Wang, G., Roux, B., Feng, F., Guy, E., Li, L., Li, N., et al. (2015b). The decoy substrate of a pathogen effector and a pseudokinase specify pathogen-induced modified-self recognition and immunity in plants. Cell Host Microbe 18, 285–295. doi: 10.1016/j.chom.2015.08.004
Wang, K., Remigi, P., Anisimova, M., Lonjon, F., Kars, I., Kajava, A., et al. (2015a). Functional assignment to positively selected sites in the core type III effector RipG7 from Ralstonia solanacearum. Mol.Plant Pathol. doi: 10.1111/mpp.12302. [Epub ahead of print].
Wang, Y.-S., Pi, L.-Y., Chen, X., Chakrabarty, P. K., Jiang, J., De Leon, A. L., et al. (2006). Rice XA21 binding protein 3 is a ubiquitin ligase required for full Xa21-mediated disease resistance. Plant Cell 18, 3635–3646. doi: 10.1105/tpc.106.046730
White, F. F., Potnis, N., Jones, J. B., and Koebnik, R. (2009). The type III effectors of Xanthomonas. Mol. Plant Pathol. 10, 749–766. doi: 10.1111/j.1364-3703.2009.00590.x
Win, J., Chaparro-Garcia, A., Belhaj, K., Saunders, D. G., Yoshida, K., Dong, S., et al. (2012a). Effector biology of plant-associated organisms: concepts and perspectives. Cold Spring Harb. Symp. Quant. Biol. 77, 235–247. doi: 10.1101/sqb.2012.77.015933
Win, J., Krasileva, K. V., Kamoun, S., Shirasu, K., Staskawicz, B. J., and Banfield, M. J. (2012b). Sequence divergent RXLR effectors share a structural fold conserved across plant pathogenic oomycete species. PLoS Pathog. 8:e1002400. doi: 10.1371/journal.ppat.1002400
Win, J., Morgan, W., Bos, J., Krasileva, K. V., Cano, L. M., Chaparro-Garcia, A., et al. (2007). Adaptive evolution has targeted the C-terminal domain of the RXLR effectors of plant pathogenic oomycetes. Plant Cell 19, 2349–2369. doi: 10.1105/tpc.107.051037
Wösten, H. A., Bohlmann, R., Eckerskorn, C., Lottspeich, F., Bolker, M., and Kahmann, R. (1996). A novel class of small amphipathic peptides affect aerial hyphal growth and surface hydrophobicity in Ustilago maydis. EMBO J. 15, 4274–4281.
Xiao, F., He, P., Abramovitch, R. B., Dawson, J. E., Nicholson, L. K., Sheen, J., et al. (2007). The N-terminal region of Pseudomonas type III effector AvrPtoB elicits Pto-dependent immunity and has two distinct virulence determinants. Plant J. 52, 595–614. doi: 10.1111/j.1365-313X.2007.03259.x
Xu, R.-Q., Blanvillain, S., Feng, J.-X., Jiang, B.-L., Li, X.-Z., Wei, H.-Y., et al. (2008). AvrACXcc8004, a type III effector with a leucine-rich repeat domain from Xanthomonas campestris pathovar campestris confers avirulence in vascular tissues of Arabidopsis thaliana ecotype Col-0. J. Bacteriol. 190, 343–355. doi: 10.1128/JB.00978-07
Yang, B., Sugio, A., and White, F. F. (2005). Avoidance of host recognition by alterations in the repetitive and C-terminal regions of AvrXa7, a type III effector of Xanthomonas oryzae pv. oryzae. Mol. Plant Microbe Interact. 18, 142–149. doi: 10.1094/MPMI-18-0142
Yang, B., Sugio, A., and White, F. F. (2006). Os8N3 is a host disease-susceptibility gene for bacterial blight of rice. Proc. Natl. Acad. Sci. U.S.A. 103, 10503–10508. doi: 10.1073/pnas.0604088103
Yang, B., and White, F. F. (2004). Diverse members of the AvrBs3/PthA family of type III effectors are major virulence determinants in bacterial blight disease of rice. Mol. Plant Microbe Interact. 17, 1192–1200. doi: 10.1094/MPMI.2004.17.11.1192
Yang, Y., and Gabriel, D. W. (1995). Intragenic recombination of a single plant pathogen gene provides a mechanism for the evolution of new host specificities. J. Bacteriol. 177, 4963–4968.
Ye, W., Wang, Y., and Wang, Y. (2015). Bioinformatics analysis reveals abundant short alpha-helices as a common structural feature of oomycete RxLR effector proteins. PLoS ONE 10:e0135240. doi: 10.1371/journal.pone.0135240
Yuan, M., Chu, Z., Li, X., Xu, C., and Wang, S. (2009). Pathogen-induced expressional loss of function is the key factor in race-specific bacterial resistance conferred by a recessive R gene xa13 in rice. Plant Cell Phys. 50, 947–955. doi: 10.1093/pcp/pcp046
Zeng, L., Velásquez, A. C., Munkvold, K. R., Zhang, J., and Martin, G. B. (2012). A tomato LysM receptor-like kinase promotes immunity and its kinase activity is inhibited by AvrPtoB. Plant J. 69, 92–103. doi: 10.1111/j.1365-313X.2011.04773.x
Keywords: repeat-containing protein effectors, plant-associated organisms, microbes, nematodes, insects
Citation: Mesarich CH, Bowen JK, Hamiaux C and Templeton MD (2015) Repeat-containing protein effectors of plant-associated organisms. Front. Plant Sci. 6:872. doi: 10.3389/fpls.2015.00872
Received: 03 August 2015; Accepted: 01 October 2015;
Published: 21 October 2015.
Edited by:
Maryam Rafiqi, Computomics GmbH, GermanyReviewed by:
Ralph Panstruga, RWTH Aachen University, GermanyFrancine Govers, Wageningen University, Netherlands
Copyright © 2015 Mesarich, Bowen, Hamiaux and Templeton. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carl H. Mesarich, Y2FybC5tZXNhcmljaEBnbWFpbC5jb20=